 Elin Bernson1,2†
Elin Bernson1,2† Karin Christenson1,2,3†
Karin Christenson1,2,3† Silvia Pesce1,2,4
Silvia Pesce1,2,4 Malin Pasanen1,2
Malin Pasanen1,2 Emanuela Marcenaro4,5
Emanuela Marcenaro4,5 Simona Sivori4,5
Simona Sivori4,5 Fredrik B. Thorén1,2*
Fredrik B. Thorén1,2*- 1TIMM Laboratory, Sahlgrenska Cancer Center, University of Gothenburg, Gothenburg, Sweden
- 2Department of Infectious Diseases, Institute of Biomedicine, University of Gothenburg, Gothenburg, Sweden
- 3Department of Oral Microbiology and Immunology, University of Gothenburg, Gothenburg, Sweden
- 4Department of Experimental Medicine, University of Genoa, Genoa, Italy
- 5Center of Excellence for Biomedical Research (CEBR), University of Genoa, Genoa, Italy
Neutrophils are potent effector cells and contain a battery of harmful substances and degrading enzymes. A silent neutrophil death, i.e., apoptosis, is therefore of importance to avoid damage to the surrounding tissue and to enable termination of the acute inflammatory process. There is a pile of evidence supporting the role for pro-inflammatory cytokines in extending the life-span of neutrophils, but relatively few studies have been devoted to mechanisms actively driving apoptosis induction in neutrophils. We have previously demonstrated that natural killer (NK) cells can promote apoptosis in healthy neutrophils. In this study, we set out to investigate how neutrophil sensitivity to NK cell-mediated cytotoxicity is regulated under inflammatory conditions. Using in vitro-activated neutrophils and a human skin chamber model that allowed collection of in vivo-transmigrated neutrophils, we performed a comprehensive characterization of neutrophil expression of ligands to NK cell receptors. These studies revealed a dramatic downregulation of HLA class I molecules in inflammatory neutrophils, which was associated with an enhanced susceptibility to NK cell cytotoxicity. Collectively, our data shed light on the complex regulation of interactions between NK cells and neutrophils during an inflammatory response and provide further support for a role of NK cells in the resolution phase of inflammation.
Introduction
Neutrophils are innate immune cells that play a key role in the defense against invading microbes. In circulation, neutrophils are in a resting state, but in response to proinflammatory signals they become alerted, extravasate and migrate toward the site of damage or infection. Neutrophils contain numerous intracellular granules containing pre-formed proteins needed to execute their functions. Thus, when neutrophils are stimulated, they rapidly mobilize specific types of granules to equip the cell with new surface structures and to release soluble factors to the extracellular space (1). The transmigration process per se is dependent on degranulation, which enables attachment to endothelial cells, and subsequently chemotaxis-driven migration (2). However, the rearrangement of surface structures does not only involve granule mobilization, but also cleavage or internalization of surface structures; CD62L, involved in attachment to endothelial cells, is for example shed from the neutrophil surface already in circulation (3).
Neutrophils have a rich arsenal of toxic substances and degrading enzymes stored in their granules, which are used to eradicate an ingested prey. The toxic content makes neutrophils a potential danger to the surrounding tissue and it is thus of importance that neutrophils are removed from the inflammatory site after they have fulfilled their tasks. A tightly controlled death process where the cell is degraded from the inside while the surface membrane remains intact, is therefore of importance for termination of the inflammatory process (4). Neutrophil apoptosis can be induced by interaction with other immune cells inducing pro-apoptotic signaling via death receptors on the neutrophil surface (5); e.g., we have previously reported that interaction between NK cells and neutrophils can promote neutrophil apoptosis (6).
Natural killer (NK) cells are cytotoxic cells that can kill aberrant cells without prior sensitization (7). Besides their undisputed role in the defense against viral infections and certain malignancies, a growing body of evidence points to a role for NK cells in immune regulation, both as an early source of cytokines but also by selective killing of immune cells (6, 8–13). The result of an encounter between an NK cell and a target cell is determined by the balance between signals originating from inhibitory and activating receptors expressed on the NK cell surface; thus, the presence of cognate ligands to NK cell receptors (NKRs) on the potential target cell determines the outcome of an NK cell—target cell interaction. When the inhibitory signaling is decreased, or the activating signaling increased, the NK cell cytotoxic machinery may be activated, resulting in release of cytotoxic granules into the immunological synapse. Among the major activating receptors are the natural-killer group 2, member D (NKG2D), recognizing MICA and MICB and different ULBPs; DNAX accessory molecule-1 (DNAM-1), recognizing polio virus receptor (PVR) and Nectin-2; 2B4 recognizing CD48; and the group of natural cytotoxicity receptors (NCRs; NKp30, NKp44, and NKp46), which in part constitute orphan receptors (14, 15), while NKp30 recognizes B7-H6 and BAG-6 (16, 17). The main inhibitory receptors are the inhibitory killer immunoglobulin-like receptors (iKIRs) and NKG2A/CD94 that bind to specific human leukocyte antigen (HLA) class I molecules on target cells. HLA-C binds to the KIR2DL receptors, while certain HLA-B and HLA-A molecules contain a Bw4 motif that is recognized by KIR3DL1. The NKG2A/CD94 heterodimer binds to the non-classical HLA-E molecule, which selectively presents the leader peptides of classical HLA molecules and thus reflects the overall expression of HLA class I in a cell. In addition, the inhibitory receptor LILRB1 recognizes HLA class I molecules. Moreover, HLA class I molecules may also bind to activating NK cell receptors, where the NKG2C/CD94 heterodimer binds to HLA-E, and activating KIRs recognize certain motifs on classical HLA class I molecules. The HLA class I molecules can be up- or downregulated from the cell surface in response to cellular signaling, and free soluble HLA class I molecules has been described in serum, either secreted from cells or shed from the cell surfaces due to proteolytic cleavage (18–20).
As mentioned above, NK cells have been ascribed immunomodulatory functions (21, 22) and previous work from our group has demonstrated that NK cells induce apoptosis in healthy neutrophils in an NCR- and Fas-dependent manner (6). Moreover, in a human in vivo blister model of sterile inflammation, NK cell entry into blisters coincided with the appearance of apoptotic neutrophils, suggesting a role for NK cells in terminating an inflammatory response (6). With this background, we hypothesized that modulation of neutrophil surface expression of NKR ligands upon neutrophil activation may lead to altered sensitivity to NK cell-induced apoptosis. In this study, we demonstrate that inflammatory neutrophils, both in vitro-stimulated blood neutrophils and in vivo-transmigrated neutrophils collected from skin chamber exudates, are more susceptible to NK cell-mediated apoptosis, and that the enhanced sensitivity is associated with a pronounced downregulation of neutrophil HLA class I expression.
