
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 28 August 2020
Sec. Multiple Sclerosis and Neuroimmunology
Volume 11 - 2020 | https://doi.org/10.3389/fimmu.2020.02163
This article is part of the Research Topic COVID-19 in CNS and PNS: Basic and Clinical Focus on the Mechanisms of Infection and New Tools for the Therapeutic Approach View all 26 articles
 Inmaculada Sanclemente-Alaman1†‡
Inmaculada Sanclemente-Alaman1†‡ Lidia Moreno-Jiménez1†‡
Lidia Moreno-Jiménez1†‡ María Soledad Benito-Martín1†
María Soledad Benito-Martín1† Alejandro Canales-Aguirre2†
Alejandro Canales-Aguirre2† Jordi A. Matías-Guiu1†
Jordi A. Matías-Guiu1† Jorge Matías-Guiu1†
Jorge Matías-Guiu1† Ulises Gómez-Pinedo1*†
Ulises Gómez-Pinedo1*†Introduction: The response to the SARS-CoV-2 coronavirus epidemic requires increased research efforts to expand our knowledge of the disease. Questions related to infection rates and mechanisms, the possibility of reinfection, and potential therapeutic approaches require us not only to use the experimental models previously employed for the SARS-CoV and MERS-CoV coronaviruses but also to generate new models to respond to urgent questions.
Development: We reviewed the different experimental models used in the study of central nervous system (CNS) involvement in COVID-19 both in different cell lines that have enabled identification of the virus’ action mechanisms and in animal models (mice, rats, hamsters, ferrets, and primates) inoculated with the virus. Specifically, we reviewed models used to assess the presence and effects of SARS-CoV-2 on the CNS, including neural cell lines, animal models such as mouse hepatitis virus CoV (especially the 59 strain), and the use of brain organoids.
Conclusion: Given the clear need to increase our understanding of SARS-CoV-2, as well as its potential effects on the CNS, we must endeavor to obtain new information with cellular or animal models, with an appropriate resemblance between models and human patients.
On 31 December 2019, the Word Health Organization reported for the first time on an epidemic of lower respiratory tract infection in Wuhan, in the Chinese province of Hubei. The causal agent was soon identified as Severe acute respiratory syndrome coronavirus (SARS-CoV-2), a coronavirus (CoV), and the associated disease was named coronavirus disease 19 (COVID-19) (1). CoVs are positive-sense single-stranded RNA viruses resembling a crown under microscopy due to the presence of spike (S) glycoproteins on the viral envelope. There are four types of CoVs: αCoVs, βCoVs, δCoVs, and γCoVs. SARS-CoV, MERS-CoV, and SARS-CoV-2 are zoonotic (2), first infecting animals and then spreading to humans. βCoVs, which include SARS-CoV-2, are thought to originate in bats (3, 4), among other species. These viruses can cause respiratory and enteric diseases in different animal species. In humans, HCoV-OC43 and HCoV-HKU1 (αCoVs), and HCoV-229E and HCoV-NL63 (βCoVs) can cause the common cold and self-limiting infection of the lower respiratory tract in immunocompetent individuals in seasonal periods (5). Two previous epidemics have, however, been caused by CoVs; these were similar to the current pandemic, though with higher mortality rates, and were caused by the severe acute respiratory syndrome coronavirus (SARS-CoV) and middle east respiratory syndrome coronavirus (MERS-CoV) and affected many countries (6). Although they led to many research studies, expanding our knowledge of the viruses, the COVID-19 epidemic has had a far greater global impact. One as-yet unknown aspect of SARS-CoV-2 is its potential short- and long-term impact on the central nervous system. In this respect, although the rate of neurological symptoms associated with infection is not high (7), the possibility of subsequent effects of central nerve system (CNS) infection has been suggested by many authors (8–17). To address these questions, we need experimental models based on advances in biomedical research both in vitro and in vivo. We review these models in the present article.
Previous studies on SARS-CoV already demonstrated that the spike protein facilitates viral invasion of the target cell by interacting with the angiotensin converting enzyme 2 (ACE2) (18). This protein is expressed on the surface of epithelial cells of the lower respiratory tract, endothelial cells of arteries and veins, intestinal mucosa cells, kidney cells, immune cells, glial cells, and neurons (19–21). In vitro studies have demonstrated that SARS-CoV-2 also uses this receptor to penetrate cells (22, 23). In fact, sequencing of the SARS-CoV-2 spike protein has shown that it contains residues that may increase its binding affinity to the ACE2 receptor 10- to 20-fold in comparison with SARS-CoV (18). When spike protein interacts with the receptor, cleavage and activation by cell proteases is necessary to enable the viral membrane to fuse with the host cell and the virus to penetrate. In vitro studies have shown that SARS-CoV-2 spike proteins are mainly processed by transmembrane protease serine 2 (TMPRSS2), by endosomal cathepsins B and L in the absence of TMPRSS2 (23, 24), and by furin (25, 26). Once the virus is internalized, viral RNA is released into the cytoplasm, and a series of translations and replications create new RNAs and viral proteins that assemble to form new virions (20). These are released from the host cell and can infect new target cells expressing the appropriate receptor on their surfaces (20, 23).
More complex biological systems are, however, needed to study the possible interactions between SARS-CoV-2 and the host. Due to these complex interactions, selecting an appropriate model should be a thoughtful and clearly defined process, in order to provide relevant and translatable scientific data enabling us to address the questions of interest and to ensure the rational use of animals. Although many animals may respond similarly to humans from physiological, pathological, and therapeutic perspectives, we must bear in mind that differences between species may lead to erroneous conclusions (27); it is therefore necessary to understand the relationship between the model and the human disease (28). In the light of the current pandemic, it is essential to have a cellular or animal model mimicking the symptoms and pathological processes identified in patients infected with SARS-CoV-2 (29).
This cell line was isolated from kidney epithelial cells extracted from an African green monkey (Chlorocebus aethiops) in 1979; the Vero E6 cell line has been shown to be very useful for viral propagation and production in vitro (30). These cells are permissive to SARS-CoV replication, as they efficiently express the ACE2 receptor (31, 32). Furthermore, they enable persistent infection in vitro (33). Vero E6 cells have therefore already been used in studies with SARS-CoV and MERS-CoV (30, 33–36) and also in the development of live-attenuated and inactivated vaccines for human use (37, 38). More recently, they have been used in research into the viral infection mechanism in COVID-19 (23, 39, 40), the effects of the virus on cells, confirmation of viral infection (41–43) and pharmacological research (44–48).
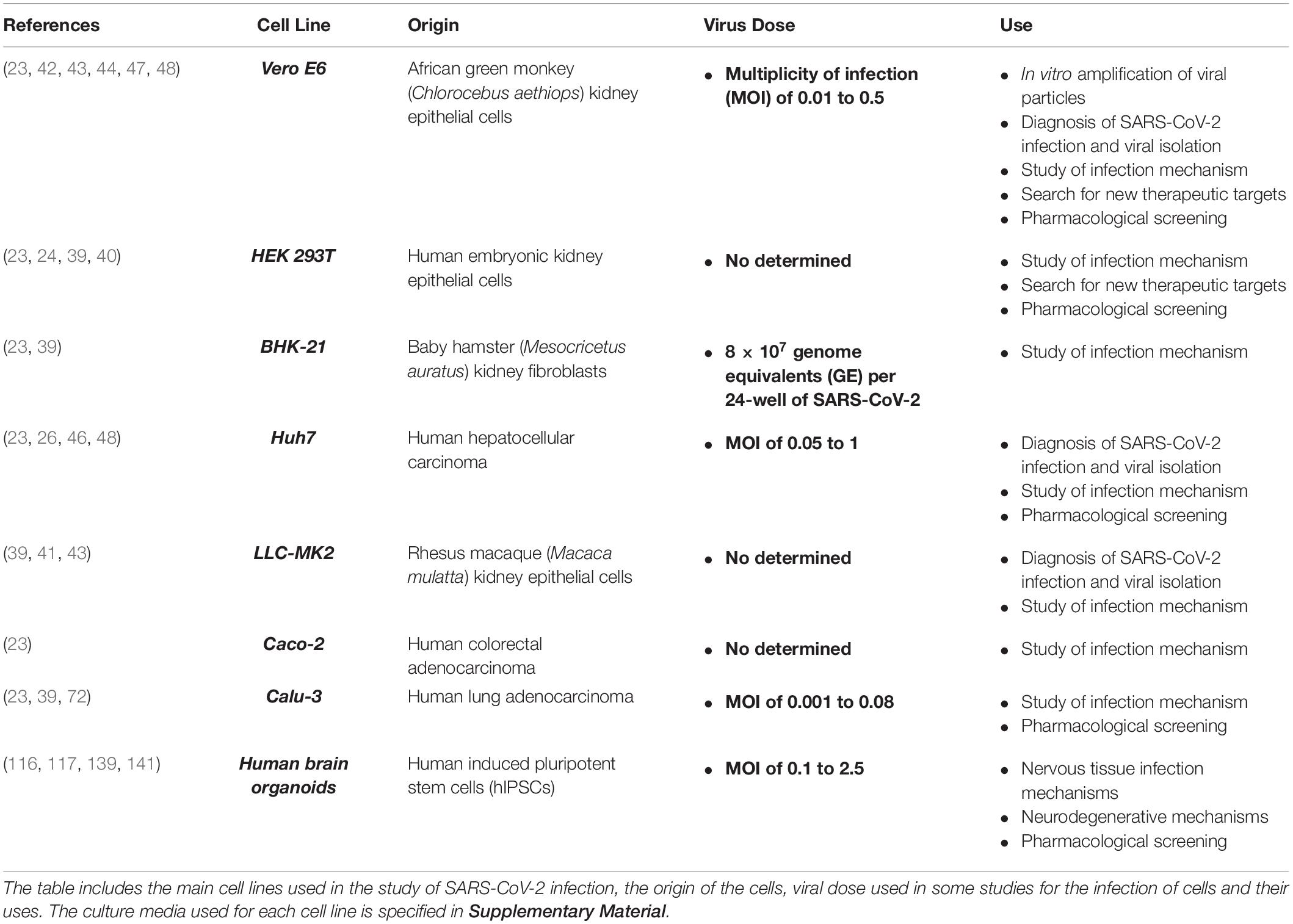
Table 1. In vitro models for the study of SARS-CoV-2.
