 Hannes Lindahl
Hannes Lindahl Tomas Olsson
Tomas Olsson
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 22 February 2021
Sec. Cytokines and Soluble Mediators in Immunity
Volume 12 - 2021 | https://doi.org/10.3389/fimmu.2021.618110
This article is part of the Research Topic Role of the IL-23/IL-17 Pathway in Chronic Immune-Mediated Inflammatory Diseases: Mechanisms and Targeted Therapies View all 21 articles
Interleukin-22 (IL-22) is secreted by a wide range of immune cells and its downstream effects are mediated by the IL-22 receptor, which is present on non-immune cells in many organs throughout the body. IL-22 is an inflammatory mediator that conditions the tissue compartment by upregulating innate immune responses and is also a homeostatic factor that promotes tissue integrity and regeneration. Interestingly, the IL-22 system has also been linked to many T cell driven inflammatory diseases. Despite this, the downstream effects of IL-22 on the adaptive immune system has received little attention. We have reviewed the literature for experimental data that suggest IL-22 mediated effects on T cells, either transduced directly or via mediators expressed by innate immune cells or non-immune cells in response to IL-22. Collectively, the reviewed data indicate that IL-22 has a hitherto unappreciated influence on T helper cell polarization, or the secretion of signature cytokines, that is context dependent but in many cases results in a reduction of the Th1 type response and to some extent promotion of regulatory T cells. Further studies are needed that specifically address these aspects of IL-22 signaling, which can benefit the understanding and treatment of a wide range of diseases.
Interleukin-22 (IL-22) is often described as a cytokine that is expressed by immune cells but that exclusively acts on non-immune cells (1, 2). Its role is best understood at so called barrier surfaces such as the skin, lungs, and gut where the effects of IL-22 ligation typically involve proliferation, regeneration, or activation of innate immune mechanisms. Interestingly, the IL-22 system has also been linked to a range of T cell driven inflammatory diseases such as rheumatoid arthritis (RA), graft versus host disease (GvHD), and multiple sclerosis (MS) (2–4). However, relatively little is known about how IL-22 influences T cell polarization. The fact that the IL-22 receptor (IL-22R) has mostly been shown to be absent on immune cells has likely contributed to this (1). Although there are an increasing number of reports of IL-22R expression on immune cells, in most cases any effect of IL-22 on T cells is likely indirect, transduced by mediators originating from IL-22 receptor expressing non-immune cells. IL-22 biology in general has been comprehensively described in excellent review articles (2, 4). Here, we have reviewed the literature for data that suggest effects of IL-22 on T helper cell polarization or secretion of the signature cytokines IFNγ and IL-17. Any experimental setup where IL-22 signaling has been specifically targeted has been screened for such data. By applying this perspective to the literature, we hope to promote further research on the role of IL-22 in shaping adaptive immunity in inflammatory and infectious diseases.
IL-22 was initially described as a cytokine produced by IL-9 activated T cells (5) and was later associated with the Th17 lineage (6). It has now become clear that IL-22 is not only frequently produced independently of IL-17 (7–9) but can also be produced by a wide range of other immune cells (Table 1). There are also reports of IL-22 being produced by non-hematopoietic cells (35, 36).
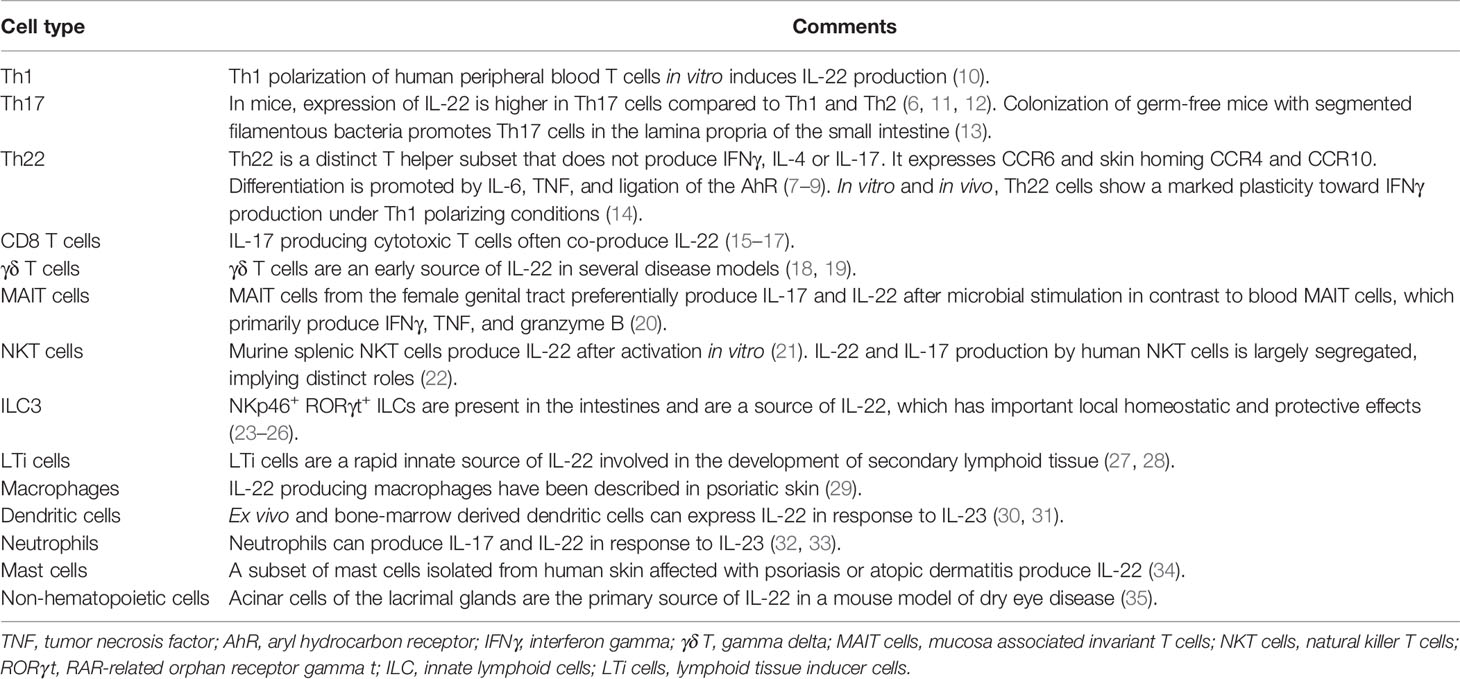
Table 1 Cell types that express IL-22.
The most well-established inducer of IL-22 is IL-23 (6, 30). The IL-23 receptor has been detected on Th17 cells, natural killer T (NKT) cells, type 3 innate lymphoid cells (ILC3), gamma delta (γδ) T cells, macrophages, dendritic cells (DC), and neutrophils. Stimulation of these cells with IL-23 can induce production of IL-22 (11, 18, 29, 31, 32, 37, 38). Moreover, IL-1β can act both independently of and synergistically with IL-23 to induce IL-22 (18, 37, 39). IL-6 and TNF polarize naïve T cells to the Th22 lineage (8). The aryl hydrocarbon receptor (AhR) promotes the differentiation of several IL-22 producing cells such as Th17 cells, Th22 cells, and ILC3 (40, 41). Ahr is located in the cytoplasm where it senses endogenous and exogenous ligands, leading to nuclear translocation and transcription of IL22 and other genes (41–43). Similarly, transcription factor RAR-related orphan receptor gamma t (RORγt) is also necessary for the differentiation of Th17 cells and ILC3 (40, 44). Furthermore, IL-22 induction by Toll-like receptor (TLR)2 ligation has also been described in innate lymphoid cells (45, 46).
Negative regulators of IL-22 production include transforming growth factor beta (TGF-β), IL-27, and Inducible T cell co-stimulator (ICOS), all of which transduce signals to the transcription factor c-Maf (47–49). Interestingly, IL-22 is one of few cytokines that has a dedicated soluble antagonist molecule to regulate its effects in vivo, IL-22 binding protein (IL-22BP), transcribed from the gene IL22RA2 (Figure 1) (50, 51). Insights about the role of IL-22 can therefore also be gained from experiments in which IL-22BP levels have been manipulated.
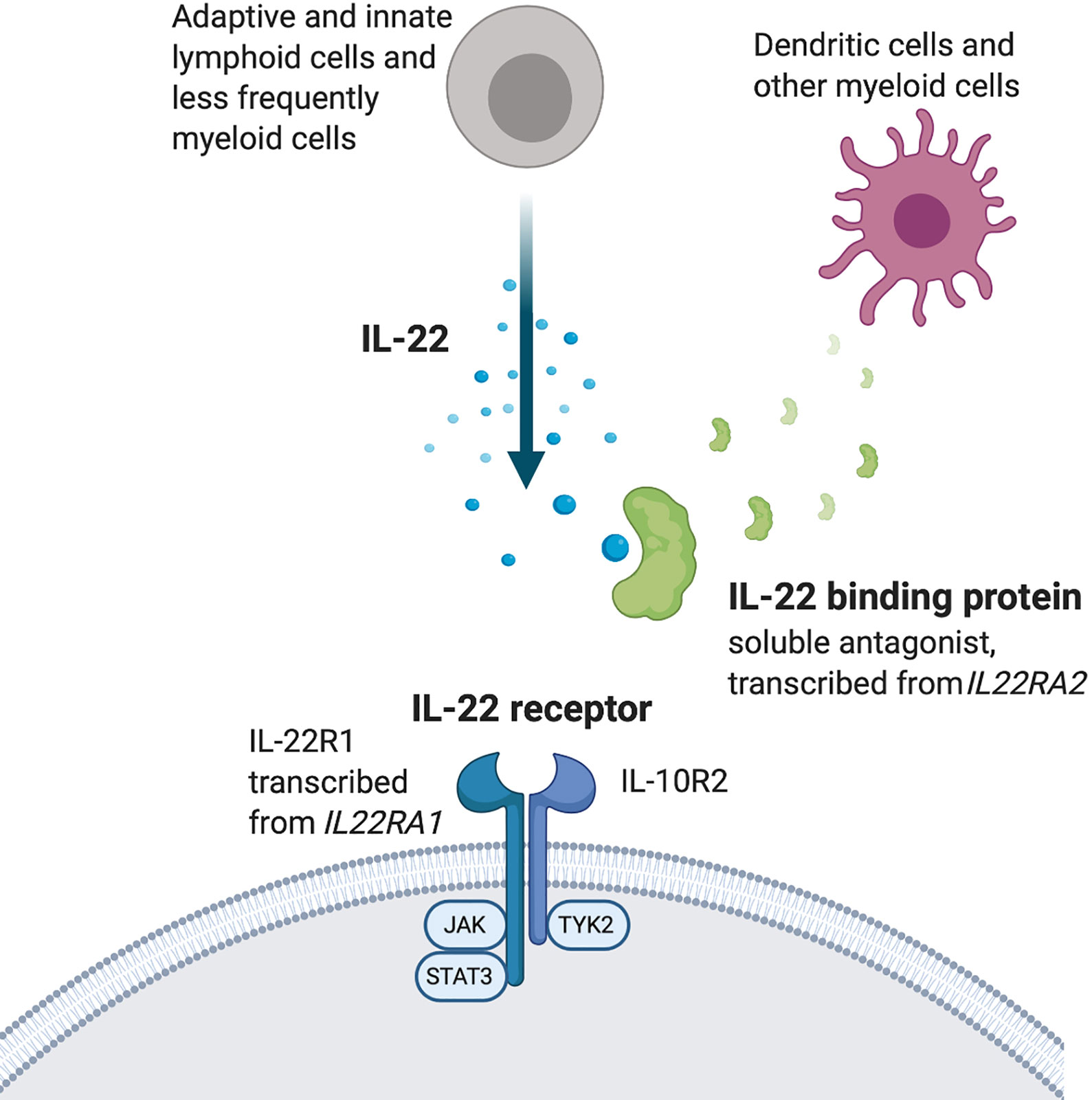
Figure 1 The IL-22 system.
