 Rubén López-Santiago1
Rubén López-Santiago1 Ana Beatriz Sánchez-Argáez1
Ana Beatriz Sánchez-Argáez1 Liliana Gabriela De Alba-Núñez1
Liliana Gabriela De Alba-Núñez1 Shantal Lizbeth Baltierra-Uribe2
Shantal Lizbeth Baltierra-Uribe2 Martha Cecilia Moreno-Lafont1*
Martha Cecilia Moreno-Lafont1*- 1Departamento de Inmunología, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, Mexico City, Mexico
- 2Departamento de Microbiología, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, Mexico City, Mexico
Brucellosis is one of the most prevalent bacterial zoonosis of worldwide distribution. The disease is caused by Brucella spp., facultative intracellular pathogens. Brucellosis in animals results in abortion of fetuses, while in humans, it frequently manifests flu-like symptoms and a typical undulant fever, being osteoarthritis a common complication of the chronic infection. The two most common ways to acquire the infection in humans are through the ingestion of contaminated dairy products or by inhalation of contaminated aerosols. Brucella spp. enter the body mainly through the gastrointestinal and respiratory mucosa; however, most studies of immune response to Brucella spp. are performed analyzing models of systemic immunity. It is necessary to better understand the mucosal immune response induced by Brucella infection since this is the main entry site for the bacterium. In this review, some virulence factors and the mechanisms needed for pathogen invasion and persistence are discussed. Furthermore, some aspects of local immune responses induced during Brucella infection will be reviewed. With this knowledge, better vaccines can be designed focused on inducing protective mucosal immune response.
Introduction
The discovery of brucellosis took place in Malta during the Crimean war in 1859. The British troops had been suffering high fever, so the Royal British medical staff was called in. David Bruce was the officer who investigated what he called Malta fever or Mediterranean fever. In 1887, he managed to isolate and cultivate the bacterium responsible for the disease (1).
Later, Themistocles Zammit found that people who lived in farms, reared goats, and drank milk from these animals showed the same symptoms as Mediterranean fever patients. Using this finding, Zammit reproduced the infection in healthy goats and successfully isolated the bacterium in blood and milk. He deduced that the British army contracted the infection by consuming milk from infected animals in the local region. Therefore, in 1906 a decision was made to ban goat's milk consumption as a preemptive measure to control the disease among the British army. However, Malta fever was not eradicated in the region and suspicions arose regarding the consumption of ice-cream, cheese, and fudge made from contaminated milk (1, 2).
Zammit's contribution proved that brucellosis is mostly transmitted orally. Later, other contagion routes were reported (respiratory, parenteral, or by contact) and considered occupational hazards.
Although it has been well-established that Brucella enters the organism orally, the bacterium has not been properly defined as enteropathogenic, which has caused certain controversy. The infection by Brucella spp. does not cause diarrhea, a characteristic symptom of enteropathogenic bacteria (3, 4). Diarrhea is caused by inflammation due to the recruitment of cells, as neutrophils, producing damage to the epithelium, and compromising the integrity of the mucosa, which triggers inflammatory diarrhea (5). Studies in vivo have demonstrated that mice intragastrically inoculated with B. melitensis do not recruit neutrophils in the small intestine, while histological sections do not show considerable inflammation (6). On the contrary, enteropathogens as Salmonella, Vibrio cholera, and Escherichia coli do cause the characteristic inflammatory reaction and trigger diarrhea (5). Additionally, Brucella seems to transit only the intestine and does not appear to invade it and create as replicative niche in it (3, 4).
Animal models in which Brucella spp. is orally or intragastrically inoculated show that the bacterium can be recovered from the small intestine, particularly from gut-associated lymphoid tissue (GALT) as Peyer's patches (PP) and mesenteric lymph nodes (MLN). The bacterium is located in these tissues from an early stage of the infection and up to 21 days afterwards, as demonstrated by plating homogenates from the organs. This suggests that Brucella remains at these sites to replicate (6). However, infection in these models is achieved using high bacterial doses, 108-1010 colony forming units (CFU), which might force gastrointestinal tract tissues to be colonized by the bacterium (3, 4).
Although Brucella spp. enters the body mainly through the mucous membranes of the gastrointestinal tract and the respiratory tract, most of the anti-Brucella immunity studies performed so far use experimental models of parenteral infection, mainly intraperitoneal, which gives easy and rapid access to the spleen and other organs of the reticuloendothelial system (Figure 1). In this review we will discuss the concepts of innate mucosal immunity involved in mucosal protection against Brucella infection. The mechanisms used by Brucella spp. to evade the elements of oral, intestinal, and repiratory mucosal immunity will also be analyzed. In addition, the recent concept of T-cell-mediated immunity operating in the intestinal mucosa, whose characteristics differ from the systemic immunity developing in the spleen, will be discussed. The understanding of Brucella's interactions with the elements of mucosal immunity and the possibilities of experimental models should allow this information to be used to explain why this infectious disease tends to be chronic, and what will be the best vaccination strategies through these pathways.
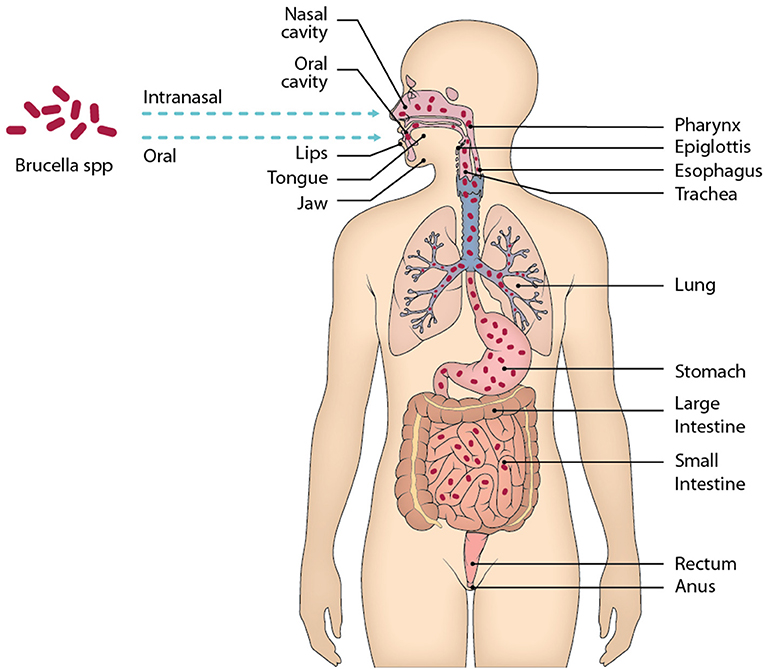
Figure 1. Main mucous membranes affected by the entry of Brucella through the oral and intranasal routes. When ingesting food contaminated with Brucella, the oral cavity is the first site of contact of the bacteria with the host, although it is very little time that remains there, in the oral cavity there are elements of the immune system belonging to MALT that should recognize the presence of Brucella and eliminate it. After the oral cavity Brucella could enter the gastrointestinal tract along with the alimentary bolus through the esophagus up to the stomach. In the stomach Brucella is apparently able to resist the pH of gastric juices, then enters the small intestine, where it will face physical and chemical barriers, as well as different cell lines and lymphoid tissue belonging to GALT. Following this route is likely to reach the large intestine and even that the bacteria was eliminated by feces, however unknown.
Oral Cavity, First Contact Site for Brucella spp.
The oral cavity is the first site of contact of Brucella spp. with the host and it is provided with an immune system mechanism belonging to mucosa-associated lymphoid tissue (MALT). Therefore, Brucella should initiate an immune recognition response in this tissue. The oral cavity is a highly hostile site: not only is it exposed to pathogens that enter by this route, as Brucella, E. coli, and Samonella, but it is also in constant contact with air antigens, food, or microbiota, and is subjected to mechanical damage by mastication. Consequently, the oral mucosa has defense mechanisms and tolerance similar to GALT (7, 8).
Initially, when Brucella enters the oral cavity, it encounters barriers such as saliva, which contains elements that eliminate or control microbial growth, such as lysozyme, lactoferrin, nystadine, peroxidases and immunoglobulins, mainly type A (IgA). Along with saliva, there is also the gingival crevicular fluid that covers the space between the teeth and the gingiva, known as the gingival sulcus. This fluid contains complement molecules, antibodies, neutrophils, and plasma cells. Therefore, the mixture of saliva and gingival crevicular fluid forms an initial strong barrier against pathogenic microorganisms (Figure 1). However, it is unknown whether Brucella is susceptible to this barrier or able to evade it (8).
In the mucosal tissue there are also phagocytic cells that recognize pathogens as Brucella. Phagocytic cells as dendritic cells and macrophages (antigen-presenting cells, APCs) able to capture antigens and migrate to the nearest regional lymph node, in this case the cervical lymph node, are distributed along the specialized tissue of the oral cavity. Once they have captured antigens from the oral mucosa, APCs migrate to the lymph node to present the antigen to the lymphocytes and send the appropriate activation signal (8, 9).
In spite of being the first contact Brucella has with the host, the response of the immune system against Brucella in this site has been poorly studied. Still, there are reports of brucellosis patients showing inflammation in the cervical lymph nodes who had apparently become infected by consuming products contaminated with Brucella. These findings led to the development of an animal model inducing infection with B. melitensis by three different routes of administration for subsequent comparison. The first one consisted of depositing the bacteria directly in the oral cavity of the mouse, the second one involved the administration of the bacteria with an oropharyngeal probe, while the third way of administration was performed intraperitoneally. It was found that, regardless of the administration route, Brucella infection induced inflammation of the cervical lymph node. The viable bacterium was also recovered from the first days after infection and Brucella was determined to persist in the GLN until 50 days after inoculation. Regardless of the administration route and because more bacteria were found in GLN than in other tissues, it is suspected that GLN may be a selective Brucella niche (10).
This suggests that from the first moment after ingesting food contaminated with Brucella, the bacterium is recognized and taken by some APCs belonging to the oral mucosa and later migrates to the cervical lymph node. Cell analysis of the inflamed cervical lymph node from the model described above indicates that there is an increase in CD68+ cells, a marker expressed by macrophages (10).
Knowledge of the passage of Brucella in the oral mucosa is practically unknown; for example, whether there is a participation of amygdales and/or adenoids, which are lymphoid tissue of the oral cavity. Similarly, it is unknown if the recognition of Brucella produces an inflammatory response in the epithelium of the oral cavity or if the response induced in the GLN extends to other sites of the host.
The oral cavity has been considered very little as the body's first point of contact with Brucella spp. Since the oral mucosa is the initial site of infection, more attention should be paid to the role of the lymph nodes draining to the head and neck. It is therefore possible that the oral route is a potential vaccination route.
Interplay of Brucella Through the Gastrointestinal Tract
The gastrointestinal tract is daily exposed to a wide variety of innocuous antigens from foods and microbiota, and, eventually, pathogenic antigens (11–14). The integrity of the intestinal tissue depends on a local immune system consisting of physical barriers, molecules, several cell strains, and specialized lymphoid tissue associated to the intestine (Figure 2) (14, 15). The epithelium is covered with a mucus barrier that protects it against direct contact with most of the bacteria, both in the stomach and the intestine. This mucus contains a glycoprotein called mucin, which has viscous and glutinous properties to trap microorganisms that are then expelled with the aid of the peristaltic movement in the intestine (11, 15). Besides mucin, the intestinal mucosa contains proteins secreted by Paneth cells, including β-defensins, lysozyme, cathelicidins, and phospholipase A2, which act as antimicrobial agents.
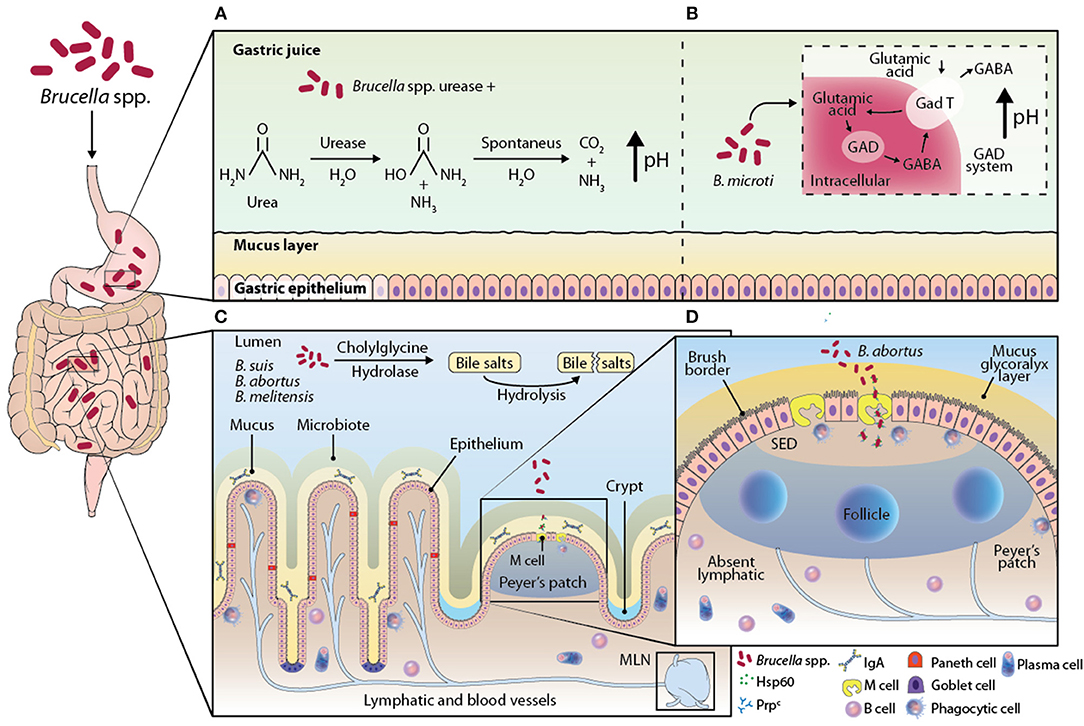
Figure 2. Illustration of the proposed models of the interaction of Brucella strains with GALT. When Brucella reaches the stomach, it encounters the main chemical barrier, the pH of gastric juices. Some Brucella spp. possess the enzyme urease, which is able to catalyze the hydrolysis of urea into carbamate and ammonia; carbamate is degraded by spontaneous hydrolysis into ammonia and carbon dioxide, creating a basic environment in the environment by increasing the pH (A). It has been described in B. microti the GAD system that consists of transporting glutamic acid into the bacterium and with the help of the enzyme glutamate decarboxylase (GAD) convert glutamic acid into gamma amino butyric acid (GABA), this reaction consumes a proton that alkalinizes the extracellular medium of the bacterium, which would help B. microti survive in an extremely acidic environment such as the pH of the stomach (B). The bile salts in the small intestine have an antimicrobial role, because they are capable of damaging the membrane of some bacteria, the strains B. suis, B. abortus, and B. melitensis possess the enzyme cholylglycine hydrolase, an enzyme that catalyzes the hydrolysis of bile salts, inhibiting their microbicidal activity (C). In addition to the chemical barriers present in the gastrointestinal tract, there is the most important physical barrier, the monolayer of epithelial cells of the intestine. It has not been fully clarified how Brucella crosses this barrier, however it has been proposed that it is through M cells, specialized enterocytes that are found above the sub-epithelial dome (SED), in the epithelium region associated with the follicle. The M cells on its apical side express highly the cellular prion protein (Prpc), this protein is a receptor and one of its ligands is the Hsp60 protein, secreted by B. abortus through its type IV secretion system, which suggests that the interaction of the receptor and its ligand promotes the entry of the bacteria into the intestine (D).
