 Vera Rocha-Perugini
Vera Rocha-Perugini Francisco Sánchez-Madrid
Francisco Sánchez-Madrid Gloria Martínez del Hoyo
Gloria Martínez del Hoyo- 1Servicio de Inmunología, Instituto de Investigación Sanitaria La Princesa, Hospital de la Princesa, Madrid, Spain
- 2Vascular Pathophysiology Area, Centro Nacional de Investigaciones Cardiovasculares Carlos III (CNIC), Madrid, Spain
Tetraspanin-enriched microdomains (TEMs) are specialized membrane platforms driven by protein–protein interactions that integrate membrane receptors and adhesion molecules. Tetraspanins participate in antigen recognition and presentation by antigen-presenting cells (APCs) through the organization of pattern-recognition receptors (PRRs) and their downstream-induced signaling, as well as the regulation of MHC-II–peptide trafficking. T lymphocyte activation is triggered upon specific recognition of antigens present on the APC surface during immunological synapse (IS) formation. This dynamic process is characterized by a defined spatial organization involving the compartmentalization of receptors and adhesion molecules in specialized membrane domains that are connected to the underlying cytoskeleton and signaling molecules. Tetraspanins contribute to the spatial organization and maturation of the IS by controlling receptor clustering and local accumulation of adhesion receptors and integrins, their downstream signaling, and linkage to the actin cytoskeleton. This review offers a perspective on the important role of TEMs in the regulation of antigen recognition and presentation and in the dynamics of IS architectural organization.
Tetraspanin-Enriched Microdomains
Tetraspanins comprise a family of small proteins with four transmembrane domains and are present on the plasma membrane and intracellular vesicles of virtually all mammalian cells. The tetraspanins CD9, CD63, CD81, CD82, and CD151 have a broad tissue distribution, whereas others are restricted to particular tissues, such as TSSC6, CD37, and CD53, in hematopoietic cells (1). Tetraspanins have small and large extracellular loops (SEL and LEL, respectively) and short N- and C-terminal intracellular tails (2). The LEL domain mediates specific protein–protein interactions with laterally associated proteins and a few known ligands, while the cytoplasmic regions provide links to cytoskeletal and signaling molecules (3). Tetraspanins organize a type of cell surface membrane microdomain, known as tetraspanin-enriched microdomains (TEMs) (2, 4), based on their exceptional ability to form multimolecular complexes. Studies using novel advanced microscopy techniques in the intact membranes of living cells have provided a more complete picture of the supramolecular organization of these microdomains (5). The diversity of TEM composition is reflected by different interaction levels, in which each tetraspanin recruits one or more partner proteins forming direct and stable primary complexes, which are assembled through tetraspanin–tetraspanin interactions to form larger complexes that can vary depending on the cell type (6). However, this classical view of TEMs has recently been challenged. Super-resolution microscopy has shown that, in B cells and dendritic cells (DCs), CD53 and CD37 single clusters overlap only to a minor extent with CD81 or CD82 clusters. Moreover, CD53 and CD81 nanoclusters are in closer proximity to their partners MHC class II (MHC-II) and CD19 than to other tetraspanins (7). Additional research using super-resolution microscopy is necessary to dissect the spatial and temporal organization of TEMs in different systems.
In the context of the immune system, TEMs regulate important processes including antigen (Ag) recognition and presentation, protein trafficking, cell proliferation, and leukocyte extravasation (1). All cells of the immune system express tetraspanins, although the tetraspanin repertoire differs between cell types (3). Several receptors responsible for immune cell functions, like the Ag receptors T-cell receptor (TCR) and B-cell receptor (BCR), pathogen receptors, and MHC molecules, are included in TEMs; furthermore, both ubiquitously expressed tetraspanins such as CD81 and immune cell-specific tetraspanins such as CD37 have been shown to be important for immunity (1). In human T lymphocytes, tetraspanins CD9, CD53, CD81, and CD82 act as costimulatory molecules (8–13), and this activity is independent of the classic CD28 costimulatory pathway (12–16). T cells from mice lacking tetraspanins CD81, CD151, CD37, or Tssc6 are hyperproliferative (17–20), and CD37- and CD81-deficient mice have impaired T-cell-dependent immune responses (17, 21–23). Moreover, CD81 expression in both T and B cells is essential for T-cell activation and proper Th2 responses (24–26).
Tetraspanins are also involved in the process of leukocyte extravasation. CD81 controls integrin α4β1 avidity, being essential for monocyte and B cell adhesion under shear flow (27), and CD9 regulates LFA-1-mediated T cell adhesion under flow conditions (28). Moreover, monocyte and T cell transmigration across brain endothelial cell monolayers is significantly reduced by monoclonal antibodies against CD81 in rodent and human in vitro models (29). This inhibitory effect was driven by CD81 expressed in both leukocytes and endothelial cells (29). Transmigrated eosinophils exhibit reduced CD9 expression levels, and their adhesion properties are inhibited by antibodies against CD9 (30, 31). In endothelial cells, various adhesion receptors are included in preassembled tetraspanin-based endothelial adhesive platforms; these platforms coalesce at docking structures for adherent leukocytes during the transmigration process (32, 33).
Immune cells, such as T cells, B cells, and DCs, can release extracellular vesicles that are an important vehicle for intercellular communication and have a role in the regulation of the immune response by different mechanisms (34). Tetraspanins, especially CD9, CD63, and CD81, are highly enriched in extracellular vesicles and have been widely used as exosomal markers. Importantly, growing evidence suggests a functional role for tetraspanins in the biogenesis, targeting, and function of extracellular vesicles (35). In particular, high throughput quantitative proteomic approaches have demonstrated that exosomes from CD81−/− mouse T lymphoblasts show an impaired inclusion of CD81 partners, including MHC molecules, BCR, ICAM-1, and Rac (36).
Together, all these observations indicate that tetraspanins influence many aspects of cellular immunity, sometimes exerting antagonistic roles, and may provide a means of manipulating the immune response for potential therapeutic applications.
The Immunological Relevance of Tetraspanins in Antigen-Presenting Cells
TEMs and Antigen Recognition: Interaction with Pattern-Recognition Receptors
The plasma membrane of antigen-presenting cells (APCs) contains specialized membrane microdomains that organize the spatial distribution of MHC and associated proteins, pattern-recognition receptors (PRRs), and integrins, which are essential for efficient Ag recognition, presentation, and ultimately the activation of the T cell. APCs express a broad repertoire of specific receptors involved in the recognition and uptake of Ags from pathogens, damaged tissues, or tumor cells. In particular, pathogen-derived Ags are recognized by different PRRs that bind to conserved microbial structures called pathogen-associated molecular patterns (PAMPs) (37). The recent identification of specific PRR interactions with tetraspanins has provided new insights into the organization of Ag receptors at the APC membrane and their subsequent downstream signaling (38). In this part, we will revise the recent data that have demonstrated tetraspanin interactions with different receptors involved in Ag recognition (Figure 1).
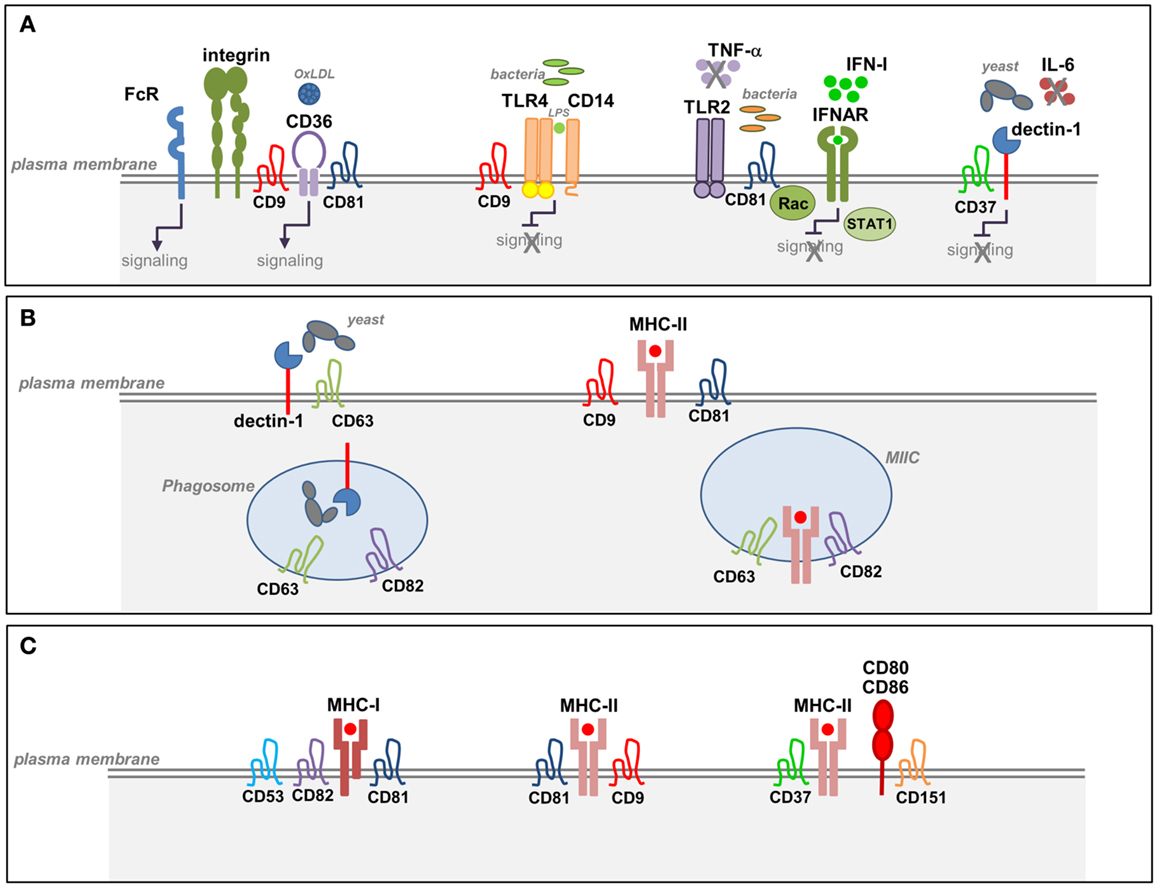
Figure 1. Tetraspanins in the function of APCs. (A) Tetraspanin interactions with pathogen-recognition receptors (PRRs) in APCs. Tetraspanins interact with specific PRRs at the plasma membrane of macrophages and DCs. CD37 associates with dectin-1 and inhibits dectin-1 mediated IL-6 production triggered by the recognition of fungal cell walls. CD9 forms a complex with CD14 and TLR4 and negatively regulates TLR4 signaling in response to LPS. CD81–Rac interaction inhibits TLR2- and IFNAR-signaling pathways and prevents the subsequent activation of STAT1 in response to Listeria monocytogenes. CD36 associates with β1 and β2 integrins and tetraspanins CD9 and CD81 forming a complex that facilitates CD36-signaling and its interaction with FcγRs. Interaction between CD9 and FcγRs promotes phagocytosis and macrophage activation. (B) Tetraspanin interactions during Ag processing and MHC-II biosynthesis. CD63 interacts with dectin-1 in immature DCs and promotes yeast phagocytosis. Both CD63 and CD82 are selectively recruited to yeast-containing phagosomes. CD82 and CD63 are highly enriched in MIIC compartments that contain newly synthesized MHC-II and accessory proteins. (C) Tetraspanin interactions during Ag presentation. Several tetraspanins associate with MHC-I and MHC-II molecules on APCs. Tetraspanins CD9, CD53, CD81, and CD37 associate with MHC-II molecules preferentially at the plasma membrane. MHC-II molecules loaded with a restricted antigenic peptide repertoire are included in TEMs together with accessory molecules and costimulatory molecules. CD9 facilitates MHC-II clustering, and CD151 is involved in the clustering of costimulatory molecules.