The IL-22R is a heterodimer of the subunit IL-10R2, which is expressed by most cells, and the more selectively distributed subunit IL-22R1 (Figure 1) (52–56). Ligation of IL-22R with IL-22 induces activation of the tyrosine kinases JAK1 and Tyk2, which in turn activates STAT3. STAT1 and STAT5 activation has also been reported (1, 57) as well as downstream signaling via the MAPK pathways (36, 58–60). IL-22R is expressed by epithelial and parenchymal cells in a wide range of tissues throughout the body. It is highly expressed at, but not limited to, barrier surfaces such as the skin, gut and lungs. Other tissues where IL-22 exerts effects include liver, thymus, pancreas, kidney, and synovium (25, 36, 61–63).
IL-22R has mostly not been detected on immune cells, which has established IL-22 as a cytokine that mediates one-way signaling from immune cells to tissue cells. However, there are reports of IL-22R expression on both myeloid and lymphoid cells and functional data on the effects of IL-22 on these cells. IL-22 has direct effects on splenic CD4 T cells, B cells and CD11b+ cells in a mouse model of autoimmune arthritis (64–67) and on splenic CD11b+ cells in a mouse model of autoimmune uveitis (68). Moreover, infiltrating mononuclear cells in the salivary glands of patients with primary Sjogren’s syndrome express IL-22R1 shown by immunohistochemistry (69). Further characterization by flow cytometry showed IL-22R expression on macrophages and, to a lesser extent, on T and B cells both in salivary glands and in the circulation from primary Sjogren’s syndrome patients but IL-22R was not detected at all in samples from non-specific chronic sialoadenitis. During the acute stage of Mycobacterium tuberculosis infection in mice, IL-22R is expressed primarily on epithelial cells. However, during the chronic stage both epithelial cells and recruited macrophages express IL-22R, which also was observed in human samples (46). IL-22 inhibits intracellular growth of M tuberculosis in human monocyte derived macrophages (70). Stimulating peripheral blood mononuclear cells from primary Sjogren’s syndrome patients in vitro with IL-22 increased production of IL-17, which was not seen in non-specific chronic sialoadenitis (69). CD14+ adipose tissue macrophages, but not circulating CD14+ cells, express IL-22R1 shown by western blot and FACS (71). IL-22 strongly induces IL-1β from CD14+ adipose tissue macrophages.
The animal model experimental autoimmune uveitis (EAU) can be induced by immunization with retinal autoantigens and disease severity is evaluated by fundoscopy. Treatment with IL-22 before onset of EAU in mice results in reduced severity of disease and delayed onset (68). The protective effect of IL-22 in this model is associated with an overall reduction in eye-infiltrating cells with a proportional decrease in T cells and neutrophils. Interestingly, the IL-22R subunit Il22ra1 is highly expressed by splenic CD11b+ cells day 12 after immunization. When stimulated in vitro with IL-22, these cells produce less IL-6, IL-12, IL-23, and IL-1β but more IL-10 and TGF-β. Consistent with a tolerogenic phenotype, they also express less MHC class II, CD80, CD86, and CD40 but more PD-L1. IL-22 treated APCs yield less antigen specific T cell proliferation in vitro and induce less IFNγ and IL-17 production but more IL-10 production.
Dry eye disease is predominantly a Th17 cell driven autoimmune disorder resulting in ocular mucosal inflammation, which in severe cases can lead to damage to the cornea and vision loss. Both IL-17 and IL-22 are elevated in tear and lacrimal fluid in persons with dry eye disease (35). IL-17 levels are positively correlated and IL-22 levels negatively correlated with severity of disease. In a mouse model of dry eye disease IL-22 neutralizing antibody or Il22 gene deletion both result in increased infiltration of Th17 cells and more severe disease, consistent with a protective effect of IL-22. IL-22R is expressed on the ocular surface and in vitro stimulation of corneal epithelial cells with IL-22 inhibits expression of inflammatory mediators, including the Th17-inducing cytokines IL-6 and IL-23.
The homeostatic role of IL-22 in the gastrointestinal tract has received much attention (72). Activation of AhR alone promotes IL-22 secretion by intestinal leukocytes but it also acts synergistically with transcription factor RORγt (40). Steady state Ahr−/− mice have an increased number of intestinal Th17 cells as a result of commensal segmented filamentous bacteria expanding and locally inducing Th17 cells (73). Administration of IL-22 to Ahr−/− reduces Th17 cells to numbers that approach normal levels. The increased intestinal Th17 numbers in Ahr−/− mice do not lead to any overt gut pathology. However, when one allele of the RORγt gene is deleted in Ahr−/− mice IL-22 production is further decreased, which is associated with occasional observations of spontaneous colitis. The authors of this study suggest that in immunocompromised patients, that potentially have impaired IL-22 production, the normally innocuous segmented filamentous bacteria may expand and cause intestinal autoimmunity through induction of pathogenic Th17 cells.
Collagen induced arthritis (CIA) is an animal model of RA. Daily administration of recombinant IL-22 starting before onset of arthritis reduces disease severity of CIA in DBA mice but does not alter incidence (74). The IL-22 treatment increases expression of IL-10 in the spleen and administration of neutralizing IL-10 antibody together with IL-22 cancels the protective effect of the latter. IL-22 induces secretion of IL-10 from CD11b+ splenocytes harvested during early disease. Addition of IL-22 decreases IFNγ secretion induced in vitro by polyclonal T cell activation of splenocytes harvested from mice with early disease as well as from naïve mice. In contrast, stimulation with IL-22 increases IL-17 secretion from splenocytes restimulated in vitro with collagen.
In a follow-up study, Justa et al. showed that IL-22 has a dual role in CIA. Treatment with a neutralizing IL-22 antibody before onset of arthritis increases disease severity consistent with the previous study, whereas treatment after onset reduces disease severity (64). Surprisingly, they show that after disease onset IL-22 actually decreases in vitro IFNγ production from splenocytes after restimulation with collagen. No effect on IL-17 or IL-10 was observed in this context. However, IL-22 causes increased proportions of Th1 cells in the draining lymph nodes but still does not influence Th17 cells. In the affected joints the proportions of Th17 cells are increased as result of IL-22 signaling. Il22ra1 mRNA and IL-22R1 protein expression is detected in CD4+ splenocytes from mice with arthritis but not at baseline or during the initiation phase. CD11c+ cells and CD4 T cells from the spleen of arthritic mice co-cultured in the presence of recombinant IL-22 results in less production of IFNγ but more IL-17. The protective effect of treatment with anti-IL-22 after disease onset is blunted when performing the experiment on Ifng−/− mice. In contrast, mice treated with anti-IL-22 before disease onset exacerbates disease, which is associated with decreased Th1 and unchanged Th17 proportions in draining lymph nodes but increased IL-17 production from cells from the affected joints. In summary, the pathogenic effect of IL-22 during late CIA is dependent on suppression of IFNγ, possibly mediated directly via IL-22R expression, which is upregulated on CD4 T cells during this phase of the disease.
A study by another group primarily focused on the influence of IL-22 on autoantibody formation in the context of CIA in C57BL/6 mice but also includes some data on T cells (75). They report that germinal center and autoantibody formation is reduced in Il22−/− mice and IL-22R expression is detected in follicular dendritic cell-like stromal cells. Furthermore, human lymphoid stromal cells produce B cell attracting chemokines CXCL12 and CXCL13 upon stimulation with IL-22. Regarding T cells, they show that, despite having less severe CIA, the Il22−/− mice have increased proportions of Th17 cells in the spleen ten days after immunization. The Th17 cells retain their pathogenic potential, which is shown by assessing secretion of IL-6 in co-cultures with synovial fibroblasts.
Psoriasis arthritis is characterized by skin lesions and articular inflammation and is often accompanied by osteoporosis. As with many of the inflammatory diseases Th17 cells have been shown to be critical in psoriasis arthritis (76). Central in the differentiation of Th17 cells is activation of transcription factor STAT3. In a novel model of psoriasis arthritis based on overactive STAT3 function specifically in CD4 T cells, the psoriatic skin phenotype and osteopenia were both ameliorated by either neutralizing IL-17 antibody or genetic deletion of Il22 (77). The Il22−/− mice had no reduction in total T helper cell infiltration in the skin but had reduced proportions of Th17 cells consistent with a disease promoting role of IL-22. Increased proportions of Tregs but also Th1 cells were seen in the inflamed skin of the Il22−/− mice. Although an improvement in the osteoporosis phenotype was seen in the Il22−/− mice the proportion of IL-17+ cells in the bone marrow was increased. Similarly, the proportions of Th1 cells and Tregs were also increased.
