
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Immunol. , 27 April 2021
Sec. Molecular Innate Immunity
Volume 12 - 2021 | https://doi.org/10.3389/fimmu.2021.656965
This article is part of the Research Topic Lysosomal Peptidases in Tumor Immunity View all 5 articles
 Abeer F. Alharbi1,2*
Abeer F. Alharbi1,2* John Parrington1*
John Parrington1*The role of endolysosomal Ca2+ signalling in immunity has been a subject of increasing interest in recent years. Here, we discuss evolving knowledge relating to the contribution of endolysosomal Ca2+ channels that include TPCs, TRPMLs, and P2X4R in physiological processes related to innate and adaptive immunity—including phagocytosis, inflammation, cytokine/chemokine release, dendritic, natural killer, and T cell activation and migration—and we underscore the paucity of clinical studies in this field. Emerging biomedical and translational data have led to important new insights into the critical roles of these channels in immune cell function and the regulation of innate and adaptive immune responses. The evolving immunological significance of endolysosomal Ca2+ signalling warrants further investigations to better characterize the roles of these channels in immunity in order to expand our knowledge about the pathology of inflammatory and autoimmune diseases and develop endolysosomal Ca2+ channels as viable biomarkers and therapeutic and preventive targets for remodelling the immune response.
Innate and adaptive immunity are two fundamental components of the immune system. The cross-talk between innate and adaptive responses is important in maintaining a functional immune system in order to protect the individual against foreign substances such as allergens, toxins, tumour cells, bacteria, and viruses. The innate immune system involves monocytes, macrophages, dendritic cells, mast cells, basophils, neutrophils, eosinophils, and natural killer cells; whereas the adaptive immune system is composed of B cells and T cells. Several studies have indicated that intracellular Ca2+ signalling is critical to maintaining various immune cell functions (1–3) and attributed the development of multiple autoimmune and inflammatory diseases to Ca2+ dysregulation (4, 5).
Ca2+ signalling mediated by endolysosomal channels is emerging as a player in processes related to immune cell functions such as phagocytosis; the release of inflammatory mediators; antigen presentation; inflammation; cellular trafficking; and T cell migration. Endo-lysosomal Ca2+ channels are localized in early, late, and recycling endosomes, lysosomes, and autophagosomes. They are comprised of two-pore channels (TPCs, also known as TPCNs); transient receptor potential cation channels; mucolipins (TRPML); and the P2X4 ATP-activated cation channel. A significant contribution of endolysosomal Ca2+ signalling has been demonstrated in phagocytosis, which is a vital physiological process in cellular immunity mediated by TPCs, TRPML1, and P2X4R (6, 7). TRPML2 is an endolysosomal Ca2+ channel that has been shown to have direct roles in the release of chemokine/cytokine (8). Additionally, TPC1 is an endolysosomal Ca2+ channel that has been reported to be involved in the development of the immune response and the release of inflammatory mediators (9). Although it has become clear that endolysosomal Ca2+ signals play pivotal roles in health and disease, the complex dynamics underlying the regulation of Ca2+ signalling via endolysosomal channels and the involvement of these in physiological processes related to immunity have remained elusive. Here, we highlight the emerging roles of endolysosomal Ca2+ channels in various physiological processes related to immunity (as shown in Figure 1 and Table 1). Our intent is to reveal their potential as key pieces in a puzzle that will help increase understanding of the pathophysiology of autoimmune and inflammatory disorders and develop endolysosomal Ca2+ channels as targets for future immunotherapy.
