 Roberto Vázquez
Roberto Vázquez Ernesto García
Ernesto García Pedro García
Pedro García- 1Centro de Investigaciones Biológicas (CSIC), Madrid, Spain
- 2Centro de Investigación Biomédica en Red de Enfermedades Respiratorias (CIBERES), Madrid, Spain
Lower respiratory tract infections and tuberculosis are responsible for the death of about 4.5 million people each year and are the main causes of mortality in children under 5 years of age. Streptococcus pneumoniae is the most common bacterial pathogen associated with severe pneumonia, although other Gram-positive and Gram-negative bacteria are involved in respiratory infections as well. The ability of these pathogens to persist and produce infection under the appropriate conditions is also associated with their capacity to form biofilms in the respiratory mucous membranes. Adding to the difficulty of treating biofilm-forming bacteria with antibiotics, many of these strains are becoming multidrug resistant, and thus the alternative therapeutics available for combating this kind of infections are rapidly depleting. Given these concerns, it is urgent to consider other unconventional strategies and, in this regard, phage lysins represent an attractive resource to circumvent some of the current issues in infection treatment. When added exogenously, lysins break specific bonds of the peptidoglycan and have potent bactericidal effects against susceptible bacteria. These enzymes possess interesting features, including that they do not trigger an adverse immune response and raise of resistance is very unlikely. Although Gram-negative bacteria had been considered refractory to these compounds, strategies to overcome this drawback have been developed recently. In this review we describe the most relevant in vitro and in vivo results obtained to date with lysins against bacterial respiratory pathogens.
The Impact of Bacterial Respiratory Diseases on Human Health
Lower respiratory tract infections remain the most deadly communicable diseases, and caused 3.2 million deaths worldwide in 2015 (1). Tuberculosis is still to date among the top 10 death causes, and community-acquired pneumonia is the single largest bacterial infectious cause of death in children worldwide (2). Streptococcus pneumoniae (pneumococcus) accounts for most of the bacterial pneumonia cases in children, followed by Haemophilus influenzae type b, and other bacterial pathogens: Streptococcus pyogenes (group A Streptococcus), non-typeable H. influenzae, Staphylococcus aureus, Mycoplasma pneumoniae, Moraxella catarrhalis, and Klebsiella pneumoniae (3). Pneumococcus is also a common cause of community-acquired pneumonia in elderly patients with comorbidities (4). On the other hand, hospital-acquired pneumonia and ventilator-associated pneumonia are among the leading nosocomial infections worldwide, with an increasing frequency of multidrug resistant (MDR) Gram-negative bacteria (G–) as the bacteriologic cause (5).
Indeed, antimicrobial resistance (AMR) and associated morbidity and mortality have been increasing globally. A recent study estimated that AMR could produce 10 million deaths a year by 2050 (6), although this prediction should be taken with care (7). Accordingly, economic simulations predict that the world will suffer an annual shortfall loss of between $1 and $3.4 trillion by 2030 because of AMR (8). In this scenario, the World Health Organization (WHO) has called for global action on AMR (9). This has encouraged several actions: (a) prevention and control actions in healthcare facilities (10); (b) widespread antimicrobial stewardship programs (11); (c) reduction of antibiotic use in livestock production and the environment (12); and (d) the search for alternatives to the currently used antibiotics (13), particularly against a group of MDR bacteria having a global impact (14). Among these priority pathogens, S. pneumoniae, H. influenzae and those referred to as “the ESKAPE bugs” (15), are of particular concern. Of note, Mycobacterium tuberculosis was not included in the above list as it is already in a globally established priority for which innovative new treatments are urgently needed (16). A few decades ago, phage therapy revived as an alternative to conventional antibiotics and, since the beginning of twenty-first century, phage lytic enzymes have also been extensively tested as antibacterials. This area of research is the focus of this review and the most relevant results of certain enzymes against respiratory pathogens will be discussed. Extensive details on the issue can be found in other recent reviews (17–26).
General Characteristics of Lysins
Endolysins, or more simply lysins, are phage-encoded enzymes capable of hydrolyzing the bacterial cell wall (CW) and that are synthesized at the end of the phage replication cycle. The peptidoglycan (PG) polymer is the basic component of the CW, and is composed of chains of a disaccharide repeat made up of N-acetylglucosamine and N-acetylmuramic acid, linked by β(1 → 4) glycosidic bonds. Glycan strands are cross-linked by tetra/pentapeptide side stems attached to muramic acid residues through amide bonds. Lysins are usually classified as glycosidases [glucosaminidases, transglycosylases, and lysozymes (or muramidases)], if they break any of the bonds of the glycan chain, N-acetylmuramoyl-L-alanine amidases (NAM-amidases), if they break the amide bonds between the glycan strands and peptide chains, or endopeptidases if they hydrolyze different bonds within peptide chains. When purified lysins are added exogenously, their CW-degrading activity can lead to rapid osmotic lysis and bacterial death. The enzymatic activity of lysins was the basis for their exploration as antibacterial agents and they were also named “enzybiotics” (27). Lysins possess several advantages over antibiotics: (a) they rapidly kill bacteria, practically upon contact; (b) they can be specific to the target pathogen, particularly against Gram-positive (G+) bacteria (28–31), which allows to preserve the normal microbiota (32); (c) development of resistance seems very unlikely (33, 34), probably because these enzymes directly target an essential and well-conserved structural component such as the PG, which cannot be easily modified without compromising fitness (35); (d) with few exceptions (36, 37), lysins are active independently of the bacterial physiological state (38, 39); (e) they are effective against MDR bacteria (20, 34, 40–42); (f) they can act synergistically with other lysins or antibiotics and thus theoretically reduce the development of resistance while increasing therapeutic efficiency; and (g) lysins are also effective killing colonizing pathogens growing on mucosal surfaces and/or in biofilms (Tables 1, 2).
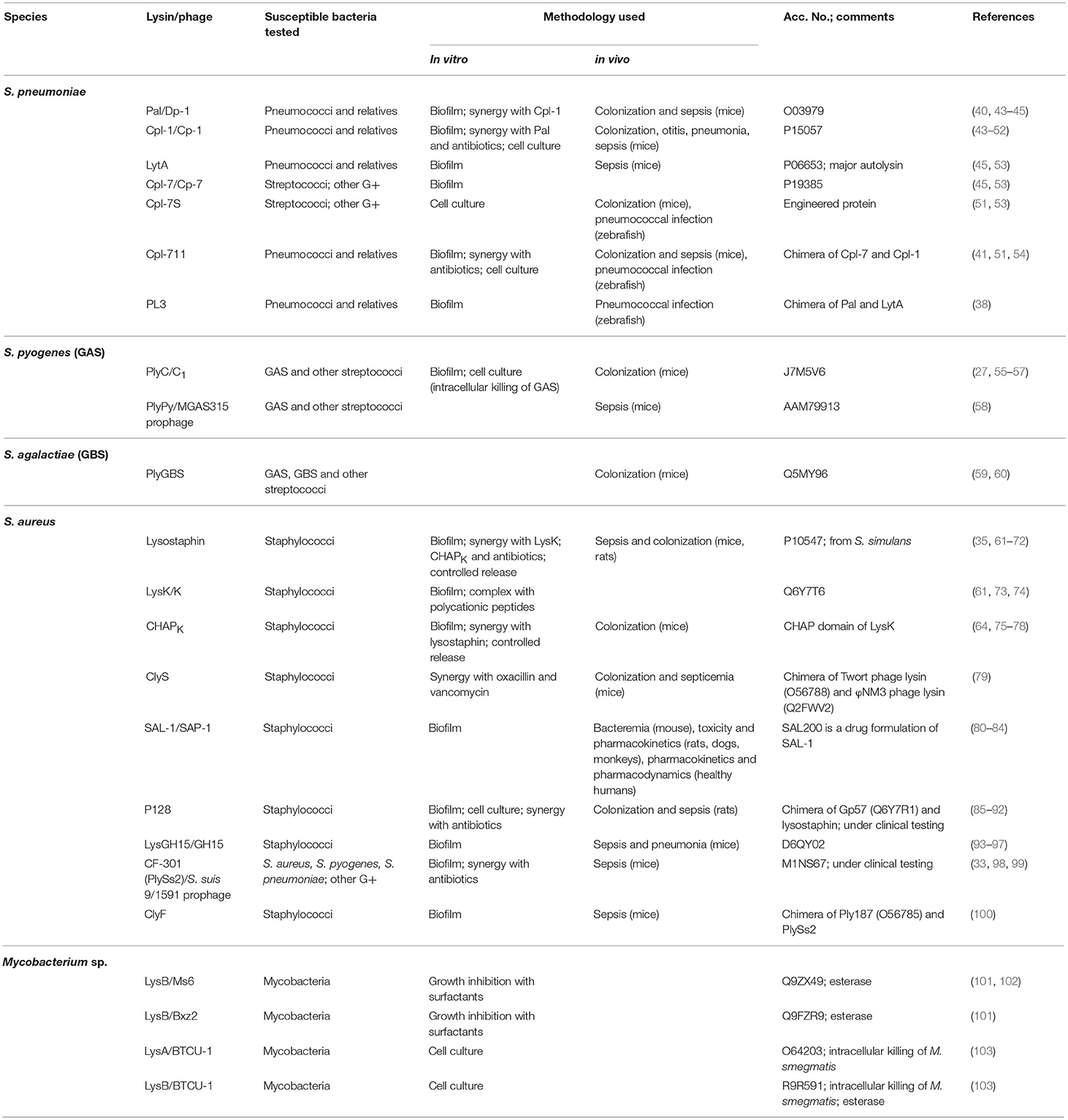
Table 1. Selected lysins active against Gram-positive bacteria and acid-fast mycobacteria.
