 Igal Gery
Igal Gery Rachel R. Caspi
Rachel R. Caspi- Laboratory of Immunology, National Eye Institute, Bethesda, MD, United States
Inflammatory intraocular eye diseases, grouped under the term uveitis are blinding conditions, believed to be mediated by pathogenic autoimmune processes that overcome the protective mechanisms of the immune privilege status of the eye. An animal model for these diseases, named experimental autoimmune uveitis (EAU), is induced by initiation of immunity against ocular-specific antigens, or it develops spontaneously in mice with T-cells that transgenically express TCR specific to the target eye antigen(s). T-Cells specific to ocular antigens are generated in the thymus and their majority are eliminated by exposure to their target antigen expressed in this organ. T-cells that escape this negative selection acquire pathogenicity by their activation with the target antigen. In spontaneous EAU, the microbiota play crucial roles in the acquisition of pathogenicity by providing both antigenic stimulation, by molecules that mimic the target ocular antigen, and an additional stimulation that allows invasion of tissues that harbor the target antigen. The pathogenic process is physiologically inhibited by the peripheral tolerance, composed of antigen-specific T-regulatory (Treg) lymphocytes. Deleting the Tregs enhances the ocular inflammation, whereas adoptively transferring them suppresses the pathogenic response. Potential usage of Treg cells for suppression of autoimmune diseases in humans is under intensive investigation.
Introduction
The clarity of ocular tissues and spaces is crucial for sight. Inflammatory processes thus have detrimental effects on vision, by causing media opacity. Furthermore, the damaging effects of inflammation in the eye are also harmful by affecting photoreceptor cells that are critical for the process of vision, and do not regenerate. To protect against immune-mediated inflammatory processes, the eye has evolved multiple layers of defense that endow it with a status of immune privileged organ (1). These layers of defense, studied, and defined by many investigators over the course of recent four decades include: (i) an efficient blood-retina barrier; (ii) an immunoinhibitory environment composed of several soluble and cell-bound immunosuppressive molecules; (iii) an active process known as “anterior chamber associated immune deviation (ACAID)” which can be demonstrated experimentally, in which foreign antigens reaching the anterior chamber of the eye initiate a complex immunosuppressive process (2, 3).
Early on, it was thought that due to their sequestration in the eye, ocular antigens, in particular those of the retina, are treated as foreign and provoke immune responses when exposed to the immune system. This notion was supported by the features of Sympathetic Ophthalmia, a disease in which trauma to one eye is followed by inflammation in the other, undamaged, eye. Autoimmunity has been proposed to play a major role in the pathogenesis of sympathetic ophthalmia, as well as in the pathogenesis of other human eye diseases in which intraocular inflammation without an accompanying infection, is the major symptom. These include birdshot chorioretinopathy, Behcet's disease, Sarcoidosis, and Vogt-Koyanagi-Harada (VKH) disease; other inflammatory eye diseases listed under the “uveitis” umbrella are not covered here.
Pathogenic autoimmunity is also assumed to be the mechanism responsible for phacoanaphylactic endophthalmitis, a rare but severe eye disease that follows damage to the lens and the release of lens antigens, proteins that are normally sequestered from the immune system (4).
As will be discussed below, the “physiological” approach to treating autoimmune-mediated diseases would be by induction of specific tolerance to the autoantigens that are the targets for the pathogenic response. We summarize here basic data concerning these issues, specifically dealing with tolerance to antigenic components of intraocular tissues of the eye. Immunity and tolerance to the cornea and external eye tissues have been investigated thoroughly and were reviewed elsewhere (5–7).
Uveitis—Current Status
Intraocular inflammatory conditions in humans are grouped under the term noninfectious uveitis, that includes the five eye diseases mentioned above (8, 9). Autoimmune processes are believed to play a major role in the pathogenesis of these diseases, a notion supported by findings of cellular and/or humoral immune responses against ocular auto-antigens in these patients (8–11). In addition, involvement of autoimmune processes is strongly supported by the similarity between the pathological changes specific to these human diseases and the ocular changes seen in animals with experimental autoimmune uveoretinitis (EAU) (8, 9, 12). This animal disease is induced by immunization with retina-specific antigens and is mediated by pathogenic T- lymphocytes (9, 13, 14). The notion that the human conditions are mediated by immunopathogenic T-cells is also supported by the finding that, like animal models (15, 16), patients with these conditions tend to respond positively to treatment with immunosuppressive agents that mainly target T cells, such as cyclosporine, rapamycin, and FK-506 (16–18) and, more recently, by daclizumab (antibody to the interleukin 2 receptor) and other biologics (19–21).
Treatment with immunosuppressive agents, however, inevitably has adverse effects on host defense. Therefore, the ideal treatment would be induction of antigen-specific tolerance. Selective tolerance to uveitogenic antigens has been achieved in animals, in which treatment with a uveitogenic protein (S-antigen/retinal arrestin) yielded inhibition of EAU induced by immunization with this same molecule (22, 23). In these studies the antigen was administered by oral gavage during the development of the experimental disease and Dick et al. (24) reported high efficiency of EAU suppression by intranasal administration of the retinal antigen. Since patients with uveitis often demonstrate immunity toward retinal S-antigen (10, 25), studies were carried out to examine the efficacy of treating such patients for induction of oral tolerance by feeding with this antigen (26), or with an HLA peptide that crossreacts with S-antigen (27). The initial results appeared encouraging, but additional studies are required to examine the therapeutic usefulness of the procedure in more depth. The main advantage of oral tolerance is the absence of known adverse effects, but its drawback, as with other antigen-specific tolerance approaches, is that it requires knowledge of the inciting antigens. However, the specific antigens that serve as molecular targets for uveitis are not certain. That said, the phenomenon of tissue specific bystander suppression, where regulatory cells induced to one antigen may suppress responses to other antigens in the same tissue environment (28) could, in theory, be exploited to get around this limitation.