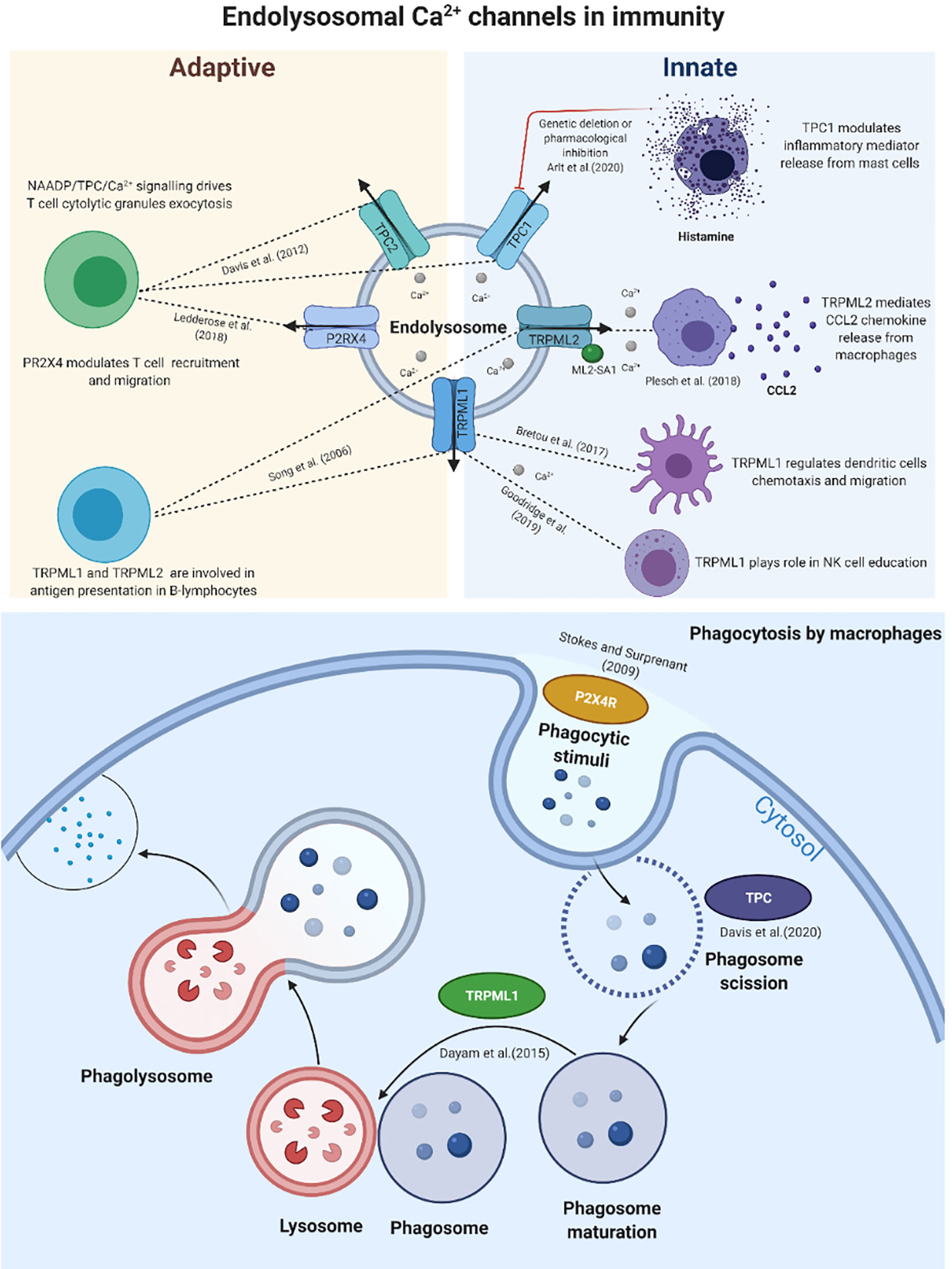
Figure 1 Schematic representation of the main calcium endolysosomal Ca2+ channels involved in immunity. The evolving contributions of TPCs, TRPMLs and P2X4R in innate and adaptive immune responses and their vital roles in various stages of phagocytosis.

Table 1 Some experimental evidence supporting endolysosomal Ca2+ signalling involvement in physiological processes attributed to immunity.
The two-pore channels (TPCs) are present as two isoforms in mammals—TPC1 and TPC2. Debate continues as to whether TPCs are primarily Ca2+ or Na+ channels (16). Data from several studies suggest that TPCs behave differently in different biological contexts. They can trigger Ca2+ or Na+ release upon binding to second messengers: nicotinic acid adenine dinucleotide phosphate (NAADP), acts directly or indirectly to release Ca2+, and phosphatidylinositol 3,5-bisphosphate [PI (3,5)P2] to release Na+ (17–20). Phagocytosis by macrophages is a physiological process initiated by our innate immune system as the first line of defines against both pathogens (bacteria, toxins, viruses) and tumour cells. Lysosomes are multifunctional organelles and play vital roles in phagocytosis, particularly in the late stages of phagosome maturation (21). A recent study by Suresh et al. (22) has uncovered the role of the tubular state of lysosomes in phagocytosis, which is known to modulate processes related to immunity, such as antigen presentation. The study found that lysosome tubular states mediate phagocytosis and enhanced phagosome-lysosome fusion in RAW 264.7 cells (an in vitro model of murine macrophages) (22). Another recent study by Freeman et al. suggested the possibility that TPC2 acts as a regulator of the lysosome tubulation process (23). The study showed that TPC2 overexpression drives lysosome tubulation in a mechanism involving phosphatidylinositol 3,5-bisphosphate activation (23). Additionally, TPC1 and TPC2 expression at the mRNA level was found to be significantly upregulated in bone marrow-derived macrophages compared to mouse embryonic fibroblasts (23). The participation of endolysosomal Ca2+-mediated phagosome-lysosome fusion was implicated in the maturation of the phagosome phase, where the phagosome fused with the lysosome, ultimately becoming a phagolysosome, which is a fundamental step of phagocytosis. Recently, Davis et al. (6) identified a role for NAADP evoked TPC-endolysosomal Ca2+ signalling from the nanodomains involving calcineurin activity and dynamine 2 activation in macrophages at the scission of phagosomes from the plasma membrane stage of phagocytosis for small and large particles (6), which suggests that TPC1 or TPC2 may act as downstream regulators of phagocytosis in macrophages.
