 Jean-Paul Soulillou1*
Jean-Paul Soulillou1* Emanuele Cozzi
Emanuele Cozzi Jean-Marie Bach
Jean-Marie Bach
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
OPINION article
Front. Immunol. , 09 June 2020
Sec. Nutritional Immunology
Volume 11 - 2020 | https://doi.org/10.3389/fimmu.2020.00834
This article is part of the Research Topic Human Antibodies Against The Dietary Non-Human Neu5Gc-carrying Glycans in Normal and Pathologic States View all 12 articles
The thematic issue of Frontiers in Immunology, entitled “Human Antibodies Against the Dietary Non-Human Neu5Gc-Carrying Glycans in Normal and Pathological States,” benefits from an extensive fundamental review of previous studies on N-glycolylneuraminic acid (Neu5Gc) in humans [(1) for recent review and citations therein] and from studies on anti-Neu5Gc antibodies (A-GcAbs) in Cmah−/− mice that, similar to humans, lack a functional cytidine monophosphate N-acetylneuraminic acid hydroxylase (CMAH), thereby allowing Neu5Gc synthesis (2). Further, “xenosialitis” assumes that the local interaction of A-GcAbs with traces of Neu5Gc residue at the surface of certain human cells may result in a chronic activation of Neu5Gc-displaying cells, which would eventually generate atheroma [for endothelial cells (ECs)] and malignancies (for epithelia) (1, 3). However, not many studies have been performed to date in humans, and these studies have not yet unambiguously demonstrated that A-GcAb-related xenosialitis could contribute to major diseases in humans.
This opinion article aims to assess the available evidence-based background of the concept of a deleterious effect of xenosialitis on human diseases, and to suggest alternative working hypotheses for future studies. As previously mentioned, xenosialitis, which is restricted to human pathologies in this opinion article, has been proposed as a “logical” basis for deleterious effects that would result from in situ interactions of A-GcAbs, which are present in human sera and diet-derived Neu5Gc deposits on certain human cells (1). However, this framework assumes the paradox that evolution would have allowed such a dangerous confrontation. Results from studies on Cmah−/− mice have been extensively reviewed (1). However, using CMAH-deficient mice—which do not benefit from the coevolution adaptation that followed the lack of Neu5Gc in humans— poses an issue, as xenosialitis models require exogenous immunizations to elicit A-GcAbs (4, 5), due to the difficulty of “humanizing” the mice with a Neu5Gc-rich diet. In addition, cases of “positive data” arguing for xenosialitis in animal models have required experimental designs that use mice which are prone to spontaneously developing endothelial injuries (5) [see also comments in (6, 7)].
The main differences regarding xenosialitis between humans and CMAH-deficient mouse models in terms of prevailing conditions are as follows: (1) the actual levels of loading among human cells with diet-derived Neu5Gc, and (2) the differences between A-GcAbs that result either from immunization by diet or from active immunization with animal-derived products and adjuvants.
The presence of Neu5Gc traces on ECs or epithelial cells from various organs in humans has been established using ten autopsy samples (8). Since unambiguously observing such deposits using anti-Neu5Gc chicken polyclonal Ab staining on frozen or fixed-histological tissue samples was difficult, we used flow cytometry to assess the binding of anti-Neu5Gc chicken Abs on living ECs from large arteries of brain-dead donors (9). Although we confirmed a faint signal on gated ECs in four samples, three other preparations were found to be negative (9). A roughly similar proportion was found in eight additional living EC preparations that were tested after sorting, of which, two were positive, two were negative, and four had extremely faint or negative staining (unpublished data). This is distinguished from the high-Neu5Gc loading among certain malignant cells (10), which may enable therapy using exogenously produced A-GcAbs and to monitor A-GcAb levels as a disease marker (11, 12). The low metabolic incorporation of Neu5Gc in humans with a Neu5Gc-rich diet can be explained by intestinal colonization of sialidases producing bacteria (13, 14).
