 Jennifer Howard1
Jennifer Howard1 Irfan Zaidi1
Irfan Zaidi1 Séverine Loizon2Odile Mercereau-Puijalon3
Séverine Loizon2Odile Mercereau-Puijalon3 Julie Déchanet-Merville2
Julie Déchanet-Merville2 Maria Mamani-Matsuda2*
Maria Mamani-Matsuda2*- 1Division of Intramural Research (DIR), National Institutes of Allergy and Infectious Diseases, Bethesda, MD, United States
- 2Univ. Bordeaux, CNRS ImmunoConcEpT UMR 5164, Bordeaux, France
- 3Parasites and Insect Vectors Department, Institut Pasteur, Paris, France
Malaria is an infectious disease caused by the protozoan parasite Plasmodium sp, the most lethal being Plasmodium falciparum. Clinical malaria is associated with the asexual replication cycle of Plasmodium parasites inside the red blood cells (RBCs) and a dysregulated immune response. Although the mechanisms of immune responses to blood—or liver-stage parasites have been extensively studied, this has not led to satisfactory leads for vaccine design. Among innate immune cells responding to infection are the non-conventional gamma-delta T-cells. The Vγ9Vδ2 T-cell subset, found only in primates, is activated in response to non-peptidic phosphoantigens produced by stressed mammalian cells or by microorganisms such as Mycobacteria, E.coli, and Plasmodium. The potential protective role of Vγ9Vδ2 T-cells against infections and cancer progression is of current research interest. Vγ9Vδ2 T-cells have been shown to play a role in the early control of P. falciparum parasitemia and to influence malaria adaptive immunity via cytokine release and antigen presentation. They are activated and expanded during a primary P. falciparum infection in response to malaria phosphoantigens and their activity is modulated upon subsequent infections. Here, we review the wide range of functions by which Vγ9Vδ2 T-cells could both contribute to and protect from malaria pathology, with a particular focus on their ability to induce both innate and adaptive responses. We discuss how the multifunctional roles of these T-cells could open new perspectives on gamma-delta T-cell-based interventions to prevent or cure malaria.
Introduction
Over the last decades, the importance of a specific subset of γδ T-cells in malaria infection is becoming increasingly apparent, namely Vγ9Vδ2 T-cells. Restricted to human and non-human primates, Vγ9Vδ2 T-cells constitute a non-conventional T-cell subset activated in a non-MHC dependent manner, by phosphorylated intermediates of isoprenoid biosynthesis pathways of mammalian cells and microorganisms, known as phospho-antigens (Ph-Ag) (1). The known most potent of these, HMBPP [(E)-4-Hydroxy-3-methyl-but-2-enyl pyrophosphate] is produced by the DOX-P pathway used by several microorganisms (2) including the parasite responsible for malaria, Plasmodium spp [reviewed (3)]. Once activated, Vγ9Vδ2 T-cells expand, produce cytokines, exert cytotoxic functions, and stimulate cells such as monocytes, resulting in improved monocyte antigen presentation capabilities (4).
Despite major global effort, malaria remains a major public health concern. Nearly half of the world's population live in malaria endemic regions, the majority in sub-saharan Africa, and it is responsible for ~216 million cases and 445,000 deaths each year (5). Efforts to create an effective vaccine are hampered by lack of understanding of the parasites interactions with our immune system.
There are five species of Plasmodium that infect humans: P. falciparum, P. vivax, P. ovale, P. malariae, and P. knowelsi. P. falciparum is the most prevalent and deadly. P. falciparum, similar to other Plasmodium, is transmitted through the bite of a female Anopheles mosquito. The extracellular, liver-invasive form, the sporozoite, is injected into the skin, where it enters the blood flow and travels to the liver. Here the parasite eventually invades hepatocytes, wherein it differentiates and divides to form the extracellular form called merozoites. Merozoites are released into the blood stream and invade red blood cells (RBCs) where they progress through a 48 h life cycle before RBC rupture and merozoite release. Clinical disease manifests during this blood stage and is characterized by cyclical episodes of fever paroxysms. Severe malaria can be fatal and presents an array of severe symptoms including severe anemia, respiratory distress caused by severe metabolic acidosis, cerebral-malaria, multi-organ failure, and in pregnant women, placental malaria (6).
For over 100 years, it has been observed that partial immunity to malaria in endemic areas is only acquired after multiple disease episodes (7–9). In endemic settings, immunity is developed first to severe malaria (usually before 5 years old) then to clinical malaria (by 10–15 years old) (8, 10–12). Acquired immunity appears to be strain- and variant-specific and in endemic areas people are frequently re-infected by novel variants with novel antigen combinations. This complicates the assessment of protective immunity, however it is commonly accepted that sterile immunity is rarely reached and low parasitemia with no clinical symptoms is instead maintained (13, 14). Malaria infection causes dysregulation of immune responses, including inhibition of DC maturation and antigen presenting capacity (15–17) and expansion of atypical memory B cells, the functionality of which is not yet understood (18–20). The role of the innate immune responses, and the cellular and humoral branches of the adaptive immune response has been excellently reviewed elsewhere (11, 21–25).
