 Andrew J. Sawyer
Andrew J. Sawyer Mathieu Garand
Mathieu Garand Damien Chaussabel
Damien Chaussabel Carl G. Feng
Carl G. Feng- 1Immunology and Host Defense Group, Discipline of Infectious Diseases and Immunology, Faculty of Medicine and Health, The University of Sydney, Sydney, NSW, Australia
- 2Tuberculosis Research Program, Centenary Institute, The University of Sydney, Sydney, NSW, Australia
- 3Immunology Program, Sidra Medicine, Doha, Qatar
Cystatin F encoded by CST7 is a cysteine peptidase inhibitor known to be expressed in natural killer (NK) and CD8+ T cells during steady-state conditions. However, little is known about its expression during inflammatory disease states in humans. We have developed an analytic approach capable of not only identifying previously poorly characterized disease-associated genes but also defining regulatory mechanisms controlling their expression. By exploring multiple cohorts of public transcriptome data comprising 43 individual datasets, we showed that CST7 is upregulated in the blood during a diverse set of infectious and non-infectious inflammatory conditions. Interestingly, this upregulation of CST7 was neutrophil-specific, as its expression was unchanged in NK and CD8+ T cells during sepsis. Further analysis demonstrated that known microbial products or cytokines commonly associated with inflammation failed to increase CST7 expression, suggesting that its expression in neutrophils is induced by an endogenous serum factor commonly present in human inflammatory conditions. Overall, through the identification of CST7 upregulation as a marker of acute inflammation in humans, our study demonstrates the value of publicly available transcriptome data in knowledge generation and potential biomarker discovery.
Introduction
Transcriptomic profiling has been widely used over the past two decades to uncover the expression and function of genes across the genome. Each transcriptomic profiling study generates enormous amounts of data, the bulk of which has been published in online repositories and is available for download free of charge. The NCBI Gene Expression Omnibus (GEO), for example, is an online database containing more than 58,000 series datasets from human studies, comprising 2 million individual samples. These include datasets from across the spectrum of human diseases, tissue types and cell types. Transcriptomic data generated from cells following stimulation or from gene-targeted cells provide additional insights into gene regulation. While transcriptomic datasets have typically been interrogated by the original researchers to identify pathways/genes of interest, most of the data remains incompletely explored and utilized. This wealth of data remains available online and represents a valuable resource for life scientists exploring a gene or pathway of interest. For example, investigations can be used to generate informed hypotheses that guide future studies.
Of particular interest are genes that are found to be differentially expressed despite no prior association with the conditions under investigation. In this case, public transcriptomic datasets can be used to validate findings, facilitate further clarification of the specificity of gene expression across cell types and diseases, and even provide insights into gene regulation. Some studies have already successfully identified a gene of interest across a small cohort of GEO datasets and been used to formulate a hypothesis for gene function (1, 2).
Here, we demonstrate an analytic pipeline that allows identification of differentially expressed genes that are yet to be characterized, then define mechanisms regulating their expression across a cohort of 43 publicly available transcriptomic datasets. We discovered that the expression of CST7 in whole blood was markedly upregulated in sepsis as well as across a range of bacterial, viral, and sterile inflammatory conditions. Cystatin F, encoded by CST7 is a cathepsin C-directed cysteine protease known to regulate cytotoxicity in natural killer (NK) cells (3, 4). While cathepsin C is involved in serine peptidase activation in NK cells and neutrophils, the significance and regulation of cystatin F expression in neutrophils remains unexplored. Our analysis revealed that this CST7 upregulation is neutrophil-specific and regulated by host factors. In this study, we demonstrate that transcriptomic profiling using publicly available datasets is a robust approach to identify associations of unknown genes with human diseases.
Methods
Literature Search
PubMed was searched to retrieve the literature associated with each gene using a query that consisted of a concatenation of its official name, symbol and aliases retrieved from NCBI gene. Using CST7 as an example, the following query was applied: CST7 [tw] OR ‘cystatin F’ [tw] OR Leukocystatin [tw]. The text word [tw] function was used to restrict the search to the title and abstract.
Single-Cohort Transcriptomic Analysis
All datasets used in this study were collected from the NCBI GEO (Supplementary Table 1). All single-cohort analysis was performed in R and datasets were downloaded using the GEOquerey (5) package. Normalized expression counts of genes of interest were exported for plotting. Where necessary, datasets were log-transformed and DEGs were identified using the Limma package (6) and P values were adjusted with a Benjamini Hochberg false discovery rate of 0.01.
Multicohort Transcriptomic Analysis
All Multicohort analyses were performed in R using the Metaintegrator package (7). Datasets were accessed from GEO using the “getGEOData” function and samples were separated into control and non-control groups using the “classFunction” function. Meta-analysis was performed using the “RunMetaAnalysis” function and forest plots were previewed with the “forestPlot” function. A summary effect size for genes was determined based on the standardized mean difference in each cohort. Effect sizes for DEGs between septic patients and controls across the nine whole blood sepsis cohorts were identified using the “filterGenes” function with an effect size threshold of 1 and false discovery rate threshold of 0.05. Heatmaps of sepsis DEG expression across other disease states were generated using the heatmapPlot function with “diplayPooled = FALSE” to maintain the original gene order.
Software
Transcriptomic analysis was performed in R 4.0.2. For single-cohort group comparisons, t-tests were performed using GraphPad Prism 7.02. All plots were generated using GraphPad Prism 7.02. Figures were prepared using Microsoft PowerPoint 2019.
Results
Cystatin F Is Upregulated in Neutrophils Following Stimulation With Septic Plasma and in Circulating Leukocytes of Sepsis Patients
We have shown previously that transcriptomic changes in public datasets can be explored for biomarker discovery by identifying previously uncharacterized genes (1, 2). Here we demonstrate a two-staged approach of literature-searches and public-data mining, which enables not only to identify a novel gene of interest but also to define regulatory mechanisms controlling its expression (Figure 1A). To identify an initial cohort of genes upregulated during inflammation, a dataset deposited in the NCBI GEO database (GSE49757) was chosen. Neutrophils from healthy donors were stimulated with plasma from either healthy controls or bacterial culture-confirmed sepsis patients and mRNA expression was measured using microarray (Figure 1B). Genes were sorted according to the fold change in expression and the top 250 genes were selected for further analysis (Figure 1C).