Experimental autoimmune encephalomyelitis (EAE) is an animal model of MS and can be induced by immunization with myelin autoantigens, which leads to a T cell driven disease characterized by ascending paralysis. Although Il22−/− mice have no apparent phenotype in EAE experiments (78), mice lacking the endogenous antagonist molecule IL-22BP have less severe disease compared to wild type mice, suggesting a protective role for IL-22 (79). These seemingly contradictory findings can be reconciled if the presence of IL-22BP blocks IL-22 in wild type mice to such a degree that they are indistinguishable from Il22−/− mice. Consistent with the reduced EAE severity, IL-22BP knockout mice have decreased infiltration of Ly6C+ inflammatory monocytes in the central nervous system (79). Although not statistically significant, a trend toward less Il-17+ T cells infiltrating the CNS in IL-22BP knockout mice is seen. In a follow-up study the role of IL-22BP in EAE was further dissected using an inducible in vivo IL-22BP knockdown rat strain. Reducing IL-22BP expression before EAE immunization results in reduced incidence and severity of disease in conjunction with increased proportions of Tregs and decreased proportions of Th1 cells in the lymph nodes that drain the site of immunization (80). Taken together, IL-22 appears to have a protective effect in autoimmune neuroinflammation but, unlike many other animal models discussed in this article, the effect of endogenous IL-22 is normally blocked by IL-22BP. Homeostatic expression of this soluble antagonist molecule is detected in many tissues including secondary lymphoid organs where immune responses are initiated and in the CNS by microglia, making it an interesting pharmacological target in MS and other neuroinflammatory diseases.
In contrast to EAE, IL-22BP knockout mice have more severe disease in an imiquimod induced model of psoriasis (81). This is consistent with the well documented pathogenic role of IL-22 in rodent models of psoriasis (4). The draining lymph nodes of the IL-22BP knockout mice have larger proportions of CD44+ CD62L− activated T cells. Also, proportions of both IFNγ producing and IL-17 producing CD8 T cells are elevated compared to wild type mice.
The role of IL-22 has been investigated in a model of atopic dermatitis and allergic asthma in which mice are epicutaneously sensitized to the model antigen ovalbumin followed by intranasal challenge (82). Sensitization alone results in increased serum levels of IL-22 and after intranasal challenge increased Il22 mRNA as well as increased eosinophil and neutrophil infiltration is detected in the airways. Applying the disease model to Il22−/− mice results in less eosinophils and neutrophils in the airways and reduces airway resistance after methacholine provocation. Cells acquired by bronchoalveolar lavage from Il22−/− mice contain more IFNγ producing type 1 ILCs compared to wild type mice. No change was observed regarding IFNγ+ CD4 and CD8 T cells. The authors go on to show that IL-22 synergizes with TNF to drive the neutrophil dominated airway inflammation.
IL-22 has well described protective and regenerative effects on the liver parenchyma after various insults (61, 83–85). In contrast, IL-22 has a disease promoting net effect on chronic hepatitis induced in a mouse strain, transgenically made to express hepatitis B virus antigens, that is treated with anti-CD137 to activate T cells. Using this model, Il22+/+ mice have increased disease severity, infiltration of granulocytes and T cells compared to Il22−/− mice (86). Moreover, Il22+/+ mice also have an altered balance between T cell responses compared to the Il22−/− mice, with higher hepatic and splenic Th17 numbers but unchanged Th1 numbers. Anti-IL-22 treatment reduces the hepatic stellate cell expression of CXCL10 and CCL20, ligands for CXCR3 and CCR6 respectively, both being chemokine receptors expressed on Th17 cells. IL-22 treated hepatic stellate cells have increased chemotactic potential in a transwell assay and specifically attracts Th17 cells.
Th1 and Th17 cells both promote atherosclerosis, whereas Tregs are protective. The role of Th2 cells is unclear. Circulating IL-22 and Th22 cells are both elevated in patients with acute coronary syndrome (87). The role of IL-22 has been investigated using a mouse model in which atherosclerosis is induced by feeding ApoE−/− mice a so called western diet (88). By administering either recombinant IL-22 or neutralizing IL-22 antibody it was shown that IL-22 aggravates atherosclerosis, which was associated with increased macrophage and T cell infiltration in the blood vessels with a proportional increase in Th17 cells. IL-22 receptor is expressed in mouse aortic tissue and its expression is further elevated in the atherosclerosis prone mice.
Chlamydial organisms are obligate intracellular gram-negative bacteria that can cause pneumonia in humans, which is modeled in mice using Chlamydia muridarum. The role of IL-22 has been investigated in this disease model using either administration of recombinant IL-22 or neutralizing IL-22 antibody (89). In both cases IL-22 was shown to improve outcome associated with increased IL-17 production in infiltrating cells in the lungs and spleen ex vivo as well as in splenocytes after antigen specific activation in vitro. Conflicting results were obtained regarding IFNγ production. Anti-IL-22 treated mice also display lower IL-10 production ex vivo and after antigen specific stimulation in vitro.
To mimic an opportunistic fungal lung infection in an immunocompromised patient, mice were treated with a neutrophil-depleting antibody prior to infection with Aspergillus fumigatus (90). Furthermore, to investigate the role of gut microbiota in this context, the mice were pre-treated with vancomycin prior to infection, which suppresses intestinal segmented filamentous bacteria and other gram-positive bacteria. Vancomycin pre-treatment did not affect disease severity in that study but did, however, reduce the presence of IL-17 and IL-22 in the lungs. They show that Il22−/− mice have increased colonization of intestinal segmented filamentous bacteria, consistent with Ivanov et al. (13). After infection with A. fumigatus, the amount of IL-17 in Il22−/− lung tissue appeared to be increased (p = 0.06), an effect that was obliterated by pre-treatment with vancomycin. Vancomycin pre-treatment of wild type mice followed by fecal transplant, neutrophil depletion, and A. fumigatus infection results in more Th17 cells infiltrating the lungs after fecal transplants from Il22−/− compared to wild type mice. Transferring serum from wild type mice colonized with segmented filamentous bacteria to uncolonized wild type mice infected with A. fumigatus results in a decrease in Th17 cells in the lungs compared to transferring serum from colonized Il22−/− mice. This effect is counteracted by premixing the serum with IL-1Ra (Anakinra). Wild type serum has decreased levels of IL-1α but similar levels of IL-1β compared to Il22−/− serum. Collectively making it plausible that IL-22 modulates serum IL-1Ra ligands leading to accumulation of Th17 cells in the lungs after fungal infection.
The immune response to Mycobacterium tuberculosis varies depending on the infecting strain. The hypervirulent W-Beijing lineage of M tuberculosis is a growing health threat that is often associated with human immunodeficiency virus and drug resistance. This infection is modeled in mice with the M. tuberculosis strain HN878 (46). In this context, Il22−/− mice have normal susceptibility to infection but have higher bacterial burden in the lungs during the chronic stage. IL-22 deficiency has no impact on alveolar macrophages, monocytes, or recruited macrophages in the lungs during the acute stage of disease but reduces the numbers of IFNγ secreting CD4 and CD8 T cells as well as IL-17 secreting CD8 T cells. However, during the chronic stage the numbers of monocytes and recruited macrophages are lower in the Il22−/− mice consistent with the impaired bacterial clearance. Interestingly, IL-17, which has been shown to be required for protective immunity to this strain (91), is increased in CD8 T cells at this stage despite the poorer outcome. This implies that the lack of IL-22 and associated reduction in IFNγ production and increased bacterial burden are not compensated for by increased IL-17 production.
Plasma levels of IL-22 are elevated in acute infection with Plasmodium falciparum in humans as well as in the murine malaria model Plasmodium berghei infection in C57BL/6 mice. Il22−/− mice have earlier onset of cerebral malaria compared to wild type mice despite similar parasite burden in the liver and decreased parasitemia (92). Similar results were seen in experiments using neutralizing IL-22 antibody in vivo. Day 3 after infection the Il22−/− mice have increased proportions of IFNγ+ cells in the spleen, evident in both CD4 and CD8 T cells as well as γδ T cells. Although no difference in IFNγ expression was observed day 6, IL-17 expression in CD4 T cells as well as γδ T cell is decreased at both time points in Il22−/− mice. APC and CD8 T cell co-cultures using bone marrow derived dendritic cells pulsed with antigen showed that both APCs and CD8 T cells from Il22−/− mice are primed to produce more IFNγ compared to cells from wild type mice. Both CD11c+ CD11b− dendritic cells ex vivo and bone marrow derived dendritic cells from Il22−/− mice have increased lipopolysaccharide induced expression of CD80 and CD86. Adoptive transfer of splenocytes from OT1 mice, that have ovalbumin specific T cell receptors, into wild type or Il22−/− recipients followed by infection with a transgenic ovalbumin-expressing malaria strain results in more antigen specific T cell proliferation in the Il22−/− recipients.
The potential of IL-22 to influence the crosstalk between the microbiota and the immune system during non-inflammatory conditions in the mouse has been examined. Staphylococcus epidermidis was applied to the skin and using both gene-deleted mice and neutralizing antibody, the authors demonstrate IL-22 dependent upregulation of MHC class II expression on keratinocytes, which results in increased numbers of S. epidermidis specific Th1 in the skin but no difference was observed regarding Th17 cells (93). Genetic deletion of MHC class II specifically in keratinocytes reduces the Th1 cells but leaves Th17 cells unchanged. The authors speculate that this may be a result of differential requirements of Th1 and Th17 cells to local co-stimulation and chemokines produced by activated keratinocytes.
The role of IL-22 in viral infections is less studied compared to bacterial infections. A study reports that IL-22 is secreted in liver and lymphoid organs within the first few days after intravenous administration of lymphocytic choriomeningitis virus (LCMV) (94). Here IL-22 is expressed mainly by γδ T cells and is dependent on the PI3K/mTOR pathway but not AhR signaling (94). IL-22R was detected on CD45− cells in both the thymus and the spleen. Infecting Il22−/− mice with the LCMV results in increased expression in splenic CD4 T cells of activation marker CD44, chemokine receptor CXCR3, and proliferation marker ki67. Antigenic restimulation of splenic and liver T cells in vitro results in higher proportions of IFNγ+ CD4 and CD8 T cells. Overexpression of IL-22 leads to the opposite results, confirming that IL-22 dampens IFNγ+ T cell responses during acute (7 days) and persistent (60 days) LCMV infection in both lymphoid organs and the liver.
In a study explicitly designed to study potential indirect effects of IL-22 on T helper cell responses, mucosal immunization with ovalbumin and the adjuvant cholera toxin was performed (95). In Il22−/− mice this results in greater antigen specific T cell responses to mucosal (intrarectal), but not systemic (intraperitoneal) immunization. Polyclonal and antigen specific restimulation in vitro of splenic T cells from the mucosally immunized Il22−/− mice results in elevated secretion of IFNγ and IL-17, but no difference was seen after systemic immunization. In this case the proposed mechanism is increased epithelial permeability in the mucosal membrane in the absence of the homeostatic trophic effects of IL-22 allowing more antigen to come in contact with the immune system.
Circulating Th17 cells as well as serum levels of IL-22 and IL-17 are elevated in multiple myeloma patients compared to healthy controls, which has spurred investigations into the role of IL-22 in this disease. When peripheral blood mononuclear cells from multiple myeloma patients are cultured under Th1 polarizing conditions in the presence of IL-22 or IL-17, no effect on IFNγ was observed but when the two cytokines are combined IFNγ production is reduced (96). The authors suggest that the elevated circulating IL-22 and IL-17 in multiple myeloma patients may dampen Th1 responses potentially contributing to the observed immune dysfunction in this patient group.