Concerning γδ T-cells, much of the early in vivo work on Vγ9Vδ2 T-cell responses to P. falciparum infection was done in primary infected adult patients, usually Caucasians living in non-endemic regions, where Vγ9Vδ2 T-cells are the dominant subset of γδ T-cells. However, it has been shown that in malaria endemic regions, where the populations are exposed to numerous malaria infections and possibly chronically infected, Vδ1 T-cells are the major subset (26, 27). It is not yet known if this is a genetic peculiarity, or different microbiota and pathogen exposure early in life that drives expansion and contraction of these subsets. An in-depth discussion on the reasons for these geographical differences, and the role played by non Vγ9Vδ2 T-cells in malaria infection is beyond the scope of this review, which focuses on Vγ9Vδ2 T-cells. Vγ9Vδ2 T-cells have features associated with both innate and adaptive T-cells, and increasing evidence suggests they act as a bridge between the innate and adaptive immune systems [reviewed (28–30)]. Vγ9Vδ2 T-cells have a wide range of effector functions [reviewed (31, 30)], and it is becoming increasingly clear that during P. falciparum infection they contribute to both protection and pathology. In this review, we discuss their role as cytotoxic killer cells and their ability to initiate both innate and adaptive immune responses against P. falciparum malaria infection via cytokine release and direct antigen presentation to CD4 and CD8 T-cells.
Vγ9vδ2 t-cells are activated during malaria infection
γδ T-cells have long been observed to expand in vivo in the peripheral blood of primary infected P. falciparum malaria patients, with the major subset being Vγ9Vδ2 T-cells (32, 33). Interestingly, expansion in the peripheral blood is greatest during recovery, after acute infection has passed (34), indicating either a delay in response, or homing to tissues during acute infection. Vγ9Vδ2 T-cells were found to be increased in human spleens during infection (35, 36), a phenomenon that was confirmed in monkey models (36). The rapid expansion of Vγ9Vδ2 T-cells during infection and their homing to sites of known importance in parasite clearance indicated that Vγ9Vδ2 T-cells could play a role in the response to infection.
Our previous work has demonstrated that the bioactive molecule released by infected red blood cells (iRBC) is a Ph-Ag of the DOX-P pathway, which is released concomitantly with iRBC rupture. We also showed that presentation of parasite Ph-Ag to Vγ9Vδ2 T-cells involves BNT3A1 on non-erythrocyte bystander cells, as RBCs and iRBCs are devoid of BNT3A1 (37). In addition to HMBPP various other signals, including IL-2, IL-15 (38, 39), CD4 T-cell interaction activation (40) and CD28 co-stimulation (41), are needed for effective Vγ9Vδ2 T-cell activation, and stimulation of Vγ9Vδ2 T-cells in different cytokine milieus emphasizes different functional behaviors (42).
Cytotoxic vγ9vδ2 t-cells directly target blood stage p. falciparum
In vitro studies have built a picture of how Vγ9Vδ2 T-cells directly inhibit the erythrocyte stage life-cycle. The first studies showed that Vγ9Vδ2 T-cells targeted the iRBCs in a contact dependent manner, and suggested that merozoites were the target, as inhibition of parasite life-cycle was not seen until after parasite reinvasion (43–45). Active granulysin release by the Vγ9Vδ2 T-cells was implied in mediating parasite growth inhibition, as granulysin production correlated with life-cycle inhibition (44). Experiments with granulysin and perforin deficient Vγ9Vδ2 T-cell lines confirmed that Vγ9Vδ2 T-cell inhibition of parasites was indeed granulysin-mediated but not perforin-dependent (46). Finally, in an experiment where Vγ9Vδ2 T-cells were co-cultured with late stage iRBC and removed before rupture there was no impact on the parasite reinvasion. This showed definitively that merozoites are the target, as schizonts are not affected by granulysin release (46).
Cytokine releasing vγ9vδ2 t-cells act as a trigger for both innate and adaptive immune responses
Vγ9Vδ2 T-cells are highly interactive, and much of their impact on the course of an immune response stems from their modulation of other innate and adaptive immune cells by cytokine release and direct cell-cell interaction (30). Existing evidence indicates that Vγ9Vδ2 T-cells are implicated in impacting the scale and nature of both innate and adaptive immune responses to P. falciparum infection. A large feature of the immune response to P. falciparum infection is the production of inflammatory cytokines. In vitro studies of schizont-activated PBMCs from naïve donors, Vγ9Vδ2 T-cells have been found to produce TNFα and be the major source of IFNγ, more than NK cells or macrophages (47–50). They have also been shown to express TNFα, TGF-β, and IL-8, and occasionally IL-10, IL-2, and IL-5 (48). In ex vivo analysis of cord blood from mothers in an endemic setting who had experienced malaria during pregnancy, the Vγ9Vδ2 T-cells produced significantly more IFNγ and TNFα than those from healthy mothers, as did the peripheral Vγ9Vδ2 T-cells from the mother (51). This inflammatory cytokine production by Vγ9Vδ2 T-cells has been associated with both protection and pathogenesis.
Vaccination studies have been performed where healthy, malaria naïve, volunteers are exposed to three doses of P. falciparum (via the bite of 12–15 infected mosquitos) with the accompaniment of chloroquine treatment. This permits the parasite to mature to blood stage, when it is then swiftly killed before disease symptoms can develop. After challenge by the bites of five infected mosquitos, the inoculated volunteers remained parasite-free, indicating that they had developed a sterilizing immunity (52, 53). Vaccinated (protected) volunteers showed increased IFNγ, TNFα, and IL-2 production compared to non-vaccinated (non-protected) when PBMCs, taken pre-challenge and 1 day post-challenge, were stimulated by iRBC in vitro (52, 53). IFNγ levels were also increased in PBMCs from vaccinated volunteers taken days 9, 35, 140, and 400 post-challenge when stimulated by both iRBC and sporozoites (53). γδ T-cells were found to be the major IFNγ contributors, with αβ T-cells the next largest. The majority of responding cells were effector memory, indicating recall responses, and IFNγ-producing γδ T-cells were demonstrated to be a major contributor to parasite-specific recall responses (53). Thus, in these vaccines, IFNγ production by lymphocytes including γδ T-cells, correlated with acquired immunity to P. falciparum infection. It should be noted that Vγ9Vδ2 T-cells were not specifically measured in this study. However, as Vγ9Vδ2T-cells are the predominant subset in the periphery of malaria naïve individuals from non-malaria endemic regions, it is reasonable to assume they were the major responding γδ T-cell subset in this study.