This cell line is a variant of the HEK 293 lineage, which was isolated from kidney epithelial cells extracted from human embryos (49). Both lines are widely used in research due to their high transfectability, gene expression, and production of proteins or recombinant retrovirus (23, 49, 50). The HEK 293T variant expresses the T SV-40 antigen, which enables the amplification of transfected plasmids containing the SV40 origin of replication and thus considerably increases the expression levels of desired gene products (50). These cells have also previously been used in studies of other such viruses as herpes simplex virus, SARS-CoV, and even CoV pseudovirions (34, 51–53). Due to its high efficiency of transfection, research with this cell line is producing significant findings, such as the confirmation of ACE2 as the cell entry receptor for SARS-CoV-2 and the potential role of CD147 as an alternative receptor (23, 24, 39, 40).
This line is a subclone of the fibroblast cell line extracted from 1-day-old Syrian hamster (Mesocricetus auratus) kidney cells (54) and is useful for studying virus propagation and plasmid transfection (55). It has been used in the study of HCoV-OC43 and SARS-CoV-2 infection mechanisms (23, 36, 56, 57).
This lineage was established in 1982 from a human hepatocellular carcinoma; it is therefore able to produce a great variety of substances secreted by the human liver (58) and has been used in the study of hepatitis C virus, SARS-CoV, and MERS-CoV (27, 34, 58, 59). In the context of SARS-CoV-2, this cell line has been used in the study of infection mechanisms (23, 39), the cytotoxicity of viruses from human hosts (22, 60), and in pharmacological research (26, 46, 48).
The LLC-MK2 cell line was established in 1955 from rhesus macaque (Macaca mulatta) kidney epithelial cells (61). It has been mainly used in the study of poliovirus, but also to study SARS-CoV, HCoV-NL63 (34, 62, 63), and SARS-CoV-2 to assess the cytopathic effects in patients infected with the virus (41, 43) and to characterize its viral infectious pathway (39).
This line was extracted in 1977 from human epithelial colorectal adenocarcinoma cells; under specific culture conditions, the cells are able to differentiate into small intestine enterocytes (64). This cell line is frequently used to study transfection, invasion, and absorption (65). It has been used in studies of MERS-CoV, SARS-CoV, and HCoV-NL63 (27, 66, 67), and in the analysis of the S protein activation in studies with SARS-CoV-2, due to the fact that these cells express the TMPRSS2 protease (23).
This line was extracted in 1975 from a human lung adenocarcinoma and is used in the study of respiratory viruses (68, 69). It has been used in the study of SARS-CoV and MERS-CoV (27, 70, 71) and infection mechanisms studies and pharmacological research into COVID-19 (23, 39, 72).
Several of the available cell lines susceptible to transfection may also be used in in vitro research. For example, the study by Zhou et al. (22) uses the HeLa cell line, which does not endogenously express ACE2, and expression plasmids for human ACE2 (hACE2) in the study of SARS-CoV-2.
Mouse and hamster strains are the main animal models used in research into SARS-CoV and MERS-CoV (73); however, in the case of SARS-CoV-2, research efforts have been focused on the study of aged mouse strains, the design of humanized mouse models that express the hACE2 receptor, and the creation of knockout mice, with the aim of replicating the mechanisms involved in human infection.
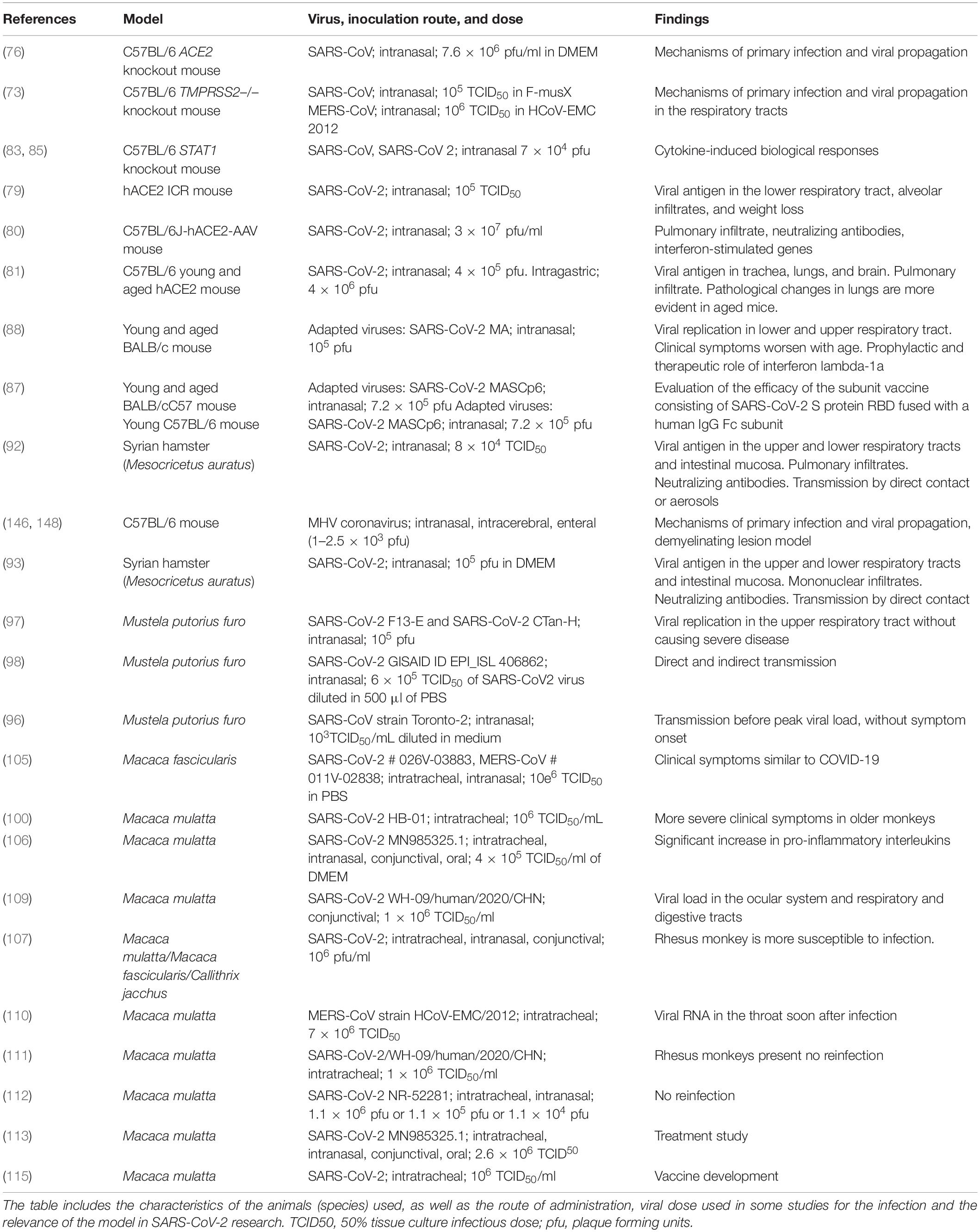
Table 2. In vivo models for the study of SARS-CoV-2 infection in the CNS.
Roberts et al. (74) reported that aged BALB/c mouse strains were able to maintain high rates of viral replication, which was associated with clinical illness and pneumonia; the study demonstrated an age-related susceptibility to SARS disease in animals that parallels the human experience. Advanced age has been identified as an independent factor of poor prognosis in COVID-19 and is considered a predictor of mortality in patients with SARS-CoV-2 infection. Other strains like C57BL/6 have also been used (75).
K18-hACE2 transgenic mice express hACE2, with regulation by the human cytokeratin 18 (K18) promoter. Specifically, these mice contain 2.5 kb of the K18 genomic sequence, including the promoter, the first intron, and a translation-promoting sequence. Expression of hACE2 is mainly observed in the epithelium of the respiratory tract, which shows a higher incidence of SARS-CoV-2 infection, and in the epithelium of other organs, including the liver, kidneys, and gastrointestinal tract (76). K18-hACE2 mice infected with SARS-CoV-2 show weight loss and viral replication in the lungs, as in humans. Furthermore, they show the typical histopathological findings of interstitial pneumonia with lymphocytic and monocytic infiltration into the alveolar interstitium and an accumulation of macrophages in the alveolar cavities. Alveolar and bronchial epithelial cells show presence of viral antigens; this is not observed in wild-type (WT) mice with SARS-CoV-2 infection. Furthermore, these mice show other pathological changes, such as vasculitis, degeneration, and necrosis of extrapulmonary organs and presence of the viral antigen in the brain (77, 78). Bao et al. (79) recently studied the pathogenicity of SARS-CoV-2 virus in hACE2-expressing ICR transgenic mice and WT mice. This model may be useful in research into drug treatments for COVID-19. Israelow et al. (80) developed an hACE2-adeno-associated virus 9 (AAV) murine model, which was subsequently intranasally infected with SARS-CoV-2. They studied the presence of coronavirus infection, the inflammatory response in the lungs, the presence of neutralizing antibodies, and the type I interferon signaling pathway. The authors reported that this model would be very useful for understanding questions related to infection, replication, and pathogenesis of SARS-CoV-2 and for testing therapeutic strategies.
Sun et al. (81) used CRISPR/Cas9 technology to create a murine hACE2 model in young and aged animals. After intranasal infection of the animals with SARS-CoV-2, the researchers observed viral replication in the lungs, trachea, and brain of both animals. However, alveolar inflammatory infiltrate and vascular lesions were more evident in aged animals; this is analogous to the pathological changes observed in older patients with COVID-19. This model also showed evidence of respiratory tract infection following intragastric inoculation.
This model was designed using a directional vector to replace exons 10–13, which codify the serine protease domain of the TMPRSS2 gene. It was constructed by electroporation of embryonic stem cells and their subsequent injection into C57BL/6 blastocysts for at least five generations (82). After experimental infection with SARS-CoV-2, TMPRSS2-deficient mouse strains showed reduced body weight loss and viral kinetics in the lungs. Absence of TMPRSS2 affected the infection sites and virus spread within the respiratory tract; therefore, this is a useful model for COVID-19 research (73).