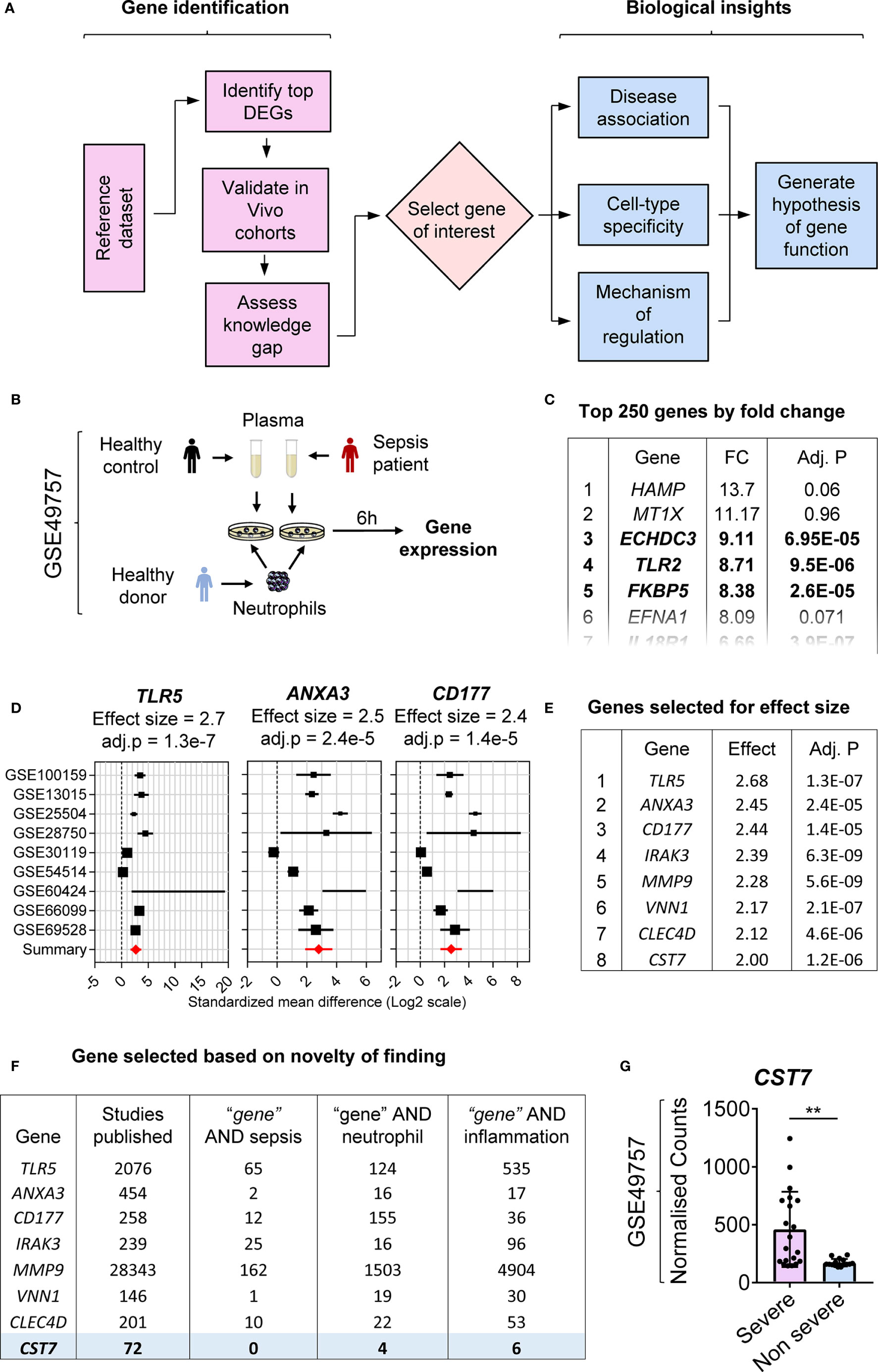
Figure 1 Identification of CST7 as a previously unknown neutrophil-specific, sepsis-associated gene. (A) Flow chart of the process employed to identify a gene of interest and assess its regulation. (B) Schematic overview of the experimental design for the reference dataset (GSE49757) used in this study. Purified neutrophils from healthy adult donors were stimulated with plasma from either septic or healthy individuals for 6 h and gene expression was measured by microarray. (C) Genes were sorted according to fold change in expression and the top 250 genes with a significant change in expression were selected for further assessment. The level of statistical significance was determined using Student’s t-test and P < 0.05 was considered to indicate statistical significance. (D) Gene expression profiles across nine transcriptomic datasets comparing whole blood samples from patients with sepsis to healthy controls are represented in forest plots. Cohort identifiers are shown on the Y-axis and closed black squares represent the standardized mean difference. Square size is proportional to the sample size and whiskers represent a 95% confidence interval. The summary (closed red diamond) represents the overall change in gene expression across cohorts and is also shown as the effect size above each plot along with its corresponding adj. P-value. (E) The top 250 genes identified in panel B were sorted according to overall effect size across sepsis cohorts. (F) For each gene, PubMed was searched to identify both the total number of articles and the number of articles linking the gene with each of the terms “sepsis”, “neutrophil” and “inflammation” in the title and abstract. Genes were searched in order from the highest effect size until a gene with adequate novelty (defined by the number of publications) was identified. (G) CST7 expression was compared between patients with severe sepsis vs. non-severe sepsis in the reference dataset (GSE49757). Each symbol represents an individual donor, while bars represent the mean ± standard deviation. The level of statistical significance was determined using Student’s t-test and P < 0.05 was considered to indicate statistical significance. **P < 0.01.
To extend the in vitro observations and identify potential candidate genes for further study, validation was performed in publicly available datasets generated using whole blood samples from healthy controls and sepsis patients. Nine datasets were selected from the NCBI GEO and multicohort analysis was performed to generate a mean fold change in expression across all cohorts. The top 250 genes were ranked according to fold change in expression in this cohort (Figure 1D). The highest ranked genes in terms of the effect size were considered as the strongest candidates for further investigation (Figure 1E). The final step in the process of selecting a gene of interest was to assess the current knowledge on each identified gene. PubMed was searched to identify the total number of articles relating to each gene and alternate names for each gene were identified through NCBI Gene and added to the search. Genes for which a very small number of articles were identified (<10) were excluded as it was considered unlikely that sufficient information about these gene would be available to inform future studies. Additional searches were carried out for each gene paired with the search terms “sepsis”, “neutrophil” and “inflammation” to identify previous gene associations. The number of articles was counted, and a score applied to each gene based on the number of related articles. Genes were examined in order starting from the gene with the highest fold change in expression and those with known associations to sepsis or neutrophils were excluded (Figure 1F). CST7 (Cystatin F) was ranked 8th in terms of fold change in expression and 72 articles were identified that referred to the gene. Among these, six publications were related to inflammation; however, none of these articles contained the term “sepsis” or linked cystatin F directly to the term neutrophil. CST7 was therefore deemed the most eligible for further investigation. We further confirmed that CST7 expression was significantly increased in neutrophils following stimulation of plasma from patients with severe sepsis compared with those with non-severe sepsis (Figure 1G).