Central and Peripheral Tolerance in Relation to the Eye
Over the past few decades, research with animals, primarily mice, has provided a large amount of information concerning the complex process of tolerance. The state of tolerance to self antigens is achieved by two separate processes: central and peripheral tolerance, which operate in tandem. When either of these fails, autoimmunity can result. Central tolerance occurs in the thymus. As part of their development, new T-lymphocytes with specificity toward a wide range of antigens are generated in the thymic cortex, and follow a stereotypic process of maturation through several well defined stages (29, 30). As part of this process, maturing T-cells with high affinity toward autoantigens undergo apoptotic death in the thymic medulla upon exposure to their cognate tissue antigens, whereas T cells with intermediate affinity may be converted into natural T regulatory cells (nTregs). The tissue antigens that trigger this process of thymic education were shown to be expressed ectopically by thymic medullary epithelial cells (29, 30).
Several groups have examined the process of central and peripheral tolerance toward retinal antigens, using transgenic mice expressing a neo-self antigen in the retina (hen egg lysozyme = HEL, or β-galactosidase = β-Gal) and/or a transgenic TCR to the neo- or a native self-antigen (31–36). With one exception, which may have been due to technical limitations (32), the conclusion reached by these studies is that there was detectable elimination of retina-specific T cells in the thymus. This was subsequently shown to be due to expression of retinal antigens (or retinal neo-antigens) in the thymus under control of the AutoImmune REgulator (AIRE) transcription factor that drives expression of tissue-specific antigens, including retinal antigens, in the thymus, and mediates negative selection of autoreactive lymphocytes (37, 38). Expression in the thymus of a natural uveitogenic retinal antigen, interphotoreceptor retinoid-binding protein (IRBP), was demonstrated by using RT-PCR (39) and immunohistochemistry (36) and was positively correlated in different mouse strains with their resistance to EAU (39) and with elimination of reactivity to specific uveitogenic epitopes from the immune repertoire (40). Importantly, testing thymic expression of ocular-specific genes (S-antigen, IRBP, RPE65, and recoverin) in thymi of several human individuals revealed a remarkable variability in the level of expression of these molecules (41). These data thus provide a possible mechanistic explanation for the differences among individuals in their susceptibility to autoimmune uveitis and to tissue specific autoimmune diseases in general, suggesting that the susceptibility is regulated at least in part by the level of thymic expression of the pathogenic autoantigens.
It is also of note that AIRE-controlled expression of self antigens plays an important role in the generation of thymus-derived natural Treg cells (nTregs) (42, 43). This likely includes also IRBP-specific nTreg cells that might account for raising the threshold of susceptibility to EAU, even though after a uveitogenic challenge, disease may be regulated also by induced Tregs (43), elicited in the periphery as a result of the immunization and disease process.
The process of thymic central tolerance alone is insufficient to eliminate all the T-lymphocytes reactive to tissue-specific antigens and consequently, a proportion of self-reactive cells manage to exit to the periphery. Such escapee cells are normally kept in check by a process known as peripheral tolerance, namely, exposure to the cognate antigen in the tissue in the absence of costimulatory danger signals, precipitates immunological paralysis (anergy), or conversion to Tregs (dubbed peripheral or induced Tregs). In a curious way, this parallels the process of central tolerance, in that it requires contact with the cognate antigen, whereupon the autoreactive cells are disarmed.
In the case of the eye, however, peripheral tolerance may not operate efficiently due to limited accessibility of the tissue antigens which are unique to the eye, and are largely sequestered behind a blood retinal barrier. In support of this notion, retinal neo-antigen presentation is not detectable in eye-draining lymph nodes, and circulating TCR Tg lymphocytes specific to retina display a largely naïve phenotype (31, 33, 44, 45). Therefore, we believe that T cells that have not been deleted in the thymus persist in the periphery in a non-tolerant state. This notion is further supported by data showing that gene expression of retinal antigens outside the eye, e.g., by transgenesis, retroviral transduction, or hydrodynamic injection, confers profound resistance to EAU (33, 43, 46). Furthermore, healthy humans have a relatively high frequency of circulating T cells specific to retinal antigens (47). These concepts are illustrated in Figure 1. In the aggregate, the data paint a picture whereby tolerance to retinal antigens is dependent mainly on thymic selection, whereas peripheral tolerance mechanisms are a “weak link” that may present an opportunity of therapeutic manipulation.