Few studies have investigated Neu5Gc loading among human cells (8, 9); however, further studies are warranted to establish whether indeed a substantial fraction of humans actually lack the fundamental component of the theoretical basis of the xenosialitis model. Although Neu5Gc exists at trace levels on ECs of positive individuals, we must also consider the myriad of surface glycoproteins or lipoproteins that display Neu5Gc and the high diversity of Neu5Gc terminal residues, which result in a huge dispersion of cell-surface epitopes that are potentially recognized by A-GcAbs (15). It is thus possible that, following interactions with A-GcAbs, the coexistence of trace levels of antigens and the high epitope dispersion results in physiological cellular signaling that is below the activation threshold. In a recent analysis of affinity-purified natural A-GcAbs detected by ELISA and Glycan arrays, the authors suggested that “specific A-GcAbs” may only represent a small minority of the pool detected in the assays (16). These data confirm our working hypothesis. In addition, whatever the molecular definition of the glycans recognized by the natural, diet induced, anti-Neu5Gc measured by ELISA or Arrays, the question of their biological effects in humans remains.
Indeed, only a few in vitro studies explored the effects of A-GcAbs on human cells. The first (17) suggests there is an activation of umbilical ECs that develop a white blood cell binding phenotype after incubation with A-GcAbs-containing whole serum. However, these first experiments used several extra-physiological conditions; for instance, the Neu5Gc loading among ECs far exceeding the levels naturally observed in human ECs and the high anti-Neu5Gc titer of the serum tested. A second study (9) used affinity-purified A-GcAbs from either normal sera (diet-induced Abs) or sera of those highly immunized by rabbit polyclonal IgGs (elicited Abs) (18). In addition, large artery ECs that undergo physiological loading levels of Neu5Gc were used (9).
Although this last study (9) was restricted to the complete transcriptomic patterns and apoptosis of stimulated ECs, it is interesting that the activation patterns triggered either by purified diet-derived human A-GcAbs or by rabbit IgG-elicited A-GcAbs in these more physiological conditions did not present a classical “inflammation-like” activation of ECs. In contrast, the observed patterns are consistent with the concept that A-GcAbs may contribute to the homeostasis of ECs (9). Moreover, purified A-GcAbs were shown to downregulate classical inflammation patterns that are induced by the presence of normal sera, added as a complement source (with components also necessary to cell homeostasis) (9). Further, purified A-GcAbs inhibited important master genes involved in EC activation (9). In conclusion, the theoretical basis of xenosialitis in humans, which involves A-GcAbs, requires an improved assessment of the actual levels of Neu5Gc loading among human cells in vivo and of the percentage of normal individuals who exhibit detectable Neu5Gc on ECs or epithelia. In addition, the effects of purified A-GcAbs on ECs or epithelial cells should be tested in vitro under experimental conditions that more closely mimic “physiological” Neu5Gc loading.
Humans develop A-GcAbs within the first few months of life after being introduced to a Neu5Gc-containing diet (19). The impact of food antigens on immunity is poorly understood; further, the apoptosis of diet-activated T cells is a hallmark of the healthy intestine (20). Whether diet/microbiota levels significantly affect A-GcAb levels in healthy adults has not yet been determined (21). In contrast, after implantation of animal biodevices (22, 23) or infusion of animal-derived molecules, such as rabbit IgGs, blood-elicited A-GcAb levels drastically increase for several months (18) and largely exceed the average normal levels in non-immunosuppressed individuals. As expected, these “exogenously” elicited Abs display a high affinity and altered repertoire (24). In contrast to diet-derived natural immunization, the elicited responses result in a vigorous, memory-type induction of A-GcAbs in young adults (18) with a significant number of individuals exhibiting extremely high titres (from 20 μg/ml up to 1 g/l). The extent to which proportion-elicited A-GcAbs stemmed from B cells that were primed by diet-derived Neu5Gcs is currently unknown. Importantly, exposure to such high titres of A-GcAbs affects drug half-life and is associated with the serum sickness disease (SSD), likely due to the A-GcAbs (25). However, SSD is linked to immune complexes that circulate (26), rather than in situ xenosialitis. Whether the increase in late renal failure in those who develop SSD (25) results from early graft injury due to immune complexes, or xenosialitis that results in long-term exposure to elicited A-GcAbs, remains unknown. The late loss of transplant function that is associated with the highest elicited A-GcAb titres in patients who received rabbit IgGs [in Supplementary Data of (25)] is yet anecdotal, generated by a small group of patients in the absence of graft histological samples. There are thus two different contexts that must be considered. Deleterious-elicited A-GcAbs (as tested experimentally in CMAH-deficient mice) do not imply that diet-induced “natural” A-GcAbs are necessarily detrimental. Since extremely high titres of elicited A-GcAbs in non-immunosuppressed patients were not associated with even a clinically detectable acute vascular insult (27), coevolution adaptation to diet-induced A-GcAbs may also operate to control elicited A-GcAb effects. Similarly, diet-induced A-GcAbs within the first year of life are not associated with vascular pathology. Thus, along with the “threshold effect” hypothesis, the presence of protective mechanisms, which are likely shaped by evolution to escape the deleterious effects of A-GcAbs, is another working hypothesis to consider.