The Significance and Regulation of Cystatin F Induction Are Poorly Defined
To inform further exploration of public data, a profile of the current knowledge of CST7 was constructed. The 72 articles that referred to CST7 were examined for major research themes and tallied (Figure 2A). The cystatins consist of three families of proteins that bind reversibly with high-affinity to inhibit multiple cathepsin family members (8) Cystatin F is unique among the cystatins in that its expression is limited to leukocytes (9) and because it is secreted in an inactive form that can be internalized and subsequently activated in target cells (10, 11). In the context of malignancy, increased levels of CST7 have been correlated with a poorer prognosis in liver metastasis following colorectal cancer (12) and with improved survival in the context of pancreatic ductal adenocarcinoma (13) and hepatocellular carinoma (14). Additionally, in the context of neuroinflammation, CST7 is upregulated during CNS demyelination and is associated with areas of remyelination (15, 16). While CST7 has not been studied in the context of other inflammatory conditions, it has been identified as playing a role in granulocyte inflammation, specifically as an inhibitor of cathepsin C (17).
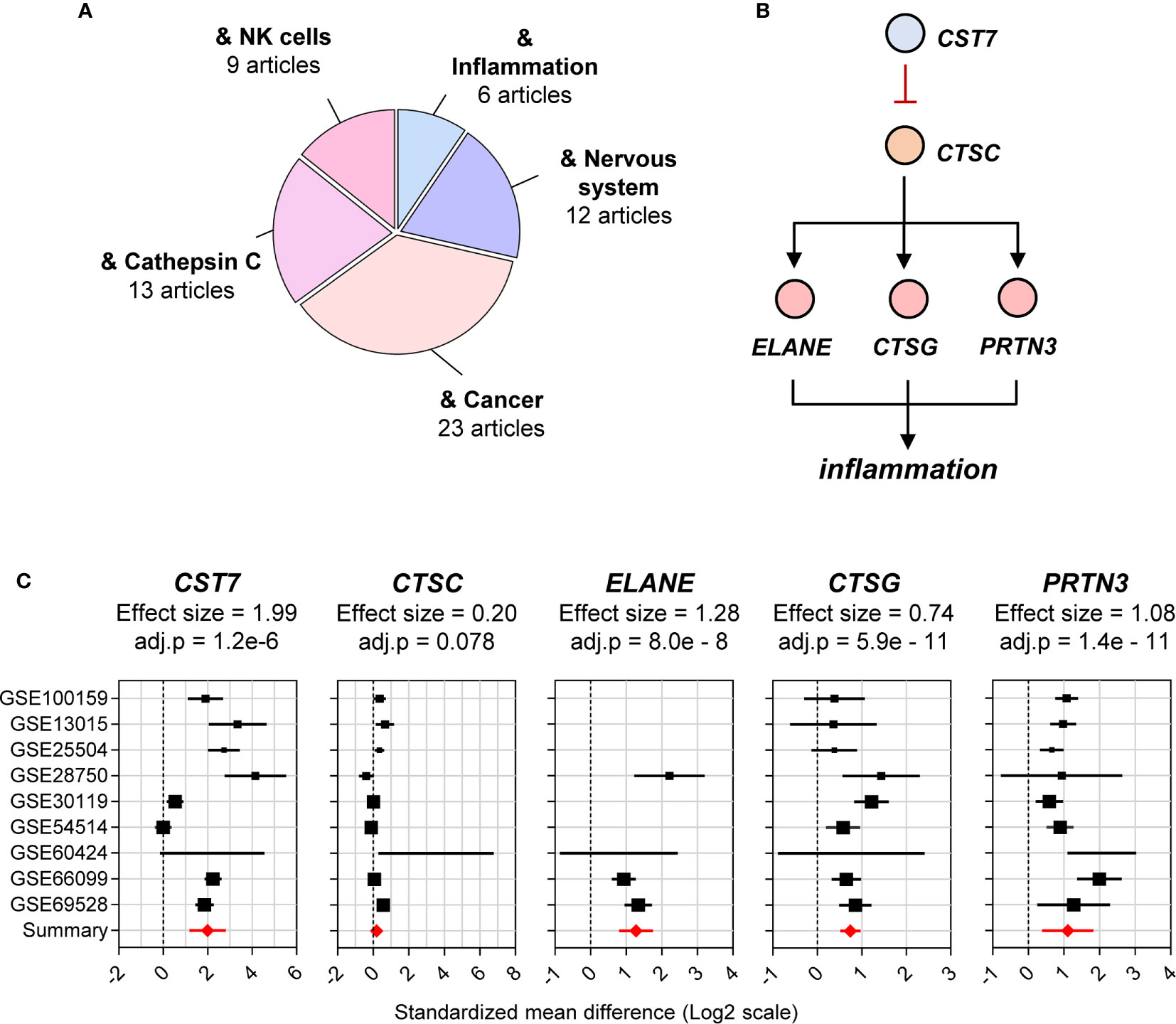
Figure 2 Cystatin F is identified as a cathepsin C-directed serine protease inhibitor. (A) The total number of articles on CST7 (72 articles) was curated and major reported biological functions were summarized. The size of each segment in the pie chart corresponds to the number of articles matching CST7 in relation to the indicated area. (B) The proposed function of the CST7 protein indicating its ability to inhibit CTSC protein. CTSC, which is known to activate the neutrophil serine proteases ELANE, PRTN3 and CTSG, can increase inflammation. (C) Forest plots of CST7, CTSC, ELANE, PRTN3 and CTSG gene expression across sepsis cohorts. The plots were constructed as described in Figure 1D.
Cathepsins consist of a large family of cysteine proteases and play a diverse set of roles in both physiological and pathophysiological conditions. In the immune system, they are involved in antigen processing and presentation, activation of pattern recognition receptors and as cytotoxic devices utilized by both the host and pathogens (18). Cathepsin C is a component of the granules in cytotoxic leukocytes and functions as a key activator of other inflammatory granules in these cells (17). Cystatin F can inhibit cathepsin C in NK cells, thereby inducing an impaired cytotoxic state known as split anergy (3, 19). High levels of cystatin F in CD8+ T cells are also correlated with decreased cytotoxicity (20).
In neutrophils, cathepsin C is required for activation of neutrophil serine proteases including Elastase, neutrophil expressed (ELANE), Cathepsin G (CTSG) and Proteinase 3 (PRTN3) in the inflammatory granules (17, 21, 22). However, whether cystatin F inhibits cathepsin C in neutrophils or exerts anti-inflammatory effects in these cells remains unknown (Figure 2B). Due to their potential connection with CST7, we also examined CTSC, ELANE, CTSG and PRTN3 in the public data cohorts in the remainder of this study. Based on the previous cohort of sepsis datasets, CST7 was significantly upregulated (effect size = 1.99) in patients with sepsis compared with healthy controls, while there was no significant difference in CTSC expression between the two groups. ELANE, CTSG and PRTN3 were significantly upregulated (effect size = 1.28, 0.74 and 1.08, respectively) (Figure 2C).
