 Jordana Batista Santana1Tarcísio Vila Verde Santana de Almeida1
Jordana Batista Santana1Tarcísio Vila Verde Santana de Almeida1 Diego Mota Lopes1
Diego Mota Lopes1 Brady Page2
Brady Page2 Sergio Costa Oliveira3,4Irismá Souza5Luís Eduardo Viana Silva Ribeiro1
Sergio Costa Oliveira3,4Irismá Souza5Luís Eduardo Viana Silva Ribeiro1 Néstor Adrián Guerrero Gutiérrez1
Néstor Adrián Guerrero Gutiérrez1 Edgar M. Carvalho1,4,6
Edgar M. Carvalho1,4,6 Luciana Santos Cardoso1,4,7*
Luciana Santos Cardoso1,4,7*- 1 Serviço de Imunologia, Hospital Universitário Professor Edgard Santos, Universidade Federal da Bahia, Salvador, Brazil
- 2Massachusetts General Hospital, Boston, MA, United States
- 3Departamento de Bioquímica e Imunologia, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil
- 4Instituto Nacional de Ciência e Tecnologia em Doenças Tropicais (INCT–DT/CNPq), Salvador, Brazil
- 5Instituto de Saúde Irismá Souza, Gandu, Brazil
- 6Laboratório de Pesquisas Clínicas, Instituto Gonçalo Moniz, Fundação Oswaldo Cruz (FIOCRUZ), Salvador, Brazil
- 7Departamento de Análises Clínicas e Toxicológicas, Faculdade de Farmácia, Universidade Federal da Bahia (UFBA), Salvador, Brazil
Schistosomiasis is a parasitic disease that affects about 166 million people around the world. It is estimated that 5%–10% of individuals with schistosomiasis develop severe forms of the disease, which are characterized by pulmonary hypertension, ascites, periportal fibrosis, and other significant complications. The chronic phase of the disease is associated with a Th2 type immune response, but evidence also suggests there are roles for Th1 and Th17 in the development of severe disease. The aim of this study was to evaluate the CD4+ T lymphocyte profile of patients with different degrees of periportal fibrosis secondary to schistosomiasis. These individuals had been treated for schistosomiasis, but since they live in a S. mansoni endemic area, they are at risk of reinfection. They were evaluated in relation to the degree of periportal fibrosis and classified into three groups: without fibrosis or with incipient fibrosis (WF/IFNE), n=12, possible periportal fibrosis/periportal fibrosis, n=13, and advanced periportal fibrosis/advanced periportal fibrosis with portal hypertension, n=4. We observed in the group without fibrosis a balance between the low expression of Th2 cytokines and high expression of T reg cells. As has already been described in the literature, we found an increase of the Th2 cytokines IL-4, IL-5, and IL-13 in the group with periportal fibrosis. In addition, this group showed higher expression of IL-17 and IL-10 but lower IL-10/IL-13 ratio than patients in the WF/IFNE group. Cells from individuals who present any level of fibrosis expressed more TGF-β compared to the WF/IFNE group and a positive correlation with left lobe enlargement and portal vein wall thickness. There was a negative correlation between IL-17 and the thickness of the portal vein wall, but more studies are necessary in order to explore the possible protective role of this cytokine. Despite the fibrosis group having presented a higher expression of pro-fibrotic molecules compared to WF/IFNE patients, it seems there is a regulation through IL-10 and T reg cells that is able to maintain the low morbidity of this group.
Introduction
Among parasitic diseases in tropical and subtropical regions, schistosomiasis is the second most important in terms of socioeconomic and public health impact. It affects about 240 million people living in developing countries, especially in rural and peri-urban areas, with an estimated 700 million people at risk worldwide (1).
It is estimated that 5%–10% of individuals infected with S. mansoni develop severe forms of the disease, which can be characterized by hepatic fibrosis and portal hypertension, ascites, as well as esophageal and gastric varices, which predispose infected individuals to gastrointestinal hemorrhage and death (2, 3). Hepatic fibrosis occurs in response to antigens present on the ova of the parasite, which become lodged in the second-order periportal venous branches, precipitating an inflammatory reaction that leads to granuloma formation with eventual tissue fibrosis (4, 5).
The immunopathogenesis of the schistosomal granuloma is predominantly Th2, characterized by the production of IL-4, IL-5, and IL-13, as reviewed by Wilson et al. (5, 6). There are few studies in the literature showing the involvement of Th1 and Th17 cytokines in the pathogenesis of fibrosis. These studies point to a dual role of IFN-γ cytokine in both pathogenesis and fibrosis protection and focus primarily on experimental models and serum levels of these cytokines in vitro (7–9). Little is known about the protective immune response in periportal fibrosis in humans (10). Unlike the results found in a murine model, where IL-10 plays a key role in controlling the inflammatory response in fibrosis (11), in vitro studies evaluating the frequency of monocytes from individuals with different degrees of periportal fibrosis do not show an increase in IL-10 production (12). The same result was shown when levels of IL-10 in serum from individuals with different degrees of periportal fibrosis were analyzed (13–15). These results suggest that there are other regulatory sources of the inflammatory process observed in the pathogenesis of fibrosis besides IL-10.
The aim of this study was to characterize the profile of TCD4+ lymphocytes expressing Th1, Th2, Th17 cytokines and molecules associated with a regulatory response by individuals with different degrees of periportal fibrosis secondary to schistosomiasis. The identification of a phenotypic profile in peripheral blood lymphocytes of individuals with periportal fibrosis could help in the discovery of immunomodulatory molecules capable of controlling the exacerbation of the fibrosis-associated inflammatory process. The results obtained here open the field for interventions with potential immunomodulatory drugs.
Methods
Selection of Participants in the Schistosomiasis-Endemic Region and Study Design
The study was conducted in a small village of Água Preta in the state of Bahia, Brazil. The sanitation conditions of the region are precarious, placing residents at high risk of parasitic infection. The main source of income within the population is agriculture, and river water is used for bathing, washing clothes and utensils, and leisure activities. The population’s access to health services is limited; there are only reports of sporadic previous treatments with anthelmintics.
In this study we included 29 patients with different degrees of periportal fibrosis due to Schistosoma mansoni infection, classified using the WHO-Niamey protocol by a trained physician. Interpretation of ultrasonographic results was performed by the combination of numerical results of IP (Image Pattern) scores, PT (Periportal Thickening) scores, and PH (Portal Hypertension).
The examination was carried out by a trained ultrasound specialist. The evaluation was carried out with a portable SONOSITE TITAN device (Medsonic) with convex transductor of 2.0–5.0 Mhz. The interpretation of the results was given by the interpolation of the numerical results of the scores from the liver parenchyma (IP) pattern, in addition to by the mean total thickness of four portal tracts after the first division from the right and left branches of portal vein (PT). The portal hypertension (PH) score was calculated by measuring the internal diameter of the portal vein and the presence of collateral circulation and ascites (1). Study carried out by Santos et al. (16) using the Niamey criteria, demonstrated moderate to substantial intra and inter-observer reproducibility in PPF classification.
Due to small sample sizes, it was also necessary to group together some categories of fibrosis for this stage of the analysis. Patients classified as without fibrosis (WF) and incipient fibrosis not excluded (IFNE) were united in a single group, as were possible periportal fibrosis (PPF) and periportal fibrosis (PF). The groups of advanced periportal fibrosis (APF) and APF with portal hypertension (APF/PH) were also analyzed in the same group. Twelve patients were classified as WF/IFNE, thirteen individuals as PPF/PF, and in the endemic areas we found only four individuals classified as APF/PH.