On the other hand, bile salts released in the small intestine create conjugates with detergent action that damage the bacterial wall and prevent microbiota growth (11, 16). Finally, in the intestinal mucus are immunoglobulins with the capacity to neutralize antigens from intestinal lumen, mostly lgA. Cells from the intestinal epithelium are joined between them by a variety of molecules. Among these intercellular interactions are close conjunctions, whose molecules prevent antigens from entering the lamina propria via the lumen (17). The main effector site of the intestinal immune response is the lamina propria; there APCs (as CD68+ macrophages and dendritic cells, DC) are located and show an immature phenotype and high phagocytic activity. Also, polymorphonuclear cells (neutrophils, mastocytes, and eosinophils) are found in the lamina propria, especially when there is an inflammatory response. The lamina propria is also populated by T and B lymphocytes that migrate from their activation site in the lymphoid tissue to the intestine (9, 11).
The GALT is constituted by PP, isolated lymphoid follicles, and MLN. These are considered the main sites inducing the adaptive immune response since they are also the location of antigen presentation to T lymphocytes and activation of B lymphocytes, which create germinal centers (13, 17).
Given that the mouth is the natural entry for Brucella spp. the gastrointestinal tract is the most exposed tissue, which the bacterium uses to disseminate and start the infection, apparently, evading the local immune response. The pH of the stomach is useful to degrade proteins and is considered a chemical barrier that pathogens must confront during their transit (15). Some bacteria, as Helicobacter pylori, produce an enzyme with urease activity that is considered a virulence factor. This is because the enzyme hydrolyzes urea, ammonia, and carbamate, creating a basic environment in the medium. This alkaline condition promotes bacterial growth while urease byproducts provide a source of nitrogen. However, urease can compromise the integrity of the epithelial tissue (18). Urease activity could be a favorable factor to Brucella spp. persistence through the stomach. In the sequencing of Brucella spp. genome, two urease operons (ure1 and ure2) were found in its chromosome 1 (19). The only Brucella species that does not produce urease is B. ovis, which is not transmitted orally but sexually in animals. This incapacity is likely to be caused by the absence of the enzyme.
The biological function of urease and its role in the pathogenesis have been evaluated in strains manipulated to eliminate operons ure1 and ure2. The mutated strains were intragastrically inoculated to BALB/c mice. The researchers observed a lower colonization in the small intestine, as compared against the one observed in the wild strain B. abortus 2308. Still, there was no evidence that the urease-deficient strains colonize the liver or the spleen. On the other hand, in the intraperitoneal inoculation, urease-deficient strains did colonize both the spleen and the liver, which suggests that urease is a key factor, although not the only one, to secure survival through the stomach (Figure 2A) (6, 19–21).
Some bacteria such as E. coli and lactic acid bacteria (Lactococcus lactis) have a system that converts glutamic acid into gamma amino-butyric acid (GABA) inside the bacterium through the action of the enzyme glutamic acid decarboxylase (GAD) (22). This reaction consumes a proton that alkalinizes the intracellular environment of the bacterium. In addition, this system allows for a GABA molecule to be transported to the extracellular medium and a glutamic acid molecule to be internalized in the bacterial cytosol, leading to an increase in the extracellular pH. This system is primarily involved in E. coli resistance in extremely acidic environments, such as stomach pH. The GAD system is present in the B. microti strain, while the gene involved in this system is identified as gadBC. In the presence of a glutamic acid medium B. microti can grow at a pH of 2.5; however, when the gadBC gene is modified into B microti its growth in an acidic medium decrease. In vivo, the oral administration of these strains with the modified genes significantly decreases their CFU in spleen and liver when compared to the B. microti strain without the modified gadBC genes. Then, it is suggested that this system is a mechanism that Brucella can employ to resist the stomach pH (Figure 2B) (23).
The presence of gadBC genes has been identified with in silico analysis present in almost all species of Brucella; still, the functionality of the GAD system is not present in more pathogenic species such as B. abortus, B. canis, B. melitensis, B. neotomae, and B. suis (24).
Bile salts are secreted in the small intestine as conjugates with detergent properties that promote dispersion and enzymatic degradation of fats. This property also has an effect on the membrane of some bacteria; therefore, bile salts are considered to play an antimicrobial role (16). It has been described that B. abortus, B. melitensis, and B. suis synthesize an enzyme hydrolyzing bile salts, called cholyglycine hydrolase, coded by gene chg. This enzyme is also found in intestinal microbiota bacteria as Lactobacillus and Bacteroidetes, which suggests that microbiota bacteria require this enzyme to prevent the detergent action of bile salt conjugates (25).
The function of Brucella spp. cholyglycine hydrolase has been studied in strains in which gene chg has been previously eliminated. Studies prove the relevance of this enzyme during transit in the intestine when the knock-out strain is orally administered to mice. A decrease in the chg−/− bacteria disseminated to the spleen was observed in comparison with wild bacteria that were orally administered. This study demonstrated that the viability of Brucella spp. transit in the gut depends on the integrity of cholyglycine hydrolase. This finding suggests cholyglycine is relevant as a virulence factor, aiding the bacteria to avoid the microbicide activity of bile salt conjugates (Figure 2C) (26).
Unlipidated 19 kDa outer membrane protein, U-Omp19, is a Brucella protein without lipid fraction. This modified protein is studied to be used in the development of a vaccine against brucellosis. During the studies, the sequence of this protein has been analyzed and found to be identical to the protease inhibitor proteins of other bacteria, mainly of the Inh family of Erwinia chrysanthemi. To demonstrate the inhibitory function of U-Omp, experiments were performed in vitro: U-Omp was incubated in a casein system and different proteases characteristic of the stomach (pepsin) and proteases secreted from the pancreas to the intestine (elastase, trypsin, and α quymotrypsin). The study indicates that U-Omp inhibits protease function by preventing casein degradation. The results suggest that Brucella can evade the proteases of the stomach and intestine of the host with this protein in order to invade the mucosal tissue and establish infection (27).
The monolayer of epithelial cells in the gut is an important physical barrier. The pathogens that reach the intestinal lumen, as Brucella, must cross this barrier to disseminate toward other tissues as liver and spleen. Evidence in animal models suggests that Brucella goes from the intestinal lumen into the small intestine and, from there, disseminates to other organs as spleen and liver (6).
The mechanism through which Brucella spp. is translocated from the intestinal lumen to tissue remains unclear. Still, it has been proposed that Brucella is introduced by enterocytes specialized in trapping lumen antigens, called M cells, which later deliver the antigen to nearby DC (6, 28). M cells are found in a region of PP, called follicle-associated epithelium. PP are distributed along the small intestine and their anatomy, cell distribution, and functions are similar to those of a node, except PP do not have capsule nor afferent lymphatic vessels (13, 29).
Studies in vitro demonstrated that B. melitensis can cross a monolayer of human epithelial cells differentiated to M cells but does not go through a monolayer of non-differentiated enterocytes, suggesting that M cells promote the transit of Brucella through the epithelial monolayer (6). Studies in vivo on the association between Brucella and PP were done in ligated loops, closing the ends of an intestinal section containing at least one PP where the bacterium as inoculated. These studies allowed for the observation of B. abortus-GFP by confocal microscopy within the follicle-associated epithelium of the Peyer's patches, a region that is rich in M cells (28). A more thorough understanding of the role M cells play was achieved studying the cellular prion protein (PrPC) highly expressed at the apical end of M cells. This protein is a receptor, one of its identified ligands is the heat-shock protein Hsp60, secreted by B. abortus through its type-IV secretion system. This suggests that the interaction between PrPC and Hsp60 promotes infection by macrophages, so PrPC could be a Brucella receptor (30). The observation that B. abortus inoculated in the ligated loop associated to PrPC is related to the high efficiency with which the bacterium is internalized in PP. In PrPC-deficient mice (prnp−/−), a lower amount of internalized B. abortus internalized in the ligated PP loop was observed. However, when Brucella is inoculated orally in prnp−/− mice, no PP were detected (31). These observations confirm the role of PrPC in the capture and internalization of the bacterium by M cells, which provide Brucella spp. An efficient penetration into the organism (Figure 2D).
It must be noted that strains of B. abortus, B. melitensis, B. suis, B. canis, and B. abortus RB51 adhere to epithelial cells CaCo-2 and HT29 inside which they can replicate. Infection of cell lines Caco-2 and HT29 with Brucella strains does not induce the secretion of pro-inflammatory cytokines as TNFα, IL-1β, MCP-1, IL-10, or TGF-β; however, they produce IL-8 and chemokine CCL-20 (32). The absence of inflammatory response to the infection by Brucella spp. has also been observed in histological sections of the small intestine from intragastrically inoculated mice. These sections do not show neutrophil infiltration, which would be favored by IL-8 and CCL-20 secretion (6).
Despite the mechanisms mentioned on the entry of Brucella spp. in the organism, it is still unclear how Brucella avoids each barrier of the intestinal immune system. Similarly, we have yet to clarify how and why Brucella remains in the gut and GALT (as PP and MLN) for so long. Submandibular maxillary lymph nodes (which drain oral cavity, eyes, and nasal mucosa) can be a niche or reservoir for Brucella to remain silent and for periods of up to 50 days (33). This proposal could also be considered for organs as PP or MLN in which viable Brucella has also been found to remain for extended periods. It is also important to elucidate how Brucella spp. virulence factors affect innate immune response and GALT adaptive response. Once this is understood, experts will be able to develop vaccines that protect the exposed tissue of the gastrointestinal mucosa.
Although Brucella cannot be considered as an enteropathogen, it passes through the intestine in a natural infection. Although this has long been known to be the pathway of entry for this pathogen, most studies of pathogenesis and the immune response against Brucella spp. have been conducted by inoculating the bacterium intraperitoneally. This route favors systemic infection in early post-infection times, with early and easy colonization of the bacteria in the spleen. However, as it is not the natural route of infection, it does not allow knowing the immunity mechanisms that could contain the infection before its systemic dissemination. It is possible that the intestine, an effective barrier to bacterial invaders, is a possible reservoir of Brucella spp. For this reason, it is essential to have a detailed knowledge of the mechanisms of innate and acquired immunity that develop in the intestinal mucous membrane toward the various components of Brucella spp. that allow its systemic dissemination.
Mucosal of Respiratory Tract. Other Portal of Entry of Brucella spp.
Oral exposure is the major route of infection and aerosol exposure is the most common source of occupationally acquired brucellosis caused by the manipulation of contaminated secretions of ill animals or by contact with laboratory cultures and tissue samples (34, 35). Brucella spp. can be easily aerosolized, with the additional hazard of its low infectious dose. The human infection has a variety of presentations, mainly affecting the reticuloendothelial system and complications had been reported; pulmonary involvement is sporadic (36, 37). Because of its potential spread by aerosols, Brucella has been classified as a biological agent of threat (38).
On the arrival of bacteria to the nasopharynx, the first step in pathogenesis is the adhesion to target molecules through different microbial ligands, most of them unknown so far. It has been reported an adhesin of B. abortus with an immunoglobulin-like domain. Although this protein has been involved in the adhesion to epithelial cells (39) the colonization of these bacteria in the respiratory tract has not been reported as is the case with species such as Bordetella pertussis, which usually enters and resides into the upper respiratory tract (40). In humans and other species, the inhalation of aerosols contaminated with Brucella leads to colonization of organs such as the lymph nodes, spleen, and liver (41).
The inhalation of aerosol or the intranasal instillation in mice is considered the most natural pathway of entry of Brucella, since the bacteria must trespass the first defense mechanisms (42). Epithelial barriers, mucus, defensins, and lysozyme avoid the colonization and entry to respiratory tract. It has been described that Brucella abortus PliC is a lysozyme inhibitor and it could participate in the brucellosis pathogenesis (43). Epithelial cells produce defensins, cytokines and chemokines in response to B. abortus. β-defensin 2 has antimicrobial activity against several pathogens and in vitro model β-defensin 2 does not seem to be effective against Brucella, but it could participate in recruiting immune cells and increase the dissemination of the bacteria (44).
In the nasal mucosa, secretory IgA and to a lesser extent IgG are other defense mechanisms. However, some evidences suggest that humoral immune response mediated by IgA may not play a central role in restraint against Brucella so the pathogen can even penetrate the epithelial barrier (45).