Cystatin F Is Consistently Upregulated Across Bacterial, Viral, and Sterile Systemic Inflammation
Having identified CST7 as an upregulated gene in whole blood during sepsis, we sought to determine whether the change was restricted to sepsis. We selected six inflammatory diseases for further examination, each of which was represented by at least two cohorts on the GEO. The diseases were categorized as bacterial, viral or sterile inflammation. All the selected cohorts compared whole blood samples from patients and healthy controls. The 2,140 genes that were found to be differentially expressed in the sepsis cohort were measured across each of the other disease cohorts. Heat maps of the similarity between the gene expression profile of each disease state and sepsis are presented in Figure 3.
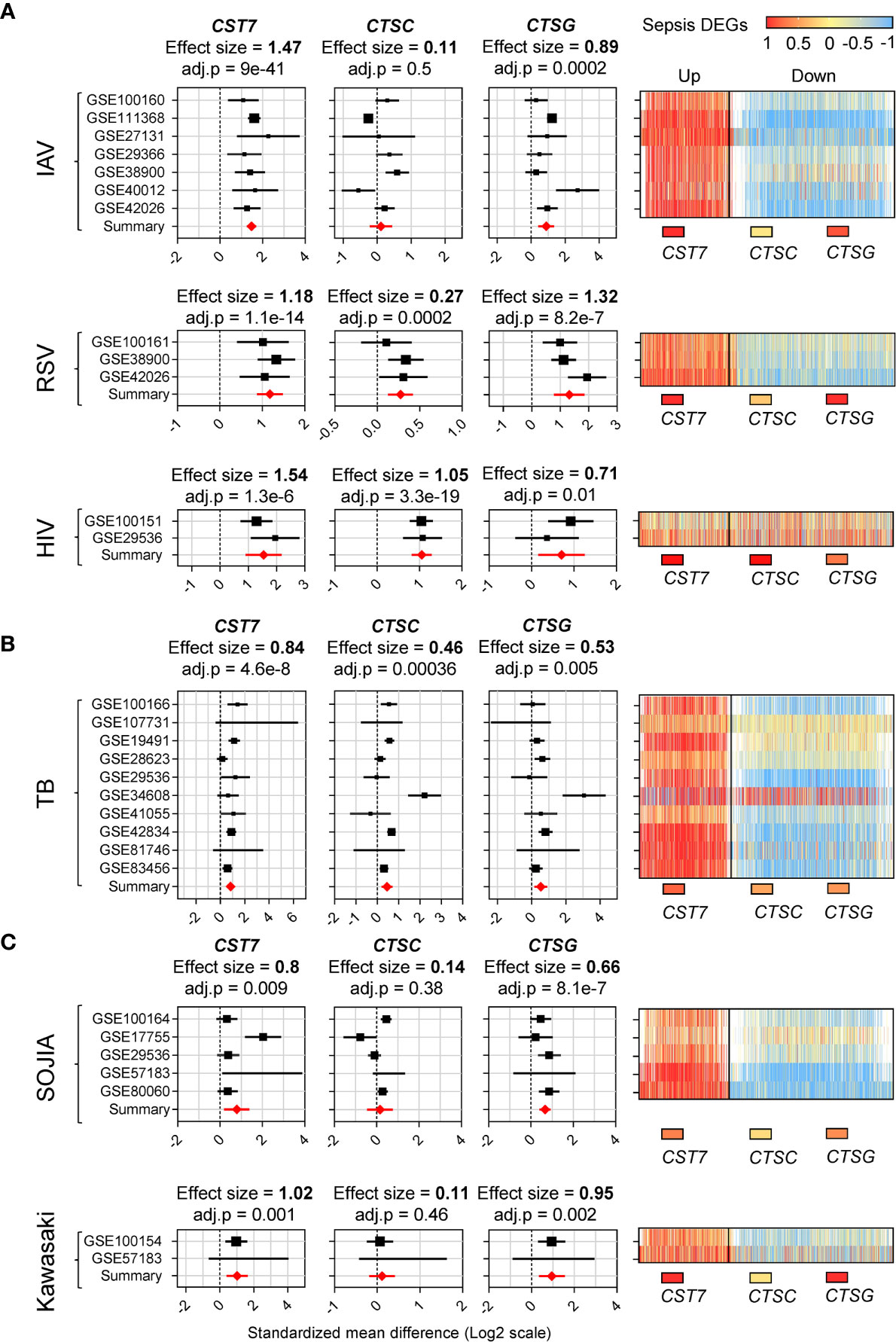
Figure 3 CST7 is upregulated across a spectrum of acute inflammatory diseases. (A) Gene expression profiles for CST7, CTSC, and CTSG across 10 transcriptomic datasets comparing whole blood samples from patients with TB to healthy controls are represented in forest plots constructed as described in Figure 1D. An accompanying heatmap represents the expression of the 2,140 DEGs across the sepsis cohorts (X-axis, separated into up/downregulated expression during sepsis) in each TB cohort (Y-axis). This was repeated in (B) with transcriptomic datasets comparing in whole blood samples from IAV patients to healthy controls, RSV patients to healthy controls and HIV patients to healthy controls. This was also repeated in (C) with transcriptomic datasets comparing whole blood samples from SOJIA patients to healthy controls and Kawasaki disease patients to healthy controls.
CST7 expression was significantly increased in all conditions, with the highest effect size during viral-induced inflammation (Figure 3A). CST7 was increased during infection with influenza A virus (IAV), respiratory syntactical virus (RSV) and human immunodeficiency virus (HIV) infection (effect size = 1.47, 1.18, 1.54 respectively). Interestingly, CTSC expression did not change during IAV infection, but was significantly increased during RSV and HIV infection. PRTN3, ELANE and CSTG were significantly increased in all three viral infection cohorts. In tuberculosis (TB), CST7 was significantly upregulated (effect size = 0.84). In contrast, there were no changes in CTSC or CTSG expression, although expression of PRTN3 and ELANE were increased (Figure 3B). Finally, in the case of sterile inflammation, CST7 expression was increased in both systemic onset idiopathic juvenile arthritis (SOJIA) and Kawasaki disease (effect size = 0.6 and 1.02 respectively). CTSC was not changed in either cohort, while CTSG and PRTN3 were significantly upregulated in both cohorts and ELANE was significantly upregulated during SOJIA (Figure 3C).