Previously, Davis et al. deciphered the biological significance of NAADP/TPC/Ca2+ signalling in T cell biology. They found that NAADP-mediated Ca2+ release is a significant pathway that drives T cell cytolytic granule exocytosis (15). Recently, Elisabeth et al. (2020) reported for the first time that endolysosomal Ca2+ signals via TPC1 mediate the development of the immune response by triggering the release of inflammatory mediators in a mechanism involving Ca2+ cross talk between TPC1-mediated and endoplasmic reticulum (ER) Ca2+ stores (9). In the in vivo TPC1KO murine model, systemic anaphylaxis was exaggerated, manifested by a profound drop in body temperature compared to WT mice (9). The study also found that TPC1 modulation either by genetic deletion or by pharmacological inhibition by trans-Ned-19 augmented mastocyte degranulation and evoked the release of inflammatory mediator (histamine) from mast cells (9), which are tissue-resident cells of the immune system that play a role in inflammatory and allergic reactions. The number and the size of the mastocytes were significantly attenuated in the TPC1-deficient murine model compared to WT controls (9). The cellular mechanisms underlying the regulation of TPC1-mediated endolysosomal Ca2+ signals in the development of inflammatory and allergic reactions warrants further investigation to aid the development of new drugs for the treatment of anaphylaxis and allergic hypersensitivity.
The NAADP/TPC/Ca2+ signalling pathway has been shown to play an important role in virus trafficking. Gunaratne et al. showed that TPC (TPC1 and TPC2) knockdown hampered Middle East Respiratory Syndrome coronavirus (MERS-CoV) infection in human embryonic kidney 293 (HEK293) cells (24). Ca2+ signalling via TPCs (involving TPC1 and TPC2) regulates Ebola virus entry and plays a significant biological role in virus trafficking and preventing the infection (25). Pharmacological inhibition or genetic knockout of TPCs diminished the capacity of the Ebola virus to infect cells in in vitro or in vivo models (25). The candidacy of these channels as druggable targets for future antiviral therapy is supported by the availability of FDA-approved drugs, such as dopamine antagonists (e.g. fluphenazine and pimozide) and selective oestrogen receptor modulators (including raloxifene, clomiphene, and tamoxifen) that inhibit TPC function and hinder Ebola virus-like particle entry into HeLa Kyoto cells in vitro (26). Functional characterisation of fluphenazine and raloxifene revealed that they block TPC2 activity by decreasing the channel opening time (26). The SARS-CoV2 outbreak led to a revisiting of the role of endolysosomal Ca2+ signalling, particularly via TPCs, in virus trafficking and infectivity. Recent evidence has shown that TPC2, phosphatidylinositol 3-phosphate 5-kinase (PIKfyve), and cathepsin L regulate SARS-CoV-2 entry in an in vitro model (HEK 293/hACE2 cells) (27). Similarly, Clementi et al. found that inhibition of TPC2 function via pharmacological means by naringenin or knockdown by siRNA attenuated SARS-CoV2 infection in vitro (28).
Despite the growing evidence linking TPC/Ca2+ signalling to physiological processes attributed to immunity, research in this area is still in its infancy. Further investigations utilizing biomedical (in vitro and in vivo) and clinical models will decode the role of TPC/Ca2+ signalling in immunity and will contribute to advancing our knowledge regarding the roles of this signalling pathway in the pathogenesis of immune system diseases and might lead to the development of therapeutic agents to treat or prevent diseases related to the immune response.