Epithelial cells and phagocytic cells are the first cells to be contacted by inhaled microorganisms. Some pathogens find temporary refuge from host immunity by sliding between epithelial cells (46). Furthermore, Brucella invades lung epithelial cells and alveolar macrophages where it avoids its degradation by inhibiting and modulating the endocytic traffic. In this way, these infected cells could promote the extrapulmonary dissemination of the bacteria within host tissues (47, 48). There is evidence that following intratracheal inoculation of Brucella in animal models, the bacteria spread to the spleen, draining cervical lymph node, tracheobronchial lymph nodes and mediastinal lymph nodes (47, 49), Therefore, immune response in mediastinal and cells homing to lungs have to be analyzed with more detail to clarify the immune responses that could mediate immune protection against mucosal infection.
Cytokines IL-1, IL-6, and TNF-α are important mediators of systemic and local inflammatory responses, but in lungs this triad do not have a key role. It has been shown that IL-1 and IL-6 deficiencies do not impact on the course of Brucella infection in the lungs, liver, or spleen. However, TCR-δ, TAP1, IL-17RA, and IFN-γR deficiencies affect Brucella control in the lungs (50). Recently, it was reported that IL-1R, probably through IL-1β action, and the NLRP3 and AIM2 inflammasomes have a protective effect on respiratory Brucella infection (51).
The induction of cellular-mediated responses is important in the clearance of Brucella from the lung, liver, and spleen, and the protection should be optimal when immunization and challenge routes are identical (45, 52). The live vaccines have shown to be effective in protection against brucellosis and these studies in animals have allowed to define the systemic immunity, but most of these studies performed parenteral vaccination and there is limited information of the mucosal response induced by the natural route of entry of Brucella (53).
Although no immunization route has shown total protection against intranasal challenge it has been found a significant reduction of bacterial load compared to the unvaccinated animals. Generally, it has been observed that intranasal immunization of Brucella induces systemic and mucosal immune responses (45, 52).
Response of T Lymphocytes to Infection by Brucella
Brucella is one of the non-commensal pathogenic microorganisms that enter the organism through the intestinal mucosa and destabilize the commensal biota. Other pathogens such as Escherichia coli (ETC, EPEC, EIEC, etc.), Shigella, Salmonella, and Yersinia (54) colonize the gut and lead to the synthesis of proinflammatory molecules as TNF-α and IL-18, which are produced by macrophages in response to the infection (55).
Brucella abortus is captured by macrophages, where it causes the inhibition of several intracellular processes, as phagosome-lysosome fusion and respiratory burst through components as lipopolysaccharide (LPS) and those of the type-IV secretion system, which are virulence factors (6, 56–58). The most relevant virulence mechanism of Brucella spp. is its ability of survival and replication within these phagocytic cells as well as its mechanisms to evade intracellular death. Brucella spp. can infect different cell strains as macrophages, monocytes, DC, epithelial cells, and B lymphocytes (3). This characteristic is shared by other bacterial species that behave as intracellular pathogens, as Mycobacterium tuberculosis and Salmonella enterica serovar Typhi (59, 60), so the mechanisms of induced immunity are partially similar.
During the severe stage of the disease, which lasts 2 weeks approximately, the bacteria multiply rapidly. In the chronic stage, lasting 5–6 weeks, the bacterial concentration stabilizes and then decreases slowly, although there are some remaining bacteria in the liver, spleen, and lymph nodes even after 5 months (57).
The lamina propria of the intestine is inhabited by a number of cell lines, including CD4+, CD8+, γδT, and activated B lymphocytes, plasmatic cells, macrophages, eosinophils, and several populations of DC that take part in the regulation of the mucosa immune response (61). Additionally, stimulated enterocytes do have the ability to produce proinflammatory cytokines IL-1, IL-6, IL-7, and IL-8 (17).
The natural invasion of the host by Brucella induces a specific immune response mediated by Th1 lymphocytes that protects against the development of the disease, similar to the one observed in infections caused by other intracellular pathogens as Salmonella and Mycobacterium tuberculosis. Th1 lymphocytes are characterized by the production of proinflammatory cytokines as IL-1β, IL-6, TNF-α, and more importantly, IFN-γ (59). These proinflammatory cytokines play a protective role since they activate macrophages to increase their bactericide capacity and cytotoxic T lymphocytes (CTL) that kill infected cells (62–65). It has been observed that IFN-γ and IL-12 production by macrophages is induced by IL-17 (66). Also, the differentiation to Th17 lymphocytes is induced by a combination of IL-1, IL-6, and the transforming growth factor β (TGF-β). The in vivo participation of IL-23 in this differentiation process is necessary to achieve a stable differentiation and proliferation of Th17 cells when activated (67).
IL-17-producing cells, found in the lamina propria, play an essential role against microorganisms infecting the gastrointestinal tract (68). IL-17 production has also been observed in the lung and oral cavity mucosa. The cells producing the cytokine migrate toward these sites due to CCR6 expression. It has been established that IL-17 response at these mucosal sites is mainly directed against extracellular bacteria and fungi (69). Its protective role in infections caused by intracellular pathogens was initially controversial. In infections caused by Listeria monocytogenes, IL-17 response is produced by γδT lymphocytes in the liver, which apparently leads to the arrival of neutrophils in the organ. Nevertheless, mice lacking IL-17 receptor (IL-17RA−/−) survive infection by L. monocytogenes (70). Contrastingly, IL-17 is necessary to generate a protective immune response against M. tuberculosis induced by vaccination.
The role of IL-17 seems to be differentially relevant with respect to Brucella species, against which protection is created. In oral vaccination of BALB/c mice with B. melitensis strain from which gene znuA (involved in zinc transportation) or B. abortus RB51 strain induced a similar protection against the intranasal challenge with B. melitensis 16M. The mice that showed IFN-γ deficiency (IFN-γ−/−) also exhibited a state of protection, albeit less significant than the one observed in BALB/c wild-type mice. The protection in both mice strains was parallel to IL-17 and IL-22 response. However, IL-17 and IL-22 production was higher in IFN-γ−/−mice than in wild type BALB/c mice, which would suggest that IL-17 and IL-22 compensate for the absence of IFN-γ. The in vivo neutralization of IL-17 with specific antibodies only affected the protection conferred by strain RB51 but did not affect the one provided by mutant ΔznuA (71). This proved that IL-17 is relevant to induce the protection provided by strain RB51 when administered orally.
Other studies have demonstrated the role that IL-17 plays in the protection against brucellosis induced by vaccination in the intestinal mucosa (72). B. abortus inhibits the host's proteases through Omp19 and limits the antigenic proteolysis of cells from PP and MLN, increasing the bacterial concentration in the gut mucosa inside monocytes and DC. However, in mice immunized with ovalbumin together with Brucella U-Omp19 as adjuvant, the anti-ovalbumin response of Th1, Th17, and CD8+ T lymphocytes was increased in the gut mucosa and system, leading to a high production of IFN-γ and IL-17 (27).
Abkar et al. (73) orally immunized BALB/c mice with protein U-Omp19 incorporated into N-trimethyl chitosan (TMC/Omp19) nanoparticles. They compared the response with that observed in mice immunized with intraperitoneal TMC/Opm19 after being challenged with virulent B. abortus strains 544 and B. melitensis 16M. The results showed that vaccination with oral TMC/Opm19 generated a protection considerably higher than the one induced by intraperitoneal vaccination. In addition, the intraperitoneal vaccination induced Th1- and Th2-type responses while the oral vaccination induced Th1 and Th17 cell response. These results demonstrate that the oral presentation of Brucella spp. antigens induce a more efficient response in which IL-17 production is deeply involved.
Lymphoid aggregates (particularly γδT lymphocytes, cryptopatches) have been identified in the lamina propria of mice. In these aggregates, Tγδ and CD8+αα lymphocytes suffer extrathymic maturation (74). Only a few studies on the role of γδT cells have been reported; however, it is well-known that they are relevant in the early stages of infection since TCRδ−/− mice are more susceptible to infection by B. abortus than wild-type C57BL/6. During the first week post infection, there was an increase in γδT cells, which abated after 2 weeks. On the other hand, IFNγ−/− mice, from which γδT cells were eliminated, showed a considerable increase in susceptibility to the infection by B. abortus (75).
In contrast, CD4+CD25+ Tregs cells are involved in the early stages of infection by Brucella, reducing the capacity of response of CD4+ effector T cells. The elimination of CD4+CD25+ Tregs cells with anti CD25 antibodies triggered an enhanced protective response against B. abortus in BALB/c mice, along with an increase in IFN-γ production compared with the response of mice without anti CDC25 treatment. In persistent infection, CD4+CD25+ Tregs cells maintain their suppressor function (76).
At the beginning of the infection by B. abortus, the number of Tregs and effector T cells was increased (76, 77). In this early stage of the infection, Tregs can negatively regulate the function of effector T cells through the production of IL-10 (78). Splenocytes can also produce IL-10 after the stimulus by PrpA protein of B. abortus and thus promoting the persistence of the infection (79). This suggests a deleterious role of the anti-Brucella immune response mediated by Treg and IL-10 production.
It is remarkable that in other infectious processes, CD4+CD25+ Treg and IL-10 protect against the infection or the disease. This has been observed in the tuberculosis murine model (80), and in infections caused by Helicobacter pylori (81), Pneumocystis carinnii (82), or trypanosomiasis (83).
It is well-established that the development of Th1 cell-mediated immune response is important in protection against Brucella spp. Particular emphasis has been placed on the role of IFN-γ in protecting against infection caused by Brucella spp. This conclusion has been drawn from studies done on animals infected by parenteral pathways, in which a systemic infection develops directly. Protective immunity in the intestine continues to be dependent on T cells, but it has become clear that other subpopulations are involved. In addition, a delicate balance must be maintained in the gut between pro-inflammatory, classically anti-Brucella, and anti-inflammatory responses. This knowledge is required for the design of vaccines that must be applied through the digestive tract to generate local immunity in the intestinal mucosa.
Antigenic Components of Brucella as Virulence Factors
Brucella spp. is an intracellular pathogen that invades and proliferates within the host's cells. The virulence of this strain is associated to the intracellular replication capacity in phagocytic and non-phagocytic cells (84). Brucella spp. lacks the typical virulence factors as toxins, fimbria, pili, and plasmids (85). Contrastingly, this intracellular pathogen has different antigenic elements that provide it with virulence and allow it to establish in the host's cells.
Lipopolysaccharide
Brucella, as genus, has two colonial morphologies: smooth and rough. In general, Brucella strains of rough morphology are attenuated or show reduced virulence, with the exception of species B. ovis and B. canis, that are virulent and naturally rough. The LPS of Brucella is one of its main virulence factors and is considered non-conventional when compared against other Gram-negative LPS, as those from the Enterobacteriaceae family. Neither B. abortus LPS nor that of B. melitensis activate complement or macrophages while their endotoxic activity is extremely reduced (86, 87).
The LPS of Brucella spp. is constituted by three main components: lipid A, core, and O side chain, which is the immunodominant component. Lipid A is inserted in the external membrane and has a backbone of diaminoglucose; acyl groups are the longest chain, C18–C19 or C29 instead of C12 to C14 of conventional LPS. The core, to which acylated chains are bound, has a polysaccharide sequence conserved among Brucella species. The bond between the acylated chain and core is through an amide linkage instead of ester and amide linkages. The O side chain of smooth LPS from Brucella is a chain of repeating units of sugars with a variability that allows for the differentiation of the species (88).
LPS from Brucella exhibits low endotoxicity due to poor macrophage recognition by heterodimer TLR-2/MD2, considered the main LPS receptor in Gram-negative bacteria that also transduces activation signals within macrophages. In consequence Brucella LPS does not induce a response in macrophages or DC in vivo or in vitro. The stimulation of macrophages or DC with Brucella spp. LPS does not induce the expression of activation markers on the cell surface nor does it induce the production of proinflammatory cytokines. In contrast, Brucella spp. LPS seems to deregulate macrophages and DC according to their function of antigen presenting cells, so it does not allow for the activation or proliferation of T lymphocytes. This low induction in the immune response mediated by cells could be the way in which Brucella evades from the beginning of the infection. It allows Brucella to establish, triggering the chronicity of the infection (89).
It has been demonstrated that the core is a virulence factor, since wadC mannosyl transferase mutants increase interaction with MD2, leading to a higher production of cytokines. On the other hand, the loss of the O-chain creates attenuated strains that allow the infected cell to carry the vesicle containing Brucella to the lysosomal compartment, where the bacterium is eliminated. Smooth strains evade from this intracellular death pathway, suggesting that the O side chain of LPS is involved in the transportation of the vesicle containing Brucella to other non-lysosomal compartments, or avoidance of transport to such lysosomal compartment (90). Brucella spp. LPS also prevents the activation of the complement, with which it also avoids opsonization by subcomponent C3b, decreasing recognition by phagocytic cells through receptors for the complement. Additionally, it prevents the generation of anaphylatoxins C3a and C5a, hampering the consequent proinflammatory response (86, 87).
Cyclic β-1,2-Glucan
Cyclic glucans are intrinsic components of Gram-negative bacteria that have a high structural variability and are responsible for maintaining homeostasis against osmolarity changes (91). Brucella spp. cyclic glucans are accumulated in the periplasm and are constituted by a backbone of glucose units bound by b-1,2 links with a polymerization degree of 17–25 monomers (92). Mutants of the genes responsible for the synthesis and transport of glucan to the periplasm show decreased survival and multiplication of the bacterium in BALB/c mice spleen. The mutant strain of the gene cgs (cyclic glucan synthase), responsible for glucan synthesis induces a response that is almost exclusive of Th1 lymphocytes (93). It has been demonstrated that the cyclic glucan of Brucella spp. acts upon the host cell membrane at the lipid rafts. Then, the bacterium controls the maturation of the vacuoles in which it is internalized, preventing lysosome fusion, and allowing to reach the replication niche (94). The cyclic β- 1,2-glucan of Brucella spp. promotes splenomegaly in mice as a consequence of monocyte, DC, and neutrophil recruitment by IL-12 and TNF-α induction (95). Studies in vitro have demonstrated that Brucella spp. induces DC activation via TLR4, MyD88 and TRIF, but not through CD14, increasing the antigen-specific response of CD8+ T lymphocytes (96).