Elevated A-GcAb levels have been determined to be inversely correlated with non-atopic asthma in farmers' children (32). Likewise, elevated A-GcAb titres have also been observed in patients with normal coronaries in Kawasaki disease (29). Both observations do not suggest a link between A-GcAb titres and inflammation.
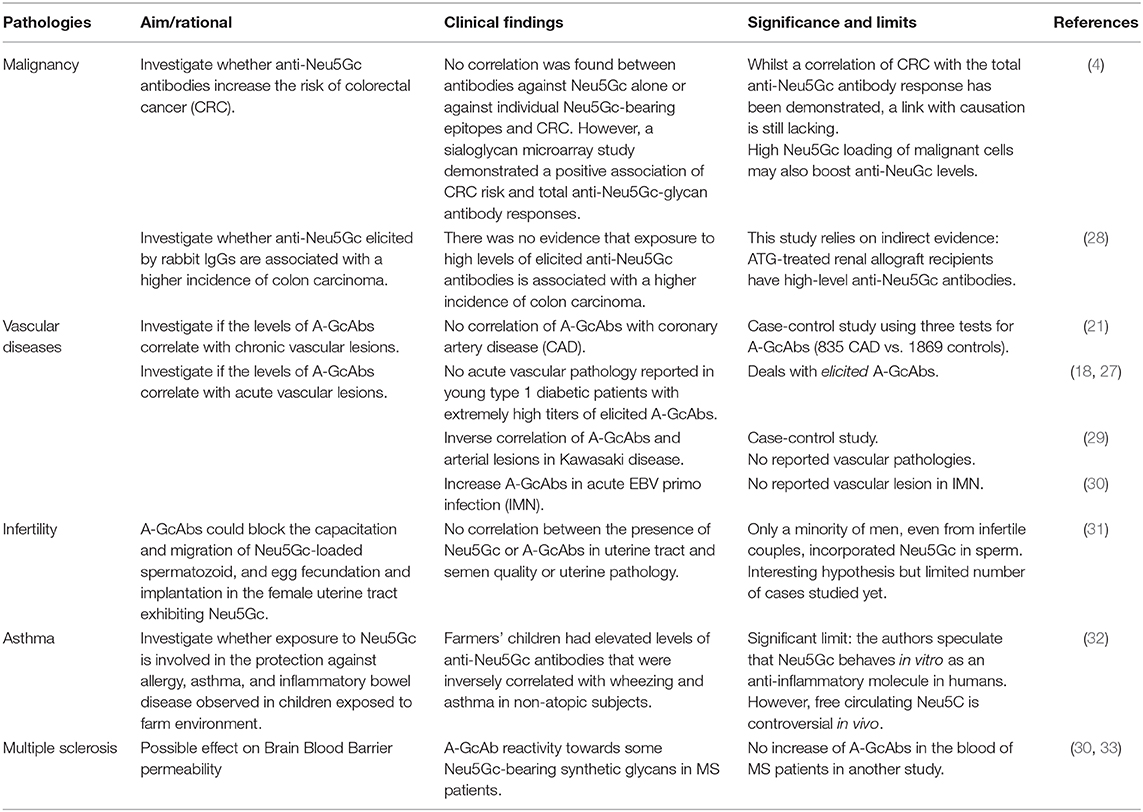
Table 1. Clinical correlations between died-induced A-GcAbs and pathologies.