Results
Surface Expression of HLA Class I Molecules Is Decreased Upon Neutrophil Activation
NK cell cytotoxicity is triggered when the signals from activating receptor-ligand interactions overcome the inhibitory signals from inhibitory receptors. Thus, in a first set of experiments we investigated to what extent activated neutrophils modulate their expression of ligands to NKRs. In in vitro experiments, we combined GM-CSF with the TLR7/8 agonist CL097 (23, 24) to activate freshly isolated neutrophils and screened the neutrophil expression of ligands to activating and inhibiting NKRs using flow cytometry. As demonstrated in Figure 1A, neutrophils stimulated with CL097 and GM-CSF for 20 min at 37°C displayed strong up-regulation of CD11b and CD66, both classical markers for neutrophil activation, as well as loss of CD62L. In parallel, we observed a significant decrease of classical HLA class I molecules after 20 min stimulation at 37°C (HLA-ABC; Figure 1A). These results were confirmed with two additional clones recognizing HLA-ABC (G46-2.6 and A6-136; data not shown). Notably, a significant downregulation of HLA class I was seen also without addition of stimuli (Figure 1A). Furthermore, we observed decreased expression of the DNAM-1 ligand PVR, and increased expression of MICA/B and ULBP-3, both ligands to NKG2D, on the neutrophil surface after 20 min of stimulation with CL097/GM-CSF (Figure 1B), while no robust changes were observed regarding expression levels of Nectin-2, ULBP-1, ULBP-2/5/6, B7-H6, or NKp46-ligand, all ligands to activating NKRs (Supplemental Figure 1). Surface expression of CD48, a ligand to 2B4, could not be detected on neither resting nor activated neutrophils (data not shown). Extended GM-CSF and TLR stimulation further decreased neutrophil surface expression of classical HLA class I molecules, while expression levels of ligands to activating NKRs displayed varying patterns (Figure 1C, Supplemental Figure 1).
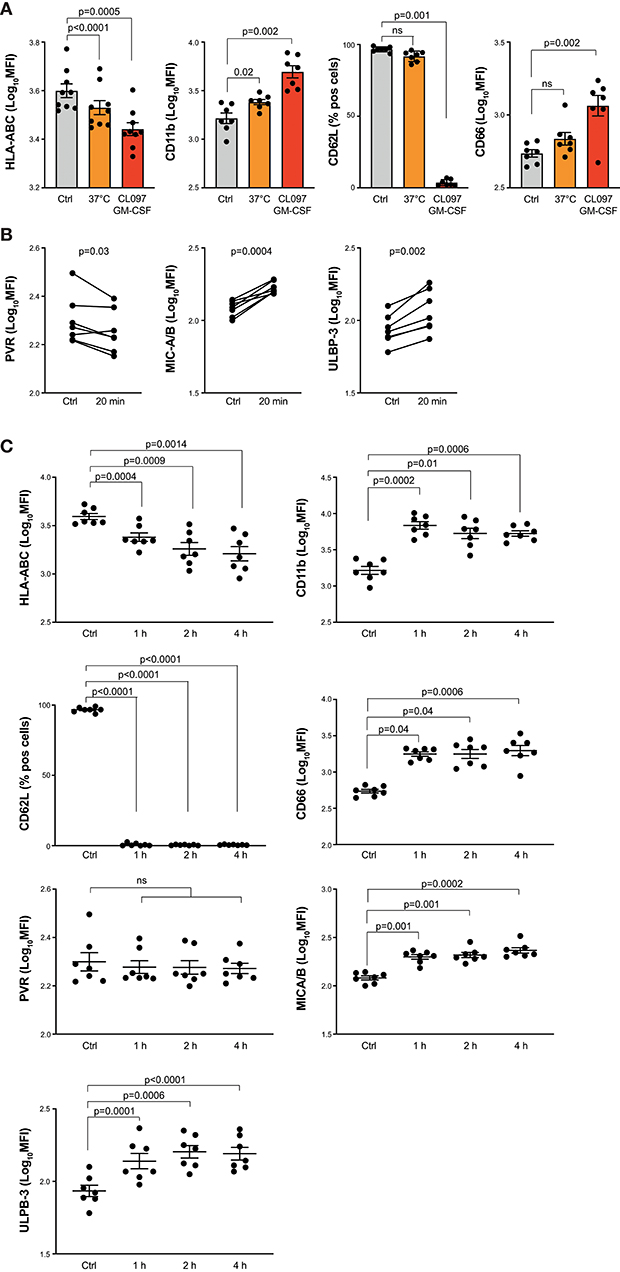
Figure 1. NKR ligand expression on in vitro-activated neutrophils. (A) Surface expression of HLA-ABC, CD11b, CD66 or CD62L as indicated, on resting neutrophils kept on ice (ctrl) or in vitro-activated neutrophils stimulated for 20 min at 37°C alone or in presence of the TLR-agonist CL097 and GM-CSF (one-way ANOVA followed by Dunnett's multiple comparisons test). (B) Surface expression of indicated ligands to NKRs on resting neutrophils (ctrl) or in vitro-activated neutrophils stimulated for 20 min at 37°C with CL097/GM-CSF (paired t-test). (C) Expression of NKR ligands and neutrophil degranulation markers on resting neutrophils (ctrl) or in vitro-activated neutrophils stimulated for 1, 2, or 4 h at 37°C with CL097/GM-CSF (one-way ANOVA followed by Dunnett's multiple comparisons test). Error bars represent SEM.
To investigate whether our observations in neutrophils activated in vitro were consistent with expression patterns on activated, extravasated neutrophils in vivo, we made use of a human in vivo model where transmigrated neutrophils are harvested from a serum-filled skin chamber placed on exposed dermis (25). Transmigrated neutrophils were highly activated as reflected by an increased CD11b expression and loss of CD62L compared to resting autologous neutrophils harvested from blood (Figure 2 and data not shown). Similar to the in vitro-activated neutrophils, transmigrated neutrophils displayed significantly decreased surface expression of classical HLA class I molecules. Consistent with our data from in vitro-stimulated neutrophils, transmigrated neutrophils did not display altered surface expression of Nectin-2 and B7-H6. However, in contrast to in vitro-stimulated neutrophils, transmigrated neutrophils did not display decreased expression of PVR.