Dectin-1 is a C-type lectin receptor (CLR) that recognizes β-glucans in fungal cell walls, triggering phagocytosis, and the antifungal immune response. Dectin-1 signaling is only activated by particulate β-glucans, which cluster the receptor in synapse-like structures from which regulatory tyrosine phosphatases are excluded (39). Two independent studies have demonstrated that CD63 and CD37 interact with dectin-1 on the APC membrane (Figure 1A; Table 1). CD63 associates with dectin-1 in immature DCs and promotes yeast phagocytosis (40) (Table 1). CD37 stabilizes dectin-1 at the APC surface, and this interaction has functional consequences since CD37 inhibits dectin-1-mediated IL-6 production in response to zymosan cell wall preparations (41) (Table 1). Moreover, CD37−/− mice are protected against systemic infection with Candida albicans (42). CD63 has also been reported to be selectively recruited to yeast-containing phagosomes (43) (Table 1), and this observation was subsequently extended to CD82 (44) (Figure 1B; Table 1). After pathogen uptake, CD82 is rapidly recruited to the membrane of nascent pathogen-containing phagosomes prior to fusion with lysosomes (44) (Figure 1B).
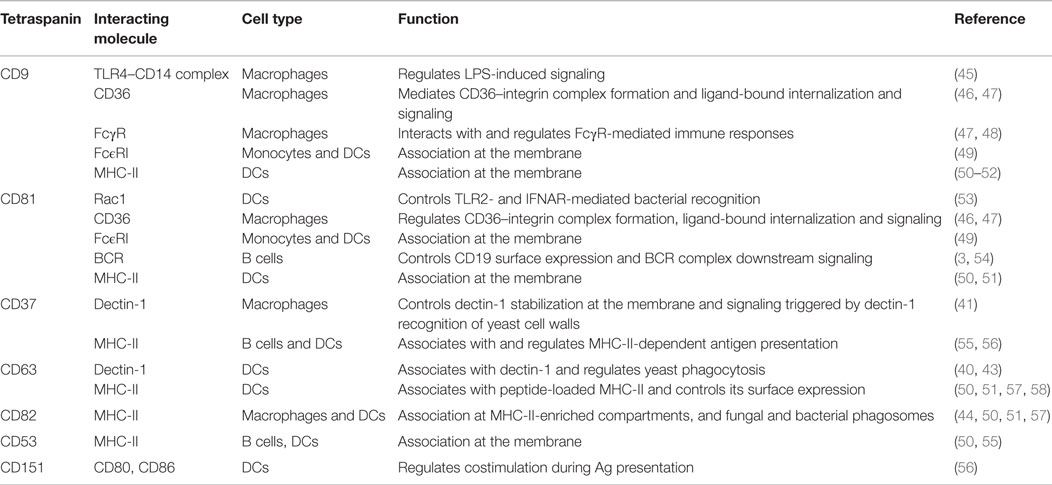
Table 1. Tetraspanin associations in pathogen-recognition receptors and APC functions.
In addition to the reported recruitment of TLR2 and TLR4 to lipid rafts (59–61), other studies demonstrate that TLR4 associates with TEMs. In macrophages, CD9 partly colocalizes with CD14 regulating its expression, its association with TLR4, and the formation of the CD14–TLR4 complex necessary for LPS-induced signaling (45) (Figure 1A; Table 1). Using the Listeria monocytogenes infection model, we recently demonstrated that CD81 is able to interfere with TLR2- and interferon-α/β receptor (IFNAR)-mediated bacterial recognition in DCs, modulating the subsequent CD8+ T cell response (53) (Figure 1A; Table 1). Importantly, CD81−/− mice are protected against lethal systemic Listeria infection. CD81−/− DCs show increased production of proinflammatory mediators and a more efficient activation of protective cytotoxic T cells. This effect is mediated specifically through direct interaction between CD81 and Rac. Indeed, inhibition of CD81–Rac interaction in wild-type DCs using CD81 C-terminal peptides, which block CD81-mediated signaling (62), promotes the same phenotype observed in CD81−/− DCs (53).
In macrophages, CD9 interacts with CD36, a scavenger receptor involved in the recognition of microbes or self-ligands, regulating CD36-mediated uptake of oxidized low-density lipoproteins (46) (Figure 1A; Table 1). CD36 clustering is necessary for the initiation of signal transduction and internalization of receptor–ligand complexes. CD36 was recently shown to form a heteromeric complex containing β1 and β2 integrins and the tetraspanins CD9 and CD81. CD36 inclusion in this complex facilitates its association with ITAM-bearing adaptor Fcγ receptors (FcγR), allowing CD36-dependent Syk activation and the internalization of ligand-bound CD36 (47) (Figure 1A; Table 1). In addition, CD9 functionally associates with FcγRs, modulating signals for phagocytosis, and FcγR-mediated immune responses (Table 1). Cross-linking of CD9-FcγRIII induces colocalization of CD9, αMβ2 integrin and F-actin, promoting macrophage activation (48) (Figure 1A). In human monocytes and skin-derived DCs, CD9 and CD81 are molecular partners of the trimeric form of FcϵRI (Figure 1; Table 1), the high-affinity receptor for IgE, and are overexpressed in patients with atopic dermatitis (49).
The tetraspanin CD81 plays an important role in Ag-induced B cell activation, B cell development, and survival. It associates functionally with CD19 and CD21, which are members of the BCR complex (3, 54) (Table 1). CD81 deficiency in humans and mice leads to antibody deficiency syndrome by preventing CD19 surface expression (21, 63). Moreover, visualization of primary B cells by super-resolution microscopy shows that CD81-enriched microdomains and the actin cytoskeleton regulate CD19 mobility and organize CD19 and BCR interactions, controlling BCR downstream signaling (64).
In the context of viral infection, CD81 was identified as a receptor for hepatitis C virus (HCV) (65), not only in hepatocytes but also in B cells, T cells, NK cells, and DCs (66). The dynamic properties of CD81 at the membrane are essential for HCV infection (67). Anti-CD81-specific antibodies mediate protection against HCV infection in vivo, further demonstrating the functional consequences of this recognition (68). Tetraspanin dynamics at the membrane are also exploited by other viruses. For example, CD9 and CD81 negatively regulate human immunodeficiency virus 1 (HIV-1)-induced membrane fusion (69).
TEMs during Antigen Processing and Presentation
T cell recognition of specific antigenic peptides bound to MHC-I and MHC-II molecules on DCs leads to T cell activation and subsequent initiation of T cell-mediated immune responses. In DCs, mechanisms regulating MHC-II intracellular transport are well known (70), and tetraspanins have a role in this process since several tetraspanin family members associate with MHC-II molecules. Interactions between MHC-I molecules and tetraspanins CD53, CD81, and CD82 have been described (71) (Figure 1C). Moreover, tetraspanins CD9, CD81, CD82, CD63, CD53, and CD37 interact with MCH-II molecules (50, 55, 57, 72) (Figures 1C and 2; Table 1). These interactions might lead to the regulation of MHC-II subcellular distribution. CD9, CD53, and CD81 associate with MHC-II at the plasma membrane (50) (Figure 1C; Table 1). In contrast, CD82 and CD63 are highly enriched in MHC-II-enriched compartments (MIIC) (Figure 1B; Table 1), particularly in intraluminal vesicles, where they associate with each other and with the chaperone HLA-DM, playing an important role in the late stages of MHC-II maturation (50, 57) (Table 1). Analysis of protein dynamics by Föster resonance energy transfer (FRET) in MIIC shows that CD63 stably associates with MHC-II and regulates MHC-II surface expression, whereas CD82 associates with HLA-DM without affecting MHC-II expression (58). Knockdown of CD63, CD82, CD9, or CD81 did not prevent MHC-II peptide loading (58). In addition, live cell imaging studies have shown differential CD63 and CD82 subcellular localization in the context of DC phagocytosis. Whereas CD63 and MHC-II are specifically recruited to yeast-containing phagosomes after phagosomal acidification (43), CD82 and MHC-II molecules are recruited to fungal and bacterial phagosomes before fusion with lysosomes and phagosomal acidification (44) (Figure 1B; Table 1). These results support a role for CD63 and CD82 in the dynamic intracellular trafficking of MHC-II after pathogen uptake, playing non-redundant roles in these processes.