Type IV Secretion System and BvrR/BvrS
Brucella spp. has a two-component system BvrR/BvrS. This system directly and indirectly controls the expression of operon virB, coding the type IV secretion system (T4SS) (97). The two-component BvrR/BvrS system of Brucella spp. is constituted by a histidine-kinase sensor located in the cytoplasmic membrane (BvrS) and a cytoplasmic regulator (BvrR). The mutant strains in the BvrR/BvrS system are avirulent in mice, show a lower capacity to invade macrophages and HeLa cells and are unable to replicate intracellularly (98).
The Brucella BvrR/BvrS two-component regulatory system is essential to detect changes in the phagosomal environment. In addition, it allows the bacterium to modify the lifestyle from extracellular to intracellular (99). The BvrR/BvrS system controls the conformation of the LPS structure and the expression of Omp (99–101). This system is also necessary to switch off extracellular survival genes and express the genes needed for invasion and intracellular survival (97).
The Brucella T4SS, constituted by 12 components, has been evaluated using in vitro and in vivo models. The in vitro infection of different cell lines demonstrated that the T4SS is essential to allow for the intracellular replication of the bacterium. In animal models it has been observed that T4SS is necessary for the onset of the infection (102). Recently it has been published that BvrR is a good candidate for a DNA vaccine in the murine brucellosis model, but many studies are missing (103).
Enzyme Superoxide Dismutase
The survival of Brucella within the macrophage has been associated to the synthesis of several proteins: antioxidant enzymes as KatE and Cu,Zn superoxide dismutase (Cu/Zn SOD). Superoxide dismutases (SODs) are metalloenzymes that catalyze the dismutation of superoxide ions, being an important mechanism of antioxidant defense (104). Periplasmic SODs are a key element of the defense mechanisms several pathogens must protect against the respiratory burst of the host's phagocytic cells. This is a previous survival step that allows for the later replication of the bacteria within phagocytes (105).
Brucella spp. Cu/Zn SOD is located in the periplasmic space of the cell wall. The bacterial count of B. abortus Cu/Zn SOD mutants recovered from BALB/c mice is much lower when compared to the amount recovered from wild B. abortus (106). This result demonstrated the key role that Cu/Zn SOD plays in the virulence of the bacterium.
Urease
This enzyme catabolizes the hydrolysis of urea in carbon and ammonic dioxide. Urease has been reported as a virulence factor for many pathogens, which induce direct toxicity in renal tissue and the formation of kidney stones. It allows for the transit of microorganisms as Yersinia, Klebsiella, and Helicobacter pylori to colonize the stomach and promotes the bacterium in an acid environment. Sangari et al. (21) evaluated urease operon mutants that were efficiently eliminated, evidencing that the presence of the enzyme promotes its survival and thus promotes infection.
Vaccines
The most effective strategy to prevent the spread of brucellosis in humans, besides the pasteurization of dairy products, has been the control of cattle through vaccination. Although there are effective vaccines to control brucellosis, the disease has not been eradicated from most of the countries around the world (107). The available vaccines present several disadvantages as the interference with the immune response induced by diagnosis methods; some cause abortions in pregnant animals while the immunity they generate does not protect the cattle vaccinated throughout their reproductive life (107, 108). To date, there is no vaccine for safe application in humans since it is considered that even those strains known for their stability can revert their attenuated state (they must be applied as live vaccine) and trigger the disease in the vaccinated population. Vaccination programs mostly focus on females because it has been suggested that some vaccines may damage the male reproductive system (109).
According to Ko and Splitter (84), Dorneles et al. (107), Nicoletti (110), and Schurig et al. (111) the ideal vaccine against brucellosis must have the following characteristics: (1) it must be constituted by living bacteria able to generate a potent Th1-type response; (2) it must not induce the production of antibodies that may interfere with diagnosis tests to detect infected animals, regardless of the route or dosage and age of animals; (3) it must be an attenuated strain that does not cause disease nor persistent infection in animals; (4) it must not be pathogenic for humans, preventing accidental contamination during the administration of the vaccine; (5) it must induce long-term protection with only one dose, without causing abortions even when administered to pregnant females; (6) it must not induce antibody response when applying boosters; (7) it must be stable and not revert to its virulent state in vivo or in vitro; and (8) it must be affordable for its massive application and easy to produce and administer (Table 1).

Table 1. Ideal Brucella vaccine (107).
Most of the vaccines currently used aim to prevent the disease caused by B. abortus (strains 19, RB51, 45/20, and SR82) and B. melitensis (strain Rev. 1) (108, 112). Vaccine strains for B. suis, B. ovis, and B. canis have been developed experimentally without reaching massive application. Table 2 summarize the classification of Brucella vaccines.
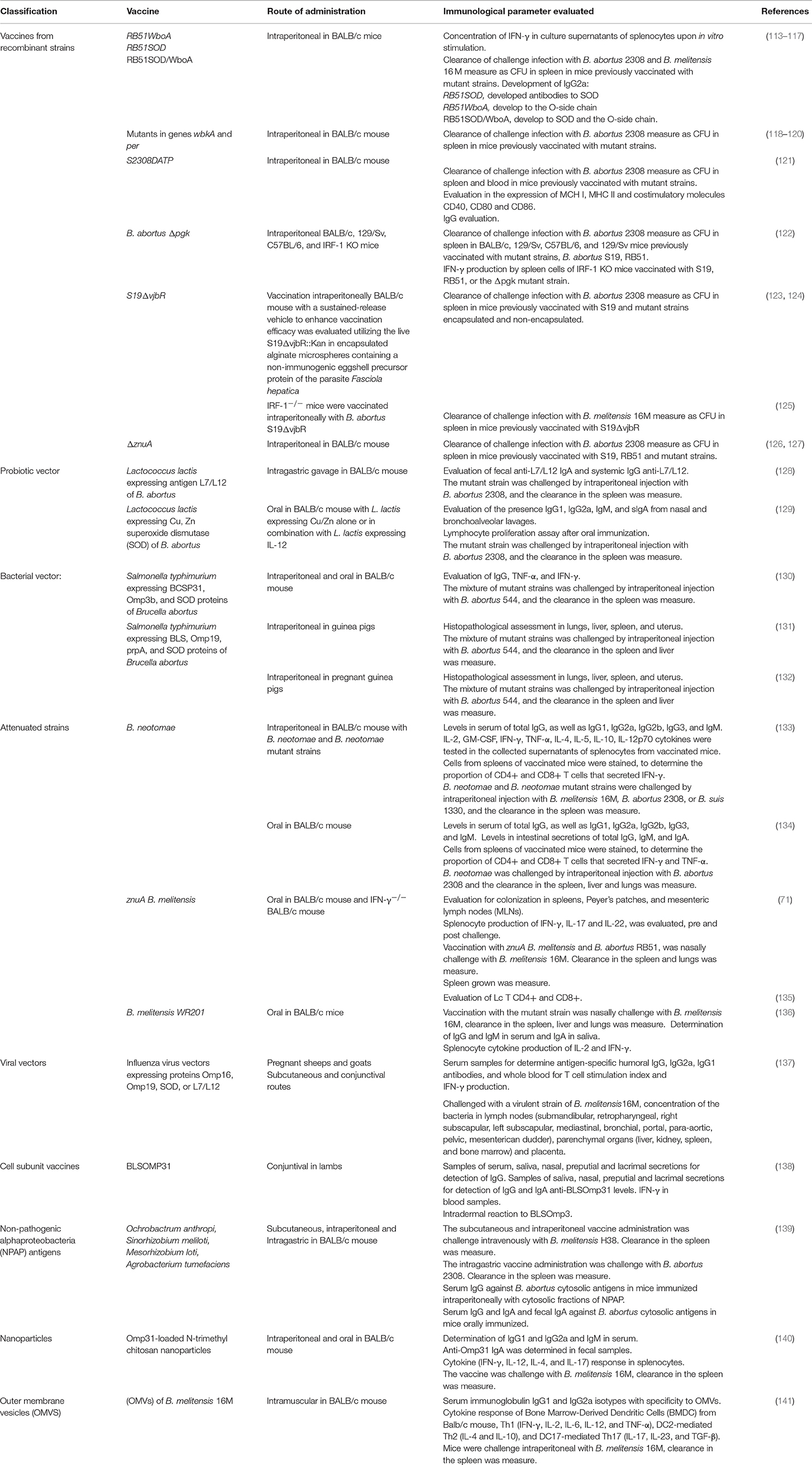
Table 2. Classification of Brucella vaccines.
Live, Attenuated Vaccines
These vaccines are given parenterally and generate immunity against systemic infection. However, as previously discussed, local mucosal immunity can precisely prevent systemic spread. It should be studied whether the use of live vaccines could maintain long-lasting immunity in the gastrointestinal and/or respiratory mucosa. Developing and maintaining local immunity will be important, as some evidence suggests that mucous membranes are reservoirs of Brucella spp.
B. abortus S19
This strain was originally isolated from milk in 1923. It was accidentally attenuated after keeping it at room temperature for a year. Its use as vaccine in cattle started in 1941 (108, 111, 142). Strain S19 is indicated for its application in calves since it causes epididymitis in male adults and is associated to abortion in pregnant females (143). Animals vaccinated with B. abortus S19 develop antibodies against LPS because the strain is smooth. Therefore, it is not possible to differentiate between animals vaccinated with this strain and naturally infected animals (108, 144). Recently, it was noticed that immune response is different among breeds of cattle, as the Shawial breed responded less to the 19S vaccine than the crossbread animals (145).
B. abortus 45/20
This strain was derived from an isolated smooth strain obtained from an infected cow (called 45) in 1922. After passages through guinea pig, strain 45 was obtained and, after 20 passages, the rough phase was isolated; therefore, the strain was named 45/20. This strain had to be administered inactivated by heating along with an adjuvant. It can revert to its virulent smooth form, so it has to be inactivated before applying it as vaccine (108, 111). Unfortunately, the protection and antibody response assays showed high variability, which questioned the efficacy of the vaccine. Additionally, some animals exhibited hypersensitivity reactions on the injection site of the vaccine. These inconveniences led to the interruption in the use of this strain as vaccine (107).
B. abortus RB51
Smooth strains commonly show phase variation into rough strains. This change in phenotype is usually accompanied by a decrease in the strain's virulence. However, some species are naturally rough, as B. ovis and B. canis. These last are pathogenic for a natural host, but the number of animal species they infect is reduced when compared against the diversity of hosts where smooth strains can be isolated. Based on these observations and the experience creating strain 45/20, experts began looking for a notably attenuated rough mutant that could colonize the host for the time needed to induce a protective immune response (111).
The strain B. abortus RB51 was obtained after successive passages of the virulent smooth strain B. abortus 2308 in a culture medium with rifampicin or penicillin. Strain RB51 shows a rough phenotype and lacks the LPS polysaccharide O-chain. It is highly stable and does not revert to the virulent smooth phenotype (142). The absence of O-chain does not induce the formation of antibodies, which allows for the differentiation between vaccinated and naturally infected animals (107). In contrast, the strain induces a strong Th1-type cell response, which confers a high protective efficiency. This effectiveness is due to the induction of IFN-γ response, which leads to the activation of cytotoxic T lymphocytes, which eliminate the infected cells, and increase the bactericide activity of macrophages (143). In 1990, the use of strain RB51 as vaccine of choice in the USA was implemented to control bovine brucellosis and is currently applied in several Latin American countries (108). The fact that it was selected as rifampicin-resistant is a considerable disadvantage since it is the first-choice antibiotic against brucellosis when streptomycin cannot be administered (146).
B. abortus SR82
Strain SR82 was developed in the former Soviet Union and was first applied as vaccine for cattle in 1988. It is recognized by the R and S phase antisera. It is as efficient as vaccine S19 but does not induce false positives in serological diagnosis tests because the presence of antibodies against R phase can be evaluated (107, 147).
B. melitensis Rev1
Strain Rev1 is smooth, behaves as attenuated strain, and is applied to goats and sheep. It was derived from B. melitensis 16M and requires the addition of streptomycin to the medium for its growth. This requirement is a disadvantage since streptomycin is the first-option antibiotic for brucellosis treatment, usually administered together with tetracycline (109, 111, 146); its administration is preferably conjunctival. This strain also induces the production of antibodies that interfere with the diagnosis: vaccinated animals cannot be differentiated from those that are naturally infected (109).
Vaccination with live strains has proven to have a highly protective efficiency in cattle and wildlife animals. This is because the bacterium remains viable in the host for a certain time and replicates. This promotes an immune response mediated by T lymphocytes against components of the pathogen, guaranteeing its effectiveness as protection-inducing agent against brucellosis (84, 148).
Mucosal Vaccines
The mucus membranes, covering the intestinal and urogenital tracts, conjunctiva, auditory canal, and exocrine gland ducts, have a highly specialized mucosal immune system that protects these surfaces against external attacks (149, 150). The design of vaccines that activate immunity in the mucus membranes seem to be promising. However, for a vaccine to be successful, it must be absorbed by the intestinal epithelium and captured by antigen-presenting cells in the intestine. It must also induce local response of B and T lymphocytes but must not create tolerance (151). Vaccines that are applied in the mucosa, particularly those that are orally administered, face dilution by mucous secretion; they may be trapped in the mucus and excluded with it or degraded by mucosal enzymes (152).
Probiotic Vectors
Several authors have explored the use of lactobacilli as vectors to generate oral vaccines. This possibility has been explored since probiotics easily survive in the gastrointestinal tract without hurting it, maintaining a close association with the epithelium. In addition, some authors have studied their immunomodulatory properties (153, 154). Because of this last property they have been applied together with vaccines for intracellular organisms. For instance, L. rhamnosus is adjuvant for vaccination with polymorphic membrane protein C (N-Pmpc) administered in the conjunctiva and increases the humoral and specific cell immune responses (155).
The following experiences have been documented when probiotics are applied together with vaccines to prevent brucellosis.
Lactococcus Lactis Expressing Antigen L7/L12 of B. abortus
L7/L12 is a ribosomal protein inducing cell-type response. Ribeiro et al. (156) transformed L. lactis using gene L7/L12. The vaccine administered orally in BALB/c mice induced significant lgA levels in feces against L7/L12, but no specific antibodies were found in serum; therefore, a systemic response was not triggered. The intraperitoneal application of the probiotic created a better protection (128).