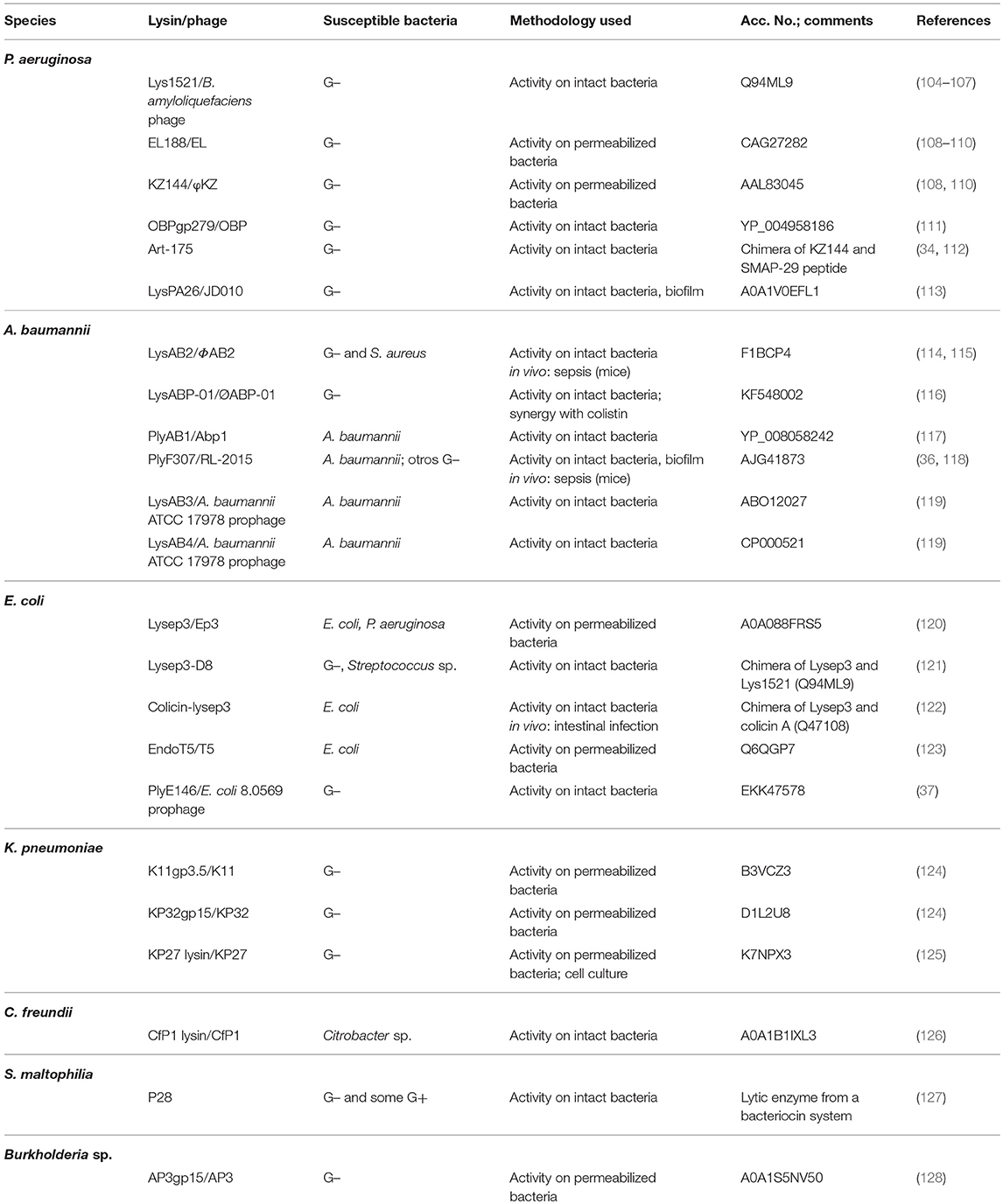
Table 2. Selected lysins active against Gram-negative bacteria.
Lysins encoded by phages infecting G+ bacteria generally display a modular structure, comprising one or more catalytic domains (CDs) and one or more CW binding domains (CWBD). Although the species specificity of a lysin is generally assigned to its CWBD, there are some data suggesting that combined interactions of CD and CWBD with unknown CW receptors may play a significant role (129). On the other hand, phages from G– bacteria usually encode globular lysins with a single CD, with several exceptions (31, 111, 128).
Concerning their systemic, therapeutic use, it has been alleged that lysins, as foreign proteins, could be expected to trigger the production of neutralizing antibodies that might hinder their antibacterial action in subsequent administrations. However, early studies addressing this potential drawback, strongly suggested that highly immune serum slows down—but does not block—lysins (46, 130). Pre-clinical and clinical trials with the antistaphylococcal lysin SAL-1 have been performed in animal models and, lately, in humans. An immune response was indeed elicited after repeated intravenous injections of SAL200, as demonstrated by the presence of specific antibodies and reduced C3 complement levels in the animal blood samples (80). Still, pharmacokinetic, pharmacodynamic, and tolerance studies of SAL200 in monkeys and humans did not show any serious adverse effects or clinically significant alterations even at the highest dose tested (81, 82). Anyhow, host immune responses to specific lysin formulations must always be considered concerning safety and improving the therapeutic potential of lysins.
The antibacterial efficacy of lysins can be improved by several means including: (a) replacement of certain amino acids to modify the net charge of the enzyme (53, 131) or allow dimerization (132); (b) deletion of entire domains (75, 133); (c) construction of chimeric proteins by domain shuffling (41); (d) fusion to cationic peptides (or other domains) to render lysins capable to cross the outer membrane (OM), a widely recognized drawback of lysin therapy against G– bacteria (122, 134, 135), or to increase CW affinity (136); (e) co-administration of lysins with membrane destabilizing agents (EDTA, carvacrol, etc.), especially in G– pathogens (53, 112).
Lysins Against Gram-Positive Bacteria
Streptococcus pneumoniae
The key aspect of the S. pneumoniae system is the role of the aminoalcohol choline in the enzymatic activity of the bacterial autolysin LytA, and the pneumococcal phage lysins. Choline forms part of the (lipo)teichoic acids and constitutes an absolute requirement for the binding of these enzymes—members of the choline-binding family of proteins (CBPs) (137)—to the CW substrate. This peculiarity explains the extreme specificity of CBPs for pneumococci. The first article reporting the use of a CBP as an enzybiotic demonstrated the capacity of the NAM-amidase Pal to kill pneumococci of every serotype tested, including penicillin-resistant isolates (40). These results were confirmed in a mouse model of nasopharyngeal carriage (27). The Cpl-1 lysozyme has also been successfully tested in several in vitro assays and in different animal models of infection (46–48), and a synergistic effect was found when Cpl-1 was used together with several antibiotics (49, 50), or in combination with Pal (43, 44). The Cpl-7 lysozyme represents an exception to choline-recognizing pneumococcal lysins, since it harbors a different CWBD (138–140) that allows it to recognize and kill a broader range of bacteria. Moreover, the bactericidal effect of Cpl-7 has been improved in the engineered Cpl-7S by inverting the net charge of its CWBD (53). To date, the most powerful killing lysins tested against S. pneumoniae are nonetheless chimeric proteins: Cpl-711, a chimera of Cpl-7 and Cpl-1 (41), and PL3, a fusion protein between Pal and LytA [Table 1; (38)]. Treatment with Cpl-711 strongly reduced the attachment of S. pneumoniae to human epithelial cells, and a single intranasal dose of Cpl-711 significantly reduced nasopharyngeal colonization in a mouse model (51).
Staphylococcus aureus
Although S. aureus is frequently carried asymptomatically in humans, it is also the cause of a variety of diseases and, particularly, methicillin-resistant strains (MRSA) are responsible for a great percentage of all infections, up to 80% in some countries (141). The S. aureus PG displays a characteristic pentaglycine interpeptide cross-linking the glycan strands (142). Most tested lysins in the S. aureus system contain two CDs (endopeptidase and NAM-amidase) together with an SH3b CWBD (61, 143, 144). Although the exact interaction between the CWBD and the structures to which these domains bind remains to be demonstrated in many cases, it has been proposed that some CWBDs recognize the pentaglycine peptide cross-bridge (145) or the CW-associated glycopolymers (79). Of note, the vast majority of studies reporting the therapeutic use of lysins are directed to fight S. aureus infections (20, 21). Together with lysostaphin (produced by Staphylococcus simulans), LysK and its derivatives seem to be the most lethal lysins against S. aureus, including MRSA (73, 76, 146, 147) as well as vancomycin-intermediate and -resistant isolates [see reference (21) and references therein]. Other examples of anti-staphylococcal lysins include several engineered proteins such as chimeric or truncated proteins (76, 85, 100, 148, 149) or fusion proteins with short cationic peptides able to cross the eukaryotic membrane and kill intracellular S. aureus (150, 151). Nevertheless, lysin-based studies that consider S. aureus as a respiratory pathogen are scarce and only include some decolonization assays (62, 63, 75, 85) and a single example of endolysin efficacy in a mouse S. aureus pneumonia model (93).
Other Gram-Positive Pathogens and Mycobacteria
S. pyogenes is a major causative agent of upper respiratory tract infections (152). The most relevant example of a lysin targeting this pathogen is PlyC, a peculiar multimeric enzyme that kills group A streptococci with high efficiency (27, 55). In addition, the ability of PlyC to penetrate respiratory tract epithelial cells to eliminate intracellular S. pyogenes cells has also been proven (56). This intracellular activity overcomes one of the major drawbacks of antibiotic therapy against streptococcal throat infections, which is bacterial self-protection by cellular invasion. Other lysins reported to kill S. pyogenes are PlyPy (58) and the broad range, pneumococcal phage-derived Cpl-7S (53). Besides, group B streptococci are known to cause severe pneumonia in newborns (153). At least one attempt has been conducted in mice toward oropharyngeal decolonization of group B streptococci using PlyGBS lysin (59).