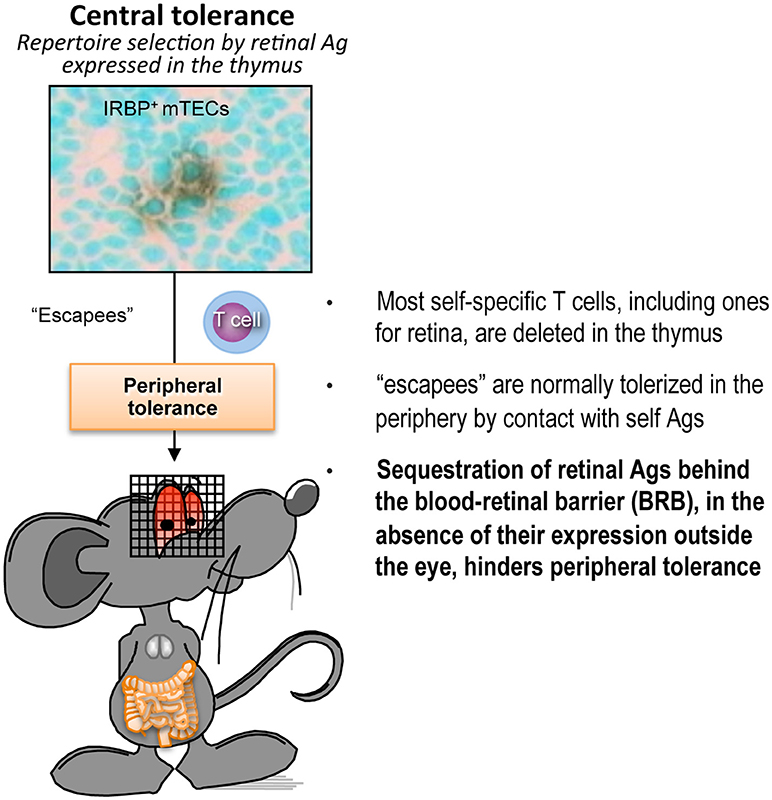
Figure 1. Peripheral tolerance is inefficient in the case of retina-specific T cells. Schematic representation of the process of self-tolerance to retinal antigens. Adapted from Horai and Caspi (48). No copyright permissions are required for the reuse of this image.
Natural and Experimental Triggers of Uveitis
Despite their presence in the circulation, naïve autoreactive T cells clearly do not cause uveitis in most individuals; perhaps, because they are unable to invade the target organs without additional activation processes. It has been well established in models of autoimmune disease that in order to elicit disease by infusion of autoimmune lymphocytes, these cells must first be activated in vitro. The notion that circulating naïve T cells are incapable of invading normal tissues, is supported by findings in mice. Thus, naïve CD4 T cells specific to a neo-self eye antigen do not induce ocular inflammation, unless they are pre-activated (in vitro) (49) and the same was true of T cells that express transgenic TCR specific for the natural retinal antigen IRBP (44). This leads to the question, where do retina-specific uveitogenic lymphocytes become activated in vivo to acquire the ability to penetrate the blood-retinal barrier and infiltrate the eye. Importantly, exposure in vivo to innate stimuli, such as TLR ligands, or complete Freund's adjuvant (which contains heat killed mycobacteria), without in vitro preactivation of the autoreactive cells, could be sufficient to support induction of uveitis in both uveitis models mentioned above (39, 40). While additional effects on the host, such as increased leakiness of the vasculature (50), could not be excluded as contributing factors, these observations strongly suggest that microbial stimuli might be involved in triggering uveitis.
Extrapolating from the knowledge that innate microbial signals (e.g., complete Freund's adjuvant) drive tissue specific T cells to a pathogenic effector phenotype, Caspi and coworkers further explored the role of commensal microbiota as a trigger of uveitis. Toward that end, they developed the R161H mouse strain, which expresses an IRBP specific TCR and develop autoimmune uveitis spontaneously (44). Rearing the mice under cover of broad spectrum antibiotics, or under germ-free conditions, strongly attenuated development of disease, supporting the notion that commensal flora can serve as a trigger of autoimmune uveitis (45). Importantly, in microbe-containing mice, the retina-specific R161H cells were seen to signal through their IRBP-specific receptors in the intestine, suggesting that they were being activated in situ by commensal flora in an antigen specific fashion to trigger disease. The proposed scenario of the gut-eye axis in uveitis is depicted in Figure 2. Interestingly, Gery and colleagues found (51, 52), that the process of pathogenicity acquisition by uveitogenic T-cells requires an additional phase of 2–3 days in the spleen, and possibly other organs, during which in vitro activated autoimmune lymphocytes are “licensed” to invade tissues where the target antigen is located. Similar findings were reported for the brain, another immune-privileged tissue (53).
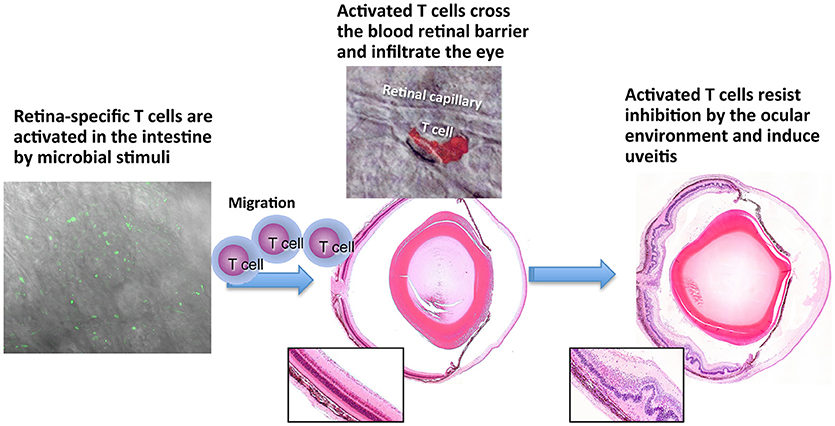
Figure 2. Persisting retina-specific T cells can be activated by peripheral stimuli. Schematic representation of EAU induction after activation of retina-specific T cells in the gut. Green cells in left panel are nurllgfP-positive T cells that are signaling through their T cell receptor [Horai et al. (45)]. Red cell in the middle top panel is a retina-specific T cell extravasating from a retinal vessel. The cell is stained by the PKH26 tracking dye and this photo was previously published in Prendergast et al. (50). No copyright permissions are required for the reuse of this image.