Transient receptor potential cation channels (TRPMLs; mucolipins) are a subfamily of the TRP channel family, and composed of TRPML1, TRPML2 and TRPML3 in mammals; they are localized in the endolysosomal compartments (29). TRPMLs play a significant role in endolysosomal biology, specifically, endolysosomal trafficking that leads to autophagy (30). Notably, the roles of TRPMLs (particularly TRPML1 and TRPML2) in physiological processes related to immune cell functions are evolving (31, 32), underscoring the importance of fully characterizing the biological and clinical functions of these channels in the immune system. Song et al. (33) reported the first evidence of the interconnection between TRPMLs (TRPML1 and TRPML2) and B-cell antigen presentation in vertebrates (33). The TRPML1-mediated Ca2+ signalling pathway has been implicated in phagocytosis (6, 13, 34). TRPML1 acts as a regulator of phagosome maturation, FYVE finger-containing phosphoinositide kinase (PIKfyve), and (PI(3,5)P2)-mediated Ca2+ signals via TRPML1-triggered phagosome-lysosome fusion (13). A previous study implicated (PI(3,5)P2)/TRPML1/Ca2+ signalling as a modulator of phagocytosis by regulating focal exocytosis, which is significant for phagosome biogenesis (34). Recently, knockout of TRPML1 was shown to attenuate the phagocytosis of large particles in murine bone marrow derived macrophages (BMDMs), indicating that lysosomal Ca2+ release via TRPML1 is necessary for large target phagocytosis (6). TRPML1/Ca2+ signalling is involved in lysosome tubulation (35); this process plays roles in phagocytosis and antigen presentation. Recently, Goodridge et al. (36) identified a pivotal role for lysosomal Ca2+ release via TRPML1 as a meditator of the natural killer (NK) cell function (36). Previous studies demonstrated that TRPML1 lysosomal Ca2+ signals were involved in dendritic RNA transportation through the modulation of Toll-like receptor 7 (TLR7) signalling (12). Additionally, the Ca2+ signalling mediated by TRPML1 regulates two important dendritic cell functions involving migration and chemotaxis (37). These findings highlight the evolution of TRPML1 as a modulator of innate and adaptive immune cell functions; thus, it warrants further investigation to reveal the molecular mechanisms of TRPML1 in immunity via in vitro and in vivo models. TRPML2/Ca2+ signalling modulates chemokine (C-C motif) ligand 2 (CCL2; also known as monocyte chemoattractant protein 1 (MCP1)) release from macrophages (8) and acts as a key regulator of monocyte and macrophage infiltrations and migration (10). Consequently, Ca2+ signalling via TRPML2 modulates the inflammation by regulating the release of CCL2; this may serve as a viable therapeutic target for patients with inflammatory diseases. In addition to the involvement of TRPML2 in chemokine release, it was shown that TRPML2-evoked endolysosomal Ca2+ signalling plays a role in viral trafficking (38). To our knowledge, there is a lack of molecular evidence characterising the role of TRPML3 in immunity. Although the evolving body of evidence highlights that endolysosomal Ca2+ signals mediated by TRPMLs play important roles in innate and adaptive immunity, further studies are required to decipher the precise mechanisms underlying the physiological processes of these channels in immunity, from chemokine/cytokine release to antigen presentation.
The P2X4 receptor belongs to the purinergic receptor family, is involved in ATP-evoked Ca2+ release, and is localized to the endolysosomal system (39). P2X4R-mediated Ca2+ signalling is recognized as a key mediator in inflammation and neuropathic pain (40–42). Recent studies have continued to shed light on the roles of P2X4R in physiological processes related to immunity. Upregulation of P2X4R protein expression was observed in the early stages of phagocytosis (with initial phagocytic stimuli) in alveolar macrophages (7), which suggested the involvement of P2X4R in phagocytosis. As discussed earlier concerning the roles of TPCs and TRPMLs in phagocytosis, we speculate that endolysosomal ion channels dynamically communicate at a molecular level to ultimately mediate phagocytosis; additionally, these channels have distinctive roles in different phases of this process. P2X4R mediates allergen-induced airway inflammation through the regulation of priming dendritic cells for T helper 2 (Th2), inducing IL-1ß (Interleukin 1 beta) secretion (43). Similarly, P2X4R was found to modulate the P2X7 receptor-mediated release of two pro-inflammatory cytokines, IL-1ß and IL18 (Interleukin-18), which mediate inflammation in murine bone marrow-derived dendritic cells (BMDCs) (11). P2X4R-deficient mice exhibited protective effects against ischemic acute kidney injury compared to WT mice; this effect was studied at the molecular level and linked to P2X4R-augmented ischemic acute kidney injury via a mechanism involving the activation of NLRP3 (NLR family pyrin domain containing 3) in inflammasome signaling (44). Inflammasome is a multiprotein complex that plays a fundamental role in inflammation of innate immune cells through the activation of caspase 1, which is responsible for cleavage of the precursor forms of two important inflammatory mediators, IL-1ß and IL18, into biologically active cytokines (44). Further research is required to determine the complex interplay between P2RX4, inflammasome, and the release of IL-1ß and IL18. Pharmacological inhibition of P2RX4 via 5-BDBD (5-(3-bromophenyl)-1,3-dihydro-2H-benzofuro[3,2-e]-1,4-diazepin-2-1) hampered T-cell migration (45). T-cell migration is a critical step in T-cell function. Additionally, Ledderose et al. (44) have provided substantial further evidence to confirm this finding. With an in vivo mouse model, they found that pharmacological modulation of P2X4R by 5-BDBD resulted in the rejection of lung transplants by impairing T-cell recruitment in allograft tissue (45). Overexpression of P2X4R was detected in CD4+ T cells from peripheral blood and adipose tissue in obese, healthy subjects, indicating a possible role for P2X4R in chronic inflammation associated with obesity (46).