(129S6/SvEv-STAT1tmRDS) contains a homozygous STAT1 mutation and completely lacks functional STAT1 proteins (Pgm1c and Gpi1b alleles of 129S6). The model was created by targeting the STAT1 gene in GS-1 ES cells and injecting target cells into blastocysts. Heterozygous models of the mutation were produced from the chimeras and were crossed over to generate homozygous models (83, 84). The JAK-STAT signaling pathway is involved in the mediation of cytokine-induced biological responses. This is therefore a useful model for determining the role of a variety of cytokines in immune responses, the role of STAT1 protein in mediating interferon-dependent responses, and its relationship with viral and bacterial pathogens (84–86); the model is also interesting in the analysis of SARS-CoV-2 inflammation mechanisms.
Studies have been recently conducted with BALB/c and C57BL/6 mice (87, 88) together with modified SARS-CoV-2 strains. Modification of the virus has led to mouse-adapted SARS-CoV-2 strains, the SARS-CoV-2 MA (88) and SARS-CoV-2 MASCp6 (87) strains, which are able to infect mice with no need for modification of the animals, as these strains efficiently bind to the murine ACE2 receptor in both young and aged mice, causing a disease resembling human COVID-19. In addition to their use in the study of pathogenesis, these models have enabled researchers to trial vaccines and treatments for the disease (87, 88).
BALB/c mice have been used in studies for the development of vaccines, and Wistar rats have been used in studies into immunization with attenuated strains of the virus (89).
This animal model (M. auratus) has previously been used in the study of SARS-CoV (90, 91), as the hamster presents an ACE2 receptor homologous to the human receptor. Sia et al. (92) intranasally inoculated animals with the β–CoV/Hong Kong/VM20001061/2020 strain and corroborated the presence of viral antigen in the epithelial cells of the nasal and bronchial mucosa with progression to pneumocytes and clearance of infectious particles by day 7 after infection. Presence of mononuclear cell infiltrates was moderate in the nasal turbinates but was greater in the lungs. The viral antigen was observed in epithelial cells of the duodenum, without signs of inflammation. No infectious particles were detected in the kidney, and no histopathological changes were observed in other organs. Neutralizing antibodies were observed on day 14 after infection. Infected animals presented clear weight loss and researchers corroborated that the main route of viral transmission was through direct contact or aerosols. The authors conclude that SARS-CoV-2 infection in Syrian hamsters presents similar characteristics to those observed in humans with mild infections and that this model represents an opportunity for understanding the transmission dynamics of this novel coronavirus.
Chan et al. (93) confirmed transmission by contact, the progressive decrease in viral load between days 2 and 7 after infection in both lower and upper respiratory tracts, and the expression of viral antigens (protein N) by epithelial cells together with presence of mononuclear infiltrates. On day 7 after infection, regenerative hyperplasia occurring in bronchioles led to the appearance of multiple irregularly arranged epithelial layers. Furthermore, they detected the viral antigen in the intestinal mucosa and observed histopathological changes in the spleen and heart. The pro-inflammatory cytokine cascade in this model normalizes at approximately day 7, and antibodies are observed at 7–14 days. This animal model reproduces the respiratory and enteric symptoms observed in patients with COVID-19.
Ferrets (Mustela putorius furo) are frequently used as animal models to study respiratory diseases caused by such viruses as influenza virus or SARS-CoV (94–96). All studies with SARS-CoV-2 in ferrets have used the intranasal route to inoculate the virus (97–99). Shi et al. (97) studied the susceptibility of ferrets to infection with 2 SARS-CoV-2 strains. In a first stage of the study, the authors detected viral RNA in samples from the nasal turbinates, palate, and tonsils, but no viral load was detected in the other organs analyzed, including the lungs and brain. This suggests that the virus can replicate in the upper respiratory tract of ferrets. In a second stage of the study, analyzing the viral replication dynamics, animals showed viral RNA in nasal washes, whereas rectal swabs showed much lower viral load. Furthermore, ferrets presented anti-SARS-CoV-2 antibodies at days 13 and 20. Histopathological studies showed altered pneumocytes, macrophages, and neutrophils. Finally, when studying replication in the lungs, the authors observed that the virus is able to replicate in the upper respiratory tract up to day 8 after infection without causing severe symptoms. Richard et al. (98) focused on the study of the direct and indirect transmission of the SARS-CoV-2. Ferrets inoculated with SARS-CoV-2 were placed in direct contact with a group of healthy ferrets 6 h after infection. Inoculated animals showed productive infection, and the peak of viral infection was reached on the third day. All animals in direct or indirect contact presented viral RNA at day 1–3 after exposure. Animals exposed to inoculated ferrets were expected to present a lower viral load than the inoculated animals; however, viral load was similar in all cases. Lastly, the authors observed that all animals presented antibodies at day 21 after exposure to the virus, and that their levels were similar, regardless of the form of infection. A similar design was employed by Kim et al. (99), who observed viral load up to 8 days after infection, both in samples from inoculated animals and in samples from animals that were in direct contact with the inoculated ferrets. Animals in indirect contact with infected ferrets showed positive results for infection at day 2 after exposure, which suggests rapid transmission, even before peak viral load was reached and in the absence of clinical symptoms; this correlates with the reported transmission by asymptomatic human patients.
Severe acute respiratory syndrome coronavirus and middle east respiratory syndrome coronavirus have previously been studied in non-human primates (75, 100–104); the use of these models has been proposed for the study of SARS-CoV-2 (105) especially with a view to the development of vaccines or antiviral treatments (106, 107). Rockx et al. (105) used a combined intratracheal and intranasal route to inoculate the SARS-CoV-2 virus to both young and old adult cynomolgus macaques (Macaca fascicularis). The results of this study showed that these animals tolerated viral infection, presenting symptoms similar to those of COVID-19 in humans. The virus efficiently replicates in epithelial cells throughout the upper respiratory tract, which correlates with the ease of transmission of the virus, whereas replication in the lower respiratory tract correlates with the development of the disease (105–108). Yu et al. (100) obtained similar results with intratracheal inoculation of young and old rhesus monkeys (Macaca mulatta). The authors report that, although viral replication was more active in older adult monkeys, both groups developed interstitial pneumonia together with edema, which was more severe and diffuse in older adult monkeys. Munster et al. (105) developed a COVID-19 model using rhesus monkeys, inoculating them with the virus through four different but combined routes (intranasal, intratracheal, conjunctival, and oral). The animals presented clinical signs compatible with COVID-19 from day 1 after the inoculation to symptom resolution, between days 9 and 17. All animals presented weight loss, low-grade fever, and pulmonary infiltrates, in addition to a significant increase in IL-6 and IL-10, among other findings; IgG antibodies were present at detectable levels from day 7 after the infection. Nasal, pharyngeal, and rectal samples showed high levels of viral RNA. Furthermore, histopathological analyses showed similar alterations to those caused by SARS-CoV and MERS-CoV (97, 98). Deng et al. (109) inoculated rhesus monkeys with SARS-CoV-2 through the conjunctival route. They did not observe significant changes in the animals’ weight or body temperature. Antibody analyses detected presence of IgG at days 14 and 21. Furthermore, radiographs of these animals showed bilateral alterations in the upper lobes and right lower lobe, which correlates with the moderate interstitial pneumonia observed microscopically. Histopathological studies showed viral load in the ocular and nasolacrimal system, as well as in the respiratory and digestive tracts. Results of a comparative study of three non-human primate species [rhesus monkey, crab-eating macaque, and common marmoset (Callithrix jacchus)], which were inoculated with the virus through three different pathways (intratracheal, intranasal, and conjunctival), showed that almost all animals presented clinical signs compatible with COVID-19, although there were differences between species in the histopathological findings (107). The rhesus monkey is the most susceptible species to viral infection, and therefore a good model for the study of COVID-19, as well as for the development of vaccines and pharmacological studies (107, 108). De Wit et al. (110) studied rhesus monkeys intratracheally inoculated with the virus, finding that viral RNA was detectable in the throat during the first days, but with levels subsequently decreasing until becoming undetectable. Bao et al. (111) and Chandrashekar et al. (112) intratracheally and intranasally inoculated rhesus monkeys with the SARS-CoV-2 virus to study the possibility of reinfection. Williamson et al. (113) used rhesus monkeys that had previously been inoculated through the intranasal, oral, ocular, and intratracheal routes for pharmacological research. These animals have also been used for the development of vaccines (106, 107, 114, 115).
Coronaviruses have been found in autopsy studies in the CNS of patients with multiple sclerosis, Parkinson’s disease, and Alzheimer disease. Experimental studies have shown that human CoVs can infect neurons, astrocytes, and microglia in primary cultures as well as immortalized human microglial cells (6). The suggestion that SARS-CoV-2 may use the brain as a reservoir (10), potentially favoring the development of neurodegenerative diseases (11), underscores the need to specifically analyze the effect of the virus on the CNS.
The information currently available is from research on SARS-CoV, which has 78% nucleotide homology with SARS-CoV-2 (18). Although CNS tropism has been described (20), no specific models of neural cell lines have been developed for the study of SARS-CoV-2; however, neural progenitor cells (NPCs), neurons and microglia derived from human induced pluripotent stem cells (hIPSCs) have already been used in in vitro studies of SARS-CoV-2 viral infection, demonstrating the virus potential to infect CNS cells (116–118). Previous studies with other neurotropic human coronaviruses showed possible neural cell lines susceptible to infection by SARS-CoV-2 that may be useful in studying the possible mechanisms by which the virus infects the CNS. Studies into neural susceptibility to SARS-CoV infection have used cell lines including HOG, a line derived from a human oligodendroglioma that expresses proteins characteristic of oligodendrocytes and is very widely used in the study of neurons (119, 120) and the C6 cell line, derived from a glioma induced in Wistar Furth rats exposed to N-nitroso-N-methylurea, which is morphologically similar to glioblastoma multiforme when injected into the brains of neonate rats (121, 122). Although both cell lines have been shown to be susceptible to SARS-CoV infection, low levels of viral replication have been observed in comparison with such other susceptible cell lines as Vero E6 or Caco-2 (123). Other cell lines used to study the virulence of HCoV-229E and HCoV-OC43 in the CNS include human H4 brain neuroglioma cells, the LA-N-5 human neuroblastoma cell line, the CHME-5 human fetal microglia cell line, and the U-373 MG and U-87 MG astrocytic lines derived from a human glioblastoma and an astrocytoma, respectively, among many others (124–129). Cultures of human primary neurons, astrocytes, oligodendrocytes, and microglia have been used to study these viruses (130, 131). All these neural cell lines may be useful in the near future to study SARS-CoV-2.