Immune Privilege and Uveitis
As briefly discussed above, the eye is protected from the immune system by the complex phenomenon of immune privilege, in which sequestration is only the first layer of defense. The internal environment of the eye contains a variety of potent soluble and cell-bound inhibitory molecules, including TGF-β, α-MSH, CGRP, VIP, and retinoic acid, as well as FasL, PD-L1, TSP-1, to name a few [reviewed in ref (3)]. Multiple studies over the last 3 decades presented evidence that ocular fluids and ocular resident cells inhibit activation and function of various immune cells in culture, and can even convert T helper (Th) cells to Foxp3+ Tregs (54). How, then, can we explain induction of uveitis in the face of immune privilege? To examine whether the living eye has the ability to control activation of uveitogenic T cells, Caspi and colleagues injected IRBP-specific T cells obtained from R161H donor mice directly into eyes of recipient mice, and followed their fate. The data showed that naïve R161H T cells recognized their cognate antigen but did not acquire effector function in the eye. Instead, they converted to Foxp3+ Treg cells, which were functional and were able to inhibit activation of fresh R161H T cells in a standard T cell suppression assay (54). In contrast, antigen-experienced R161H T cells, isolated from IRBP immunized R161H mice by selection for activation/memory markers, that had been exposed to IRBP before being placed in the eye, could not be inhibited by the suppressive intraocular environment and caused extensive photoreceptor damage (54). This can explain why T cells that have been activated outside the eye—whether as a result of trauma or activation in the gut—and have acquired the ability to penetrate the blood-retinal barrier, can induce uveitis despite ocular immune privilege.
Natural and Therapeutic Regulation of Uveitis
Peripheral tolerance against pathogenic autoimmunity is executed by several types of regulatory cells, including, but are not limited to, natural, and induced Tregs. Treg cells are capable of killing and/or suppressing T-lymphocytes specific to autoantigens, that escaped the negative selection process in the thymus and migrated into the blood and lymphoid organs [reviewed in ref (55)]. In early studies, the CD4 Treg cells were identified mainly by their strong surface expression of CD25 (56), but their identification has been greatly improved by the finding that their majority express the transcription factor FoxP3. Depletion of Foxp3+ Tregs caused the mice to develop more severe uveitis upon IRBP challenge, indicating that preexisting (likely thymic-derived) Tregs raise the threshold of susceptibility to uveitis (57). In mice that already developed uveitis, depletion of Foxp3+ Tregs at the peak of disease prevented resolution, and depletion after resolution induced a relapse, indicating that Foxp3+ Tregs naturally bring about and maintain resolution of EAU (43). The suppressive activity of Treg cells in the EAU system was also examined by Gregerson group, who found that Tregs may be generated in the eye and protect the retina from EAU induced by active immunization or adoptively transferred pathogenic Th cells. Interestingly, the generation of local Treg cells is inhibited by selective elimination of dendritic cells (DC) and microglia cannot replace DC in this function (58).
Treg cells that do not express Foxp3 (Tr1) have also been reported and characterized (59). Tr1 cells may be important in uveitis, but less is known about them, as they are more difficult to identify, and therefore to study. Tr1 cells are induced by IL-27 (60), as well as by c-Maf, Il-21 and ICOS (61). Caspi group demonstrated that IL-27 is potently induced in DC by IFN-γ-producing natural killer (NK) cells, in a self-amplifying feedback loop that takes place in the draining lymph node of mice immunized for uveitis. This interaction dampens the immune response that leads to uveitis, thus identifying NK cells as a novel regulatory cell that controls the magnitude of the autoimmune response (62). Other cells identified to be immunosuppressive in the context of EAU are IL-35 producing B-cells that were recently found by Egwuagu and colleagues to suppress EAU development, in part through induction of IL-35-producing Tregs (63, 64). Interestingly, this group showed that the IL-12p35 subunit has immunoregulatory functions that were hitherto attributed to IL-35 (65). IL-35 is also expressed by Treg cells and, interestingly, Wei et al. reported that that different subpopulations of Tregs produce IL-35 or IL-10 (66).
Based on the data described above, the group of Caspi set out to examine whether therapeutic induction of tolerance could regulate the pathogenic process of EAU. Treatment of mice with an IRBP expression plasmid, in the form of naked DNA administered by hydrodynamic injection, markedly suppressed the induction of EAU in the treated mice and analysis of the inhibitory process revealed that the inhibition of EAU was mostly due to CD4+CD25+FoxP3+ regulatory cells (43). Notably, Treg cells from hydrodynamically injected mice could be expanded into functionally suppressive Treg cell lines that, when adoptively transferred to mice immunized with IRBP, inhibited EAU development in the recipient mice (43).
In view of the specific and potent inhibitory capacity of Treg cells, the notion of using these cells in suppression of autoimmune disease in humans seems very attractive (67). One approach is to isolate antigen-specific Treg cells, increase their numbers ex vivo and inject them back to the patient (68). Another approach, proposed by the Salomon group (69), offers uveitis as the model disease, and uses preactivated polyclonal Treg cells that would exert bystander suppression in the target tissue. The system was tested in mice with EAU and suppression of disease was achieved, but only when the cells were injected into the vitreous. So far, no data have been reported to show successful treatment of uveitis patients with Treg cells. However, the notion that Treg cells are involved in suppression of the pathogenic process of uveitis is supported by the finding of a correlation between increase in the proportions of Treg cells in blood of uveitic patients and remission of the disease (70). Furthermore, the frequency of Treg cells was found to decline in parallel with increase in the severity of the uveitic changes (71).