Individuals of both genders, aged between 10 and 60 years, who already tested positive for S. mansoni were included in the immunological evaluation. Subjects less than 10 years of age or older than 60, pregnant women, individuals with a history of chronic alcoholism, individuals with diseases that may interfere with the results of the immune response, such as hepatitis B and C, diabetes mellitus and HIV, and individuals who were on immunosuppressive drugs or undergoing chemotherapy of any kind were excluded from the study. Children younger than 10 and adults over 60 were not included in the immunological evaluation because of the potential of aberrant immune responses in these age groups. The mean age of the APF/PH group was higher (60.7 ± 4.2) than the WF/IFNE group (30.7 ± 10.2; p <0.05). There were no significant differences in gender distribution.
Determination of parasite infection was carried out using the spontaneous sedimentation technique (Hoffmann-Pons-Janer or Lutz method) and determination of the parasite load of Schistosoma infection was accomplished by the Kato-Katz method. The majority of individuals reported that they had been treated for schistosomiasis for less than 1 year (98%). This report may justify the low prevalence of infection (15%) at the time of recruitment for ultrasound analysis (USG). Additionally, the prevalence of specific soluble eggs antigen (SEA)-IgE levels was 89% among all groups evaluated (Table 1).
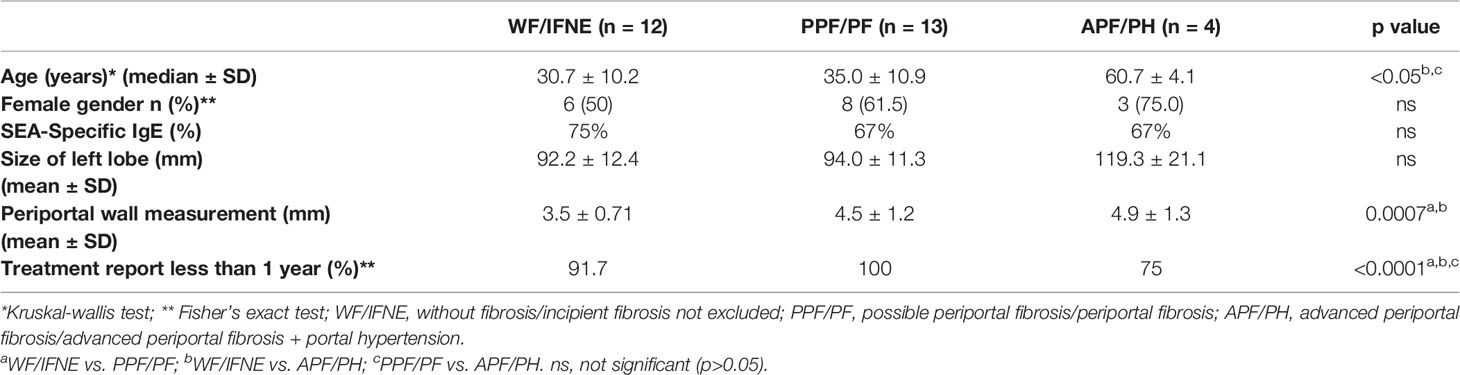
Table 1 Demographical and clinical characteristics of studied population.
Ethics Statement
The present study is part of a study approved by the Ethics Committee of the School of Nursing, Federal University of Bahia, entitled: ‘‘Identification of biomarkers associated with the development of severe forms of schistosomiasis’’ (License number: 1,374.864). All individuals who agreed to participate in the study signed the Informed Consent Form.
Preparation of Peripheral Blood Mononuclear Cells and Flow Cytometry Experiments
Peripheral blood mononuclear cells of patients different degrees of periportal fibrosis due to schistosomiasis were obtained using the Ficoll-Hypaque density gradient technique (GE Healthcare, Uppsala) and adjusted to a concentration of 1 × 107 cells/ml in complete RPMI 1640 (100 μl/ml gentamicin, 2mM L-glutamine, 30mM HEPES) containing 10% heat-inactivated fetal bovine serum (FBS) (Life Technologies GIBCO BRL, Gaithersburg, MD). Cells were stimulated with 10μg of soluble egg antigen (SEA) and maintained at 37°C and 5% CO2 in 96-well culture plates for 16h.
After incubation, brefeldin A (10 μg/ml; Sigma, St. Louis, MO) was added for 4 h at 37°C and 5% CO2. Labeling of PBMC surface markers was then performed with 20 μl of solution containing the specific conjugated antibodies for CD4 T lymphocytes and activation and regulation markers: CD3 (clone OKT3, eBioscience), CD4 (clone OKT4, eBioscience), CD25 (clone BC96, eBioscience) and CTLA-4 (clone 14D3, eBioscience). After this step, the plates were incubated with 150 μl/well of permeabilization buffer for 10 min at room temperature. After incubation, intracellular labeling was done using anti-cytokine and anti-transcription factor monoclonal antibodies: IL-4 (clone 8D4-8, BD Bioscience), IL-5 (clone TRFK5, eBioscience), IFN-γ (clone 45.B3, eBioscience), IL-17 (clone eBio64DEC17, eBioscience), FOXP3 (clone 236A/E7, eBioscience), IL-13 (clone PVM13-1, eBioscience), IL-10 (JES3-9D7, eBioscience), and TGF-β (clone 9016, R&D System). The acquisition was performed using the FACSCanto (Becton Dickinson) apparatus, for a total of 200,000 events.
The Flow Jo V10 (Tree Star, BD) program was used for analysis of lymphocytes. These cells were analyzed according to the frequency of expression of cell surface markers. Cell populations were defined by non-specific fluorescence from frontal (FSC) and lateral light (SSC) dispersion by parameters of particle size/volume and particle complexity, respectively. A population of lymphocytes was selected by gating based on cellular characteristics in this population. A specific region was delineated in the graph corresponding to the lymphocyte area, then the T lymphocyte population was selected by the presence of CD3 (Figure 1A), and within this population the CD4+ T lymphocyte population was subsequently identified. We finally evaluated the expression surface markers and intracellular cytokines in this population (Figures 1B–F).