Tetraspanins are also involved in the clustering of MHC molecules (Figure 1C). APCs express very small amounts of relevant MHC-II–peptide complexes on the plasma membrane. These MHC-II–peptide complexes are organized and clustered on the cell surface, allowing efficient cross-linking of TCRs and promoting Ag-specific T cell activation (73). It is widely accepted that MHC-II molecules are concentrated into two types of membrane microdomains, TEMs, and lipid rafts (74). The composition and dynamics of these microdomains are essential factors in the outcome of T cell activation. Evidence from a model of raft disruption in B cells suggests that MHC-II association with lipid rafts is important for presentation of Ag at low concentrations (75). Other studies report that TEMs contain MHC-II molecules loaded with a restricted antigenic peptide repertoire, together with HLA-DM and the costimulatory molecule CD86. In contrast, raft-associated MHC-II molecules display a highly diverse set of peptides (51) (Table 1). However, these results are controversial, since the MHC-II determinant CDw78, which is used to identify selectively tetraspanin-associated MHC-II, also defines a conformation of peptide-bound MHC-II acquired through the trafficking to lysosomal compartments (76). Moreover, TEM-induced MHC-II clustering is also supported by evidence that CD9 is required to facilitate the formation of I-A/I-E MHC-II multimers, which are responsible for enhancing the T cell stimulatory capacity of DCs (52) (Table 1). However, a subsequent study showed that cholesterol depletion disrupts MHC-II I-A/I-E interactions, whereas the absence of CD9 or CD81 has no effect (77). This controversy might be due to the differential sensitivity of microdomains to cholesterol depletion. Although TEMs are more resistant to cholesterol depletion than lipid rafts, partial disruption is also observed under certain conditions. Therefore, it is possible that rafts and TEMs both contribute to MHC clustering.
Studies derived from tetraspanin-deficient mice have shown that certain tetraspanin members do not promote MHC multimerization, being rather involved in Ag presentation. DCs from CD37−/− or CD151−/− mice induce hyperstimulation of T cells (56), and similar results were obtained with DCs from Tssc6−/− mice and CD37−/− Tssc6−/− double knockout mice (78). CD37−/− DCs induce T cell hyperstimulation through a mechanism that regulates MHC-dependent Ag presentation, whereas CD151 in DCs regulates T cell costimulation (56) (Figures 1C and 2; Table 2). DC maturation is required for effective T-cell costimulation and involves the upregulation of costimulatory and adhesion molecules (79, 80). In contrast to conventional DCs, plasmacytoid DCs lack CD9 surface expression, which could be responsible for their significant low expression of MHC-II and limited T cell stimulatory potential (80). TEMs thus play a well-documented role in the regulation of different aspects of the MHC-II lifecycle in APCs, including MHC-II clustering and intracellular trafficking of peptide–MHC-II complexes to the APC plasma membrane.
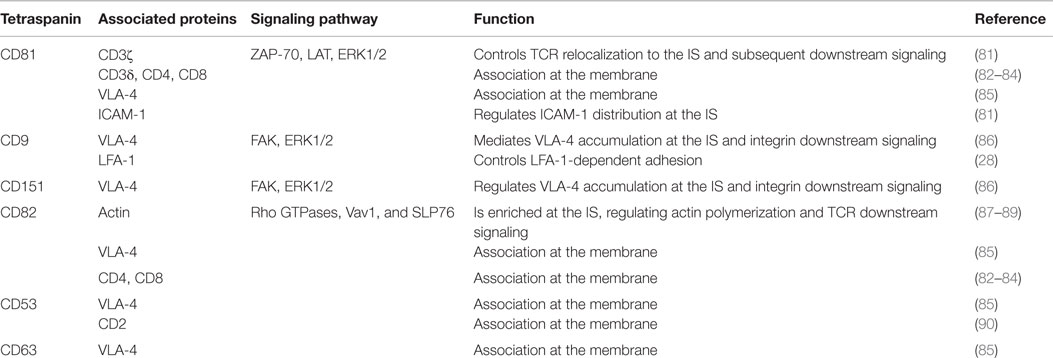
Table 2. Tetraspanin associations in T cells and their role at the immunological synapse.
Role of Tetraspanins in the Organization of T-Cell Immunological Synapses
The Immunological Synapse
The initiation of T cell activation mediated by APCs, mainly DCs, requires the establishment of a dynamic structure formed at the cell–cell contact called the immunological synapse (IS) (Figure 2). This structure is characterized by a dynamic spatiotemporal recruitment of Ag receptors, costimulatory molecules, and adhesion proteins to specific zones at the T cell–APC interface. At the T cell side of mature IS, TCR microclusters are clustered together with costimulatory proteins, signaling molecules, and other signaling adaptors at the central supramolecular activation complex (cSMAC) (91–96). More specifically, preexisting TCR nanoclusters (97) concatenate into microclusters, as demonstrated with high-resolution imaging techniques like photoactivated localization microscopy (PALM) and stimulated emission depletion (STED) (98–100). These microclusters form in the periphery of the IS and are translocated toward the cSMAC in a process dependent on the actin cytoskeleton (93, 94, 98, 101, 102). The central area is surrounded by a peripheral SMAC (pSMAC), where integrins and adhesion receptors are localized (81, 91, 102–104). The super-resolution optical techniques near-field scanning optical microscopy (NSOM) and single-dye tracking (SDT) revealed that, like the TCR, LFA-1 is preorganized into nanoclusters that coalesce into microclusters after ligand binding (105, 106). The stability of the IS depends on the binding of integrins, not only lymphocyte function-associated antigen 1 (LFA-1; αLβ2) but also very late antigen 4 (VLA-4; α4β1), to their ligands, the adhesion receptors intercellular adhesion molecule-1, -3 (ICAM) in the case of LFA-1 (91, 103, 104, 107–109). The VLA-4 ligand at the T-cell–APC interface remains unknown (104). In resting T lymphocytes, integrins are mostly in an inactive bent conformation, with low affinity and avidity for ligands. TCR stimulation triggers intracellular signaling that leads integrins to adopt an intermediate-affinity conformation, and then the extended high-affinity conformation (110). These conformational changes induced by TCR signaling modify integrin avidity through a process called inside-out signaling (111), which ultimately regulates integrin affinity for their ligands (112, 113). LFA-1 engagement by its ligand ICAM-1 triggers outside-in signaling, inducing cytoskeletal reorganization that recruits T cell signaling proteins to the IS (113–115).
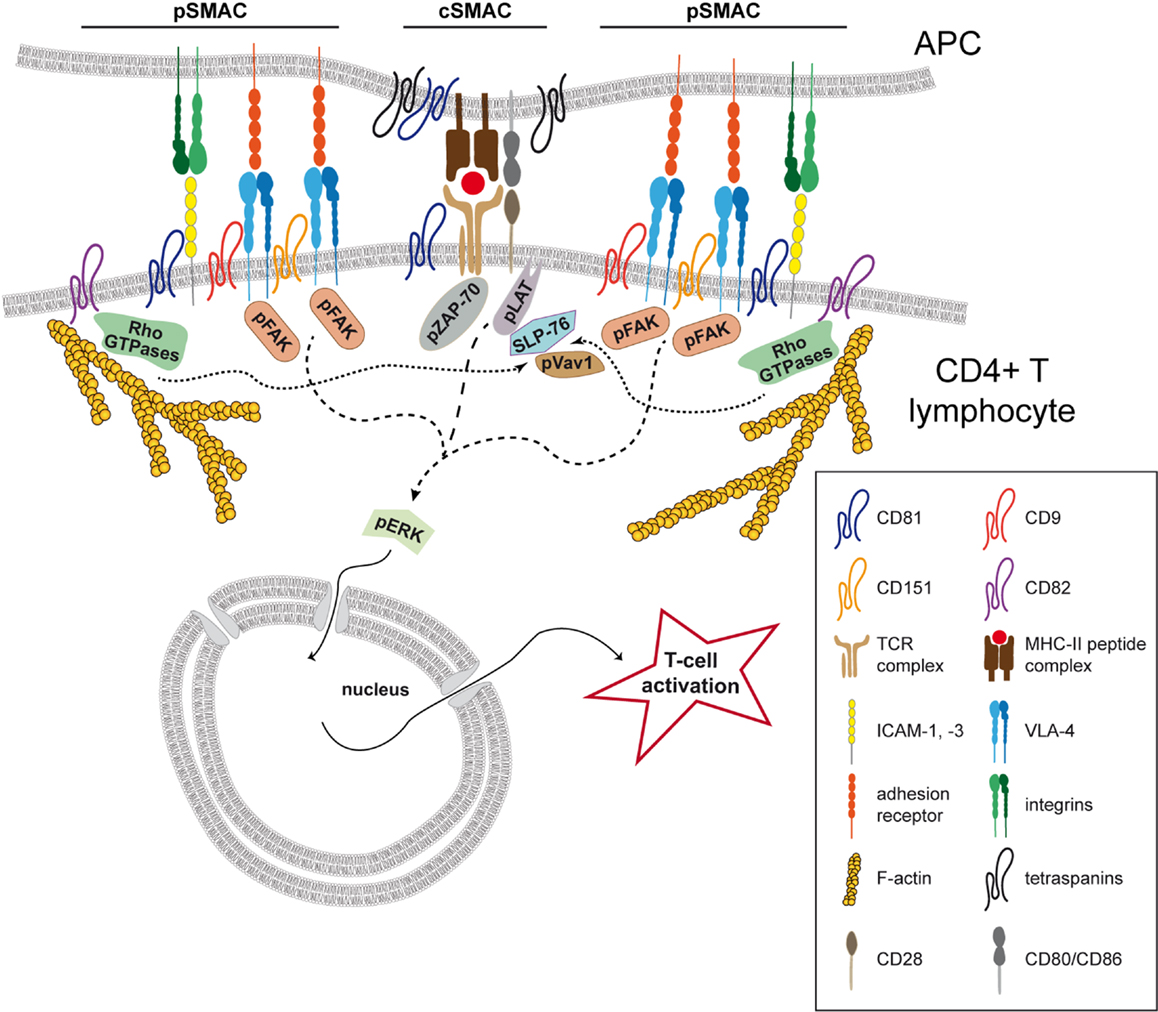
Figure 2. Tetraspanins organize the T-cell immunological synapse. Tetraspanin CD81 regulates the organization of the immunological synapse (IS) in CD4+ T lymphocytes through the association with CD3ζ at the central SMAC (cSMAC). CD81 controls the localization of the TCR complex and its downstream signaling, positively modulating the phosphorylation of ZAP-70, LAT, and ERK1/2 (dashed line). At the peripheral area of the cell–cell contact (pSMAC), tetraspanins CD9, and CD151 are important for integrin VLA-4 relocalization and activation, positively regulating the integrin downstream phosphorylation of FAK and ERK1/2 (lines with small dashes). At this location, CD81 also interacts with the adhesion receptor ICAM-1, regulating its segregation during IS maturation. Tetraspanin CD82 accumulates at the pSMAC and triggers actin polymerization and the activation of the Rho GTPase pathway (RhoA, Rac1, and Cdc42). The activation of this pathway induces the phosphorylation and the association of Vav1 and SLP76 (dotted lines), potentiating the phosphorylation of the TCR signaling molecules LAT and ZAP-70. In APCs, CD81 is enriched at the IS and several tetraspanins are described to associate with MHC-II. Moreover, CD151, CD37, and Tssc6 were described to regulate antigen presentation by DCs.