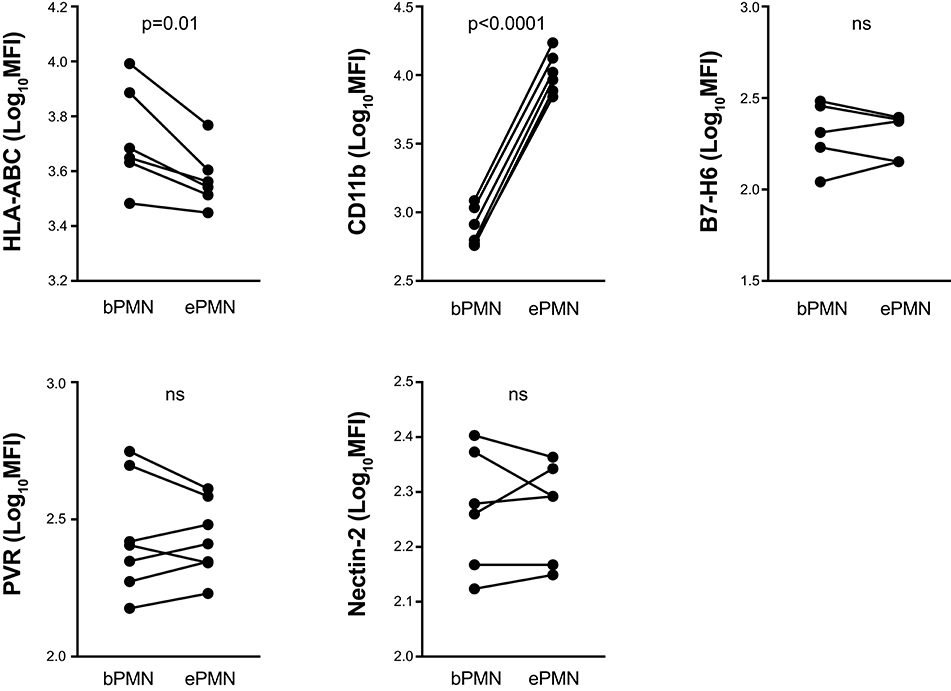
Figure 2. Surface expression of ligands to inhibitory and activating NKRs on transmigrated neutrophils. Surface staining of indicated ligands to NKRs on transmigrated neutrophils (ePMN), collected from exudates in the skin chamber model, and autologous blood neutrophils (bPMN). Paired t-test, error bars represent SEM.
Downmodulation of NK Cell-Regulatory HLA Molecules on Activated Neutrophils
HLA class I expression on hematopoietic cells is dominated by HLA-A and -B, which are approximately 15 times more abundant than HLA-C (26), which in turn is more abundant than the non-classical HLA class I molecule, HLA-E. Given the important role for HLA-C and HLA-E in regulation of NK cell effector functions (27), we next investigated to what extent these HLA molecules were also downregulated in response to neutrophil stimulation. As shown in Figures 3A,B, we observed a strong downregulation of both HLA-C and HLA-E already after 20 min of stimulation. The decreased HLA-C expression was observed also after longer neutrophil stimulation (Figures 3C,D; Supplemental Figure 2). Consistently, we observed a similar picture on transmigrated neutrophils in vivo (Figures 3E,F).
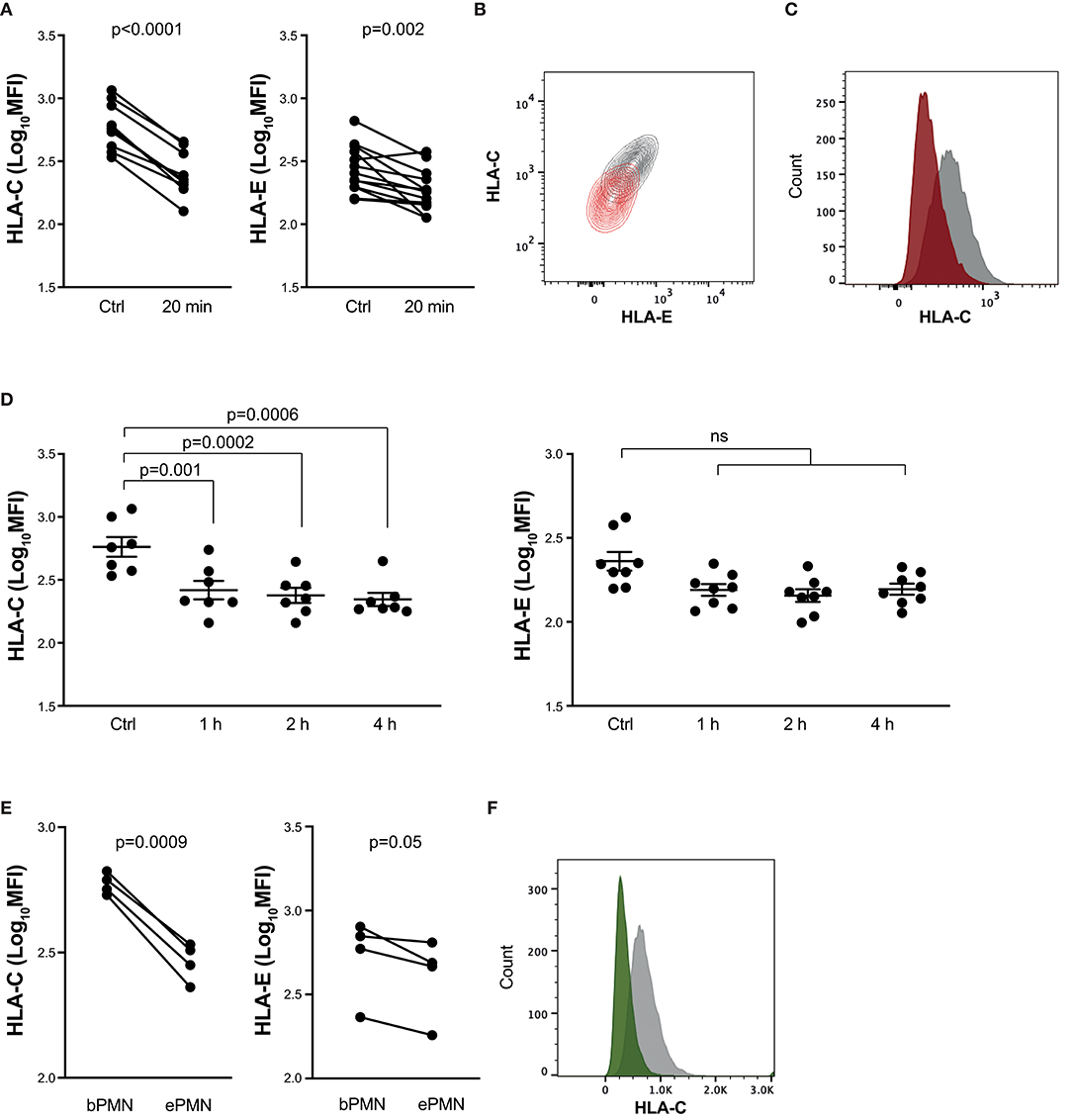
Figure 3. Downmodulation of HLA-C and HLA-E on activated neutrophils. (A–D) Neutrophil staining of indicated HLA class I cell surface structures (HLA-C or HLA-E) binding to inhibitory NKRs. Resting neutrophils were kept on ice (ctrl) or stimulated for 20 min with the TLR-agonist CL097 and GM-CSF at 37°C (A; paired t-test) or for 1, 2, or 4 h under the same conditions (D; one-way ANOVA followed by Dunnett's multiple comparisons test). (B) Representative staining of HLA-C and HLA-E of resting neutrophils (gray) or neutrophils that had been stimulated with CL097/GM-CSF for 20 min (orange). (C) Representative staining from one experiment showing expression of HLA-C on resting neutrophils (gray) or neutrophils that had been stimulated with CL097/GM-CSF for 1 h (red). (E) Neutrophil staining of indicated HLA class I molecules on transmigrated neutrophils from skin chamber exudates (ePMN) or resting autologous neutrophils (bPMN; paired t-test). (F) Representative staining from one experiment showing the expression of HLA-C on resting neutrophils (gray) or transmigrated neutrophils (green). Error bars represent SEM.