IL-22 produced by local lymphoid tissue inducer (LTi) cells after radiation induced thymic injury is essential for the regeneration of thymopoiesis (97). In this disease model, the upregulation of IL-22 in the thymus is dependent on IL-23 from dendritic cells, primarily of the CD103+ subset, and may be triggered by the loss of double positive thymocytes. IL-22R subunit Il22ra1 is expressed on thymic epithelial cells and in vitro treatment of these cells with IL-22 results in improved survival and increased proliferation. Administration of IL-22 to irradiated mice with or without subsequent hematopoietic stem cell transplantation results in increased thymic cellularity, including all developing thymocyte subsets and thymic epithelial subsets.
IL-22 and Th22 cells are elevated in sera and tumor samples from patients with lung cancer and high IL22R1 expression is an indicator of poor prognosis in non-small cell lung cancer. In a study aimed at elucidating the role of IL-22 in tumor-promoting inflammation a Kras-induced mouse lung cancer model was used in combination with genetic deletion of Il22 (98). In the absence of IL-22 lung tumor burden is reduced. Characterization of the bronchoalveolar lavage fluid T cells showed reduced proportions of Tregs and increased proportions of IFNγ+ CD4 and CD8 T cells. IL-22 is known to induce STAT3 activation, which has also been observed in non-small cell lung cancer and is furthermore associated with poor prognosis, thus being a plausible mechanism for the effects on tumor burden in the mouse model. However, this would not explain the observed effects of IL-22 on T cell phenotype. The authors propose that pharmacologic targeting of IL-22 may have potential as an add-on therapy to conventional treatments of KRAS-mutant lung cancer.
Both adaptive and innate immune cells influence acute liver transplant rejection. IFNγ and IL-17 secretion is involved but the role of IL-22 is largely unknown. In a rat model of acute liver allograft rejection treatment with IL-22 neutralizing antibody 12 h before sacrificing the animal day 1, representing ischemia-reperfusion-injury, results in worse liver function (99). In contrast, treatment with IL-22 antibody 24 h before sacrificing the animal on day 7, representing acute rejection, results in improved liver function. At both timepoints IL-22 promoted expression of anti-apoptosis and pro-regeneration associated genes, implying that another mechanism overrides these effects day 7 when clinical outcome is better compared to controls. IL-22 neutralization is associated with increased proportions of Tregs and decreased proportions of Th17 cells in the liver allografts day 7 but no difference was observed day 1. The authors propose that at both timepoints the effect of IL-22 is mediated via STAT3. During the ischemia-reperfusion-injury stage the protective effect on hepatocytes by induction of anti-apoptotic and reparative factors lead to better clinical outcome but at the acute rejection stage day 7 induction of hepatocyte chemokine secretion and Th17 type inflammation dominate leading to worse clinical outcome.
Several studies have examined the role of IL-22 in GvHD. In a model of acute GvHD performed by injecting C57BL/6 splenocytes into F1 progeny of C57BL/6 and D2 mice, neutralizing IL-22 antibody was administered simultaneously and results in increased survival as well as suppressed expansion of donor CD8 T cells and decreased disease-associated depletion of host cells, particularly B cells (100). This is associated with increased proportions of Tregs in the spleen and decreased proportions of IFNγ, IL-4, and TNF secreting CD4 T cells. Splenic CD11b+ cells harvested from acute GvHD mice have upregulated IL-22R mRNA as well as protein and cells from IL-22 antibody treated mice have decreased expression of co-stimulatory molecules. Co-cultures with CD11b+ cells from IL-22 antibody treated mice with normal C57BL/6 CD4+ CD25− T cells promote Treg induction.
The role of IL-22 has also been investigated in a bone marrow transplantation model in which recipient mice were exposed to total body irradiation followed by injection of T cell depleted allogenic bone marrow (101). Treatment with recombinant IL-22 in this model accelerates thymic reconstitution. To induce GvHD the recipient mice were injected with allogenic T cells from the same donor mouse strain. Treatment with recombinant IL-22 has no effect on acute GvHD (day 7) but reduces severity of chronic GvHD (day 60), which is associated with increased numbers of natural Tregs in the thymus and the spleen and decreased numbers of Th1 cells in the spleen. No effect was seen on induced Tregs. The same group has shown using the same acute GvHD model with Il22−/− mice that recipient derived IL-22 reduces severity of disease and improves survival day 30 (102). The Il22−/− mice has higher proportions of Th1 cells in liver, spleen, and intestines. Tregs are reduced but no effect was seen on Th17 cells. IL-22R mRNA was detected in bone marrow derived dendritic cells and stimulation of these with recombinant IL-22 results in reduced expression of CD80 and IFNγ. Compared to wild type bone marrow derived dendritic cells Il22−/− counterparts co-cultured with wild type T cells results in higher proportions of Th1 cells and lower proportions of Tregs.
Published data that demonstrate direct or indirect effects of IL-22 on T cell polarization or secretion of signature cytokines in the context of a wide range of diseases are compiled in this review (Table 2, Figures 2 and 3). The observed effects are not uniform, but the data suggest that IL-22 signaling often results in reduced Th1 type responses and it may also contribute to resolution of inflammation by promoting Tregs or IL-10 secretion.
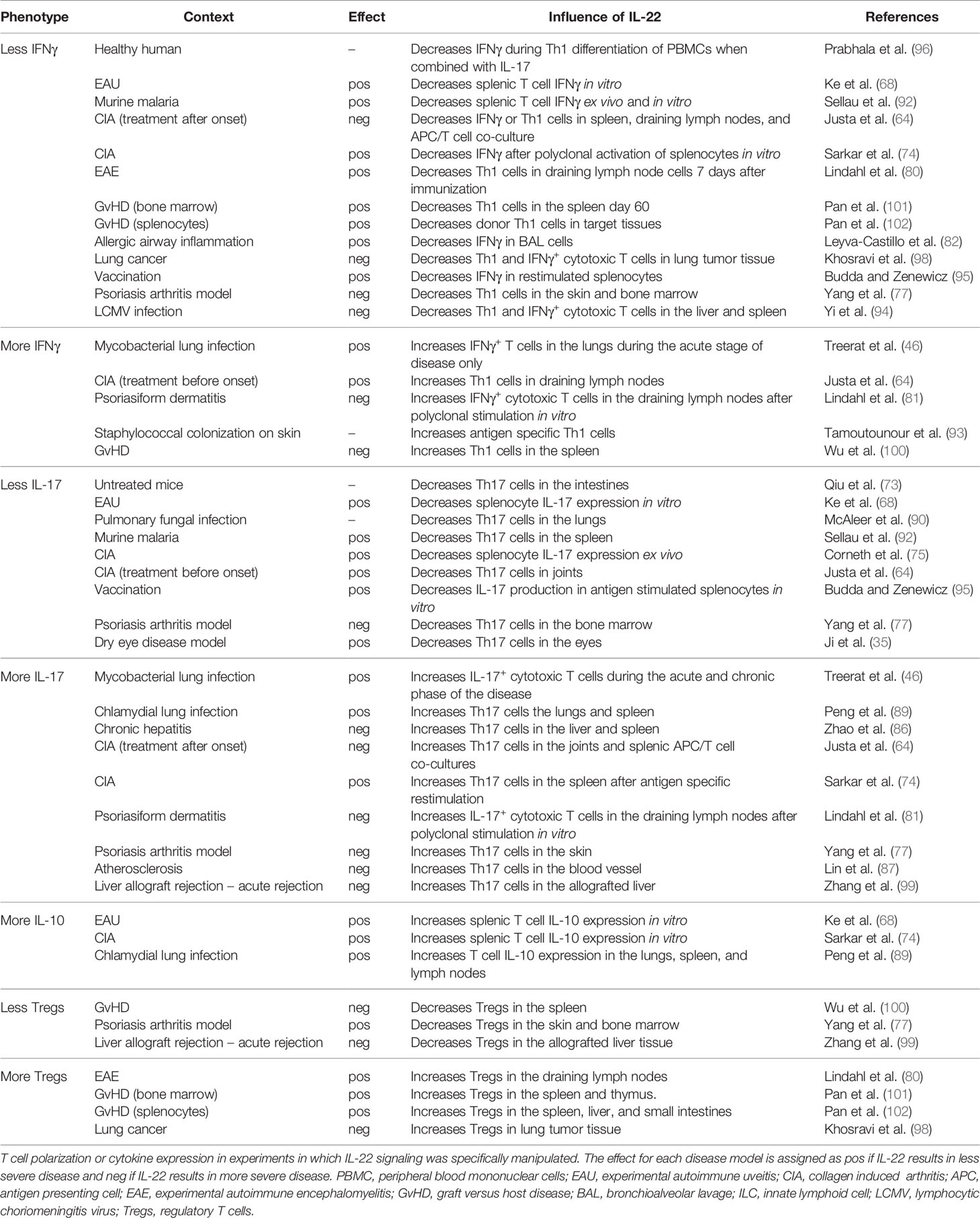
Table 2 The influence of IL-22 on the Th1/Th17 axis.