Overall, these findings underline the immunological significance of P2X4R in innate and adaptive immunity and warrant further investigations to biologically and clinically characterize the multi-functional role of P2X4R in immunity. This may further the development of immunomodulators to treat inflammatory diseases and prevent graft rejection and transplantation complications.
Although there is a growing scientific interest in the role of endolysosomal Ca2+ channels in immunity, there is a paucity of studies that categorize these channels clinically. Recently, a genome-wide association study in the Han Chinese population identified TPCN2 as one of four gene signatures attributed to systemic lupus erythematosus (SLE) susceptibility, which is characterized as a chronic autoimmune disease (14). Significantly overexpressed P2X4 at the protein level was discovered in tissues of patients with hepatitis C virus-induced hepatocellular carcinoma compared to non-hepatitis C virus-induced hepatocellular carcinoma (47). This finding raises a clinical question regarding the possibility of targeting P2X4 to modulate the immune response that contributes to hepatitis C virus-induced hepatocellular carcinoma, which warrants further investigation to understand the role of P2X4 in hepatitis C virus-induced hepatocellular carcinoma pathology.
An evolving body of evidence continues to uncover the function of endolysosomal Ca2+ signalling in innate and adaptive immune cell responses. It has become clear that endolysosomal Ca2+ channels, mainly TPC2 and TRPML-1, serve a critical role in phagocytosis at a global level, with distinctive roles at different stages of the phagocytic process. Importantly, TRPML-2 and P2X4R are implicated in modulating chemokine and cytokine release and consequently their effect on inflammation; however, the precise mechanisms underlying the action of these channels remain elusive and require further investigation to define a specific upstream or downstream target to overcome problems posed by the ubiquity of Ca2+ signals in our cells and to modulate the innate immune response. Recent studies shed light on the roles of TRPML-1 and P2X4 in adaptive immune cell function and raise questions regarding their candidacy as valuable targets for modulation of adaptive immune responses. Regardless of the paucity of clinical evidence, GWAS revealed the potential applications of TPC2 as a biomarker in the definition of SLE susceptibility in the Chinese population and warrants validation in prospective cohorts of a diverse population. Despite the exploratory nature of the evidence highlighting the role of endolysosomal Ca2+ signalling in various processes related to immunity, this mini-review offers some insights into the pivotal roles of these channels in the specific mechanisms of innate and adaptive immunity that lead to inflammation and disorders related to the immune system. Additionally, it raises questions regarding the clinical utility of these channels as biomarkers or immunotherapy targets to modulate innate and adaptive immune responses.
AA has contributed to the mini-review-research question, data collection, and interpretation, and written the manuscript. JP has revised the manuscript. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
AA is a recipient of a Saudi Ministry of Education scholarship. Furthermore, the Saudi Ministry of Education supported AA’s graduate studies. The figure was created with BioRender.com.