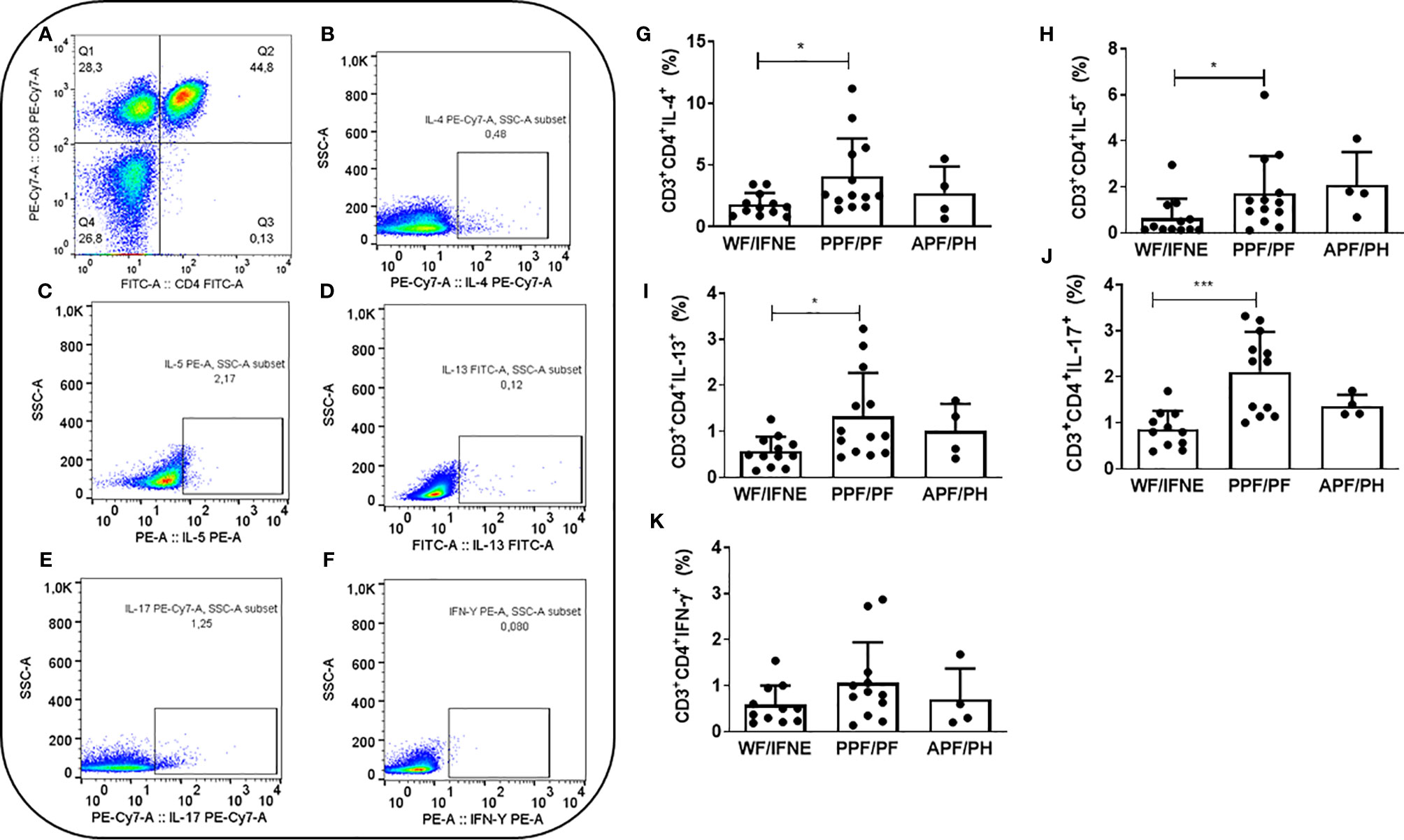
Figure 1 Representative plots of one experiment showing the frequency of CD4+ T lymphocytes (A) and the expression of IL-4 (B), IL-5 (C), IL-13 (D), IL-17 (E), and IFN-γ (F). Frequency of CD4+ T lymphocytes expressing IL-4 (G), IL-5 (H), IL-13 (I), IL-17 (J), IFN-γ (K) after SEA stimulation from individuals without fibrosis and with different degrees of periportal fibrosis secondary to schistosomiasis *p <0.05, ***p <0.005 (Mann-Whitney test).The histogram and bars represent the mean + SD.
Schistosoma mansoni Soluble Egg Antigen
The S. mansoni soluble egg antigen (SEA) used in this study were prepared as previously described (17) and kindly provided by Dr. Alfredo M. Góes.
Determination of Cytokine Levels
Levels of IL-10 and IL-13 were evaluated in the supernatants of PBMC cultures stimulated with SEA antigen (10 µg/ml) for 16 h according to the manufacturer’s instructions (Pharmingen, San Diego, CA). Briefly, plates (Nunc-Immuno Plate MaxiSorp Surface, Denmark) were sensitized overnight at 4°C with human anti-cytokine monoclonal antibody (anti-IL-10 or anti-IL-13). The following day, after washing the plates with PBS/Tween 0.05%, blockade of non-specific binding was performed with PBS + 0.01% bovine albumin for 2 h at room temperature. Three washes were performed with PBS/Tween 0.05% and then the samples, blanks, and standards were added and the plate was incubated at room temperature for 2 h. The plate was washed again 3 times and biotinylated human anti-cytokine detection antibody (2 μg/ml) was added. After incubating for 1 h at room temperature the plates were washed 4 times and the conjugate (streptavidin-conjugated peroxidase) was added. The plate was incubated for 30 min at room temperature. After washing, the substrate (3,3’,5,5’-tetramethylbenzidine + H2O2 + dimethyl sulfoxide) was added and the plate was incubated for 20 min at room temperature. The reaction was interrupted by the addition of H2SO4 (8M). Optical density (OD) was read at 450 nm (SpectraMax, Molecular Devices Corporation, Sunnyvale, CA) and the values were converted to pg/ml based on the standard curve (Soft Max Pro 5.0 Molecular Devices Corporation, Sunnyvale, CA).
Sample Size and Statistical Analysis
Sample size was calculated based on previous studies in which the degree of periportal fibrosis was evaluated by the Cairo protocol. We observed that 25% of individuals presented some degree of periportal fibrosis (18). A study power of 80% and an α error of 0.05% were taken into account. One blood sample was collected from each individual. The samples were collected in different trips to the endemic area and the data were plot in the same graph. Statistical analysis was performed in GraphPad PRISM 5.0 (La Jolla, CA, USA). Before the analysis of each data set, the D’Agostino-Pearson normality test was performed. Therefore, for comparison between two or more groups, parametric and non-parametric tests were used according to the nature of the data generated (ANOVA, Kruskal Wallis). Spearman correlation (rs) was calculated in the correlation analysis. All tests were two-tailed and statistical significance was established in the 95% confidence interval. P values <0.05 were considered significant.
Results
Cytokine Profile Expressed by CD4+ T Lymphocytes From Patients With Different Degrees of Periportal Fibrosis Secondary to Schistosomiasis
To investigate the contribution of Th1, Th2, and Th17 cytokines, we analyzed the intracellular expression of these molecules in CD4+ T lymphocytes from individuals with periportal fibrosis secondary to schistosomiasis. We first evaluated the frequency of T CD4+ lymphocytes expressing IL-4, IL-5, and IL-13 after stimulation with SEA, since these cytokines are the main molecules involved in the pathogenesis of periportal fibrosis related to Schistosoma mansoni infection (19). We observed an increase in the frequency of CD4+ T cells expressing these cytokines in the group of individuals with periportal fibrosis (PPF/PF), compared to individuals without fibrosis (WF/IFNE) (Figures 1G–I).
There was an increase in the frequency of CD4+ T lymphocytes expressing IL-17 in the PPF/PF group (p<0.01), when compared to WF/IFNE group (Figure 1J). In order to try to understand the possible role of this cytokine we proceeded to correlate its expression with ultrasonographic specific measurements. We performed a correlation between IL-17 and the measurement of the periportal wall and the size of the left lobe (Supplementary Figure 1A) and observed a negative correlation between the frequency of TCD4+IL-17+ cells and the measurement of the periportal wall (rs =0.53; p <0.05), but we did not observe any correlation with the size of the left lobe (Supplementary Figure 1B). The intracellular expression of IFN-γ in CD4 T lymphocytes did not differ between all evaluated groups (Figure 1K).