Organoids are miniaturized, simplified, three-dimensional versions of an organ produced in vitro, partially recreating the cellular structure and the functioning of that organ (132, 133). Classic cell culture systems present certain limitations, such as the inability to study complex and dynamic responses or cell-cell interactions. The use of organoids enables us to study complex physiological or pathological processes in structures bearing much greater resemblance to in vivo conditions, including SARS-CoV-2 infection, tropisms and potential treatments (133–135). To date, SARS-CoV-2 infection has been studied in human organoids of lung, liver, intestine, blood vessels, and kidney (42, 118, 136–138). Human brain organoids have also been used; these present strong cellular and structural similarities to some mammalian brain regions, such as a neural epithelium containing NPCs, that align to form a ventricular zone-like layer, cortical neurons, that contribute to the formation of a cortical plate-like layer and glial cells, such as astrocytes or oligodendrocytes (139–141). These organoids are useful for the study of early stages of human neurodevelopment and network formation, key cellular processes such as proliferation, differentiation, apoptosis, synaptogenesis or myelination, CNS function such as electrophysiological activity, neurodegenerative diseases, potential treatments, and have been already used for the study of other virus such as ZIKA virus or HIV (139, 141–144).
Ramani et al. (116) observed that in these brain organoids, the virus mainly infects mature cortical neurons and presents a perinuclear distribution within these cells. Furthermore, neurodegenerative effects have been observed in cells infected by SARS-CoV-2, including cell death and hyperphosphorylation, as well as mislocation of Tau protein; these alterations are observed in such conditions as tauopathies or Alzheimer disease (116). However, no productive replication of the virus was observed in these cells, at least in the first 4 days after infection (116), which would support the hypothesis that the CNS may act as a long-term reservoir of the virus (10). In contrast, Bullen et al. (139) observed an incremented accumulation of viral particles in neural cells of brain organoids between 6 and 72 h after SARS-CoV-2 infection, suggesting an active replication and productive infection of the virus in neural cells during the first days. Viral particles were detected mainly in the neuronal soma and, in some cells, also into the neurites (139). Similar to Ramani et al., Mesci et al. (141) used these brain organoids and observed that the virus was able to infect neurons, including NPCs and mature cortical neurons, and cause cell death accompanied by the impairment of excitatory synapses. Furthermore, this work tested the efficiency of Sofosbuvir, an FDA-approved brain-penetrant antiviral drug for positive-sense single-stranded RNA viruses (145), as a treatment for the SARS-CoV-2 infection and observed that this drug was able to rescue the altered synaptogenesis and decrease neuronal death and viral accumulation in these brain organoids (141). Song et al. (117) also demonstrated that SARS-CoV-2 has neuroinvasive capacity in human brain organoids, particularly of NPCs and mature cortical neurons. Infected cells showed a hypermetabolic state and viral particles were accumulated within endoplasmic reticulum-like structures, indicating the virus ability to use the neural cell machinery to replicate (117). In addition, a hypoxic environment and extensive neuronal cell death were observed in high density SARS-CoV-2 infected areas, suggesting that virus infection could promote death of nearby cells (117). Finally, this study detected IgG antibodies against SARS-CoV-2 in the cerebrospinal fluid of a COVID-19 patient that were able to block SARS-CoV-2 infection in brain organoids (117). All these studies show that SARS-CoV-2 can directly infect neural cells and trigger damaging consequences that could cause neurologic symptoms. Also, these studies expose the great potential of the human brain organoids for the study of the SARS-CoV-2 effects in the CNS.
Mouse hepatitis virus (MHV) is a βCoV that poses no risk to humans but presents a great similarity with other viruses from the same family, such as SARS-CoV, MERS-CoV, and SARS-CoV-2. It penetrates the CNS, causing white matter lesions; it has therefore been proposed as a viral model of demyelinating disease (146, 147). The virus has been shown to remain in the white matter and to be able to replicate in the CNS (148); therefore, it is a good model for the study of CNS infection by coronaviruses. Neurotropic strains of MHV-CoV have been used extensively to induce acute and chronic demyelinating disease mediated by neuroinflammation (149). Depending on the inoculation route and the MHV-CoV strain, different CNS regions are affected. Inoculation with experimental neurotropic strains, especially MHV-A59, induces a biphasic disease of acute meningoencephalitis at 10–14 days after inoculation, followed by a disease causing subacute, chronic inflammatory demyelination in the brain; spinal cord involvement is more pronounced (150). Virus translocation from the initial site of inoculation in the brain to the spinal cord is caused by the transit of virus particles in neural and glial cells, as well as mechanisms that involve the fusion of lipid membranes, probably during the virus internalization step (151). Intranasal and intracranial inoculation of JHM-CoV induces similar symptoms in BALB/c mice to those caused by MHV-A59. After intranasal inoculation of mice, MHV-CoV accesses the CNS through the olfactory nerve and propagates from the olfactory system to limbic system structures and their connections with the brainstem (152).
In order to study the immune system role in demyelination induction caused by MHV infection, Wang et al. (153) treated infected animals with gamma radiation to cause immunosuppression and, subsequently, reconstituted immunity by transferring cells from other immunocompetent animals. The results showed that demyelination was prevented by radiation and was present again when the immunity was restored, indicating that immunity is directly involved in the demyelination process (153). Moreover, CD4 and CD8 T cells have been observed to play a critical role in the development of the demyelinating process, with γδ T cells being the most important for this process (154, 155). In contrast, B cells, the most abundant cell type in the spleen, and NK cells are not involved in demyelination as nude animals without spleen do not present demyelination (154), MHV offers a unique model for studying host defense-mediated demyelination during chronic infection in a phase acute viral infection and immune response (156).
Research on SARS-CoV-2 has become a necessity due to the magnitude of its spread worldwide. Such aspects as infection rate and mechanisms, the possibility of reinfection, and possible therapeutic approaches make it necessary not only to use the experimental models previously employed to study the SARS-CoV and MERS-CoV coronaviruses, but also to generate new models to respond to urgent questions. The potential involvement of the CNS due to SARS-CoV-2 infection should be studied specifically, and research efforts must focus on obtaining information with cellular or animal models to expand our understanding of the virus.
JM-G and UG-P: lead researchers. All authors: research project group, manuscript drafting, and critical review of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors would like to thank the Spanish Society of Neurology’s Research Operations Office for assisting with the English-language version of the manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2020.02163/full#supplementary-material
1. Rothan HA, Byrareddy, SN. The epidemiology and pathogenesis of coronavirus disease (COVID-19) outbreak. J Autoimmun. (2020) 109:102433. doi: 10.1016/j.jaut.2020.102433
2. Mackenzie JS, Smith, DW. COVID-19: a novel zoonotic disease caused by a coronavirus from China: what we know and what we don’t [published online ahead of print, 2020 Mar 17]. Microbiol Aust. (2020) 41:MA20013. doi: 10.1071/MA20013
3. Tang X, Wu C, Li X, Song Y, Yao X, Wu X, et al. On the origin and continuing evolution of SARS-CoV-2. Natl Sci Rev. (2020) 2020:1012–23. doi: 10.1093/nsr/nwaa036
4. Forster P, Forster L, Renfrew C, Forster, M. Phylogenetic network analysis of SARS-CoV-2 genomes. Proc Natl Acad Sci USA. (2020) 117:9241–3. doi: 10.1073/pnas.2004999117
5. Carneiro LJ, Paula de Lima Gusmão T, Machado ZA, Leao FJ, Barkokebas Santos de Faria A, Morais SIH, et al. Coronaviridae - old friends, new enemy! Oral Dis. (2020). doi: 10.1111/odi.13447 [Epub ahead of print].
6. Rabaan AA, Al-Ahmed SH, Haque S, Sah R, Tiwari R, Malik YS, et al. SARS-CoV-2, SARS-CoV, and MERS-COV: a comparative overview. Infez Med. (2020) 28:174–84.
7. Matías-Guiu J, Gomez-Pinedo U, Montero-Escribano P, Gomez-Iglesias P, Porta-Etessam J, Matias-Guiu, JA. ?‘Es esperable que haya cuadros neurológicos por la pandemia por SARS-CoV-2? Neurologia. (2020) 35:170–75. doi: 10.1016/j.nrl.2020.03.001
8. Paniz-Mondolfi A, Bryce C, Grimes Z, Gordon RE, Reidy J, Lednicky J, et al. Central nervous system involvement by severe acute respiratory syndrome coronavirus -2 (SARS-CoV-2). J Med Virol. (2020). doi: 10.1002/jmv.25915 [Epub ahead of print].
9. Bulfamante G, Chiumello D, Canevini MP, Priori A, Mazzanti M, Centanni S, et al. First ultrastructural autoptic findings of SARS-Cov-2 in olfactory pathways and brainstem. Minerva Anestesiol. (2020): doi: 10.23736/S0375-9393.20.14772-2 [Epub ahead of print].
10. Gomez-Pinedo U, Matias-Guiu J, Sanclemente-Alaman I, Moreno-Jimenez L, Montero-Escribano P, Matias-Guiu, JA. Is the brain a reservoir organ for SARS-CoV2? J Med Virol. (2020). doi: 10.1002/jmv.26046 [Epub ahead of print].
11. Gomez-Pinedo U, Matias-Guiu J, Sanclemente-Alaman I, Moreno-Jimenez L, Montero-Escribano P, Matias-Guiu, JA. SARS-CoV2 as a potential trigger of neurodegenerative diseases. Mov Disord. (2020). doi: 10.1002/mds.28179 [Epub ahead of print].
