 Josué da Costa Lima-Junior
Josué da Costa Lima-Junior Lilian Rose Pratt-Riccio
Lilian Rose Pratt-Riccio
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 27 January 2016
Sec. Microbial Immunology
Volume 7 - 2016 | https://doi.org/10.3389/fimmu.2016.00013
This article is part of the Research Topic Immune evasion strategies in protozoan-host interactions View all 50 articles
The importance of host and parasite genetic factors in malaria resistance or susceptibility has been investigated since the middle of the last century. Nowadays, of all diseases that affect man, malaria still plays one of the highest levels of selective pressure on human genome. Susceptibility to malaria depends on exposure profile, epidemiological characteristics, and several components of the innate and adaptive immune system that influences the quality of the immune response generated during the Plasmodium lifecycle in the vertebrate host. But it is well known that the parasite’s enormous capacity of genetic variation in conjunction with the host genetics polymorphism is also associated with a wide spectrum of susceptibility degrees to complicated or severe forms of the disease. In this scenario, variations in genes of the major histocompatibility complex (MHC) associated with host resistance or susceptibility to malaria have been identified and used as markers in host–pathogen interaction studies, mainly those evaluating the impact on the immune response, acquisition of resistance, or increased susceptibility to infection or vulnerability to disease. However, due to the intense selective pressure, number of cases, and mortality rates, the majority of the reported associations reported concerned Plasmodium falciparum malaria. Studies on the MHC polymorphism and its association with Plasmodium vivax, which is the most widespread Plasmodium and the most prevalent species outside the African continent, are less frequent but equally important. Despite punctual contributions, there are accumulated evidences of human genetic control in P. vivax infection and disease. Herein, we review the current knowledge in the field of MHC and derived molecules (HLA Class I, Class II, TNF-α, LTA, BAT1, and CTL4) regarding P. vivax malaria. We discuss particularly the results of P. vivax studies on HLA class I and II polymorphisms in relation to host susceptibility, naturally acquired immune response against specific antigens and the implication of this knowledge to overcome the parasite immune evasion. Finally, the potential impact of such polymorphisms on the development of vaccine candidate antigens against P. vivax will be studied.
Caused by blood-borne apicomplexan parasites of the genus Plasmodium, malaria remains a major public health problem. Malaria transmission occurs in 96 countries and territories, and according to the latest estimates, 3.3 billion people are at risk of infection. Approximately 214 million cases and 438,000 deaths due to malaria occur worldwide, mainly of children under 5 years. The great majority of cases (88%) and deaths (90%) occurs in Africa, followed by Southeast Asia (10%) and Eastern Mediterranean region (2%) (1). Of the five Plasmodium species that affect humans, Plasmodium vivax is responsible for about 6% of the world estimated cases. However, outside sub-Saharan Africa, P. vivax accounts for 51% of all malaria cases, being the most widespread Plasmodium species (1).
Plasmodium parasites have a complex lifecycle, which includes the development of a sexual cycle in the invertebrate vector, the female of the Anopheles mosquitos, and an asexual cycle in the vertebrate hosts. Infection with Plasmodium parasites presents an asymptomatic stage, pre-erythrocytic, which occurs in the liver followed by a symptomatic erythrocytic stage, when merozoites arisen during pre-erythrocytic stage invade red blood cells. The rupture of the erythrocytic schizont is typically accompanied by clinical symptoms, because of the release of parasite derived toxins, such as phospholipids, that can activate immune cells leading to the production of inflammatory cytokines that can, directly or indirectly, contribute to the elimination of the parasite and complications associated with infection.
At first, humans are susceptible to malaria, even those who have already contracted the disease several times. However, young children, pregnant women, and adults from non-endemic areas are particularly susceptible to develop severe malaria. In high endemic malaria areas, with repeated exposure, older children and adults develop considerable degree of protection from death and severe malaria, and thus, the clinical manifestations are milder, or even absent, although sterile immunity is probably never achieved and the infected immune individuals continue to present parasites in the blood for long periods, probably in the presence of very mild symptomatology (2, 3). It has been proposed that these changes reflect the parasitological and clinical immunity collectively referred to as naturally acquired immunity, which generally determines not only the age-specific incidence and prevalence of infections but also the expression of pathological processes that underlie the clinical manifestations of infection.
The spectrum of malaria clinical manifestations generally differs between adults and children and from person to person, ranging from asymptomatic infection to clinical symptoms as fever, nausea, headache, and muscle pain, chills and vomiting and, in 1–2% of the cases, to severe malaria, leading to multiorgan system involvement, severe anemia, and death (4–6). Plasmodium falciparum is the most virulent agent and responsible for the majority of severe malaria deaths (1). Severe malaria due to P. falciparum may present as confusion, drowsiness, excitement, convulsions, delirium, and coma. The differences observed in the clinical forms of the disease as well as the underlying pathophysiological processes are still under investigation, but it is now clear that the genetic factors influence the spectrum of clinical manifestations and the evolution and severity of the disease (7, 8).
The classical framework of the influence of genetic factors in malaria evolution and severity is the protective effect of certain hemoglobinopathies. The first observations were postulated in the late 40s by Haldane, known as one of the three founders of population genetics and acknowledged as the first person to suggest that disease could be an important evolutionary force in humans (9). Based on the distribution of thalassemia in the Mediterranean, Haldane proposed that certain hemoglobinopathies are highly prevalent in regions where malaria is endemic due to the protection against the severe disease (10). According to the Haldane’s malaria hypothesis, this could result in a “balanced polymorphism” where the homozygote disadvantage for inherited erythrocyte disease is compensated through the resistance of the heterozygote where malaria is endemic (11–14). Thus, it has been proposed that malaria is associated to gene selective pressure in the human genome, and it has been associated with some genetic diseases. After that, several reports have shown that genetic disorders, such as thalassemias, sickle-cell trait, glucose-6-phosphate dehydrogenase (G6PD) deficiency, ovalocytosis, Hemoglobin (Hb) S, HbC, HbE, and complement receptor-1 (CR1) deficiency, are associated with malaria susceptibility or resistance. Case–control studies have shown that these polymorphisms reduce the risk of severe and complicated malaria. Among the mechanisms involved in the protection against P. falciparum severe malaria are reduced invasion of erythrocyte by the parasite, decreased intracellular parasite growth, increased phagocytosis, and enhanced immune response against parasite-infected erythrocyte (14–17). Besides these genetic disorders, other polymorphisms in genes encoding the immune system molecules may also be involved in malaria outcome (Table 1).
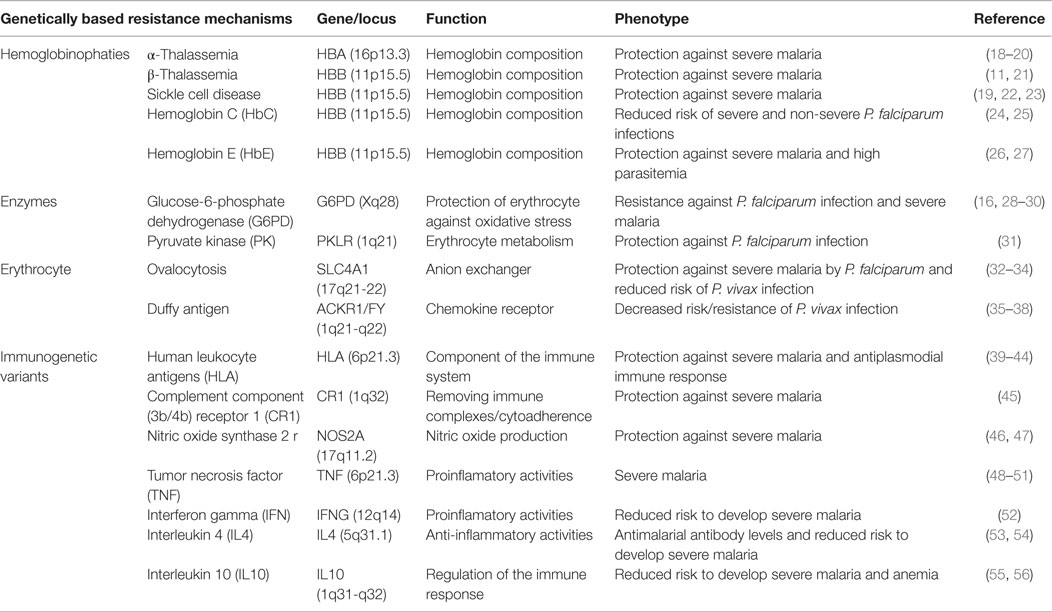
Table 1. Genetic polymorphisms of the vertebrate host and associations with the natural resistance to malaria.
Considering the intense selective pressure, the number of cases and the mortality rates associated with P. falciparum infection, specific studies of association between genetic factors and P. vivax are less frequent, even though this species is the most widespread Plasmodium, the most prevalent species outside the African continent, and with increasing evidences of associated death (57, 58). The observation that P. vivax malaria is rare in West Africa and that most sub-Saharan Africans are negative to blood group Duffy was the first evidence regarding P. vivax natural resistance. It led to the discovery that P. vivax uses the Duffy blood group antigen as a receptor to invade erythrocytes (59). Populations with the null phenotype of Duffy, although susceptible to the hepatic malaria stage, are less susceptible to P. vivax merozoite invasion. Moreover, there are accumulated evidences of the relationship between immune response to P. vivax antigens and major histocompatibility complex (MHC) genes. Therefore, in the present study, we review the current knowledge in the field of MHC molecules regarding P. vivax malaria.