In longitudinal studies of semi-immune children from Papua New Guinea, the in vitro response of PMBCs to iRBC was measured, and subsequent malaria incidence recorded. Increased IFNγ production by PBMCs correlated with reduced risk of future moderate and high-density P. falciparum infection. Further, though there was much donor heterogeneity, γδ T-cells were the predominant IFNγ producing cell population (54). However, a different longitudinal study of children from Papua New Guinea suggests that γδ T-cell cytokine production is involved in severe malaria. Ex vivo stimulation of PBMCs from children with either severe or uncomplicated malaria or healthy controls showed that γδ T-cells and monocytes were responsible for inflammatory cytokines associated with ‘high odds' of severe malaria (55). Several studies together have shown that Vγ9Vδ2 T-cell cytokine production is abrogated with repeat malaria exposure, and this contributes to decreasing clinical symptoms in subsequent infections.
Decreased peripheral activity of Vγ9Vδ2 T-cells has been found during the acute stage of infection in primary P. falciparum infected adults. Vγ9Vδ2 T-cells taken from the peripheral blood during paroxysms were found to expand less and produce less TNFα in response to IPP stimulation than Vγ9Vδ2 T-cells taken during recovery, post-treatment (though still expanded compared to uninfected controls). It was also found that there are less Vγ9Vδ2 T-cells [particularly Vγ2Jγ1.2γδ T-cells (US nomenclature), the TCR subset that is particularly reactive to Ph-Ags] in circulation during P. falciparum paroxysm than during recovery (34).
In a longitudinal study of Ugandan children, the percentage of Vγ9Vδ2 T-cells in peripheral blood was found to be inversely correlated with prior incidence of malaria infections. Ex vivo, Vγ9Vδ2 T-cell proliferation, TNFα, and IFNγ production and immune-modulatory gene expression was also negatively associated with prior malaria episodes—indicating decreased peripheral blood Vγ9Vδ2 T-cell activity with increasing exposure to the parasite. Lower in vitro Vγ9Vδ2 T-cell responsiveness to iRBC correlated with lower subsequent incidences of symptomatic infection, but to increased probability of higher parasitemia (56). This Vγ9Vδ2 T-cell dysfunction was shown to occur because of frequent malaria episodes in childhood, an effect that was abrogated by chemoprevention in early childhood (57). The mechanism of Vγ9Vδ2 T-cell regulation is as yet unknown. Vγ9Vδ2 T-cells are very susceptible to activation-induced cell death by Fas-Fas-L interaction as demonstrated for M. tuberculosis (58), though active regulation cannot be ruled out.
Together, these studies indicate that while Vγ9Vδ2 T-cell inflammatory cytokine responses can control parasitemia, excessive stimulation of these cells may also result in pathology suggesting that clinical immunity to malaria may be associated with reduced Vγ9Vδ2 responses.
Several accumulated data in mice, where the equivalent of human Vγ9Vδ2 T-cell subset is not yet certain, also show the importance of the cytokine secretion activity of murine γδ T-cells (59, 60). A recent study (61) showed that clonal expansion of a subset of γδ T-cells producing macrophage colony stimulating factor (M-CSF), prevents parasitemic recurrence. While it is perhaps a stretch to expect a direct murine equivalent of Vγ9Vδ2 T-cells, certainly one is not yet identified, it is likely that one or more murine γδ T-cell subsets have evolved which perform the same protective and/or pathologic functions in malaria infection as human Vγ9Vδ2 T-cells. “TγδM” cells are a good candidate for one such functional equivalent of Vγ9Vδ2 T-cells.
Antigen presenting vγ9vδ2 t-cells stimulate adaptive immune responses
Another way in which Vγ9Vδ2 T-cells influence the course of an immune response is by antigen presentation to αβ T-cells. Over the last 12 years it has been demonstrated that Vγ9Vδ2 T-cells can take up, process and present exogenous Ag, both via the classical pathway to CD4 T-cells and the cross-presentation pathway to CD8 T-cells. They even have shown the ability to act as professional antigen presenting cells (APCs) and stimulate naive CD4 and CD8 T-cells (62–70). γδ T-APC resembling cells are present in malaria infected individuals, and in vitro iRBC stimulated Vγ9Vδ2 T-cells not only take on an APC phenotype but also can cross-present Ag to a memory cell line and activate naïve CD4 and CD8 T-cells in a mixed-lymphocyte reaction (71). Where this might be occurring in vivo, or what the implication of repeated malaria infection could be is worth investigated. However, interesting work from liver stage malaria vaccines could shed some light on this (see below).
Overall, the data allow us to propose a global model of how peripheral Vγ9Vδ2 T-cells could control parasitemia and initiate both innate and adaptive responses (Figure 1). Whether the same cells are responsible for these functions or whether different subsets of Vγ9Vδ2 T-cell are concerned is still to be worked out.