The acid-fast M. tuberculosis is still rather unexplored for the development of lysin-based therapy. This might be due to the peculiarity of Mycobacterium CW structure, which comprises a thick PG layer covalently attached to arabinogalactan sterified with mycolic acids (154). Because of this architecture, the lytic cassette of mycobacteriophages comprises two different lytic enzymes: a classical PG hydrolase (usually named LysA) and mycolyl-arabinogalactan esterase (LysB), which cleaves the ester bond linking mycolic acid to the arabinogalactan-PG layer. As a result, the mycolic acid layer detaches from the cell, rendering vulnerable to osmotic shock and, finally, lysis (155). Some in vitro assays have been conducted with both mycobacteriophage-derived hydrolases, yielding, in general, promising results that show either growth arrest (101) or a bactericidal effect (103), but further research is still required. The mycobacterial endolysins and their therapeutical potential have been recently reviewed (156).
Lysins Against Gram-Negative Bacteria
Pseudomonas aeruginosa
The first lysins tested against P. aeruginosa, for example, EL188, only killed bacteria when membrane permeabilizers (e.g., polycationic agents, EDTA) were co-administered (108, 109). Due to the potential difficulties of therapies based on the co-administration of lysins and permeabilizing agents, some of the most recent efforts have been directed toward the engineering of the enzymes themselves, giving rise to the “artilysin” concept (134). In this study, lysins were fused to cationic, antimicrobial peptides (AMPs), and these fusions were able to exert a permeabilizing activity that allowed them to cross P. aeruginosa OM to degrade the PG layer both in vitro and in vivo (134). Art-175 is an artilysin that was constructed by fusing lysin KZ144 and the sheep myeloid AMP 29 (SMAP-29), and further optimizing the thermostability of the resulting chimera by point mutation of several cysteine residues (34). Art-175 was able to efficiently kill either antibiotic-susceptible or MDR P. aeruginosa strains. Of note, Art-175 also controlled the appearance of persisters, i.e., bacterial subpopulations transiently tolerant to antibiotics that often appear upon antiinfective chemotherapy (157).
Despite the engineering efforts mentioned above, lysins able to lyse G– bacteria on their own are also currently available. Typically, this intrinsic activity from without relies on non-enzymatic mechanisms, which were first described for the T4 phage lysozyme (158) and then in several P. aeruginosa phage lysins (159). These lysins harbor AMP-like elements (peptides with an amphipathic secondary structure and a positive net charge) that destabilize the OM. In some cases, as for T4 lysozyme, these regions account for the bactericidal activity of the enzyme to a higher extent than the enzymatic activity itself (158). One of the first examples of a lysin with a natural cationic peptide exploited as an enzybiotic was the Bacillus amyloliquefaciens phage lysin Lys1521, which was indeed able to lyse P. aeruginosa cells (104). Other examples of P. aeruginosa lysins with intrinsic anti-G– activity include OBPgp279 (124) and LysPA26 (113). Although active research is being performed to deal with the OM barrier issue, no extensive in vivo experimental evidence has been provided for the clearance, upon lysin treatment, of P. aeruginosa from respiratory infections.
Acinetobacter baumannii
In general, lysins against G– bacteria appear to be less specific than their G+ counterparts, possibly due to the (apparently) simpler organization of the former sacculi (160). This broader spectrum allows some lysins to kill several pathogenic genera, like the already mentioned lysin LysPA26, which besides P. aeruginosa can also lyse other G– pathogens such as E. coli, K. pneumoniae or A. baumannii (113), or Art-175, which also kills A. baumannii (112). This bacterium is a potential respiratory pathogen (particularly for immunocompromised and debilitated patients) that is receiving great attention in recent years due to its worrisome increased antibiotic resistance (161). Thus, several enzybiotics have also been developed with emphasis in their A. baumannii killing capacity, such as LysAB3 and LysAB4 (119), PlyAB1 (117), and LysABP-01 (116).
PlyF307 was capable of killing A. baumannii isolates, including MDR strains, both in planktonic and biofilm cultures (36) and represents the first example of an intact lysin with intrinsic anti-G– activity tested in a mammalian (mouse bacteremia) model. Unsurprisingly, it was later determined that such intrinsic activity from without partly resided in a cationic peptide located in the C-terminal domain of the lysin (118). Further studies revealed that this region contains sub-domain structural motifs with membrane permeabilizing ability, but lacking enzymatic activity; similar motifs have also been found in other lysins. For example, lysin LysAB2 (114) represents a broad-spectrum enzybiotic, both active against G+ and G– bacteria (A. baumannii, Escherichia coli and, surprisingly, S. aureus). Based on its permeabilizing properties (114), AMPs based on the C-terminal region of LysAB2 have been synthesized and demonstrated high antimicrobial activity when tested in mice infected with A. baumannii (115).
Other Gram-Negative Pathogens
In spite of being a prominent member of the ESKAPE group (162), there are only few reports of lysins active against K. pneumoniae. As already mentioned, LysPA26 also showed bactericidal activity against K. pneumoniae (113). Consequently, it is conceivable that some of the other broad spectrum anti-G– lysins would kill K. pneumoniae. As for specific Klebsiella phage lysins, some examples of lysins with proven lytic activity are those from phages K11, KP32, and KP27 (124, 125, 163), but only KP32 and KP27 were tested for their anti-Klebsiella activity. Although usually associated with intestinal infections, E. coli is also a frequent cause of nosocomial pneumonia (164). Again, some of the other G– lysins are also active against E. coli (105, 113, 114, 116, 124). Specifically from an E. coli phage, Lysep3 lysin has demonstrated noticeable activity against permeabilized E. coli cells (120). Moreover, a chimeric construction between Lysep3 and a colicin was able to traverse the OM via specific recognition by OM transporters (122, 165).
Concluding Remarks and Future Trends
As MDR bacterial respiratory pathogens are increasingly prevalent, alternative therapeutics are urgently needed. Lysins represent more than a hope in this scenario and may be a perfect counterpart to therapies based on standard antibiotics. The potential for lysin development is seemingly endless. For example, thousands of putative lysins, many of which displaying novel domain architectures, have been recently described using bioinformatic techniques (166). All this huge amount of information, together with the crystal structures of lysins and a more detailed knowledge on the bacterial CW structure, will provide better insights to design and construct “tailor-made lysins” potentially directed against any desired pathogen. Drug delivery and other added-value systems involving lysins are now also being researched by setting up different approaches (167–170). Several polymers have been studied as potential drug release vehicles not only for research but also for clinical purposes. Particularly interesting is the case of poly(N-isopropylacrylamide) (PNIPAM) that has been used for the coadministration of the CHAPK lysin and lysostaphin through a thermally triggered release event (the temperature increase due to infection) (64).
Although a limited number of endolysins have entered clinical trials and some of them are already available in the market [reviewed in reference (18)], phages and phage-based products are subjected to strict regulatory measures (171). Moreover, in spite of their demonstrated specificity and lack of resistance development, the use of phage endolysins in humans raises several concerns. Among them, the relatively short plasma life of lysins, their immunogenicity and possible toxicity, the proinflammatory response to bacterial debris, and the difficulties to attack intracellular bacteria have been mentioned. Although only limited data of phage lysin interactions with the human body, e.g., pharmacokinetic/pharmacodynamic studies, have been published, it is encouraging that most (if not all) of the above mentioned potential limitations lack current experimental support (18, 23, 25). Although this scenario seems favorable toward hitting the clinic in the short term, further evidence is still due, especially when bacterial respiratory diseases—in particular, those caused by G– bacteria—are considered. Additional efforts to cover the currently unmet therapeutic requirements are warranted.
Author Contributions
RV, EG, and PG wrote, edited, and approved the final manuscript.
Funding
The authors are supported by a grant from the Ministerio de Economía, Industria y Competitividad (MEICOM) (SAF2017-88664-R). Additional funding was provided by CIBER de Enfermedades Respiratorias (CIBERES), an initiative of the Instituto de Salud Carlos III (ISCIII). RV was the recipient of a predoctoral fellowship from CIBERES.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge support of the publication fee by the CSIC Open Access Publication Support Initiative through its Unit of Information Resources for Research (URICI).
References
1. World Health Organization. The Top 10 Causes of Death (2017). Available online at: http://www.who.int/mediacentre/factsheets/fs310/en/ (Accessed April 18, 2018).
2. World Health Organization. Pneumonia (2016). Available online at: http://www.who.int/mediacentre/factsheets/fs331/en (Accessed April 24, 2018).
3. Rodrigues CMC, Groves H. Community-acquired pneumonia in children: the challenges of microbiological diagnosis. J Clin Microbiol. (2018) 56:e01318–17. doi: 10.1128/JCM.01318-17
4. Torres A, Cillóniz C, Blasi F, Chalmers JD, Gaillat J, Dartois N, et al. Burden of pneumococcal community-acquired pneumonia in adults across Europe: a literature review. Respir Med. (2018) 137:6–13. doi: 10.1016/j.rmed.2018.02.007
5. Kidd JM, Kuti JL, Nicolau DP. Novel pharmacotherapy for the treatment of hospital-acquired and ventilator-associated pneumonia caused by resistant Gram-negative bacteria. Expert Opin Pharmacother. (2018) 19:397–408. doi: 10.1080/14656566.2018.1438408
6. O'Neill J. Review on Antimicrobial Resistance. Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations (2014). Available online at: https://amr-review.org/sites/default/files/AMR%20Review%20Paper%20-%20Tackling%20a%20crisis%20for%20the%20health%20and%20wealth%20of%20nations_1.pdf. (Accessed April 24, 2018).