Xenosialitis has been proposed as a contributor to colon cancer, due to also being associated with high red meat intake (21). However, no association between A-GcAb levels, red meat intake, or coronary artery disease (CAD) risk, has been evidenced in adults, as assessed by several types of ELISA (21). In comparison, colorectal cancer (CRC) was found to be significantly associated with total A-GcAb responses using a Sialoglycan Microarray that measures the Ab repertoire against Neu5Gc-glycans (21). Nevertheless, the increased A-GcAb titres observed may merely represent an immunogenic marker of the strong Neu5Gc loading of malignant cells (10, 12, 34). Another study compared the incidence of colon cancer between kidney recipients (including 212,465 patients and 522 with colon cancer) who either received or did not receive rabbit anti-T cell IgGs that are able to induce long-term elevated A-GcAbs (28). While relying indirectly on inducing elicited A-GcAbs by rabbit IgGs in immunosuppressed patients (25), a long-term survey showed no difference in colon cancer incidence (28).
In further studies, the deleterious effects of A-GcAbs from females (and potentially males) have been reported on spermatozoids and egg implantation in the female uterine tract that contain Neu5Gc from diet, which thus may imply that xenosialitis is involved in certain cases of infertility (31). However, neither the presence of Neu5Gc nor that of A-GcAbs have been correlated with differences in semen quality or the presence of uterine pathology (31).
There are also several instances in the clinical arena in which a low antigen density on ECs does not result in identifiable deleterious effects, such as when blood group A2 organs are transplanted in ABO-incompatible recipients (35). Moreover, normal individual sera display a high diversity of Abs that cross-react with self-determinants (36, 37), which may provide anti-apoptotic signals and shape the immune repertoire by enabling more efficient cognitive responses. Significantly increased levels of A-GcAbs have also been reported in EBV acute infectious mononucleosis (IMN), which is likely related to the concomitant high percentage of EBV-infected B cells (30). As IMN is associated with high incidence of multiple sclerosis (MS) (38), it has been hypothesized that A-GcAb levels could enhance the migration of anti-EBV T cells through the blood-brain barrier (39). However, as previously mentioned, transcriptomic studies in vitro do not reveal patterns that are classically associated with EC inflammation (9). Using a semi-quantitative synthetic glycan array, a recent study reported a specific pattern of IgG reactivity for some Neu5Gc epitopes in MS patients compared to other neurologic diseases (33).
Thus, whether the concomitant traces of Neu5Gc on ECs and of diet-induced circulating A-GcAbs theoretically trigger inflammation at the site of the antigens, either by direct or complement-mediated effects, or by bridging CD16 positive blood mononucleated cells onto ECs, remains to be explored. Due to the absence of convincing and statistically-powered clinical evidence of xenosialitis, we recommend critically revisiting associated concepts and exploring the possibility that diet-derived A-GcAbs may contribute to EC homeostasis.
As Galileo said, experiments are “questions asked to nature,” and so scientists routinely encounter subjectivity in their designs. We are aware that this limitation also exists when elaborating on the putative role of diet-induced “natural” A-GcAbs in the clinical arena, especially following Descartes' “de omnibus dubitandum” seminal warning. We suggest that a revisiting of the role of A-GcAbs in human biology with new tools and innovative working hypotheses will benefit scientific understanding and clinical application.
All authors thoroughly discussed all assertions of the correspondence and wrote this opinion paper.
J-PS and J-MB are cofounders of the Xenothera start-up.
The remaining author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
A-GcAbs, Anti-Neu5Gc antibodies; CAD, Coronary artery disease; CMAH, Cytidine monophosphate N-acetylneuraminic acid hydroxylase; CRC, Colorectal cancer; EBV, Epstein-Barr virus; IMN, Infectious mononucleosis; ECs, Endothelial cells; Neu5Gc, N-glycolylneuraminic acid; MS, Multiple sclerosis; SSD, Serum sickness disease.