Like in other species of the Plasmodium genus, P. vivax lifecycle is a complex process and requires an invertebrate and a vertebrate host for survival and perpetuation (Figure 1). Therefore, during its entire life cycle in humans, P. vivax undergoes multiple morphological and antigenically distinct stages and can be attacked by different immune mechanisms, depending on the stage and whether the parasite is within or outside the host cell. During the migration through the bloodstream to the liver, antibodies can block sporozoite migration and/or invasion of hepatocytes, repressing lifecycle progression (60–63). In the liver stage, infected hepatocytes are potential targets of CD4+ and CD8+ T cells, although the immune response mediated by NK cells and T gamma-delta T cells also participates in the immune response against pre-erythrocytic forms stimulating other cell populations secreting cytokines or acting directly on the infected hepatocyte (64–66). After being released from merosomes, free merozoites are susceptible to host immune responses. Merozoites can be the target of opsonizing antibodies, triggering cell-mediated merozoite killing or blocking merozoite proteins responsible for the initial interaction with the molecules on the surface of erythrocytes, preventing invasion (67, 68). Considering that the MHCs Class I and II antigens are absent on the surface of the erythrocytes, the immune response against blood stage forms involves mainly antibodies. During the intraerythrocytic stage, antibodies may coalesce merozoites at or just before the rupture of erythrocytes, preventing their release and spread into the bloodstream, essential for the clearance of parasitemia in the later stages of the infection (2, 67, 68). Although antibodies have a critical role in the development of immunity against erythrocytic forms, studies indicate that the development of the immune response also involves monocytes, neutrophils, CD4+ T cells, NK cells, and NKT cells (68).
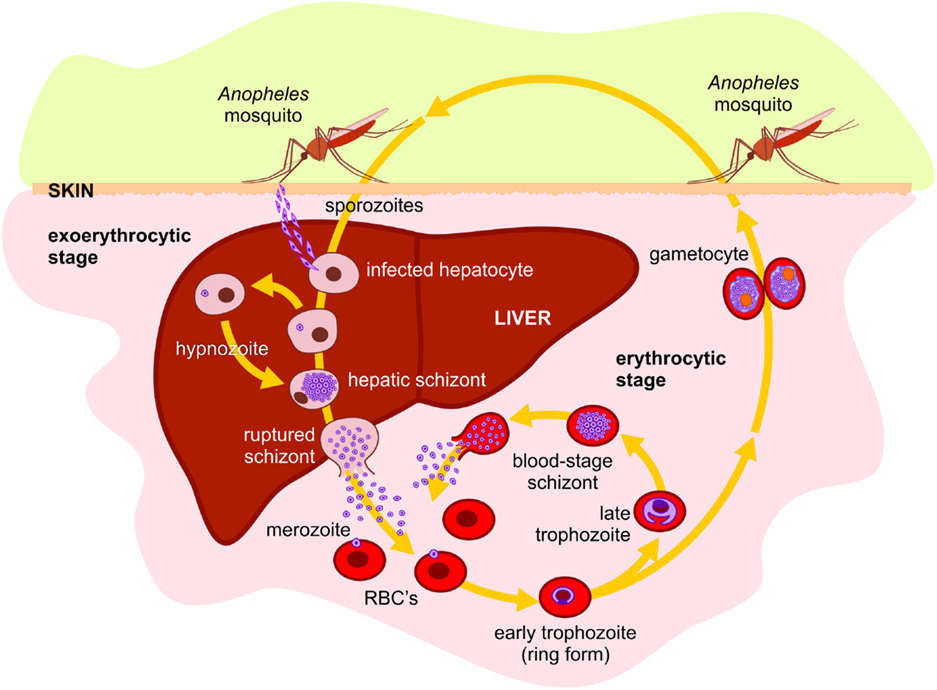
Figure 1. Plasmodium vivax lifecycle in human host: P. vivax is transmitted to humans by the bite of an infected female of the Anopheles mosquito, releasing the salivary fluid carrying sporozoites into the tissues or directly into the bloodstream. From the tissues, the motile sporozoites can penetrate small blood vessels. In hepatic sinusoids, they penetrate through Kupffer cells into Space of Disse and invade hepatocytes to begin the exo-erythrocytic or liver-stage cycle. The sporozoite differentiates into mature liver-stage schizont with thousands of uninucleated merozoites surrounded by a parasitophorous membrane. The hepatocyte containing mature liver schizonts ruptures releasing merosomes. These merosomes are transported into the general blood circulation and break, releasing merozoites which invade young red blood cells (reticulocytes), beginning the erythrocytic or blood-stage cycle. P. vivax has dormant liver hypnozoite stages, which can reactivate and lead to blood-stage relapses. Within the erythrocyte, the merozoite differentiates in erythrocytic trophozoite. When fully mature, the infected erythrocyte ruptures, releasing the merozoites, which then invade new erythrocytes, initiating the entire intraerythrocytic-stage cycle, rupture, and reinvasion. Alternatively, some merozoites can develop gametocytes. During blood feeding, female mosquito of a susceptible Anopheles species can ingest the gametocytes, beginning the sexual stage of the life cycle. In the midgut of the mosquito, gametocytes escape from erythrocytes and become sexually stimulated. The male gamete fuses with the female, forming a diploid zygote. Therefore, the zygote is transformed into an invasive parasite stage ookinete. The ookinete traverses the midgut wall by passing through epithelial cells and comes to rest adjacent to the basal lamina where it transforms into an oocyst that undergoes multiple nuclear divisions producing several thousand sporozoites. At maturity, the oocyst breaks open and the sporozoites are released into the hemocele of the mosquito, migrating and penetrating the salivary glands. In the salivary glands, the sporozoites become infectious to humans, completing the life cycle.
Overall, one may say that the cellular immune response is more important in the control of the hepatic forms of the parasite, whereas the humoral immune response seems to be more important to the control of its erythrocytic stage. Since P. vivax stimulates various components of the immune system, the balance of this activation can represent a fine line between inhibition of the parasite growth and immunopathology. Thus, it is acceptable to consider that polymorphisms in genes encoding immune system molecules, especially those located at MHC locus, could be involved in P. vivax malaria outcome.
The MHC, referred as human leukocyte antigen (HLA) system in humans, is an extremely polymorphic region encoding for the major molecules in charge of antigen presentation on the cell surface, and it has been one of the most intensively studied areas in the human genome (69, 70). Located in the short arm of the chromosome 6, HLA complex consists of more than 200 genes categorized into three basic groups: class I, class II, and class III (Figure 2). Class I molecule is a heterodimer consisting of a heavy chain and a light chain, the beta-2 microglobulin. HLA Class I genetic region encodes the heavy chain of the classics HLA-A, -B, and -C molecules, besides HLA-E, -F, -G, and the MHC class I polypeptide-related sequence A (MICA) and MICB. Class I molecules are expressed in nearly all cells and play a central role in the immune system by presenting peptides derived from the endoplasmic reticulum lumen. Class II molecules are heterodimers formed by α and β chains. HLA class II genetic region, initially called Immune response (Ir) genes due to its role in controlling the immune response, encodes the α and β chains of the HLA-DR, -DQ, -DP, -DM, and -DO molecules and peptide transporter proteins (TAP) 1 and (TAP) 2 (69–71). Class II molecules are predominantly expressed on antigen-presenting cells (APC), such as macrophages, dendritic cells, B cells, Langerhans cells, and Kupffer cells, although some cells may express class II molecules during inflammatory process (70, 72, 73). The proteins produced from HLA class III genes have somewhat different functions, some of which involve participation in inflammation processes and other immune system activities. HLA-Class III genetic region encodes C2 and C4 complement components and tumor necrosis factor (TNF) superfamily (70). The functions of some HLA genes are unknown.
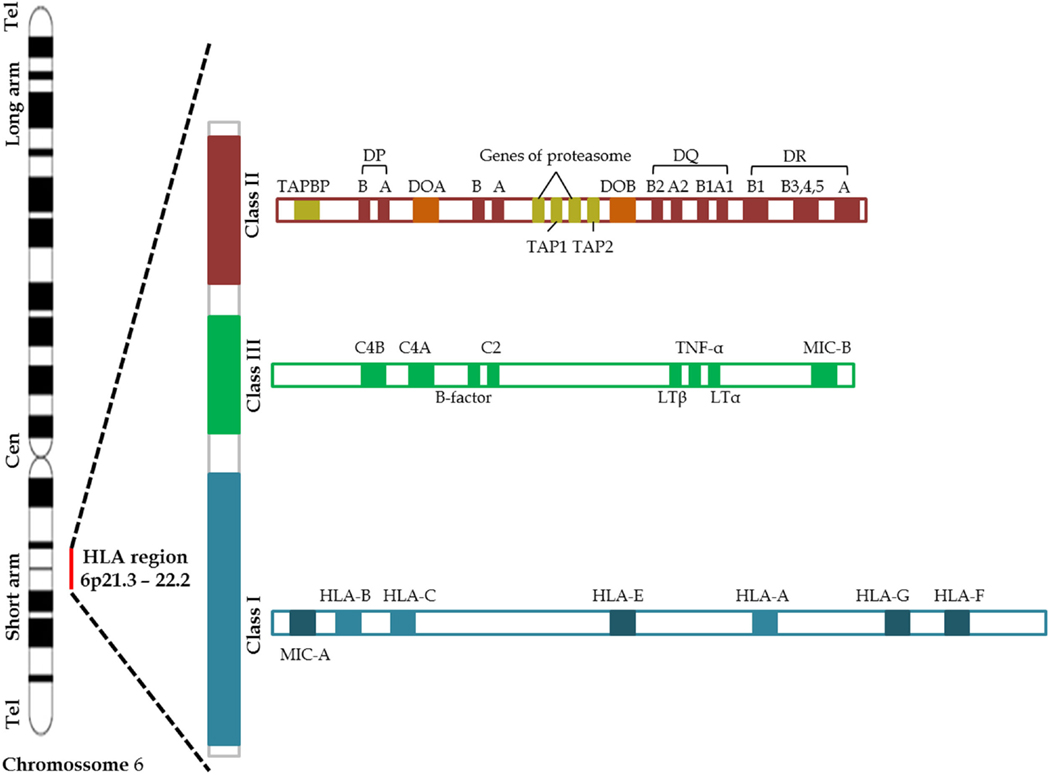
Figure 2. Schematic representation of the human Chromosome 6 including the main MHC genes (69).