Both the TCR and LFA-1 modulate cytoskeletal dynamics. TCR signaling triggers actin polymerization enabling the extension of the actin network downstream of LFA-1 (116). VLA-4 costimulation regulates the cytoskeletal movements that drive TCR microclusters associated with signaling complexes to the central area of the IS (109). Moreover, it has been suggested that TCR microcluster formation is dependent on actin polymerization (94, 101); however, other investigators claim that actin is necessary only for microcluster maintenance (98). Continuous actin retrograde flow sustains T cell signaling and signal termination at the central area of the IS (94, 101, 102, 117). Actin filaments are also important for the segregation of adhesion molecules to the pSMAC (91, 102, 103), and actin centripetal flow is essential for the maintenance of LFA-1 in a high-affinity conformation at this location (118, 119).
The T cell cytoskeletal network thus plays an essential role in the spatial organization of the IS. However, the precise mechanisms by which molecules are specifically partitioned into central and peripheral areas of the IS remain an open question. It has been proposed that this segregation is supported by size differences in the ectodomains of immune surface interacting proteins; e.g., LFA-1-ICAM-1 (40 nm) and CD2-CD58 (15 nm) (120–122). In agreement with this view, evidence suggests size-dependent exclusion from the cSMAC of large phosphatases such as CD45, thus allowing the initiation of TCR signaling (94, 123). Recent data show that CD45 is already excluded from preexisting TCR microclusters (124). Given that the TCR in naïve T cells is already clustered with signaling molecules, and that numerous proteins that are translocated to, rearranged and accumulated at the IS are known to associate with tetraspanins, we postulate that protein–protein interactions driven by TEMs actively contribute to IS architectural organization.
Tetraspanins and the Distribution of Receptors at the T-cell IS
Tetraspanin CD81 accumulates at the IS in both T lymphocytes and APCs (125) (Figure 2), and we recently found that CD81 is an important molecular organizer of the IS structure at the T cell side (81). Fluorescence recovery after photobleaching (FRAP) experiments indicate that CD81 is mostly confined to the cSMAC in the early IS (81), where it colocalizes with the CD3ζ component of the TCR complex (81, 125) (Figure 2). Analyses by phasor fluorescence-lifetime imaging microscopy (phasorFLIM)-FRET reveal that CD81 associates with CD3ζ at the cSMAC of the early IS (81) (Figure 2). In the late IS, CD81 and CD3ζ spread throughout the cell–cell contact and CD81 diffusion decreases, suggesting stable protein–protein interactions throughout the IS. In agreement with this view, CD81 and CD3ζ interaction increases with the IS maturation (81). As a molecular organizer, CD81 controls CD3ζ relocalization to the cSMAC, and the efficient maintenance of the CD3 signaling complex at the cell–cell contact (Figure 2). Hence, CD81 knockdown reduces the number of CD3ζ microclusters at the cSMAC, as detected by total internal reflection microscopy (TIRFM), and impairs TCR downstream signaling, reducing the phosphorylation of CD3ζ, ZAP-70, LAT, and ERK1/2 (81) (Figure 2; Table 2). Moreover, pretreatment of T cells with soluble GST-LEL-CD81 (81), which decreases membrane diffusion of the protein (33), increases T cell activation (81), further indicating that CD81 regulates T cell activation by controlling the duration of TCR signaling at the membrane. A direct CD81-mediated signaling does not seem to be involved in this process, since CD81 C-terminal peptides do not affect T cell activation (81). Thus, by organizing TEMs CD81 regulates spatial molecular organization during the maturation of the IS.
In T lymphocytes, different tetraspanins associate with receptors that are enriched at the IS. In addition to CD3ζ (81), CD81 also interacts with the CD3δ subunit of the TCR complex (84) (Table 2). CD9 localizes with TCR signaling molecules in lipid microdomains (10), CD81 and CD82 associate with CD4 and CD8 coreceptors (82, 83) (Table 2), and CD53 interacts with the costimulatory receptor CD2 (90) (Table 2). It is therefore conceivable that the IS architectural organization of these receptors depends on their inclusion in TEMs through interaction with different tetraspanins. Further research is required to address this notion.
Adhesion Molecules, Tetraspanins, and the Stabilization of the T-cell IS
Integrins and adhesion receptors are also included in TEMs. In T cells, CD9 interact with LFA-1 (28), CD81, CD82, and CD53 with VLA-4 (85), and CD81 with ICAM-1 (81) (Table 2). In the immune system, tetraspanins regulate cell–cell adhesion through LFA-1 and ICAM-1: CD81 and CD82 promote T-APC cell–cell interaction (126, 127); CD81 induces thymocyte aggregation (128); and CD53 modulates NK and B cell aggregation (129, 130). Conversely, leukocyte LFA-1-dependent adhesion is negatively regulated by CD9 (28) (Table 2). Integrin adhesiveness can be regulated by several mechanisms, such as alterations in the affinity of individual integrin molecules or changes in their clustering on the cell surface or their interactions with ligands. Tetraspanins can modulate integrin activity through various mechanisms. For example, CD81 modulates VLA-4 avidity for its ligand VCAM-1, and CD151 stabilizes α3β1 integrin in its active conformation and regulates α6 integrin diffusion at the plasma membrane (27, 131, 132). CD9 promotes β1 activation, LFA-1 aggregation, and in leukocytes it seems to be essential for a balanced regulation of β1 and β2 integrin activity: it increases β1 adhesion to fibronectin but diminishes LFA-1-mediated adhesion (28, 133).
At the IS, CD81 regulates pSMAC organization through association with the adhesion receptor ICAM-1, controlling ICAM-1 segregation at the cell–cell contact during IS maturation (81) (Figure 2; Table 2). CD81 knockdown decreases the proportion of early synapses, in which ICAM-1 is confined to the pSMAC, and increases the proportion of late synapses (81). During maturation of the IS, ICAM-1 redistributes throughout the entire cell–cell contact, with increasing colocalization and molecular interaction with CD81 (81). T cell activation is also regulated by other tetraspanins. CD9 and CD151 modulate VLA-4 accumulation at the IS (86) (Figure 2; Table 2). Interestingly, the IS enrichment of β1 integrins in a high-affinity conformation is impaired in T cells knocked-down for CD9 and CD151, suggesting that integrin activation upon IS formation occurs within TEMs (86). The conformational changes of β integrin extracellular domains can be controlled by the actin linker protein talin (134), which accumulates at the pSMAC (91) and is required for LFA-1 activation mediated by the TCR (135). However, CD9 and CD151 knockdown does not alter talin relocalization to the IS, indicating that these tetraspanins are not involved in the regulation of integrin inside-out signaling (86). Integrins and adhesion molecules can act as signaling receptors. Integrin or ICAM-1 costimulation triggers T cell activation (136–138), and LFA-1 coengagement with the TCR lowers the T cell activation threshold (139, 140). VLA-4 ligation also costimulates T cells in a TCR-dependent manner (141), and polarizes T lymphocytes toward Th1 responses (104). LFA-1 and VLA-4 activation is controlled by the interaction with a cascade of adaptor and signaling proteins (142, 143), and these downstream signaling can be modulated by tetraspanins. CD151 supports the phosphorylation of FAK, Src, and p130CAS (144) and promotes the activation of small GTPases and ERK1/2 in an integrin-dependent manner (145, 146). ERK1/2 signaling is also increased by CD9 (147). During T-APC cognate cell–cell interactions, CD9 and CD151 knockdown reduces FAK and ERK1/2 phosphorylation, and impairs the enrichment of phosphorylated FAK at the IS (86) (Figure 2; Table 2). Tetraspanins CD9 and CD151 are therefore important for integrin enrichment at the IS, modulating integrin downstream signaling.