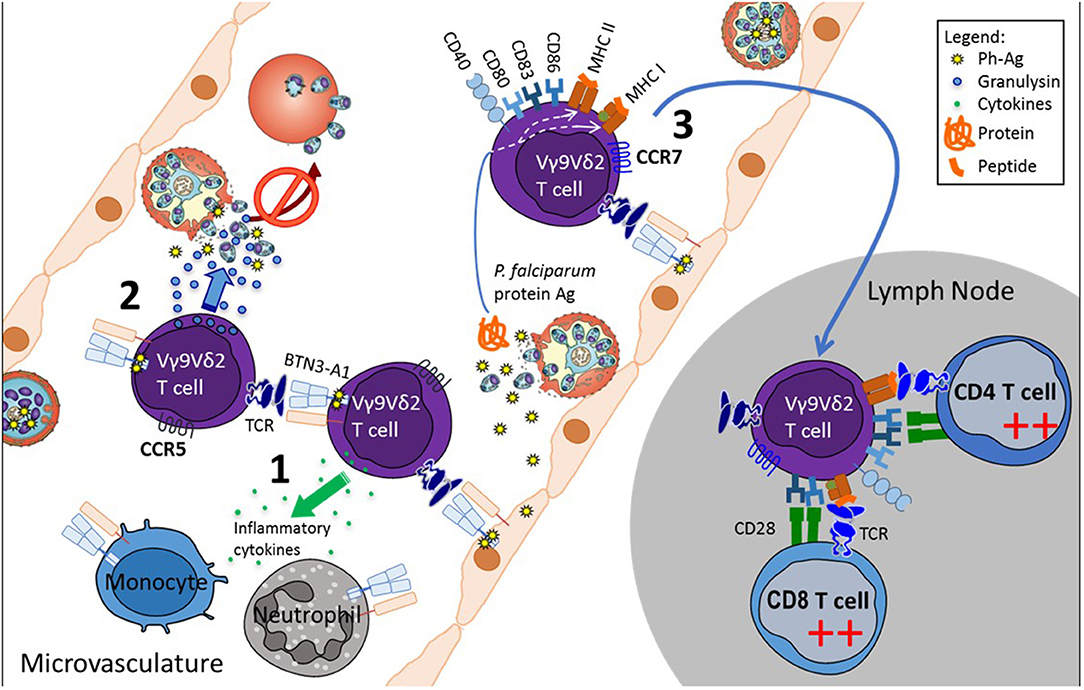
Figure 1. Proposed model of Vγ9Vδ2 T-cell functions in the microvasculature during P. falciparum infection. Plasmodium-infected red blood cells sequester to the endothelium in the microvasculature, where they release phosphoantigens concomitantly with the red blood cell rupture. Phosphoantigens stimulate Vγ9Vδ2 T-cells via BTN3A1 available on neighboring cells, including Vγ9Vδ2 T-cells, the endothelial cells and innate immune cells. Activated Vγ9Vδ2 T-cells (1) modulate innate cells by cytokine secretion, (2) inhibit free parasite reinvasion of red blood cells, by releasing the cytotoxic granulysin, and (3) acquire APC phenotype and the capability to migrate to lymph nodes where they initiate an adaptive immune response.
Vγ9vδ2 t-cells: correlates of protection for whole organism malaria vaccine?
Vγ9Vδ2 T-cells have been implicated in protection against liver stage immunity after vaccination with whole sporozoites.
In a mouse model of irradiated sporozoite vaccinations, it was clear that the γδ T-cells were required for induction of protective CD8 T-cell responses, but not antibodies, and were not acting as effectors in controlling liver stage parasite replication (72). An as yet undefined subset of mouse γδ T-cells are able to function by inducing downstream αβ T-cell responses. Further studies are required to establish which mouse γδ T-cell subsets mirror the various activities of Vγ9Vδ2 γδ T-cells and explore the effect of irradiated sporozoite vaccination dose on these cells. In humans, in the first field trial of the Sanaria® PfSPZ vaccine in Mali, it was demonstrated that the Vγ9Vδ2 T-cells were highest in vaccines that remained uninfected throughout an intense malaria transmission season, compared to infected vaccines or the placebo group (72). These findings were comparable to those observed in malaria naïve individuals vaccinated with either the PfSPZ vaccine or a chemoprophylaxis vaccination, who also had a remarkable increase in Vγ9Vδ2 T-cells (73). Overall, these findings are intriguing in that they suggest that liver-stages growth of P. falciparum can stimulate Vγ9Vδ2 T-cell activation. This activation could have several explanations: first, locally in an infected liver, hepatocytes displaying BNT3A1 or other presentation molecules could activate Vγ9Vδ2 T-cells in situ. Second, Vγ9Vδ2 T-cells could be activated in the draining lymph nodes of the site of infection where a substantial fraction of the sporozoites migrate, as shown by Amino et al. in mouse model (74). Third, HMBPP produced by liver stages of Plasmodium could be sensed in the periphery by exquisitely sensitive Vγ9Vδ2 T-cells, as seen during blood stage P. falciparum infections (37). Finally, the activation of Vγ9Vδ2 T-cells could be due to recognition of other antigens or metabolites.
It should be noted that in subsequent trials which used a higher dose of the PfSPZ vaccine, Vγ9Vδ2 T-cell expansion did not distinguish protected vs. unprotected vaccines (75, 76). Interestingly, liver stage induced Vγ9Vδ2 T-cell expansion has not been observed in volunteers undergoing controlled human malaria infections (77). The reasons behind this are not yet understood, but given the plasticity of Vγ9Vδ2 T-cells, it may be that varying antigen loads modulate the phenotype and function of these cells.
Concluding remarks
In conclusion, the Vγ9Vδ2 T-cell is an enigmatic cell, with a wide range of functions that can both contribute to and protect from malaria pathology. It is important to better consider this subset of γδ T-cells, especially their role in malaria vaccine protection. Given their sensitivity to Ph-Ag's such as HMBPP and apparent functional plasticity under different cytokines and stimuli dose, a cocktail of Ph-Ag and cytokines could be envisioned as an adjuvant to boost efficacy of both liver and blood stage malaria vaccines.