The polymorphism of HLA has been useful in the search of donors with compatible grafts in tracing population migration as well as in its potential relationship to pathogen-mediated selection. Further, assessing and comparing the polymorphism of HLA allows to better define the extent of the genetic variability in humans as well as the reasons of this diversity. The HLA region is associated with more diseases (mainly autoimmune and infectious diseases) than any other region of the genome (74–78).
The importance of HLA genes influencing malaria outcome has been demonstrated since studies conducted by Hill and colleagues who elegantly illustrated the influence of HLA genes in the protection against an intracellular pathogen and how the polymorphism of HLA genes may have evolved through selection of molecules induced by the pathogen. In a study of over 2000 children in West Africa, Hill et al. showed that carriers of HLA Class I Bw53 and HLA class II DRB1*1302-DQB1*0501, frequently occurring in sub-Saharan Africa, were protected against severe malaria (39). Later studies showed that HLA-B53 restricted cytotoxic T cells recognize peptides corresponding to regions of P. falciparum Liver Stage Antigen-1 (79). Thus, HLA molecules have been used as genetic markers in an attempt to determine the presence of genetic modulation of the immune response during malaria infection. Considering the increasing focus on the development of subunit malaria vaccines, studies on the influence of HLA molecules in the immune response in ethnically diverse populations are important before the implementation of vaccine trials. This is particularly relevant for P. vivax, which affects populations with high diversity of genetic backgrounds.
In this scenario, the circumsporozoite surface protein (CSP) is the most abundant polypeptide present in the sporozoite covering. This protein is involved in the motility and invasion of the sporozoite during its entrance in the hepatocyte (80). The csp gene encodes a protein, which has in its central portion, a highly immunogenic repetitive region. Based on the csp gene, two variants, VK247 and P. vivax-like, have been described. They differ from the classical form (VK210) by sequence variations in the central region of the gene (81, 82). A study performed by Oliveira-Ferreira and others with 108 individuals living in Rondonia State, in the Southwestern part of the Brazilian Amazon, observed a significant association between the antibody response to the CSP repeats of VK247and the presence of HLA-DRB1*16 and between the presence of HLA-DRB1*07 and the absence of antibody responses to the CSP repeats of VK210 (83). More recently, Storti-Mello and co-workers described a significant association between the absence of antibody response to the CSP amino-terminal region and the presence of HLA-DRB1*03 and DR5 in a study with 55 individuals from different regions of the Brazilian Amazon (84) (Table 2).
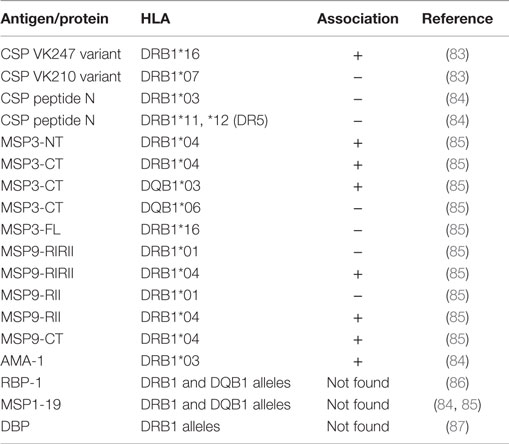
Table 2. Associations between HLA-DRB1 and HLA-DQB1 allelic groups and antibody response to P. vivax antigens.
Merozoite surface proteins (MSPs) have been reported as abundantly expressed on the surface of merozoites and can contribute to the initial recognition of erythrocytes. MSP-1, MSP-3, and MSP-9 have been considered important vaccine candidates based on their location, on their recognition by antibodies from individuals naturally exposed to P. vivax, their immunogenic properties in animal models, and evidence of the induction of antibodies able to inhibit parasite-growth (88–95). Therefore, considering the importance of the antibodies against MSPs in the development of anti-parasite immunity, studies have also focused the evaluation of the genetic restriction of the anti-MSP humoral response. Storti-Melo et al. analyzed the influence of the HLA-DRB1 alleles on antibody levels against the amino-terminal region of the MSP-1 in individuals from the Brazilian Amazon and observed significant association between high levels of antibodies for MSP-1 and the presence of HLA-DRB1*03 (84). In contrast, no evidence of a specific HLA-DR or HLA-DQ restriction for the antibody response to MSP-1 was observed in a study carried out by Lima-Junior et al. in 276 individuals living in Rondonia State in the Brazilian Amazon (85). However, in that paper, the authors showed HLA associations with IgG antibody response against different regions of MSP-3 and MSP-9 proteins. A high frequency of responders to carboxy-terminal (CT) and amino-terminal (NT) regions of MSP-3 were defined in HLA-DRB1*04 carriers and to MSP-3CT also defined in HLA-DQB1*03 carriers. Additionally, a high frequency of non-responders to MSP-3CT and the presence of HLA-DQB1*06 and to a recombinant protein representing the full length (FL) of MSP-3 with the presence of the HLA-DRB1*16 allele were observed. Regarding MSP-9, the presence of HLA-DRB1*04 was positively associated with the IgG immune response against all constructions used in the study, the amino-terminal domain (NT) and the C-terminal blocks of tandem repeats (RII and RIRII), while the presence of the HLA-DRB1*01 was associated with the high frequency of non-responders only to the repeated region (Table 2).
However, other studies did not find associations between HLA-DR or HLA-DQ alleles and antibody response to P. vivax antigens. In a study performed by Ferreira and co-workers, no genetic restriction mediated by HLA-DRB1* and HLA-DQB1* against two constructions of P. vivax Reticulocyte Binding Protein-1 (PvRBP1) was verified in more than 500 HLA alleles from different individuals from communities in the Amazon region of Brazil (86). Moreover, regarding the cellular response, Arevalo-Herrera et al. also did not observe association between HLA and cellular immune response of healthy volunteers vaccinated with CSP derived long synthetic peptides (96) and Lima-junior et al. describe five promiscuous peptides from MSP-9 which also presented no association between HLA-DRB1 alleles and the cellular immune response (97).
Several genes of the immune system have proved to be important in relation to the susceptibility or resistance to malaria, especially those associated with severe malaria. Therefore, a common strategy is to identify the mutations in such genes and observe their possible association with the disease outcome. Since Kwiatkowski et al. showed that the TNF was associated with the susceptibility to cerebral malaria (98), numerous mutations have already been identified in the promoter of this gene, which can influence on TNF production rate. In one of those vanguard studies, McGuire and colleagues showed that mutation at position -308 of the TNF promoter region is associated with increased risk of death from cerebral malaria in Africa (51). Analysis of other clinical complications experienced in African children with severe malaria also showed that severe anemia due to malaria is associated with the mutation at position -238 suggesting that the clinical manifestations could also be influenced by genetic determinants located near the TNF gene. In fact, the guanine-to-adenine transition at position -308 in the TNF promoter, which defines the rare allele TNF2 is strongly associated with the MHC haplotypes HLA-A1, B8, DR3 and was also reported to influence the TNF promoter activity, enhancing TNF-α production (99). In patients with cerebral, severe malaria and mucocutaneous leishmaniasis, the TNF-α -308G/A polymorphism has been shown to be associated with the outcome and clinical course of the disease (100). However, only in the last years, the influence of these polymorphisms on P. vivax infection began to be investigated. On the one hand, in patients with P. vivax malaria from India, two single nucleotide polymorphisms (SNP) in the TNF promoter (–308G > A and –1031C > T) were associated with cytokine levels and temperature, but no association related to susceptibility were reported (101). On the other hand, there was neither association between six different TNF SNP polymorphisms and P. vivax malaria in Thailand nor differences in allelic distribution among the three distinct ethnic groups assessed by the study: Thai, Burmese, and Karen (102). In Brazil, even TNF-308 GA genotype or A allele carriers presented higher levels of TNF than those with the GG genotype or G allele, no association related to susceptibility was observed in P. vivax infected individuals (103, 104). In fact, we tend to reinforce the idea that a SNP is often not sufficient for predicting the susceptibility or resistance of individuals to P. vivax malaria. Therefore, the usual approach when investigating the differences in response to malaria infection should be the haplotype analysis. For example, Sortica et al. reported the association of TNF haplotype with a lower susceptibility to P. vivax infections, since an uninfected group presented a significantly higher frequency of a specific haplotype (T1031/A863/C857/G308/G238) when compared to P. vivax infected individuals (104). However, despite these several evidences of polymorphism in TNF gene in relation to malaria susceptibility in the studies, a larger number of samples and different clinical and epidemiological scenarios are necessary to confirm the associations.
Aside the TNF association studies, the associations between malaria and polymorphisms in other genes located at HLA locus were also the focus of investigations. For example, the nuclear protein HLA-B-associated transcript 1 (BAT1), which is an RNA helicase encoded by the DDX39B gene, has been described as a negative regulator of inflammation by modulating expression of proinflammatory cytokines (such as TNF) (105). Therefore, using mutations in two MHC genes located approximately at 150 kb from each other (TNF and DDX39B) Mendonça et al. reported associations between DDX39B haplotypes and complicated P. vivax malaria. Participants with DDX39B-22/DDX39B-348/TNF-308/IL6-176 genotype combinations GC/CC/GG/GG and GG/CT/GG/GG had reduced and increased risk, respectively, of developing malaria symptoms (103).
Lastly, other HLA-Class III host candidate gene polymorphisms were also associated with susceptibility/resistance to Plasmodium infection. However, the absence of studies using only P. vivax infected/exposed individuals makes the definition of genetic polymorphism of HLA-class III genes associated specifically to this species particularly difficult. In P. falciparum studies conducted in Africa, a trend of association between LTA polymorphism with antimalarial IgG subclass levels was found but not confirmed by statistical tests (106). Moreover, no LTA polymorphisms were associated with severe malaria in cohorts in Kenya and Malawi in a large study involving >10,000 individuals from three African populations. In Brazilian endemic areas, recently, a study with a large number of candidate gene polymorphisms was performed and the association with susceptibility/resistance to Plasmodium infection with clinical (mild) malaria in a population infected with P. falciparum or P. vivax was investigated. Although no differences between species were found, the results showed, for the first time, an association between alleles of CTL4 gene with malaria, which displayed a significant association with reduced risk for clinical malaria. In addition, two other associations with cytokines were identified, both within MHC class III region, that included TNF and the lymphotoxin alpha (LT-α/LTA) and beta (LT-β/LTB) genes, which are closely related (107).