In addition to T and B cells, myeloid cells can also regulate EAU. Myeloid-derived suppressor cells (MDSC) are a rather heterogeneous myeloid cell population that may include monocyte/macrophage- and granulocyte-like populations. Evidence reported by several groups indicates that MDSC could act at the systemic as well as local levels to curb disease (72, 73). Importantly, MDSC have also been identified in association with human uveitis (73). Additionally, dendritic cells (DC) having a tolerogenic phenotype may also play a regulatory role. Forrester and collaborators reported that treatment of mice with LPS-induced tolerogenic DCs, which produced IL-2 and suppressed uveitis by multiple mechanisms (74).
Finally, ocular inflammation may also be regulated by non-lymphoid cells, known as mesenchymal stem/stromal cells (MSCs). The mechanisms may involve induction of MDSC-like cells (75) as well as induction of Treg cells (76).
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Funding
This work was supported by NEI Intramural funding, project numbers EY000184 (RRC) and EY00069 (IG).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Medawar PB III. The fate of skin homografts transplanted to the brain, to subcutaneous tissue, and to the anterior chamber of the eye. Br J Exp Pathol. (1948) 29:58–69.
2. Streilein JW. Ocular immune privilege: therapeutic opportunities from an experiment of nature. Nat Rev Immunol. (2003) 3:879–89. doi: 10.1038/nri1224
3. Taylor AW. Ocular immune privilege and transplantation. Front Immunol. (2016) 7:37. doi: 10.3389/fimmu.2016.00037
5. Cursiefen C, Chen L, Dana MR, Streilein JW. Corneal lymphangiogenesis: evidence, mechanisms, and implications for corneal transplant immunology. Cornea (2003) 22:273–81. doi: 10.1097/00003226-200304000-00021
6. Tahvildari M, Amouzegar A, Foulsham W, Dana R. Therapeutic approaches for induction of tolerance and immune quiescence in corneal allotransplantation. Cell Mol Life Sci. (2018) 75:1509–20. doi: 10.1007/s00018-017-2739-y
7. Pflugfelder SC, de Paiva CS. The Pathophysiology of dry eye disease: what we know and future directions for research. Ophthalmology (2017) 124:S4–13. doi: 10.1016/j.ophtha.2017.07.010
8. Nussenblatt RB, Gery I. Experimental autoimmune uveitis and its relationship to clinical ocular inflammatory disease. J Autoimmun. (1996) 9:575–85. doi: 10.1006/jaut.1996.0077
9. Caspi RR. A look at autoimmunity and inflammation in the eye. J Clin Invest. (2010) 120:3073–83. doi: 10.1172/JCI42440
10. Doekes G, van der Gaag R, Rothova A, van Kooyk Y, Broersma L, Zaal MJ, et al. Humoral and cellular immune responsiveness to human S-antigen in uveitis. Curr Eye Res. (1987) 6:909–19.
11. Forrester JV. Duke-elder lecture: new concepts on the role of autoimmunity in the pathogenesis of uveitis. Eye (1992) 6 (Pt 5) 433–46.
12. Hirose S, Kuwabara T, Nussenblatt RB, Wiggert B, Redmond TM, Gery I. Uveitis induced in primates by interphotoreceptor retinoid-binding protein. Arch Ophthalmol. (1986) 104:1698–702. doi: 10.1001/archopht.1986.01050230136049
13. Mochizuki M, Kuwabara T, McAllister C, Nussenblatt RB, Gery I. Adoptive transfer of experimental autoimmune uveoretinitis in rats. Immunopathogenic mechanisms and histologic features. Invest Ophthalmol Vis Sci. (1985) 26:1–9.
14. Caspi RR, Roberge FG, McAllister CG, Mel-Saied, Kuwabara T, Gery I, et al. T cell lines mediating experimental autoimmune uveoretinitis (EAU) in the rat. J Immunol. (1986) 136:928–33.
15. Nussenblatt RB, Rodrigues MM, Salinas-Carmona MC, Gery I, Cevario S, Wacker W. Modulation of experimental autoimmune uveitis with cyclosporin A. Arch Ophthalmol. (1982) 100:1146–9. doi: 10.1001/archopht.1982.01030040124022
16. Hennig M, Bauer D, Wasmuth S, Busch M, Walscheid K, Thanos S, et al. Everolimus improves experimental autoimmune uveoretinitis. Exp Eye Res. (2012) 105:43–52. doi: 10.1016/j.exer.2012.09.003
17. Nussenblatt RB, Palestine AG, Rook AH, Scher I, Wacker WB, Gery I. Treatment of intraocular inflammatory disease with cyclosporin A. Lancet (1983) 2:235–8. doi: 10.1016/S0140-6736(83)90230-1
18. Pleyer U, Thurau SR. Sirolimus for the treatment of noninfectious uveitis. Expert Opin Pharmacother (2016) 17:127–35. doi: 10.1517/14656566.2016.1124855
19. Heiligenhaus A, Thurau S, Hennig M, Grajewski RS, Wildner G. Anti-inflammatory treatment of uveitis with biologicals: new treatment options that reflect pathogenetic knowledge of the disease. Graefes Arch Clin Exp Ophthalmol. (2010) 248:1531–51. doi: 10.1007/s00417-010-1485-8
20. Larson T, Nussenblatt RB, Sen HN. Emerging drugs for uveitis. Expert Opin Emerg Drugs (2011) 16:309–22. doi: 10.1517/14728214.2011.537824
21. Mushtaq B, Saeed T, Situnayake RD, Murray PI. Adalimumab for sight-threatening uveitis in Behcet's disease. Eye (Lond) (2007) 21:824–5. doi: 10.1038/sj.eye.6702352
22. Nussenblatt RB, Caspi RR, Mahdi R, Chan CC, Roberge F, Lider O, et al. Inhibition of S-antigen induced experimental autoimmune uveoretinitis by oral induction of tolerance with S-antigen. J Immunol. (1990) 144:1689–95.