7. de Kraker MEA, Stewardson AJ, Harbarth S. Will 10 million people die a year due to antimicrobial resistance by 2050? PLoS Med. (2016) 13:e1002184. doi: 10.1371/journal.pmed.1002184
8. World Bank. Drug-Resistant Infections: A Threat to our Economic Future (2017). Available online at: http://www.worldbank.org/en/topic/health/publication/drug-resistant-infections-a-threat-to-our-economic-future (Accessed September 24, 2018).
9. World Health Organization. Antimicrobial Resistance (2018). Available online at: http://www.who.int/en/news-room/fact-sheets/detail/antimicrobial-resistance (Accessed April 25, 2018).
10. Pires D, de Kraker MEA, Tartari E, Abbas M, Pittet D. 'Fight antibiotic resistance—It's in your hands': call from the World Health Organization for 5th May 2017. Clin Infect Dis. (2017) 64:1780–3. doi: 10.1093/cid/cix226
11. Pulcini C. Antibiotic stewardship: a European perspective. FEMS Microbiol Lett. (2017) 364, fnx230. doi: 10.1093/femsle/fnx230
12. Holmes AH, Moore LSP, Sundsfjord A, Steinbakk M, Regmi S, Karkey A, et al. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet (2016) 387:176–87. doi: 10.1016/S0140-6736(15)00473-0
13. Tacconelli E, Sifakis F, Harbarth S, Schrijver R, van Mourik M, Voss A, et al. Surveillance for control of antimicrobial resistance. Lancet Infect Dis. (2018) 18:E99–106. doi: 10.1016/S1473-3099(17)30485-1
14. World Health Organization. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics (2017). Available online at: http://www.who.int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf (Accessed April, 26 2018).
15. Rice LB. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J Infect Dis. (2008) 197:1079–81. doi: 10.1086/533452
16. World Health Organization. Global Tuberculosis Report 2018 (2018). Available online at: http://www.who.int/tb/publications/global_report/en/ (Accessed September 24, 2018).
17. Cooper CJ, Koonjan S, Nilsson AS. Enhancing whole phage therapy and their derived antimicrobial enzymes through complex formulation. Pharmaceuticals (2018) 11:34. doi: 10.3390/ph11020034
18. Gerstmans H, Criel B, Briers Y. Synthetic biology of modular endolysins. Biotechnol Adv. (2018) 36:624–40. doi: 10.1016/j.biotechadv.2017.12.009
19. Gu J, Xi H, Cheng M, Han W. Phage-derived lysins as therapeutic agents against multidrug-resistant Enterococcus faecalis. Future Microbiol. (2018) 13:275–8. doi: 10.2217/fmb-2017-0235
20. Gutiérrez D, Fernández L, Rodríguez A, García P. Are phage lytic proteins the secret weapon to kill Staphylococcus aureus? mBio (2018) 9:e01923–17. doi: 10.1128/mBio.01923-17
21. Haddad Kashani H, Schmelcher M, Sabzalipoor H, Seyed Hosseini E, Moniri R. Recombinant endolysins as potential therapeutics against antibiotic-resistant Staphylococcus aureus: current status of research and novel delivery strategies. Clin Microbiol Rev. (2018) 31:e00071–17. doi: 10.1128/CMR.00071-17
22. Love MJ, Bhandari D, Dobson RCJ, Billington C. Potential for bacteriophage endolysins to supplement or replace antibiotics in food production and clinical care. Antibiotics (2018) 7:17. doi: 10.3390/antibiotics7010017
23. Oliveira H, São-José C, Azeredo J. Phage-derived peptidoglycan degrading enzymes: challenges and future prospects for in vivo therapy. Viruses (2018) 10:292. doi: 10.3390/v10060292
24. São-José C. Engineering of phage-derived lytic enzymes: improving their potential as antimicrobials. Antibiotics (2018) 7:29. doi: 10.3390/antibiotics7020029
25. Sharma U, Vipra A, Channabasappa S. Phage-derived lysins as potential agents for eradicating biofilms and persisters. Drug Discov Today (2018) 23:848–56. doi: 10.1016/j.drudis.2018.01.026
26. Fischetti V. Development of phage lysins as novel therapeutics: a historical perspective. Viruses (2018) 10:310. doi: 10.3390/v10060310
27. Nelson D, Loomis L, Fischetti VA. Prevention and elimination of upper respiratory colonization of mice by group A streptococci by using a bacteriophage lytic enzyme. Proc Natl Acad Sci USA (2001) 98:4107–12. doi: 10.1073/pnas.061038398
28. López R, García E, García P, García JL. The pneumococcal cell wall degrading enzymes: a modular design to create new lysins? Microb Drug Resist. (1997) 3:199–211. doi: 10.1089/mdr.1997.3.199
29. Loessner MJ, Kramer K, Ebel F, Scherer S. C-terminal domains of Listeria monocytogenes bacteriophage murein hydrolases determine specific recognition and high-affinity binding to bacterial cell wall carbohydrates. Mol Microbiol. (2002) 44:335–49. doi: 10.1046/j.1365-2958.2002.02889.x
30. Schuch R, Nelson D, Fischetti VA. A bacteriolytic agent that detects and kills Bacillus anthracis. Nature (2002) 418:884–9. doi: 10.1038/nature01026
31. Briers Y, Schmelcher M, Loessner MJ, Hendrix J, Engelborghs Y, Volckaert G, et al. The high-affinity peptidoglycan binding domain of Pseudomonas phage endolysin KZ144. Biochem Biophys Res Commun. (2009) 383:187–91. doi: 10.1016/j.bbrc.2009.03.161
32. Cheng M, Zhang Y, Li X, Liang J, Hu L, Gong P, et al. Endolysin LysEF-P10 shows potential as an alternative treatment strategy for multidrug-resistant Enterococcus faecalis infections. Sci Rep. (2017) 7:10164. doi: 10.1038/s41598-017-10755-7
33. Gilmer DB, Schmitz JE, Euler CW, Fischetti VA. Novel bacteriophage lysin with broad lytic activity protects against mixed infection by Streptococcus pyogenes and methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother. (2013) 57:2743–50. doi: 10.1128/AAC.02526-12
34. Briers Y, Walmagh M, Grymonprez B, Biebl M, Pirnay J-P, Defraine V, et al. Art-175 is a highly efficient antibacterial against multidrug-resistant strains and persisters of Pseudomonas aeruginosa. Antimicrob Agents Chemother. (2014) 58:3774–84. doi: 10.1128/AAC.02668-14
35. Kusuma C, Jadanova A, Chanturiya T, Kokai-Kun JF. Lysostaphin-resistant variants of Staphylococcus aureus demonstrate reduced fitness in vitro and in vivo. Antimicrob Agents Chemother. (2007) 51:475–82. doi: 10.1128/AAC.00786-06
36. Lood R, Winer BY, Pelzek AJ, Diez-Martinez R, Thandar M, Euler CW, et al. Novel phage lysin capable of killing the multidrug-resistant gram-negative bacterium Acinetobacter baumannii in a mouse bacteremia model. Antimicrob Agents Chemother. (2015) 59:1983–91. doi: 10.1128/AAC.04641-14
37. Larpin Y, Oechslin F, Moreillon P, Resch G, Entenza JM, Mancini S. In vitro characterization of PlyE146, a novel phage lysin that targets Gram-negative bacteria. PLoS ONE (2018) 13:e0192507. doi: 10.1371/journal.pone.0192507
38. Blázquez B, Fresco-Taboada A, Iglesias-Bexiga M, Menéndez M, García P. PL3 amidase, a tailor-made lysin constructed by domain shuffling with potent killing activity against pneumococci and related species. Front Microbiol. (2016) 7:1156. doi: 10.3389/fmicb.2016.01156
39. Vázquez R, Domenech M, Iglesias-Bexiga M, Menéndez M, García P. Csl2, a novel chimeric bacteriophage lysin to fight infections caused by Streptococcus suis, an emerging zoonotic pathogen. Sci Rep. (2017) 7:16506. doi: 10.1038/s41598-017-16736-0
40. Loeffler JM, Nelson D, Fischetti VA. Rapid killing of Streptococcus pneumoniae with a bacteriophage cell wall hydrolase. Science (2001) 294:2170–2. doi: 10.1126/science.1066869
41. Díez-Martínez R, De Paz HD, García-Fernández E, Bustamante N, Euler CW, Fischetti VA, et al. A novel chimeric phage lysin with high in vitro and in vivo bactericidal activity against Streptococcus pneumoniae. J Antimicrob Chemother. (2015) 70:1763–73. doi: 10.1093/jac/dkv038
42. Yang H, Wang M, Yu J, Wei H. Antibacterial activity of a novel peptide-modified lysin against Acinetobacter baumannii and Pseudomonas aeruginosa. Frontiers Microbiol. (2015) 6:1471. doi: 10.3389/fmicb.2015.01471
43. Loeffler JM, Fischetti VA. Synergistic lethal effect of a combination of phage lytic enzymes with different activities on penicillin-sensitive and -resistant Streptococcus pneumoniae strains. Antimicrob Agents Chemother. (2003) 47:375–7. doi: 10.1128/AAC.47.1.375-377.2003