Evaluation of Regulatory Molecules by CD4 T Lymphocytes in Individuals Without Fibrosis and With Different Degrees of Periportal Fibrosis
We next evaluated the frequency of the main regulatory molecules involved in the control of schistosomiasis fibrosis (6) CTLA-4, IL-10, besides regulatory T cells (CD4+CD25hi) expressing the regulatory molecules FOXP3 and IL-10 (Figure 2A).
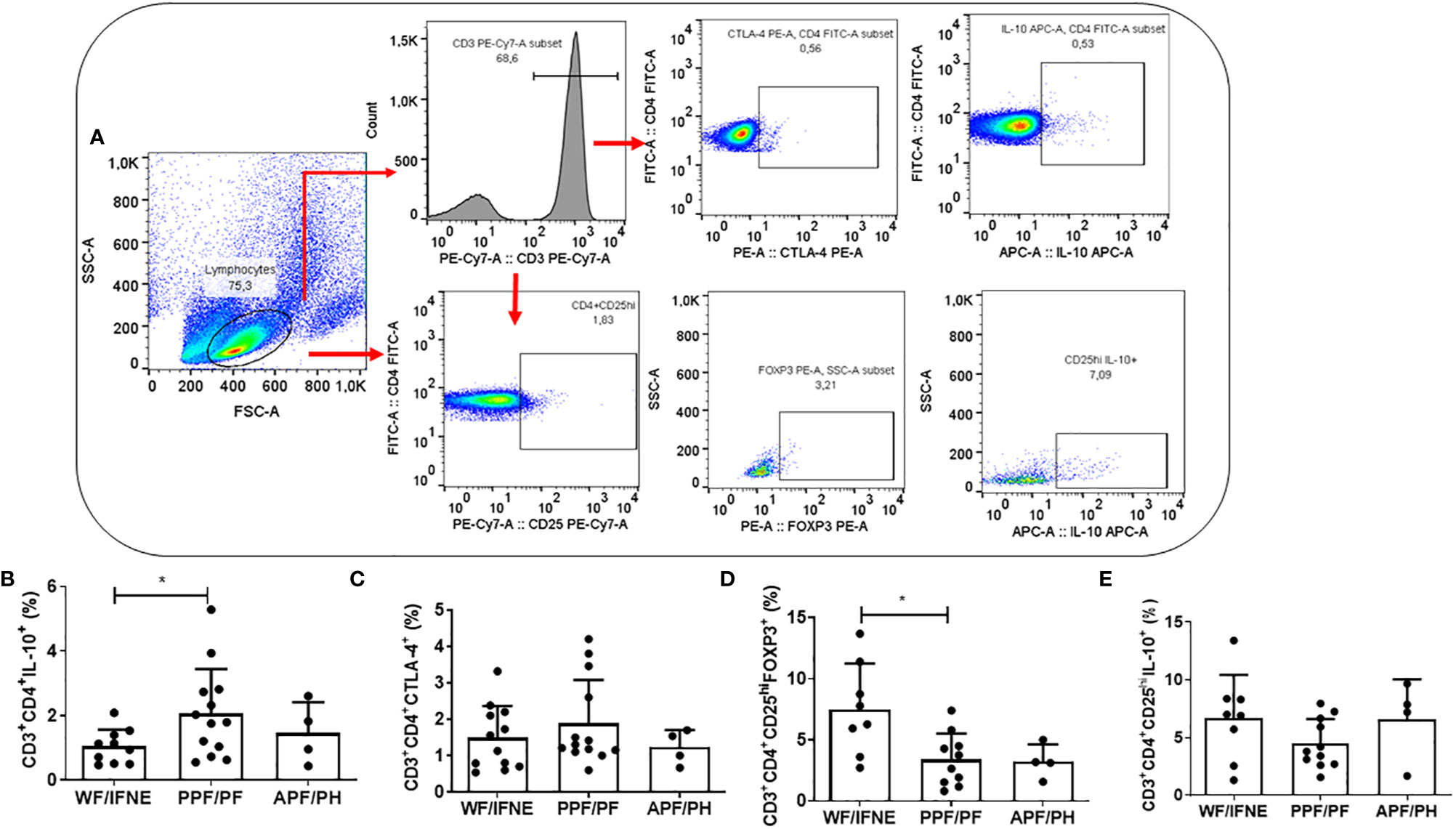
Figure 2 Flow cytometry gating strategy used in order to analyze the regulatory molecules, CTLA-4, FOXP3 and IL-10 (A). Frequency of CD4 T lymphocytes expressing IL-10 (B), CTLA-4 (C), and the CD4+CD25+ that express FOXP3 (D) or IL-10 (E) after SEA stimulation from individuals without fibrosis and with different degrees of periportal fibrosis secondary to schistosomiasis *p < 0.05, (Mann-Whitney test). The histogram and bars represent the mean + SD.
The frequency of CD4+IL-10+ T lymphocytes was higher in the PPF/PF group, compared to the WF/FINE group (p<0.01; Figure 2B). We did not observe any difference in the CD4+CTLA-4+ T cells among all groups (Figure 2C). The frequency of T reg FOXP3+ cells was higher in the group of individuals WF/FINE compared to PPF/PF (p<0.01; Figure 2D). Regarding the expression of IL-10 by the regulatory cells, in our study we did not find any statistical difference among all groups (Figure 2E).
Since we observed in the group without fibrosis a lower frequency of TCD4+ lymphocytes expressing IL-10, due to its role in controlling the pathogenesis of the disease, we evaluated the levels of this cytokine in the culture supernatant (Supplementary Figure 2A. The levels of IL-10 did not differ between the WF and PPF/PF groups, while they were lower in the APF/PH group, compared to the PPF/PF group (p<0,01; Supplementary Figure 2A).
We performed a correlation between CD4+ T cells expressing IL-10 with CD4+ T lymphocytes expressing IL-4 or IL-13. We observed a positive correlation only with CD4+IL-4+ T cells (p=0.0007, r=0.79) (Supplementary Figure 3A).
We then evaluated the levels of IL-13 in the culture supernatants (Supplementary Figure 2B), one of the main cytokines related to the fibrosis process (13, 19, 20); the results were similar to the expression of IL-13 by CD4+ T lymphocytes (Figure 1I). However, the IL-10/IL-13 ratio was higher in the group without fibrosis, compared to the PPF/PF group (p <0.01; Supplementary Figure 2C). These findings suggest that IL-10 produced by other cellular sources, such as macrophages, dendritic cells and CD8+ T lymphocytes, would be of fundamental importance in the regulation of this group without fibrosis.
The Expression of Pro-Fibrotic TGF-β by CD4+ T Lymphocytes and Correlation With the Pathogenesis of Periportal Fibrosis Secondary to Schistosomiasis
Since TGF-β is the main pro-fibrotic cytokine produced by CD4+ T lymphocytes, we decided to evaluate the frequency of these cells producing TGF-β (Figures 3A–D) and the correlation between this cytokine and ultrasonography measurements (Figures 3E, F).