23. Thurau SR, Chan CC, Suh E, Nussenblatt RB. Induction of oral tolerance to S-antigen induced experimental autoimmune uveitis by a uveitogenic 20mer peptide. J Autoimmun. (1991) 4:507–16. doi: 10.1016/0896-8411(91)90162-6
24. Dick AD, Cheng YF, Liversidge J, Forrester JV. Intranasal administration of retinal antigens suppresses retinal antigen-induced experimental autoimmune uveoretinitis. Immunology (1994) 82:625–31.
25. Nussenblatt RB, Gery I, Ballintine EJ, Wacker WB. Cellular immune responsiveness of uveitis patients to retinal S-antigen. Am J Ophthalmol. (1980) 89:173–9. doi: 10.1016/0002-9394(80)90108-7
26. Nussenblatt RB, Gery I, Weiner HL, Ferris FL, Shiloach J, Remaley N, et al. Treatment of uveitis by oral administration of retinal antigens: results of a phase I/II randomized masked trial. Am J Ophthalmol. (1997) 123:583–92. doi: 10.1016/S0002-9394(14)71070-0
27. Thurau SR, Wildner G. An HLA-peptide mimics organ-specific antigen in autoimmune uveitis: its role in pathogenesis and therapeutic induction of oral tolerance. Autoimmun Rev. (2003) 2:171–6. doi: 10.1016/S1568-9972(03)00011-9
28. Yeh WI, Seay HR, Newby B, Posgai AL, Moniz FB, Michels A, et al. Avidity and bystander suppressive capacity of human regulatory T cells expressing de novo autoreactive T-cell receptors in type 1 diabetes. Front Immunol. (2017) 8:1313. doi: 10.3389/fimmu.2017.01313
29. Klein L, Kyewski B, Allen PM, Hogquist KA. Positive and negative selection of the T cell repertoire: what thymocytes see (and don't see). Nat Rev Immunol. (2014) 14:377–91. doi: 10.1038/nri3667
30. Richards DM, Kyewski B, Feuerer M. Re-examining the nature and function of self-reactive T cells. Trends Immunol. (2016) 37:114–25. doi: 10.1016/j.it.2015.12.005
31. Lambe T, Leung JC, Ferry H, Bouriez-Jones T, Makinen K, Crockford TL, et al. Limited peripheral T cell anergy predisposes to retinal autoimmunity. J Immunol. (2007) 178:4276–83. doi: 10.4049/jimmunol.178.7.4276
32. Gregerson DS, Heuss ND, Lehmann U, McPherson SW. Peripheral induction of tolerance by retinal antigen expression. J Immunol. (2009) 183:814–22. doi: 10.4049/jimmunol.0803748
33. Gregerson DS, Torseth JW, McPherson SW, Roberts JP, Shinohara T, Zack DJ. Retinal expression of a neo-self antigen, beta-galactosidase, is not tolerogenic and creates a target for autoimmune uveoretinitis. J Immunol. (1999) 163:1073–80.
34. Lai JC, Fukushima A, Wawrousek EF, Lobanoff MC, Charukamnoetkanok P, Smith-Gill SJ, et al. Immunotolerance against a foreign antigen transgenically expressed in the lens. Invest Ophthalmol Vis Sci. (1998) 39:2049–57.
35. Zhang M, Vacchio MS, Vistica BP, Lesage S, Egwuagu CE, Yu CR, et al. T cell tolerance to a neo-self antigen expressed by thymic epithelial cells: the soluble form is more effective than the membrane-bound form. J Immunol. (2003) 170:3954–62. doi: 10.4049/jimmunol.170.8.3954
36. Avichezer D, Grajewski RS, Chan CC, Mattapallil MJ, Silver PB, Raber JA, et al. An immunologically privileged retinal antigen elicits tolerance: major role for central selection mechanisms. J Exp Med. (2003) 198:1665–76. doi: 10.1084/jem.20030413
37. Anderson MS, Venanzi ES, Klein L, Chen Z, Berzins SP, Turley SJ, et al. Projection of an immunological self shadow within the thymus by the aire protein. Science (2002) 298:1395–401. doi: 10.1126/science.1075958
38. DeVoss J, Hou Y, Johannes K, Lu W, Liou GI, Rinn J, et al. Spontaneous autoimmunity prevented by thymic expression of a single self-antigen. J Exp Med. (2006) 203:2727–35. doi: 10.1084/jem.20061864
39. Egwuagu CE, Charukamnoetkanok P, Gery I. Thymic expression of autoantigens correlates with resistance to autoimmune disease. J Immunol. (1997) 159:3109–12.