As previously mentioned, the actin cytoskeleton plays a crucial role in the regulation of the spatial organization of TCRs and adhesion molecules at the IS. The links between tetraspanins, membrane receptors, adhesion proteins, and the actin cytoskeleton suggest a possible regulation of this process by TEMs. CD81 and CD9 are connected to the actin cytoskeleton through α-actinin and ezrin-radixin-moesin (ERM) proteins (148, 149). CD151, CD81, and CD82 regulate the actin cytoskeleton through RhoA and Rac1 signaling molecules (62, 150–152). In T lymphocytes, CD82 costimulation triggers actin polymerization and T-cell activation by stabilizing signaling downstream of TCR/CD3 (87, 88) (Figure 2; Table 2). T cell morphological changes induced by CD82 engagement depend on the activity of Rho GTPases (RhoA, Rac1, and Cdc42), involving the association of Vav1 and the adapter molecule SLP76 with the Rho GTPase pathway (88). Importantly, CD82 is enriched at the IS in an actin-dependent manner (89) (Figure 2; Table 2). CD82-dependent regulation of the actin cytoskeleton during T cell activation may involve its interaction with LFA-1. CD82 regulates T cell-APC adhesion-dependent signaling (153), through its interaction with LFA-1 (126), and like LFA-1, CD82 localizes at the pSMAC (89) (Figure 2). At the IS, CD82 seems to stabilize interactions with the actin cytoskeleton, favoring the formation of signaling complexes. It would be interesting to determine whether CD82 dynamics depend on its association with LFA-1, and whether CD82 can modulate LFA-1 functions.
Thus, at the IS, tetraspanins CD9, CD81, CD82, and CD151 mediate the connections between adhesion molecules, the actin cytoskeleton and signaling complexes. Increasing evidence highlights the importance of TEMs in the organization of the temporal and spatial molecular distribution at the IS, generating the context that allows full T cell activation.
Concluding Remarks
In APCs, different receptors involved in pathogen recognition and Ag presentation are associated with tetraspanins. Further investigations are necessary to determine the spatial distribution and segregation of receptors within TEMs, as well as the importance of these microdomains in the regulatory mechanisms of receptor functions and downstream signaling. The establishment of long-lasting T cell–APC contacts, which lead to the formation of the IS and ultimately promote an efficient T cell activation, are required for the initiation of T cell-mediated immune responses. IS stability depends on the binding of integrins to adhesion receptors upon TCR ligation, triggering downstream signaling. The complex IS architectural organization depends on the inclusion of the receptors concentrated at the IS into TEMs, through their dynamic and spatiotemporal interactions with different tetraspanins. The important role of TEMs in the regulation of the dynamic process of IS formation has been recently emphasized. These specialized membrane domains allow the compartmentalization of receptors and adhesion molecules and connect them to the cytoskeleton and signaling complexes that induce T cell activation. The development of advanced microscopy techniques will provide further insight into IS dynamics and the contribution of TEMs and other microdomains to this process. Considering the plasticity of the interactions that take place in TEMs, strategies that regulate IS organization by targeting tetraspanins could allow therapeutic manipulation of the final outcome of T cell activation and the subsequent immune response.
Author Contributions
VR-P, FS-M, and GMH had scientific discussion for this work and wrote the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank S. Bartlett (CNIC) for manuscript editing. This study was supported by grants to FS-M from the Spanish Ministry of Economy and Competitiveness (SAF2011-25834 and SAF2014-55579-R), the Comunidad de Madrid (INDISNET-S2011/BMD-2332), the Instituto de Salud Carlos III (Cardiovascular Network RD12-0042-0056), and the European Commission (ERC-2011-AdG 294340-GENTRIS); and by grant PI11/00939 from the Instituto de Salud Carlos III to GMH. GMH is supported by the Ramon y Cajal program (RYC-2010-05920) from the Spanish Ministry of Economy and Competitiveness. The Centro Nacional de Investigaciones Cardiovasculares (CNIC) is supported by the Spanish Ministry of Economy and Competitiveness and the Pro-CNIC Foundation.
Funding
The cost of this publication has been paid in part by FEDER funds.
References
1. Tarrant JM, Robb L, Van Spriel AB, Wright MD. Tetraspanins: molecular organisers of the leukocyte surface. Trends Immunol (2003) 24:610–7. doi: 10.1016/j.it.2003.09.011
2. Hemler ME. Tetraspanin functions and associated microdomains. Nat Rev Mol Cell Biol (2005) 6:801–11. doi:10.1038/nrm1736
3. Levy S, Shoham T. The tetraspanin web modulates immune-signalling complexes. Nat Rev Immunol (2005) 5:136–48. doi:10.1038/nri1548
4. Berditchevski F, Odintsova E, Sawada S, Gilbert E. Expression of the palmitoylation-deficient CD151 weakens the association of alpha 3 beta 1 integrin with the tetraspanin-enriched microdomains and affects integrin-dependent signaling. J Biol Chem (2002) 277:36991–7000. doi:10.1074/jbc.M205265200
5. Yanez-Mo M, Barreiro O, Gordon-Alonso M, Sala-Valdes M, Sanchez-Madrid F. Tetraspanin-enriched microdomains: a functional unit in cell plasma membranes. Trends Cell Biol (2009) 19:434–46. doi:10.1016/j.tcb.2009.06.004
6. Boucheix C, Rubinstein E. Tetraspanins. Cell Mol Life Sci (2001) 58:1189–205. doi:10.1007/PL00000933
7. Zuidscherwoude M, Gottfert F, Dunlock VM, Figdor CG, Van Den Bogaart G, Spriel AB. The tetraspanin web revisited by super-resolution microscopy. Sci Rep (2015) 5:12201. doi:10.1038/srep12201
8. Lagaudriere-Gesbert C, Le Naour F, Lebel-Binay S, Billard M, Lemichez E, Boquet P, et al. Functional analysis of four tetraspans, CD9, CD53, CD81, and CD82, suggests a common role in costimulation, cell adhesion, and migration: only CD9 upregulates HB-EGF activity. Cell Immunol (1997) 182:105–12. doi:10.1006/cimm.1997.1223
9. Tai XG, Toyooka K, Yashiro Y, Abe R, Park CS, Hamaoka T, et al. CD9-mediated costimulation of TCR-triggered naive T cells leads to activation followed by apoptosis. J Immunol (1997) 159:3799–807.
10. Yashiro-Ohtani Y, Zhou XY, Toyo-Oka K, Tai XG, Park CS, Hamaoka T, et al. Non-CD28 costimulatory molecules present in T cell rafts induce T cell costimulation by enhancing the association of TCR with rafts. J Immunol (2000) 164:1251–9. doi:10.4049/jimmunol.164.3.1251
11. Tseng CT, Miskovsky E, Klimpel GR. Crosslinking CD81 results in activation of TCRgammadelta T cells. Cell Immunol (2001) 207:19–27. doi:10.1006/cimm.2000.1744
12. Pfistershammer K, Majdic O, Stockl J, Zlabinger G, Kirchberger S, Steinberger P, et al. CD63 as an activation-linked T cell costimulatory element. J Immunol (2004) 173:6000–8. doi:10.4049/jimmunol.173.10.6000
13. Sagi Y, Landrigan A, Levy R, Levy S. Complementary costimulation of human T-cell subpopulations by cluster of differentiation 28 (CD28) and CD81. Proc Natl Acad Sci U S A (2012) 109:1613–8. doi:10.1073/pnas.1121307109
14. Lebel-Binay S, Lagaudriere C, Fradelizi D, Conjeaud H. CD82, member of the tetra-span-transmembrane protein family, is a costimulatory protein for T cell activation. J Immunol (1995) 155:101–10.