Lactococcus Lactis Expressing Cu, Zn Superoxide Dismutase (SOD) of B. abortus
Bermúdez-Humarán et al. (157) showed that when administered in C57BL/6 mice by nasal route, L. lactis transformed to produce IL-12 in conjunction with L. lactis, and E7 antigen of the human papillomavirus type 16 (human papillomavirus type 16 E7 antigen) presented a Th1-type response induced with significant secretion of IFN-γ in splenocytes. Based on this study two recombinant strains of L. lactis, one expressing Cu/Zn SOD and another expressing IL-12, were administered to BALB/c mice orally. The animals produced significant SOD-specific slgA in nasal and bronchoalveolar lavage and T cell proliferation as a response to re-stimulation with SOD or crude Brucella protein extract. Protection against challenge with B. abortus 2308 was similar to the one observed in mice vaccinated with RB51 and it was better when administered together with the two recombinant L. lactis strains (129).
Bacterial Vectors
Salmonella typhimurium Expressing BCSP31, Omp3b, and SOD Proteins of Brucella abortus
BALB/c mice were intraperitoneally and orally immunized with the mix of S. typhimurium recombinant strains. The concentration of lgG, TNF-α, and IFN-γ in serum of mice immunized in both routes (except the recombinant one expressing Omp3b, orally administered) was significantly higher when compared against the control inoculated with wild-type S. typhimurium. Protection against challenge with intraperitoneal B. abortus 544 was higher in mice vaccinated with the mix of recombinant bacteria administered orally (130).
Salmonella typhimurium Expressing BLS, Omp19, prpA, and SOD Proteins of Brucella abortus
A vaccine using an attenuated S. typhimurium strain was designed and proved to be well-tolerated when administered orally or subcutaneously; it induces low production of TNF-α and lower cytotoxicity (158). S. typhimurium transformed to express Cu/Zn SOD, lumazine synthase (BLS), Omp-19, and proline racemase protein A (prpA) (131, 158) induces a good level of protection against challenge with B. abortus 544 when orally administered. This protection correlated with Th1 and Th17 response (72).
In addition, researchers have conducted experiments in which the vaccine additioned with B. abortus LPS is intraperitoneally administered to guinea pigs. Protection against challenge with B. abortus 544 was acceptable and did not generate histopathological damage in lungs, liver, spleen, and uterus (132, 159).
Attenuated Strains
B. neotomae
One of the options evaluated in the creation of new vaccines against brucellosis is the use of Brucella strains that are non-pathogenic for humans or cattle. One of these naturally attenuated strains is B. neotomae, a smooth strain isolated from desert mice.
Moustafa et al. (133) designed four vaccine strains from B. neotomae: B. neotomae irradiated with gamma rays, which maintains the bacterium metabolically active but prevents its replication, B. neotomae SOD, and B. neotomae Bp26 (26 kDa). The strains were intraperitoneally administered to BALB/c mice that were challenged with B. abortus 2308, B. melitensis 16M, and B. suis 1330. The highest protection was provided by the irradiated strain, which also induced the production of IFN-γ, IL-12p70, IL-5, and IL-10. Based on this result, Dabral et al. (134) orally vaccinated BALB/c mice with the irradiated B. neotomae strain and B. abortus RB51 as reference. In both groups of mice, antigen-specific CD4+ and CD8+ T cells secreting IFN-γ and TNF-α were induced. The challenge with intraperitoneal and intranasal B. abortus demonstrated that protection induced by B. neotomae was better than the one by RB51.
znuA B. melitensis
Yang et al. (126) demonstrated that B. abortus strain znuA showed a deficiency while transporting zinc but generated a good protective response. They generated the same gene deletion in B. melitensis and orally vaccinated BALB/c mice that were later challenged nasally with B. melitensis 16M. The vaccine induced Th1 response in the spleen and respiratory lymph nodes, inducing 10-fold more CD8+ T lymphocytes producing IFN-γ than CD4+ T lymphocytes. Additionally, a systemic and mucosal Th-17 response was generated (71, 135). Two weeks after the administration, no colonies of the mutant strain were detected in spleen, PP or MLN. In contrast, mice immunized with RB51 and S19 still showed the bacterium in the organs after 2 weeks. This result indicates that B. melitensis znuA is less virulent (160, 161).
B. melitensis WR201
This is a purine-auxotrophic strain harboring operon purEK deletion. The orally administered bacteria are no longer isolated from spleen, lungs, and liver 8 weeks after immunization. It induces a good protection against nasal challenge with B. melitensis 16M (136).
Viral Vectors
Influenza virus vectors expressing proteins Omp16, Omp19, SOD, or L7/L12 were administered subcutaneously and through the conjunctiva along with the adjuvant Montanide™ Gel 01 in goats and sheep simultaneously. The vaccination induced 70% protection in pregnant animals against challenge with B. melitensis 16M (137).
Cell Subunit Vaccines
BLSOMP31
Decameric Brucella lumazine synthase (BLS) has been proven to be a protein able to carry antigens without altering their conformation due to its physicochemical and immunogenic properties. In addition, it induces activation of CD8+ T lymphocyte response and IFN-γ production (162). Estein et al. (163) used BLS and added an exposed loop of Omp31 and proved that this combination induced protection against challenge with B. ovis administered parenterally to goats. The same antigen was administered at the conjunctiva to 5-month-old muttons, using a thermoresponsive and mucoadhesive gel and chitosan as carriers (BLSOmp31-P407-Ch). The antigen generated specific lgG- and lgA-specific response, increased IFN-γ in serum, and development of an intradermal reaction at 90 days post infection (138).
Alphaproteobacteria
Since it has been proven that there is a cross-reactivity between Brucella and non-pathogenic alphaproteobacteria (NPAP) antigens, Delpino et al. (26) vaccinated BALB/c mice with Ochrobactrum anthropi, Sinorhizobium meliloti, Mesorhizobium loti, Agrobacterium tumefaciens, or Brucella melitensis H38 as positive control. The best response to the intravenous challenge with B. abortus 2308 was found in mice vaccinated with O. anthropi, M. loti, and B. melitensis H38, compared against the response in mice administered PBS and that in mice vaccinated with cytosolic extracts.
Delpino et al. (26) also orally immunized using live NPAP and heat-killed B. abortus 2308, while they orally challenged using B. abortus 2308. The results showed that O. anthropi induced a strong lgA anti-Brucella response in serum and feces. Oral vaccination with NPAP induced high protection levels but reduced ones in mice vaccinated with heat-killed B. abortus 2308.
Nanoparticles
The oral administration with Omp31-loaded N-trimethyl chitosan nanoparticles in BALB/c mice induced a combination of Th1-Th17 responses, and a strong response of IFN-γ, IL-12, and IL-17. The mice challenged with B. melitensis 16M showed a significant level of protection in the spleen 4 weeks after vaccination (140).
Outer Membrane Vesicles
It has been observed that Gram-negative bacteria can secrete membrane vesicles, which carry molecular components associated to the virulence of the bacterium they are derived from Dorneles et al. (107). Vesicles of Gram-negative bacteria derive from the outer membrane. The proteomic analysis of the outer membrane vesicles (OMVs) of B. melitensis 16M proved that they contain, among others, Omp16, Omp19, Omp25, Omp31, Cu/Zn SOD, IalB, InvB GroES, and bp26 (141, 164). OMVs are recognized by membrane and cytoplasmic receptors as NOD receptors (nucleotide binding oligomerization domain-like receptors) in eukaryotic cells and induce immune response. OMVs obtained from strain VTRM1 induced IL-12, TNF-α, and IFN-γ response significantly higher than that induced by strain 16M in DC derived from bone marrow. OMVs obtained from B. melitensis 16M were intraperitoneally applied as vaccine to BALB/c mice and induced a slightly reduced protection level against challenge with 16M, when compared with the challenge with B. melitensis Rev1 (2.64 vs. 1.9 logarithmic units). Interestingly, OMVs obtained from mutant rough B. melitensis VTRM1 conferred a much higher protection level (3.08 logarithmic units) (141). These results are promising since they suggest the possibility of obtaining an acellular vaccine to be applied in humans. However, its efficacy must be proven orally as an edible vaccine to induce local immunity, which must stop infection at the entry site.
Concluding Remarks
The studies discussed in this review indicate current research on interactions between Brucella and its host mucosa. Brucella spp. is a pathogen that enters its host preferably through the mucosa of the gastrointestinal tract. Infection is eventually reported through the respiratory tract, either in the laboratory by inhalation of contaminated aerosols, or in the field by inhalation of dust particles including bacteria. Systemic immunity is well-known, since most experimental studies administer the bacteria intraperitoneally, where it establishes interactions with cellular populations different from those found in mucous membranes. As discussed, some studies suggest that in the intestine Brucella spp. interacts with dendritic cells, and that these represent a variety of populations with antagonistic functions. It has been observed that an important population of dendritic cells induces tolerogenic responses, so the role of dendritic cells with proinflammatory function should be studied.
In systemic infection the proinflammatory response is fundamental to establish the anti Brucella immunity, as it activates Th1 cells, which through the IFN-g stimulate the bactericidal activity of macrophages. In the mucosa a reaction must be subject to strong control as it can be deleterious. The role of cytotoxic T lymphocytes should be explored in more detail, as in systemic immunity studies they are essential to eliminate cells infected with Brucella spp. Preliminary results suggest their activation in the gut, but the role of Th17 cells in the gut as well as in the oral cavity and gut should be confirmed. It is possible that these cellular populations have a critical participation in these sites in view of the evidence suggesting that mucous membranes are reservoir sites for Brucella spp. which could be an important basis for the chronicity of Brucella infection. The study in some of these areas is growing exponentially and may translate into new vaccines and therapies. The new vaccines are involving the use of compounds derived from specific bacteria or bacteria as probiotics that protect mucosal surfaces and induce immunity.
New vaccines should increase protection against brucellosis, an intracellular bacterial infection that is difficult to treat. Much remains to be learned from the study of the interaction between Brucella and his host. The knowledge gained from these studies should also be translated into better therapeutic treatments to protect against this important chronic infectious disease.
Author Contributions
RL-S contributed with the T-cell immunity and compilation of the paper's chapters. AS-A contributed with the chapter related to gut immunity. LD contributed with the chapter related to Brucella vaccines. SB-U contributed with the chapter related to respiratory tract immunity. MM-L contributed with the whole organization, the critical review and edition.
Funding
This work was partially supported by the Instituto Politécnico Nacional grants 20195796 and 20181218. RL-S and MM-L are COFAA and EDI fellows. RL-S, SB-U, and MM-L are SNI fellows.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Wyatt HV. Lessons from the history of brucellosis. Rev Sci Tech. (2013) 32:17–25. doi: 10.20506/rst.32.1.2181
2. Wyatt HV. How themistocles zammit found malta fever (brucellosis) to be transmitted by the milk of goats. J R Soc Med. (2005) 98:451–4. doi: 10.1177/014107680509801009
3. Gorvel JP, Moreno E. Brucella intracellular life: from invasion to intracellular replication. Vet Microbiol. (2002) 90:281–97. doi: 10.1016/S0378-1135(02)00214-6
4. Gorvel JP, Moreno E, Moriyó N. Is Brucella an enteric pathogen? Nat Rev Microbiol. (2009) 7:250. doi: 10.1038/nrmicro2012-c1
5. Tsolis RM, Young GM, Solnick JV, Baumler AJ. From bench to bedside: stealth of enteroinvasive pathogens. Nat Rev Microbiol. (2008) 6:883–92. doi: 10.1038/nrmicro2012
6. Paixao TA, Roux CM, den Hartigh AB, Sankaran-Walters S, Dandekar S, Santos RL, et al. Establishment of systemic Brucella melitensis infection through the digestive tract requires urease, the type IV secretion system, and lipopolysaccharide O antigen. Infect Immun. (2009) 77:4197–208. doi: 10.1128/IAI.00417-09
7. Feller L, Altini M, Khammissa RA, Chandran R, Bouckaert M, Lemmer J. Oral mucosal immunity. Oral Surg Oral Med Oral Pathol Oral Radiol. (2013) 116:576–83. doi: 10.1016/j.oooo.2013.07.013
8. Meyle J, Dommisch H, Groeger S, Giacaman RA, Costalonga M, Herzberg M. The innate host response in caries and periodontitis. J Clin Periodontol. (2017) 44:1215–25. doi: 10.1111/jcpe.12781
10. von Bargen K, Gagnaire A, Arce-Gorvel V, de Bovis B, Baudimont F, Chasson L, et al. Cervical lymph nodes as a selective niche for Brucella during oral infections. PLoS ONE. (2014) 10:e0121790. doi: 10.1371/journal.pone.0121790
11. Acheson D, Luccioli S. Microbial-gut interactions in health and disease. Mucosal immune responses. Best Pract Res Clin Gastroenterol. (2004) 18:387–404. doi: 10.1016/j.bpg.2003.11.002
12. Forchielli ML, Walker WA. The effect of protective nutrients on mucosal defense in the immature intestine. Acta Paediatr Suppl. (2005) 94:74–83. doi: 10.1080/08035320510043592
13. Forchielli M, Walker W. The role of gut-associated lymphoid tissues and mucosal defence. Br J Nutr. (2005) 93:S41–8. doi: 10.1079/BJN20041356
14. Jeon MK, Klaus C, Kaemmerer E, Gassler N. Intestinal barrier: molecular pathways and modifiers. World J Gastrointest Pathophysiol. (2013) 4:94–9. doi: 10.4291/wjgp.v4.i4.94
15. Schenk M, Mueller C. The mucosal immune system at the gastrointestinal barrier. Best Pract Res Clin Gastroenterol. (2008) 22:91–409. doi: 10.1016/j.bpg.2007.11.002
16. Hofmann AF, Eckman L. How bile acids confer gut mucosal protection against bacteria. Proc Natl Acad Sci USA. (2006) 103:4333–4. doi: 10.1073/pnas.0600780103
17. Mowat AM, Agace WW. Regional specialization within the intestinal immune system. Nat Rev Immunol. (2014) 14:667–85. doi: 10.1038/nri3738
18. Smoot DT, Mobley HLT, Chippendale GR, Lewison JF, Resau JH. Helicobacter pylori urease activity is toxic to human gastric epithelial cells. Infect Immun. (1990) 58:1992–4.