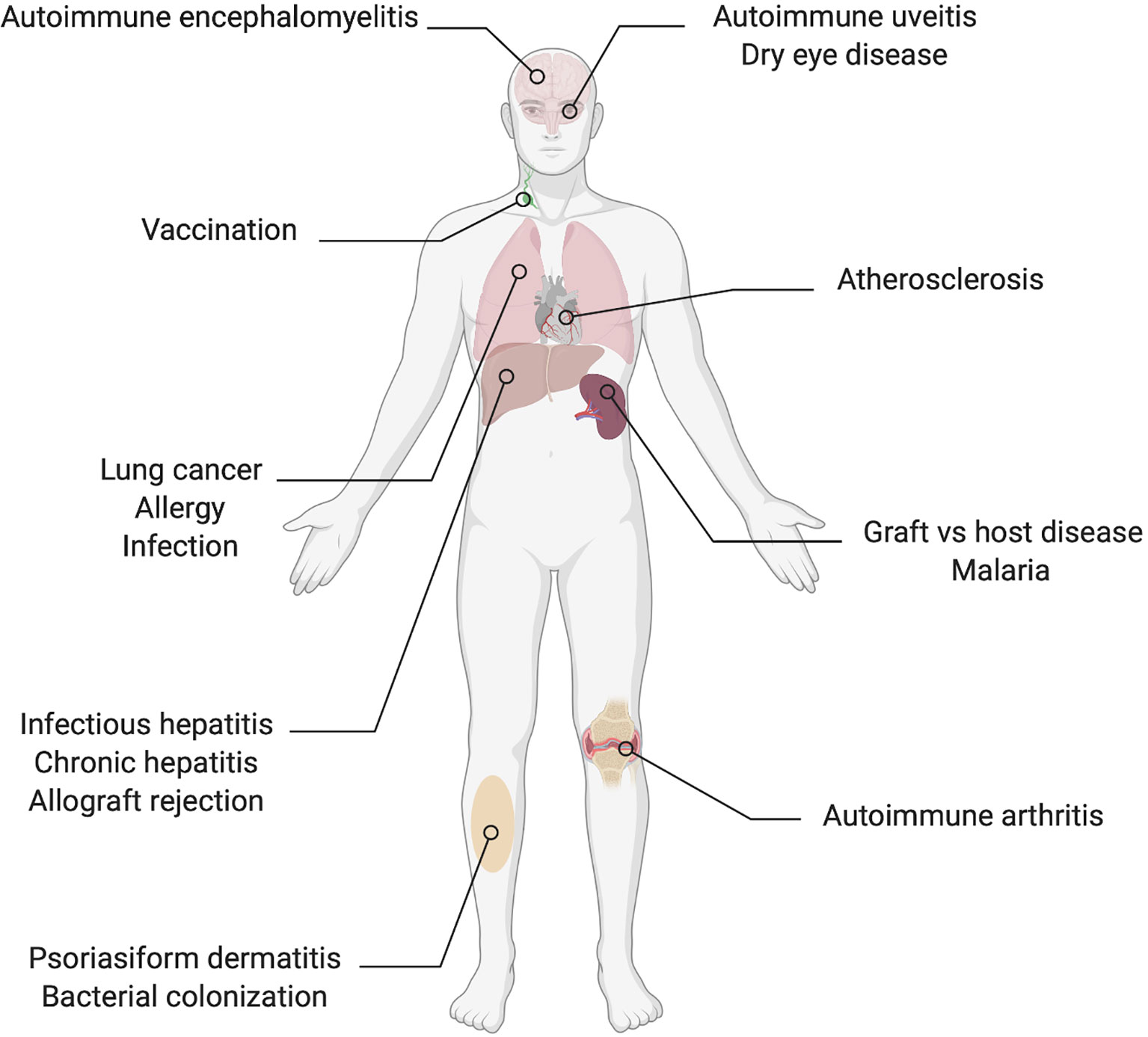
Figure 2 Organs and conditions in which there are reported effects of IL-22 on the Th1/Th17 axis.
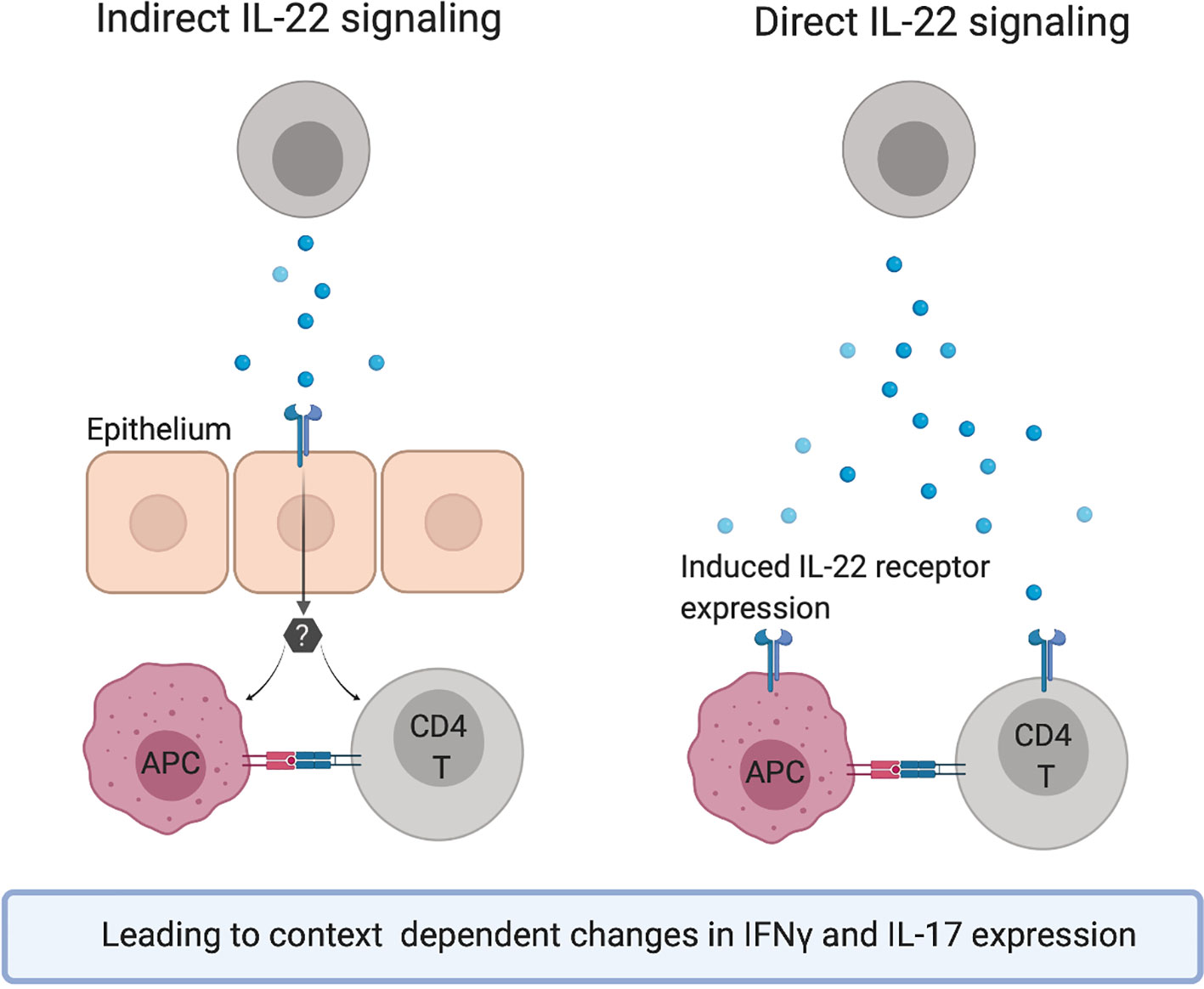
Figure 3 Hypothetical mechanisms of action for the observed effects of IL-22 on the Th1/Th17 axis.
IL-22 has an important role in the response to bacterial pathogens in several models of infectious diseases including Citrobacter rodentium colitis (30, 40) and Klebsiella pneumonia (103) as well as Mycobacterium tuberculosis infections of the lungs (46). The proposed mechanisms have primarily been maintenance of physiological barriers and induction of antimicrobial peptides. Considering this, it is somewhat surprising that available data frequently describe an inhibitory effect of IL-22 on Th1 immune responses. In contrast, if one considers the inflammatory diseases, an inhibitory effect of IL-22 on inflammatory T cell responses is consistent with the observed net effect of IL-22 on most models. The notable exception is psoriasis and other dermatitides where IL-22 has a pathogenic effect, which likely involves direct actions of IL-22 on keratinocytes leading to excessive proliferation, aberrant maturation as well as induction of inflammatory mediators (6, 11, 104–106).
Although not covered in this review, several studies have shown that IL-22 can promote B cell responses. CXCL13 is a central chemokine in B cell immune responses and is expressed in follicles of lymphoid tissues where it attracts B cells via the receptor CXCR5. In vivo neutralization of CXCL13 in mice reduces B cell recruitment to lymphoid follicles and inhibits formation of germinal centers (107). CXCL13 can be induced by IL-22 in tertiary lymphoid follicles (108) offering a plausible explanation to the elevated antibody levels in response to IL-22. In mice, IFNγ promotes isotype switching to IgG2a or IgG2c (109). The IL-22 mediated decrease in Th1 responses would thus be expected to be associated with a reduction in these isotypes which was the case in the study of Geboes et al. (66), but not in Justa et al. (64) or Corneth et al. (75). Although Th2 cells have not frequently been assessed in the included studies one can speculate that IL-22 induces a shift in the Th1/Th2 balance toward a Th2 and humoral immune response.
Both the proliferative and anti-apoptotic effects on epithelia and other tissues commonly attributed to IL-22 signaling as well as a potential inhibitory effect on T cell responses may lead to a permissive environment for tumor growth. IL-22 has been reported to both increase and decrease formation of tumors depending on tissue and model system but a tumor-promoting effect is more commonly observed (72). Interestingly, constitutive deletion of the IL-22 antagonist molecule IL-22BP in mice does not result in more tumor formation in steady state conditions but in the context of chronic colitis the unrestrained IL-22 signaling results in increased incidence of colon tumors. On the other hand, another study demonstrated a protective role of IL-22 in inflammation-induced colon tumors by improving the cellular response to DNA damage (110).
In summary, IL-22 signaling often reduces Th1 type immune responses. This characteristic of IL-22 may act in synergy with its protective effects on IL-22R expressing tissue cells to reduce collateral damage in the context of infection and inflammation. The long-term risk of tumor growth needs to be counterbalanced, which is the likely role of the endogenous IL-22 antagonist IL-22BP that is constitutively expressed in many tissues.
HL reviewed the literature and drafted the manuscript. TO revised the manuscript and approved the final version. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All figures were created with BioRender.com.
IL, interleukin; RA, rheumatoid arthritis; MS, multiple sclerosis; IL-22R, IL-22 receptor; NKT cell, natural killer T cell; ILC, innate lymphoid cell; γδ T cell, gamma delta T cell; DC, dendritic cell; AhR, aryl hydrocarbon receptor; RORγt, RAR-related orphan receptor gamma t; TLR, Toll-like receptor; TGF-β, transforming growth factor beta; ICOS, inducible T cell co-stimulator; IL-22BP, IL-22 binding protein; TNF, tumor necrosis factor; IFNγ, interferon gamma; MAIT cell, mucosa associated invariant T cell; LTi cell, lymphoid tissue inducer cell; EAU, experimental autoimmune uveitis; APC, antigen presenting cell; CIA, collagen induced arthritis; LCMV, lymphocytic choriomeningitis virus; GvHD, graft versus host disease.