The ultimate goal of MHC binding antigenic peptide prediction is to identify epitopes that activate T-cells and mediate cell-mediated immunity without HLA genotype/haplotype restriction. Recognition of peptide bound to an MHC molecule by a T-cell receptor is a critical step and for T-cell activation binding of peptide to the MHC molecule is a necessary requirement (108). The association of immunogenic fragments (epitopes) to the HLA molecules of class I or II determines what type of cell is to be stimulated and, consequently, what kind of response will be generated. Conventional vaccinology approaches accumulate successes and failures aiming at experimental screening methods to evaluate the presence of HLA restriction in immune response to vaccine candidates. But this conventional process is still laborious, expensive, and time-consuming. Computational prediction methods complement experimental studies, minimize the number of validation experiments, and significantly speed up the epitope mapping process (109). The bioinformatics tools have already e helped identifying promiscuous epitopes within Leishmania (110), Mycobacterium tuberculosis (111) and HIV (112) antigens. In malaria, epitope identification is particularly challenging, as more than 5000 proteins are encoded by the genome (113, 114), which could generate hundreds of thousands of possible CD4+ T cell epitopes. On the other hand, the identification of CD4+ and CD8+ epitopes from malaria is urgently required to track various vaccine approaches, mainly to evaluate candidates for compositions of subunit vaccines. For example, in P. falciparum vaccine research, Doolan et al. first used proteomic approaches to identify 27 highly expressed candidate antigens, and then used HLA-DR binding predictions to identify 723 predicted HLA-DR binders. Of these, 39 peptides binding tightly to HLA-DR variants derived from four newly identified antigenic targets were identified (115). Beyond antigen identification, this application of proteomics and bioinformatics was confirmed as particularly powerful and is likely to prove useful in other applications, particularly as consensus motif prediction approaches.
Despite several T-cell epitopes from pre-erythrocytic (116–118), asexual blood stage (119–121), and gametocyte (122) antigens have been predicted and/or experimentally confirmed for P. falciparum, the use of bioinformatics strategies to identify potentially important epitopes in P. vivax is still restricted. The majority of the studies focusing on the detection of B or T-cell epitopes have used conventional screening methods (94, 123–125). Only few studies have already used prediction servers to trial the most promising epitopes to be used in validation assays (Table 3). One of the first prediction studies on P. vivax reported the results of in silico analysis of PvMSP-1 vaccine candidates in relation to potential HLA restricted or promiscuous CD4 and CD8 epitopes (126). More recently, Kumar et al. using several computational screening methods analyzed 10 protein sequences of P. vivax proteins, including vaccine candidates, such as MSP-1, MSP-9, Pvs25, and PvS28 in relation to potential antigenicity, promiscuity and binding to several HLA class I and II alleles. The best scored T-CD4 and T-CD8 epitopes for each antigen were also identified (127). Even with promising results, these bioinformatics approach reported is still dependent on experimental validation. In this scenario, our previous studies reported that along all PvMSP-9 N-terminal 11 peptides were highly predicted by the ProPred algorithm to be promiscuous, of which only five of them were recognized at high frequency by PBMCs from individuals living in malaria endemic areas presenting a large variety of HLA class II allelic groups (97). If the conventional screening methods had been applied, at least 40 overlapping peptides should have been synthetized and tested individually in order to select these promiscuous epitopes; on the other hand, if we had used only prediction approaches, five non-immunogenic peptides could have been selected. Although bioinformatics approach has lately accumulated more successes than failures, the confidence level (approximately 50%) for predicting epitopes to MHC class II molecules is far from perfection and in some cases can cause mismatches between predicted versus experimental results. This can happen mainly because these molecules accept a wider range of peptides in size and binding registers (16). For example, two universal epitopes were described in PvDBP sequence using conventional vaccinology experiments; however, the SYFPEITHI-binding prediction for the HLA-DRB1*0101 molecule was not in accordance with the experimental results (128). This comparison between experimental and theoretical data sets suggests that class II binding prediction tools are useful, but they have to be used with caution. Therefore, by different ways, both PvMSP-9 and PvDBP studies highlighted the combination of in silico analysis and the experimental confirmation as the ideal method. Therefore, actually there are accumulated evidences of successful use of bioinformatics on P. vivax vaccine research. For example, peptide sequences of PvRBP1 promiscuous for binding to HLA class II molecules were selected by ProPred algorithm and the IEDB server (http://www.iedb.org/) for allele binding score and population coverage. The most promising peptide sequences were included in a PvRBP-1 chimeric antigen containing the predicted promiscuous T-cell epitopes and known B-cell epitopes and presented no HLA restriction in naturally acquired immune response of exposed individuals (86). Moreover, Cespedes et al. also used the identification and selection of novel antigens by structure, binding predictions and protein motifs. A total of 50 P. vivax antigens were selected based on proteome and transcriptome data of P. falciparum orthologs. After immunological confirmation, four peptides were experimentally confirmed as truly immunogenic peptides and were preselected for further preclinical trials (129). Lastly, beyond the vaccine field, the prediction tools have also been used in a comparative analysis of epitopes from lactate dehydrogenase (LDH) protein (130). Based on P. vivax and P. falciparum LDH sequences, T-cell epitope prediction indicated that 28 HLA alleles could recognize pLDH antigen epitopes. Interestingly, despite a large number of potentially common or similar epitopes, specific Pv-LDH and Pf-LDH epitopes were also predicted and, if experimentally confirmed, could be involved in future specific diagnostic rapid tests.
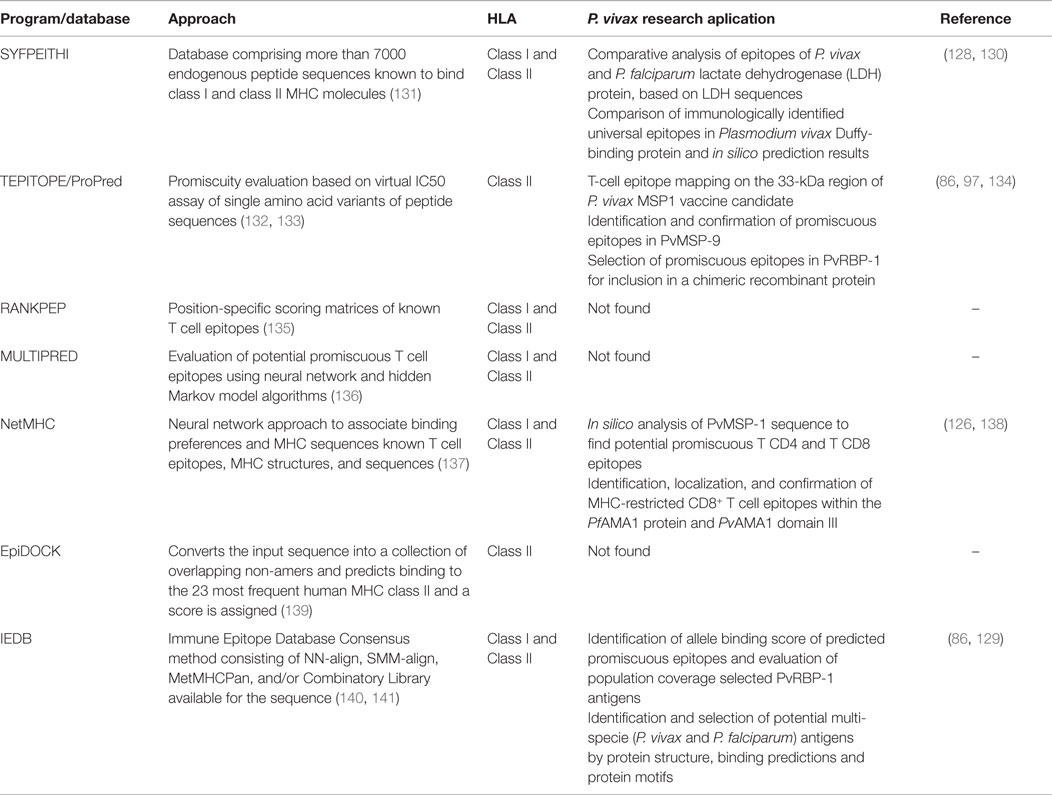
Table 3. Bioinformatics approaches applied to epitope selection in relation to MHC alleles.
In summary, with the concomitant advent of whole-genome sequencing and advances in bioinformatics, the vaccinology field changed in the last few decades, providing the opportunity of describing novel antigens and improving the already known. Consequently, the focus in vaccine design shifted to explore antigens susceptible to antibody recognition and T-cell induction through comparative pan genome reverse vaccinology. Even though, in most of cases, experimental confirmation is necessary, high-accuracy predictions are available for any HLA known, non-human primates, mouse strains, and other mammals. Therefore, those “reverse immunology” systems have become highly accessible, and they can be a fast and efficient alternative when some conventional vaccinology strategies are difficult, especially when dealing with non-culturable microorganisms, as P. vivax.
The number of studies involving MHC polymorphism and P. vivax specific immune response and clinical outcome are still increasing, and there are several similarities and disparities among these association studies. Despite the variation of MHC genes, alleles and/or haplotypes in different clinical and epidemiological scenarios, the association between MHC genes and P. vivax has been demonstrated in the majority of studies presented. We believe that the inconsistency of some data may derive from the fact that a large number of potential risk factors, such as nutritional status, coinfections, and relapses, which could influence the specific immune response, are almost impossible to be controlled in malaria endemic areas. Other issues are the small sample size, the heterogeneity of human populations in different endemic areas, and of course, the complexity of MHC genes. Therefore, since population-based cohorts with a single P. vivax infection represent a valuable but uncommon resource for genetic studies, more sophisticated analytical approaches are needed to study the expression of MHC genes in such different exposure conditions to determine the precise role of such polymorphisms as determinant for P. vivax susceptibility, immune response, and its outcome in disease progression.