44. Jado I, López R, García E, Fenoll A, Casal J, García P, et al. Phage lytic enzymes as therapy of antibiotic-resistant Streptococcus pneumoniae infection in a murine sepsis model. J Antimicrob Chemother. (2003) 52:967–73. doi: 10.1093/jac/dkg485
45. Domenech M, García E, Moscoso M. In vitro destruction of Streptococcus pneumoniae biofilms with bacterial and phage peptidoglycan hydrolases. Antimicrob Agents Chemother. (2011) 55:4144–8. doi: 10.1128/AAC.00492-11
46. Loeffler JM, Djurkovic S, Fischetti VA. Phage lytic enzyme Cpl-1 as a novel antimicrobial for pneumococcal bacteremia. Infect Immun. (2003) 71:6199–204. doi: 10.1128/IAI.71.11.6199-6204.2003
47. McCullers JA, Karlström Å, Iverson AR, Loeffler JM, Fischetti VA. Novel strategy to prevent otitis media caused by colonizing Streptococcus pneumoniae. PLoS Pathog. (2007) 3:e28. doi: 10.1371/journal.ppat.0030028
48. Doehn JM, Fischer K, Reppe K, Gutbier B, Tschernig T, Hocke AC, et al. Delivery of the endolysin Cpl-1 by inhalation rescues mice with fatal pneumococcal pneumonia. J Antimicrob Chemother. (2013) 68:2111–7. doi: 10.1093/jac/dkt131
49. Vouillamoz J, Entenza JM, Giddey M, Fischetti VA, Moreillon P, Resch G. Bactericidal synergism between daptomycin and the phage lysin Cpl-1 in a mouse model of pneumococcal bacteraemia. Int J Antimicrob Agents (2013) 42:416–21. doi: 10.1016/j.ijantimicag.2013.06.020
50. Djurkovic S, Loeffler JM, Fischetti VA. Synergistic killing of Streptococcus pneumoniae with the bacteriophage lytic enzyme Cpl-1 and penicillin or gentamicin depends on the level of penicillin resistance. Antimicrob Agents Chemother. (2005) 49:1225–8. doi: 10.1128/AAC.49.3.1225-1228.2005
51. Corsini B, Díez-Martínez R, Aguinagalde L, González-Camacho F, García-Fernández E, Letrado P, et al. Chemotherapy with phage lysins reduces pneumococcal colonization of the respiratory tract. Antimicrob Agents Chemother. (2018) 62:e02212–17. doi: 10.1128/AAC.02212-17
52. Witzenrath M, Schmeck B, Doehn JM, Tschernig T, Zahlten J, Loeffler JM, et al. Systemic use of the endolysin Cpl-1 rescues mice with fatal pneumococcal pneumonia. Crit Care Med. (2009) 37:642–9. doi: 10.1097/CCM.0b013e31819586a6
53. Díez-Martínez R, de Paz HD, Bustamante N, García E, Menéndez M, García P. Improving the lethal effect of Cpl-7, a pneumococcal phage lysozyme with broad bactericidal activity, by inverting the net charge of its cell wall-binding module. Antimicrob Agents Chemother. (2013) 57:5355–65. doi: 10.1128/AAC.01372-13
54. Letrado P, Corsini B, Díez-Martínez R, Bustamante N, Yuste JE, García P. Bactericidal synergism between antibiotics and phage endolysin Cpl-711 to kill multidrug-resistant pneumococcus. Future Microbiol. (2018) (in press).
55. Nelson D, Schuch R, Chahales P, Zhu S, Fischetti VA. PlyC: a multimeric bacteriophage lysin. Proc Natl Acad Sci USA (2006) 103:10765–70. doi: 10.1073/pnas.0604521103
56. Shen Y, Barros M, Vennemann T, Gallagher DT, Yin Y, Linden SB, et al. A bacteriophage endolysin that eliminates intracellular streptococci. eLife (2016) 5:e13152. doi: 10.7554/eLife.13152
57. Shen Y, Koller T, Kreikemeyer B, Nelson DC. Rapid degradation of Streptococcus pyogenes biofilms by PlyC, a bacteriophage-encoded endolysin. J Antimicrob Chemother. (2013) 68:1818–24. doi: 10.1093/jac/dkt104
58. Lood R, Raz A, Molina H, Euler CW, Fischetti VA. A highly active and negatively charged Streptococcus pyogenes lysin with a rare D-alanyl-L-alanine endopeptidase activity protects mice against streptococcal bacteremia. Antimicrob Agents Chemother. (2014) 58:3073–84. doi: 10.1128/AAC.00115-14
59. Cheng Q, Nelson D, Zhu S, Fischetti VA. Removal of group B streptococci colonizing the vagina and oropharynx of mice with a bacteriophage lytic enzyme. Antimicrob Agents Chemother. (2005) 49:111–7. doi: 10.1128/AAC.49.1.111-117.2005
60. Cheng Q, Fischetti VA. Mutagenesis of a bacteriophage lytic enzyme PlyGBS significantly increases its antibacterial activity against group B streptococci. Appl Microbiol Biotechnol. (2007) 74:1284–91. doi: 10.1007/s00253-006-0771-1
61. Schmelcher M, Shen Y, Nelson DC, Eugster MR, Eichenseher F, Hanke DC, et al. Evolutionarily distinct bacteriophage endolysins featuring conserved peptidoglycan cleavage sites protect mice from MRSA infection. J Antimicrob Chemother. (2015) 70:1453–65. doi: 10.1093/jac/dku552
62. Kokai-Kun JF, Walsh SM, Chanturiya T, Mond JJ. Lysostaphin cream eradicates Staphylococcus aureus nasal colonization in a cotton rat model. Antimicrob Agents Chemother. (2003) 47:1589–97. doi: 10.1128/AAC.47.5.1589-1597.2003
63. Walsh S, Kokai-Kun J, Shah A, Mond J. Extended nasal residence time of lysostaphin and an anti-staphylococcal monoclonal antibody by delivery in semisolid or polymeric carriers. Pharm Res. (2004) 21:1770–5. doi: 10.1023/B:PHAM.0000045227.16829.37
64. Hathaway H, Ajuebor J, Stephens L, Coffey A, Potter U, Sutton JM, et al. Thermally triggered release of the bacteriophage endolysin CHAPK and the bacteriocin lysostaphin for the control of methicillin resistant Staphylococcus aureus (MRSA). J Control Release (2017) 245:108–15. doi: 10.1016/j.jconrel.2016.11.030
65. Polak J, Della Latta P, Blackburn P. In vitro activity of recombinant lysostaphin-antibiotic combinations toward methicillin-resistant Staphylococcus aureus. Diagn Microbiol Infect Dis. (1993) 17:265–70. doi: 10.1016/0732-8893(93)90034-5
66. Climo MW, Ehlert K, Archer GL. Mechanism and suppression of lysostaphin resistance in oxacillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother. (2001) 45:1431–7. doi: 10.1128/AAC.45.5.1431-1437.2001
67. Kiri N, Archer G, Climo MW. Combinations of lysostaphin with β-lactams are synergistic against oxacillin-resistant Staphylococcus epidermidis. Antimicrob Agents Chemother. (2002) 46:2017–20. doi: 10.1128/AAC.46.6.2017-2020.2002
68. Wu JA, Kusuma C, Mond JJ, Kokai-Kun JF. Lysostaphin disrupts Staphylococcus aureus and Staphylococcus epidermidis biofilms on artificial surfaces. Antimicrob Agents Chemother. (2003) 47:3407–14. doi: 10.1128/AAC.47.11.3407-3414.2003
69. Walsh S, Shah A, Mond J. Improved pharmacokinetics and reduced antibody reactivity of lysostaphin conjugated to polyethylene glycol. Antimicrob Agents Chemother. (2003) 47:554–8. doi: 10.1128/AAC.47.2.554-558.2003
70. Kokai-Kun JF, Chanturiya T, Mond JJ. Lysostaphin as a treatment for systemic Staphylococcus aureus infection in a mouse model. J Antimicrob Chemother. (2007) 60:1051–9. doi: 10.1093/jac/dkm347
71. Becker SC, Foster-Frey J, Donovan DM. The phage K lytic enzyme LysK and lysostaphin act synergistically to kill MRSA. FEMS Microbiol Lett. (2008) 287:185–91. doi: 10.1111/j.1574-6968.2008.01308.x
72. Aguinaga A, Francés ML, Del Pozo JL, Alonso M, Serrera A, Lasa I, et al. Lysostaphin and clarithromycin: a promising combination for the eradication of Staphylococcus aureus biofilms. Int J Antimicrob Agents (2011) 37:585–7. doi: 10.1016/j.ijantimicag.2011.02.009
73. O'Flaherty S, Coffey A, Meaney W, Fitzgerald GF, Ross RP. The recombinant phage lysin LysK has a broad spectrum of lytic activity against clinically relevant staphylococci, including methicillin-resistant Staphylococcus aureus. J Bacteriol. (2005) 187:7161–4. doi: 10.1128/JB.187.20.7161-7164.2005
74. Filatova LY, Donovan DM, Becker SC, Lebedev DN, Priyma AD, Koudriachova HV, et al. Physicochemical characterization of the staphylolytic LysK enzyme in complexes with polycationic polymers as a potent antimicrobial. Biochimie (2013) 95:1689–96. doi: 10.1016/j.biochi.2013.04.013
75. Fenton M, Casey PG, Hill C, Gahan CGM, Ross RP, McAuliffe O, et al. The truncated phage lysin CHAPk eliminates Staphylococcus aureus in the nares of mice. Bioeng Bugs (2010) 1:404–7. doi: 10.4161/bbug.1.6.13422
76. Horgan M, O'Flynn G, Garry J, Cooney J, Coffey A, Fitzgerald GF, et al. Phage lysin LysK can be truncated to its CHAP domain and retain lytic activity against live antibiotic-resistant staphylococci. Appl Environ Microbiol. (2009) 75:872–4. doi: 10.1128/AEM.01831-08