Author contributions
JH, IZ, and MM-M wrote the manuscript. SL and OM-P contributed to the manuscript. JD-M secured fundings.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This review includes content from the Doctoral thesis of JH, which is archived at Bordeaux University library (78). This work was supported in part by grants from the Centre National de la Recherche Scientifique, the French Research Agency [ANR-12-Blanc-GDSTRESS] and the Fondation pour la Recherche Médicale (FRM, équipe labellisée). JH and IZ are supported by the Intramural Research Program of the National Institute of Allergy and Infectious Disease, NIH.
References
1. Morita CT, Jin C, Sarikonda G, Wang H. Nonpeptide antigens, presentation mechanisms, and immunological memory of human Vgamma2Vdelta2 T cells: discriminating friend from foe through the recognition of prenyl pyrophosphate antigens. Immunol Rev. (2007) 215:59–76. doi: 10.1111/j.1600-065X.2006.00479.x
2. Liuzzi AR, McLaren JE, Price DA, Eberl M. Early innate responses to pathogens: pattern recognition by unconventional human T-cells. Curr Opin Immunol. (2015) 36:31–7. doi: 10.1016/j.coi.2015.06.002
3. Guggisberg AM, Amthor RE, Odom AR. Isoprenoid biosynthesis in Plasmodium falciparum . Eukaryot Cell. (2014) 13:1348–59. doi: 10.1128/EC.00160-14
4. Eberl M, Moser B. Monocytes and gammadelta T cells: close encounters in microbial infection. Trends Immunol. (2009) 30:562–8. doi: 10.1016/j.it.2009.09.001
5. World Health Organisation (WHO) (2017). WHO | World Malaria Report 2017. WHO. Available online at: http://www.who.int/malaria/publications/world-malaria-report-2017/en/ (Accessed February 26, 2018).
6. Murray CJL, Rosenfeld LC, Lim SS, Andrews KG, Foreman KJ, Haring D, et al. Global malaria mortality between 1980 and 2010: a systematic analysis. Lancet (2012) 379:413–31. doi: 10.1016/S0140-6736(12)60034-8
8. Marsh K, Kinyanjui S. Immune effector mechanisms in malaria. Parasite Immunol. (2006) 28:51–60. doi: 10.1111/j.1365-3024.2006.00808.x
9. Trape JF, Rogier C. Combating malaria morbidity and mortality by reducing transmission. Parasitol Today (1996) 12:236–40. doi: 10.1016/0169-4758(96)10015-6
10. Doolan DL, Dobaño C, Baird JK. Acquired immunity to malaria. Clin Microbiol Rev. (2009) 22:13–36. doi: 10.1128/CMR.00025-08
11. Langhorne J, Ndungu FM, Sponaas AM, Marsh K. Immunity to malaria: more questions than answers. Nat Immunol. (2008) 9:725–32. doi: 10.1038/ni.f.205
12. Struik SS, Riley EM. Does malaria suffer from lack of memory? Immunol Rev. (2004) 201:268–90. doi: 10.1111/j.0105-2896.2004.00181.x
13. Bousema T, Okell L, Felger I, Drakeley C. Asymptomatic malaria infections: detectability, transmissibility and public health relevance. Nat Rev Microbiol. (2014) 12:833–40. doi: 10.1038/nrmicro3364
14. Tran TM, Li S, Doumbo S, Doumtabe D, Huang CY, Dia S, et al. An intensive longitudinal cohort study of Malian children and adults reveals no evidence of acquired immunity to Plasmodium falciparum infection. Clin Infect Dis. (2013) 57:40–7. doi: 10.1093/cid/cit174
15. Arama C, Giusti P, Boström S, Dara V, Traore B, Dolo A, et al. Interethnic differences in antigen-presenting cell activation and TLR responses in Malian children during Plasmodium falciparum malaria. PLoS ONE (2011) 6:e18319. doi: 10.1371/journal.pone.0018319
16. Pinzon-Charry A, Woodberry T, Kienzle V, McPhun V, Minigo G, Lampah DA, et al. Apoptosis and dysfunction of blood dendritic cells in patients with falciparum and vivax malaria. J Exp Med. (2013) 210:1635–46. doi: 10.1084/jem.20121972
17. Urban BC, Cordery D, Shafi MJ, Bull PC, Newbold CI, Williams TN, et al. The frequency of BDCA3-positive dendritic cells is increased in the peripheral circulation of Kenyan children with severe malaria. Infect Immun. (2006) 74:6700–6. doi: 10.1128/IAI.00861-06
18. Illingworth J, Butler NS, Roetynck S, Mwacharo J, Pierce SK, Bejon P, et al. Chronic exposure to Plasmodium falciparum is associated with phenotypic evidence of B and T cell exhaustion. J Immunol. (2013) 190:1038–47. doi: 10.4049/jimmunol.1202438
19. Muellenbeck MF, Ueberheide B, Amulic B, Epp A, Fenyo D, Busse CE, et al. Atypical and classical memory B cells produce Plasmodium falciparum neutralizing antibodies. J Exp Med. (2013) 210:389–99. doi: 10.1084/jem.20121970
20. Weiss GE, Crompton PD, Li S, Walsh LA, Moir S, Traore B, et al. Atypical memory B cells are greatly expanded in individuals living in a malaria-endemic area. J Immunol. (2009) 183:2176–82. doi: 10.4049/jimmunol.0901297
21. Crompton PD, Moebius J, Portugal S, Waisberg M, Hart G, Garver LS, et al. Malaria immunity in man and mosquito: insights into unsolved mysteries of a deadly infectious disease. Annu Rev Immunol. (2014) 32:157–87. doi: 10.1146/annurev-immunol-032713-120220