12. Cheng Q, Yang Y, Gao, J. Infectivity of human coronavirus in the brain [published online ahead of print, 2020 May 28]. EBioMedicine. (2020) 56:102799. doi: 10.1016/j.ebiom.2020.102799
13. Pinzon RT, Wijaya VO, Buana RB, Al Jody A, Nunsio, PN. Neurologic Characteristics in coronavirus disease 2019 (COVID-19): a systematic review and meta-analysis. Front Neurol. (2020) 11:565. doi: 10.3389/fneur.2020.00565
14. Fiani B, Covarrubias C, Desai A, Sekhon M, Jarrah, RA. Contemporary Review of Neurological Sequelae of COVID-19. Front Neurol. (2020) 11:640. doi: 10.3389/fneur.2020.00640
15. Huang J, Zheng M, Tang X, Chen Y, Tong A, Zhou, L. Potential of SARS-CoV-2 to cause CNS infection: biologic fundamental and clinical experience. Front Neurol. (2020) 11:659. doi: 10.3389/fneur.2020.00659
16. Torre-Fuentes L, Matías-Guiu J, Hernández-Lorenzo L, Montero-Escribano P, Pytel V, Porta-Etessam J, et al. ACE2, TMPRSS2, Furin variants and SARS-CoV-2 infection in Madrid, Spain. J Med Virol. (2020). doi: 10.1002/jmv.26319 [Epub ahead of print].
17. Gómez-Iglesias P, Porta-Etessam J, Montalvo T, Valls-Carbó, A Gajate, V, Matías-Guiu JA, et al. An online observational study of patients with olfactory and gustory alterations secondary to SARS-CoV-2 infection. Front Public Health. (2020) 8:243. doi: 10.3389/fpubh.2020.00243
18. Wan Y, Shang J, Graham R, Baric RS, Li, F. Receptor recognition by the novel coronavirus from wuhan: an analysis based on decade-long structural studies of SARS coronavirus. J Virol. (2020) 94:e00127-20. doi: 10.1128/JVI.00127-20
19. Lukassen S, Chua RL, Trefzer T, Kahn NC, Schneider MA, Muley T, et al. SARS-CoV-2 receptor ACE2 and TMPRSS2 are primarily expressed in bronchial transient secretory cells. EMBO J. (2020) 39:e105114. doi: 10.15252/embj.20105114
20. Song Z, Xu Y, Bao L, Zhang L, Yu P, Qu Y, et al. From SARS to MERS, thrusting coronaviruses into the spotlight. Viruses. (2019) 11:59. doi: 10.3390/v11010059
21. Baig AM, Khaleeq A, Ali U, Syeda, H. Evidence of the COVID-19 Virus Targeting the CNS: tissue distribution, host-virus interaction, proposed neurotropic mechanisms. ACS Chem Neurosci. (2020) 11:995–98. doi: 10.1021/acschemneuro.0c00122
22. Zhou P, Yang XL, Wang XG, Hu B, Zhang L, Zhang W, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. (2020) 579:270–73. doi: 10.1038/s41586-020-2012-7
23. Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, et al. SARS-CoV-2 Cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell. (2020) 181:271–80.e8. doi: 10.1016/j.cell.2020.02.052
24. Zhu Y, Yu D, Yan H, Chong H, He, Y. Design of potent membrane fusion inhibitors against SARS-CoV-2, an emerging coronavirus with high fusogenic activity. J Virol. (2020) 94:e00635-20. doi: 10.1128/JVI.00635-20
25. Coutard B, Valle C, de Lamballerie X, Canard B, Seidah NG, Decroly, E. The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade. Antiviral Res. (2020) 176:104742. doi: 10.1016/j.antiviral.2020.104742
26. Xia S, Liu M, Wang C, Xu W, Lan Q, Feng S, et al. Inhibition of SARS-CoV-2 (previously 2019-nCoV) infection by a highly potent pan-coronavirus fusion inhibitor targeting its spike protein that harbors a high capacity to mediate membrane fusion. Cell Res. (2020) 30:343–55. doi: 10.1038/s41422-020-0305-x
27. Chan JF, Chan KH, Choi GK, To KW, Tse H, Cai JP, et al. Differential cell line susceptibility to the emerging novel human betacoronavirus 2c EMC/2012: Implications for disease pathogenesis and clinical manifestation. Infest Dis. (2013) 207:1743–52. doi: 10.1093/infdis/jit123
28. Swearengen, JR. Choosing the right animal model for infectious disease research. Animal Model Exp Med. (2018) 1:100–08. doi: 10.1002/ame2.12020
29. Lai CC, Shih TP, Ko WC, Tang HJ, Hsueh, PR. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and coronavirus disease-2019 (COVID-19): The epidemic and the challenges. Int J Antimicrob Agents. (2020) 55:105924. doi: 10.1016/j.ijantimicag.2020.105924
30. Guo D, Zhu Q, Zhang H, Sun, D. Proteomic analysis of membrane proteins of vero cells: exploration of potential proteins responsible for virus entry. DNA Cell Biol. (2014) 33:20–8. doi: 10.1089/dna.2013.2193
31. Li W, Moore M, Vasilieva N, Sui J, Wong SK, Berneet MA, et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature. (2003) 426:450–4. doi: 10.1038/nature02145
32. Ng ML, Tan SH, See EE, Ooi EE, Ling, AE. Early events of SARS coronavirus infection in vero cells. J Med Virol. (2003) 71:323–31. doi: 10.1002/jmv.10499
33. Yamate M, Yamashita M, Goto T, Tsuji S, Li YG, Warachit J, et al. Establishment of vero E6 cell clones persistently infected with severe acute respiratory syndrome coronavirus. Microbes Infect. (2005) 7:1530–40. doi: 10.1016/j.micinf.2005.05.013
34. Kaye, M. SARS-associated coronavirus replication in cell lines. Emerg Infect Dis. (2006) 12:128–33. doi: 10.3201/eid1201.050496
35. Lin SC, Ho CT, Chuo WH, Li S, Wang TT, Lin, CC. Effective inhibition of MERS-CoV infection by resveratrol. BMC Infect Dis. (2017) 17:144. doi: 10.1186/s12879-017-2253-8
36. Sudeep AB, Vyas PB, Parashar D, Shil, P. Differential susceptibility & replication potential of Vero E6, BHK-21, RD, A-549, C6/36 cells & Aedes aegypti mosquitoes to three strains of chikungunya virus. Indian J Med Res. (2019) 149:771–7. doi: 10.4103/ijmr.IJMR_453_17
37. Barrett PN, Mundt W, Kistner O, Howard, MK. Vero cell platform in vaccine production: moving towards cell culture-based viral vaccines. Expert Rev Vaccines. (2009) 8:607–18. doi: 10.1586/erv.09.19
38. Chen HL, Chang JK, Tang, RB. Current recommendations for the Japanese encephalitis vaccine. J Chin Med Assoc. (2015) 78:271–75. doi: 10.1016/j.jcma.2014.12.009
39. Ou X, Liu Y, Lei X, Li P, Mi D, Ren L, et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat Commun. (2020) 11:1620. doi: 10.1038/s41467-020-15562-9
40. Wang K, Chen W, Zhou Y, Lian J, Zhang Z, Du P, et al. SARS-CoV-2 invades host cells via a novel route: CD147-spike protein. bioRxiv. (2020). [Preprint]. doi: 10.1101/2020.03.14.988345
41. Liu WD, Chang SY, Wang JT, Tsai MJ, Hung CC, Hsuet CL, et al. Prolonged virus shedding even after seroconversion in a patient with COVID-19. J Infect. (2020). doi: 10.1016/j.jinf.2020.03.063 [Epub ahead of print].
42. Zhou J, Li C, Liu X, Chiu MC, Zhao X, Wang D, et al. Infection of bat and human intestinal organoids by SARS-CoV-2. Nat Med. (2020) 26:1077–83. doi: 10.1038/s41591-020-0912-6
43. Colson P, Lagier JC, Baudoin JP, Bou Khalil J, La Scola B, Raoult, D. Ultrarapid diagnosis, microscope imaging, genome sequencing, and culture isolation of SARS-CoV-2. Eur J Clin Microbiol Infect Dis. (2020). doi: 10.1007/s10096-020-03869-w [Epub ahead of print].