40. Avichezer D, Liou GI, Chan CC, Lewis GM, Wiggert B, Donoso LA, et al. Interphotoreceptor retinoid-binding protein (IRBP)-deficient C57BL/6 mice have enhanced immunological and immunopathogenic responses to IRBP and an altered recognition of IRBP epitopes. J Autoimmun. (2003) 21:185–94. doi: 10.1016/j.jaut.2003.08.004
41. Takase H, Yu CR, Mahdi RM, Douek DC, Dirusso GB, Midgley FM, et al. Thymic expression of peripheral tissue antigens in humans: a remarkable variability among individuals. Int Immunol. (2005) 17:1131–40. doi: 10.1093/intimm/dxh275
42. Malchow S, Leventhal DS, Lee V, Nishi S, Socci ND, Savage PA. Aire enforces immune tolerance by directing autoreactive T cells into the regulatory T cell lineage. Immunity (2016) 44:1102–13. doi: 10.1016/j.immuni.2016.02.009
43. Silver PB, Agarwal RK, Su SB, Suffia I, Grajewski RS, Luger D, et al. Hydrodynamic vaccination with DNA encoding an immunologically privileged retinal antigen protects from autoimmunity through induction of regulatory T cells. J Immunol. (2007) 179:5146–58. doi: 10.4049/jimmunol.179.8.5146
44. Horai R, Silver PB, Chen J, Agarwal RK, Chong WP, Jittayasothorn Y, et al. Breakdown of immune privilege and spontaneous autoimmunity in mice expressing a transgenic T cell receptor specific for a retinal autoantigen. J Autoimmun. (2013) 44:21–33. doi: 10.1016/j.jaut.2013.06.003
45. Horai R, Zarate-Blades CR, Dillenburg-Pilla P, Chen J, Kielczewski JL, Silver PB, et al. Microbiota-dependent activation of an autoreactive T cell receptor provokes autoimmunity in an immunologically privileged site. Immunity (2015) 43:343–53. doi: 10.1016/j.immuni.2015.07.014
46. Agarwal RK, Kang Y, Zambidis E, Scott DW, Chan CC, Caspi RR. Retroviral gene therapy with an immunoglobulin-antigen fusion construct protects from experimental autoimmune uveitis. J Clin Invest. (2000) 106:245–52. doi: 10.1172/JCI9168
47. Caspi RR. Ocular autoimmunity: the price of privilege? Immunol Rev. (2006) 213:23–35. doi: 10.1111/j.1600-065X.2006.00439.x
48. Horai R, Caspi RR. Chapter 11: Retinal Inflammation: uveitis/uveoretinitis. In: Pang LH, Clark AF editors. Neuromethods: Animal Models of Retinal Disease. Totowa, NJ: Humana Press (2010). p. 207–25.
49. Fujimoto C, Yu CR, Shi G, Vistica BP, Wawrousek EF, Klinman DM, et al. Pertussis toxin is superior to TLR ligands in enhancing pathogenic autoimmunity, targeted at a neo-self antigen, by triggering robust expansion of Th1 cells and their cytokine production. J Immunol. (2006) 177:6896–903. doi: 10.4049/jimmunol.177.10.6896
50. Prendergast RA, Iliff CE, Coskuncan NM, Caspi RR, Sartani G, Tarrant TK, et al. T cell traffic and the inflammatory response in experimental autoimmune uveoretinitis. Invest Ophthalmol Vis Sci. (1998) 39:754–62.
51. Tan C, Wandu WS, Lee RS, Hinshaw SH, Klinman DM, Wawrousek E, et al. Shedding new light on the process of “Licensing” for pathogenicity by th lymphocytes. J Immunol. (2017) 198:681–90. doi: 10.4049/jimmunol.1502108
52. Chen J, Vistica BP, Takase H, Ham DI, Fariss RN, Wawrousek EF, et al. A unique pattern of up- and down-regulation of chemokine receptor CXCR3 on inflammation-inducing Th1 cells. Eur J Immunol. (2004) 34:2885–94. doi: 10.1002/eji.200425318
53. Flugel A, Berkowicz T, Ritter T, Labeur M, Jenne DE, Li Z, et al. Migratory activity and functional changes of green fluorescent effector cells before and during experimental autoimmune encephalomyelitis. Immunity (2001) 14:547–60. doi: 10.1016/S1074-7613(01)00143-1
54. Zhou R, Horai R, Silver PB, Mattapallil MJ, Zarate-Blades CR, Chong WP, et al. The living eye “disarms” uncommitted autoreactive T cells by converting them to Foxp3(+) regulatory cells following local antigen recognition. J Immunol. (2012) 188:1742–50. doi: 10.4049/jimmunol.1102415
55. Sakaguchi S. Regulatory T cells: history and perspective. Methods Mol Biol. (2011) 707:3–17. doi: 10.1007/978-1-61737-979-6_1
56. Sakaguchi S, Ono M, Setoguchi R, Yagi H, Hori S, Fehervari Z, et al. Foxp3+ CD25+ CD4+ natural regulatory T cells in dominant self-tolerance and autoimmune disease. Immunol Rev. (2006) 212:8–27. doi: 10.1111/j.0105-2896.2006.00427.x
57. Grajewski RS, Silver PB, Agarwal RK, Su SB, Chan CC, Liou GI, et al. Endogenous IRBP can be dispensable for generation of natural CD4+CD25+ regulatory T cells that protect from IRBP-induced retinal autoimmunity. J Exp Med. (2006) 203:851–6. doi: 10.1084/jem.20050429
58. McPherson SW, Heuss ND, Pierson MJ, Gregerson DS. Retinal antigen-specific regulatory T cells protect against spontaneous and induced autoimmunity and require local dendritic cells. J Neuroinflammation (2014) 11:205. doi: 10.1186/s12974-014-0205-4