77. Fenton M, Ross RP, McAuliffe O, O'Mahony J, Coffey A. Characterization of the staphylococcal bacteriophage lysin CHAPK. J Appl Microbiol. (2011) 111:1025–35. doi: 10.1111/j.1365-2672.2011.05119.x
78. Fenton M, Keary R, McAuliffe O, Ross RP, O'Mahony J, Coffey A. Bacteriophage-derived peptidase CHAPK eliminates and prevents staphylococcal biofilms. Int J Microbiol. (2013) 2013:625341. doi: 10.1155/2013/625341
79. Daniel A, Euler C, Collin M, Chahales P, Gorelick KJ, Fischetti VA. Synergism between a novel chimeric lysin and oxacillin protects against infection by methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother. (2010) 54:1603–12. doi: 10.1128/AAC.01625-09
80. Jun SY, Jung GM, Yoon SJ, Choi Y-J, Koh WS, Moon KS, et al. Preclinical safety evaluation of intravenously administered SAL200 containing the recombinant phage endolysin SAL-1 as a pharmaceutical ingredient. Antimicrob Agents Chemother. (2014) 58:2084–8. doi: 10.1128/AAC.02232-13
81. Jun SY, Jung GM, Yoon SJ, Youm SY, Han H-Y, Lee J-H, et al. Pharmacokinetics of the phage endolysin-based candidate drug SAL200 in monkeys and its appropriate intravenous dosing period. Clin Exp Pharmacol Physiol. (2016) 43:1013–6. doi: 10.1111/1440-1681.12613
82. Jun SY, Jang IJ, Yoon S, Jang K, Yu K-S, Cho JY, et al. Pharmacokinetics and tolerance of the phage endolysin-based candidate drug SAL200 after a single intravenous administration among healthy volunteers. Antimicrob Agents Chemother. (2017) 61:e02629–16. doi: 10.1128/AAC.02629-16
83. Jun SY, Jung GM, Son J-S, Yoon SJ, Choi Y-J, Kang SH. Comparison of the antibacterial properties of phage endolysins SAL-1 and LysK. Antimicrob Agents Chemother. (2011) 55:1764–7. doi: 10.1128/AAC.01097-10
84. Jun SY, Jung GM, Yoon SJ, Oh M-D, Choi Y-J, Lee WJ, et al. Antibacterial properties of a pre-formulated recombinant phage endolysin, SAL-1. Int J Antimicrob Agents (2013) 41:156–61. doi: 10.1016/j.ijantimicag.2012.10.011
85. Paul VD, Rajagopalan SS, Sundarrajan S, George SE, Asrani JY, Pillai R, et al. A novel bacteriophage Tail-Associated Muralytic Enzyme (TAME) from phage K and its development into a potent antistaphylococcal protein. BMC Microbiol. (2011) 11:226. doi: 10.1186/1471-2180-11-226
86. Vipra AA, Desai SN, Roy P, Patil R, Raj JM, Narasimhaswamy N, et al. Antistaphylococcal activity of bacteriophage derived chimeric protein P128. BMC Microbiol. (2012) 12:41. doi: 10.1186/1471-2180-12-41
87. George SE, Chikkamadaiah R, Durgaiah M, Joshi AA, Thankappan UP, Madhusudhana SN, et al. Biochemical characterization and evaluation of cytotoxicity of antistaphylococcal chimeric protein P128. BMC Res Notes (2012) 5:280. doi: 10.1186/1756-0500-5-280
88. Drilling AJ, Cooksley C, Chan C, Wormald PJ, Vreugde S. Fighting sinus-derived Staphylococcus aureus biofilms in vitro with a bacteriophage-derived muralytic enzyme. Int Forum Allergy Rhinol. (2016) 6:349–55. doi: 10.1002/alr.21680
89. Poonacha N, Nair S, Desai S, Tuppad D, Hiremath D, Mohan T, et al. Efficient killing of planktonic and biofilm-embedded coagulase-negative staphylococci by bactericidal protein P128. Antimicrob Agents Chemother. (2017) 61:e00457–17. doi: 10.1128/AAC.00457-17
90. Nair S, Desai S, Poonacha N, Vipra A, Sharma U. Antibiofilm activity and synergistic inhibition of Staphylococcus aureus biofilms by bactericidal protein P128 in combination with antibiotics. Antimicrob Agents Chemother. (2016) 60:7280–9. doi: 10.1128/AAC.01118-16
91. Nair S, Poonacha N, Desai S, Hiremath D, Tuppad D, Mohan T, et al. Restoration of sensitivity of a diverse set of drug-resistant Staphylococcus clinical strains by bactericidal protein P128. J Med Microbiol. (2018) 67:296–307. doi: 10.1099/jmm.0.000697
92. Channabasappa S, Durgaiah M, Chikkamadaiah R, Kumar S, Joshi A, Sriram B. Efficacy of novel antistaphylococcal ectolysin P128 in a rat model of methicillin-resistant Staphylococcus aureus bacteremia. Antimicrob Agents Chemother. (2018) 62:e01358–17. doi: 10.1128/AAC.01358-17
93. Xia F, Li X, Wang B, Gong P, Xiao F, Yang M, et al. Combination therapy of LysGH15 and apigenin as a new strategy for treating pneumonia caused by Staphylococcus aureus. Appl Environ Microbiol. (2016) 82:87–94. doi: 10.1128/AEM.02581-15
94. Gu J, Xu W, Lei L, Huang J, Feng X, Sun C, et al. LysGH15, a novel bacteriophage lysin, protects a murine bacteremia model efficiently against lethal methicillin-resistant Staphylococcus aureus infection. J Clin Microbiol. (2011) 49:111–7. doi: 10.1128/JCM.01144-10
95. Gu J, Zuo J, Lei L, Zhao H, Sun C, Feng X, et al. LysGH15 reduces the inflammation caused by lethal methicillin-resistant Staphylococcus aureus infection in mice. Bioeng Bugs (2011) 2:96–9. doi: 10.4161/bbug.2.2.14883
96. Zhang L, Li D, Li X, Hu L, Cheng M, Xia F, et al. LysGH15 kills Staphylococcus aureus without being affected by the humoral immune response or inducing inflammation. Sci Rep. (2016) 6:29344. doi: 10.1038/srep29344
97. Zhang Y, Cheng M, Zhang H, Dai J, Guo Z, Li X, et al. Antibacterial effects of phage lysin LysGH15 on planktonic cells and biofilms of diverse staphylococci. Appl Environ Microbiol. (2018) 84:e00886–18. doi: 10.1128/AEM.00886-18
98. Schuch R, Lee HM, Schneider BC, Sauve KL, Law C, Khan BK, et al. Combination therapy with lysin CF-301 and antibiotic is superior to antibiotic alone for treating methicillin-resistant Staphylococcus aureus-induced murine bacteremia. J Infect Dis. (2014) 209:1469–78. doi: 10.1093/infdis/jit637
99. Schuch R, Khan BK, Raz A, Rotolo JA, Wittekind M. Bacteriophage lysin CF-301, a potent antistaphylococcal biofilm agent. Antimicrob Agents Chemother. (2017) 61:e02666–16. doi: 10.1128/AAC.02666-16
100. Yang H, Zhang H, Wang J, Yu J, Wei H. A novel chimeric lysin with robust antibacterial activity against planktonic and biofilm methicillin-resistant Staphylococcus aureus. Sci Rep. (2017) 7:40182. doi: 10.1038/srep40182
101. Grover N, Paskaleva EE, Mehta KK, Dordick JS, Kane RS. Growth inhibition of Mycobacterium smegmatis by mycobacteriophage-derived enzymes. Enzyme Microb Technol. (2014) 63:1–6. doi: 10.1016/j.enzmictec.2014.04.018
102. Gil F, Grzegorzewicz AE, Catalão MJ, Vital J, McNeil MR, Pimentel M. Mycobacteriophage Ms6 LysB specifically targets the outer membrane of Mycobacterium smegmatis. Microbiology (2010) 156:1497–504. doi: 10.1099/mic.0.032821-0
103. Lai M-J, Liu C-C, Jiang S-J, Soo P-C, Tu M-H, Lee J-J, et al. Antimycobacterial activities of endolysins derived from a mycobacteriophage, BTCU-1. Molecules (2015) 20:19277–90. doi: 10.3390/molecules201019277
104. Morita M, Tanji Y, Mizoguchi K, Soejima A, Orito Y, Unno H. Antibacterial activity of Bacillus amyloliquefaciens phage endolysin without holin conjugation. J Biosci Bioeng. (2001) 91:469–73. doi: 10.1016/S1389-1723(01)80275-9
105. Morita M, Tanji Y, Orito Y, Mizoguchi K, Soejima A, Unno H. Functional analysis of antibacterial activity of Bacillus amyloliquefaciens phage endolysin against Gram-negative bacteria. FEBS Lett. (2001) 500:56–9. doi: 10.1016/S0014-5793(01)02587-X
106. Orito Y, Morita M, Hori K, Unno H, Tanji Y. Bacillus amyloliquefaciens phage endolysin can enhance permeability of Pseudomonas aeruginosa outer membrane and induce cell lysis. Appl Microbiol Biotechnol. (2004) 65:105–9. doi: 10.1007/s00253-003-1522-1
107. Muyombwe A, Tanji Y, Unno H. Cloning and expression of a gene encoding the lytic functions of Bacillus amyloliquefaciens phage: evidence of an auxiliary lysis system. J Biosci Bioeng. (1999) 88:221–5. doi: 10.1016/S1389-1723(99)80206-0
108. Briers Y, Volckaert G, Cornelissen A, Lagaert S, Michiels CW, Hertveldt K, et al. Muralytic activity and modular structure of the endolysins of Pseudomonas aeruginosa bacteriophages φKZ and EL. Mol Microbiol. (2007) 65:1334–44. doi: 10.1111/j.1365-2958.2007.05870.x
109. Briers Y, Walmagh M, Lavigne R. Use of bacteriophage endolysin EL188 and outer membrane permeabilizers against Pseudomonas aeruginosa. J Appl Microbiol. (2011) 110:778–85. doi: 10.1111/j.1365-2672.2010.04931.x