JL-J and LP-R wrote the review.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We are grateful to Heloisa Maria Nogueira Diniz for the figure of Plasmodium vivax lifecycle and to Rodrigo Nunes Rodrigues da Silva for the figure of Map of the human Chromosome 6. We are also grateful to Dr. Claudio Tadeu Daniel-Ribeiro for the critical reading of this review.
The authors are supported by the Brazilian National Research Council – CNPq/PAPES (Conselho Nacional de Desenvolvimento Científico e Tecnológico/Programa de Apoio Pesquisa Estratégica em Saúde), Fiocruz. JL-J is recipient of a FAPERJ-APQ1 (E-26/111.248/2014) and CPNq-Universal research grants (445150/2014-9).
2. Langhorne J, Ndungu FM, Sponaas AM, Marsh K. Immunity to malaria: more questions than answers. Nat Immunol (2008) 9:725–32. doi:10.1038/ni.f.205
3. White NJ, Pukrittayakamee S, Hien TT, Faiz MA, Mokuolu OA, Dondorp AM. Malaria. Lancet (2014) 383:723–35. doi:10.1016/S0140-6736(13)60024-0
4. Schumacher RF, Spinelli E. Malaria in children. Mediterr J Hematol Infect Dis (2012) 4:e2012073. doi:10.4084/MJHID.2012.073
5. Maitland K, Marsh K. Pathophysiology of severe malaria in children. Acta Trop (2004) 90:131–40. doi:10.1016/j.actatropica.2003.11.010
6. Menendez C, Fleming AF, Alonso PL. Malaria-related anaemia. Parasitol Today (2000) 16:469–76. doi:10.1016/S0169-4758(00)01774-9
7. Mangano VD, Modiano D. Host genetics and parasitic infections. Clin Microbiol Infect (2014) 20:1265–75. doi:10.1111/1469-0691.12793
8. Mangano VD, Modiano D. An evolutionary perspective of how infection drives human genome diversity: the case of malaria. Curr Opin Immunol (2014) 30:39–47. doi:10.1016/j.coi.2014.06.004
9. Kwiatkowski DP. How malaria has affected the human genome and what human genetics can teach us about malaria. Am J Hum Genet (2005) 77:171–92. doi:10.1086/432519
10. Haldane J. Disease and Evolution. La Ricerca Scientifica Suppl. A. Roma, Italy (1949). p. 68–76.
11. Yuthavong Y, Wilairat P. Protection against malaria by thalassaemia and haemoglobin variants. Parasitol Today (1993) 9:241–5. doi:10.1016/0169-4758(93)90065-N
12. Cooke BM, Mohandas N, Coppel RL. Malaria and the red blood cell membrane. Semin Hematol (2004) 41:173–88. doi:10.1053/j.seminhematol.2004.01.004
13. Richer J, Chudley AE. The hemoglobinopathies and malaria. Clin Genet (2005) 68:332–6. doi:10.1111/j.1399-0004.2005.00503.x
14. Duffy PE, Fried M. Red blood cells that do and red blood cells that don’t: how to resist a persistent parasite. Trends Parasitol (2006) 22:99–101. doi:10.1016/j.pt.2006.01.009
15. Pasvol G, Weatherall DJ, Wilson RJ. Cellular mechanism for the protective effect of haemoglobin S against P. falciparum malaria. Nature (1978) 274:701–3. doi:10.1038/274701a0
16. Cappadoro M, Giribaldi G, O’Brien E, Turrini F, Mannu F, Ulliers D, et al. Early phagocytosis of glucose-6-phosphate dehydrogenase (G6PD)-deficient erythrocytes parasitized by Plasmodium falciparum may explain malaria protection in G6PD deficiency. Blood (1998) 92:2527–34.
17. Ayi K, Turrini F, Piga A, Arese P. Enhanced phagocytosis of ring-parasitized mutant erythrocytes: a common mechanism that may explain protection against falciparum malaria in sickle trait and beta-thalassemia trait. Blood (2004) 104:3364–71. doi:10.1182/blood-2003-11-3820
18. Allen SJ, O’Donnell A, Alexander ND, Alpers MP, Peto TE, Clegg JB, et al. alpha+-Thalassemia protects children against disease caused by other infections as well as malaria. Proc Natl Acad Sci U S A (1997) 94:14736–41. doi:10.1073/pnas.94.26.14736
19. Williams TN, Mwangi TW, Wambua S, Alexander ND, Kortok M, Snow RW, et al. Sickle cell trait and the risk of Plasmodium falciparum malaria and other childhood diseases. J Infect Dis (2005) 192:178–86. doi:10.1086/430744
20. Wambua S, Mwangi TW, Kortok M, Uyoga SM, Macharia AW, Mwacharo JK, et al. The effect of alpha+-thalassaemia on the incidence of malaria and other diseases in children living on the coast of Kenya. PLoS Med (2006) 3:e158. doi:10.1371/journal.pmed.0030158
21. Willcox M, Bjorkman A, Brohult J. Falciparum malaria and beta-thalassaemia trait in northern Liberia. Ann Trop Med Parasitol (1983) 77:335–47.
22. Friedman MJ. Erythrocytic mechanism of sickle cell resistance to malaria. Proc Natl Acad Sci U S A (1978) 75:1994–7. doi:10.1073/pnas.75.4.1994
23. Ackerman H, Usen S, Jallow M, Sisay-Joof F, Pinder M, Kwiatkowski DP. A comparison of case-control and family-based association methods: the example of sickle-cell and malaria. Ann Hum Genet (2005) 69:559–65. doi:10.1111/j.1529-8817.2005.00180.x
24. Friedman MJ, Roth EF, Nagel RL, Trager W. The role of hemoglobins C, S, and Nbalt in the inhibition of malaria parasite development in vitro. Am J Trop Med Hyg (1979) 28:777–80.
25. Rihet P, Flori L, Tall F, Traore AS, Fumoux F. Hemoglobin C is associated with reduced Plasmodium falciparum parasitemia and low risk of mild malaria attack. Hum Mol Genet (2004) 13:1–6. doi:10.1093/hmg/ddh002
26. Bunyaratvej A, Butthep P, Sae-Ung N, Fucharoen S, Yuthavong Y. Reduced deformability of thalassemic erythrocytes and erythrocytes with abnormal hemoglobins and relation with susceptibility to Plasmodium falciparum invasion. Blood (1992) 79:2460–3.
27. Chotivanich K, Udomsangpetch R, Pattanapanyasat K, Chierakul W, Simpson J, Looareesuwan S, et al. Hemoglobin E: a balanced polymorphism protective against high parasitemias and thus severe P falciparum malaria. Blood (2002) 100:1172–6.
28. Roth EF Jr, Raventos-Suarez C, Rinaldi A, Nagel RL. Glucose-6-phosphate dehydrogenase deficiency inhibits in vitro growth of Plasmodium falciparum. Proc Natl Acad Sci U S A (1983) 80:298–9. doi:10.1073/pnas.80.1.298
29. Guindo A, Fairhurst RM, Doumbo OK, Wellems TE, Diallo DA. X-linked G6PD deficiency protects hemizygous males but not heterozygous females against severe malaria. PLoS Med (2007) 4:e66. doi:10.1371/journal.pmed.0040066
30. Ouattara AK, Bisseye C, Bazie BV, Diarra B, Compaore TR, Djigma F, et al. Glucose-6-phosphate dehydrogenase (G6PD) deficiency is associated with asymptomatic malaria in a rural community in Burkina Faso. Asian Pac J Trop Biomed (2014) 4:655–8. doi:10.12980/APJTB.4.2014APJTB-2014-0100
31. Ayi K, Min-Oo G, Serghides L, Crockett M, Kirby-Allen M, Quirt I, et al. Pyruvate kinase deficiency and malaria. N Engl J Med (2008) 358:1805–10. doi:10.1056/NEJMoa072464
32. Allen SJ, O’Donnell A, Alexander ND, Mgone CS, Peto TE, Clegg JB, et al. Prevention of cerebral malaria in children in Papua New Guinea by southeast Asian ovalocytosis band 3. Am J Trop Med Hyg (1999) 60:1056–60.
33. Genton B, al-Yaman F, Mgone CS, Alexander N, Paniu MM, Alpers MP, et al. Ovalocytosis and cerebral malaria. Nature (1995) 378:564–5. doi:10.1038/378564a0
34. Rosanas-Urgell A, Lin E, Manning L, Rarau P, Laman M, Senn N, et al. Reduced risk of Plasmodium vivax malaria in Papua New Guinean children with Southeast Asian ovalocytosis in two cohorts and a case-control study. PLoS Med (2012) 9:e1001305. doi:10.1371/journal.pmed.1001305
35. Miller LH, Mason SJ, Clyde DF, McGinniss MH. The resistance factor to Plasmodium vivax in blacks. The Duffy-blood-group genotype, FyFy. N Engl J Med (1976) 295:302–4. doi:10.1056/NEJM197608052950602
36. Spencer HC, Miller LH, Collins WE, Knud-Hansen C, McGinnis MH, Shiroishi T, et al. The Duffy blood group and resistance to Plasmodium vivax in Honduras. Am J Trop Med Hyg (1978) 27:664–70.