1. Rao A, Hogan PG. Calcium Signaling in Cells of the Immune and Hematopoietic Systems. Immunol Rev (2009) 231(1):5–9. doi: 10.1111/j.1600-065X.2009.00823.x
2. Trebak M, Kinet JP. Calcium Signalling in T Cells. Nat Rev Immunol (2019) 19(3):154–69. doi: 10.1038/s41577-018-0110-7
3. Hann J, Bueb JL, Tolle F, Bréchard S. Calcium Signaling and Regulation of Neutrophil Functions: Still a Long Way to Go. J Leukocyte Biol (2020) 107(2):285–97. doi: 10.1002/JLB.3RU0719-241R
4. Park YJ, Yoo SA, Kim M, Kim WU. The Role of Calcium–Calcineurin–NFAT Signaling Pathway in Health and Autoimmune Diseases. Front Immunol (2020) 11:195. doi: 10.3389/fimmu.2020.00195
5. Wajdner HE, Farrington J, Barnard C, Peachell PT, Schnackenberg CG, Marino JP Jr, et al. Orai and TRPC Channel Characterization in Fcϵri-Mediated Calcium Signaling and Mediator Secretion in Human Mast Cells. Physiol Rep (2017) 5(5):e13166. doi: 10.14814/phy2.13166
6. Davis LC, Morgan AJ, Galione A. NAADP-Regulated Two-Pore Channels Drive Phagocytosis Through Endo-Lysosomal Ca2+ Nanodomains, Calcineurin, and Dynamin. EMBO J (2020) 8:e104058. doi: 10.15252/embj.2019104058
7. Stokes L, Surprenant A. Dynamic Regulation of the P2X4 Receptor in Alveolar Macrophages by Phagocytosis and Classical Activation. Eur J Immunol (2009) 39(4):986–95. doi: 10.1002/eji.200838818
8. Plesch E, Chen CC, Butz E, Rosato AS, Krogsaeter EK, Yinan H, et al. Selective Agonist of TRPML2 Reveals Direct Role in Chemokine Release From Innate Immune Cells. Elife (2018) 7:e39720. doi: 10.7554/eLife.39720
9. Arlt E, Fraticelli M, Tsvilovskyy V, Nadolni W, Breit A, O’Neill TJ, et al. TPC1 Deficiency or Blockade Augments Systemic Anaphylaxis and Mast Cell Activity. Proc Natl Acad Sci (2020) 117(30):18068–78. doi: 10.1073/pnas.1920122117
10. Deshmane SL, Kremlev S, Amini S, Sawaya BE. Monocyte Chemoattractant Protein-1 (MCP-1): An Overview. J Interferon Cytokine Res (2009) 29(6):313–26. doi: 10.1089/jir.2008.0027
11. Sakaki H, Fujiwaki T, Tsukimoto M, Kawano A, Harada H, Kojima S. P2X4 Receptor Regulates P2X7 Receptor-Dependent IL-1β and IL-18 Release in Mouse Bone Marrow-Derived Dendritic Cells. Biochem Biophys Res Commun (2013) 432(3):406–11. doi: 10.1016/j.bbrc.2013.01.135
12. Li X, Saitoh SI, Shibata T, Tanimura N, Fukui R, Miyake K. Mucolipin 1 Positively Regulates TLR7 Responses in Dendritic Cells by Facilitating RNA Transportation to Lysosomes. Int Immunol (2015) 27(2):83–94. doi: 10.1093/intimm/dxu086
13. Dayam RM, Saric A, Shilliday RE, Botelho RJ. The Phosphoinositide-Gated Lysosomal Ca2+ Channel, TRPML1, is Required for Phagosome Maturation. Traffic (2015) 16(9):1010–26. doi: 10.1111/tra.12303
14. Wen L, Zhu C, Zhu Z, Yang C, Zheng X, Liu L, et al. Exome-Wide Association Study Identifies Four Novel Loci for Systemic Lupus Erythematosus in Han Chinese Population. Ann Rheum Dis (2018) 77(3):417.