44. Caly L, Druce JD, Catton MG, Jans DA, Wagstaff, KM. The FDA-approved drug ivermectin inhibits the replication of SARS-CoV-2 in vitro [published online ahead of print, 2020 Apr 3]. Antiviral Res. (2020) 178:104787. doi: 10.1016/j.antiviral.2020.104787
45. Liu J, Cao R, Xu M, Wang X, Zhang H, Hu H, et al. Hydroxychloroquine, a less toxic derivative of chloroquine, is effective in inhibiting SARS-CoV-2 infection in vitro. Cell Discov. (2020) 6:16. doi: 10.1038/s41421-020-0156-0
46. Wang M, Cao R, Zhang L, Yang X, Liu J, Xu M, et al. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. (2020) 30:269–71. doi: 10.1038/s41422-020-0282-0
47. Yao X, Ye F, Zhang M, Cui C, Huang B, Niu P, et al. In vitro antiviral activity and projection of optimized dosing design of hydroxychloroquine for the treatment of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) [published online ahead of print, 2020 Mar 9]. Clin Infect Dis. (2020) 71:ciaa237. doi: 10.1093/cid/ciaa237
48. Runfeng L, Yunlong H, Jicheng H, Weiqi P, Qinhai M, Yongxiaet S, et al. Lianhuaqingwen exerts anti-viral and anti-inflammatory activity against novel coronavirus (SARS-CoV-2). Pharmacol Res. (2020) 156:104761. doi: 10.1016/j.phrs.2020.104761
49. Stepanenko AA, Dmitrenko, VV. HEK293 in cell biology and cancer research: phenotype karyotype, tumorigenicity, and stress-induced genome-phenotype evolution. Gene. (2015) 569:182–90. doi: 10.1016/j.gene.2015.05.065
50. He N, Sun Y, Yang M, Lu Q, Wang J, Xiao C, et al. Analysis of circular RNA expression profile in HEK 293T cells exposed to ionizing radiation. Dose Response. (2019) 17:1559325819837795. doi: 10.1177/1559325819837795
51. Millet JK, Tang T, Nathan L, Jaimes J, Hsu H, Dan S, et al. Production of pseudotyped particles to study highly pathogenic coronaviruses in a biosafety level 2 setting. J Vis Exp. (2019) 1:145. doi: 10.3791/59010
52. Wan W, Wang L, Chen X, Zhu S, Shang W, Xiao G, et al. A subcellular quantitative proteomic analysis of herpes simplex virus type 1-infected HEK 293T Cells. Molecules. (2019) 24:4215. doi: 10.3390/molecules24234215
53. Yuan Y, Wang Z, Tian B, Zhou M, Fu ZF, Zhao, L. Cholesterol 25-hydroxylase suppresses rabies virus infection by inhibiting viral entry. Arch Virol. (2019) 164:2963–74. doi: 10.1007/s00705-019-04415-6
54. Capstick PB, Telling RC, Chapman WG, Stewart, DL. Growth of a cloned strain of hamster kidney cells in suspended cultures and their susceptibility to the virus of foot-and-mouth disease. Nature. (1962) 195:1163–64. doi: 10.1038/1951163a0
55. Hernandez R, Brown, DT. Growth and maintenance of baby hamster kidney (BHK) cells. Curr Protoc Microbiol. (2010) 17:A.4H.1–7. doi: 10.1002/9780471729259.mca04hs17
56. Amadori M, Volpe G, Defilippi P, Berneri, C. Phenotypic features of BHK-21 cells used for production of foot-and-mouth disease vaccine. Biologicals. (1997) 25:65–73. doi: 10.1006/biol.1996.0061
57. Shen L, Yang Y, Ye F, Liu G, Desforges M, Talbot PJ, et al. Safe and sensitive antiviral screening platform based on recombinant human coronavirus OC43 expressing the luciferase reporter gene. Antimicrob Agents Chemother. (2016) 60:5492–503. doi: 10.1128/AAC.00814-16
58. Kasai F, Hirayama N, Ozawa M, Satoh M, Kohara, A. HuH-7 reference genome profile: complex karyotype composed of massive loss of heterozygosity. Hum Cell. (2018) 31:261–67. doi: 10.1007/s13577-018-0212-3
59. Behnam MA, Nitsche C, Boldescu V, Klein, CD. The Medicinal chemistry of dengue virus. J Med Chem. (2016) 59:5622–49. doi: 10.1021/acs.jmedchem.5b01653
60. Ciotti M, Angeletti S, Minieri M, Giovannetti M, Benvenuto D, Pascarella S, et al. COVID-19 outbreak: an overview [published online ahead of print, 2020 Apr 7]. Chemotherapy. (2020) 64:215–23. doi: 10.1159/000507423
61. Evans VJ, Kerr HA, McQuilkin WT, Earle WR, Hull, RN. Growth in vitro of a long-term strain of monkey-kidney cells in medium NCTC 109 free of any added protein. Am J Hyg. (1959) 70:297–302. doi: 10.1093/oxfordjournals.aje.a120078
62. Hull RN, Cherry WR, Tritch, OJ. Growth characteristics of monkey kidney cell strains LLC-MK1, LLC-MK2, and LLC-MK2(NCTC-3196) and their utility in virus research. J Exp Med. (1962) 115:903–18. doi: 10.1084/jem.115.5.903
63. Milewska A, Nowak P, Owczarek K, Szczepanski A, Zarebski M, Hoang A, et al. Entry of human coronavirus NL63 into the cell. J Virol. (2018) 92:e01933-17. doi: 10.1128/JVI.01933-17
64. Hidalgo IJ, Raub TJ, and Borchardt, RT. Characterization of the human colon carcinoma cell line (Caco-2) as a model system for intestinal epithelial permeability. Gastroenterology. (1989) 96:736–49. doi: 10.1016/s0016-5085(89)80072-1
65. Van Breemen RB, and Li, Y. Caco-2 cell permeability assays to measure drug absorption. Expert Opin Drug Metab Toxicol. (2005) 1:175–85. doi: 10.1517/17425255.1.2.175
66. Carbajo-Lozoya J, Müller MA, Kallies S, Thiel V, Drosten C, and von Brunn, A. Replication of human coronaviruses SARS-CoV, HCoV-NL63 and HCoV-229E is inhibited by the drug FK506. Virus Res. (2012) 165:112–7. doi: 10.1016/j.virusres.2012.02.002
67. Carbajo-Lozoya J, Ma-Lauer Y, Malešević M, Theuerkorn M Kahlert V, Prell E, et al. Human coronavirus NL63 replication is cyclophilin A-dependent and inhibited by non-immunosuppressive cyclosporine A-derivatives including Alisporivir. Virus Res. (2014) 184:44–53. doi: 10.1016/j.virusres.2014.02.010
68. Kreft ME, Jerman UD, Lasiè, E, Hevir-Kene N, Rižner TL, Peternel L, et al. The characterization of the human cell line Calu-3 under different culture conditions and its use as an optimized in vitro model to investigate bronchial epithelial function. Eur J Pharm Sci. (2015) 69:1–9. doi: 10.1016/j.ejps.2014.12.017
69. Zhu Y, Chidekel A, Shaffer, TH. Cultured human airway epithelial cells (calu-3): a model of human respiratory function structure, and inflammatory responses. Crit Care Res Pract. (2010) 2010:394578. doi: 10.1155/2010/394578
70. Sims AC, Burkett SE, Yount B, Pickles, RJ. SARS-CoV replication and pathogenesis in an in vitro model of the human conducting airway epithelium. Virus Res. (2008) 133:33–44. doi: 10.1016/j.virusres.2007.03.013
71. Tseng CT, Tseng J, Perrone L, Worthy M, Popov V, Peters, CJ. Apical entry and release of severe acute respiratory syndrome-associated coronavirus in polarized Calu-3 lung epithelial cells. J Virol. (2005) 79:9470–9. doi: 10.1128/JVI.79.15.9470-9479.2005
72. Sheahan TP, Sims AC, Zhou S, Graham RL, Pruijssers AJ, Agostini ML, et al. An orally bioavailable broad-spectrum antiviral inhibits SARS-CoV-2 in human airway epithelial cell cultures and multiple coronaviruses in mice. Sci Transl Med. (2020) 12:eabb5883. doi: 10.1126/scitranslmed.abb5883
73. Iwata-Yoshikawa N, Okamura T, Shimizu Y, Hasegawa H, Takeda M, Nagata, N. TMPRSS2 contributes to virus spread and immunopathology in the airways of murine models after coronavirus infection. J Virol. (2019) 93:e01815-18. doi: 10.1128/JVI.01815-18
74. Roberts A, Paddock C, Vogel L, Butler E, Zaki S, Subbarao, K. Aged BALB/c mice as a model for increased severity of severe acute respiratory syndrome in elderly humans. J Virol. (2005) 79:5833–8. doi: 10.1128/JVI.79.9.5833-5838.2005
75. Gong SR, Bao, LL. The battle against SARS and MERS coronaviruses: reservoirs and animal models. Animal Model Exp Med. (2018) 1:125–33. doi: 10.1002/ame2.12017
76. McCray PB Jr., Pewe L, Wohlford-Lenane C, Hickey M, Manzel L, Shi L, et al. Lethal infection of K18-hACE2 mice infected with severe acute respiratory syndrome coronavirus. J Virol. (2007) 81:813–21. doi: 10.1128/JVI.02012-06
77. Yang XH, Deng W, Tong Z, Liu XY, Zhang LF, Zhu H, et al. Mice transgenic for human angiotensin-converting enzyme 2 provide a model for SARS coronavirus infection. Comp Med. (2007) 57:450–59.