22. Hafalla JC, Silvie O, Matuschewski K. Cell biology and immunology of malaria. Immunol Rev. (2011) 240:297–316. doi: 10.1111/j.1600-065X.2010.00988.x
23. Hviid L, Barfod L, Fowkes FJI. Trying to remember: immunological B cell memory to malaria. Trends Parasitol. (2015) 31:89–94. doi: 10.1016/j.pt.2014.12.009
24. Schwenk RJ, Richie TL. Protective immunity to pre-erythrocytic stage malaria. Trends Parasitol. (2011) 27:306–14. doi: 10.1016/j.pt.2011.02.002
25. Stanisic DI, Barry AE, Good MF. Escaping the immune system: how the malaria parasite makes vaccine development a challenge. Trends Parasitol. (2013) 29:612–22. doi: 10.1016/j.pt.2013.10.001
26. Hviid L, Akanmori BD, Loizon S, Kurtzhals JA, Ricke CH, Lim A, et al. High frequency of circulating gamma delta T cells with dominance of the v(delta)1 subset in a healthy population. Int Immunol. (2000) 12:797–805. doi: 10.1093/intimm/12.6.797
27. Hviid L, Kurtzhals JA, Adabayeri V, Loizon S, Kemp K, Goka BQ, et al. Perturbation and proinflammatory type activation of V delta 1(+) gamma delta T cells in African children with Plasmodium falciparum malaria. Infect Immun. (2001) 69:3190–6. doi: 10.1128/IAI.69.5.3190-3196.2001
28. Meraviglia S, El Daker S, Dieli F, Martini F, Martino A. γδ T cells cross-link innate and adaptive immunity in Mycobacterium tuberculosis infection. Clin Dev Immunol. (2011) 2011:1–11. doi: 10.1155/2011/587315
29. Scotet E, Nedellec S, Devilder MC, Allain S, Bonneville M. Bridging innate and adaptive immunity through gammadelta T-dendritic cell crosstalk. Front Biosci. (2008) 13:6872–85. doi: 10.2741/3195
30. Tyler CJ, Doherty DG, Moser B, Eberl M. Human Vγ9/Vδ2 T cells: innate adaptors of the immune system. Cell Immunol. (2015) 296:10–21. doi: 10.1016/j.cellimm.2015.01.008
31. Chen ZW. Multifunctional immune responses of HMBPP-specific Vγ2Vδ2 T cells in M. tuberculosis and other infections . Cell Mol Immunol. (2013) 10:58–64. doi: 10.1038/cmi.2012.46
32. Chang WL, van der Heyde H, Maki DG, Malkovsky M, Weidanz WP. Subset heterogeneity among gamma delta T cells found in peripheral blood during Plasmodium falciparum malaria. Immunol Lett. (1992) 32:273–4. doi: 10.1016/0165-2478(92)90061-R
33. Roussilhon C, Agrapart M, Guglielmi P, Bensussan A, Brasseur P, Ballet JJ. Human TcR gamma delta+ lymphocyte response on primary exposure to Plasmodium falciparum . Clin Exp Immunol. (1994) 95:91–7. doi: 10.1111/j.1365-2249.1994.tb06020.x
34. Martini F, Paglia MG, Montesano C, Enders PJ, Gentile M, Pauza CD, et al. V gamma 9V delta 2 T-cell anergy and complementarity-determining region 3-specific depletion during paroxysm of nonendemic malaria infection. Infect Immun. (2003) 71:2945–9. doi: 10.1128/IAI.71.5.2945-2949.2003
35. Bordessoule D, Gaulard P, Mason DY. Preferential localisation of human lymphocytes bearing gamma delta T cell receptors to the red pulp of the spleen. J Clin Pathol. (1990) 43:461–4. doi: 10.1136/jcp.43.6.461
36. Nakazawa S, Brown AE, Maeno Y, Smith CD, Aikawa M. Malaria-induced increase of splenic gamma delta T cells in humans, monkeys, and mice. Exp Parasitol. (1994) 79:391–8.
37. Guenot M, Loizon S, Howard J, Costa G, Baker DA, Mohabeer SY, et al. Phosphoantigen burst upon Plasmodium falciparum schizont rupture can distantly activate Vγ9Vδ2 T cells. Infect Immun. (2015) 83:3816–24. doi: 10.1128/IAI.00446-15
38. Elloso MM, van der Heyde HC, Troutt A, Manning DD, Weidanz WP. Human gamma delta T cell subset-proliferative response to malarial antigen in vitro depends on CD4+ T cells or cytokines that signal through components of the IL-2R. J Immunol. (1996) 157:2096–102.
39. Elloso MM, Wallace M, Manning DD, Weidanz WP. The effects of interleukin-15 on human gammadelta T cell responses to Plasmodium falciparum in vitro . Immunol Lett. (1998) 64:125–32. doi: 10.1016/S0165-2478(98)00088-1
40. Jones SM, Goodier MR, Langhorne J. The response of gamma delta T cells to Plasmodium falciparum is dependent on activated CD4+ T cells and the recognition of MHC class I molecules. Immunology (1996) 89:405–12. doi: 10.1046/j.1365-2567.1996.d01-762.x
41. Ribot JC, Debarros A, Mancio-Silva L, Pamplona A, Silva-Santos B. B7-CD28 costimulatory signals control the survival and proliferation of murine and human γδ T cells via IL-2 production. J Immunol. (2012) 189:1202–8. doi: 10.4049/jimmunol.1200268
42. Vermijlen D, Ellis P, Langford C, Klein A, Engel R, Willimann K, et al. Distinct cytokine-driven responses of activated blood gammadelta T cells: insights into unconventional T cell pleiotropy. J Immunol. (2007) 178:4304–14. doi: 10.4049/jimmunol.178.7.4304
43. Elloso MM, Van Der Heyde HC, Vande Waa JA, Manning DD, Weidanz WP. Inhibition of Plasmodium falciparum in vitro by human gamma delta T cells. J Immunol. (1994) 153:1187–94.