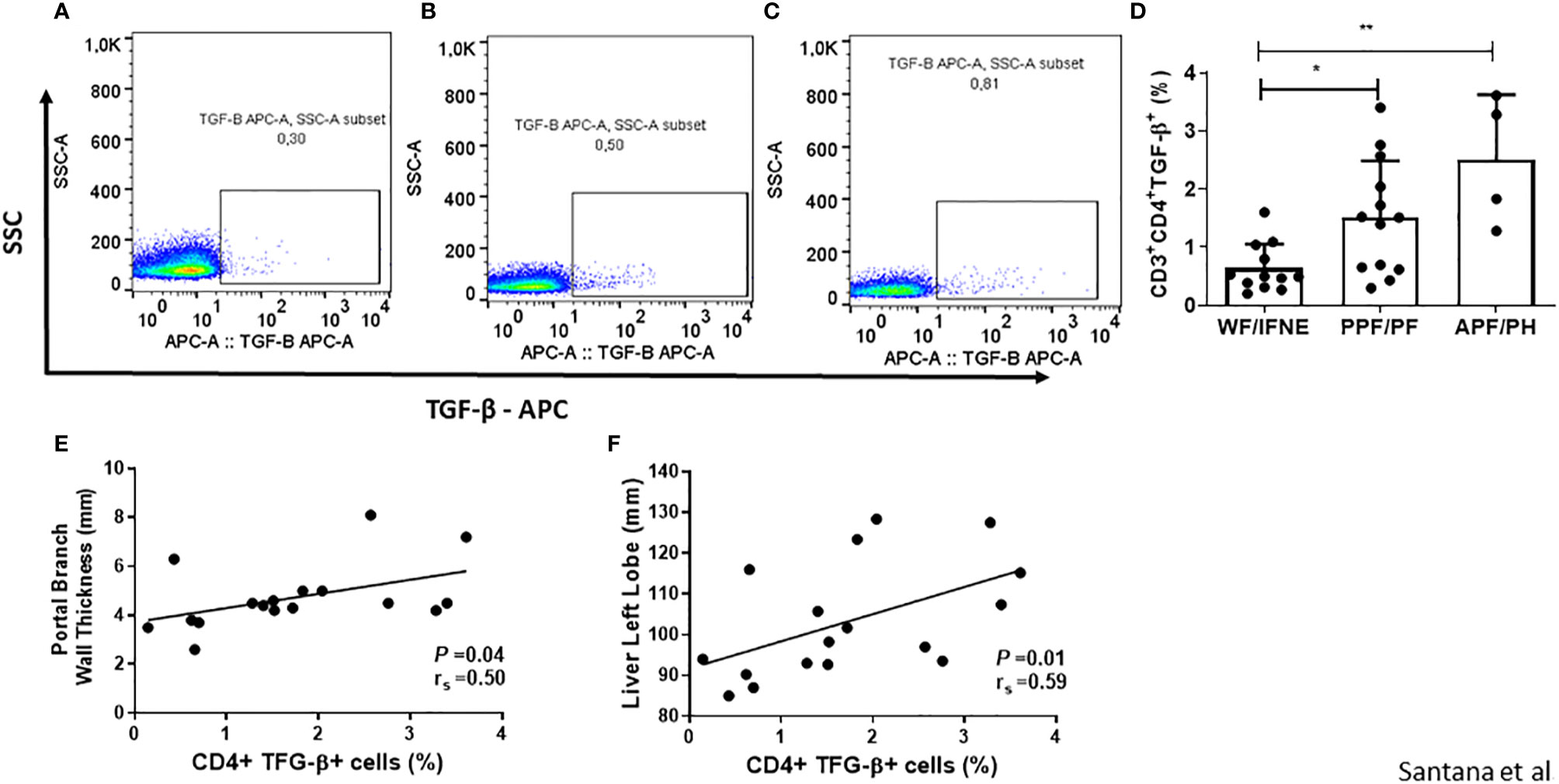
Figure 3 Representative plots of one experiment showing the frequency of CD4+TGF-β+ T lymphocytes in the different groups (A–C). Frequency of CD4 T lymphocytes expressing TGF-β after SEA stimulation from individuals without fibrosis and with different degrees of periportal fibrosis secondary to schistosomiasis (D). Correlations between this value and the portal branch wall thickness or the size of the liver left lobe of the patients that shows some level of fibrosis (E, F). Spearman’s rank correlation coefficient (rs) and P value is shown. *p < 0.05 **p < 0.01.
We observed a higher frequency of CD4+TGF-β+ T lymphocytes in the cultures of individuals with any grade of periportal fibrosis, both in the PPF/PF and APF/PH groups, compared to the group without fibrosis (p<0.01; Figure 3A). We also performed a correlation between TCD4+ TGF-β+ T lymphocytes from cultures of individuals with some degree of periportal fibrosis with the measurement of the periportal wall and the size of the left lobe (Figures 3E, F). And we observed a positive correlation both with the measurement of the periportal wall (r=0.5; p<0.05) and the size of the left lobe (r=0.59; p<0.05). We did not observe a significant difference in correlation of the frequency of CD4+T cells expressing TGF-β with CD4+ IL-4+ and CD4+IL-13+ T cells in the group of individuals with periportal fibrosis (Supplementary Figure 3B).
Discussion
The hepatic fibrosis associated with chronic S. mansoni infection contributes to the development of the most severe forms of the disease and is responsible for the morbidity and mortality observed in individuals living in endemic areas (6). In this study, we identified the complex network of cytokines expressed by CD4+T lymphocytes in individuals with periportal fibrosis secondary to schistosomiasis who reside in an endemic area.
Epidemiological studies in populations living in endemic areas suggest that the development of hepatic fibrosis is associated with several factors such as disease duration, parasite load, sex, as well as genetic and immunological factors of the study population (4, 21). Demographic analysis of the study population showed no difference in gender ratio between individuals evaluated by ultrasonography and those selected for immunologic evaluation. Some studies, however, have demonstrated the importance of gender in the development of periportal fibrosis (22, 23). In fact, a previous study by our group in the Água Preta population showed that women were more likely than men to be exposed to contaminated water, probably due to gendered activities taking place in river water such as washing clothes, dishes, and food preparation. Despite this, no significant differences in the development of hepatic fibrosis between genders in this population were found (10).
As far as age, the APF/PH group presented a higher median age in relation to the WF/IFNE group. These results corroborate the data presented by Alves Oliveira et al. (15) in which the majority of individuals with the most advanced degrees of fibrosis were over 50 years of age (15). This could be explained by repeated parasite re-exposure in the older age group or by the slow process of fibrotic tissue generation; younger individuals have likely not been exposed enough time for the cumulative effects of the excessive secretion of collagen in the peripheral hepatic parenchyma or near the periportal tract to be observed (24).
The present study demonstrates a greater production of IL-4, IL-5, and IL-13 by the lymphocytes of individuals with periportal fibrosis. IL-4 is responsible for the differentiation of the Th2 response and the blockade of this cytokine in S. mansoni-infected mice leads to a decrease in hepatic collagen deposition (25). In animals reinfected by S. mansoni, IL-4 is a risk factor for the development of fibrosis and in humans this cytokine shows a positive correlation with fibrosis score (26, 27). There are studies that also strengthen the association between a higher degree of fibrosis and higher levels of IL-5, a cytokine whose main function is eosinophil activation (13). Additionally, IL-5 knockout is able to reduce hepatic fibrosis in S. mansoni-infected mice (28). Many studies have pointed to IL-13 as the main pro-fibrotic cytokine, since its blockade is capable of both preventing the onset of periportal fibrosis and of reducing already established fibrosis (17, 29, 30). IL-13 levels in the supernatant of PBMCs cultures from patients also show a positive association with the presence and severity of periportal fibrosis (15). In this study, the group with fibrosis showed a higher frequency of CD4+ T lymphocytes expressing IL-13 when compared to individuals without fibrosis, as well as higher levels of this cytokine in the supernatant of PBMCs cultures stimulated with SEA. The source of Th2-type cytokines are not only produced by Th2 cells, but are also secreted by other innate lymphocytes, such as type 2 innate lymphoid cells (ILC2s), which could be contributing to the higher levels of this cytokine in the culture supernatant.