1. Wolk K, Kunz S, Witte E, Friedrich M, Asadullah K, Sabat R. IL-22 increases the innate immunity of tissues. Immunity (2004) 21:241–54. doi: 10.1016/j.immuni.2004.07.007
2. Dudakov JA, Hanash AM, van den Brink MRM. Interleukin-22: immunobiology and pathology. Annu Rev Immunol (2015) 33:747–85. doi: 10.1146/annurev-immunol-032414-112123
3. Kreymborg K, Becher B. IL-22 vs. IL-22: The Tissue Matters. Open Autoimmun J (2010) 2:181–6. doi: 10.2174/1876894601002040181
4. Sabat R, Ouyang W, Wolk K. Therapeutic opportunities of the IL-22-IL-22R1 system. Nat Rev Drug Discovery (2014) 13:21–38. doi: 10.1038/nrd4176
5. Dumoutier L, Louahed J, Renauld JC. Cloning and characterization of IL-10-related T cell-derived inducible factor (IL-TIF), a novel cytokine structurally related to IL-10 and inducible by IL-9. J Immunol (2000) 164:1814–9. doi: 10.4049/jimmunol.164.4.1814
6. Liang SC, Tan X-Y, Luxenberg DP, Karim R, Dunussi-Joannopoulos K, Collins M, et al. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. J Exp Med (2006) 203:2271–9. doi: 10.1084/jem.20061308
7. Trifari S, Kaplan CD, Tran EH, Crellin NK, Spits H. Identification of a human helper T cell population that has abundant production of interleukin 22 and is distinct from T(H)-17, T(H)1 and T(H)2 cells. Nat Immunol (2009) 10:864–71. doi: 10.1038/ni.1770
8. Duhen T, Geiger R, Jarrossay D, Lanzavecchia A, Sallusto F. Production of interleukin 22 but not interleukin 17 by a subset of human skin-homing memory T cells. Nat Immunol (2009) 10:857–63. doi: 10.1038/ni.1767
9. Eyerich S, Eyerich K, Pennino D, Carbone T, Nasorri F, Pallotta S, et al. Th22 cells represent a distinct human T cell subset involved in epidermal immunity and remodeling. J Clin Invest (2009) 119:3573–85. doi: 10.1172/JCI40202
10. Wolk K, Kunz S, Asadullah K, Sabat R. Cutting Edge: Immune Cells as Sources and Targets of the IL-10 Family Members? J Immunol (2002) 168:5397–402. doi: 10.4049/jimmunol.168.11.5397
11. Zheng Y, Danilenko DM, Valdez P, Kasman I, Eastham-Anderson J, Wu J, et al. Interleukin-22, a T(H)17 cytokine, mediates IL-23-induced dermal inflammation and acanthosis. Nature (2007) 445:648–51. doi: 10.1038/nature05505
12. Chung Y, Yang X, Chang SH, Ma L, Tian Q, Dong C. Expression and regulation of IL-22 in the IL-17-producing CD4+ T lymphocytes. Cell Res (2006) 16:902–7. doi: 10.1038/sj.cr.7310106
13. Ivanov II, Atarashi K, Manel N, Brodie EL, Shima T, Karaoz U, et al. Induction of Intestinal Th17 Cells by Segmented Filamentous Bacteria. Cell (2009) 139:485–98. doi: 10.1016/j.cell.2009.09.033
14. Plank MW, Kaiko GE, Maltby S, Weaver J, Tay HL, Shen W, et al. Th22 Cells Form a Distinct Th Lineage from Th17 Cells In Vitro with Unique Transcriptional Properties and Tbet-Dependent Th1 Plasticity. J Immunol (2017) 198:2182–90. doi: 10.4049/jimmunol.1601480
15. Ortega C, Fernández-A S, Carrillo JM, Romero P, Molina IJ, Moreno JC, et al. IL-17-producing CD8+ T lymphocytes from psoriasis skin plaques are cytotoxic effector cells that secrete Th17-related cytokines. J Leukoc Biol (2009) 86:435–43. doi: 10.1189/JLB.0109046
16. Ciric B, El-behi M, Cabrera R, Zhang G, Rostami A. IL-23 drives pathogenic IL-17-producing CD8+ T cells. J Immunol (2009) 182:5296–305. doi: 10.4049/jimmunol.0900036
17. Billerbeck E, Kang Y, Walker L, Lockstone H, Grafmueller S, Fleming V, et al. Analysis of CD161 expression on human CD8+ T cells defines a distinct functional subset with tissue-homing properties. Proc Natl Acad Sci USA (2010) 107:3006–11. doi: 10.1073/pnas.0914839107
18. Sutton CE, Lalor SJ, Sweeney CM, Brereton CF, Lavelle EC, Mills KHG. Interleukin-1 and IL-23 Induce Innate IL-17 Production from gd T Cells, Amplifying Th17 Responses and Autoimmunity. Immunity (2009) 31:331–41. doi: 10.1016/j.immuni.2009.08.001
19. Martin B, Hirota K, Cua DJ, Stockinger B, Veldhoen M. Interleukin-17-producing gammadelta T cells selectively expand in response to pathogen products and environmental signals. Immunity (2009) 31:321–30. doi: 10.1016/j.immuni.2009.06.020
20. Gibbs A, Leeansyah E, Introini A, Paquin-Proulx D, Hasselrot K, Andersson E, et al. MAIT cells reside in the female genital mucosa and are biased towards IL-17 and IL-22 production in response to bacterial stimulation. Mucosal Immunol (2016) 10:1–11. doi: 10.1038/mi.2016.30
21. Goto M, Murakawa M, Kadoshima-Yamaoka K, Tanaka Y, Nagahira K, Fukuda Y, et al. Murine NKT cells produce Th17 cytokine interleukin-22. Cell Immunol (2009) 254:81–4. doi: 10.1016/j.cellimm.2008.10.002
22. Moreira-Teixeira L, Resende M, Coffre M, Devergne O, Herbeuval J, Hermine O, et al. Proinflammatory environment dictates the IL-17-producing capacity of human invariant NKT cells. J Immunol (2011) 186:5758–65. doi: 10.4049/jimmunol.1003043
23. Vonarbourg C, Mortha A, Bui VL, Hernandez PP, Kiss EA, Hoyler T, et al. Regulated expression of nuclear receptor RORγt confers distinct functional fates to NK cell receptor-expressing RORγt+ innate lymphocytes. Immunity (2010) 33:736–51. doi: 10.1016/j.immuni.2010.10.017
24. Geremia A, Arancibia-Cárcamo CV, Fleming MPP, Rust N, Singh B, Mortensen NJ, et al. IL-23–responsive innate lymphoid cells are increased in inflammatory bowel disease. J Exp Med (2011) 208:1127–33. doi: 10.1084/jem.20101712
25. Sonnenberg GF, Monticelli LA, Alenghat T, Fung TC, Hutnick NA, Kunisawa J, et al. Innate lymphoid cells promote anatomical containment of lymphoid-resident commensal bacteria. Science (2012) 336:1321–5. doi: 10.1126/science.1222551
26. Satpathy AT, Briseño CG, Lee JS, Ng D, Manieri NA, Kc W, et al. Notch2-dependent classical dendritic cells orchestrate intestinal immunity to attaching-and-effacing bacterial pathogens. Nat Immunol (2013) 14:937–48. doi: 10.1038/ni.2679
27. Takatori H, Kanno Y, Watford WT, Tato CM, Weiss G, Ivanov II, et al. Lymphoid tissue inducer-like cells are an innate source of IL-17 and IL-22. J Exp Med (2009) 206:35–41. doi: 10.1084/jem.20072713
28. Cupedo T, Crellin NK, Papazian N, Rombouts EJ, Weijer K, Grogan JL, et al. Human fetal lymphoid tissue-inducer cells are interleukin 17-producing precursors to RORC+ CD127+ natural killer-like cells. Nat Immunol (2009) 10:66–74. doi: 10.1038/ni.1668
29. Res PCM, Piskin G, de Boer OJ, van der Loos CM, Teeling P, Bos JD, et al. Overrepresentation of IL-17A and IL-22 producing CD8 T cells in lesional skin suggests their involvement in the pathogenesis of psoriasis. PloS One (2010) 5:e14108. doi: 10.1371/journal.pone.0014108
30. Zheng Y, Valdez PA, Danilenko DM, Hu Y, Sa SM, Gong, et al. Interleukin-22 mediates early host defense against attaching and effacing bacterial pathogens. Nat Med (2008) 14:282–9. doi: 10.1038/nm1720
31. Fumagalli S, Torri A, Papagna A, Citterio S, Mainoldi F, Foti M. IL-22 is rapidly induced by Pathogen Recognition Receptors Stimulation in Bone-Marrow-derived Dendritic Cells in the Absence of IL-23. Sci Rep (2016) 6:33900. doi: 10.1038/srep33900
32. Chen F, Cao A, Yao S, Evans-Marin HL, Liu H, Wu W, et al. mTOR Mediates IL-23 Induction of Neutrophil IL-17 and IL-22 Production. J Immunol (2016) 196:4390–9. doi: 10.4049/jimmunol.1501541
33. Zindl CL, Lai J, Lee YK, Maynard CL, Harbour SN, Ouyang W, et al. IL-22-producing neutrophils contribute to antimicrobial defense and restitution of colonic epithelial integrity during colitis. Proc Natl Acad Sci USA (2013) 110:12768–73. doi: 10.1073/pnas.1300318110
34. Mashiko S, Bouguermouh S, Rubio M, Baba N, Bissonnette R, Sarfati M. Human mast cells are major IL-22 producers in patients with psoriasis and atopic dermatitis. J Allergy Clin Immunol (2015) 136:351–9.e1. doi: 10.1016/j.jaci.2015.01.033
35. Ji YW, Mittal SK, Hwang HS, Chang E, Lee JH, Seo, et al. Lacrimal gland–derived IL-22 regulates IL-17-mediated ocular mucosal inflammation. Mucosal Immunol (2017) 10:1202–10. doi: 10.1038/mi.2016.119
36. Ikeuchi H, Kuroiwa T, Hiramatsu N, Kaneko Y, Hiromura K, Ueki K, et al. Expression of interleukin-22 in rheumatoid arthritis: potential role as a proinflammatory cytokine. Arthritis Rheum (2005) 52:1037–46. doi: 10.1002/art.20965
37. Cavaillon B, Marie JC, Couillin I, Blanchet K, Zitvogel L, Ryffel B, et al. Cutting Edge: Crucial Role of IL-1 and IL-23 in the Innate IL-17 Response of Peripheral Lymph Node NK1.1 2 Invariant NKT Cells to Bacteria. J Immunol (2011) 186:662–6. doi: 10.4049/jimmunol.1002725
38. Sonnenberg GF, Monticelli LA, Elloso MM, Fouser LA, Artis D. CD4(+) lymphoid tissue-inducer cells promote innate immunity in the gut. Immunity (2011) 34:122–34. doi: 10.1016/j.immuni.2010.12.009