In an attempt to understand the mechanism of HLA downmodulation, we investigated whether HLA was internalized upon neutrophil activation. Neutrophils were stained with a FITC-conjugated anti-HLA-ABC antibody prior to stimulation. After 20 min CL097/GM-CSF stimulation, an anti-FITC mAb was added, quenching the extracellular FITC signal. However, as shown in Figure 4, the fluorescence detected from activated neutrophils was quenched to a similar extent as resting neutrophils, suggesting that internalization of HLA is not the main explanation to the HLA downregulation observed on activated neutrophils. Activated neutrophils release granules containing various proteolytic enzymes, such as serine proteases, e.g., elastase, proteinase 3, cathepsin G, and different matrix metalloproteases, and we next addressed to what extent inhibition of specific enzymes could restore HLA expression in activated neutrophils. However, Batimastat, an inhibitor of matrix metalloproteases, did not affect the neutrophil surface expression of HLA class I and neither did α1-antitrypsin nor specific inhibitors of cathepsin G and PR3 (data not shown).
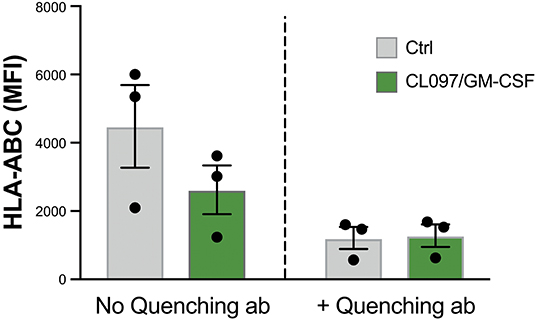
Figure 4. Decrease of surface HLA class I molecules is not explained by internalization. Graph shows resting neutrophils stained with FITC-conjugated HLA-ABC antibody prior to addition of buffer (ctrl) or CL097/GM-CSF for 20 min at 37°C, after which FITC-quenching antibody was added.
Reduced HLA Class I Expression on Activated Neutrophils Translates Into Enhanced Sensitivy to NK Cell Cytotoxicity
As activated neutrophils displayed lower expression of ligands to inhibitory NKRs, and no or modest modulation of their expression of ligands to activating NKRs was detected, we speculated that the outcome of interactions between NK cells and activated vs. resting neutrophils would differ. To investigate the neutrophil sensitivity to NK cell cytotoxicity, the viability of resting or pre-activated neutrophils was determined after co-culture with pre-activated NK cells. As demonstrated in Figures 5A,B, both transmigrated skin chamber neutrophils and in vitro-stimulated neutrophils were more sensitive to NK cell cytotoxicity compared to resting neutrophils, as reflected by increased binding of Annexin V to extracellular phosphatidyl serine and nuclear staining of cells with compromised plasma membrane integrity with To-Pro-3 (Figure 5C). Notably, addition of an HLA antibody, blocking the inhibitory interaction between HLA and KIRs or NKG2A/CD94, resulted in a significantly higher NK cell cytotoxicity against resting neutrophils (Figure 5A). By contrast, the cytotoxicity against activated neutrophils, which display low expression of HLA class I molecules, did not further increase upon HLA class I blockade. Blocking of NCR signaling via NKp46 resulted in decreased NK cell-mediated killing of in vitro-activated neutrophils (Supplemental Figure 3), consistent with our previous findings for resting neutrophils (6).
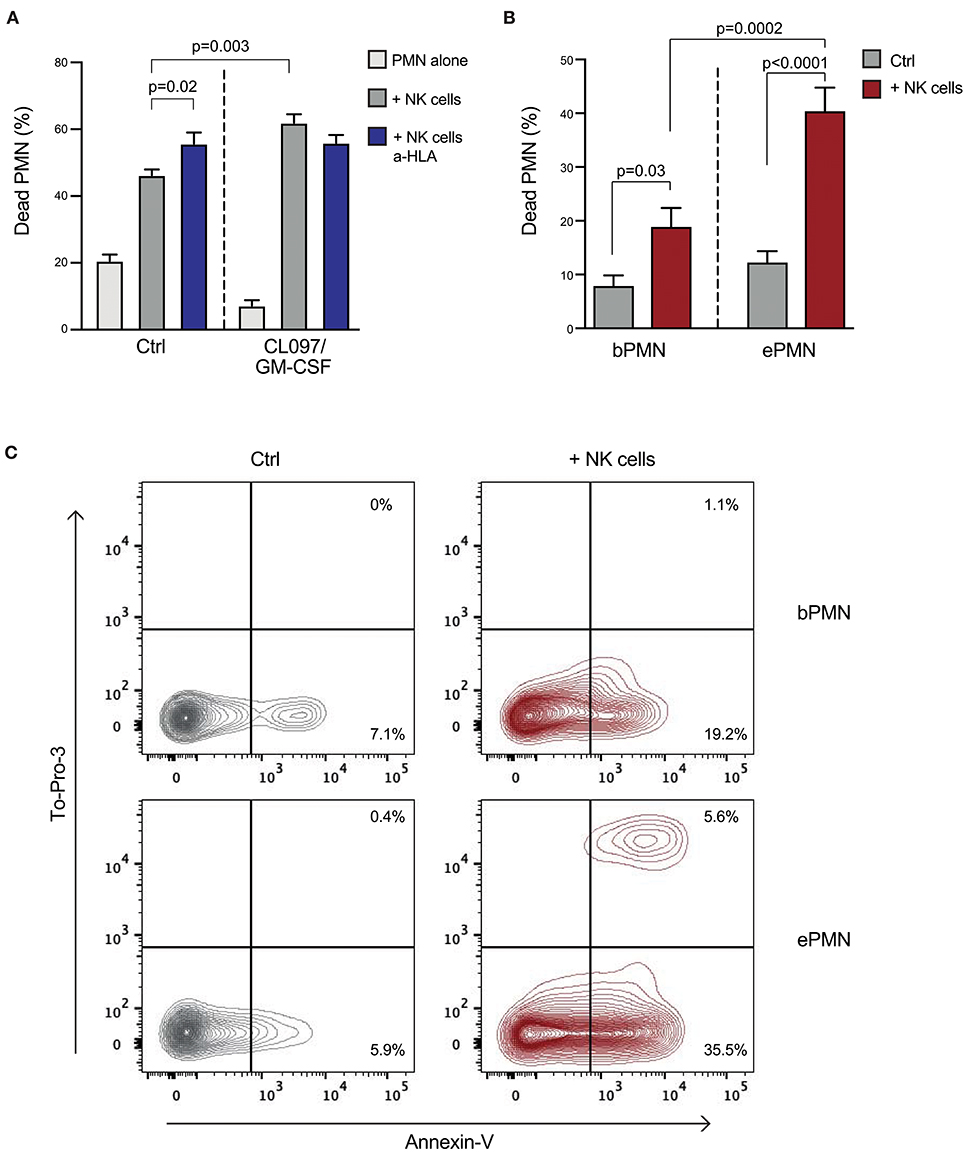
Figure 5. Activated neutrophils are more sensitive to NK cell cytotoxicity compared to resting neutrophils. (A) Percentage of dead neutrophils (PMN) after a 3 h co-culture of neutrophils [pre-stimulated in vitro with CL097 and GM-CSF, or kept on ice (ctrl)] and bulk-NK cells at an E:T ratio of 10:1, with addition of an HLA antibody as indicated (n = 7; one-way ANOVA followed by Dunnett's multiple comparisons test). (B) Graph shows percentage of dead neutrophils after a 4 h cytotoxicity assay performed using autologous NK effector cells toward transmigrated (ePMN) or blood neutrophils (bPMN; E:T ratio 10:1; n = 5; one-way ANOVA followed by Dunnett's multiple comparisons test). Error bars represent SEM. (C) Representative FACS plots showing percentage of dead neutrophils (transmigrated or resting) measured using AnnexinV and To-Pro-3, after a 4 h co-culture with autologous NK cells.