19. Sangari FJ, Seoane A, Cruz-Rodríguez M, Agüero J, García-Lobo JM. Characterization of the urease operon of Brucella abortus and assessment of its role in virulence of the bacterium. Infect Immun. (2007) 75:774–80. doi: 10.1128/IAI.01244-06
20. Bandara AB, Contreras A, Contreras-Rodríguez A, Martins AM, Dobrean V, Poff-Reichow S, et al. Brucella suis urease encoded by ure1 but not ure2 is necessary for intestinal infection of BALB/c mice. BMC Microbiol. (2007) 7:57. doi: 10.1186/1471-2180-7-57
21. Sangari FJ, Cayon AM, Seoane A, García-Lobo JM. Brucella abortus ure2 region contains an acid-activated urea transporter and a nickel transport system. BMC Microbiol. (2010) 10:107. doi: 10.1186/1471-2180-10-107
22. De Biase D, Tramonti A, Bossa F, Visca P. The response to stationary-phase stress conditions in Escherichia coli: role and regulation of the glutamic acid decarboxylase system. Mol Microbiol. (1999) 32:1198–211.
23. Occhialini A, Jimenez de Bagues MP, Saadeh B, Bastianelli D, Hanna N, De Biase D, et al. The glutamic acid decarboxylase system of the new species Brucella microti contributes to its acid resistance and to oral infection of mice. J Infect Dis. (2012) 206:1424–32. doi: 10.1093/infdis/jis522
24. Damiano MA, Bastianelli D, Al Dahouk S, Kohler S, Cloeckaert A, De Biase D, et al. Glutamate decarboxylase-dependent acid resistance in Brucella spp.: distribution and contribution to fitness under extremely acidic conditions. Appl Environ Microbiol. (2015) 81:578–86. doi: 10.1128/AEM.02928-14
25. Tanaka H, Hashiba H, Kok J, Mierau I. Bile salt hydrolase of Bifidobacterium longum—Biochemical and genetic characterization. Appl Environ Microbiol. (2000) 66:2502–12. doi: 10.1128/AEM.66.6.2502-2512.2000
26. Delpino MV, Marchesini MI, Estein SM, Comerci DJ, Cassataro J, Fossati CA, et al. A bile salt hydrolase of Brucella abortus contributes to the establishment of a successful infection through the oral route in mice. Infect Immun. (2007) 75:299–305. doi: 10.1128/IAI.00952-06
27. Ibañez AE, Coria LM, Carabajal MV, Delpino MV, Risso GS, González-Cobiello P, et al. A bacterial protease inhibitor protects antigens delivered in oral vaccines from digestion while triggering specific mucosal immune responses. J Control Release. (2015) 220:18–28. doi: 10.1016/j.jconrel.2015.10.011
28. Salcedo SP, Marchesini MI, Lelouard H, Fugier E, Jolly G, Balor S, et al. Brucella control of dendritic cells maturation is dependent on the TIR-containing protein Btp1. PLoS Pathogen. (2008) 4:e21. doi: 10.1371/journal.ppat.0040021
29. Man AL, Prieto-García ME, Nicoletti C. Improving M cell mediated transport across mucosal barriers: do certain bacteria hold the keys? Immunology. (2004) 113:15–22. doi: 10.1111/j.1365-2567.2004.01964.x
30. Watarai M, Kim S, Erdenebaatar J, Makino SI, Horiuchi M, Shirahata T, et al. Cellular prion protein promotes Brucella infection into macrophages. J Exp Med. (2003) 198:5–17. doi: 10.1084/jem.20021980
31. Nakato G, Hase K, Suzuki M, Kimura M, Ato M, Hanazato M, et al. Cutting edge: Brucella abortus exploits a cellular prion protein on intestinal M cells as an invasive receptor. J Immunol. (2012) 189:540–1544. doi: 10.4049/jimmunol.1103332
32. Ferrero MC, Fossati CA, Rumbo M, Baldi PC. Brucella invasion of human intestinal epithelial cells elicits a weak proinflammatory response but a significant CCL20 secretion. FEMS Immunol Med Microbiol. (2012) 66:45–57. doi: 10.1111/j.1574-695X.2012.00985.x
33. von Bargen K, Gorvel JP, Salcedo SP. Internal affairs: investigating the Brucella intracellular lifestyle. FEMS Microbiol Rev. (2012) 36:533–62. doi: 10.1111/j.1574-6976.2012.00334.x
35. Traxler RM, Lehman MW, Bosserman EA, Guerra MA, Smith TL. A literature review of laboratory-acquired brucellosis. J Clin Microbiol. (2013) 51:3055–62. doi: 10.1128/JCM.00135-13
36. Solera J, Solís García del Pozo J. Treatment of pulmonary brucellosis: a systematic review. Exp Rev Anti Infect Ther. (2017) 15:33–42. doi: 10.1080/14787210.2017.1254042
37. Uluğ M, Can-Uluğ N. Pulmonary involvement in brucellosis. Can J Infect Dis Med Microbiol. (2012) 23:e13–5. doi: 10.1155/2012/164892
38. Pappas G, Panagopoulou P, Christou L, Akritidis N. Brucella as a biological weapon. Cell Mol Life Sci. (2006) 63:2229–36. doi: 10.1007/s00018-006-6311-4
39. Czibener C, Merwaiss F, Guaimas F, Del Giudice MG, Serantes DAR, Spera JM, et al. BigA is a novel adhesin of Brucella that mediates adhesion to epithelial cells. Cell Microbiol. (2016) 18:500–13. doi: 10.1111/cmi.12526
40. Massillo C, Gorgojo J, Lamberti Y, Rodriguez ME. Bordetella pertussis entry into respiratory epithelial cells and intracellular survival. Path Dis. (2013) 69:194–204. doi: 10.1111/2049-632X.12072
41. Henning LN, Gillum KT, Fisher DA, Barnewall RE, Krile RT, Anderson MS, et al. The pathophysiology of inhalational brucellosis in BALB/c mice. Sci Rep. (2012) 2:495–495. doi: 10.1038/srep00495
42. Smither SJ, Perkins SD, Davies C, Stagg AJ, Nelson M, Atkins HS. Development and characterization of mouse Models of infection with aerosolized Brucella melitensis and Brucella suis. Clin Vaccine Immunol. (2009) 16:779–83. doi: 10.1128/CVI.00029-09
43. Um SH, Kim JS, Kim K, Kim N, Cho HS, Ha NC. Structural basis for the inhibition of human Lysozyme by PliC from Brucella abortus. Biochemistry. (2013) 52:9385–93. doi: 10.1021/bi401241c
44. Hielpos MS, Ferrero MC, Fernández AG, Bonetto J, Giambartolomei GH, Fossati CA, et al. CCL20 and beta-Defensin 2 production by human lung epithelial cells and macrophages in response to Brucella abortus infection. PLoS ONE. (2015) 10:e0140408–e0140408. doi: 10.1371/journal.pone.0140408
45. Surendran N, Sriranganathan N, Lawler H, Boyle SM, Hiltbold EM, Heid B, et al. Efficacy of vaccination strategies against intranasal challenge with Brucella abortus in BALB/c mice. Vaccine. (2011) 29:2749–55. doi: 10.1016/j.vaccine.2011.01.090
46. Virji M. Bacterial adherence and tropism in the human respiratory tract. In: Nataro JP, Cohen PS, Mobley HLT, Weiser JN, editors. Colonization of Mucosal Surfaces. Washington, DC: ASM Press (2005) p. 97–116.
47. Archambaud C, Salcedo SP, Lelouard H, Devilard E, de Bovis B, Van Rooijen N, et al. Contrasting roles of macrophages and dendritic cells in controlling initial pulmonary Brucella infection. Eur J Immunol. (2010) 40:3458–71. doi: 10.1002/eji.201040497
48. Ferrero MC, Fossati CA, Baldi PC. Smooth Brucella strains invade and replicate in human lung epithelial cells without inducing cell death. Microbes Infect. (2009) 11:476–83. doi: 10.1016/j.micinf.2009.01.010
49. Hensel ME, Garcia-Gonzalez DG, Chaki SP, Samuel J, Arenas-Gamboa AM. Characterization of an intratracheal aerosol challenge model of Brucella melitensis in guinea pigs. PLoS ONE. (2019) 14:e0212457. doi: 10.1371/journal.pone.0212457
50. Hanot Mambres D, Machelart A, Potemberg G, De Trez C, Ryffel B, Letesson J-J, et al. Identification of immune effectors essential to the control of primary and secondary intranasal infection with Brucella melitensis in mice. J Immunol. (2016) 196:3780–93. doi: 10.4049/jimmunol.1502265
51. Hielpos MS, Fernández AG, Falivene J, Alonso Paiva IM, Muñoz González F, Ferrero MC, et al. IL-1R and inflammasomes mediate early pulmonary protective mechanisms in respiratory Brucella abortus infection. Front Cell Infect Microbiol. (2018) 8:391. doi: 10.3389/fcimb.2018.00391
52. Surendran N, Sriranganathan N, Boyle SM, Hiltbold EM, Tenpenny N, Walker M, et al. Protection to respiratory challenge of Brucella abortus strain 2308 in the lung. Vaccine. (2013) 31:4103–10. doi: 10.1016/j.vaccine.2013.06.078
53. Lalsiamthara J, Lee JH. Development and trial of vaccines against Brucella. J Vet Sci. (2017) 18:281–90. doi: 10.4142/jvs.2017.18.S1.281
54. Hooper LV. Bacterial contributions to mammalian gut development. Trends Microbiol. (2004) 12:129–34. doi: 10.1016/j.tim.2004.01.001
55. Sansonetti PJ. War and peace at mucosal surfaces. Nat Rev Immunol. (2004) 4:953–64. doi: 10.1038/nri1499
56. Billard E, Cazeivielle C, Dornand J, Gross A. High susceptibility of human dendritic cells to invasion by the intracellular pathogens Brucella suis, B. abortus and B. melitensis. Infect. Immun. (2005) 73:8418–24. doi: 10.1128/IAI.73.12.8418-8424.2005
57. Hong PC, Tsolis RM, Ficht TA. Identification of genes required for chronic persistence of Brucella abortus in mice. Infect Immun. (2000) 68:4102–7. doi: 10.1128/IAI.68.7.4102-4107.2000
58. Roux CM, Rolan HG, Santos RL, Beremand PD, Thomas TL, Adams LG, et al. Brucella requires a functional Type IV secretion system to elicit innate immune responses in mice. Cell Microbiol. (2007) 9:1851–69. doi: 10.1111/j.1462-5822.2007.00922.x
59. Byndloss MX, Tsolis RM. Brucella spp. virulence factors and immunity. Annu Rev Anim Biosci. (2016) 4:111–27. doi: 10.1146/annurev-animal-021815-111326
60. Byndloss MX, Rivera-Chavez F, Tsolis RM, Baumler AJ. How bacterial pathogens use type III and type IV secretion systems to facilitate their transmission. Curr Op Microbiol. (2017) 35:1–7. doi: 10.1016/j.mib.2016.08.007
61. Kato T, Owen RL. Absence of EGF receptors and of EGF uptake in Peyer's patch dome epithelium. Adv Exp Med Biol. (1994) 355:295–301. doi: 10.1007/978-1-4615-2492-2_50
62. Brandao AP, Oliveira FS, Carvalho NB, Vieira LQ, Azevedo V, Macedo GC, et al. Host susceptibility to Brucella abortus infection is more pronounced in IFN-γ knockout than IL-12/β2-microglobulin double deficient mice. Clin Dev Immunol. (2012) 2012:589494. doi: 10.1155/2012/589494
63. Murphy EA, Sathiyaseelan J, Parent MA, Zou B, Baldwin CL. Interferon gamma is crucial for surviving a Brucella abortus infection in both resistant C57BL/6 and susceptible BALB/ mice. Immunology. (2001) 103:511–8. doi: 10.1046/j.1365-2567.2001.01258.x
64. Zhan Y, Cheers C. Endogenous interleukin-12 is involved in resistance to Brucella abortus infection. Infect Immun. (1995) 63:1387–90.
65. Zhan Y, Cheers C. Control of IL-12 and IFN-gamma production in response to live or dead bacteria by TNF and other factors. J Immunol. (1998) 161:1447–53.