39. Lee Y, Kumagai Y, Jang MS, Kim J, Yang B, Lee E, et al. Intestinal Lin- c-Kit+ NKp46- CD4- population strongly produces IL-22 upon IL-1β stimulation. J Immunol (2013) 190:5296–305. doi: 10.4049/jimmunol.1201452
40. Qiu J, Heller JJ, Guo X, Chen ZME, Fish K, Fu Y-X, et al. The Aryl Hydrocarbon Receptor Regulates Gut Immunity through Modulation of Innate Lymphoid Cells. Immunity (2012) 36:92–104. doi: 10.1016/j.immuni.2011.11.011
41. Veldhoen M, Hirota K, Westendorf AM, Buer J, Dumoutier L, Renauld J-C, et al. The aryl hydrocarbon receptor links TH17-cell-mediated autoimmunity to environmental toxins. Nature (2008) 453:106–9. doi: 10.1038/nature06881
42. Zelante T, Iannitti RG, Cunha C, DeLuca A, Giovannini G, Pieraccini G, et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity (2013) 39:372–85. doi: 10.1016/j.immuni.2013.08.003
43. Lowe MM, Mold JE, Kanwar B, Huang Y, Louie A, Pollastri MP, et al. Identification of cinnabarinic acid as a novel endogenous aryl hydrocarbon receptor ligand that drives IL-22 production. PloS One (2014) 3(9):e87877. doi: 10.1371/journal.pone.0087877
44. Nurieva R, Yang XO, Martinez G, Zhang Y, Panopoulos AD, Ma L, et al. Essential autocrine regulation by IL-21 in the generation of inflammatory T cells. Nature (2007) 448:480–3. doi: 10.1038/nature05969
45. Crellin NK, Trifari S, Kaplan CD, Satoh-Takayama N, Di Santo JP, Spits H. Regulation of cytokine secretion in human CD127+ LTi-like innate lymphoid cells by toll-like receptor 2. Immunity (2010) 33:752–64. doi: 10.1016/j.immuni.2010.10.012
46. Treerat P, Prince O, Cruz-Lagunas A, Muñoz-Torrico M, Salazar-Lezama MA, Selman M, et al. Novel role for IL-22 in protection during chronic Mycobacterium tuberculosis HN878 infection. Mucosal Immunol (2017) 10:1069–81. doi: 10.1038/mi.2017.15
47. Rutz S, Noubade R, Eidenschenk C, Ota N, Zeng W, Zheng Y, et al. Transcription factor c-Maf mediates the TGF-β-dependent suppression of IL-22 production in TH17 cells. Nat Immunol (2011) 12:1238–45. doi: 10.1038/ni.2134
48. Liu H, Rohowsky-Kochan C. Interleukin-27-mediated suppression of human Th17 cells is associated with activation of STAT1 and suppressor of cytokine signaling protein 1. J Interferon Cytokine Res (2011) 31:459–69. doi: 10.1089/jir.2010.0115
49. Paulos CM, Carpenito C, Plesa G, Suhoski MM, Varela-Rohena A, Golovina TN, et al. The inducible costimulator (ICOS) is critical for the development of human T(H)17 cells. Sci Transl Med (2010) 2:55ra78. doi: 10.1126/scitranslmed.3000448
50. Xu W, Presnell SR, Parrish-Novak J, Kindsvogel W, Jaspers S, Chen Z, et al. A soluble class II cytokine receptor, IL-22RA2, is a naturally occurring IL-22 antagonist. Proc Natl Acad Sci USA (2001) 98:9511–6. doi: 10.1073/pnas.171303198
51. Dumoutier L, Lejeune D, Colau D, Renauld JC. Cloning and characterization of IL-22 binding protein, a natural antagonist of IL-10-related T cell-derived inducible factor/IL-22. J Immunol (2001) 166:7090–5. doi: 10.4049/jimmunol.166.12.7090
52. Dumoutier L, Van Roost E, Colau D, Renauld JC. Human interleukin-10-related T cell-derived inducible factor: molecular cloning and functional characterization as an hepatocyte-stimulating factor. Proc Natl Acad Sci USA (2000) 97:10144–9. doi: 10.1073/pnas.170291697
53. Xie MH, Aggarwal S, Ho WH, Foster J, Zhang Z, Stinson J, et al. Interleukin (IL)-22, a novel human cytokine that signals through the interferon receptor-related proteins CRF2-4 and IL-22R. J Biol Chem (2000) 275:31335–9. doi: 10.1074/jbc.M005304200
54. Li J, Tomkinson KN, Tan X-Y, Wu P, Yan G, Spaulding V, et al. Temporal associations between interleukin 22 and the extracellular domains of IL-22R and IL-10R2. Int Immunopharmacol (2004) 4:693–708. doi: 10.1016/j.intimp.2004.01.010
55. Kotenko SV, Krause CD, Izotova LS, Pollack BP, Wu W, Pestka S. Identification and functional characterization of a second chain of the interleukin-10 receptor complex. EMBO J (1997) 16:5894–903. doi: 10.1093/emboj/16.19.5894
56. Jones BC, Logsdon NJ, Walter MR. Structure of IL-22 bound to its high-affinity IL-22R1 chain. Structure (2008) 16:1333–44. doi: 10.1016/j.str.2008.06.005
57. Lejeune D, Dumoutier L, Constantinescu S, Kruijer W, Schuringa JJ, Renauld J-CC. Interleukin-22 (IL-22) activates the JAK/STAT, ERK, JNK, and p38 MAP kinase pathways in a rat hepatoma cell line. Pathways that are shared with and distinct from IL-10. J Biol Chem (2002) 277:33676–82. doi: 10.1074/jbc.M204204200
58. Andoh A, Zhang Z, Inatomi O, Fujino S, Deguchi Y, Araki Y, et al. Interleukin-22, a member of the IL-10 subfamily, induces inflammatory responses in colonic subepithelial myofibroblasts. Gastroenterology (2005) 129:969–84. doi: 10.1053/j.gastro.2005.06.071
59. Mitra A, Raychaudhuri SK, Raychaudhuri SP. IL-22 induced cell proliferation is regulated by PI3K/Akt/mTOR signaling cascade. Cytokine (2012) 60:38–42. doi: 10.1016/j.cyto.2012.06.316
60. Wolk K, Witte E, Wallace E, Döcke W-DD, Kunz S, Asadullah K, et al. IL-22 regulates the expression of genes responsible for antimicrobial defense, cellular differentiation, and mobility in keratinocytes: a potential role in psoriasis. Eur J Immunol (2006) 36:1309–23. doi: 10.1002/eji.200535503
61. Radaeva S, Sun R, Pan H-N, Hong F, Gao B. Interleukin 22 (IL-22) plays a protective role in T cell-mediated murine hepatitis: IL-22 is a survival factor for hepatocytes via STAT3 activation. Hepatology (2004) 39:1332–42. doi: 10.1002/hep.20184
62. Aggarwal S, Xie MH, Maruoka M, Foster J, Gurney AL. Acinar cells of the pancreas are a target of interleukin-22. J Interferon Cytokine Res (2001) 21:1047–53. doi: 10.1089/107999001317205178
63. Xu M-J, Feng D, Wang H, Guan Y, Yan X, Gao B. IL-22 Ameliorates Renal Ischemia-Reperfusion Injury by Targeting Proximal Tubule Epithelium. J Am Soc Nephrol (2014) 25:967–77. doi: 10.1681/ASN.2013060611
64. Justa S, Zhou X, Sarkar S. Endogenous IL-22 plays a dual role in arthritis: regulation of established arthritis via IFN-γ responses. PloS One (2014) 9:e93279. doi: 10.1371/journal.pone.0093279
65. Sarkar S, Zhou X, Justa S, Bommireddy SR. IL-22 reduces the severity of collagen arthritis in association with increased levels of IL-10. Arthritis Rheum (2013) 65(4):960–71. doi: 10.1002/art.37849
66. Geboes L, Dumoutier L, Kelchtermans H, Schurgers E, Mitera T, Renauld J-C, et al. Proinflammatory role of the Th17 cytokine interleukin-22 in collagen-induced arthritis in C57BL/6 mice. Arthritis Rheum (2009) 60:390–5. doi: 10.1002/art.24220
67. Pfeifle R, Rothe T, Ipseiz N, Scherer HU, Culemann S, Harre U, et al. Regulation of autoantibody activity by the IL-23-TH17 axis determines the onset of autoimmune disease. Nat Immunol (2017) 18:104–13. doi: 10.1038/ni.3579
68. Ke Y, Sun D, Jiang G, Kaplan HJ, Shao H. IL-22-induced regulatory CD11b+ APCs suppress experimental autoimmune uveitis. J Immunol (2011) 187:2130–9. doi: 10.4049/jimmunol.1100482
69. Ciccia F, Guggino G, Rizzo A, Bombardieri M, Raimondo S, Carubbi F, et al. Interleukin (IL)-22 receptor 1 is over-expressed in primary Sjögren’s syndrome and Sjögren-associated non-Hodgkin lymphomas and is regulated by IL-18. Clin Exp Immunol (2015) 181:219–29. doi: 10.1111/cei.12643
70. Dhiman R, Venkatasubramanian S, Paidipally P, Barnes PF, Tvinnereim A. Interleukin 22 Inhibits Intracellular Growth of Mycobacterium tuberculosis by Enhancing Calgranulin A Expression. J Infect Dis (2014) 209(4):578–87. doi: 10.1093/infdis/jit495
71. Dalmas E, Venteclef N, Caer C, Poitou C, Cremer I, Aron-Wisnewsky J, et al. T cell-derived IL-22 amplifies IL-1β-driven inflammation in human adipose tissue: relevance to obesity and type 2 diabetes. Diabetes (2014) 63:1966–77. doi: 10.2337/db13-1511
72. Hernandez P, Gronke K, Diefenbach A. A catch-22: Interleukin-22 and cancer. Eur J Immunol (2018) 48:15–31. doi: 10.1002/eji.201747183
73. Qiu J, Guo X, Chen ZE, He L, Sonnenberg GF, Artis D, et al. Group 3 innate lymphoid cells inhibit T-cell-mediated intestinal inflammation through aryl hydrocarbon receptor signaling and regulation of microflora. Immunity (2013) 39:386–99. doi: 10.1016/j.immuni.2013.08.002
74. Sarkar S, Zhou X, Justa S, Bommireddy SR. Interleukin-22 reduces the severity of collagen-induced arthritis in association with increased levels of interleukin-10. Arthritis Rheum (2013) 65:960–71. doi: 10.1002/art.37849
75. Corneth OBJ, Reijmers RM, Mus AMC, Asmawidjaja PS, van Hamburg JP, Papazian N, et al. Loss of IL-22 inhibits autoantibody formation in collagen-induced arthritis in mice. Eur J Immunol (2016) 46:1404–14. doi: 10.1002/eji.201546241