15. Davis LC, Morgan AJ, Chen JL, Snead CM, Bloor-Young D, Shenderov E, et al. NAADP Activates Two-Pore Channels on T Cell Cytolytic Granules to Stimulate Exocytosis and Killing. Curr Biol (2012) 22(24):2331–7. doi: 10.1016/j.cub.2012.10.035
16. Morgan AJ, Galione A. Two-Pore Channels (TPC s): Current Controversies. Bioessays (2014) 36(2):173–83. doi: 10.1002/bies.201300118
17. Wang X, Zhang X, Dong XP, Samie M, Li X, Cheng X, et al. TPC Proteins are Phosphoinositide-Activated Sodium-Selective Ion Channels in Endosomes and Lysosomes. Cell (2012) 151:372–83:2. doi: 10.1016/j.cell.2012.08.036
18. Brailoiu E, Churamani D, Cai X, Schrlau MG, Brailoiu GC, Gao X, et al. Essential Requirement for Two-Pore Channel 1 in NAADP-mediated Calcium Signaling. J Cell Biol (2009) 186:201–9:2. doi: 10.1083/jcb.200904073
19. Zong X, Schieder M, Cuny H, Fenske S, Gruner C, Rötzer K, et al. The Two-Pore Channel TPCN2 Mediates NAADP-dependent Ca 2+-Release From Lysosomal Stores. Pflügers Archiv-Eur J Physiol (2009) 458.5:891–9. doi: 10.1007/s00424-009-0690-y
20. Calcraft PJ, Ruas M, Pan Z, Cheng X, Arredouani A, Hao X, et al. NAADP Mobilizes Calcium From Acidic Organelles Through Two-Pore Channels. Nature (2009) 459.7246:596–600. doi: 10.1038/nature08030
21. Westman J, Freeman SA, Grinstein S. Unconventional Role of Lysosomes in Phagocytosis. Cell Calcium (2020) 91:102269. doi: 10.1016/j.ceca.2020.102269
22. Suresh B, Saminathan A, Chakraborty K, Cui C, Becker L, Krishnan Y. Tubular Lysosomes Harbor Active Ion Gradients and Poise Macrophages for Phagocytosis. bioRxiv (2020). doi: 10.1101/2020.12.05.413229
23. Freeman SA, Uderhardt S, Saric A, Collins RF, Buckley CM, Mylvaganam S, et al. Lipid-Gated Monovalent Ion Fluxes Regulate Endocytic Traffic and Support Immune Surveillance. Science (2020) 367(6475):301–5. doi: 10.1126/science.aaw9544
24. Gunaratne GS, Yang Y, Li F, Walseth TF, Marchant JS. NAADP-Dependent Ca2+ Signaling Regulates Middle East Respiratory Syndrome-Coronavirus Pseudovirus Translocation Through the Endolysosomal System. Cell Calcium (2018) 75:30–41. doi: 10.1016/j.ceca.2018.08.003
25. Sakurai Y, Kolokoltsov AA, Chen CC, Tidwell MW, Bauta WE, Klugbauer N, et al. Two-Pore Channels Control Ebola Virus Host Cell Entry and are Drug Targets for Disease Treatment. Science (2015) 347(6225):995–8. doi: 10.1126/science.1258758
26. Penny CJ, Vassileva K, Jha A, Yuan Y, Chee X, Yates E, et al. Mining of Ebola Virus Entry Inhibitors Identifies Approved Drugs as Two-Pore Channel Pore Blockers. Biochim Et Biophys Acta (BBA)-Mol Cell Res (2019) 1866(7):1151–61. doi: 10.1016/j.bbamcr.2018.10.022
27. Ou X, Liu Y, Lei X, Li P, Mi D, Ren L, et al. Characterization of Spike Glycoprotein of SARS-CoV-2 on Virus Entry and its Immune Cross-Reactivity With SARS-Cov. Nat Commun (2020) 11(1):1–2. doi: 10.1038/s41467-020-15562-9
28. Clementi N, Scagnolari C, D’Amore A, Palombi F, Criscuolo E, Frasca F, et al. Naringenin is a Powerful Inhibitor of SARS-CoV-2 Infection In Vitro. Pharmacol Res (2020). doi: 10.1016/j.phrs.2020.105255
29. Rosato AS, Tang R, Grimm C. Two-Pore and TRPML Cation Channels: Regulators of Phagocytosis, Autophagy and Lysosomal Exocytosis. Pharmacol Ther (2020) 220:107713. doi: 10.1016/j.pharmthera.2020.107713
30. Venkatachalam K, Wong CO, Zhu MX. The Role of TRPMLs in Endolysosomal Trafficking and Function. Cell Calcium (2015) 58(1):48–56. doi: 10.1016/j.ceca.2014.10.008
31. Spix B, Chao YK, Abrahamian C, Chen CC, Grimm C. TRPML Cation Channels in Inflammation and Immunity. Front Immunol (2020) 11:225. doi: 10.3389/fimmu.2020.00225
32. Santoni G, Morelli MB, Amantini C, Santoni M, Nabissi M, Marinelli O, et al. “Immuno-Transient Receptor Potential Ion Channels”: The Role in Monocyte-and Macrophage-Mediated Inflammatory Responses. Front Immunol (2018) 9:1273. doi: 10.3389/fimmu.2018.01273