Cystatin F Upregulation Is Neutrophil-Specific
Having established that CST7 is upregulated in bulk leukocyte preparations during inflammation, we next determined whether this change is restricted to certain leukocyte populations in the blood. Given the initial finding that septic plasma induced upregulation of CST7 in neutrophils, we identified datasets comparing purified neutrophils from healthy controls and sepsis patients. Two datasets were identified and CST7 was found to be significantly upregulated in neutrophils from sepsis patients compared to those from healthy controls (effect size = 2.3). However, there were no changes in the expression levels of CTSC, CTSG, PRTN3 and ELANE in neutrophils during sepsis compared to those from healthy controls (Figure 4A).
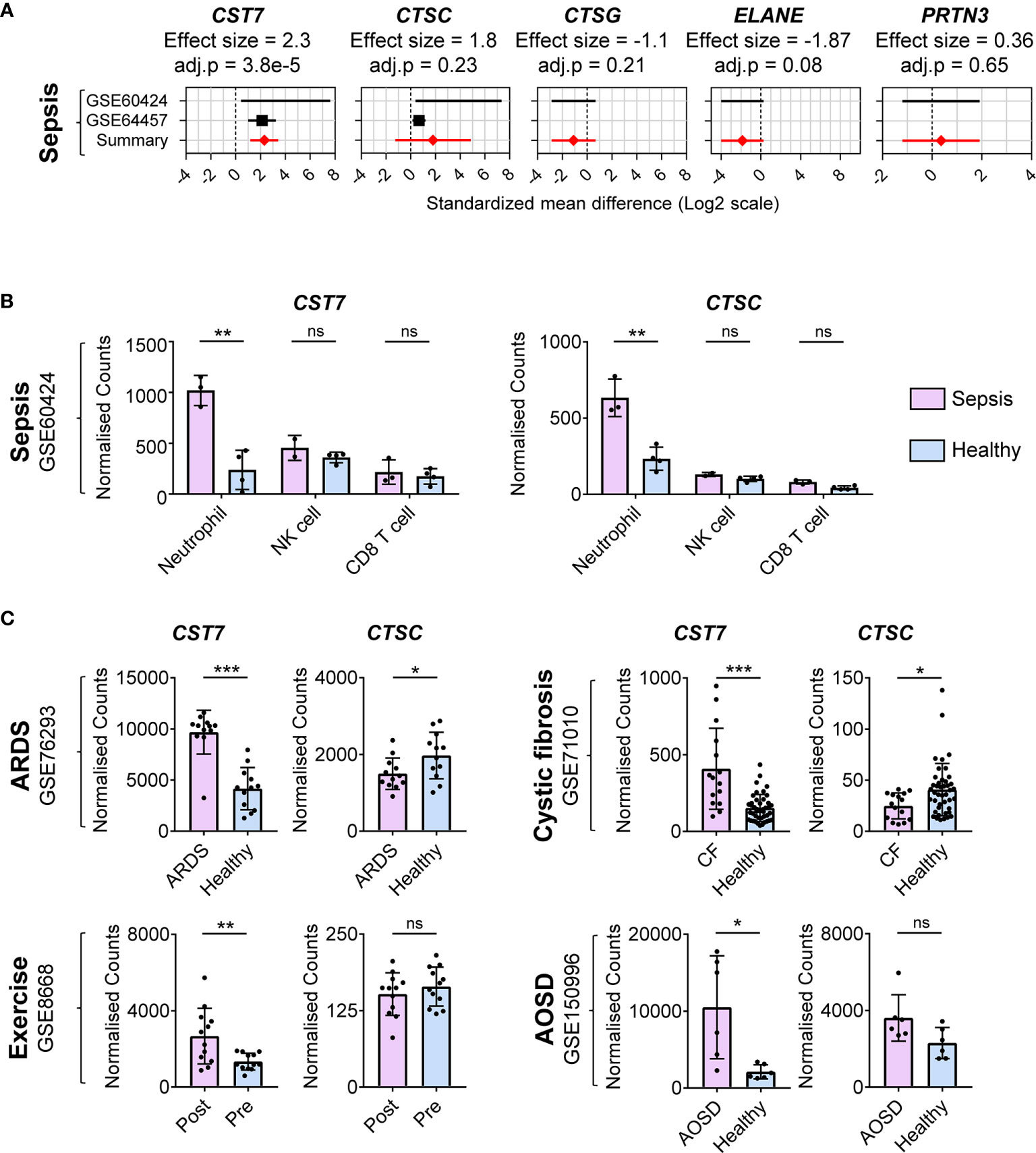
Figure 4 CST7 upregulation during inflammation is neutrophil-specific. (A) Gene expression profiles for CST7, CTSC, CTSG, ELANE, and PRTN3 across two datasets comparing purified neutrophil populations from patients with sepsis and healthy controls are represented in forest plots constructed as described in Figure 1D. (B) Normalized expression counts of CST7 and CTSC in purified neutrophils, NK cells and CD8+ T from healthy controls compared to those from patients with sepsis. (C) Expression of CST7 and CTSC in purified neutrophils comparing patients with ARDS (GSE76293), CF (GSE71010) and SOJIA (GSE103170) to healthy controls as well as comparing healthy individuals pre- and post-exercise (GSE8668). In panel B and C each symbol represents an individual donor, while bars represent the mean ± standard deviation. The level of statistical significance is indicated on the plot: *P < 0.05 **P < 0.01 ***P < 0.001, ns, not significant.
We next examined whether expression of CST7 is modulated in other leukocyte populations during sepsis using one cohort from the previous analysis that also included purified CD8+ T cells, NK cells, CD4+ T cell and monocytes as well as neutrophils. While NK cells expressed the highest CST7 levels under healthy conditions, there was a significant increase in CST7 expression in neutrophils during sepsis and these cells became the predominant contributor of CST7 expression (Figure 4B). There was no change in CST7 expression in NK cells or CD8+ T cells during sepsis. CST7 was expressed lowly in in CD4+ T cells and monocytes and its expression in these cells did not change during sepsis (Data not shown). Given this neutrophil-specific increase during sepsis, we next sought to determine whether CST7 levels were increased in neutrophils under other inflammatory conditions. We examined four cohorts comparing gene expression from purified neutrophils during acute respiratory distress syndrome (ARDS), adult-onset Still’s disease (AOSD) and cystic fibrosis (CF) compared to purified neutrophils from healthy controls as well pre- and post-exercise. In all these conditions, CST7 expression was significantly increased, suggesting neutrophil-specific upregulation of CST7 is positively associated with inflammatory conditions (Figure 4C).