78. Alenina N, Bader, M. ACE2 in brain physiology and pathophysiology: evidence from transgenic animal models. Neurochem Res. (2019) 44:1323–29. doi: 10.1007/s11064-018-2679-4
79. Bao L, Deng W, Huang B, Gao H, Liu J, Ren L, et al. The pathogenicity of SARS-CoV-2 in hACE2 transgenic mice. Nature. (2020) 583:830–3. doi: 10.1038/s41586-020-2312-y
80. Israelow B, Song E, Mao T, Lu P, Meir A, Liu F, et al. Mouse model of SARS-CoV-2 reveals inflammatory role of type I interferon signaling. bioRxiv. (2020). [Preprint]. doi: 10.1101/2020.05.27.118893
81. Sun SH, Chen Q, Gu HJ, Yang G, Wang YX, Huang XY, et al. A Mouse Model of SARS-CoV-2infection and pathogenesis. Cell Host Microbe. (2020) 28:1–10. doi: 10.23880/vij-16000242
82. Kim TS, Heinlein C, Hackman RC, Nelson, PS. Phenotypic analysis of mice lacking the Tmprss2-encoded protease. Mol Cell Biol. (2006) 26:965–75. doi: 10.1128/MCB.26.3.965-975.2006
83. Sugawara I, Yamada H, Mizuno, S. STAT1 knockout mice are highly susceptible to pulmonary mycobacterial infection. Tohoku J Exp Med. (2004) 202:41–50. doi: 10.1620/tjem.202.41
84. Meraz MA, White JM, Sheehan KC, Bach A, Rodig SJ, Dighe AS, et al. Targeted disruption of the Stat1 gene in mice reveals unexpected physiologic specificity in the JAK-STAT signaling pathway. Cell. (1996) 84:431–42. doi: 10.1016/s0092-867481288-x
85. Mahlakõiv T, Ritz D, Mordstein M, DeDiego ML, Enjuanes L, Müller MA, et al. Combined action of type I and type III interferon restricts initial replication of severe acute respiratory syndrome coronavirus in the lung but fails to inhibit systemic virus spread. J Gen Virol. (2012) 93(Pt 12):2601–5. doi: 10.1099/vir.0.046284-0
86. Frieman MB, Chen J, Morrison TE, Whitmore A, Funkhouser W, Ward JM, et al. SARS-CoV pathogenesis is regulated by a STAT1 dependent but a type I, II and III interferon receptor independent mechanism. PLoS Pathog. (2010) 6:e1000849. doi: 10.1371/journal.ppat.1000849
87. Gu H, Chen Q, Yang G, He L, Fan H, Deng YQ, et al. Rapid adaptation of SARS-CoV-2 in BALB/c mice: Novel mouse model for vaccine efficacy. bioRxiv. (2020). [Preprint]. doi: 10.1101/2020.05.02.073411
88. Dinnon IIIKH, Leist SR, Schäfer A, Edwards CE, Martinez DR, Montgomery SA, et al. A mouse-adapted SARS-CoV-2 model for the evaluation of COVID-19 medical countermeasures. bioRxiv. (2020). [Preprint]. doi: 10.1101/2020.05.06.081497
89. Gao Q, Bao L, Mao H, Wang L, Xu K, Yang M, et al. Rapid development of an inactivated vaccine for SARS-CoV-2. bioRxiv. (2020). [Preprint] doi: 10.1101/2020.04.17.046375
90. Roberts A, Vogel L, Guarner J, Hayes N, Murphy B, Zaki S, et al. Severe acute respiratory syndrome coronavirus infection of golden Syrian hamsters. J Virol. (2005) 79:503–11. doi: 10.1128/JVI.79.1.503-511.2005
91. Schaecher SR, Stabenow J, Oberle C, Schriewer J, Buller RM, Sagartz JE, et al. An immunosuppressed Syrian golden hamster model for SARS-CoV infection. Virology. (2008) 380:312–21. doi: 10.1016/j.virol.2008.07.026
92. Sia SF, Yan LM, Chin AWH, Fung K, Choy KT, Wong AYL, et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature. (2020) 583:834–8. doi: 10.1038/s41586-020-2342-5
93. Chan JF, Zhang AJ, Yuan S, Poon VK, Chan CS, Lee AC, et al. Simulation of the clinical and pathological manifestations of Coronavirus Disease 2019 (COVID-19) in golden Syrian hamster model: implications for disease pathogenesis and transmissibility [published online ahead of print, 2020 Mar 26]. Clin Infect Dis. (2020) 20:ciaa325. doi: 10.1093/cid/ciaa325
94. Zhang Q, Shi J, Deng G, Guo J, Zeng X, He X, et al. H7N9 influenza viruses are transmissible in ferrets by respiratory droplet. Science. (2013) 341:410–14. doi: 10.1126/science.1240532
95. van den Brand JM, Haagmans BL, Leijten L, van Riel D, Martina BEE, Osterhauset ADME, et al. Pathology of experimental SARS coronavirus infection in cats and ferrets. Vet Pathol. (2008) 45:551–62. doi: 10.1354/vp.45-4-551
96. Chu YK, Ali GD, Jia F, Li Q, Kelvin D, Couch RC, et al. The SARS-CoV ferret model in an infection-challenge study. Virology. (2008) 374:151–63. doi: 10.1016/j.virol.2007.12.032
97. Shi J, Wen Z, Zhong G, Yang H, Wang C, Huang B, et al. Susceptibility of ferrets cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science. (2020) 368:1016–20. doi: 10.1126/science.abb7015
98. Richard M, Kok A, de Meulder D, Bestebroer T, Lamers M, Okba N, et al. SARS-CoV-2 is transmitted via contact and via the air between ferrets. bioRxiv. (2020). [Preprint]. doi: 10.1101/2020.04.16.044503
99. Kim YI, Kim SG, Kim SM, Kim EH, Park SJ, Yu KM, et al. Infection and Rapid Transmission of SARS-CoV-2 in Ferrets. Cell Host Microbe. (2020) 27:704–9.e2. doi: 10.1016/j.chom.2020.03.023
100. Yu P, Qi F, Xu Y, Li F, Liu P, Liu J, et al. Age-related rhesus macaque models of COVID-19. Animal Model Exp Med. (2020) 3:93–7. doi: 10.1002/ame2.12108
101. Roberts A, Subbarao, K. Animal models for SARS. Adv Exp Med Biol. (2006) 581:463–71. doi: 10.1007/978-0-387-33012-9_83
102. Roberts A, Lamirande EW, Vogel L, Jackson JP, Paddock CD, Guarner J, et al. Animal models and vaccines for SARS-CoV infection. Virus Res. (2008) 133:20–32. doi: 10.1016/j.virusres.2007.03.025
103. Smits SL, de Lang A, van den Brand JM, Leijten JM, van IJcken WF, Eijkemans M, et al. Exacerbated innate host response to SARS-CoV in aged non-human primates. PLoS Pathog. (2010) 6:e1000756. doi: 10.1371/journal.ppat.1000756
104. Gretebeck LM, Subbarao, K. Animal models for SARS and MERS coronaviruses. Curr Opin Virol. (2015) 13:123–9. doi: 10.1016/j.coviro.2015.06.009
105. Rockx B, Kuiken T, Herfst S, Bestebroer T, Lamers MM, de Meulder D, et al. Comparative pathogenesis Of COVID-19, MERS And SARS in a non-human primate model. Science. (2020) 368:1012–5. doi: 10.1126/science.abb7314
106. Munster VJ, Feldmann F, Williamson BN, van Doremalen N, Pérez-Pérez L, Schulzet J, et al. Respiratory disease in rhesus macaques inoculated with SARS-CoV-2. Nature. (2020). doi: 10.1038/s41586-020-2324-7 [Epub ahead of print].
107. Lu S, Zhao Y, Yu W, Yang Y, Gao J, Wang J, et al. Comparison of SARS-CoV-2 infections among 3 species of non-human primates. bioRxiv. (2020). [Preprint]. doi: 10.1101/2020.04.08.031807
108. Le Bras, A. SARS-CoV-2 causes COVID-19-like disease in cynomolgus macaques. Lab Anim (N Y). (2020) 49:174. doi: 10.1038/s41684-020-0571-8
109. Deng W, Bao L, Gao H, Xiang Z, Qu Y, Song Z, et al. Ocular conjunctival inoculation of SARS-CoV-2 can cause mild COVID-19 in Rhesus macaques. bioRxiv. (2020). [Preprint]. doi: 10.1101/2020.03.13.990036
110. de Wit E, Feldmann F, Cronin J, Jordan R, Okumura A, Thomas T, et al. Prophylactic and therapeutic remdesivir (GS-5734) treatment in the rhesus macaque model of MERS-CoV infection. Proc Natl Acad Sci USA. (2020) 117:6771–6. doi: 10.1073/pnas.1922083117
111. Bao L, Deng W, Gao H, Xiao C, Liu J, Xue J, et al. Lack of Reinfection in Rhesus Macaques Infected with SARS-CoV-2. bioRxiv. (2020). [Preprint]. doi: 10.1101/2020.03.13.990226
112. Chandrashekar A, Liu J, Martinot A, Mcmahan K, Mercado N, Peter L, et al. SARS-CoV-2 infection protects against rechallenge in rhesus macaques. Science. (2020) 20:eabc4776. doi: 10.1126/science.abc4776
113. Williamson B, Feldman F, Meade-White K, Porter DR, Schulz J, et al. Clinical benefit of remdesivir in rhesus macaques infected with SARS-CoV-2. bioRxiv. (2020). [Preprint]. doi: 10.1101/2020.04.15.043166
114. Zhang J, Zeng H, Gu J, Li H, Zheng L, Zou, Q. Progress and prospects on vaccine development against SARS-CoV-2. Vaccines (Basel). (2020) 8:E153. doi: 10.3390/vaccines8020153
115. Gao Q, Bao L, Mao H, Wang L, Xu K, Yang M, et al. Development of an inactivated vaccine candidate for SARS-CoV-2. Science. (2020) 6:eabc1932. doi: 10.1126/science.abc1932
116. Ramani A, Müller L, Ostermann PN, Gabriel E, Abida-Islam P, Müller-Schiffmann A, et al. SARS-CoV-2 targets cortical neurons of 3D human brain organoids and shows neurodegeneration-like effects. bioRxiv. (2020). [Preprint]. doi: 10.1101/2020.05.20.106575
117. Song E, Zhang C, Israelow B, Weizman OE, Liu F, Dai Y, et al. Neuroinvasive potential of SARS-CoV-2 revealed in a human brain organoid model. bioRxiv. (2020). [Preprint]. doi: 10.1101/2020.06.25.169946
118. Yang L, Han Y, Nilsson-Payant BE, Gupta V, Wang H, Duan K, et al. A Human pluripotent stem cell-based platform to study SARS-CoV-2 tropism and model virus infection in human cells and organoids. Cell Stem Cell. (2020) 27:125–36.e7. doi: 10.1016/j.stem.2020.06.015
119. De Kleijn KMA, Zuure WA, Peijnenborg J, Heuvelmans JM, Martens, GJM. Reappraisal of human HOG and MO3.13 cell lines as a model to study oligodendrocyte functioning. Cells. (2019) 8:1096. doi: 10.3390/cells8091096
120. Post GR, Dawson, G. Characterization of a cell line derived from a human oligodendroglioma. Mol Chem Neuropathol. (1992) 16:303–17. doi: 10.1007/BF03159976
121. Benda P, Lightbody J, Sato G, Levine L, Sweet, W. Differentiated rat glial cell strain in tissue culture. Science. (1839) 16:1968.