110. Briers Y, Cornelissen A, Aertsen A, Hertveldt K, Michiels CW, Volckaert G, et al. Analysis of outer membrane permeability of Pseudomonas aeruginosa and bactericidal activity of endolysins KZ144 and EL188 under high hydrostatic pressure. FEMS Microbiol Lett. (2008) 280:113–9. doi: 10.1111/j.1574-6968.2007.01051.x
111. Walmagh M, Briers Y, dos Santos SB, Azeredo J, Lavigne R. Characterization of modular bacteriophage endolysins from Myoviridae phages OBP, 201j2-1 and PVP-SE1. PLoS ONE (2012) 7:e36991. doi: 10.1371/journal.pone.0036991
112. Defraine V, Schuermans J, Grymonprez B, Govers SK, Aertsen A, Fauvart M, et al. Efficacy of artilysin Art-175 against resistant and persistent Acinetobacter baumannii. Antimicrob Agents Chemother. (2016) 60:3480–8. doi: 10.1128/AAC.00285-16
113. Guo M, Feng C, Ren J, Zhuang X, Zhang Y, Zhu Y, et al. A novel antimicrobial endolysin, LysPA26, against Pseudomonas aeruginosa. Front Microbiol. (2017) 8:293. doi: 10.3389/fmicb.2017.00293
114. Lai M-J, Lin N-T, Hu A, Soo P-C, Chen L-K, Chen L-H, et al. Antibacterial activity of Acinetobacter baumannii phage ΦFB2 endolysin (LysAB2) against both Gram-positive and Gram-negative bacteria. Appl Microbiol Biotechnol. (2011) 90:529–39. doi: 10.1007/s00253-011-3104-y
115. Peng S-Y, You R-I, Lai M-J, Lin N-T, Chen L-K, Chang K-C. Highly potent antimicrobial modified peptides derived from the Acinetobacter baumannii phage endolysin LysAB2. Sci Rep. (2017) 7:11477. doi: 10.1038/s41598-017-11832-7
116. Thummeepak R, Kitti T, Kunthalert D, Sitthisak S. Enhanced antibacterial activity of Acinetobacter baumannii bacteriophage ØABP-01 endolysin (LysABP-01) in combination with colistin. Front Microbiol. (2016) 7:1402. doi: 10.3389/fmicb.2016.01402
117. Huang G, Shen X, Gong Y, Dong Z, Zhao X, Shen W, et al. Antibacterial properties of Acinetobacter baumannii phage Abp1 endolysin (PlyAB1). BMC Infect Dis. (2014) 14:681. doi: 10.1186/s12879-014-0681-2
118. Thandar M, Lood R, Winer BY, Deutsch DR, Euler CW, Fischetti VA. Novel engineered peptides of a phage lysin as effective antimicrobials against multidrug-resistant Acinetobacter baumannii. Antimicrob Agents Chemother. (2016) 60:2671–9. doi: 10.1128/AAC.02972-15
119. Lai M-J, Soo P-C, Lin N-T, Hu A, Chen Y-J, Chen L-K, et al. Identification and characterisation of the putative phage-related endolysins through full genome sequence analysis in Acinetobacter baumannii ATCC 17978. Int J Antimicrob Agents (2013) 42:141–8. doi: 10.1016/j.ijantimicag.2013.04.022
120. Lv M, Wang S, Yan G, Sun C, Feng X, Gu J, et al. Genome sequencing and analysis of an Escherichia coli phage vB_EcoM-ep3 with a novel lysin, Lysep3. Virus Genes (2015) 50:487–97. doi: 10.1007/s11262-015-1195-8
121. Wang S, Gu J, Lv M, Guo Z, Yan G, Yu L, et al. The antibacterial activity of E. coli bacteriophage lysin lysep3 is enhanced by fusing the Bacillus amyloliquefaciens bacteriophage endolysin binding domain D8 to the C-terminal region. J Microbiol. (2017) 55:403–8. doi: 10.1007/s12275-017-6431-6
122. Yan G, Liu J, Ma Q, Zhu R, Guo Z, Gao C, et al. The N-terminal and central domain of colicin A enables phage lysin to lyse Escherichia coli extracellularly. Antonie van Leeuwenhoek (2017) 110:1627–35. doi: 10.1007/s10482-017-0912-9
123. Shavrina MS, Zimin AA, Molochkov NV, Chernyshov SV, Machulin AV, Mikoulinskaia GV. In vitro study of the antibacterial effect of the bacteriophage T5 thermostable endolysin on Escherichia coli cells. J Appl Microbiol. (2016) 121:1282–90. doi: 10.1111/jam.13251
124. Walmagh M, Boczkowska B, Grymonprez B, Briers Y, Drulis-Kawa Z, Lavigne R. Characterization of five novel endolysins from Gram-negative infecting bacteriophages. Appl Microbiol Biotechnol. (2013) 97:4369–75. doi: 10.1007/s00253-012-4294-7
125. Maciejewska B, Roszniowski B, Espaillat A, Kesik-Szeloch A, Majkowska-Skrobek G, Kropinski AM, et al. Klebsiella phages representing a novel clade of viruses with an unknown DNA modification and biotechnologically interesting enzymes. Appl Microbiol Biotechnol. (2017) 101:673–84. doi: 10.1007/s00253-016-7928-3
126. Oliveira H, Pinto G, Oliveira A, Oliveira C, Faustino MA, Briers Y, et al. Characterization and genome sequencing of a Citrobacter freundii phage CfP1 harboring a lysin active against multidrug-resistant isolates. Appl Microbiol Biotechnol. (2016) 100:10543–53. doi: 10.1007/s00253-016-7858-0
127. Dong H, Zhu C, Chen J, Ye X, Huang Y-P. Antibacterial activity of Stenotrophomonas maltophilia endolysin P28 against both Gram-positive and Gram-negative bacteria. Front Microbiol. (2015) 6:1299. doi: 10.3389/fmicb.2015.01299
128. Maciejewska B, Zrubek K, Espaillat A, Wiśniewska M, Rembacz KP, Cava F, et al. Modular endolysin of Burkholderia AP3 phage has the largest lysozyme-like catalytic subunit discovered to date and no catalytic aspartate residue. Sci Rep. (2017) 7:14501. doi: 10.1038/s41598-017-14797-9
129. Becker SC, Foster-Frey J, Stodola AJ, Anacker D, Donovan DM. Differentially conserved staphylococcal SH3b_5 cell wall binding domains confer increased staphylolytic and streptolytic activity to a streptococcal prophage endolysin domain. Gene (2009) 443:32–41. doi: 10.1016/j.gene.2009.04.023
130. López R, García E, García P. Enzymes for anti-infective therapy: phage lysins. Drug Discov Today Ther Strateg. (2004) 1:469–74. doi: 10.1016/j.ddstr.2004.09.002
131. Low LY, Yang C, Perego M, Osterman A, Liddington R. Role of net charge on the catalytic domain and the influence of the cell-wall binding domain on the bactericidal activity, specificity, and host-range of phage lysins. J Biol Chem. (2011) 286:34391–403. doi: 10.1074/jbc.M111.244160
132. Resch G, Moreillon P, Fischetti VA. A stable phage lysin (Cpl-1) dimer with increased antipneumococcal activity and decreased plasma clearance. Int J Antimicrob Agents (2011) 38:516–21. doi: 10.1016/j.ijantimicag.2011.08.009
133. Donovan DM, Foster-Frey J. LambdaSa2 prophage endolysin requires Cpl-7-binding domains and amidase-5 domain for antimicrobial lysis of streptococci. FEMS Microbiol Lett. (2008) 287:22–33. doi: 10.1111/j.1574-6968.2008.01287.x
134. Briers Y, Walmagh M, Van Puyenbroeck V, Cornelissen A, Cenens W, Aertsen A, et al. Engineered endolysin-based “Artilysins” to combat multidrug-resistant Gram-negative pathogens. mBio (2014) 5:e01379–14. doi: 10.1128/mBio.01379-14
135. Antonova NP, Balabanyan VY, Tkachuk AP, Makarov VV, Gushchin VA. Physical and chemical properties of recombinant KPP10 phage lysins and their antimicrobial activity against Pseudomonas aeruginosa. Bull RSMU (2018) 7:22–9. doi: 10.24075/brsmu.2018.010
136. Rodríguez-Rubio L, Chang W-L, Gutiérrez D, Lavigne R, Martínez B, Rodríguez A, et al. 'Artilysation' of endolysin lSa2lys strongly improves its enzymatic and antibacterial activity against streptococci. Sci Rep. (2016) 6:35382. doi: 10.1038/srep35382
137. López R, García E. Recent trends on the molecular biology of pneumococcal capsules, lytic enzymes, and bacteriophage. FEMS Microbiol Rev. (2004) 28:553–80. doi: 10.1016/j.femsre.2004.05.002
138. García P, García JL, García E, Sánchez-Puelles JM, López R. Modular organization of the lytic enzymes of Streptococcus pneumoniae and its bacteriophages. Gene (1990) 86:81–8. doi: 10.1016/0378-1119(90)90116-9
139. Bustamante N, Campillo NE, García E, Gallego C, Pera B, Diakun GP, et al. Cpl-7, a lysozyme encoded by a pneumococcal bacteriophage with a novel cell wall-binding motif. J Biol Chem. (2010) 285:33184–96. doi: 10.1074/jbc.M110.154559
140. Bustamante N, Iglesias-Bexiga M, Bernardo-García N, Silva-Martín N, García G, Campanero-Rhodes MA, et al. Deciphering how Cpl-7 cell wall-binding repeats recognize the bacterial peptidoglycan. Sci Rep. (2017) 7:16494. doi: 10.1038/s41598-017-16392-4
141. World Health Organization. Antimicrobial Resistance: Global Report on Surveillance (2014). Available online at: http://www.who.int/drugresistance/documents/surveillancereport/en/ (Accessed September 24, 2018).