Cystatin F Expression During Inflammation Is Regulated by Host Factor(s)
Having identified upregulation of CST7 during inflammation, we next examined whether public transcriptomic data can be used to shed light on the regulatory mechanisms underpinning this increase in expression. Given that CST7 is increased under a diverse range of inflammatory conditions, we hypothesized that a broadly acting inflammatory mediator may regulate CST7 expression changes. Indeed, examination of whole blood transcriptomic data from one study showed that defective Toll-like receptor (TLR) and IL-1 signaling due to MYD88 or IRAK deficiency is associated with decreased levels of CST7 under baseline conditions (Figure 5A), suggesting a role for an innate signaling pathway in CST7 upregulation. Therefore, we analyzed another dataset in which whole blood cells were stimulated with a range of TLR ligands, heat killed pathogens and major pro-inflammatory cytokines. However, none of these factors stimulated any significant change in CST7 expression (Figure 5B). Finally, we analyzed a dataset in which neutrophil gene expression was compared following endotoxin exposure in vivo and in vitro. We found that when neutrophils were purified from the blood of healthy donors and exposed to endotoxin in vitro, no change in CST7 expression was observed (Figure 5C), confirming that microbial products do not contribute to CST7 induction directly. However, when the donors were exposed to endotoxin by bronchoscopic instillation in vivo and circulating neutrophils were purified from the blood and examined 16 h later, CST7 was significantly upregulated (Figure 5D). Together, these findings suggest that an endogenous rather than exogenous factor contributes to the CST7 upregulation during inflammation.
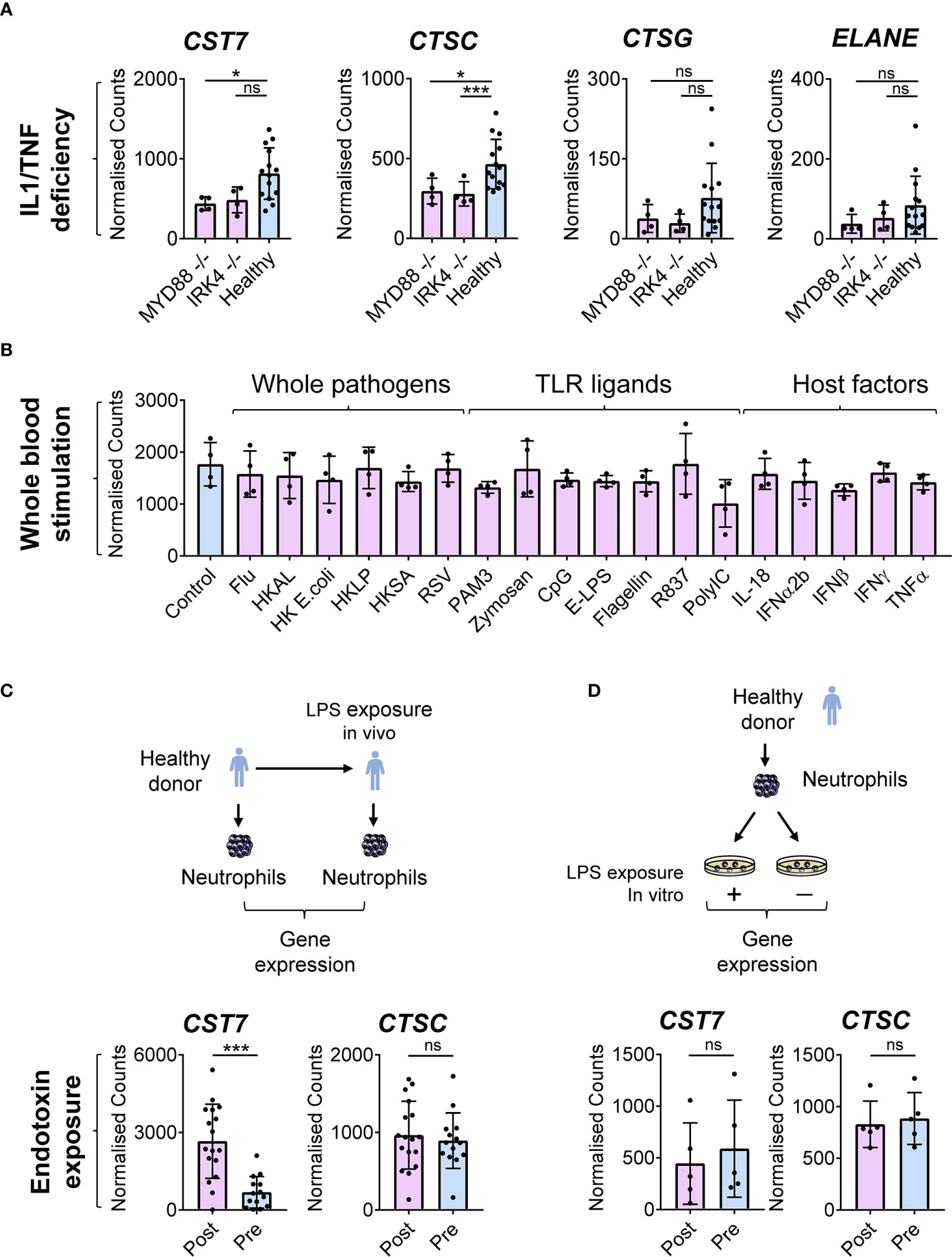
Figure 5 CST7 upregulation during inflammation is regulated by host factors (A) Baseline gene expression of CST7, CTSC, CTSG and ELANE in whole blood samples form a cohort of healthy controls or patients with either complete MYD88 deficiency or complete IRAK-4 deficiency. (B) CST7 expression in whole blood cells following stimulation in vitro with heat-killed (HK) whole pathogens, purified TLR ligands and host cytokines (GSE30101). Whole blood from four healthy donors was incubated for 6 h with one of the listed stimuli before gene expression was measured by microarray. (C) Expression of CST7 and CTSC in a circulating neutrophils from healthy individuals taken prior to or 16 h after bronchoscopic instillation of endotoxin (LPS). (D) Expression of CST7 and CTSC in purified neutrophils isolated from healthy donors following stimulation with endotoxin in vitro. (A–D), each symbol represents an individual donor, while bars represent the mean ± standard deviation. The level of statistical significance is indicated on the plot: *P < 0.05 ***P < 0.001, ns, not significant.