122. Gao J, Hu L, Peng HH, Shi, JG. The complete mitochondrial genome sequence of the rat C6 glioma cell line. Mitochondrial DNA Part A, DNA mapping. Seq Anal. (2016) 27:2188–9. doi: 10.3109/19401736.2014.982619
123. Yamashita M, Yamate M, Li GM, Ikuta, K. Susceptibility of human and rat neural cell lines to infection by SARS-coronavirus. Biochem Biophys Res Commun. (2005) 334:79–85. doi: 10.1016/j.bbrc.2005.06.061
124. Arbour N, Talbot, PJ. Persistent infection of neural cell lines by human coronaviruses. Adv Exp Med Biol. (1998) 440:575–81. doi: 10.1007/978-1-4615-5331-1_75
125. Arbour N, Côté, G Lachance, C Tardieu, M Cashman, NR, Talbot, PJ. Acute and persistent infection of human neural cell lines by human coronavirus OC43. J Virol. (1999) 73:3338–50. doi: 10.1128/jvi.73.4.3338-3350.1999
126. Favreau DJ, Meessen-Pinard M, Desforges M, Talbot, PJ. Human coronavirus-induced neuronal programmed cell death is cyclophilin D dependent and potentially caspase dispensable. J Virol. (2012) 86:81–93. doi: 10.1128/jvi.06062-11
127. Lachance C, Arbour N, Cashman NR, Talbot, PJ. Involvement of aminopeptidase N (CD13) in Infection of human neural cells by human coronavirus 229E. J Virol. (1998) 72:6511–9. doi: 10.1128/jvi.72.8.6511-6519.1998
128. Meessen-Pinard M, Le Coupanec A, Desforges M, Talbot, PJ. Pivotal role of receptor-interacting protein kinase 1 and mixed lineage kinase domain-like in neuronal cell death induced by the human neuroinvasive coronavirus OC43. J Virol. (2017) 91:1–20. doi: 10.1128/jvi.01513-16
129. St-Jean JR, Desforges M, Talbot, PJ. Genetic evolution of human coronavirus OC43 in neural cell culture. Adv Exp Med Biol. (2006) 581:499–502. doi: 10.1007/978-0-387-33012-9_88
130. Bonavia A, Arbour N, Yong VW, Talbot, PJ. Infection of primary cultures of human neural cells by human coronaviruses 229E and OC43. J Virol. (1997) 71:800–6. doi: 10.1128/jvi.71.1.800-806.1997
131. Talbot PJ, Ekande S, Cashman NR, Mounir S, Stewart, JN. Neurotropism of human coronavirus 229E. Adv Exp Med Biol. (1994) 342:339–46. doi: 10.1007/978-1-4615-2996-5_52
132. Dutta D, Heo I, Clevers, H. Disease modeling in stem cell-derived 3D organoid systems. Trends Mol Med. (2017) 23:393–410. doi: 10.1016/j.molmed.2017.02.007
133. Rossi G, Manfrin A, Lutolf, MP. Progress and potential in organoid research. Nat Rev Genet. (2018) 19:671–87. doi: 10.1038/s41576-018-0051-9
134. Busquet F, Hartung T, Pallocca G, Rovida C, Leist, M. Harnessing the power of novel animal-free test methods for the development of COVID-19 drugs and vaccines. Arch Toxicol. (2020) 94:2263–72. doi: 10.1007/s00204-020-02787-2
135. Takayama, K. In vitro and animal models for SARS-CoV-2 research. Trends Pharmacol Sci. (2020) 41:513–7. doi: 10.1016/j.tips.2020.05.005
136. Han Y, Yang L, Duan X, Duan F, Payant N, Tomer MY, et al. Identification of candidate COVID-19 therapeutics using hPSC-derived lung organoids. bioRxiv. (2020). [Preprint]. doi: 10.1101/2020.05.05.079095
137. Zhao B, Ni C, Gao R, Wang G, Li Y, Jinsong W, et al. Recapitulation of SARS-CoV-2 infection and cholangiocyte damage with human liver ductal organoids. Protein Cell. (2020) 2:6. doi: 10.1007/s13238-020-00718-6
138. Monteil V, Kwon H, Prado P, Hagelkrüys A, Wimmer RA, Stahlet M, et al. Inhibition of SARS-CoV-2 infections in engineered human tissues using clinical-grade soluble human ACE2. Cell. (2020) 181:905–13.e7. doi: 10.1016/j.cell.2020.04.004
139. Bullen CK, Hogberg HT, Bahadirli-Talbott A, Bishai WR, Hartung T, Keuthanet C, et al. Infectability of human BrainSphere neurons suggests neurotropism of SARS-CoV-2. ALTEX. (2020) 20:111. doi: 10.14573/altex.2006111
140. Mansour AAF, Schafer ST, Gage, FH. Cellular complexity in brain organoids: Current progress and unsolved issues. Semin Cell Dev Biol. (2020) 20:13. doi: 10.1016/j.semcdb.2020.05.013
141. Mesci P, Macia A, Saleh A, Martin-Sancho L, Yin Y, Snethlage C, et al. Sofosbuvir protects human brain organoids against SARS-CoV-2. bioRxiv. (2020). [Preprint]. doi: 10.1101/2020.05.30.125856
142. Kim H, Park HJ, Choi H, Chang Y, Park H, Shin J, et al. Modeling G2019S-LRRK2 sporadic Parkinson’s disease in 3D midbrain organoids. Stem Cell Rep. (2019) 12:518–31. doi: 10.1016/j.stemcr.2019.01.020
143. Lin YT, Seo J, Gao F, Feldman HM, Wen HL, Penney J, et al. APOE4 causes widespread molecular and cellular alterations associated with Alzheimer’s disease phenotypes in human iPSC-derived brain cell types [published correction appears in Neuron. 2018 Jun 27;98 (6):1294]. Neuron. (2018) 98:1141–54. doi: 10.1016/j.neuron.2018.05.008
144. Smits LM, Reinhardt L, Reinhardt P, Glatza M, Monzel AS, Stanslowskyet N, et al. Modeling Parkinson’s disease in midbrain-like organoids. NPJ Parkinsons Dis. (2019) 5:5. doi: 10.1038/s41531-019-0078-4
145. Bhatia HK, Singh H, Grewal N, Natt, NK. Sofosbuvir: a novel treatment option for chronic hepatitis C infection. J Pharmacol Pharmacother. (2014) 5:278–84. doi: 10.4103/0976-500X.142464
146. Walsh KB, Edwards RA, Romero KM, Kotlajich MV, Stohlman SA, Lane, TE. Expression of CXC chemokine ligand 10 from the mouse hepatitis virus genome results in protection from viral-induced neurological and liver disease. J Immunol. (2007) 179:1155–65. doi: 10.4049/jimmunol.179.2.1155
147. Savarin C, Bergmann, CC. Fine tuning the cytokine storm by IFN and IL-10 following neurotropic coronavirus encephalomyelitis. Front Immunol. (2018) 9:3022. doi: 10.3389/fimmu.2018.03022
148. Wege H, Schluesener H, Meyermann R, Barac-Latas V, Suchanek G, Lassmann, H. Coronavirus infection and demyelination. Development of inflammatory lesions in Lewis rats. Adv Exp Med Biol. (1998) 440:437–44. doi: 10.1007/978-1-4615-5331-1_55
149. Zalinger ZB, Elliott R, Weiss, SR. Role of the inflammasome-related cytokines Il-1 and Il-18 during infection with murine coronavirus. J Neurovirol. (2017) 23:845–54. doi: 10.1007/s13365-017-0574-4
150. Gruslin E, Moisan S, St-Pierre Y, Desforges M, Talbot, PJ. Transcriptome profile within the mouse central nervous system and activation of myelin-reactive T cells following murine coronavirus infection. J Neuroimmunol. (2005) 162:60–70. doi: 10.1016/j.jneuroim.2005.01.007
151. Choi KS, Aizaki H, Lai, MM. Murine coronavirus requires lipid rafts for virus entry and cell-cell fusion but not for virus release. J Virol. (2005) 79:9862–71. doi: 10.1128/JVI.79.15.9862-9871.2005
152. Singh M, Khan RS, Dine K, Das Sarma J, Shindler, KS. Intracranial inoculation is more potent than intranasal inoculation for inducing optic neuritis in the mouse hepatitis virus-induced model of multiple sclerosis. Front Cell Infect Microbiol. (2018) 8:311. doi: 10.3389/fcimb.2018.00311
153. Wang FI, Stohlman SA, Fleming, JO. Demyelination induced by murine hepatitis virus JHM strain (MHV-4) is immunologically mediated. J Neuroimmunol. (1990) 30:31–41. doi: 10.1016/0165-5728(90)90050-w
154. Dandekar AA, Perlman, S. Virus-induced demyelination in nude mice is mediated by gamma delta T cells. Am J Pathol. (2002) 161:1255–63. doi: 10.1016/s0002-9440(10)64402-1
155. Glass WG, Chen BP, Liu MT, Lane, TE. Mouse hepatitis virus infection of the central nervous system: chemokine-mediated regulation of host defense and disease. Viral Immunol. (2002) 15:261–72. doi: 10.1089/08828240260066215
Keywords: SARS-CoV-2, experimental models, COVID-19, central nervous system, neurodegenerative disease, multiple sclerosis
Citation: Sanclemente-Alaman I, Moreno-Jiménez L, Benito-Martín MS, Canales-Aguirre A, Matías-Guiu JA, Matías-Guiu J and Gómez-Pinedo U (2020) Experimental Models for the Study of Central Nervous System Infection by SARS-CoV-2. Front. Immunol. 11:2163. doi: 10.3389/fimmu.2020.02163
Received: 19 June 2020; Accepted: 10 August 2020;
Published: 28 August 2020.
Edited by:
Bruno Stankoff, Sorbonne Université, FranceReviewed by:
Yoshiro Ohara, Kanazawa Medical University, JapanCopyright © 2020 Sanclemente-Alaman, Moreno-Jiménez, Benito-Martín, Canales-Aguirre, Matías-Guiu, Matías-Guiu and Gómez-Pinedo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ulises Gómez-Pinedo, dS5nb21lei5waW5lZG9AZ21haWwuY29t
†ORCID: Inmaculada Sanclemente-Alaman, orcid.org/0000-0003-1847-547X; Lidia Moreno-Jiménez, orcid.org/0000-0002-2730-4130; María Soledad Benito-Martín orcid.org/0000-0002-6131-6426; Alejandro Canales-Aguirre, orcid.org/0000-0003-0918-788X; Jordi A. Matías-Guiu, orcid.org/0000-0001-5520-2708; Jorge Matías-Guiu, orcid.org/0000-0001-8162-9043; Ulises Gómez-Pinedo, orcid.org/0000-0003-3097-5557
‡Both authors are considered lead authors with the same responsibility
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.