37. Mendes C, Dias F, Figueiredo J, Mora VG, Cano J, de Sousa B, et al. Duffy negative antigen is no longer a barrier to Plasmodium vivax – molecular evidences from the African West Coast (Angola and Equatorial Guinea). PLoS Negl Trop Dis (2011) 5:e1192. doi:10.1371/journal.pntd.0001192
38. Woldearegai TG, Kremsner PG, Kun JF, Mordmuller B. Plasmodium vivax malaria in Duffy-negative individuals from Ethiopia. Trans R Soc Trop Med Hyg (2013) 107:328–31. doi:10.1093/trstmh/trt016
39. Hill AV, Allsopp CE, Kwiatkowski D, Anstey NM, Twumasi P, Rowe PA, et al. Common west African HLA antigens are associated with protection from severe malaria. Nature (1991) 352:595–600. doi:10.1038/352595a0
40. Hill AV, Bennett S, Allsopp CE, Kwiatkowski D, Anstey NM, Twumasi P, et al. HLA, malaria and dominant protective associations. Parasitol Today (1992) 8:57. doi:10.1016/0169-4758(92)90089-K
41. Wilkinson RJ, Pasvol G. Host resistance to malaria runs into swampy water. Trends Microbiol (1997) 5:213–5. doi:10.1016/S0966-842X(97)01053-6
42. Gilbert SC, Plebanski M, Gupta S, Morris J, Cox M, Aidoo M, et al. Association of malaria parasite population structure, HLA, and immunological antagonism. Science (1998) 279:1173–7. doi:10.1126/science.279.5354.1173
43. Banic DM, Goldberg AC, Pratt-Riccio LR, De Oliveira-Ferreira J, Santos F, Gras-Masse H, et al. Human leukocyte antigen class II control of the immune response to p126-derived amino terminal peptide from Plasmodium falciparum. Am J Trop Med Hyg (2002) 66:509–15.
44. Pratt-Riccio LR, Lima-Junior JC, Carvalho LJ, Theisen M, Espindola-Mendes EC, Santos F, et al. Antibody response profiles induced by Plasmodium falciparum glutamate-rich protein in naturally exposed individuals from a Brazilian area endemic for malaria. Am J Trop Med Hyg (2005) 73:1096–103.
45. Thathy V, Moulds JM, Guyah B, Otieno W, Stoute JA. Complement receptor 1 polymorphisms associated with resistance to severe malaria in Kenya. Malar J (2005) 4:54. doi:10.1186/1475-2875-4-54
46. Hobbs MR, Udhayakumar V, Levesque MC, Booth J, Roberts JM, Tkachuk AN, et al. A new NOS2 promoter polymorphism associated with increased nitric oxide production and protection from severe malaria in Tanzanian and Kenyan children. Lancet (2002) 360:1468–75. doi:10.1016/S0140-6736(02)11474-7
47. Planche T, Macallan DC, Sobande T, Borrmann S, Kun JF, Krishna S, et al. Nitric oxide generation in children with malaria and the NOS2G-954C promoter polymorphism. Am J Physiol Regul Integr Comp Physiol (2010) 299:R1248–53. doi:10.1152/ajpregu.00390.2010
48. Clark TG, Diakite M, Auburn S, Campino S, Fry AE, Green A, et al. Tumor necrosis factor and lymphotoxin-alpha polymorphisms and severe malaria in African populations. J Infect Dis (2009) 199:569–75. doi:10.1086/596320
49. Hananantachai H, Patarapotikul J, Ohashi J, Naka I, Krudsood S, Looareesuwan S, et al. Significant association between TNF-alpha (TNF) promoter allele (-1031C, -863C, and -857C) and cerebral malaria in Thailand. Tissue Antigens (2007) 69:277–80. doi:10.1111/j.1399-0039.2006.00756.x
50. Wattavidanage J, Carter R, Perera KL, Munasingha A, Bandara S, McGuinness D, et al. TNFalpha*2 marks high risk of severe disease during Plasmodium falciparum malaria and other infections in Sri Lankans. Clin Exp Immunol (1999) 115:350–5. doi:10.1046/j.1365-2249.1999.00804.x
51. McGuire W, Hill AV, Allsopp CE, Greenwood BM, Kwiatkowski D. Variation in the TNF-alpha promoter region associated with susceptibility to cerebral malaria. Nature (1994) 371:508–10. doi:10.1038/371508a0
52. Kanchan K, Jha P, Pati SS, Mohanty S, Mishra SK, Sharma SK, et al. Interferon-gamma (IFNG) microsatellite repeat and single nucleotide polymorphism haplotypes of IFN-alpha receptor (IFNAR1) associated with enhanced malaria susceptibility in Indian populations. Infect Genet Evol (2015) 29:6–14. doi:10.1016/j.meegid.2014.10.030
53. Cabantous S, Ranque S, Poudiougou B, Traore A, Berbache S, Vitte J, et al. Genotype combinations of two IL4 polymorphisms influencing IL-4 plasma levels are associated with different risks of severe malaria in the Malian population. Immunogenetics (2015) 67:283–8. doi:10.1007/s00251-015-0836-3
54. Luoni G, Verra F, Arca B, Sirima BS, Troye-Blomberg M, Coluzzi M, et al. Antimalarial antibody levels and IL4 polymorphism in the Fulani of West Africa. Genes Immun (2001) 2:411–4. doi:10.1038/sj.gene.6363797
55. Wilson JN, Rockett K, Jallow M, Pinder M, Sisay-Joof F, Newport M, et al. Analysis of IL10 haplotypic associations with severe malaria. Genes Immun (2005) 6:462–6. doi:10.1038/sj.gene.6364227
56. Ouma C, Davenport GC, Were T, Otieno MF, Hittner JB, Vulule JM, et al. Haplotypes of IL-10 promoter variants are associated with susceptibility to severe malarial anemia and functional changes in IL-10 production. Hum Genet (2008) 124:515–24. doi:10.1007/s00439-008-0578-5
57. Baird JK. Evidence and implications of mortality associated with acute Plasmodium vivax malaria. Clin Microbiol Rev (2013) 26:36–57. doi:10.1128/CMR.00074-12
58. Galinski MR, Barnwell JW. Plasmodium vivax: who cares? Malar J (2008) 7(Suppl 1):S9. doi:10.1186/1475-2875-7-S1-S9
59. Welch SG, McGregor IA, Williams K. The Duffy blood group and malaria prevalence in Gambian West Africans. Trans R Soc Trop Med Hyg (1977) 71:295–6. doi:10.1016/0035-9203(77)90102-X
60. Nardin E, Zavala F, Nussenzweig V, Nussenzweig RS. Pre-erythrocytic malaria vaccine: mechanisms of protective immunity and human vaccine trials. Parassitologia (1999) 41:397–402.
61. Luo S, Liu D, Ye B, Shu H, Fu R. Effect of monoclonal antibodies on the entry and development of Plasmodium vivax sporozoite in cultured cells. Zhongguo Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi (1995) 13:284–9.
62. Millet P, Chizzolini C, Wirtz RA, Bathurst I, Broderson JR, Campbell GH, et al. Inhibitory activity against sporozoites induced by antibodies directed against nonrepetitive regions of the circumsporozoite protein of Plasmodium vivax. Eur J Immunol (1992) 22:519–24. doi:10.1002/eji.1830220234
63. Hollingdale MR, Nardin EH, Tharavanij S, Schwartz AL, Nussenzweig RS. Inhibition of entry of Plasmodium falciparum and P. vivax sporozoites into cultured cells; an in vitro assay of protective antibodies. J Immunol (1984) 132:909–13.
64. Doolan DL, Hoffman SL. The complexity of protective immunity against liver-stage malaria. J Immunol (2000) 165:1453–62. doi:10.4049/jimmunol.165.3.1453
65. Cockburn IA, Amino R, Kelemen RK, Kuo SC, Tse SW, Radtke A, et al. In vivo imaging of CD8+ T cell-mediated elimination of malaria liver stages. Proc Natl Acad Sci U S A (2013) 110:9090–5. doi:10.1073/pnas.1303858110
66. Chia WN, Goh YS, Renia L. Novel approaches to identify protective malaria vaccine candidates. Front Microbiol (2014) 5:586. doi:10.3389/fmicb.2014.00586
67. Herrera S, Corradin G, Arevalo-Herrera M. An update on the search for a Plasmodium vivax vaccine. Trends Parasitol (2007) 23:122–8. doi:10.1016/j.pt.2007.01.008
68. Mueller I, Galinski MR, Tsuboi T, Arevalo-Herrera M, Collins WE, King CL. Natural acquisition of immunity to Plasmodium vivax: epidemiological observations and potential targets. Adv Parasitol (2013) 81:77–131. doi:10.1016/B978-0-12-407826-0.00003-5
69. Beck S, Geraghty D, Inoko H, Rowen L, Aguado B, Bahram S, et al. Complete sequence and gene map of a human major histocompatibility complex. The MHC sequencing consortium. Nature (1999) 401:921–3.
70. Goldberg AC, Rizzo LV. MHC structure and function – antigen presentation. Part 1. Einstein (Sao Paulo) (2015) 13:153–6. doi:10.1590/S1679-45082015RB3122
72. Hershberg RM, Cho DH, Youakim A, Bradley MB, Lee JS, Framson PE, et al. Highly polarized HLA class II antigen processing and presentation by human intestinal epithelial cells. J Clin Invest (1998) 102:792–803. doi:10.1172/JCI3201
73. Wu Z, Biro PA, Mirakian R, Hammond L, Curcio F, Ambesi-Impiombato FS, et al. HLA-DMB expression by thyrocytes: indication of the antigen-processing and possible presenting capability of thyroid cells. Clin Exp Immunol (1999) 116:62–9. doi:10.1046/j.1365-2249.1999.00831.x
74. Price P, Calder DM, Witt CS, Allcock RJ, Christiansen FT, Davies GR, et al. Periodontal attachment loss in HIV-infected patients is associated with the major histocompatibility complex 8.1 haplotype (HLA-A1,B8,DR3). Tissue Antigens (1999) 54:391–9. doi:10.1034/j.1399-0039.1999.540409.x
75. Trowsdale J, Knight JC. Major histocompatibility complex genomics and human disease. Annu Rev Genomics Hum Genet (2013) 14:301–23. doi:10.1146/annurev-genom-091212-153455
76. Li CP, Zhou Y, Xiang X, Zhou Y, He M. Relationship of HLA-DRB1 gene polymorphism with susceptibility to pulmonary tuberculosis: updated meta-analysis. Int J Tuberc Lung Dis (2015) 19:841–9. doi:10.5588/ijtld.14.0521
77. Magira EE, Papasteriades C, Kanterakis S, Toubis M, Roussos C, Monos DS. HLA-A and HLA-DRB1 amino acid polymorphisms are associated with susceptibility and protection to pulmonary tuberculosis in a Greek population. Hum Immunol (2012) 73:641–6. doi:10.1016/j.humimm.2012.03.008
78. Haag S, Tuncel J, Thordardottir S, Mason DE, Yau AC, Dobritzsch D, et al. Positional identification of RT1-B (HLA-DQ) as susceptibility locus for autoimmune arthritis. J Immunol (2015) 194:2539–50. doi:10.4049/jimmunol.1402238
79. Hill AV, Elvin J, Willis AC, Aidoo M, Allsopp CE, Gotch FM, et al. Molecular analysis of the association of HLA-B53 and resistance to severe malaria. Nature (1992) 360:434–9. doi:10.1038/360434a0
80. Zavala F, Cochrane AH, Nardin EH, Nussenzweig RS, Nussenzweig V. Circumsporozoite proteins of malaria parasites contain a single immunodominant region with two or more identical epitopes. J Exp Med (1983) 157:1947–57. doi:10.1084/jem.157.6.1947
81. Qari SH, Shi YP, Goldman IF, Udhayakumar V, Alpers MP, Collins WE, et al. Identification of Plasmodium vivax-like human malaria parasite. Lancet (1993) 341:780–3. doi:10.1016/0140-6736(93)90559-Y
82. Qari SH, Goldman IF, Povoa MM, Oliveira S, Alpers MP, Lal AA. Wide distribution of the variant form of the human malaria parasite Plasmodium vivax. J Biol Chem (1991) 266:16297–300.