44. Farouk SE, Mincheva-Nilsson L, Krensky AM, Dieli F, Troye-Blomberg M. Gamma delta T cells inhibit in vitro growth of the asexual blood stages of Plasmodium falciparum by a granule exocytosis-dependent cytotoxic pathway that requires granulysin. Eur J Immunol. (2004) 34:2248–56. doi: 10.1002/eji.200424861
45. Troye-Blomberg M, Worku S, Tangteerawatana P, Jamshaid R, Söderström K, Elghazali G, et al. Human gamma delta T cells that inhibit the in vitro growth of the asexual blood stages of the Plasmodium falciparum parasite express cytolytic and proinflammatory molecules. Scand J Immunol. (1999) 50:642–50.
46. Costa G, Loizon S, Guenot M, Mocan I, Halary F, de Saint-Basile G, et al. Control of Plasmodium falciparum erythrocytic cycle: γδ T cells target the red blood cell-invasive merozoites. Blood (2011) 118:6952–62. doi: 10.1182/blood-2011-08-376111
47. D'Ombrain MC, Hansen DS, Simpson KM, Schofield L. gammadelta-T cells expressing NK receptors predominate over NK cells and conventional T cells in the innate IFN-gamma response to Plasmodium falciparum malaria. Eur J Immunol. (2007) 37:1864–73. doi: 10.1002/eji.200636889
48. Goodier MR, Lundqvist C, Hammarström ML, Troye-Blomberg M, Langhorne J. Cytokine profiles for human V gamma 9+ T cells stimulated by Plasmodium falciparum . Parasite Immunol. (1995) 17:413–23.
49. Pichyangkul S, Saengkrai P, Yongvanitchit K, Stewart A, Heppner DG. Activation of gammadelta T cells in malaria: interaction of cytokines and a schizont-associated Plasmodium falciparum antigen. J Infect Dis. (1997) 176:233–41. doi: 10.1086/514029
50. Scragg IG, Hensmann M, Bate CA, Kwiatkowski D. Early cytokine induction by Plasmodium falciparum is not a classical endotoxin-like process. Eur J Immunol. (1999) 29:2636–44.
51. Engelmann I, Santamaria A, Kremsner PG, Luty AJF. Activation status of cord blood gamma delta T cells reflects in utero exposure to Plasmodium falciparum antigen. J Infect Dis. (2005) 191:1612–22. doi: 10.1086/429336
52. Roestenberg M, McCall M, Hopman J, Wiersma J, Luty AJF, van Gemert GJ, et al. Protection against a malaria challenge by sporozoite inoculation. N Engl J Med. (2009) 361:468–77. doi: 10.1056/NEJMoa0805832
53. Teirlinck AC, McCall MBB, Roestenberg M, Scholzen A, Woestenenk R, de Mast Q, et al. Longevity and composition of cellular immune responses following experimental Plasmodium falciparum malaria infection in humans. PLoS Pathog. (2011) 7:e1002389. doi: 10.1371/journal.ppat.1002389
54. D'Ombrain MC, Robinson LJ, Stanisic DI, Taraika J, Bernard N, Michon P, et al. Association of early interferon-gamma production with immunity to clinical malaria: a longitudinal study among Papua New Guinean children. Clin Infect Dis. (2008) 47:1380–7. doi: 10.1086/592971
55. Stanisic DI, Cutts J, Eriksson E, Fowkes FJI, Rosanas-Urgell A, Siba P, et al. γδ T cells and CD14+ monocytes are predominant cellular sources of cytokines and chemokines associated with severe malaria. J Infect Dis. (2014) 210:295–305. doi: 10.1093/infdis/jiu083
56. Jagannathan P, Kim CC, Greenhouse B, Nankya F, Bowen K, Eccles-James I, et al. Loss and dysfunction of V 2+ T cells are associated with clinical tolerance to malaria. Sci Transl Med. (2014) 6:251ra117. doi: 10.1126/scitranslmed.3009793
57. Farrington LA, Jagannathan P, McIntyre TI, Vance HM, Bowen K, Boyle MJ, et al. Frequent malaria drives progressive Vδ2 T-cell loss, dysfunction, and CD16 up-regulation during early childhood. J Infect Dis. (2016) 213:1483–90. doi: 10.1093/infdis/jiv600
58. Li B, Bassiri H, Rossman MD, Kramer P, Eyuboglu AF, Torres M, et al. Involvement of the Fas/Fas ligand pathway in activation-induced cell death of mycobacteria-reactive human gamma delta T cells: a mechanism for the loss of gamma delta T cells in patients with pulmonary tuberculosis. J Immunol. (1998) 161:1558–67.