Contrary to what was expected, lymphocytes from individuals with advanced fibrosis did not present a higher production of Th2 and proinflammatory cytokines when compared to the other groups. This fact may be associated with the phenomenon of immune exhaustion, which is characterized by the gradual and progressive loss of specific T-cell effector functions due to antigenic persistence in chronic infections (31). Despite being initially described in models of viral infections, exhaustion has also been documented in parasitic infections, including schistosomiasis in a murine model, as reviewed by Rodrigues et al. (32). Another fact corroborating this hypothesis is the advanced age of patients with advanced fibrosis, which may indicate a longer period of infection and antigenic exposure, that would lead to hyposensitivity to the parasite. However, we must consider the small number of individuals with the most severe forms of fibrosis in the endemic area.
Regarding the expression of IL-17 by CD4+ T lymphocytes; interestingly, lymphocytes of individuals with fibrosis had a higher production of IL-17 (6). However, the role of IL-17 in periportal fibrosis secondary to schistosomiasis is still poorly understood. IL-17 is able to exacerbate fibrosis in the murine model through activation of Kupffer cells and direct induction of collagen production by hepatic stellate cells (33). Neutralization of IL-17 is able to reduce granuloma formation and liver damage in mice infected with S. japonicum (34). This evidence, when viewed in conjunction with our data, leads us to believe that IL-17 acts as a pro-fibrotic cytokine in schistosomiasis, as could be the case with S. haematobium (35). In addition to the expression of IL-17A by CD4+ T lymphocytes, we expanded the focus of our study by assessing the correlation between this cytokine and parameters obtained by ultrasonography. There is a negative correlation between CD4+T IL-17+ and the thickness of the portal vein wall, suggesting a protective role of this cytokine in fibrogenesis. This could be due to the fact that Th17 is not the only source of the cytokine (36). Further studies are needed to clarify the role of this cytokine in the protection or pathogenesis of periportal fibrosis in humans.
The expression of the proinflammatory cytokine IFN-γ by lymphocytes of individuals did not differ among all groups. The role of this cytokine in periportal fibrosis secondary to schistosomiasis remains controversial, as the majority of studies show a profibrotic role for this cytokine, as reviewed by Zheng et al. (6).
The cytokine IL-10 seems to have an important protective role in the pathogenesis of periportal fibrosis. We observed a higher frequency of CD4+IL-10+ T cells in the PPF/PF group compared to individuals without fibrosis. The increased expression of these cellular markers in the PPF/PF groups could be explained by an attempt to control the greater activation and production of Th2 and proinflammatory cytokines observed in the lymphocytes of this group of patients. The smaller ratio of secreted cytokines IL-10/IL-13 could indicate that the regulatory response is not strong enough to prevent the fibrosis. Different studies have shown that the low level of IL-10 secreted by PBMCs was associated with severe fibrosis (24, 37). And there could be additional sources of IL-10-like monocytes. We have found a correlation between the frequency of CD4+ T cells expressing IL-4+ with CD4+ T cells expressing IL-10+ or TGF-β. A study evaluating PBMC from patients with filariasis, showed that CD4+ T cells are the main source of IL-10 and the majority of these cells co-produced neither IL-4 nor IFN-γ. In this study, only 22% of them stained positively for IL-4 (38). In our study, the persistence of the SEA antigen in the second periportal branches could justify the positive correlation observed between IL-4 and IL-10. And IL-10 could also be produced by Th2 cells and other cells, like macrophages. Further investigations, analyzing in detail which specific cell populations are expressing each cytokine, using a flow cytometry approach that could include these types of analysis, may help us to understand the role of these populations.
Few studies have focused on the profile of T reg cells in individuals with periportal fibrosis secondary to schistosomiasis, and how they become activated and migrate to exert their functions has been reviewed elsewhere (39). In our study, we observed a higher frequency of T reg lymphocytes in the group without fibrosis compared to the PPF/PF group. In a previous work we have shown a higher frequency of CD4+CD25hi T cells in individuals with moderate/severe fibrosis (10). Other studies have associated the presence of T reg FOXP3+ lymphocytes in peripheral blood with the worsening of the disease, suggesting a failure in the recruitment of this cell population to the liver tissue (40, 41).
Regarding TGF-β, an important cytokine involved in fibrosis due to its ability to induce fibroblast proliferation and collagen deposition, we observed a higher frequency of CD4+ T lymphocytes expressing this cytokine in groups with some degree of fibrosis. Studies evaluating PBMCs stimulated with SEA or serum levels of TGF-β from individuals with different degrees of fibrosis did not report differences between the studied groups (13, 14). Nonetheless, in a study published by our group in 2014 evaluating subpopulations of monocytes in patients with different degrees of fibrosis secondary to schistosomiasis, we observed higher intracellular levels of TGF-β in classic, intermediate, and non-classic monocytes in individuals with moderate to severe fibrosis compared to individuals without fibrosis or individuals with incipient fibrosis (12). Our findings suggest that like monocytes, the CD4+ T lymphocytes are an important source of TGF-β and play an essential role in fibrogenesis. Our results also show a positive correlation between TGF-β and the measurement of the left lobe and the portal vein wall thickness. The article published by Li et al. (42) analyzing individuals with periportal fibrosis due to Schistosoma japonicum infections shows a positive correlation between TGF-β mRNA levels with spleen thickness and liver stiffness (42), parameters which are correlated with the severity of the disease. Furthermore, an association between the expression of this molecule and the degree of inflammation was observed; therefore, TGF-β could participate in both the inflammatory process and the fibrotic process (42–44).
In light of the above, we conclude that in the PPF/PF group, despite intense production of cytokines associated with the inflammatory response, there is a regulation through IL-10 and T reg cells that could maintain the low morbidity of this group. To our knowledge, this is the first study showing the complex network of cytokines expressed by CD4+ T cells in endemic area patients classified by ultrasonography. Interventions focused on the control of the cytokines related to periportal fibrosis could be useful to reduce the morbidity associated with schistosomiasis.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The studies involving human participants were reviewed and approved by the Ethics Committee of the School of Nursing, Federal University of Bahia, License number: 1,374.864. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
LC, EC, and SO contributed to conception and design of the study. JS, TA, DL, LR and BP carried out most of the experiments. IS performed the ultrasound evaluations of the patients and helped in the interpretation of the results. JS, BP, NG, and LC wrote the manuscript and carried out the statistical analysis and prepared the figures. LC submitted this paper. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the FAPESB/EDITAL UNIVERSAL, grant number APP0051/2016. This work was also supported by the CNPq/MST/INCT-DT, grant number 465229/2014-0. This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES)—Finance Code 001.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank the patients of Água Preta for participating in the study as well as all the teams of professionals who work in these endemic areas for their dedication to the clinical work of the study. EC, SO, and LC are investigators supported by The Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). We would also like to thank A.E. Carrasco for English language assistance.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.605235/full#supplementary-material
Supplementary Figure 1 | Correlations between the frequency of CD4+CD17+ T lymphocytes and the portal branch wall thickness or the size of the liver left lobe of the patients that show some level of fibrosis. Spearman’s rank correlation coefficient (rs) and P value is shown.