15. Tai XG, Yashiro Y, Abe R, Toyooka K, Wood CR, Morris J, et al. A role for CD9 molecules in T cell activation. J Exp Med (1996) 184:753–8. doi:10.1084/jem.184.2.753
16. Witherden DA, Boismenu R, Havran WL. CD81 and CD28 costimulate T cells through distinct pathways. J Immunol (2000) 165:1902–9. doi:10.4049/jimmunol.165.4.1902
17. Miyazaki T, Muller U, Campbell KS. Normal development but differentially altered proliferative responses of lymphocytes in mice lacking CD81. EMBO J (1997) 16:4217–25. doi:10.1093/emboj/16.14.4217
18. Tarrant JM, Groom J, Metcalf D, Li R, Borobokas B, Wright MD, et al. The absence of Tssc6, a member of the tetraspanin superfamily, does not affect lymphoid development but enhances in vitro T-cell proliferative responses. Mol Cell Biol (2002) 22:5006–18. doi:10.1128/MCB.22.14.5006-5018.2002
19. Van Spriel AB, Puls KL, Sofi M, Pouniotis D, Hochrein H, Orinska Z, et al. A regulatory role for CD37 in T cell proliferation. J Immunol (2004) 172:2953–61. doi:10.4049/jimmunol.172.5.2953
20. Wright MD, Geary SM, Fitter S, Moseley GW, Lau LM, Sheng KC, et al. Characterization of mice lacking the tetraspanin superfamily member CD151. Mol Cell Biol (2004) 24:5978–88. doi:10.1128/MCB.24.13.5978-5988.2004
21. Maecker HT, Levy S. Normal lymphocyte development but delayed humoral immune response in CD81-null mice. J Exp Med (1997) 185:1505–10. doi:10.1084/jem.185.8.1505
22. Tsitsikov EN, Gutierrez-Ramos JC, Geha RS. Impaired CD19 expression and signaling, enhanced antibody response to type II T independent antigen and reduction of B-1 cells in CD81-deficient mice. Proc Natl Acad Sci U S A (1997) 94:10844–9. doi:10.1073/pnas.94.20.10844
23. Knobeloch KP, Wright MD, Ochsenbein AF, Liesenfeld O, Lohler J, Zinkernagel RM, et al. Targeted inactivation of the tetraspanin CD37 impairs T-cell-dependent B-cell response under suboptimal costimulatory conditions. Mol Cell Biol (2000) 20:5363–9. doi:10.1128/MCB.20.15.5363-5369.2000
24. Maecker HT, Do MS, Levy S. CD81 on B cells promotes interleukin 4 secretion and antibody production during T helper type 2 immune responses. Proc Natl Acad Sci U S A (1998) 95:2458–62. doi:10.1073/pnas.95.5.2458
25. Deng J, Dekruyff RH, Freeman GJ, Umetsu DT, Levy S. Critical role of CD81 in cognate T-B cell interactions leading to Th2 responses. Int Immunol (2002) 14:513–23. doi:10.1093/intimm/14.5.513
26. Maecker HT. Human CD81 directly enhances Th1 and Th2 cell activation, but preferentially induces proliferation of Th2 cells upon long-term stimulation. BMC Immunol (2003) 4:1. doi:10.1186/1471-2172-4-1
27. Feigelson SW, Grabovsky V, Shamri R, Levy S, Alon R. The CD81 tetraspanin facilitates instantaneous leukocyte VLA-4 adhesion strengthening to vascular cell adhesion molecule 1 (VCAM-1) under shear flow. J Biol Chem (2003) 278:51203–12. doi:10.1074/jbc.M303601200
28. Reyes R, Monjas A, Yanez-Mo M, Cardenes B, Morlino G, Gilsanz A, et al. Different states of integrin LFA-1 aggregation are controlled through its association with tetraspanin CD9. Biochim Biophys Acta (2015) 1853(10 Pt A):2464–80. doi:10.1016/j.bbamcr.2015.05.018
29. Dijkstra S, Kooij G, Verbeek R, Van Der Pol SM, Amor S, Geisert EE Jr, et al. Targeting the tetraspanin CD81 blocks monocyte transmigration and ameliorates EAE. Neurobiol Dis (2008) 31:413–21. doi:10.1016/j.nbd.2008.05.018
30. Fernvik E, Gronneberg R, Lundahl J, Raud J, Zetterstrom O, Van Hage-Hamsten M, et al. Characterization of eosinophils and detection of eotaxin in skin chamber fluid after challenge with relevant allergen in patients with mild asthma. Clin Exp Allergy (1999) 29:1516–25. doi:10.1046/j.1365-2222.1999.00572.x
31. Fernvik E, Lundahl J, Hallden G. The impact of eotaxin- and IL-5-induced adhesion and transmigration on eosinophil activity markers. Inflammation (2000) 24:73–87. doi:10.1023/A:1006940109869
32. Barreiro O, Yanez-Mo M, Sala-Valdes M, Gutierrez-Lopez MD, Ovalle S, Higginbottom A, et al. Endothelial tetraspanin microdomains regulate leukocyte firm adhesion during extravasation. Blood (2005) 105:2852–61. doi:10.1182/blood-2004-09-3606
33. Barreiro O, Zamai M, Yanez-Mo M, Tejera E, Lopez-Romero P, Monk PN, et al. Endothelial adhesion receptors are recruited to adherent leukocytes by inclusion in preformed tetraspanin nanoplatforms. J Cell Biol (2008) 183:527–42. doi:10.1083/jcb.200805076
34. Gutierrez-Vazquez C, Villarroya-Beltri C, Mittelbrunn M, Sanchez-Madrid F. Transfer of extracellular vesicles during immune cell-cell interactions. Immunol Rev (2013) 251:125–42. doi:10.1111/imr.12013
35. Andreu Z, Yanez-Mo M. Tetraspanins in extracellular vesicle formation and function. Front Immunol (2014) 5:442. doi:10.3389/fimmu.2014.00442
36. Perez-Hernandez D, Gutierrez-Vazquez C, Jorge I, Lopez-Martin S, Ursa A, Sanchez-Madrid F, et al. The intracellular interactome of tetraspanin-enriched microdomains reveals their function as sorting machineries toward exosomes. J Biol Chem (2013) 288:11649–61. doi:10.1074/jbc.M112.445304
37. Mogensen TH. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin Microbiol Rev (2009) 22:240–73. doi:10.1128/CMR.00046-08
38. Zuidscherwoude M, De Winde CM, Cambi A, Van Spriel AB. Microdomains in the membrane landscape shape antigen-presenting cell function. J Leukoc Biol (2014) 95:251–63. doi:10.1189/jlb.0813440
39. Goodridge HS, Reyes CN, Becker CA, Katsumoto TR, Ma J, Wolf AJ, et al. Activation of the innate immune receptor dectin-1 upon formation of a ‘phagocytic synapse’. Nature (2011) 472:471–5. doi:10.1038/nature10071
40. Mantegazza AR, Barrio MM, Moutel S, Bover L, Weck M, Brossart P, et al. CD63 tetraspanin slows down cell migration and translocates to the endosomal-lysosomal-MIICs route after extracellular stimuli in human immature dendritic cells. Blood (2004) 104:1183–90. doi:10.1182/blood-2004-01-0104
41. Meyer-Wentrup F, Figdor CG, Ansems M, Brossart P, Wright MD, Adema GJ, et al. Dectin-1 interaction with tetraspanin CD37 inhibits IL-6 production. J Immunol (2007) 178:154–62. doi:10.4049/jimmunol.178.1.154
42. Van Spriel AB, Sofi M, Gartlan KH, Van Der Schaaf A, Verschueren I, Torensma R, et al. The tetraspanin protein CD37 regulates IgA responses and anti-fungal immunity. PLoS Pathog (2009) 5:e1000338. doi:10.1371/journal.ppat.1000338
43. Artavanis-Tsakonas K, Love JC, Ploegh HL, Vyas JM. Recruitment of CD63 to Cryptococcus neoformans phagosomes requires acidification. Proc Natl Acad Sci U S A (2006) 103:15945–50. doi:10.1073/pnas.0607528103
44. Artavanis-Tsakonas K, Kasperkovitz PV, Papa E, Cardenas ML, Khan NS, Van Der Veen AG, et al. The tetraspanin CD82 is specifically recruited to fungal and bacterial phagosomes prior to acidification. Infect Immun (2011) 79:1098–106. doi:10.1128/IAI.01135-10
45. Suzuki M, Tachibana I, Takeda Y, He P, Minami S, Iwasaki T, et al. Tetraspanin CD9 negatively regulates lipopolysaccharide-induced macrophage activation and lung inflammation. J Immunol (2009) 182:6485–93. doi:10.4049/jimmunol.0802797
46. Huang W, Febbraio M, Silverstein RL. CD9 tetraspanin interacts with CD36 on the surface of macrophages: a possible regulatory influence on uptake of oxidized low density lipoprotein. PLoS One (2011) 6:e29092. doi:10.1371/journal.pone.0029092
47. Heit B, Kim H, Cosio G, Castano D, Collins R, Lowell CA, et al. Multimolecular signaling complexes enable Syk-mediated signaling of CD36 internalization. Dev Cell (2013) 24:372–83. doi:10.1016/j.devcel.2013.01.007
48. Kaji K, Takeshita S, Miyake K, Takai T, Kudo A. Functional association of CD9 with the Fc gamma receptors in macrophages. J Immunol (2001) 166:3256–65. doi:10.4049/jimmunol.166.5.3256
49. Peng WM, Yu CF, Kolanus W, Mazzocca A, Bieber T, Kraft S, et al. Tetraspanins CD9 and CD81 are molecular partners of trimeric FcvarepsilonRI on human antigen-presenting cells. Allergy (2011) 66:605–11. doi:10.1111/j.1398-9995.2010.02524.x
50. Engering A, Pieters J. Association of distinct tetraspanins with MHC class II molecules at different subcellular locations in human immature dendritic cells. Int Immunol (2001) 13:127–34. doi:10.1093/intimm/13.2.127
51. Kropshofer H, Spindeldreher S, Rohn TA, Platania N, Grygar C, Daniel N, et al. Tetraspan microdomains distinct from lipid rafts enrich select peptide-MHC class II complexes. Nat Immunol (2002) 3:61–8. doi:10.1038/ni750
52. Unternaehrer JJ, Chow A, Pypaert M, Inaba K, Mellman I. The tetraspanin CD9 mediates lateral association of MHC class II molecules on the dendritic cell surface. Proc Natl Acad Sci U S A (2007) 104:234–9. doi:10.1073/pnas.0609665104
53. Martinez Del Hoyo G, Ramirez-Huesca M, Levy S, Boucheix C, Rubinstein E, Minguito De La Escalera M, et al. CD81 controls immunity to Listeria infection through Rac-dependent inhibition of proinflammatory mediator release and activation of cytotoxic T cells. J Immunol (2015) 194:6090–101. doi:10.4049/jimmunol.1402957
54. Levy S. Function of the tetraspanin molecule CD81 in B and T cells. Immunol Res (2014) 58:179–85. doi:10.1007/s12026-014-8490-7
55. Angelisova P, Hilgert I, Horejsi V. Association of four antigens of the tetraspans family (CD37, CD53, TAPA-1, and R2/C33) with MHC class II glycoproteins. Immunogenetics (1994) 39:249–56. doi:10.1007/BF00188787
56. Sheng KC, Van Spriel AB, Gartlan KH, Sofi M, Apostolopoulos V, Ashman L, et al. Tetraspanins CD37 and CD151 differentially regulate Ag presentation and T-cell co-stimulation by DC. Eur J Immunol (2009) 39:50–5. doi:10.1002/eji.200838798
57. Hammond C, Denzin LK, Pan M, Griffith JM, Geuze HJ, Cresswell P. The tetraspan protein CD82 is a resident of MHC class II compartments where it associates with HLA-DR, -DM, and -DO molecules. J Immunol (1998) 161:3282–91.
58. Hoorn T, Paul P, Janssen L, Janssen H, Neefjes J. Dynamics within tetraspanin pairs affect MHC class II expression. J Cell Sci (2012) 125:328–39. doi:10.1242/jcs.088047
59. Pfeiffer A, Bottcher A, Orso E, Kapinsky M, Nagy P, Bodnar A, et al. Lipopolysaccharide and ceramide docking to CD14 provokes ligand-specific receptor clustering in rafts. Eur J Immunol (2001) 31:3153–64. doi:10.1002/1521-4141(200111)31:11<3153::AID-IMMU3153>3.0.CO;2-0
60. Triantafilou M, Miyake K, Golenbock DT, Triantafilou K. Mediators of innate immune recognition of bacteria concentrate in lipid rafts and facilitate lipopolysaccharide-induced cell activation. J Cell Sci (2002) 115:2603–11.