Discussion
After executing their tasks in peripheral tissues, neutrophils, with their cargo of toxins and degrading enzymes, must undergo apoptosis to prevent prolonged inflammation and resulting tissue damage. During the last decade, a growing pile of evidence has identified an important role for NK cells as modulators of the immune response (21, 22, 24) and we have earlier demonstrated that NK cells can induce apoptosis in neutrophils in an NKp46-dependent manner (6). The outcome of NK cell—target cell interactions is dependent not only on cell signaling via activating receptors, but also on signaling via inhibitory receptors. In this study we investigated the neutrophil surface expression of ligands to inhibitory, as well as activating NKRs, on both in vitro-activated neutrophils and in vivo-transmigrated neutrophils isolated using a skin chamber model (25). With the exception of certain NKG2D ligands, the expression of ligands to activating NKRs remained largely unchanged upon neutrophil activation. By contrast, we observed a substantial downregulation of HLA class I molecules on activated neutrophils, which included also less abundant HLA structures of importance for NK cell regulation, HLA-C, and HLA-E.
HLA downregulation is a commonly used immunoevasive strategy among virus-infected cells or tumor cells to avoid T cell recognition and elimination. However, the decreased HLA expression will reduce inhibitory signaling in NK cells and thus allow missing-self recognition by NK cells (27–29). The dramatic downregulation of HLA class I molecules observed in this study necessarily involves downregulation of the abundant HLA subtypes, HLA-A and -B which according to previous reports comprise 95% of total HLA class I (26). Interestingly, our analysis demonstrated that the downregulation of HLA class I on activated neutrophils was not confined to the most abundant HLA class I molecules but also included pronounced downregulation of HLA-C and HLA-E. As HLA class I molecules act as ligands to inhibitory NK cell receptors, we hypothesized that the reduced surface expression of NK cell-specific HLA-C and HLA-E would result in higher sensitivity to NK cell cytotoxicity in inflammatory neutrophils. Indeed, the loss of surface HLA class I molecules rendered inflammatory neutrophils more susceptible to NK cell-mediated killing.
Our data suggest that HLA downregulation on inflammatory neutrophils may serve the purpose to increase their susceptibility to NK cell cytotoxicity and thus contribute to resolution of the inflammatory process by NK-induced neutrophil apoptosis. Chronic inflammatory diseases, such as rheumatoid arthritis, are characterized by an ongoing inflammation and presence of high numbers of inflammatory neutrophils (30). A focus of further studies should be to what extent aberrant HLA expression patterns or absence of certain NK cell phenotypes may be related to disease grade and characteristics. In this regard, it has been reported that the immature, less cytotoxic CD56bright NK cells predominate in the inflamed synovium (31). This finding is in line with our data, as the less cytotoxic CD56bright NK cell population may rather fuel the inflammatory process by producing inflammatory cytokines than kill inflammatory neutrophils.
We were not able to pinpoint the exact mechanism for neutrophil HLA downregulation in this study. It is possible that HLA molecules are internalized upon neutrophil activation. To test this hypothesis, we made use of an anti-FITC antibody, which quenches the FITC. As the neutrophils were stained with FITC-conjugated anti-HLA-ABC antibody before priming, internalization would mean that also the FITC-conjugate was internalized. The quenching antibody can only bind to extracellular FITC, and thus, any internal FITC signal would still fluoresce when hit by light in the right wave length. However, the FITC signal was not higher in activated neutrophils than in resting neutrophils, and we could thus not conclude that HLA was internalized. Alternatively, HLA molecules could be shed from the cell surface. Neutrophil granules contain enzymes, such as the serine proteases, neutrophil elastase, cathepsin G, and proteinase 3, which may be released upon neutrophil transmigration to an inflammatory site (32, 33). Shedding of surface B7-H6 from tumor cells has previously been described as a cause of matrix metalloprotease-mediated shedding (34), and presence of a soluble form of B7-H6 has been reported in supernatants from in vitro-stimulated neutrophils (35). This suggests that also neutrophil modulation of HLA molecules may involve proteolytic shedding. Supporting this hypothesis, the metalloprotease inhibitor Batimastat has been reported to inhibit release of soluble HLA class I in activated lymphocytes (19). In this study, however, we were unable to inhibit HLA class I downregulation on neutrophils with Batimastat, although the Batimastat readily inhibited shedding of CD62L from the activated neutrophils. Notably, a fraction of serine proteases from azurophil granules remain associated with the neutrophil cell membrane after granule-mobilization (33), and there are reports suggesting that membrane-bound forms of these enzymes are relatively insensitive to protease inhibitors (36). Thus, the inability of protease inhibitors to prevent HLA class I downregulation does not preclude a role for these proteases in this process.
A third alternative for the observed downregulation of HLA expression could be that neutrophil activation leads to a change in the HLA conformation, disabling antibody recognition of HLA. Several studies have reported open conformers of HLA, with the conformation change leading to a shift in receptor—ligand interactions (37–39). Surface staining of resting and activated neutrophils using an antibody recognizing an open conformation of HLA class I did not reveal any increase in HLA class I open conformers on activated neutrophils (data not shown). Nevertheless, in the study we observed HLA class I downregulation on activated neutrophils using five different antibody clones detecting HLA molecules (clones G46-2.6, W6/32 and A6-136 against HLA-ABC, clone DT9 against HLA-C, and clone 3D12 against HLA-E) strongly suggesting that HLA class I is decreased on the surface of inflammatory neutrophils.
In conclusion, we have demonstrated that both in vitro-activated and in vivo-transmigrated neutrophils display decreased surface expression of HLA class I molecules. In line with the decreased HLA class I expression, inflammatory neutrophils were more sensitive to NK cell-mediated cytotoxicity, as the loss of inhibitory signaling tips the balance toward NK cell activation and cytotoxicity. Collectively, our study demonstrates that neutrophils dynamically modulate their expression and release of NK cell receptor ligands, and that NK cells may contribute to a controlled resolution of the inflammatory process by killing activated neutrophils.