66. Onishi RM, Gaffen SL. Interleukin-17 and its target genes: mechanisms of interleukin-17 function in disease. Immunology. (2010) 129:311–21. doi: 10.1111/j.1365-2567.2009.03240.x
67. Korn T, Bettelli E, Oukka M, Kuchroo VK. IL-17 and Th17 cells. Annu Rev Immunol. (2009) 27:485–517. doi: 10.1146/annurev.immunol.021908.132710
68. Marks BR, Craft J. Barrier immunity and IL-17. Semin Immunol. (2009) 21:164–71. doi: 10.1016/j.smim.2009.03.001
69. O'Quinn DB, Palmer MT, Lee YK, Weaver CT. Emergence of the Th17 pathway and its role in host defense. Adv Immunol. (2008) 99:115–63. doi: 10.1016/S0065-2776(08)00605-6
70. Hamada S, Umemura M, Shiono T, Tanaka K, Yahagi A, Begum MD, et al. IL-17A produced by gamma delta T cells plays a critical role in innate immunity against Listeria monocytogenes infection in the liver. J Immunol. (2008) 181:3456–63. doi: 10.4049/jimmunol.181.5.3456
71. Clapp B, Skyberg JA, Yang X, Thornburg T, Walters N, Pascual DW. Protective live oral brucellosis vaccines stimulate Th1 and Th17 cell responses. Infect Immun. (2011) 79:4165–74. doi: 10.1128/IAI.05080-11
72. Pasquevich KA, Ibanez AE, Coria LM, Garcia-Samartino C, Estein S, Zwerdling M, et al. An oral vaccine based on U-Omp19 induces protection against B. abortus mucosal challenge by inducing an adaptive IL-17 immune response in mice. PLoS ONE. (2011) 6:e16203. doi: 10.1371/journal.pone.0016203
73. Abkar M, Lotfi AS, Amani J, Eskandari K, Ramandi MF, Salimian J, et al. Survey of Omp19 immunogenicity against Brucella abortus and Brucella melitensis: influence of nanoparticulation versus traditional immunization. Vet Res Comm. (2015) 39:217–28. doi: 10.1007/s11259-015-9645-2
74. Kanamori Y, Ishimaru K, Nanno M, Maki K, Ikuta K, Nariuchi H, et al. Identification of novel lymphoid tissues in murine intestinal mucosa where clusters of c-kit+IL-7R+Thy1+ lympho-hemopoietic progenitors develop. J Exp Med. (1996) 184:1449–59. doi: 10.1084/jem.184.4.1449
75. Skyberg JA, Thornburg T, Rollins M, Huarte E, Jutila MA, Pascual DW. Murine and bovine γδ T cells enhance innate immunity against Brucella abortus infections. PLoS ONE. (2011) 6:e21978. doi: 10.1371/journal.pone.0021978
76. Pasquali P, Thornton AM, Vendetti S, Pistoia C, Petrucci P, Tarantino M, et al. CD4+CD25+ T regulatory cells limit effector T cells and favor the progression of brucellosis in BALB/c mice. Microbes Infect. (2010) 12:3–10. doi: 10.1016/j.micinf.2009.09.005
77. Belkaid Y, Rouse BT. Natural regulatory T cells in infectious disease. Nat Immunol. (2005) 6:353–60. doi: 10.1038/ni1181
78. Xavier MN, Winter MG, Spees AM, Nguyen K, Atluri VL, Silva TM, et al. CD4+ T cell-derived IL-10 promotes Brucella abortus persistence via modulation of macrophage function. PLoS Pathog. (2013) 9:e1003454. doi: 10.1371/journal.ppat.1003454
79. Spera JM, Ugalde JE, Mucci J, Comerci DJ, Ugalde RA. A B lymphocyte mitogen is a Brucella abortus virulence factor required for persistent infection. Proc Natl Acad Sci USA. (2006) 103:16514–9. doi: 10.1073/pnas.0603362103
80. Kursar M, Koch M, Mittrucker HW, Nouailles G, Bonhagen K, Kamradt T, et al. Regulatory T cells prevent efficient clearance of Mycobacterium tuberculosis. J Immunol. (2007) 178:2661–5. doi: 10.4049/jimmunol.178.5.2661
81. Raghavan S, Fredriksson M, Svennerholm AM, Holmgren J, Suri-Payer E. Absence of CD4+CD25+ regulatory T cells is associated with a loss of regulation leading to increased pathology in Helicobacter pylori-infected mice. Clin Exp Immunol. (2003) 132:393–400. doi: 10.1046/j.1365-2249.2003.02177.x
82. Hori S, Carvalho TL, Demengeot J. CD25+ CD4+ regulatory T cells suppress CD4+ T cell-mediated pulmonary hyperinflammation driven by Pneumocystis carinii in immunodeficient mice. Eur J Immunol. (2002) 32:1282–91. doi: 10.1002/1521-4141(200205)32:5<1282::AID-IMMU1282>3.0.CO;2-#
83. Guilliams M, Oldenhove G, Noel W, Herin M, Brys L, Loi P, et al. African trypanosomiasis: naturally occurring regulatory T cells favor trypanotolerance by limiting pathology associated with sustained type 1 inflammation. J Immunol. (2007) 179:2748–57. doi: 10.4049/jimmunol.179.5.2748
84. Ko J, Splitter GA. Molecular host-pathogen interaction in brucellosis: current understanding and future approaches to vaccine development for mice and humans. Clin Microbiol Rev. (2003) 16:65–78. doi: 10.1128/CMR.16.1.65-78.2003
85. Moreno E, Moriyon I. Brucella melitensis: a nasty bug with hidden credentials for virulence. Proc Natl Acad Sci USA. (2002) 99:1–3. doi: 10.1073/pnas.022622699
86. Barquero-Calvo E, Chaves-Olarte E, Weiss DS, Guzman-Verri C, Chacon-Diaz C, Rucavado A, et al. Brucella abortus uses a stealthy strategy to avoid activation of the innate immune system during the onset of infection. PLoS ONE. (2007) 2:e631. doi: 10.1371/journal.pone.0000631
87. Tumurkhuu G, Koide N, Takahashi K, Hassan F, Islam S, Ito H, et al. Characterization of biological activities of Brucella melitensis lipopolysaccharide. Microbiol Immunol. (2006) 50:421–7. doi: 10.1111/j.1348-0421.2006.tb03810.x
88. Lapaque N, Moriyon I, Moreno E, Gorvel JP. Brucella lipopolysaccharide acts as a virulence factor. Curr Op Microbiol. (2005) 8:60–6. doi: 10.1016/j.mib.2004.12.003
89. Conde-Álvarez R, Arce-Gorvel V, Iriarte M, Manček-Keber M, Barquero-Calvo E, Palacios-Chaves L, et al. The lipopolysaccharide core of Brucella abortus acts as a shield against innate immunity recognition. PLoS ONE. (2012) 8:e1002675. doi: 10.1371/journal.ppat.1002675
90. Fontana C, Conde-Alvarez R, Stahle J, Holst O, Iriarte M, Zhao Y, et al. Structural studies of lipopolysaccharide-defective mutants from Brucella melitensis identify a core oligosaccharide critical in virulence. J Biol Chem. (2016) 291:7727–41. doi: 10.1074/jbc.M115.701540
91. Breedveld MW, Miller KJ. Cyclic beta-glucans of members of the family Rhizobiaceae. Microbiol Rev. (1994) 58:145–61.
92. Iñón de Iannino N, Briones G, Tolmasky M, Ugalde RA. Molecular cloning and characterization of cgs, the Brucella abortus cyclic beta(1–2) glucan synthetase gene: genetic complementation of Rhizobium meliloti ndvB and Agrobacterium tumefaciens chvB mutants. J Bacteriol. (1998) 180:4392–400.
93. Briones G, Iñón de Ianninol N, Roset M, Vigliocco A, Silva-Paulo P, Ugalde RA. Brucella abortus cyclic b-1,2-glucan mutants have reduced virulence in mice and are defective in intracellular replication in HeLa cells. Infect Immun. (2001) 69:44528–35. doi: 10.1128/IAI.69.7.4528-4535.2001
94. Arellano-Reynoso B, Lapaque N, Salcedo S, Briones G, Ciocchini AE, Ugalde R, et al. Cyclic beta-1,2-glucan is a Brucella virulence factor required for intracellular survival. Nat Immunol. (2005) 6:618–25. doi: 10.1038/ni1202
95. Roset MS, Ibañez AE, de Souza Filho JA, Spera JM, Minatel L, Oliveira SC, et al. Brucella cyclic β-1,2-glucan plays a critical role in the induction of splenomegaly in mice. PLoS ONE. (2014) 9:e101279. doi: 10.1371/journal.pone.0101279
96. Martirosyan A, Pérez-Gutiérrez C, Banchereau R, Dutartre H, Lecine P, Dullaers M, et al. Brucella β1,2 cyclic glucan is an activator of human and mouse dendritic cells. PLoS Pathog. (2012) 8:e1002983. doi: 10.1371/journal.ppat.1002983
97. Martínez-Núñez C, Altamirano-Silva P, Alvarado-Guillén F, Moreno E, Guzman-Verri C, Chaves-Olarte E. The two-component system BvrR/BvrS regulates the expression of the type IV secretion system VirB in Brucella abortus. J Bacteriol. (2010) 192:5603–8. doi: 10.1128/JB.00567-10
98. Pizarro-Cerdá J, Meresse S, Parton RG, van der Goot G, Sola-Landa A, Lopez-Goñi I, et al. Brucella abortus transits through the autophagic pathway and replicates in the endoplasmic reticulum of nonprofessional phagocytes. Infect Immun. (1998) 66:5711–24.
99. Guzman-Verri C, Manterola L, Sola-Landa A, Parra A, Cloeckaert A, Garin J, et al. The two-component system BvrR/BvrS essential for Brucella abortus virulence regulates the expression of outer membrane proteins with counterparts in members of the Rhizobiaceae. Proc Natl Acad Sci USA. (2002) 99:12375–80. doi: 10.1073/pnas.192439399
100. Lamontagne J, Butler H, Chaves-Olarte E, Hunter J, Schirm M, Paquet C, et al. Extensive cell envelope modulation is associated with virulence in Brucella abortus. J Proteome Res. (2007) 6:1519–29. doi: 10.1021/pr060636a
101. Manterola L, Moriyon I, Moreno E, Sola-Landa A, Weiss DS, Koch MHJ, et al. The lipopolysaccharide of Brucella abortus BvrS/BvrR mutants contains lipid A modifications and has higher affinity for bactericidal cationic peptides. J Bacteriol. (2005) 187:5631–9. doi: 10.1128/JB.187.16.5631-5639.2005
102. Ke Y, Wang Y, Li W, Chen Z. Type IV secretion system of Brucella spp. and its effectors. Front Cell Infec Microbiol. (2015) 5:72. doi: 10.3389/fcimb.2015.00072
103. Chen B, Liu B, Zhao Z, Wang G. Evaluation of a DNA vaccine encoding Brucella BvrR in BALB/c mice. Mol Med Rep. (2019) 19:1302–8. doi: 10.3892/mmr.2018.9735
104. Broxton CN, Culotta VC. SOD enzymes and microbial pathogens: surviving the oxidative storm of infection. PLoS Pathog. (2016) 12:e1005295. doi: 10.1371/journal.ppat.1005295
105. Pratt AJ, DiDonato M, Shin DS, Cabelli DE, Bruns CK, Belzer CA, et al. Structural, functional, and immunogenic insights on Cu,Zn superoxide dismutase pathogenic virulence factors from Neisseria meningitidis and Brucella abortus. J Bacteriol. (2015) 197:3834–47. doi: 10.1128/JB.00343-15
106. Tatum FM, Detilleux PG, Sacks JM, Halling SM. Construction of Cu-Zn superoxide dismutase deletion mutants of Brucella abortus: analysis of survival in vitro in epithelial and phagocytic cells and in vivo in mice. Infect Immun. (1992) 60:2863–9.
107. Dorneles EMS, Sriranganathan N, Lage AP. Recent advances in Brucella abortus vaccines. Vet Res. (2015) 2015:46:76. doi: 10.1186/s13567-015-0199-7
108. Olsen SC, Stoffregen WS. Essential role of vaccines in brucellosis control and eradication programs for livestock. Exp Rev Vaccines. (2005) 4:915–28. doi: 10.1586/14760584.4.6.915
109. Avila-Calderón ED, Lopez-Merino A, Sriranganathan N, Boyle SM, Contreras-Rodríguez A. A history of the development of Brucella vaccines. Biomed Res Int. (2013) 2013:743509. doi: 10.1155/2013/743509
111. Schurig GG, Sriranganathan N, Corbel MJ. Brucellosis vaccines: past, present and future. Vet Microbiol. (2002) 90:479–96. doi: 10.1016/S0378-1135(02)00255-9
112. Pandey A, Cabello A, Akoolo L, Rice-Ficht A, Arenas-Gamboa A, McMurray D, et al. The case for live attenuated vaccines against the neglected zoonotic diseases brucellosis and bovine tuberculosis. PLoS Negl Trop Dis. (2016) 10:e0004572. doi: 10.1371/journal.pntd.0004572
113. Vemulapalli R, Contreras A, Sanakkayala N, Sriranganathan N, Boyle SM, Schurig GG. Enhanced efficacy of recombinant Brucella abortus RB51 vaccines against B. melitensis infection in mice. Vet Microbiol. (2004) 102:237–45. doi: 10.1016/j.vetmic.2004.07.001
114. Vemulapalli R, McQuiston JR, Schurig GG, Sriranganathan N, Halling SM, Boyle SM. Identification of and IS711 element interrupting the wboA gene of Brucella abortus vaccine strain RB51 and a PCR assay to distinguish strain RB51 from other Brucella species and strains. Clin Diagn Lab Immunol. (1999) 6:760–4.
115. Vemulapalli R, He Y, Cravero S, Sriranganathan N, Boyle SM, Schurig GG. Overexpression of protective antigen as a novel approach to enhance vaccine efficacy of Brucella abortus strain RB51. Infect Immun. (2000) 68:3286–9. doi: 10.1128/IAI.68.6.3286-3289.2000
116. Vemulapalli R, He Y, Buccolo LS, Boyle SM, Sriranganathan N, Schurig GG. Complementation of Brucella abortus RB51 with a functional wboA gene results in O antigen synthesis and enhanced vaccine efficacy but no change in rough phenotype and attenuation. Infect Immun. (2000) 68:3927–32. doi: 10.1128/IAI.68.7.3927-3932.2000
117. Dabral N, Burcham GN, Jain-Gupta N, Sriranganathan N, Vemulapalli R. Overexpression of wbkF gene in Brucella abortus RB51WboA leads to increased O-polysaccharide expression and enhanced vaccine efficacy against B. abortus 2308, B. melitensis 16M, and B. suis 1330 in a murine brucellosis model. PLoS ONE. (2019) 14:e0213587. doi: 10.1371/journal.pone.0213587
118. Dabral N, Jain-Gupta N, Seleem MN, Sriranganathan N, Vemulapalli R. Overexpression of Brucella putative glycosyltransferase WbkA in B. abortus RB51 leads to production of exopolysaccharide. Front Cell Infect Microbiol. (2015) 5:54. doi: 10.3389/fcimb.2015.00054
119. Monreal D, Grillo MJ, Gonzalez D, Marin CM, de Miguel MJ, Lopez-Goñi I, et al. Characterization of Brucella abortus O Polysaccharide and core lipopolysaccharide mutants and demonstration that a complete core is required for rough vaccines to be efficient against Brucella abortus and Brucella ovis in the mouse model. Infect Immun. (2003) 71:3261–71. doi: 10.1128/IAI.71.6.3261-3271.2003
120. Godfroid F, Taminiau B, Danese I, Denoel PA, Tibor A, Weynants VE, et al. Identification of the perosamine synthetase gene of Brucella melitensis 16M and involvement of lipopolysaccharide O side chain in Brucella survival in mice and in macrophages. Infect Immun. (1998) 66:5485–93.