61. Soong G, Reddy B, Sokol S, Adamo R, Prince A. TLR2 is mobilized into an apical lipid raft receptor complex to signal infection in airway epithelial cells. J Clin Invest (2004) 113:1482–9. doi:10.1172/JCI20773
62. Tejera E, Rocha-Perugini V, Lopez-Martin S, Perez-Hernandez D, Bachir AI, Horwitz AR, et al. CD81 regulates cell migration through its association with Rac GTPase. Mol Biol Cell (2013) 24:261–73. doi:10.1091/mbc.E12-09-0642
63. Van Zelm MC, Smet J, Adams B, Mascart F, Schandene L, Janssen F, et al. CD81 gene defect in humans disrupts CD19 complex formation and leads to antibody deficiency. J Clin Invest (2010) 120:1265–74. doi:10.1172/JCI39748
64. Mattila PK, Feest C, Depoil D, Treanor B, Montaner B, Otipoby KL, et al. The actin and tetraspanin networks organize receptor nanoclusters to regulate B cell receptor-mediated signaling. Immunity (2013) 38:461–74. doi:10.1016/j.immuni.2012.11.019
65. Pileri P, Uematsu Y, Campagnoli S, Galli G, Falugi F, Petracca R, et al. Binding of hepatitis C virus to CD81. Science (1998) 282:938–41. doi:10.1126/science.282.5390.938
66. Van Spriel AB, Figdor CG. The role of tetraspanins in the pathogenesis of infectious diseases. Microbes Infect (2010) 12:106–12. doi:10.1016/j.micinf.2009.11.001
67. Rocha-Perugini V, Lavie M, Delgrange D, Canton J, Pillez A, Potel J, et al. The association of CD81 with tetraspanin-enriched microdomains is not essential for hepatitis C virus entry. BMC Microbiol (2009) 9:111. doi:10.1186/1471-2180-9-111
68. Meuleman P, Hesselgesser J, Paulson M, Vanwolleghem T, Desombere I, Reiser H, et al. Anti-CD81 antibodies can prevent a hepatitis C virus infection in vivo. Hepatology (2008) 48:1761–8. doi:10.1002/hep.22547
69. Gordon-Alonso M, Yanez-Mo M, Barreiro O, Alvarez S, Munoz-Fernandez MA, Valenzuela-Fernandez A, et al. Tetraspanins CD9 and CD81 modulate HIV-1-induced membrane fusion. J Immunol (2006) 177:5129–37. doi:10.4049/jimmunol.177.8.5129
70. Roche PA, Furuta K. The ins and outs of MHC class II-mediated antigen processing and presentation. Nat Rev Immunol (2015) 15:203–16. doi:10.1038/nri3818
71. Szollosi J, Horejsi V, Bene L, Angelisova P, Damjanovich S. Supramolecular complexes of MHC class I, MHC class II, CD20, and tetraspan molecules (CD53, CD81, and CD82) at the surface of a B cell line JY. J Immunol (1996) 157:2939–46.
72. Rubinstein E, Le Naour F, Lagaudriere-Gesbert C, Billard M, Conjeaud H, Boucheix C. CD9, CD63, CD81, and CD82 are components of a surface tetraspan network connected to HLA-DR and VLA integrins. Eur J Immunol (1996) 26:2657–65. doi:10.1002/eji.1830261117
73. Turley SJ, Inaba K, Garrett WS, Ebersold M, Unternaehrer J, Steinman RM, et al. Transport of peptide-MHC class II complexes in developing dendritic cells. Science (2000) 288:522–7. doi:10.1126/science.288.5465.522
74. Vogt AB, Spindeldreher S, Kropshofer H. Clustering of MHC-peptide complexes prior to their engagement in the immunological synapse: lipid raft and tetraspan microdomains. Immunol Rev (2002) 189:136–51. doi:10.1034/j.1600-065X.2002.18912.x
75. Anderson HA, Hiltbold EM, Roche PA. Concentration of MHC class II molecules in lipid rafts facilitates antigen presentation. Nat Immunol (2000) 1:156–62. doi:10.1038/77842
76. Poloso NJ, Denzin LK, Roche PA. CDw78 defines MHC class II-peptide complexes that require Ii chain-dependent lysosomal trafficking, not localization to a specific tetraspanin membrane microdomain. J Immunol (2006) 177:5451–8. doi:10.4049/jimmunol.177.8.5451
77. Khandelwal S, Roche PA. Distinct MHC class II molecules are associated on the dendritic cell surface in cholesterol-dependent membrane microdomains. J Biol Chem (2010) 285:35303–10. doi:10.1074/jbc.M110.147793
78. Gartlan KH, Belz GT, Tarrant JM, Minigo G, Katsara M, Sheng KC, et al. A complementary role for the tetraspanins CD37 and Tssc6 in cellular immunity. J Immunol (2010) 185:3158–66. doi:10.4049/jimmunol.0902867
79. Benvenuti F, Lagaudriere-Gesbert C, Grandjean I, Jancic C, Hivroz C, Trautmann A, et al. Dendritic cell maturation controls adhesion, synapse formation, and the duration of the interactions with naive T lymphocytes. J Immunol (2004) 172:292–301. doi:10.4049/jimmunol.172.1.292
80. Mittelbrunn M, Martinez Del Hoyo G, Lopez-Bravo M, Martin-Cofreces NB, Scholer A, Hugues S, et al. Imaging of plasmacytoid dendritic cell interactions with T cells. Blood (2009) 113:75–84. doi:10.1182/blood-2008-02-139865
81. Rocha-Perugini V, Zamai M, Gonzalez-Granado JM, Barreiro O, Tejera E, Yanez-Mo M, et al. CD81 controls sustained T cell activation signaling and defines the maturation stages of cognate immunological synapses. Mol Cell Biol (2013) 33:3644–58. doi:10.1128/MCB.00302-13
82. Imai T, Yoshie O. C33 antigen and M38 antigen recognized by monoclonal antibodies inhibitory to syncytium formation by human T cell leukemia virus type 1 are both members of the transmembrane 4 superfamily and associate with each other and with CD4 or CD8 in T cells. J Immunol (1993) 151:6470–81.
83. Imai T, Kakizaki M, Nishimura M, Yoshie O. Molecular analyses of the association of CD4 with two members of the transmembrane 4 superfamily, CD81 and CD82. J Immunol (1995) 155:1229–39.
84. Cevik SI, Keskin N, Belkaya S, Ozlu MI, Deniz E, Tazebay UH, et al. CD81 interacts with the T cell receptor to suppress signaling. PLoS One (2012) 7:e50396. doi:10.1371/journal.pone.0050396
85. Mannion BA, Berditchevski F, Kraeft SK, Chen LB, Hemler ME. Transmembrane-4 superfamily proteins CD81 (TAPA-1), CD82, CD63, and CD53 specifically associated with integrin alpha 4 beta 1 (CD49d/CD29). J Immunol (1996) 157:2039–47.
86. Rocha-Perugini V, Gonzalez-Granado JM, Tejera E, Lopez-Martin S, Yanez-Mo M, Sanchez-Madrid F. Tetraspanins CD9 and CD151 at the immune synapse support T-cell integrin signaling. Eur J Immunol (2014) 44(7):1967–75. doi:10.1002/eji.201344235
87. Lagaudriere-Gesbert C, Lebel-Binay S, Hubeau C, Fradelizi D, Conjeaud H. Signaling through the tetraspanin CD82 triggers its association with the cytoskeleton leading to sustained morphological changes and T cell activation. Eur J Immunol (1998) 28:4332–44. doi:10.1002/(SICI)1521-4141(199812)28:12<4332::AID-IMMU4332>3.0.CO;2-8
88. Delaguillaumie A, Lagaudriere-Gesbert C, Popoff MR, Conjeaud H. Rho GTPases link cytoskeletal rearrangements and activation processes induced via the tetraspanin CD82 in T lymphocytes. J Cell Sci (2002) 115:433–43.