1. Dhar C, Sasmal A, Varki A. From “Serum Sickness” to “Xenosialitis”: past, present, and future significance of the non-human sialic acid Neu5Gc. Front Immunol. (2019) 10:807. doi: 10.3389/fimmu.2019.00807
2. Schauer R, Schoop HJ, Faillard H. [On biosynthesis of the glycolyl groups of N-glycolylneuraminic acid. Oxidative conversion of N-acetyl groups to glycolyl groups]. Hoppe Seylers Z Physiol Chem. (1968) 349:645–52. doi: 10.1515/bchm2.1968.349.1.645
3. Pearce OM, Läubli H. Sialic acids in cancer biology and immunity. Glycobiology. (2016) 26:111–28. doi: 10.1093/glycob/cwv097
4. Samraj AN, Pearce OM, Läubli H, Crittenden AN, Bergfeld AK, Banda K, et al. A red meat-derived glycan promotes inflammation and cancer progression. Proc Natl Acad Sci USA. (2015) 112:542–7. doi: 10.1073/pnas
5. Kawanishi K, Dhar C, Do R, Varki N, Gordts PLSM, Varki A. Human species-specific loss of CMP- N -acetylneuraminic acid hydroxylase enhances atherosclerosis via intrinsic and extrinsic mechanisms. Proc Natl Acad Sci USA. (2019) 116:16036–45. doi: 10.1073/pnas.1902902116
6. Soulillou JP, Cozzi E, Galli C, Bach JM. Can we extrapolate from a Cmah (-/-) Ldlr (-/-) mouse model a susceptibility for atherosclerosis in humans? Proc Natl Acad Sci USA. (2020) 117:1845–6. doi: 10.1073/pnas.1915658117
7. Kawanishi K, Dhar C, Varki A, Gordts PLSM. Reply to soulillou et al.: difficulties in extrapolating from animal models exemplify unusual human atherosclerosis susceptibility and mechanisms via CMAH loss. Proc Natl Acad Sci USA. (2020). 117:1847–8. doi: 10.1073/pnas.1917278117
8. Tangvoranuntakul P, Gagneux P, Diaz S, Bardor M, Varki N, Varki A, et al. Human uptake and incorporation of an immunogenic nonhuman dietary sialic acid. Proc Natl Acad Sci USA. (2003) 100:12045–50. doi: 10.1073/pnas.2131556100
9. Le Berre L, Danger R, Mai HL, Amon R, Leviatan Ben-Arye S, Bruneau S, et al. Elicited and pre-existing anti-Neu5Gc antibodies differentially affect human endothelial cells transcriptome. Xenotransplantation. (2019) 26:e12535. doi: 10.1111/xen.12535
10. Inoue S, Sato C, Kitajima K. Extensive enrichment of N-glycolylneuraminic acid in extracellular sialoglycoproteins abundantly synthesized and secreted by human cancer cells. Glycobiology. (2010) 20:752–62. doi: 10.1093/glycob/cwq030
11. Labrada M, Dorvignit D, Hevia G, Rodríguez-Zhurbenko N, Hernández AM, Vázquez AM, et al. GM3(Neu5Gc) ganglioside: an evolution fixed neoantigen for cancer immunotherapy. Semin Oncol. (2018) 45:41–51. doi: 10.1053/j.seminoncol.2018.04.003
12. Padler-Karavani V, Hurtado-Ziola N, Pu M, Yu H, Huang S, Muthana S, et al. Human xeno-autoantibodies against a non-human sialic acid serve as novel serum biomarkers and immunotherapeutics in cancer. Cancer Res. (2011) 71:3352–63. doi: 10.1158/0008-5472.CAN-10-4102
13. Banda K, Gregg CJ, Chow R, Varki NM, Varki A. Metabolism of vertebrate amino sugars with N-glycolyl groups: mechanisms underlying gastrointestinal incorporation of the non-human sialic acid xeno-autoantigen N-glycolylneuraminic acid. J Biol Chem. (2012) 287:28852–64. doi: 10.1074/jbc.M112.364182
14. Zaramela LS, Martino C, Alisson-Silva F, Rees SD, Diaz SL, Chuzel L, et al. Gut bacteria responding to dietary change encode sialidases that exhibit preference for red meat-associated carbohydrates. Nat Microbiol. (2019) 4:2082–9. doi: 10.1038/s41564-019-0564-9