Materials and Methods
Antibodies and Reagents
Following mAbs were used for detection of surface markers: anti-CD11b APC (clone: ICRF44), CD48 BV421 (TÜ145), HLA-ABC FITC (G46-2.6), CD62L APC (DREG-56), CD66 PE (B1.1; all from BD Biosciences, CA, USA), HLA-C (DT9, Merck Millipore), HLA-ABC Alexa Fluor 647 (W6/32) and HLA-E PE-Cy7 (3D12; both from Biolegend), B7-H6 (875002), ULBP1 PerCP (170818), ULBP-3 PE (166510), ULBP-2/5/6 APC (165903; all from R&D Systems, MN, USA), CD56 PE-Cy7 (N901, Beckman Coulter, IN, USA), MICA/B viobright FITC (6D4, Miltenyi Biotech, Bergisch Gladbach, Germany), and Alexa Fluor 488-conjugated goat anti-mouse IgG (Invitrogen, CA, USA). Anti-B7-H6 (17B1.3) was kindly provided by Prof E Vivier (CMIL, Marseille, France), and anti-PVR (CD155; M5A10) and Nectin-2 (CD112; L14) were received from the lab of Prof. A Moretta (Genova, Italy). Goat serum and α1-antitrypsin were obtained from Sigma-Aldrich (MO, USA), CL097 from InvivoGen (San Diego, CA), GM-CSF from R&D Systems (MN, USA), anti-PR3 from Abcam (UK), and Batimastat and cathepsin G inhibitor from Calbiochem (CA, USA). In experiments where the extracellular FITC signal was quenched, a Fluorescin/Oregon green polyclonal antibody (ThermoFisher, Sweden) was used.
Isolation of Human Leukocytes
Buffy coats or fresh blood obtained from healthy donors were mixed 1:1 with 2% dextran. After sedimentation of erythrocytes, granulocytes and PBMCs were separated by density gradient centrifugation (40, 41). Remaining erythrocytes were lysed in distilled water to obtain a pure granulocyte population. This method provides a granulocyte population of >95% purity. NK cells were isolated from PBMCs using an NK cell isolation kit (Miltenyi Biotec, Bergisch Gladbach, Germany), according to the manufacturer's protocol. To obtain activated, polyclonally expanded NK cells (bulk), freshly isolated NK cells were cultured on irradiated feeder cells in the presence of 100 U/mL recombinant human IL-2 (Proleukin; Chiron) and 1.5 ng/ml phytohemagglutinin (PHA) (GIBCO Ltd.).
Isolation of Transmigrated Neutrophils
In order to isolate in vivo-transmigrated neutrophils, a skin chamber technique was used, described in detail elsewhere (25, 42). In short, blisters were introduced on the forearm on healthy volunteers using negative pressure. After 2 h, the blister roofs were removed and collection chambers were placed on top of the lesions. Autologous serum was added to the chamber wells. After 24 h, the chambers were removed and in vivo-transmigrated neutrophils were harvested from the skin chamber fluid, with a granulocyte purity >90%, and with mainly viable neutrophils (42). Autologous blood neutrophils were isolated from peripheral blood as described above and used as control cells. All neutrophils were diluted in Krebs Ringer Glucose (KRG) supplemented with Ca2+, and stored on melting ice until use. The study was approved by the regional ethical board in Gothenburg (543-07) and voluntary donors gave written informed consent in accordance with the declaration of Helsinki.
Generation of Fusion Proteins for Detection of NKp46 Ligand
HEK293T cells, transfected with a construct encoding for a fusion protein consisting of the extracellular part of NKp46 fused to an Fc-portion of human IgG, were kindly provided by Prof. O Mandelboim (Jerusalem, Israel). Details of the transfection procedure has been described elsewhere (43, 44). Cells were cultured in DMEM (Sigma-Aldrich) supplemented with 10% heat inactivated fetal calf serum (FCS), 1% PEST, 1% L-glutamine and Puromycin dihydrochloride (5 μg/ml; Sigma-Aldrich). Cell cultures were kept at 37°C in a humidified, 5% CO2 atmosphere. When reaching 70% confluency, medium was exchanged to low protein medium supplemented with non-essential amino acids, 1% Penicillin-Streptomycin, 1% L-glutamine and 1% Sodium Pyruvate. After 3 days, the cell-free supernatant was collected and fusion proteins were purified with affinity chromatography using a HiTrap Protein G column (GE Healthcare). Proteins were eluted with 0.1 M Glycine/HCl (pH 2,7-3), neutralized in 1 M TRIS/HCl (pH 8), concentrated using an Amicon Ultra 15 centrifugation tube (Millipore) and stored in buffered NaCl in −20°C until use.
Expression of Surface Ligands on Neutrophils
Freshly isolated neutrophils were stimulated with CL097/GM-CSF (1 μg/ml and 100 U/ml, respectively), at 37°C for indicated time. In some experiments, the matrix metalloproteinase (MMP) inhibitor Batimastat (10 μM), α1-antitrypsin (1 μg/ml), cathepsin G inhibitor (1 μM) or anti-PR3 antibody (100 ng/ml) were added to the cells. Supernatants were collected after priming for detection of soluble HLA. Cells were stained with antibodies for 30 min at 4°C. Degranulation/activation was monitored using antibodies directed against the surface markers CD11b, CD62L, and CD66. For detection of unconjugated antibodies (HLA-C, B7-H6, PVR and Nectin-2), cells were saturated in 20% goat serum for 10 min prior to incubation with Alexa Fluor-488 conjugated goat anti-mouse IgG. Samples were collected on a three-laser FACSAria (405, 488, and 633 nm) and data was analyzed using FlowJo software (TreeStar; version 9.7.2 or later).
For detection of NKp46 ligand, neutrophils were treated with the streptoccal enzyme EndoS, kindly provided by Dr. Mattias Collin (Lund University, Sweden), at 37°C for 20 min to digest endogenous IgG bound to the surface of the cells (45). Cells were subsequently incubated with fusion protein (10 μg/ml) for 30 min at 4°C. Cells were saturated in 20% goat serum prior to staining with FITC-conjugated Fcγ-specific goat anti-human IgG (Jackson Immunoresearch Laboratories, PA, USA).
Detection of NK Cell-Induced Neutrophil Apoptosis
Freshly isolated neutrophils were stimulated with CL097/GM-CSF (1 μg/ml and 100 U/ml, respectively), or kept at 4°C without stimuli, for 1 h. After stimulation the neutrophils were co-cultured with polyclonal IL-2 activated NK cells at a 10:1 E:T ratio for 3 h at 37°C in the presence or absence of anti-HLA-I mAb (clone A6136, IgM), or anti-NKp46 (clone KL247, IgM). Transmigrated neutrophils collected from skin chamber exudates or autologous neutrophils isolated from blood were co-cultured with autologous NK cells, stimulated in IL-12 (1 ng/ml) or IL-15 (10 ng/ml) overnight. Apoptosis and lysis of neutrophils were detected with FITC-conjugated AnnexinV (BD Biosciences, CA, USA) and To-Pro-3 (Invitrogen) using flow cytometry (46).
Statistics
For single pairwise comparisons, paired t-test was used. For multiple comparisons within a data set, one-way ANOVA followed by Dunnett's multiple comparisons test, was used.