121. Zhang M, Han X, Liu H, Tian M, Ding C, Song J, et al. Inactivation of the ABC transporter ATPase gene in Brucella abortus strain 2308 attenuated the virulence of the bacteria. Vet Microbiol. (2013) 164:322–9. doi: 10.1016/j.vetmic.2013.02.017
122. Trant CGMC, Lacerda TLS, Carvalho NB, Azevedo V, Rosinha GMS, Salcedo SP, et al. The Brucella abortus phosphoglycerate kinase mutant is highly attenuated and induces protection superior to that of vaccine strain in immunocompromised and immunocompetent mice. Infect immun. (2010) 78:2283–91. doi: 10.1128/IAI.01433-09
123. Arenas-Gamboa AM, Ficht TA, Kahl-McDonagh MM, Gomez G, Rice-Ficht AC. The Brucella abortus S19 DeltavjbR live vaccine candidate is safer than S19 and confers protection against wild-type challenge in BALB/c mice when delivered in a sustained-release vehicle. Infect Immun. (2009) 77:877–84. doi: 10.1128/IAI.01017-08
124. Khalaf OH, Chaki SP, Garcia-Gonzalez DG, Ficht TA, Arenas-Gamboa AM. NOD-scid IL2rynull mouse model is suitable to study osteoarticular brucellosis and vaccine safety. Infect Immun. (2019) 87:e00901–18. doi: 10.1128/IAI.00901-18
125. Arenas-Gamboa AM, Rice-Ficht AC, Fan Y, Kahl-McDonagh MM, Ficht TA. Extended safety and efficacy studies of the attenuated Brucella vaccine candidates 16MΔvjbR and S19ΔvjbR in the immunocompromised IRF-1−/− mouse model. Clin Vaccine Immunol. (2012) 19:249–60. doi: 10.1128/CVI.05321-11
126. Yang X, Becker T, Walters N, Pascual DW. Deletion of znuA virulence factor attenuates Brucella abortus and confers protection against wild-type challenge. Infect Immun. (2006) 74:3874–9. doi: 10.1128/IAI.01957-05
127. Yang X, Clapp B, Thornburg T, Hoffman C, Pascual DW. Vaccination with a ΔnorD ΔznuA Brucella abortus mutant confers potent protection against virulent challenge. Vaccine. (2016) 34:5290–7. doi: 10.1016/j.vaccine.2016.09.004
128. Pontes DS, Dorella FA, Ribeiro LA, Miyoshi A, Loir YL, Gruss A, et al. Induction of partial protection in mice after oral administration o flactococcus lactis producing Brucella abortus L7/L12 antigen. J Drug Target. (2003) 11:489–93. doi: 10.1080/10611860410001670035
129. Sáez D, Fernández P, Rivera A, Andrews E, Oñate A. Oral immunization of mice with recombinant Lactococcus lactis expressing Cu,Zn superoxide dismutase of Brucella abortus triggers protective immunity. Vaccine. (2012) 30:1283–90. doi: 10.1016/j.vaccine.2011.12.088
130. Kim WK, Moon JY, Kim S, Hur J. Comparison between immunization routes of live attenuated Salmonella Typhimurium strains expressing BCSP31, Omp3b, and SOD of Brucella abortus in murine model. Front Microbiol. (2016) 7:550. doi: 10.3389/fmicb.2016.00550
131. Lalsiamthara J, Lee JH. Immunization of guinea pigs with Salmonella delivered anti-Brucella formulation reduces organs bacterial load and mitigates histopathological consequences of Brucella abortus 544 challenge. Vet Immunol Immunopathol. (2018) 195:40–5. doi: 10.1016/j.vetimm.2017.11.006
132. Lalsiamthara J, Senevirathne A, Lee JH. Partial protection induced by Salmonella based Brucella vaccine candidate in pregnant guinea pigs. Vaccine. (2019) 37:899–902. doi: 10.1016/j.vaccine.2019.01.020
133. Moustafa D, Garg VK, Jain N, Sriranganathan N, Vemulapalli R. Immunization of mice with gamma-irradiated Brucella neotomae and its recombinant strains induces protection against virulent B. abortus, B melitensis, and B suis challenge. Vaccine. (2011) 29:784–94. doi: 10.1016/j.vaccine.2010.11.018
134. Dabral N, Moreno-Lafont M, Sriranganathan N, Vemulapalli R. Oral immunization of mice with gamma-irradiated Brucella neotomae induces protection against intraperitoneal and intranasal challenge with virulent B. abortus 2308. PLoS ONE. (2014) 9:e107180. doi: 10.1371/journal.pone.0107180
135. Clapp B, Yang X, Thornburg T, Walters N, Pascual DW. Nasal vaccination stimulates CD8+ T cells for potent protection against mucosal Brucella melitensis challenge. Immunol Cell Biol. (2016) 94:496–508. doi: 10.1038/icb.2016.5
136. Izadjoo MJ, Bhattacharjee AK, Paranavitana CM, Hadfield TL, Hoover DL. Oral vaccination with Brucella melitensis WR201 protects mice against intranasal challenge with virulent Brucella melitensis 16M. Infect Immun. (2004) 72:4031–9. doi: 10.1128/IAI.72.7.4031-4039.2004
137. Mailybayeva A, Yespembetov B, Ryskeldinova S, Zinina N, Sansyzbay A, Renukaradhya GJ, et al. Improved influenza viral vector-based Brucella abortus vaccine induces robust B and T-cell responses and protection against Brucella melitensis infection in pregnant sheep and goats. PLoS ONE. (2017) 12:e0186484. doi: 10.1371/journal.pone.0186484
138. Díaz AG, Quinteros DA, Gutiérrez SE, Rivero MA, Palma SD, Allemandi DA, et al. Immune response induced by conjunctival immunization with polymeric antigen BLSOmp31 using a thermoresponsive and mucoadhesive in situ gel as vaccine delivery system for prevention of ovine brucellosis. Vet Immunol Immunopathol. (2016) 178:50–6. doi: 10.1016/j.vetimm.2016.07.004
139. Delpino MV, Estein SM, Fossati CA, Baldi PC. Partial protection against Brucella infection in mice by immunization with non-pathogenic alphaproteobacteria. Clin Vaccine Immunol. (2007) 14:1296–301. doi: 10.1128/CVI.00459-06
140. Abkar M, Fasihi-Ramandi M, Kooshki H, Lotfi AS. Oral immunization of mice with Omp31-loaded N-trimethyl chitosan nanoparticles induces high protection against Brucella melitensis infection. Int J Nanomed. (2017) 12:8769–78. doi: 10.2147/IJN.S149774
141. Avila-Calderón ED, Lopez-Merino A, Jain N, Peralta H, López-Villegas EO, Sriranganathan N, et al. Characterization of outer membrane vesicles from Brucella melitensis and protection induced in mice. Clin Dev Immunol. (2012) 201:352493. doi: 10.1155/2012/352493
142. Schurig GG, Roop IIRM, Bagchu T, Boyle S, Buhrman D, Sriranganathan N. Biological properties of RB51; a stable rough strain of Brucella abortus. Vet Microbiol. (1991) 28:171–88. doi: 10.1016/0378-1135(91)90091-S
143. He Y, Vemulapalli R, Zeytun A, Schurig GG. Induction of specific cytotoxic lymphocytes in mice vaccinated with Brucella abortus RB51. Infect Immun. (2001) 69:5502–8. doi: 10.1128/IAI.69.9.5502-5508.2001
144. Simpson GJG, Mercotty T, Rouille E, Chilundo A, Letteson JJ, Godfroid J. Immunological response to Brucella abortus strain 19 vaccination of cattle in a communal area in South Africa. J S Afr Vet Assoc. (2018) 89:e1–7. doi: 10.4102/jsava.v89i0.1527
145. Kumar DR, Sivalingam J, Mishra SK, Kumar A, Vineeth MR, Chaudhuri P, et al. Differential expression of cytokines in PBMC of Bos indicus and Bos Taurus x Bos indicus cattle due to Brucella abortus S19 antigen. Anim. Biotechnol. (2019) 5:1–7. doi: 10.1080/10495398.2018.1555167
146. Moriyón I, Grillo MJ, Monreal D, González D, Marín C, López-Goñi I, et al. Rough vaccines in animal brucellosis: structural and genetic basis and present status. Vet Res. (2004) 35:1–38. doi: 10.1051/vetres:2003037
147. Ivanov AV, Salmakov KM, Olsen SC, Plumb GE. A live vaccine from Brucella abortus strain 82 for control of cattle brucellosis in the Russian Federation. Anim Health Res Rev. (2011) 12:113–21. doi: 10.1017/S1466252311000028
148. Zabalza-Baranguá A, San-Román B, Chacón-Díaz C, de Miguel MJ, Muñoz PM, Iriarte M, et al. GFP tagging of Brucella melitensis Rev1 allows the identification of vaccinated sheep. Transbound Emerg. Dis. (2019) 66:505–16. doi: 10.1111/tbed.13053
149. Holmgren J, Czerkinsky C. Mucosal immunity and vaccines. Nat Med. (2005) 11:S45–S53. doi: 10.1038/nm1213
150. Czerkinsky C, Holmgren J. Vaccines against enteric infections for the developing world. Philos Trans R Soc Lond B Biol Sci. (2015) 370:20150142. doi: 10.1098/rstb.2015.0142
151. Czerkinsky C, Holmgren J. Enteric vaccines for the developing world: a challenge for mucosal immunology. Mucosal Immunol. (2009) 2:284–7. doi: 10.1038/mi.2009.22
152. Neutra MR, Kozlowski PA. Mucosal vaccines: the promise and the challenge. Nat Rev Immunol. (2006) 6:148–58. doi: 10.1038/nri1777
153. Yu Q, Zhu L, Kang H, Yang Q. Mucosal Lactobacillus vectored vaccines. Hum Vaccin Immunother. (2013) 9:805–7. doi: 10.4161/hv.23302
154. Pouwels PH, Leer RJ, Shaw M, Heijne den Bak-Glashouwer MJ, Tielen FD, Smit E, et al. Lactic acid bacteria as antigen delivery vehicles for oral immunization purposes. Int J Food Microbiol. (1998) 41:155–67.
155. Inic-Kanada A, Stojanovic M, Marinkovic E, Becker E, Stein E, Lukic I, et al. A probiotic adjuvant Lactobacillus rhamnosus enhances specific immune responses after ocular mucosal immunization with chlamydial polymorphic membrane protein C. PLoS ONE. (2016) 11:e0157875. doi: 10.1371/journal.pone.0157875
156. Ribeiro LA, Azevedo V, Le Loir Y, Oliveira SC, Dieye Y, Piard JV, et al. Production and targeting of the Brucella abortus antigen L7/L12 in Lactococcus lactis: a first step towards food-grade live vaccines against brucellosis. Appl Environ Microbiol. (2002) 68:910–6. doi: 10.1128/AEM.68.2.910-916.2002
157. Bermúdez-Humarán LG, Langella P, Cortes-Perez NG, Gruss A, Tamez-Guerra RS, Oliveira SC, et al. Intranasal immunization with recombinant Lactococcus lactis secreting murine interleukin-12 enhances antigen-specific Th1 cytokine production. Infect Immun. (2003) 71, 1887–1896.
158. Lalsiamthara J, Senevirathne A, So MY, Lee JH. Safety implication of Salmonella based Brucella vaccine candidate in mice and in vitro human cell culture. Vaccine. (2018) 36:1837–45. doi: 10.1016/j.vaccine.2018.02.069
159. Lalsiamthara J, Lee JH. Immuno-profiles of BALB/c mice inoculated with Salmonella vector delivering B-cell mitogen hydroxyproline epimerase. Mol Immunol. (2018) 95:114–21. doi: 10.1016/j.molimm.2018.02.004
160. Wang Z, Wu Q. Research progress in live attenuated Brucela vaccine development. Curr Pharm Biotchnol. (2013) 14:887–96. doi: 10.2174/1389201014666131226123016
161. Yang X, Skyberg JA, Cao L, Clapp B, Thornburg T, Pascual DW. Progress in Brucella vaccine development. Front Biol. (2013) 8:60–7. doi: 10.1007/s11515-012-1196-0
162. Rosas G, Fragoso G, Ainciart N, Esquivel-Guadarrama F, Santana A, Bobes RJ, et al. Brucella spp. lumazine synthase: a novel adjuvant and antigen delivery system to effectively induce oral immunity. Microbes Infect. (2006) 8:1277–86. doi: 10.1016/j.micinf.2005.12.006
163. Estein SM, Fiorentino MA, Paolicchi FA, Clausse M, Manazza J, Cassataro J, et al. The polymeric antigen BLSOmp31 confers protection against Brucella ovis infection in rams. Vaccine. (2009) 27:6704–11. doi: 10.1016/j.vaccine.2009.08.097
Keywords: brucellosis, gut immunity, Brucella vaccines, intracellular infection, respiratory tract immunity
Citation: López-Santiago R, Sánchez-Argáez AB, De Alba-Núñez LG, Baltierra-Uribe SL and Moreno-Lafont MC (2019) Immune Response to Mucosal Brucella Infection. Front. Immunol. 10:1759. doi: 10.3389/fimmu.2019.01759
Received: 01 May 2019; Accepted: 11 July 2019;
Published: 20 August 2019.
Edited by:
Luis F. Garcia, University of Antioquia, ColombiaReviewed by:
Maryam Dadar, Razi Vaccine and Serum Research Institute, IranSunil Joshi, University of Miami, United States
Copyright © 2019 López-Santiago, Sánchez-Argáez, De Alba-Núñez, Baltierra-Uribe and Moreno-Lafont. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Martha Cecilia Moreno-Lafont, bW1sYWZvbnRAZ21haWwuY29t; bWxhZm9udEBpcG4ubXg=