Discussion
Cystatin F has been poorly characterized as a protease inhibitor and has not been linked to neutrophils until now. By using public transcriptome data, we discovered that CST7 expression is robustly increased in neutrophils cultured in vitro in the presence of plasma from sepsis patients and also in the circulating neutrophils of sepsis patients in vivo. Further analysis using additional datasets established that this neutrophil-specific CST7 upregulation occurs across a diverse range of sterile and pathogen-driven inflammatory conditions. Publicly available transcriptomic data also showed that this induction is independent of a broad range of microbial products, suggesting that CST7 upregulation during inflammation is regulated by a circulating endogenous soluble factor. These novel findings suggest that CST7 is a previously unrecognized major inflammation-responsive gene in humans. Given the known anti-cytotoxic actions of cystatin F in NK cells (3), further in-depth investigations focusing on its expression patterns and functions may reveal fundamental mechanisms regulating inflammation in human disease.
Our literature search of the PubMed database revealed that no link has been established between CST7 expression in the context of sepsis or in neutrophils. This approach, which has been used in previous studies (1, 2), is a simple and rapid method for establishing novel gene expression in the context of a condition or cell of interest. Subsequent full text searches using Google Scholar revealed that CST7 was identified as a DEG during sepsis in three studies (23–25). However, in each of these studies CST7 appeared among dozens of other DEGs and none of these studies commented on the biological relevance of CST7 upregulation during sepsis. Thus, we consider that CST7 upregulation in neutrophils is a novel biological finding in the context of sepsis.
Throughout this study we performed multicohort analyses of genes of interest to determine an overall fold change in the expression of a gene across any number of compatible datasets. This type of multicohort gene expression analysis has been used to validate biomarker gene signatures across inflammatory diseases (26, 27), including sepsis (28). Here we apply this approach to a single gene across a variety of different diseases and conditions. Comprehensive exploration of the NCBI GEO was performed to identify biologically relevant datasets for each disease and cell-type explored in this study, which led to the discovery of the novel association between neutrophil-expressed CST7 and acute inflammation in humans. Using our analysis pathway, we additionally identified novel neutrophil-specific changes in the expression of two other genes, SAMSN1 and BATF during inflammation, and each of these genes could form the basis of an additional study. Overall, the multicohort analysis process is straightforward and provides a wealth of information on gene regulation to inform hypotheses for future experiments. Moreover, the entire process is free and uses publicly available data and software.
With the current knowledge of CST7 in the literature, the new findings generated from this study can be used to inform hypotheses for future research. Of particular interest, was the ability of cystatin F to inhibit cathepsin C and thus, inhibit the activation of serine proteases (17). While cystatin F can function in an autocrine manner, it can also act in trans, being secreted and taken up by other cells. If the upregulation of CST7 in neutrophils corresponds to an increase in cystatin F protein, it can be speculated that it acts on the neutrophils themselves, other cells, or both. If neutrophilic cystatin F acts in an autocrine manner, this would be of great interest given its known function to suppress cathepsin C.
We hypothesize that cystatin F suppression of cathepsin C in neutrophils may be an as-yet unknown anti-inflammatory function in neutrophils. If neutrophilic cystatin F is secreted, then it may act to suppress cathepsin C in other cells. Furthermore, if cystatin F is secreted by neutrophils in a tumor microenvironment (14), then it may contribute to pro-tumor immunosuppression. We identified a dataset (GSE101584) for tumor-associated neutrophils in a mouse cancer model (29) and found a significant increase in CST7 expression compared to that in naïve neutrophils. Given that cystatin F also inhibits cathepsin C in mice (30), this supports the hypothesis that cystatin F plays a role in the tumor microenvironment. The ability of cystatin F to inhibit cathepsin C in neutrophils is a key area for future investigation. To test these hypotheses, future bench-based research is required to determine first whether the increase in CST7 in neutrophils corresponds to an increase in cystatin F protein and whether the protein is secreted.
In this study we have both identified CST7 as a gene of interest and assessed its regulation across a variety of public transcriptomic datasets. To date, no role for CST7 has been established in neutrophils or in any inflammatory disease. Having identified CST7 upregulation across inflammatory diseases and in a neutrophil-specific manner, we were able to formulate hypotheses about the potential inhibitory activity of cystatin F. Our findings suggest that cystatin F or CST7 may potentially be used, either independently or in conjunction with C-reactive protein, as an indicator of acute inflammation in the clinic.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics Statement
Ethical review and approval was not required for the study on human participants in accordance with the local legislation and institutional requirements. Written informed consent from the participants’ legal guardian/next of kin was not required to participate in this study in accordance with the national legislation and the institutional requirements.
Author Contributions
AJS conceived the study design, performed literature analysis, performed transcriptomic analyses, and wrote the manuscript. MG assisted with analysis conceptualization. DC and CF supervised the work. All authors contributed to the article and approved the submitted version.
Funding
AJS is supported by an Australian Government Research Training Program (RTP) Scholarship.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be constructed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.634119/full#supplementary-material
Supplementary Table 1 | Characteristics of the public datasets used in this study.
References
1. Rinchai D, Kewcharoenwong C, Kessler B, Lertmemongkolchai G, Chaussabel D. Increased abundance of ADAM9 transcripts in the blood is associated with tissue damage. F1000Research (2015) 4:89. doi: 10.12688/f1000research.6241.2
2. Toufiq M, Roelands J, Alfaki M, Syed Ahamed Kabeer B, Saadaoui M, Lakshmanan AP, et al. Annexin A3 in sepsis: novel perspectives from an exploration of public transcriptome data. Immunology (2020) 161:291–302. doi: 10.1111/imm.13239
3. Nanut MP, Sabotič J, Švajger U, Jewett A, Kos J. Cystatin F Affects Natural Killer Cell Cytotoxicity. Front Immunol (2017) 8:1459. doi: 10.3389/fimmu.2017.01459
4. Kos J, Nanut M, Prunk M, Sabotič J, Dautović E, Jewett A. Cystatin F as a regulator of immune cell cytotoxicity. Cancer Immunol Immunother (2018) 67:1931–8. doi: 10.1007/s00262-018-2165-5
5. Davis S, Meltzer PS. GEOquery: a bridge between the Gene Expression Omnibus (GEO) and BioConductor. Bioinformatics (2007) 23:1846–7. doi: 10.1093/bioinformatics/btm254
6. Ritchie ME, Phipson B, Wu D, Hu Y, Law CW, Shi W, et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res (2015) 43:e47. doi: 10.1093/nar/gkv007
7. Haynes WA, Vallania F, Liu C, Bongen E, Tomczak A, Andres-Terrè M, et al. Empowering Multi-Cohort Gene Expression Analysis To Increase Reproducibility. Pac Symp Biocomput (2017) 22:144–53. doi: 10.1142/9789813207813_0015
9. Halfon S, Ford J, Foster J, Dowling L, Lucian L, Sterling M, et al. Leukocystatin, a new Class II cystatin expressed selectively by hematopoietic cells. J Biol Chem (1998) 273:16400–8. doi: 10.1074/jbc.273.26.16400
10. Colbert JD, Matthews SP, Miller G, Watts C. Diverse regulatory roles for lysosomal proteases in the immune response. Eur J Immunol (2009) 39:2955–65. doi: 10.1002/eji.200939650
11. Colbert JD, Matthews SP, Kos J, Watts C. Internalization of exogenous cystatin F supresses cysteine proteases and induces the accumulation of single-chain cathepsin L by multiple mechanisms. J Biol Chem (2011) 286:42082–90. doi: 10.1074/jbc.M111.253914
12. Utsunomiya T, Hara Y, Kataoka A, Morita M, Arakawa H, Mori M, et al. Cystatin-like metastasis-associated protein mRNA expression in human colorectal cancer is associated with both liver metastasis and patient survival. Clin Cancer Res (2002) 8:2591–4.