Data Availability Statement
The raw data supporting the conclusions of this manuscript will be made available by the authors, without undue reservation, to any qualified researcher.
Ethics Statement
The studies involving human participants were reviewed and approved by The Regional Ethics Board in Gothenburg. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
EB, KC, SP, and FT designed the research. EB, KC, SP, and MP performed the research and analyzed the data. EM, SS, and FT supervised the study. EB, KC, and FT drafted the manuscript. All authors contributed to the final version of the manuscript.
Funding
This work was supported by the Swedish Research Council (Vetenskapsrådet, 2016-03181), Swedish Cancer Society (Cancerfonden, CAN 2016/813), the Swedish state via the ALF agreement (ALFGBG-724861), IngaBritt and Arne Lundberg Foundation (2016-0036), Assar Gabrielsson Foundation (FB-18-32), Wilhelm and Martina Lundgren Research Foundation (2017-1948), OE och Edla Johanssons vetenskapliga stiftelse, Magnus Bergwalls stiftelse, SU fonder, BioCARE, the Sahlgrenska Academy at University of Gothenburg, and Fondazione Associazione Italiana per la Ricerca sul Cancro and Fondazione Roche (Progetto Roche per la Ricerca). SP is recipient of the post-doctoral fellowship from the Fondazione Umberto Veronesi.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2019.02444/full#supplementary-material
References
1. Borregaard N, Cowland JB. Granules of the human neutrophilic polymorphonuclear leukocyte. Blood. (1997) 89:3503–21.
2. Ley K, Laudanna C, Cybulsky MI, Nourshargh S. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nat Rev Immunol. (2007) 7:678–89. doi: 10.1038/nri2156
3. Kuhns DB, Long Priel DA, Gallin JL. Loss of L-selectin (CD62L) on human neutrophils following exudation in vivo. Cell Immunol. (1995) 164:306–10. doi: 10.1006/cimm.1995.1174
4. Fox S, Leitch AE, Duffin R, Haslett C, Rossi AG. Neutrophil apoptosis: relevance to the innate immune response and inflammatory disease. J Innate Immun. (2010) 2:216–27. doi: 10.1159/000284367
5. El Kebir D, Filep JG. Role of neutrophil apoptosis in the resolution of inflammation. Sci World J. (2010) 10:1731–48. doi: 10.1100/tsw.2010.169
6. Thoren FB, Riise RE, Ousback J, Della Chiesa M, Alsterholm M, Marcenaro E, et al. Human NK Cells induce neutrophil apoptosis via an NKp46- and Fas-dependent mechanism. J Immunol. (2012) 188:1668–74. doi: 10.4049/jimmunol.1102002
7. Kiessling R, Klein E, Pross H, Wigzell H. Natural killer cells in the mouse. II Cytotoxic cells with specificity for mouse Moloney leukemia cells Characteristics of the killer cell. Eur J Immunol. (1975) 5:117–21. doi: 10.1002/eji.1830050209
8. Orange JS, Wang B, Terhorst C, Biron CA. Requirement for natural killer cell-produced interferon gamma in defense against murine cytomegalovirus infection and enhancement of this defense pathway by interleukin 12 administration. J Exp Med. (1995) 182:1045–56. doi: 10.1084/jem.182.4.1045
9. Martin-Fontecha A, Thomsen LL, Brett S, Gerard C, Lipp M, Lanzavecchia A, et al. Induced recruitment of NK cells to lymph nodes provides IFN-gamma for T(H)1 priming. Nat Immunol. (2004) 5:1260–5. doi: 10.1038/ni1138
10. Nedvetzki S, Sowinski S, Eagle RA, Harris J, Vély F, Pende D, et al. Reciprocal regulation of human natural killer cells and macrophages associated with distinct immune synapses. Blood. (2007) 109:3776–85. doi: 10.1182/blood-2006-10-052977
11. Vivier E, Raulet DH, Moretta A, Caligiuri MA, Zitvogel L, Lanier LL, et al. Innate or adaptive immunity? The example of natural killer cells Science. (2011) 331:44–9. doi: 10.1126/science.1198687
12. Waggoner SN, Cornberg M, Selin LK, Welsh RM. Natural killer cells act as rheostats modulating antiviral T cells. Nature. (2011) 481:394–8. doi: 10.1038/nature10624
13. Aydin E, Johansson J, Nazir FH, Hellstrand K, Martner A. Role of NOX2-derived reactive oxygen species in NK cell-mediated control of murine melanoma metastasis. Cancer Immunol Res. (2017) 5:804–11. doi: 10.1158/2326-6066.CIR-16-0382
14. Narni-Mancinelli E, Gauthier L, Baratin M, Guia S, Fenis A, Deghmane AE, et al. Complement factor P is a ligand for the natural killer cell-activating receptor NKp46. Sci Immunol. (2017) 2:eaam9628. doi: 10.1126/sciimmunol.aam9628
15. Gaggero S, Bruschi M, Petretto A, Parodi M, Del Zotto G, Lavarello C, et al. Nidogen-1 is a novel extracellular ligand for the NKp44 activating receptor. Oncoimmunology. (2018) 7:e1470730. doi: 10.1080/2162402X.2018.1470730
16. Pogge von Strandmann E, Simhadri VR, von Tresckow B, Sasse S, Reiners KS, Hansen HP, et al. Human leukocyte antigen-B-associated transcript 3 is released from tumor cells and engages the NKp30 receptor on natural killer cells. Immunity. (2007) 27:965–4. doi: 10.1016/j.immuni.2007.10.010
17. Brandt CS, Baratin M, Yi EC, Kennedy J, Gao Z, Fox B, et al. The B7 family member B7-H6 is a tumor cell ligand for the activating natural killer cell receptor NKp30 in humans. J Exp Med. (2009) 206:1495–503. doi: 10.1084/jem.20090681
18. Krangel MS. Secretion of HLA-A and -B antigens via an alternative RNA splicing pathway. J Exp Med. (1986) 163:1173–90. doi: 10.1084/jem.163.5.1173
19. DeVito-Haynes LD, Demaria S, Bushkin Y, Burlingham WJ. The metalloproteinase-mediated pathway is essential for generation of soluble HLA class I proteins by activated cells in vitro: proposed mechanism for soluble HLA release in transplant rejection. Hum Immunol. (1998) 59:426–34. doi: 10.1016/S0198-8859(98)00032-9
20. DeVito-Haynes LD, Jankowska-Gan E, Meyer KC, Cornwell RD, Zeevi A, Griffith B, et al. Soluble donor HLA class I and beta 2m-free heavy chain in serum of lung transplant recipients: steady-state levels and increases in patients with recurrent CMV infection, acute rejection episodes, and poor outcome. Hum Immunol. (2000) 61:1370–82. doi: 10.1016/S0198-8859(00)00210-X