83. Oliveira-Ferreira J, Pratt-Riccio LR, Arruda M, Santos F, Ribeiro CT, Goldberg AC, et al. HLA class II and antibody responses to circumsporozoite protein repeats of P. vivax (VK210, VK247 and P. vivax-like) in individuals naturally exposed to malaria. Acta Trop (2004) 92:63–9. doi:10.1016/j.actatropica.2004.02.011
84. Storti-Melo LM, da Costa DR, Souza-Neiras WC, Cassiano GC, Couto VS, Povoa MM, et al. Influence of HLA-DRB-1 alleles on the production of antibody against CSP, MSP-1, AMA-1, and DBP in Brazilian individuals naturally infected with Plasmodium vivax. Acta Trop (2012) 121:152–5. doi:10.1016/j.actatropica.2011.10.009
85. Lima-Junior JC, Rodrigues-da-Silva RN, Banic DM, Jiang J, Singh B, Fabricio-Silva GM, et al. Influence of HLA-DRB1 and HLA-DQB1 alleles on IgG antibody response to the P. vivax MSP-1, MSP-3alpha and MSP-9 in individuals from Brazilian endemic area. PLoS One (2012) 7:e36419. doi:10.1371/journal.pone.0036419
86. Ferreira AR, Singh B, Cabrera-Mora M, Magri De Souza AC, Queiroz Marques MT, Porto LC, et al. Evaluation of naturally acquired IgG antibodies to a chimeric and non-chimeric recombinant species of Plasmodium vivax reticulocyte binding protein-1: lack of association with HLA-DRB1*/DQB1* in malaria exposed individuals from the Brazilian Amazon. PLoS One (2014) 9:e105828. doi:10.1371/journal.pone.0105828
87. Martinez P, Lopez C, Saravia C, Vanegas M, Patarroyo MA. Evaluation of the antigenicity of universal epitopes from PvDBPII in individuals exposed to Plasmodium vivax malaria. Microbes Infect (2010) 12:1188–97. doi:10.1016/j.micinf.2010.08.007
88. Soares IS, Levitus G, Souza JM, Del Portillo HA, Rodrigues MM. Acquired immune responses to the N- and C-terminal regions of Plasmodium vivax merozoite surface protein 1 in individuals exposed to malaria. Infect Immun (1997) 65:1606–14.
89. Galinski MR, Corredor-Medina C, Povoa M, Crosby J, Ingravallo P, Barnwell JW. Plasmodium vivax merozoite surface protein-3 contains coiled-coil motifs in an alanine-rich central domain. Mol Biochem Parasitol (1999) 101:131–47. doi:10.1016/S0166-6851(99)00063-8
90. de Oliveira CI, Wunderlich G, Levitus G, Soares IS, Rodrigues MM, Tsuji M, et al. Antigenic properties of the merozoite surface protein 1 gene of Plasmodium vivax. Vaccine (1999) 17:2959–68. doi:10.1016/S0264-410X(99)00176-0
91. Barnwell JW, Galinski MR, DeSimone SG, Perler F, Ingravallo P. Plasmodium vivax, P. cynomolgi, and P. knowlesi: identification of homologue proteins associated with the surface of merozoites. Exp Parasitol (1999) 91:238–49. doi:10.1006/expr.1998.4372
92. Oliveira-Ferreira J, Vargas-Serrato E, Barnwell JW, Moreno A, Galinski MR. Immunogenicity of Plasmodium vivax merozoite surface protein-9 recombinant proteins expressed in E. coli. Vaccine (2004) 22:2023–30. doi:10.1016/j.vaccine.2003.07.021
93. Soares IS, Rodrigues MM. Immunogenic properties of the Plasmodium vivax vaccine candidate MSP1(19) expressed as a secreted non-glycosylated polypeptide from Pichia pastoris. Parasitology (2002) 124:237–46. doi:10.1017/S003118200100110X
94. Lima-Junior JC, Jiang J, Rodrigues-da-Silva RN, Banic DM, Tran TM, Ribeiro RY, et al. B cell epitope mapping and characterization of naturally acquired antibodies to the Plasmodium vivax merozoite surface protein-3alpha (PvMSP-3alpha) in malaria exposed individuals from Brazilian Amazon. Vaccine (2011) 29:1801–11. doi:10.1016/j.vaccine.2010.12.099
95. Lima-Junior JC, Tran TM, Meyer EV, Singh B, De-Simone SG, Santos F, et al. Naturally acquired humoral and cellular immune responses to Plasmodium vivax merozoite surface protein 9 in Northwestern Amazon individuals. Vaccine (2008) 26:6645–54. doi:10.1016/j.vaccine.2008.09.029
96. Arevalo-Herrera M, Soto L, Perlaza BL, Cespedes N, Vera O, Lenis AM, et al. Antibody-mediated and cellular immune responses induced in naive volunteers by vaccination with long synthetic peptides derived from the Plasmodium vivax circumsporozoite protein. Am J Trop Med Hyg (2011) 84:35–42. doi:10.4269/ajtmh.2011.09-0507
97. Lima-Junior JC, Banic DM, Tran TM, Meyer VS, De-Simone SG, Santos F, et al. Promiscuous T-cell epitopes of Plasmodium merozoite surface protein 9 (PvMSP9) induces IFN-gamma and IL-4 responses in individuals naturally exposed to malaria in the Brazilian Amazon. Vaccine (2010) 28:3185–91. doi:10.1016/j.vaccine.2010.02.046
98. Kwiatkowski D, Hill AV, Sambou I, Twumasi P, Castracane J, Manogue KR, et al. TNF concentration in fatal cerebral, non-fatal cerebral, and uncomplicated Plasmodium falciparum malaria. Lancet (1990) 336:1201–4. doi:10.1016/0140-6736(90)92827-5
99. Wilson AG, Symons JA, McDowell TL, McDevitt HO, Duff GW. Effects of a polymorphism in the human tumor necrosis factor alpha promoter on transcriptional activation. Proc Natl Acad Sci U S A (1997) 94:3195–9. doi:10.1073/pnas.94.7.3195
100. Cabrera M, Shaw MA, Sharples C, Williams H, Castes M, Convit J, et al. Polymorphism in tumor necrosis factor genes associated with mucocutaneous leishmaniasis. J Exp Med (1995) 182:1259–64. doi:10.1084/jem.182.5.1259
101. Sohail M, Kaul A, Bali P, Raziuddin M, Singh MP, Singh OP, et al. Alleles -308A and -1031C in the TNF-alpha gene promoter do not increase the risk but associated with circulating levels of TNF-alpha and clinical features of vivax malaria in Indian patients. Mol Immunol (2008) 45:1682–92. doi:10.1016/j.molimm.2007.10.002
102. Kuesap J, Hirayama K, Kikuchi M, Ruangweerayut R, Na-Bangchang K. Study on association between genetic polymorphisms of haem oxygenase-1, tumour necrosis factor, cadmium exposure and malaria pathogenicity and severity. Malar J (2010) 9:260. doi:10.1186/1475-2875-9-260
103. Mendonca VR, Souza LC, Garcia GC, Magalhaes BM, Lacerda MV, Andrade BB, et al. DDX39B (BAT1), TNF and IL6 gene polymorphisms and association with clinical outcomes of patients with Plasmodium vivax malaria. Malar J (2014) 13:278. doi:10.1186/1475-2875-13-278
104. Sortica VA, Cunha MG, Ohnishi MD, Souza JM, Ribeiro-Dos-Santos AK, Santos NP, et al. IL1B, IL4R, IL12RB1 and TNF gene polymorphisms are associated with Plasmodium vivax malaria in Brazil. Malar J (2012) 11:409. doi:10.1186/1475-2875-11-409
105. Allcock RJ, Williams JH, Price P. The central MHC gene, BAT1, may encode a protein that down-regulates cytokine production. Genes Cells (2001) 6:487–94. doi:10.1046/j.1365-2443.2001.00435.x
106. Afridi S, Atkinson A, Garnier S, Fumoux F, Rihet P. Malaria resistance genes are associated with the levels of IgG subclasses directed against Plasmodium falciparum blood-stage antigens in Burkina Faso. Malar J (2012) 11:308. doi:10.1186/1475-2875-11-308
107. da Silva Santos S, Clark TG, Campino S, Suarez-Mutis MC, Rockett KA, Kwiatkowski DP, et al. Investigation of host candidate malaria-associated risk/protective SNPs in a Brazilian Amazonian population. PLoS One (2012) 7:e36692. doi:10.1371/journal.pone.0036692
108. van der Merwe PA, Davis SJ. Molecular interactions mediating T cell antigen recognition. Annu Rev Immunol (2003) 21:659–84. doi:10.1146/annurev.immunol.21.120601.141036
109. Lin HH, Zhang GL, Tongchusak S, Reinherz EL, Brusic V. Evaluation of MHC-II peptide binding prediction servers: applications for vaccine research. BMC Bioinformatics (2008) 9(Suppl 12):S22. doi:10.1186/1471-2105-9-S12-S22
110. Alves CR, Pontes de Carvalho LC, Souza AL, De Simone SG. A strategy for the identification of T-cell epitopes on Leishmania cysteine proteinases. Cytobios (2001) 104:33–41.