15. Kooner AS, Yu H, Chen X. Synthesis of N-glycolylneuraminic acid (Neu5Gc) and Its glycosides. Front Immunol. (2019) 10:2004. doi: 10.3389/fimmu.2019.02004
16. Obukhova P, Tsygankova S, Chinarev A, Shilova N, Nokel A, Kosma P, et al. Are there specific antibodies against Neu5Gc epitopes in the blood of healthy individuals? Glycobiology. (2020). doi: 10.1093/glycob/cwz107
17. Pham T, Gregg CJ, Karp F, Chow R, Padler-Karavani V, Cao H, et al. Evidence for a novel human-specific xeno-auto-antibody response against vascular endothelium. Blood. (2009) 114:5225–35. doi: 10.1182/blood-2009-05-220400
18. Salama A, Evanno G, Lim N, Rousse J, Le Berre L, Nicot A, et al. Anti-Gal and anti-Neu5Gc responses in nonimmunosuppressed patients after treatment with rabbit antithymocyte polyclonal IgGs. Transplantation. (2017) 101:2501–7. doi: 10.1097/TP.0000000000001686
19. Taylor RE, Gregg CJ, Padler-Karavani V, Ghaderi D, Yu H, Huang S, et al. Novel mechanism for the generation of human xeno-autoantibodies against the nonhuman sialic acid N-glycolylneuraminic acid. J Exp Med. (2010) 207:1637–46. doi: 10.1084/jem.20100575
20. Visekruna A, Hartmann S, Sillke YR, Glauben R, Fischer F, Raifer H, et al. Intestinal development and homeostasis require activation and apoptosis of diet-reactive T cells. J Clin Invest. (2019) 129:1972–83. doi: 10.1172/JCI98929
21. Samraj AN, Bertrand KA, Luben R, Khedri Z, Yu H, Nguyen D, et al. Polyclonal human antibodies against glycans bearing red meat-derived non-human sialic acid N-glycolylneuraminic acid are stable, reproducible, complex and vary between individuals: total antibody levels are associated with colorectal cancer risk. PLoS ONE. (2018) 13:e0197464. doi: 10.1371/journal.pone.0197464
22. Scobie L, Padler-Karavani V, Le Bas-Bernardet S, Crossan C, Blaha J, Matouskova M, et al. Long-term IgG response to porcine Neu5Gc antigens without transmission of PERV in burn patients treated with porcine skin xenografts. J Immunol. (2013) 191:2907–15. doi: 10.4049/jimmunol.1301195
23. Salama A, Evanno G, Harb J, Soulillou JP. Potential deleterious role of anti-Neu5Gc antibodies in xenotransplantation. Xenotransplantation. (2015) 22:85–94. doi: 10.1111/xen.12142
24. Amon R, Ben-Arye SL, Engler L, Yu H, Lim N, Berre LL, et al. Glycan microarray reveal induced IgGs repertoire shift against a dietary carbohydrate in response to rabbit anti-human thymocyte therapy. Oncotarget. (2017) 8:112236–44. doi: 10.18632/oncotarget.23096
25. Couvrat-Desvergnes G, Salama A, Le Berre L, Evanno G, Viklicky O, Hruba P, et al. Rabbit antithymocyte globulin-induced serum sickness disease and human kidney graft survival. J Clin Invest. (2015) 125:4655–65. doi: 10.1172/JCI82267
26. Dixon FJ, Mauer PH. Effects of large infusions of heterologous serum proteins on the serum protein metabolism of rabbits. J Exp Med. (1955) 101:233–44. doi: 10.1084/jem.101.3.233
27. Gitelman SE, Gottlieb PA, Rigby MR, Felner EI, Willi SM, Fisher LK, et al. Antithymocyte globulin treatment for patients with recent-onset type 1 diabetes: 12-month results of a randomised, placebo-controlled, phase 2 trial. Lancet Diabetes Endocrinol. (2013) 1:306–16. doi: 10.1016/S2213-8587(13)70065-2