Supplementary Figure 2 | Levels of IL-10 (A) and IL-13 (B) in the supernatant of PBMC stimulated with SEA antigen from individuals without fibrosis and with different degrees of periportal fibrosis secondary to schistosomiasis. (C) The IL-10/IL-13 ratio. *p <0.05, (Mann-Whitney test). The bars represent the mean + SD. WF/IFNE, without fibrosis and incipient fibrosis not excluded (N=12); PF/PPF, periportal fibrosis and possible periportal fibrosis (N=12). APF/PH, advanced periportal fibrosis and APF with portal hypertension (N=4).
Supplementary Figure 3 | Correlations between the frequency of CD4+ T cells from individuals with any degree of periportal fibrosis expressing different cytokines. (A) CD4+IL4+ T cells vs. CD4+ IL-10 +; (B) CD4+IL13+ vs. CD4+ IL-10+; (C) CD4+IL4+ T cells vs. CD4+TGF-β + T cells; (D) CD4+IL-13+ T cells vs. CD4+TGF-β + T cells. Spearman’s rank correlation coefficient (rs) and P value is shown.
References
1. World Health Organization (WHO). Schistosomiasis of WHO (2019). Available at: http://www.who.int/health-topics/schistosomiasis#tab=tab_1 (Accessed August 30, 2020).
2. Bina JC, Prata A. Schistosomiasis in hyperendemic area of Taquarendi: I-Schistosoma mansoni infection and severe clinical forms. Rev Soc Bras Med Trop (2003) 36:211–6. doi: 10.1590/S0037-86822003000200003
3. Henri S, Chevillard C, Mergani A, Paris P, Gaudart J, Camilla C, et al. Cytokine regulation of periportal fibrosis in humans infected with Schistosoma mansoni: IFN-γ is associated with protection against fibrosis and TNF-α with aggravation of disease. J Immunol (2002) 169:929–36. doi: 10.4049/jimmunol.169.2.929
4. Gryseels B, Polman K, Clerinx J, Kestens L. Human schistosomiasis. Lancet (2006) 368:1106–18. doi: 10.1016/S0140-6736(06)69440-3
5. Wilson MS, Mentink - Kane MM, Pesce JT, Ramalingam TR, Thompson R, Wynn TA. Immunopathology of schistosomiasis. Immunol Cell Biol (2007) 85:148–54. doi: 10.1038/sj.icb.7100014
6. Zheng B, Zhang J, Chen H, Nie H, Miller H, Gong Q, et al. T Lymphocyte-Mediated Liver Immunopathology of Schistosomiasis. Front Immunol (2020) 11:61. doi: 10.3389/fimmu.2020.00061
7. Araújo MI, de Jesus AR, Bacellar O, Sabin E, Pearce E, Carvalho EM. Evidence of a T helper type 2 activation in human schistosomiasis. Eur J Immunol (1996) 26:1399–403. doi: 10.1002/eji.1830260633
8. Rutitzky LI, Hernandez HJ, Stadecker MJ. Th1-polarizing immunization with egg antigens correlates with severe exacerbation of immunopathology and death in schistosome infection. Proc Natl Acad Sci (2001) 98:13243–8. doi: 10.1073/pnas.231258498
9. Rutitzky LI, Bazzone L, Shainheit MG, Joyce-Shaikh B, Cua DJ, Stadecker MJ. IL-23 is required for the development of severe egg-induced immunopathology in schistosomiasis and for lesional expression of IL-17. J Immunol (2008) 180:2486–95. doi: 10.4049/jimmunol.180.4.2486
10. Cardoso LS, Barreto ADSR, Fernandes JS, Oliveira RR, Souza RDPD, Carvalho EM, et al. Impaired lymphocyte profile in schistosomiasis patients with periportal fibrosis. Clin Dev Immunol (2013) 2013:1–8. doi: 10.1155/2013/710647
11. Louis H, Van Laethem JL, Wu W, Quertinmont E, Degraef C, Van den Berg K, et al. Interleukin-10 controls neutrophilic infiltration, hepatocyte proliferation, and liver fibrosis induced by carbon tetrachloride in mice. Hepatology (1998) 28:1607–15. doi: 10.1002/hep.510280621
12. Fernandes JS, Araujo MI, Lopes DM, Souza RDPD, Carvalho EM, Cardoso LS. Monocyte subsets in schistosomiasis patients with periportal fibrosis. Mediators Inflamm (2014) 2014:1–12. doi: 10.1155/2014/703653
13. de Jesus AR, Magalhaes A, Miranda DG, Miranda RG, Araújo MI, de Jesus AA, et al. Association of type 2 cytokines with hepatic fibrosis in human Schistosoma mansoni infection. Infect Immun (2004) 72:3391–7. doi: 10.1128/IAI.72.6.3391-3397.2004
14. De Souza RDP, Cardoso LS, Lopes GTV, Almeida MCF, Oliveira RR, Alcântara LM, et al. Cytokine and chemokine profile in individuals with different degrees of periportal fibrosis due to Schistosoma mansoni infection. J Parasitol Res (2012) 2012:1–10. doi: 10.1155/2012/394981
15. Alves Oliveira LF, Moreno EC, Gazzinelli G, Martins-Filho OA, Silveira AMSD, Gazzinelli A, et al. Cytokine production associated with periportal fibrosis during chronic schistosomiasis mansoni in humans. Infect Immun (2006) 74:1215–21. doi: 10.1128/IAI.74.2.1215-1221.2006
16. Santos GT, Sales DM, Leão ARS, Santos JEM, Aguiar LAK, Brant PE, et al. Reprodutibilidade da classificação ultra-sonográfica de Niamey na avaliação da fibrose periportal na esquistossomose mansônica. Radiol Bras (2007) 40:377–81. doi: 10.1590/S0100-39842007000600005
17. Hirsch C, Goes AM. Characterization of fractionated Schistosoma mansoni soluble adult worm antigens that elicit human cell proliferation and granuloma formation in vitro. Parasitology (1996) 112:529–35. doi: 10.1017/S0031182000066105
18. De Jesus AR, Miranda DG, Miranda RG, Araújo I, Magalhães A, Bacellar O, et al. Morbidity associated with Schistosoma mansoni infection determined by ultrasound in an endemic area of Brazil, Caatinga do Moura. Am J Trop Med Hyg (2000) 63:1–4. doi: 10.4269/ajtmh.2000.63.1
19. Fallon PG, Richardson EJ, McKenzie GJ, McKenzie NA. Schistosome infection of transgenic mice defines distinct and contrasting pathogenic roles for IL-4 and IL-13: IL-13 is a profibrotic agent. J Immunol (2000) 164:2585–91. doi: 10.4049/jimmunol.164.5.2585
20. Wynn TA, Thompson RW, Cheever AW, Mentink Kane MM. Immunopathogenesis of schistosomiasis. Immunol Rev (2004) 201:156–67. doi: 10.1111/j.0105-2896.2004.00176.x
21. Gryseels B. Schistosomiasis. Infect Dis Clin North Am (2012) 26:383–97. doi: 10.1016/j.idc.2012.03.004
22. Silva-Teixeira DN, Contigli C, Lambertucci JR, Serufo JC, Rodrigues V. Gender-related cytokine patterns in sera of schistosomiasis patients with Symmers’ fibrosis. Clin Diagn Lab Immunol (2004) 11:627–30. doi: 10.1128/CDLI.11.3.627-630.2004
23. Dessein AJ, Couissinier P, Demeure C, Rihet P, Kohlstaedt S, Carneiro-Carvalho D, et al. Environmental, genetic and immunological factors in human resistance to Schistosoma mansoni. Immunol Invest (1992) 21:423–53. doi: 10.3109/08820139209069383
24. Booth M, Mwatha JK, Joseph S, Jones FM, Kadzo H, Ireri E, et al. Periportal fibrosis in human Schistosoma mansoni infection is associated with low IL-10, low IFN-γ, high TNF-α, or low RANTES, depending on age and gender. J Immunol (2004) 172:1295–303. doi: 10.4049/jimmunol.172.2.1295
25. Cheever AW, Williams ME, Wynn TA, Finkelman FD, Seder RA, Cox TM, et al. Anti-IL-4 treatment of Schistosoma mansoni-infected mice inhibits development of T cells and non-B, non-T cells expressing Th2 cytokines while decreasing egg-induced hepatic fibrosis. J Immunol (1994) 153:753–9.