142. Rajagopal M, Walker S. Envelope structures of Gram-positive bacteria. Curr Top Microbiol Immunol. (2017) 404:1–44. doi: 10.1007/82_2015_5021
143. Sass P, Bierbaum G. Lytic activity of recombinant bacteriophage φ11 and φ12 endolysins on whole cells and biofilms of Staphylococcus aureus. Appl Environ Microbiol. (2007) 73:347–52. doi: 10.1128/AEM.01616-06
144. Pritchard DG, Dong S, Baker JR, Engler JA. The bifunctional peptidoglycan lysin of Streptococcus agalactiae bacteriophage B30. Microbiology (2004) 150:2079–87. doi: 10.1099/mic.0.27063-0
145. Gründling A, Schneewind O. Cross-linked peptidoglycan mediates lysostaphin binding to the cell wall envelope of Staphylococcus aureus. J Bacteriol. (2006) 188:2463–72. doi: 10.1128/JB.188.7.2463-2472.2006
146. Idelevich EA, von Eiff C, Friedrich AW, Iannelli D, Xia G, Peters G, et al. In vitro activity against Staphylococcus aureus of a novel antimicrobial agent, PRF-119, a recombinant chimeric bacteriophage endolysin. Antimicrob Agents Chemother. (2011) 55:4416–9. doi: 10.1128/AAC.00217-11
147. Idelevich EA, Schaumburg F, Knaack D, Scherzinger AS, Mutter W, Peters G, et al. The recombinant bacteriophage endolysin HY-133 exhibits in vitro activity against different African clonal lineages of the Staphylococcus aureus complex, including Staphylococcus schweitzeri. Antimicrob Agents Chemother. (2016) 60:2551–3. doi: 10.1128/AAC.02859-15
148. Schmelcher M, Powell AM, Becker SC, Camp MJ, Donovan DM. Chimeric phage lysins act synergistically with lysostaphin to kill mastitis-causing Staphylococcus aureus in murine mammary glands. Appl Environ Microbiol. (2012) 78:2297–305. doi: 10.1128/AEM.07050-11
149. Mao J, Schmelcher M, Harty WJ, Foster-Frey J, Donovan DM. Chimeric Ply187 endolysin kills Staphylococcus aureus more effectively than the parental enzyme. FEMS Microbiol Lett. (2013) 342:30–6. doi: 10.1111/1574-6968.12104
150. Becker SC, Roach DR, Chauhan VS, Shen Y, Foster-Frey J, Powell AM, et al. Triple-acting lytic enzyme treatment of drug-resistant and intracellular Staphylococcus aureus. Sci Rep. (2016) 6:25063. doi: 10.1038/srep25063
151. Wang Z, Kong L, Liu Y, Fu Q, Cui Z, Wang J, et al. A phage lysin fused to a cell-penetrating peptide kills intracellular methicillin-resistant Staphylococcus aureus in keratinocytes and has potential as a treatment for skin infections in mice. Appl Environ Microbiol. (2018) 84:e00380–18. doi: 10.1128/AEM.00380-18
152. Brouwer S, Barnett Timothy C, Rivera-Hernandez T, Rohde M, Walker Mark J. Streptococcus pyogenes adhesion and colonization. FEBS Lett. (2016) 590:3739–57. doi: 10.1002/1873-3468.12254
153. Heath PT, Culley FJ, Jones CE, Kampmann B, Le Doare K, Nunes MC, et al. Group B streptococcus and respiratory syncytial virus immunisation during pregnancy: a landscape analysis. Lancet Infect Dis. (2017) 17:e223–34. doi: 10.1016/S1473-3099(17)30232-3
154. Squeglia F, Ruggiero A, Berisio R. Chemistry of peptidoglycan in Mycobacterium tuberculosis life cycle: an off-the-wall balance of synthesis and degradation. Chemistry (2018) 24:2533–46. doi: 10.1002/chem.201702973
155. Payne KM, Hatfull GF. Mycobacteriophage endolysins: diverse and modular enzymes with multiple catalytic activities. PLoS ONE (2012) 7:e34052. doi: 10.1371/journal.pone.0034052
156. Catalão MJ, Pimentel M. Mycobacteriophage lysis enzymes: targeting the mycobacterial cell envelope. Viruses (2018) 10:428. doi: 10.3390/v10080428
157. Fisher RA, Gollan B, Helaine S. Persistent bacterial infections and persister cells. Nat Rev Microbiol. (2017) 15:453–64. doi: 10.1038/nrmicro.2017.42
158. Düring K, Porsch P, Mahn A, Brinkmann O, Gieffers W. The non-enzymatic microbicidal activity of lysozymes. FEBS Lett. (1999) 449:93–100. doi: 10.1016/S0014-5793(99)00405-6
159. Rotem S, Radzishevsky I, Inouye RT, Samore M, Mor A. Identification of antimicrobial peptide regions derived from genomic sequences of phage lysins. Peptides (2006) 27:18–26. doi: 10.1016/j.peptides.2005.07.001
160. de Pedro MA, Cava F. Structural constraints and dynamics of bacterial cell wall architecture. Front Microbiol. (2015) 6:449. doi: 10.3389/fmicb.2015.00449
161. Elhosseiny NM, Attia AS. Acinetobacter: an emerging pathogen with a versatile secretome. Emerg Microb Infect. (2018) 7:33. doi: 10.1038/s41426-018-0030-4
162. Porreca AM, Sullivan KV, Gallagher JC. The epidemiology, evolution, and treatment of KPC-producing organisms. Curr Infect Dis Rep. (2018) 20:13. doi: 10.1007/s11908-018-0617-x
163. Junn HJ, Youn J, Suh KH, Lee SS. Cloning and expression of Klebsiella phage K11 lysozyme gene. Protein Expr Purif. (2005) 42:78–84. doi: 10.1016/j.pep.2005.03.026
164. Koulenti D, Tsigou E, Rello J. Nosocomial pneumonia in 27 ICUs in Europe: perspectives from the EU-VAP/CAP study. Eur J Clin Microbiol Infect Dis. (2017) 36:1999–2006. doi: 10.1007/s10096-016-2703-z
165. Lukacik P, Barnard TJ, Keller PW, Chaturvedi KS, Seddiki N, Fairman JW, et al. Structural engineering of a phage lysin that targets gram-negative pathogens. Proc Natl Acad Sci USA (2012) 109:9857–62. doi: 10.1073/pnas.1203472109
166. Fernández-Ruiz I, Coutinho FH, Rodríguez-Valera F. Thousands of novel endolysins discovered in uncultured phage genomes. Front Microbiol. (2018) 9:1033. doi: 10.3389/fmicb.2018.01033
167. Huang WC, Tsai PJ, Chen YC. Multifunctional Fe3O4@Au nanoeggs as photothermal agents for selective killing of nosocomial and antibiotic-resistant bacteria. Small (2009) 5:51–6. doi: 10.1002/smll.200801042
168. Kell AJ, Stewart G, Ryan S, Peytavi R, Boissinot M, Huletsky A, et al. Vancomycin-modified nanoparticles for efficient targeting and preconcentration of Gram-positive and Gram-negative bacteria. ACS Nano (2008) 2:1777–88. doi: 10.1021/nn700183g
169. Li Y, Krentz TM, Wang L, Benicewicz BC, Schadler LS. Ligand engineering of polymer nanocomposites: from the simple to the complex. ACS Appl Mater Interfaces (2014) 6:6005–21. doi: 10.1021/am405332a
170. Choi K-H, Lee H-J, Park BJ, Wang K-K, Shin EP, Park J-C, et al. Photosensitizer and vancomycin-conjugated novel multifunctional magnetic particles as photoinactivation agents for selective killing of pathogenic bacteria. Chem Commun. (2012) 48:4591–3. doi: 10.1039/c2cc17766h
Keywords: phage lysins, pneumonia, respiratory infection, antibacterials, antibiotic resistance, endolysins
Citation: Vázquez R, García E and García P (2018) Phage Lysins for Fighting Bacterial Respiratory Infections: A New Generation of Antimicrobials. Front. Immunol. 9:2252. doi: 10.3389/fimmu.2018.02252
Received: 15 June 2018; Accepted: 11 September 2018;
Published: 16 October 2018.
Edited by:
Junkal Garmendia, Consejo Superior de Investigaciones Científicas (CSIC), SpainReviewed by:
Theo Araújo-Santos, Universidade Federal do Oeste da Bahia, BrazilMark Ambrose, University of Tasmania, Australia
Copyright © 2018 Vázquez, García and García. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pedro García, cGdhcmNpYUBjaWIuY3NpYy5lcw==