13. Yang C, Yu T, Liu Z, Ye X, Liao X, Wang X, et al. Cystatin F as a key family 2 cystatin subunit and prognostic biomarker for earlystage pancreatic ductal adenocarcinoma. Oncol Rep (2019) 42:79–90. doi: 10.3892/or.2019.7135
14. Zhou X, Wang X, Huang K, Liao X, Yang C, Yu T, et al. Investigation of the clinical significance and prospective molecular mechanisms of cystatin genes in patients with hepatitis B virusrelated hepatocellular carcinoma. Oncol Rep (2019) 42:189–201. doi: 10.3892/or.2019.7154
15. Ma J, Tanaka KF, Shimizu T, Bernard CC, Kakita A, Takahashi H, et al. Microglial cystatin F expression is a sensitive indicator for ongoing demyelination with concurrent remyelination. J Neurosci Res (2011) 89:639–49. doi: 10.1002/jnr.22567
16. Liang J, Li N, Zhang Y, Hou C, Yang X, Shimizu T, et al. Disinhibition of Cathepsin C Caused by Cystatin F Deficiency Aggravates the Demyelination in a Cuprizone Model. Front Mol Neurosci (2016) 9:152. doi: 10.3389/fnmol.2016.00152
17. Hamilton G, Colbert JD, Schuettelkopf AW, Watts C. Cystatin F is a cathepsin C-directed protease inhibitor regulated by proteolysis. EMBO J (2008) 27:499–508. doi: 10.1038/sj.emboj.7601979
18. Bird PI, Trapani JA, Villadangos JA. Endolysosomal proteases and their inhibitors in immunity. Nat Rev Immunol (2009) 9:871. doi: 10.1038/nri2671
19. Magister Š, Tseng H-C, Bui VT, Kos J, Jewett A. Regulation of split anergy in natural killer cells by inhibition of cathepsins C and H and cystatin F. Oncotarget (2015) 6:22310–27. doi: 10.18632/oncotarget.4208
20. Prunk M, Nanut MP, Sabotic J, Svajger U, Kos J. Increased cystatin F levels correlate with decreased cytotoxicity of cytotoxic T cells. Radiol Oncol (2019) 53:57–68. doi: 10.2478/raon-2019-0007
21. McGuire MJ, Lipsky PE, Thiele DL. Generation of active myeloid and lymphoid granule serine proteases requires processing by the granule thiol protease dipeptidyl peptidase I. J Biol Chem (1993) 268:2458–67. doi: 10.1016/S0021-9258(18)53798-4
22. Adkison AM, Raptis SZ, Kelley DG, Pham CT. Dipeptidyl peptidase I activates neutrophil-derived serine proteases and regulates the development of acute experimental arthritis. J Clin Invest (2002) 109:363–71. doi: 10.1172/jci13462
23. Chen H, Li Y, Li T, Sun H, Tan C, Gao M, et al. Identification of Potential Transcriptional Biomarkers Differently Expressed in Both S. aureus- and E. coli-Induced Sepsis via Integrated Analysis. BioMed Res Int (2019) 2019:2487921. doi: 10.1155/2019/2487921
24. Ahmad S, Singh P, Sharma A, Arora S, Shriwash N, Rahmani AH, et al. Transcriptome Meta-Analysis Deciphers a Dysregulation in Immune Response-Associated Gene Signatures during Sepsis. Genes (Basel) (2019) 10. doi: 10.3390/genes10121005
25. Xie K, Kong S, Li F, Zhang Y, Wang J, Zhao W. Bioinformatics-Based Study to Investigate Potential Differentially Expressed Genes and miRNAs in Pediatric Sepsis. Med Sci Monit (2020) 26:e923881. doi: 10.12659/msm.923881
26. Andres-Terre M, McGuire HM, Pouliot Y, Bongen E, Sweeney TE, Tato CM, et al. Integrated, Multi-cohort Analysis Identifies Conserved Transcriptional Signatures across Multiple Respiratory Viruses. Immunity (2015) 43:1199–211. doi: 10.1016/j.immuni.2015.11.003
27. Sweeney TE, Braviak L, Tato CM, Khatri P. Genome-wide expression for diagnosis of pulmonary tuberculosis: a multicohort analysis. Lancet Respir Med (2016) 4:213–24. doi: 10.1016/s2213-26001600048-5
28. Sweeney TE, Shidham A, Wong HR, Khatri P. A comprehensive time-course-based multicohort analysis of sepsis and sterile inflammation reveals a robust diagnostic gene set. Sci Trans Med (2015) 7:287ra271. doi: 10.1126/scitranslmed.aaa5993
29. Shaul ME, Levy L, Sun J, Mishalian I, Singhal S, Kapoor V, et al. Tumor-associated neutrophils display a distinct N1 profile following TGFβ modulation: A transcriptomics analysis of pro- vs. antitumor TANs. Oncoimmunology (2016) 5:e1232221. doi: 10.1080/2162402x.2016.1232221
Keywords: neutrophil, transcriptome, cystatin F, data mining, meta-analysis, literature analysis
Citation: Sawyer AJ, Garand M, Chaussabel D and Feng CG (2021) Transcriptomic Profiling Identifies Neutrophil-Specific Upregulation of Cystatin F as a Marker of Acute Inflammation in Humans. Front. Immunol. 12:634119. doi: 10.3389/fimmu.2021.634119
Received: 27 November 2020; Accepted: 17 March 2021;
Published: 01 April 2021.
Edited by:
Antonio Cappuccio, Mount Sinai Hospital, United StatesReviewed by:
Carlos Alberto Moreira-Filho, University of São Paulo, BrazilWinston Haynes, Serimmune, United States
Copyright © 2021 Sawyer, Garand, Chaussabel and Feng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Damien Chaussabel, ZGNoYXVzc2FiZWxAc2lkcmEub3Jn; Carl G. Feng, Y2FybC5mZW5nQHN5ZG5leS5lZHUuYXU=
†These authors have contributed equally to this work