26. Farah IO, Mola PW, Kariuki TM, Nyindo M, Blanton RE, King CL. Repeated exposure induces periportal fibrosis in Schistosoma mansoni-infected baboons: role of TGF-β and IL-4. J Immunol (2000) 164:5337–43. doi: 10.4049/jimmunol.164.10.5337
27. Kamal SM, Bianchi L, Al Tawil A, Koziel M, El Sayed Khalifa K, Peter T, et al. Specific cellular immune response and cytokine patterns in patients coinfected with hepatitis C virus and Schistosoma mansoni. J Infect Dis (2001) 184:972–82. doi: 10.1086/323352
28. Reiman RM, Thompson RW, Feng CG, Hari D, Knight R, Cheever AW, et al. Interleukin-5 (IL-5) augments the progression of liver fibrosis by regulating IL-13 activity. Infect Immun (2006) 74:1471–9. doi: 10.1128/IAI.74.3.1471-1479.2006
29. Chiaramonte MG, Donaldson DD, Cheever AW, Wynn TA. An IL-13 inhibitor blocks the development of hepatic fibrosis during a T-helper type 2–dominated inflammatory response. J Clin Invest (1999) 104:777–85. doi: 10.1172/JCI7325
30. Ramalingam TR, Gieseck RL, Acciani TH, M Hart K, Cheever AW, Mentink-Kane MM, et al. Enhanced protection from fibrosis and inflammation in the combined absence of IL-13 and IFN-γ. J Pathol (2016) 239:344–54. doi: 10.1002/path.4733
31. Cardoso LS, Oliveira SC, Góes AMD, Oliveira RR, Pacifico LG, Marinho FV, et al. Schistosoma mansoni antigens modulate the allergic response in a murine model of ovalbumin-induced airway inflammation. Clin Exp Immunol (2010) 160:266–74. doi: 10.1111/j.1365-2249.2009.04084.x
32. Rodrigues V, Cordeiro-da-Silva A, Laforge M, Ouaissi A, Akharid K, Silvestre R, et al. Impairment of T cell function in parasitic infections. PloS Negl Trop Dis (2014) 8:e2567. doi: 10.1371/journal.pntd.0002567
33. Meng YC, Jiang HX, Zhang JH, Lu ZF, Qin SY, Ning L. Activation of hepatocyte growth factor-induced apoptosis in hepatic stellate cells. Zhonghua Gan Zang Bing Za Zhi (2012) 20:698. doi: 10.3760/cma.j.issn.1007-3418.2012.09.012
34. Zhang Y, Chen L, Gao W, Hou X, Gu Y, Gui L, et al. IL-17 neutralization significantly ameliorates hepatic granulomatous inflammation and liver damage in Schistosoma japonicum infected mice. Eur J Immunol (2012) 42:1523–35. doi: 10.1002/eji.201141933
35. Mbow M, Larkin BM, Meurs L, Wammes LJ, De Jong SE, Labuda LA, et al. T-helper 17 cells are associated with pathology in human schistosomiasis. J Infect Dis (2013) 207:186–95. doi: 10.1093/infdis/jis654
36. Cua DJ, Tato CM. Innate IL-17-producing cells: the sentinels of the immune system. Nat Rev Immunol (2010) 10:479–89. doi: 10.1038/nri2819
37. Arnaud V, Li J, Wang Y, Fu X, Mengzhi S, Luo X, et al. Regulatory role of interleukin-10 and interferon-γ in severe hepatic central and peripheral fibrosis in humans infected with Schistosoma japonicum. J Infect Dis (2008) 198:418–26. doi: 10.1086/588826
38. Mitre E, Chien D, Nutman TB. CD4+ (and Not CD25+) T Cells Are the Predominant Interleukin-10-Producing Cells in the Circulation of Filaria-Infected Patients. J Infect Dis (2008) 197:94–101. doi: 10.1086/524301
39. Kamdem SD, Moyou-Somo R, Brombacher F, Nono JK. Host regulators of liver fibrosis during human schistosomiasis. Front Immunol (2018) 9:2781. doi: 10.3389/fimmu.2018.02781
40. Watanabe K, Mwinzi PN, Black CL, Muok EM, Karanja DM, Secor WE, et al. T regulatory cell levels decrease in people infected with Schistosoma mansoni on effective treatment. Am J Trop Med Hyg (2007) 77:676–82. doi: 10.4269/ajtmh.2007.77.676
41. Romano A, Hou X, Sertorio M, Dessein H, Cabantous S, Oliveira P, et al. FOXP3+ regulatory T cells in hepatic fibrosis and splenomegaly caused by Schistosoma japonicum: the spleen may be a major source of Tregs in subjects with splenomegaly. PLoS Negl Trop Dis (2016) 10:e0004306. doi: 10.1371/journal.pntd.0004306
42. Li L, Wu T, Huang J, Ma K, Xu L, Wang H, et al. Expression of heat shock protein 47, transforming growth factor-beta 1, and connective tissue growth factor in liver tissue of patients with Schistosoma japonicum-induced hepatic fibrosis. Parasitology (2015) 142:341. doi: 10.1017/S0031182014001115
43. Fabregat I, Moreno-Càceres J, Sánchez A, Dooley S, Dewidar B, Giannelli G, et al. TGF-β signalling and liver disease. FEBS J (2016) 283:2219–32. doi: 10.1111/febs.13665
Keywords: schistosomiasis, periportal fibrosis, Schistosoma mansoni, CD4+ T lymphocytes, fibrosis
Citation: Santana JB, de Almeida TVVS, Lopes DM, Page B, Oliveira SC, Souza I, Ribeiro LEVS, Gutiérrez NAG, Carvalho EM and Cardoso LS (2021) Phenotypic Characterization of CD4+ T Lymphocytes in Periportal Fibrosis Secondary to Schistosomiasis. Front. Immunol. 12:605235. doi: 10.3389/fimmu.2021.605235
Received: 11 September 2020; Accepted: 04 January 2021;
Published: 22 February 2021.
Edited by:
Thiago Almeida Pereira, Stanford University, United StatesReviewed by:
Keke Celeste Fairfax, The University of Utah, United StatesMargaret M. Mentink-Kane, Biomedical Research Institute, United States
Copyright © 2021 Santana, de Almeida, Lopes, Page, Oliveira, Souza, Ribeiro, Gutiérrez, Carvalho and Cardoso. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luciana Santos Cardoso, bHVjaWFuYWNAdWZiYS5icg==