21. Moretta A. Natural killer cells and dendritic cells: rendezvous in abused tissues. Nat Rev Immunol. (2002) 2:957–64. doi: 10.1038/nri956
22. Vivier E, Tomasello E, Baratin M, Walzer T, Ugolini S. Functions of natural killer cells. Nat Immunol. (2008) 9:503–10. doi: 10.1038/ni1582
23. Wang JP, Bowen GN, Padden C, Cerny A, Finberg RW, Newburger PE, et al. Toll-like receptor-mediated activation of neutrophils by influenza A virus. Blood. (2008) 112:2028–34. doi: 10.1182/blood-2008-01-132860
24. Riise RE, Bernson E, Aurelius J, Martner A, Pesce S, Della Chiesa M, et al. TLR-stimulated neutrophils instruct NK cells to trigger dendritic cell maturation and promote adaptive T cell responses. J Immunol. (2015) 195:1121–8. doi: 10.4049/jimmunol.1500709
25. Follin P. Skin chamber technique for study of in vivo exudated human neutrophils. J Immunol Methods. (1999) 232:55–65. doi: 10.1016/S0022-1759(99)00153-2
26. Apps R, Meng Z, Del Prete GQ, Lifson JD, Zhou M, Carrington M. Relative expression levels of the HLA class-I proteins in normal and HIV-infected cells. J Immunol. (2015) 194:3594–600. doi: 10.4049/jimmunol.1403234
27. Long EO, Kim HS, Liu D, Peterson ME, Rajagopalan S. Controlling natural killer cell responses: integration of signals for activation and inhibition. Annu Rev Immunol. (2013) 31:227–58. doi: 10.1146/annurev-immunol-020711-075005
28. Jost S, Altfeld M. Control of human viral infections by natural killer cells. Annu Rev Immunol. (2013) 31:163–94. doi: 10.1146/annurev-immunol-032712-100001
29. Korner C, Simoneau CR, Schommers P, Granoff M, Ziegler M, Holzemer A, et al. HIV-1-mediated downmodulation of HLA-C impacts target cell recognition and antiviral activity of NK cells. Cell Host Microbe. (2017) 22:111–9.e114. doi: 10.1016/j.chom.2017.06.008
30. Wright HL, Moots RJ, Edwards SW. The multifactorial role of neutrophils in rheumatoid arthritis. Nat Rev Rheumatol. (2014) 10:593–601. doi: 10.1038/nrrheum.2014.80
31. Dalbeth N, Callan MF. A subset of natural killer cells is greatly expanded within inflamed joints. Arthritis Rheum. (2002) 46:1763–72. doi: 10.1002/art.10410
32. Scheja A, Forsgren A, Ohlsson K. Kinetics of enzymes released from polymorphonuclear leucocytes in a skin chamber. Clin Exp Rheumatol. (1986) 4:37–41.
33. Pham CT. Neutrophil serine proteases: specific regulators of inflammation. Nat Rev Immunol. (2006) 6:541–50. doi: 10.1038/nri1841
34. Schlecker E, Fiegler N, Arnold A, Altevogt P, Rose-John S, Moldenhauer G, et al. Metalloprotease-mediated tumor cell shedding of B7-H6, the ligand of the natural killer cell-activating receptor NKp30. Cancer Res. (2014) 74:3429–40. doi: 10.1158/0008-5472.CAN-13-3017
35. Matta J, Baratin M, Chiche L, Forel JM, Cognet C, Thomas G, et al. Induction of B7-H6, a ligand for the natural killer cell-activating receptor NKp30, in inflammatory conditions. Blood. (2013) 122:394–404. doi: 10.1182/blood-2013-01-481705
36. Campbell EJ, Campbell MA, Owen CA. Bioactive proteinase 3 on the cell surface of human neutrophils: quantification, catalytic activity, and susceptibility to inhibition. J Immunol. (2000) 165:3366–74. doi: 10.4049/jimmunol.165.6.3366
37. Arosa FA, Santos SG, Powis SJ. Open conformers: the hidden face of MHC-I molecules. Trends Immunol. (2007) 28:115–23. doi: 10.1016/j.it.2007.01.002
38. Burian A, Wang KL, Finton KA, Lee N, Ishitani A, Strong RK, et al. HLA-F and MHC-I open conformers bind natural killer cell Ig-like receptor KIR3DS1. PLoS ONE. (2016) 11:e0163297. doi: 10.1371/journal.pone.0163297
39. Garcia-Beltran WF, Holzemer A, Martrus G, Chung AW, Pacheco Y, Simoneau CR, et al. Open conformers of HLA-F are high-affinity ligands of the activating NK-cell receptor KIR3DS1. Nat Immunol. (2016) 17:1067–74. doi: 10.1038/ni.3513
40. Bøyum A. Isolation of mononuclear cells and granulocytes from human blood. Isolation of monuclear cells by one centrifugation, and of granulocytes by combining centrifugation and sedimentation at 1 g. Scand J Clin Lab Invest Suppl. (1968) 97:77–89.
41. Bøyum A, Løvhaug D, Tresland L, Nordie EM. Separation of leucocytes: improved cell purity by fine adjustments of gradient medium density and osmolality. Scand J Immunol. (1991) 34:697–712. doi: 10.1111/j.1365-3083.1991.tb01594.x
42. Christenson K, Bjorkman L, Karlsson J, Sundqvist M, Movitz C, Speert DP, et al. In vivo-transmigrated human neutrophils are resistant to antiapoptotic stimulation. J Leukoc Biol. (2011) 90:1055–63. doi: 10.1189/jlb.0111051
43. Mandelboim O, Malik P, Davis DM, Jo CH, Boyson JE, Strominger JL. Human CD16 as a lysis receptor mediating direct natural killer cell cytotoxicity. Proc Natl Acad Sci USA. (1999) 96:5640–4. doi: 10.1073/pnas.96.10.5640
44. Mandelboim O, Lieberman N, Lev M, Paul L, Arnon TI, Bushkin Y, et al. Recognition of haemagglutinins on virus-infected cells by NKp46 activates lysis by human NK cells. Nature. (2001) 409:1055–60. doi: 10.1038/35059110
45. Collin M, Olsén A. EndoS, a novel secreted protein from Streptococcus pyogenes with endoglycosidase activity on human IgG. EMBO J. (2001) 20:3046–55. doi: 10.1093/emboj/20.12.3046
Keywords: neutrophil, NK cell, HLA class I, immunoregulation, neutrophil apoptosis
Citation: Bernson E, Christenson K, Pesce S, Pasanen M, Marcenaro E, Sivori S and Thorén FB (2019) Downregulation of HLA Class I Renders Inflammatory Neutrophils More Susceptible to NK Cell-Induced Apoptosis. Front. Immunol. 10:2444. doi: 10.3389/fimmu.2019.02444
Received: 22 May 2019; Accepted: 01 October 2019;
Published: 15 October 2019.
Edited by:
Christian Jan Lood, University of Washington, United StatesReviewed by:
Amir Horowitz, Icahn School of Medicine at Mount Sinai, United StatesMatthew Parsons, Emory University, United States
Copyright © 2019 Bernson, Christenson, Pesce, Pasanen, Marcenaro, Sivori and Thorén. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fredrik B. Thorén, ZnJlZHJpay50aG9yZW5AZ3Uuc2U=
†These authors shared first authorship