33. Song Y, Dayalu R, Matthews SA, Scharenberg AM. TRPML Cation Channels Regulate the Specialized Lysosomal Compartment of Vertebrate B-Lymphocytes. Eur J Cell Biol (2006) 85(12):1253–64. doi: 10.1016/j.ejcb.2006.08.004
34. Samie M, Wang X, Zhang X, Goschka A, Li X, Cheng X, et al. A TRP Channel in the Lysosome Regulates Large Particle Phagocytosis Via Focal Exocytosis. Dev Cell (2013) 26(5):511–24. doi: 10.1016/j.devcel.2013.08.003
35. Li X, Rydzewski N, Hider A, Zhang X, Yang J, Wang W, et al. A Molecular Mechanism to Regulate Lysosome Motility for Lysosome Positioning and Tubulation. Nat Cell Biol (2016) 18(4):404–17. doi: 10.1038/ncb3324
36. Goodridge JP, Jacobs B, Saetersmoen ML, Clement D, Hammer Q, Clancy T, et al. Remodeling of Secretory Lysosomes During Education Tunes Functional Potential in NK Cells. Nat Commun (2019) 10(1):1–5. doi: 10.1038/s41467-019-08384-x
37. Bretou M, Sáez PJ, Sanséau D, Maurin M, Lankar D, Chabaud M, et al. Lysosome Signaling Controls the Migration of Dendritic Cells. Sci Immunol (2017) 2(16). doi: 10.1126/sciimmunol.aak9573
38. Rinkenberger N, Schoggins JW. Mucolipin-2 Cation Channel Increases Trafficking Efficiency of Endocytosed Viruses. MBio (2018) 9(1). doi: 10.1128/mBio.02314-17
39. Murrell-Lagnado RD, Frick M. P2X4, and Lysosome Fusion. Curr Opin Pharmacol (2019) 47:126–32. doi: 10.1016/j.coph.2019.03.002
40. Suurväli J, Boudinot P, Kanellopoulos J, Boudinot SR. P2x4: A Fast and Sensitive Purinergic Receptor. Biomed J (2017) 40(5):245–56. doi: 10.1016/j.bj.2017.06.010
41. Trang T, Salter MW. P2X4 Purinoceptor Signaling in Chronic Pain. Purinerg Signaling (2012) 8(3):621–8. doi: 10.1007/s11302-012-9306-7
42. Kang L, Yayi H, Fang Z, Bo Z, Zhongyuan X. Dexmedetomidine attenuates P2X4 and NLRP3 expression in the spine of rats with diabetic neuropathic pain. Acta Cirurg Bras (2019) 34(11). doi: 10.1590/s0102-865020190110000005
43. Zech A, Wiesler B, Ayata CK, Schlaich T, Dürk T, Hoßfeld M, et al. P2rx4 Deficiency in Mice Alleviates Allergen-Induced Airway Inflammation. Oncotarget (2016) 7(49):80288. doi: 10.18632/oncotarget.13375
44. Han SJ, Lovaszi M, Kim M, D’Agati V, Haskó G, Lee HT. P2X4 Receptor Exacerbates Ischemic AKI and Induces Renal Proximal Tubular NLRP3 Inflammasome Signaling. FASEB J (2020) 34(4):5465–82. doi: 10.1096/fj.201903287R
45. Ledderose C, Liu K, Kondo Y, Slubowski CJ, Dertnig T, Denicoló S, et al. Purinergic P2X4 Receptors and Mitochondrial ATP Production Regulate T Cell Migration. J Clin Invest (2018) 128(8):3583–94. doi: 10.1172/JCI120972
46. Ruiz-Rodríguez VM, Cortes-García JD, de Jesús Briones-Espinoza M, Rodríguez-Varela E, Vega-Cárdenas M, Gómez-Otero A, et al. P2X4 Receptor as a Modulator in the Function of P2X Receptor in CD4+ T Cells From Peripheral Blood and Adipose Tissue. Mol Immunol (2019) 112:369–77. doi: 10.1016/j.molimm.2019.06.009
Keywords: endolysosomal, calcium, Ca2+ signals, immunity, innate response, adaptive response
Citation: Alharbi AF and Parrington J (2021) Deciphering the Role of Endolysosomal Ca2+ Channels in Immunity. Front. Immunol. 12:656965. doi: 10.3389/fimmu.2021.656965
Received: 21 January 2021; Accepted: 09 April 2021;
Published: 27 April 2021.
Edited by:
Jagadeesh Bayry, Institut National de la Santé et de la Recherche Médicale (INSERM), FranceReviewed by:
Francesco Moccia, University of Pavia, ItalyCopyright © 2021 Alharbi and Parrington. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Abeer F. Alharbi, YWJlZXIuYWxoYXJiaUBwaGFybS5veC5hYy51aw==; John Parrington, am9obi5wYXJyaW5ndG9uQHdvcmMub3guYWMudWs=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.