111. Mustafa AS. Th1 cell reactivity and HLA-DR binding prediction for promiscuous recognition of MPT63 (Rv1926c), a major secreted protein of Mycobacterium tuberculosis. Scand J Immunol (2009) 69:213–22. doi:10.1111/j.1365-3083.2008.02221.x
112. Fonseca SG, Coutinho-Silva A, Fonseca LA, Segurado AC, Moraes SL, Rodrigues H, et al. Identification of novel consensus CD4 T-cell epitopes from clade B HIV-1 whole genome that are frequently recognized by HIV-1 infected patients. AIDS (2006) 20:2263–73. doi:10.1097/01.aids.0000253353.48331.5f
113. Carlton JM, Escalante AA, Neafsey D, Volkman SK. Comparative evolutionary genomics of human malaria parasites. Trends Parasitol (2008) 24:545–50. doi:10.1016/j.pt.2008.09.003
114. Carlton JM, Adams JH, Silva JC, Bidwell SL, Lorenzi H, Caler E, et al. Comparative genomics of the neglected human malaria parasite Plasmodium vivax. Nature (2008) 455:757–63. doi:10.1038/nature07327
115. Doolan DL, Southwood S, Freilich DA, Sidney J, Graber NL, Shatney L, et al. Identification of Plasmodium falciparum antigens by antigenic analysis of genomic and proteomic data. Proc Natl Acad Sci U S A (2003) 100:9952–7. doi:10.1073/pnas.1633254100
116. Doolan DL, Southwood S, Chesnut R, Appella E, Gomez E, Richards A, et al. HLA-DR-promiscuous T cell epitopes from Plasmodium falciparum pre-erythrocytic-stage antigens restricted by multiple HLA class II alleles. J Immunol (2000) 165:1123–37. doi:10.4049/jimmunol.165.2.1123
117. Moreno A, Clavijo P, Edelman R, Davis J, Sztein M, Sinigaglia F, et al. CD4+ T cell clones obtained from Plasmodium falciparum sporozoite-immunized volunteers recognize polymorphic sequences of the circumsporozoite protein. J Immunol (1993) 151:489–99.
118. Parra-Lopez C, Calvo-Calle JM, Cameron TO, Vargas LE, Salazar LM, Patarroyo ME, et al. Major histocompatibility complex and T cell interactions of a universal T cell epitope from Plasmodium falciparum circumsporozoite protein. J Biol Chem (2006) 281:14907–17. doi:10.1074/jbc.M511571200
119. Daubenberger CA, Nickel B, Ciatto C, Grutter MG, Poltl-Frank F, Rossi L, et al. Amino acid dimorphism and parasite immune evasion: cellular immune responses to a promiscuous epitope of Plasmodium falciparum merozoite surface protein 1 displaying dimorphic amino acid polymorphism are highly constrained. Eur J Immunol (2002) 32:3667–77. doi:10.1002/1521-4141(200212)32:12<3667::AID-IMMU3667>3.0.CO;2-C
120. Ohta N, Iwaki K, Itoh M, Fu J, Nakashima S, Hato M, et al. Epitope analysis of human T-cell response to MSP-1 of Plasmodium falciparum in malaria-nonexposed individuals. Int Arch Allergy Immunol (1997) 114:15–22. doi:10.1159/000237637
121. Vargas LE, Parra CA, Salazar LM, Guzman F, Pinto M, Patarroyo ME. MHC allele-specific binding of a malaria peptide makes it become promiscuous on fitting a glycine residue into pocket 6. Biochem Biophys Res Commun (2003) 307:148–56. doi:10.1016/S0006-291X(03)01129-X
122. Contreras CE, Ploton IN, Siliciano RF, Karp CL, Viscidi R, Kumar N. Mapping of specific and promiscuous HLA-DR-restricted T-cell epitopes on the Plasmodium falciparum 27-kilodalton sexual stage-specific antigen. Infect Immun (1998) 66:3579–90.
123. Caro-Aguilar I, Rodriguez A, Calvo-Calle JM, Guzman F, De la Vega P, Patarroyo ME, et al. Plasmodium vivax promiscuous T-helper epitopes defined and evaluated as linear peptide chimera immunogens. Infect Immun (2002) 70:3479–92. doi:10.1128/IAI.70.7.3479-3492.2002
124. Herrera MA, de Plata C, Gonzalez JM, Corradin G, Herrera S. Immunogenicity of multiple antigen peptides containing Plasmodium vivax CS epitopes in BALB/c mice. Mem Inst Oswaldo Cruz (1994) 89(Suppl 2):71–6. doi:10.1590/S0074-02761994000600017
125. Fern J, Good MF. Promiscuous malaria peptide epitope stimulates CD45Ra T cells from peripheral blood of nonexposed donors. J Immunol (1992) 148:907–13.
126. Wiwanitkit V. Predicted epitopes of malarial merozoite surface protein 1 by bioinformatics method: a clue for further vaccine development. J Microbiol Immunol Infect (2009) 42:19–21.
127. Nanda Kumar Y, Jeyakodi G, Gunasekaran K, Jambulingam P. Computational screening and characterization of putative vaccine candidates of Plasmodium vivax. J Biomol Struct Dyn (2015) 29:1–59. doi:10.1080/07391102.2015.1090344
128. Saravia C, Martinez P, Granados DS, Lopez C, Reyes C, Patarroyo MA. Identification and evaluation of universal epitopes in Plasmodium vivax Duffy binding protein. Biochem Biophys Res Commun (2008) 377:1279–83. doi:10.1016/j.bbrc.2008.10.153
129. Cespedes N, Habel C, Lopez-Perez M, Castellanos A, Kajava AV, Servis C, et al. Plasmodium vivax antigen discovery based on alpha-helical coiled coil protein motif. PLoS One (2014) 9:e100440. doi:10.1371/journal.pone.0100440
130. Jiang L, Wang ZY, Ma XJ, Zhang XP, Cai L. [Comparative analysis of nucleotide sequences of lactate dehydrogenase (LDH) gene and LDH epitopes of Plasmodium vivax and Plasmodium falciparum]. Zhongguo Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi (2010) 28:103–7.
131. Rammensee H, Bachmann J, Emmerich NP, Bachor OA, Stevanovic S. SYFPEITHI: database for MHC ligands and peptide motifs. Immunogenetics (1999) 50:213–9. doi:10.1007/s002510050595
132. Sturniolo T, Bono E, Ding J, Raddrizzani L, Tuereci O, Sahin U, et al. Generation of tissue-specific and promiscuous HLA ligand databases using DNA microarrays and virtual HLA class II matrices. Nat Biotechnol (1999) 17:555–61. doi:10.1038/9858
133. Singh H, Raghava GP. ProPred: prediction of HLA-DR binding sites. Bioinformatics (2001) 17:1236–7. doi:10.1093/bioinformatics/17.12.1236
134. Rosa DS, Iwai LK, Tzelepis F, Bargieri DY, Medeiros MA, Soares IS, et al. Immunogenicity of a recombinant protein containing the Plasmodium vivax vaccine candidate MSP1(19) and two human CD4+ T-cell epitopes administered to non-human primates (Callithrix jacchus jacchus). Microbes Infect (2006) 8:2130–7. doi:10.1016/j.micinf.2006.03.012
135. Reche PA, Glutting JP, Reinherz EL. Prediction of MHC class I binding peptides using profile motifs. Hum Immunol (2002) 63:701–9. doi:10.1016/S0198-8859(02)00432-9
136. Zhang GL, Khan AM, Srinivasan KN, August JT, Brusic V. MULTIPRED: a computational system for prediction of promiscuous HLA binding peptides. Nucleic Acids Res (2005) 33:W172–9. doi:10.1093/nar/gki452
137. Nielsen M, Lundegaard C, Blicher T, Peters B, Sette A, Justesen S, et al. Quantitative predictions of peptide binding to any HLA-DR molecule of known sequence: NetMHCIIpan. PLoS Comput Biol (2008) 4:e1000107. doi:10.1371/journal.pcbi.1000107
138. Sedegah M, Kim Y, Peters B, McGrath S, Ganeshan H, Lejano J, et al. Identification and localization of minimal MHC-restricted CD8+ T cell epitopes within the Plasmodium falciparum AMA1 protein. Malar J (2010) 9:241. doi:10.1186/1475-2875-9-241
139. Atanasova M, Patronov A, Dimitrov I, Flower DR, Doytchinova I. EpiDOCK: a molecular docking-based tool for MHC class II binding prediction. Protein Eng Des Sel (2013) 26:631–4. doi:10.1093/protein/gzt018
140. Kim Y, Ponomarenko J, Zhu Z, Tamang D, Wang P, Greenbaum J, et al. Immune epitope database analysis resource. Nucleic Acids Res (2012) 40:W525–30. doi:10.1093/nar/gks438
Keywords: malaria, P.vivax, MHC, HLA, vaccine
Citation: Lima-Junior JC and Pratt-Riccio LR (2016) Major Histocompatibility Complex and Malaria: Focus on Plasmodium vivax Infection. Front. Immunol. 7:13. doi: 10.3389/fimmu.2016.00013
Received: 17 November 2015; Accepted: 12 January 2016;
Published: 27 January 2016
Edited by:
Alexandre Morrot, Federal University of Rio de Janeiro, BrazilReviewed by:
Hari Mohan Saxena, Guru Angad Dev Veterinary and Animal Sciences University, IndiaCopyright: © 2016 Lima-Junior and Pratt-Riccio. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Josué da Costa Lima-Junior, am9zdWVAaW9jLmZpb2NydXouYnI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.