28. Soulillou JP, Süsal C, Döhler B, Opelz G. No increase in colon cancer risk following induction with Neu5Gc-bearing rabbit Anti-T Cell IgG (ATG) in recipients of kidney transplants. Cancers. (2018) 10:324. doi: 10.3390/cancers10090324
29. Padler-Karavani V, Tremoulet AH, Yu H, Chen X, Burns JC, Varki A. A simple method for assessment of human anti-Neu5Gc antibodies applied to Kawasaki disease. PLoS ONE. (2013) 8:e58443. doi: 10.1371/journal.pone.0058443
30. Le Berre L, Rousse J, Gourraud PA, Imbert-Marcille BM, Salama A, Evanno G, et al. Decrease of blood anti-α1,3 galactose abs levels in multiple sclerosis (MS) and clinically isolated syndrome (CIS) patients. Clin Immunol. (2017) 180:128–35. doi: 10.1016/j.clim.2017.05.006
31. Sroga JM, Wu DH, Ma F, Tecle E, Sequoyah Reynoso H, Ressler IB, et al. Detection of the dietary xenoglycan N-glycolylneuraminic acid (Neu5Gc) and anti-Neu5Gc antibodies within reproductive tracts of male and female infertility subjects. Clin Obstet Gynecol Reprod Med. (2015) 1:72–8. doi: 10.15761/COGRM.1000120
32. Frei R, Roduit C, Ferstl R, O'Mahony L, Lauener RP. Exposure of children to rural lifestyle factors associated with protection against allergies induces an anti-Neu5Gc antibody response. Front Immunol. (2019) 10:1628. doi: 10.3389/fimmu.2019.01628
33. Boligan KF, Oechtering J, Keller CW, Peschke B, Rieben R, Bovin N, et al. Xenogeneic Neu5Gc and self-glycan Neu5Ac epitopes are potential immune targets in MS. Neurol Neuroimmunol Neuroinflamm. (2020) 7:e676. doi: 10.1212/NXI.0000000000000676.
34. Bousquet PA, Sandvik JA, Jeppesen Edin NF, Krengel U. Hypothesis: hypoxia induces de novo synthesis of NeuGc gangliosides in humans through CMAH domain substitute. Biochem Biophys Res Commun. (2018) 495:1562–6. doi: 10.1016/j.bbrc.2017.11.183
35. Bryan CF, Cherikh WS, Sesok-Pizzini DA. A2 /A2 B to B renal transplantation: past, present, and future directions. Am J Transplant. (2016) 16:11–20. doi: 10.1111/ajt.13499
36. Bussone G, Dib H, Dimitrov JD, Camoin L, Broussard C, Tamas N, et al. Identification of target antigens of self-reactive IgG in intravenous immunoglobulin preparations. Proteomics. (2009) 9:2253–62. doi: 10.1002/pmic.200800819
37. Avrameas S. Autopolyreactivity confers a holistic role in the immune system. Scand J Immunol. (2016) 83:227–34. doi: 10.1111/sji.12414
38. Ascherio A, Munger KL, Lennette ET, Spiegelman D, Hernán MA, Olek MJ, et al. Epstein-barr virus antibodies and risk of multiple sclerosis: a prospective study. JAMA. (2001) 286:3083–8. doi: 10.1001/jama.286.24.3083
Keywords: NeuGc, anti-Neu5Gc, autoantibodies, endothelial cells, epithelial cells, atheroma, cancers, humans
Citation: Soulillou J-P, Cozzi E and Bach J-M (2020) Challenging the Role of Diet-Induced Anti-Neu5Gc Antibodies in Human Pathologies. Front. Immunol. 11:834. doi: 10.3389/fimmu.2020.00834
Received: 08 January 2020; Accepted: 14 April 2020;
Published: 09 June 2020.
Edited by:
Wilson Savino, Oswaldo Cruz Foundation (Fiocruz), BrazilReviewed by:
Yaqing Qie, University of Texas MD Anderson Cancer Center, United StatesCopyright © 2020 Soulillou, Cozzi and Bach. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jean-Paul Soulillou, amVhbi1wYXVsLnNvdWxpbGxvdUB1bml2LW5hbnRlcy5mcg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.