59. Groux H, O'Garra A, Bigler M, Rouleau M, Antonenko S, de Vries JE, et al. A CD4+ T-cell subset inhibits antigen-specific T-cell responses and prevents colitis. Nature (1997) 389:737–42. doi: 10.1038/39614
60. Awasthi A, Carrier Y, Peron JP, Bettelli E, Kamanaka M, Flavell RA, et al. A dominant function for interleukin 27 in generating interleukin 10-producing anti-inflammatory T cells. Nat Immunol. (2007) 8:1380–9. doi: 10.1038/ni1541
61. Pot C, Jin H, Awasthi A, Liu SM, Lai CY, Madan R, et al. Cutting edge: IL-27 induces the transcription factor c-Maf, cytokine IL-21, and the costimulatory receptor ICOS that coordinately act together to promote differentiation of IL-10-producing Tr1 cells. J Immunol. (2009) 183:797–801. doi: 10.4049/jimmunol.0901233
62. Chong WP, van Panhuys N, Chen J, Silver PB, Jittayasothorn Y, Mattapallil MJ, et al. NK-DC crosstalk controls the autopathogenic Th17 response through an innate IFN-gamma-IL-27 axis. J Exp Med. (2015) 212:1739–52. doi: 10.1084/jem.20141678
63. Egwuagu CE, Yu CR. Interleukin 35-Producing B Cells (i35-Breg): a new mediator of regulatory B-cell functions in CNS autoimmune diseases. Crit Rev Immunol. (2015) 35:49–57. doi: 10.1615/CritRevImmunol.2015012558
64. Wang RX, Yu CR, Dambuza IM, Mahdi RM, Dolinska MB, Sergeev YV, et al. Interleukin-35 induces regulatory B cells that suppress autoimmune disease. Nat Med. (2014) 20:633–41. doi: 10.1038/nm.3554
65. Dambuza IM, He C, Choi JK, Yu CR, Wang R, Mattapallil MJ, et al. IL-12p35 induces expansion of IL-10 and IL-35-expressing regulatory B cells and ameliorates autoimmune disease. Nat Commun. (2017) 8:719. doi: 10.1038/s41467-017-00838-4
66. Wei X, Zhang J, Gu Q, Huang M, Zhang W, Guo J, et al. Reciprocal Expression of IL-35 and IL-10 defines two distinct effector treg subsets that are required for maintenance of immune tolerance. Cell Rep. (2017) 21:1853–69. doi: 10.1016/j.celrep.2017.10.090
67. Foussat A, Gregoire S, Clerget-Chossat N, Terrada C, Asnagli H, Lemoine FM, et al. Regulatory T cell therapy for uveitis: a new promising challenge. J Ocul Pharmacol Ther. (2017) 33:278–84. doi: 10.1089/jop.2016.0165
68. Chandran S, Tang Q, Sarwal M, Laszik ZG, Putnam AL, Lee K, et al. Polyclonal regulatory T cell therapy for control of inflammation in kidney transplants. Am J Transplant (2017) 17:2945–54. doi: 10.1111/ajt.14415
69. Gregoire S, Terrada C, Martin GH, Fourcade G, Baeyens A, Marodon G, et al. Treatment of Uveitis by in situ administration of ex vivo-activated polyclonal regulatory T cells. J Immunol. (2016) 196:2109–18. doi: 10.4049/jimmunol.1501723
70. Gilbert RM, Zhang X, Sampson RD, Ehrenstein MR, Nguyen DX, Chaudhry M, et al. Clinical remission of sight-threatening non-infectious uveitis is characterized by an upregulation of peripheral T-regulatory cell polarized towards T-bet and TIGIT. Front Immunol. (2018) 9:907. doi: 10.3389/fimmu.2018.00907
71. Zhuang Z, Wang Y, Zhu G, Gu Y, Mao L, Hong M, et al. Imbalance of Th17/Treg cells in pathogenesis of patients with human leukocyte antigen B27 associated acute anterior uveitis. Sci Rep. (2017) 7:40414. doi: 10.1038/srep40414
72. Tu Z, Li Y, Smith D, Doller C, Sugita S, Chan CC, et al. Myeloid suppressor cells induced by retinal pigment epithelial cells inhibit autoreactive T-cell responses that lead to experimental autoimmune uveitis. Invest Ophthalmol Vis Sci. (2012) 53:959–66. doi: 10.1167/iovs.11-8377
73. Jeong HJ, Lee HJ, Ko JH, Cho BJ, Park SY, Park JW, et al. Myeloid-derived suppressor cells mediate inflammation resolution in humans and mice with autoimmune uveoretinitis. J Immunol. (2018) 200:1306–15. doi: 10.4049/jimmunol.1700617
74. Klaska IP, Muckersie E, Martin-Granados C, Christofi M, Forrester JV. Lipopolysaccharide-primed heterotolerant dendritic cells suppress experimental autoimmune uveoretinitis by multiple mechanisms. Immunology (2017) 150:364–77. doi: 10.1111/imm.12691
75. Ko JH, Lee HJ, Jeong HJ, Kim MK, Wee WR, Yoon SO, et al. Mesenchymal stem/stromal cells precondition lung monocytes/macrophages to produce tolerance against allo- and autoimmunity in the eye. Proc Natl Acad Sci USA. (2016) 113:158–63. doi: 10.1073/pnas.1522905113
Keywords: experimental autoimmune uveitis (EAU), ocular inflammation (uveitis), tolerance process, T-helper (Th) cells, T-regulatory cells (Treg), microbiota
Citation: Gery I and Caspi RR (2018) Tolerance Induction in Relation to the Eye. Front. Immunol. 9:2304. doi: 10.3389/fimmu.2018.02304
Received: 24 May 2018; Accepted: 17 September 2018;
Published: 09 October 2018.
Edited by:
Herman Waldmann, University of Oxford, United KingdomReviewed by:
Bruce Milne Hall, University of New South Wales, AustraliaAndrew David Dick, University College London, United Kingdom
Copyright © 2018 Gery and Caspi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Igal Gery, Z2VyeWlAbmVpLm5paC5nb3Y=