59. Inoue SI, Niikura M, Mineo S, Kobayashi F. Roles of IFN-γ and γδ T cells in protective immunity against blood-stage malaria. Front Immunol. (2013) 4:258. doi: 10.3389/fimmu.2013.00258
60. Inoue S-I, Niikura M, Asahi H, Iwakura Y, Kawakami Y, Kobayashi F. Preferentially expanding Vγ1 + γδ T cells are associated with protective immunity against Plasmodium infection in mice. Eur J Immunol. (2017) 47:685–91. doi: 10.1002/eji.201646699
61. Mamedov MR, Scholzen A, Nair RV, Cumnock K, Kenkel JA, Oliveira JHM, et al. A macrophage colony-stimulating-factor-producing γδ T cell subset prevents malarial parasitemic recurrence. Immunity (2018) 48:350–63.e7. doi: 10.1016/j.immuni.2018.01.009
62. Altvater B, Pscherer S, Landmeier S, Kailayangiri S, Savoldo B, Juergens H, et al. Activated human gammadelta T cells induce peptide-specific CD8+ T-cell responses to tumor-associated self-antigens. Cancer Immunol Immunother (2012) 61:385–96. doi: 10.1007/s00262-011-1111-6
63. Brandes M, Willimann K, Moser B. Professional antigen-presentation function by human gammadelta T Cells. Science (2005) 309:264–8. doi: 10.1126/science.1110267
64. Himoudi N, Morgenstern DA, Yan M, Vernay B, Saraiva L, Wu Y, et al. Human γδ T lymphocytes are licensed for professional antigen presentation by interaction with opsonized target cells. J Immunol. (2012) 188:1708–16. doi: 10.4049/jimmunol.1102654
65. Khan MWA, Curbishley SM, Chen HC, Thomas AD, Pircher H, Mavilio D, et al. Expanded human blood-derived γδT cells display potent antigen-presentation functions. Front Immunol. (2014) 5:344. doi: 10.3389/fimmu.2014.00344
66. Landmeier S, Altvater B, Pscherer S, Juergens H, Varnholt L, Hansmeier A, et al. Activated human gammadelta T cells as stimulators of specific CD8+ T-cell responses to subdominant Epstein Barr virus epitopes: potential for immunotherapy of cancer. J Immunother. (2009) 32:310–21. doi: 10.1097/CJI.0b013e31819b7c30
67. Meuter S, Eberl M, Moser B. Prolonged antigen survival and cytosolic export in cross-presenting human gammadelta T cells. Proc Natl Acad Sci USA. (2010) 107:8730–5. doi: 10.1073/pnas.1002769107
68. Muto M, Baghdadi M, Maekawa R, Wada H, Seino KI. Myeloid molecular characteristics of human γδ T cells support their acquisition of tumor antigen-presenting capacity. Cancer Immunol Immunother. (2015) 64:941–9. doi: 10.1007/s00262-015-1700-x
69. Wu Y, Wu W, Wong WM, Ward E, Thrasher AJ, Goldblatt D, et al. Human gamma delta T cells: a lymphoid lineage cell capable of professional phagocytosis. J Immunol. (2009) 183:5622–9. doi: 10.4049/jimmunol.0901772
70. Brandes M, Willimann K, Bioley G, Lévy N, Eberl M, Luo M, et al. Cross-presenting human gammadelta T cells induce robust CD8+ alphabeta T cell responses. Proc Natl Acad Sci USA (2009) 106:2307–12. doi: 10.1073/pnas.0810059106
71. Howard J, Loizon S, Tyler CJ, Duluc D, Moser B, Mechain M, et al. The antigen-presenting potential of Vγ9Vδ2 T cells during Plasmodium falciparum blood-stage infection. J Infect Dis. (2017) 215:1569–79. doi: 10.1093/infdis/jix149
72. Zaidi I, Diallo H, Conteh S, Robbins Y, Kolasny J, Orr-Gonzalez S, et al. γδ T cells are required for the induction of sterile immunity during irradiated sporozoite vaccinations. J Immunol. (2017) 199:3781–8. doi: 10.4049/jimmunol.1700314
73. Ishizuka AS, Lyke KE, DeZure A, Berry AA, Richie TL, Mendoza FH, et al. Protection against malaria at 1 year and immune correlates following PfSPZ vaccination. Nat Med. (2016) 22:614–23. doi: 10.1038/nm.4110
74. Amino R, Thiberge S, Martin B, Celli S, Shorte S, Frischknecht F, et al. Quantitative imaging of Plasmodium transmission from mosquito to mammal. Nat Med. (2006) 12:220–4. doi: 10.1038/nm1350
75. Lyke KE, Ishizuka AS, Berry AA, Chakravarty S, DeZure A, Enama ME, et al. Attenuated PfSPZ vaccine induces strain-transcending T cells and durable protection against heterologous controlled human malaria infection. Proc Natl Acad Sci USA. (2017) 114:2711–6. doi: 10.1073/pnas.1615324114
76. Mordmüller B, Surat G, Lagler H, Chakravarty S, Ishizuka AS, Lalremruata A, et al. Sterile protection against human malaria by chemoattenuated PfSPZ vaccine. Nature (2017) 542:445–9. doi: 10.1038/nature21060
Keywords: gamma-delta T cells, malaria, falciparum, immunity to malaria, antigen presenting cell (APC), cytotoxicity
Citation: Howard J, Zaidi I, Loizon S, Mercereau-Puijalon O, Déchanet-Merville J and Mamani-Matsuda M (2018) Human Vγ9Vδ2 T Lymphocytes in the Immune Response to P. falciparum Infection. Front. Immunol. 9:2760. doi: 10.3389/fimmu.2018.02760
Received: 31 March 2018; Accepted: 09 November 2018;
Published: 27 November 2018.
Edited by:
Kenth Gustafsson, University College London, United KingdomReviewed by:
Jean Langhorne, Francis Crick Institute, United KingdomLars Hviid, University of Copenhagen, Denmark
Copyright © 2018 Howard, Zaidi, Loizon, Mercereau-Puijalon, Déchanet-Merville and Mamani-Matsuda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maria Mamani-Matsuda, bWFyaWEubWFtYW5pLW1hdHN1ZGFAdS1ib3JkZWF1eC5mcg==