76. Karczewski J, Dobrowolska A, Rychlewska-Hańczewska A, Adamski Z. New insights into the role of T cells in pathogenesis of psoriasis and psoriatic arthritis. Autoimmunity (2016) 49:435–50. doi: 10.3109/08916934.2016.1166214
77. Yang L, Fanok MH, Mediero-Munoz A, Fogli LK, Corciulo C, Abdollahi S, et al. Augmented Th17 Differentiation Leads to Cutaneous and Synovio-Entheseal Inflammation in a Novel Model of Psoriatic Arthritis. Arthritis Rheumatol (Hoboken NJ) (2018) 70:855–67. doi: 10.1002/art.40447
78. Kreymborg K, Etzensperger R, Dumoutier L, Haak S, Rebollo A, Buch T, et al. IL-22 is expressed by Th17 cells in an IL-23-dependent fashion, but not required for the development of autoimmune encephalomyelitis. J Immunol (2007) 179:8098–104. doi: 10.4049/jimmunol.179.12.8098
79. Laaksonen H, Guerreiro-Cacais AO, Adzemovic MZ, Parsa R, Zeitelhofer M, Jagodic M, et al. The multiple sclerosis risk gene IL22RA2 contributes to a more severe murine autoimmune neuroinflammation. Genes Immun (2014) 15:457–65. doi: 10.1038/gene.2014.36
80. Lindahl H, Guerreiro-Cacais AO, Bedri SK, Linnerbauer M, Lindén M, Abdelmagid N, et al. IL-22 Binding Protein Promotes the Disease Process in Multiple Sclerosis. J Immunol (2019) 203:888–98. doi: 10.4049/jimmunol.1900400
81. Lindahl H, Martini E, Brauner S, Nikamo P, Gallais Serezal I, Guerreiro-Cacais AO, et al. IL-22 binding protein regulates murine skin inflammation. Exp Dermatol (2017) 26:444–6. doi: 10.1111/exd.13225
82. Leyva-Castillo JM, Yoon J, Geha RS. IL-22 promotes allergic airway inflammation in epicutaneously sensitized mice. J Allergy Clin Immunol (2019) 143:619–630.e7. doi: 10.1016/j.jaci.2018.05.032
83. Zenewicz LA, Yancopoulos GD, Valenzuela DM, Murphy AJ, Karow M, Flavell RA. Interleukin-22 but not interleukin-17 provides protection to hepatocytes during acute liver inflammation. Immunity (2007) 27:647–59. doi: 10.1016/j.immuni.2007.07.023
84. Ren X, Hu B, Colletti LM. IL-22 is involved in liver regeneration after hepatectomy. Am J Physiol Gastrointest Liver Physiol (2010) 298:G74–80. doi: 10.1152/ajpgi.00075.2009
85. Ki SH, Park O, Zheng M, Morales-Ibanez O, Kolls JK, Bataller R, et al. Interleukin-22 treatment ameliorates alcoholic liver injury in a murine model of chronic-binge ethanol feeding: role of signal transducer and activator of transcription 3. Hepatology (2010) 52:1291–300. doi: 10.1002/hep.23837
86. Zhao J, Zhang Z, Luan Y, Zou Z, Sun Y, Li Y, et al. Pathological Functions of Interleukin-22 in Chronic Liver Inflammation and Fibrosis With Hepatitis B Virus Infection by Promoting T Helper 17 Cell Recruitment. Hepatology (2013) 59:1331–42. doi: 10.1002/hep.26916
87. Lin YZ, Wu BW, De L, Huang Y, Shi Y, Liu H, et al. Circulating Th22 and Th9 levels in patients with acute coronary syndrome. Mediators Inflammation (2013) 2013:635672. doi: 10.1155/2013/635672
88. Shi L, Ji Q, Liu L, Shi Y, Lu Z, Ye J, et al. IL-22 produced by Th22 cells aggravates atherosclerosis development in ApoE–/– mice by enhancing DC-induced Th17 cell proliferation. J Cell Mol Med (2020) 24:3064–78. doi: 10.1111/jcmm.14967
89. Peng Y, Gao X, Yang J, Shekhar S, Wang S, Fan Y, et al. Interleukin-22 promotes T helper 1 (Th1)/Th17 immunity in chlamydial lung infection. Mol Med (2014) 20:109–19. doi: 10.2119/molmed.2013.00115
90. McAleer JP, Nguyen NLH, Chen K, Kumar P, Ricks DM, Binnie M, et al. Pulmonary Th17 Antifungal Immunity Is Regulated by the Gut Microbiome. J Immunol (2016) 197:97–107. doi: 10.4049/jimmunol.1502566
91. Gopal R, Monin L, Slight S, Uche U, Blanchard E, Junecko BAF, et al. Unexpected Role for IL-17 in Protective Immunity against Hypervirulent Mycobacterium tuberculosis HN878 Infection. PLos Pathog (2014) 10(5):e1004099. doi: 10.1371/journal.ppat.1004099
92. Sellau J, Alvarado CF, Hoenow S, Mackroth MS, Kleinschmidt D, Huber S, et al. IL-22 dampens the T cell response in experimental malaria. Sci Rep (2016) 6:28058. doi: 10.1038/srep28058
93. Tamoutounour S, Han SJ, Deckers J, Constantinides MG, Hurabielle C, Harrison OJ, et al. Keratinocyte-intrinsic MHCII expression controls microbiota-induced Th1 cell responses. Proc Natl Acad Sci USA (2019) 116:23643–52. doi: 10.1073/pnas.1912432116
94. Yi P, Liang Y, Yuan DMK, Jie Z, Kwota Z, Chen Y, et al. A tightly regulated IL-22 response maintains immune functions and homeostasis in systemic viral infection. Sci Rep (2017) 7:3857. doi: 10.1038/s41598-017-04260-0
95. Budda SA, Zenewicz LA. IL-22 deficiency increases CD4 T cell responses to mucosal immunization. Vaccine (2018) 36:3694–700. doi: 10.1016/j.vaccine.2018.05.011
96. Prabhala RH, Pelluru D, Fulciniti M, Prabhala HK, Nanjappa P, Song W, et al. Elevated IL-17 produced by T H 17 cells promotes myeloma cell growth and inhibits immune function in multiple myeloma. Blood (2010) 115:5385–92. doi: 10.1182/blood-2009-10-246660
97. Dudakov JA, Hanash AM, Jenq RR, Young LF, Ghosh A, Singer NV. Interleukin-22 drives endogenous thymic regeneration in mice. Science (2012) 336:91–5. doi: 10.1126/science.1218004
98. Khosravi N, Caetano MS, Cumpian AM, Unver N, De la Garza Ramos C, Noble O, et al. IL22 promotes kras-mutant lung cancer by induction of a protumor immune response and protection of stemness properties. Cancer Immunol Res (2018) 6:788–97. doi: 10.1158/2326-6066.CIR-17-0655
99. Zhang Y, Wang X, Mao L, Yang D, Gao W, Tian Z, et al. Dual roles of IL-22 at ischemia-reperfusion injury and acute rejection stages of rat allograft liver transplantation. Oncotarget (2017) 8:115384–97. doi: 10.18632/oncotarget.23266
100. Wu J, Gu J, Zhou S, Lu H, Lu Y, Lu L, et al. Anti-IL-22 antibody attenuates acute graft-versus-host disease via increasing Foxp3+ T cell through modulation of CD11b+ cell function. J Immunol Res (2018) 2018:1605341. doi: 10.1155/2018/1605341
101. Pan B, Wang D, Li L, Shang L, Xia F, Zhang F, et al. IL-22 Accelerates Thymus Regeneration via Stat3/Mcl-1 and Decreases Chronic Graft-versus-Host Disease in Mice after Allotransplants. Biol Blood Marrow Transplant (2019) 25:1911–9. doi: 10.1016/j.bbmt.2019.06.002
102. Pan B, Xia F, Wu Y, Zhang F, Lu Z, Fu R, et al. Recipient-derived IL-22 alleviates murine acute graft-versus-host disease in association with reduced activation of antigen presenting cells. Cytokine (2018) 111:33–40. doi: 10.1016/j.cyto.2018.08.010
103. Aujla SJ, Chan YR, Zheng M, Fei M, Askew DJ, Pociask DA, et al. IL-22 mediates mucosal host defense against Gram-negative bacterial pneumonia. Nat Med (2008) 14:275–81. doi: 10.1038/nm1710
104. Boniface K, Guignouard E, Pedretti N, Garcia M, Delwail A, Bernard F-X, et al. A role for T cell-derived interleukin 22 in psoriatic skin inflammation. Clin Exp Immunol (2007) 150:407–15. doi: 10.1111/j.1365-2249.2007.03511.x
105. Wolk K, Haugen HS, Xu W, Witte E, Waggie K, Anderson M, et al. IL-22 and IL-20 are key mediators of the epidermal alterations in psoriasis while IL-17 and IFN-gamma are not. J Mol Med (Berl) (2009) 87:523–36. doi: 10.1007/s00109-009-0457-0
106. Ma H-L, Liang S, Li J, Napierata L, Brown T, Benoit S, et al. IL-22 is required for Th17 cell-mediated pathology in a mouse model of psoriasis-like skin inflammation. J Clin Invest (2008) 118:597–607. doi: 10.1172/JCI33263
107. Klimatcheva E, Pandina T, Reilly C, Torno S, Bussler H, Scrivens M, et al. CXCL13 antibody for the treatment of autoimmune disorders. BMC Immunol (2015) 16:6. doi: 10.1186/s12865-015-0068-1
108. Barone F, Nayar S, Campos J, Cloake T, Withers DR, Toellner K, et al. IL-22 regulates lymphoid chemokine production and assembly of tertiary lymphoid organs. Proc Natl Acad Sci USA (2015) 112:11024–9. doi: 10.1073/pnas.1503315112
109. Snapper CM, Paul WE. Interferon-gamma and B cell stimulatory factor-1 reciprocally regulate Ig isotype production. Science (1987) 236:944–7. doi: 10.1126/science.3107127
Keywords: interleukin 22 (IL-22), interferon gamma, interleukin 17 (IL-17), animal models, inflammatory disease, infectious disease
Citation: Lindahl H and Olsson T (2021) Interleukin-22 Influences the Th1/Th17 Axis. Front. Immunol. 12:618110. doi: 10.3389/fimmu.2021.618110
Received: 16 October 2020; Accepted: 07 January 2021;
Published: 22 February 2021.
Edited by:
Lars Rogge, Institut Pasteur, FranceReviewed by:
Angela Bonura, National Research Council (CNR), ItalyCopyright © 2021 Lindahl and Olsson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hannes Lindahl, aGFubmVzLmxpbmRhaGxAa2kuc2U=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.