89. Delaguillaumie A, Harriague J, Kohanna S, Bismuth G, Rubinstein E, Seigneuret M, et al. Tetraspanin CD82 controls the association of cholesterol-dependent microdomains with the actin cytoskeleton in T lymphocytes: relevance to co-stimulation. J Cell Sci (2004) 117:5269–82. doi:10.1242/jcs.01380
90. Bell GM, Seaman WE, Niemi EC, Imboden JB. The OX-44 molecule couples to signaling pathways and is associated with CD2 on rat T lymphocytes and a natural killer cell line. J Exp Med (1992) 175:527–36. doi:10.1084/jem.175.2.527
91. Monks CR, Freiberg BA, Kupfer H, Sciaky N, Kupfer A. Three-dimensional segregation of supramolecular activation clusters in T cells. Nature (1998) 395:82–6. doi:10.1038/25764
92. Bunnell SC, Hong DI, Kardon JR, Yamazaki T, McGlade CJ, Barr VA, et al. T cell receptor ligation induces the formation of dynamically regulated signaling assemblies. J Cell Biol (2002) 158:1263–75. doi:10.1083/jcb.200203043
93. Yokosuka T, Sakata-Sogawa K, Kobayashi W, Hiroshima M, Hashimoto-Tane A, Tokunaga M, et al. Newly generated T cell receptor microclusters initiate and sustain T cell activation by recruitment of Zap70 and SLP-76. Nat Immunol (2005) 6:1253–62. doi:10.1038/ni1272
94. Varma R, Campi G, Yokosuka T, Saito T, Dustin ML. T cell receptor-proximal signals are sustained in peripheral microclusters and terminated in the central supramolecular activation cluster. Immunity (2006) 25:117–27. doi:10.1016/j.immuni.2006.04.010
95. Huse M, Klein LO, Girvin AT, Faraj JM, Li QJ, Kuhns MS, et al. Spatial and temporal dynamics of T cell receptor signaling with a photoactivatable agonist. Immunity (2007) 27:76–88. doi:10.1016/j.immuni.2007.05.017
96. Yokosuka T, Kobayashi W, Sakata-Sogawa K, Takamatsu M, Hashimoto-Tane A, Dustin ML, et al. Spatiotemporal regulation of T cell costimulation by TCR-CD28 microclusters and protein kinase C theta translocation. Immunity (2008) 29:589–601. doi:10.1016/j.immuni.2008.08.011
97. Schamel WW, Alarcon B. Organization of the resting TCR in nanoscale oligomers. Immunol Rev (2013) 251:13–20. doi:10.1111/imr.12019
98. Lillemeier BF, Mortelmaier MA, Forstner MB, Huppa JB, Groves JT, Davis MM. TCR and Lat are expressed on separate protein islands on T cell membranes and concatenate during activation. Nat Immunol (2010) 11:90–6. doi:10.1038/ni.1832
99. Sherman E, Barr V, Manley S, Patterson G, Balagopalan L, Akpan I, et al. Functional nanoscale organization of signaling molecules downstream of the T cell antigen receptor. Immunity (2011) 35:705–20. doi:10.1016/j.immuni.2011.10.004
100. Sherman E, Barr V, Samelson LE. Super-resolution characterization of TCR-dependent signaling clusters. Immunol Rev (2013) 251:21–35. doi:10.1111/imr.12010
101. Campi G, Varma R, Dustin ML. Actin and agonist MHC-peptide complex-dependent T cell receptor microclusters as scaffolds for signaling. J Exp Med (2005) 202:1031–6. doi:10.1084/jem.20051182
102. Kaizuka Y, Douglass AD, Varma R, Dustin ML, Vale RD. Mechanisms for segregating T cell receptor and adhesion molecules during immunological synapse formation in Jurkat T cells. Proc Natl Acad Sci U S A (2007) 104:20296–301. doi:10.1073/pnas.0710258105
103. Grakoui A, Bromley SK, Sumen C, Davis MM, Shaw AS, Allen PM, et al. The immunological synapse: a molecular machine controlling T cell activation. Science (1999) 285:221–7. doi:10.1126/science.285.5425.221
104. Mittelbrunn M, Molina A, Escribese MM, Yanez-Mo M, Escudero E, Ursa A, et al. VLA-4 integrin concentrates at the peripheral supramolecular activation complex of the immune synapse and drives T helper 1 responses. Proc Natl Acad Sci U S A (2004) 101:11058–63. doi:10.1073/pnas.0307927101
105. Van Zanten TS, Cambi A, Koopman M, Joosten B, Figdor CG, Garcia-Parajo MF. Hotspots of GPI-anchored proteins and integrin nanoclusters function as nucleation sites for cell adhesion. Proc Natl Acad Sci U S A (2009) 106:18557–62. doi:10.1073/pnas.0905217106
106. Bakker GJ, Eich C, Torreno-Pina JA, Diez-Ahedo R, Perez-Samper G, Van Zanten TS, et al. Lateral mobility of individual integrin nanoclusters orchestrates the onset for leukocyte adhesion. Proc Natl Acad Sci U S A (2012) 109:4869–74. doi:10.1073/pnas.1116425109
107. Montoya MC, Sancho D, Bonello G, Collette Y, Langlet C, He HT, et al. Role of ICAM-3 in the initial interaction of T lymphocytes and APCs. Nat Immunol (2002) 3:159–68. doi:10.1038/ni753
108. De La Fuente H, Mittelbrunn M, Sanchez-Martin L, Vicente-Manzanares M, Lamana A, Pardi R, et al. Synaptic clusters of MHC class II molecules induced on DCs by adhesion molecule-mediated initial T-cell scanning. Mol Biol Cell (2005) 16:3314–22. doi:10.1091/mbc.E05-01-0005
109. Nguyen K, Sylvain NR, Bunnell SC. T cell costimulation via the integrin VLA-4 inhibits the actin-dependent centralization of signaling microclusters containing the adaptor SLP-76. Immunity (2008) 28:810–21. doi:10.1016/j.immuni.2008.04.019
110. Kim M, Carman CV, Springer TA. Bidirectional transmembrane signaling by cytoplasmic domain separation in integrins. Science (2003) 301:1720–5. doi:10.1126/science.1084174
111. Kinashi T. Intracellular signalling controlling integrin activation in lymphocytes. Nat Rev Immunol (2005) 5:546–59. doi:10.1038/nri1646
112. Carman CV, Springer TA. Integrin avidity regulation: are changes in affinity and conformation underemphasized? Curr Opin Cell Biol (2003) 15:547–56. doi:10.1016/j.ceb.2003.08.003
113. Dustin ML, Bivona TG, Philips MR. Membranes as messengers in T cell adhesion signaling. Nat Immunol (2004) 5:363–72. doi:10.1038/ni1057
114. Wulfing C, Davis MM. A receptor/cytoskeletal movement triggered by costimulation during T cell activation. Science (1998) 282:2266–9. doi:10.1126/science.282.5397.2266
115. Zhang Y, Wang H. Integrin signalling and function in immune cells. Immunology (2012) 135:268–75. doi:10.1111/j.1365-2567.2011.03549.x
116. Tabdanov E, Gondarenko S, Kumari S, Liapis A, Dustin ML, Sheetz MP, et al. Micropatterning of TCR and LFA-1 ligands reveals complementary effects on cytoskeleton mechanics in T cells. Integr Biol (Camb) (2015) 7(10):1272–84. doi:10.1039/c5ib00032g
117. Babich A, Li S, O’connor RS, Milone MC, Freedman BD, Burkhardt JK. F-actin polymerization and retrograde flow drive sustained PLCgamma1 signaling during T cell activation. J Cell Biol (2012) 197:775–87. doi:10.1083/jcb.201201018
118. Yi J, Wu XS, Crites T, Hammer JA. Actin retrograde flow and actomyosin II arc contraction drive receptor cluster dynamics at the immunological synapse in Jurkat T cells. Mol Biol Cell (2012) 23:834–52. doi:10.1091/mbc.E11-08-0731
119. Comrie WA, Babich A, Burkhardt JK. F-actin flow drives affinity maturation and spatial organization of LFA-1 at the immunological synapse. J Cell Biol (2015) 208:475–91. doi:10.1083/jcb.201406121
120. Springer TA. Adhesion receptors of the immune system. Nature (1990) 346:425–34. doi:10.1038/346425a0
121. Burroughs NJ, Kohler K, Miloserdov V, Dustin ML, Van Der Merwe PA, Davis DM. Boltzmann energy-based image analysis demonstrates that extracellular domain size differences explain protein segregation at immune synapses. PLoS Comput Biol (2011) 7:e1002076. doi:10.1371/journal.pcbi.1002076
122. Dustin ML. Visualization of cell-cell interaction contacts: synapses and kinapses. Self Nonself (2011) 2:85–97. doi:10.4161/self.2.2.17931
123. Choudhuri K, Wiseman D, Brown MH, Gould K, Van Der Merwe PA. T-cell receptor triggering is critically dependent on the dimensions of its peptide-MHC ligand. Nature (2005) 436:578–82. doi:10.1038/nature03843
124. Crites TJ, Padhan K, Muller J, Krogsgaard M, Gudla PR, Lockett SJ, et al. TCR Microclusters pre-exist and contain molecules necessary for TCR signal transduction. J Immunol (2014) 193:56–67. doi:10.4049/jimmunol.1400315
125. Mittelbrunn M, Yanez-Mo M, Sancho D, Ursa A, Sanchez-Madrid F. Cutting edge: dynamic redistribution of tetraspanin CD81 at the central zone of the immune synapse in both T lymphocytes and APC. J Immunol (2002) 169:6691–5. doi:10.4049/jimmunol.169.12.6691
126. Shibagaki N, Hanada K, Yamashita H, Shimada S, Hamada H. Overexpression of CD82 on human T cells enhances LFA-1/ICAM-1-mediated cell-cell adhesion: functional association between CD82 and LFA-1 in T cell activation. Eur J Immunol (1999) 29:4081–91. doi:10.1002/(SICI)1521-4141(199912)29:12<4081::AID-IMMU4081>3.3.CO;2-9
127. Vancompernolle SE, Levy S, Todd SC. Anti-CD81 activates LFA-1 on T cells and promotes T cell-B cell collaboration. Eur J Immunol (2001) 31:823–31. doi:10.1002/1521-4141(200103)31:3<823::AID-IMMU823>3.0.CO;2-D
128. Todd SC, Lipps SG, Crisa L, Salomon DR, Tsoukas CD. CD81 expressed on human thymocytes mediates integrin activation and interleukin 2-dependent proliferation. J Exp Med (1996) 184:2055–60. doi:10.1084/jem.184.5.2055
129. Cao L, Yoshino T, Kawasaki N, Sakuma I, Takahashi K, Akagi T. Anti-CD53 monoclonal antibody induced LFA-1/ICAM-1-dependent and -independent lymphocyte homotypic cell aggregation. Immunobiology (1997) 197:70–81. doi:10.1016/S0171-2985(97)80058-7
130. Todros-Dawda I, Kveberg L, Vaage JT, Inngjerdingen M. The tetraspanin CD53 modulates responses from activating NK cell receptors, promoting LFA-1 activation and dampening NK cell effector functions. PLoS One (2014) 9:e97844. doi:10.1371/journal.pone.0097844
131. Nishiuchi R, Sanzen N, Nada S, Sumida Y, Wada Y, Okada M, et al. Potentiation of the ligand-binding activity of integrin alpha3beta1 via association with tetraspanin CD151. Proc Natl Acad Sci U S A (2005) 102:1939–44. doi:10.1073/pnas.0409493102
132. Yang XH, Mirchev R, Deng X, Yacono P, Yang HL, Golan DE, et al. CD151 restricts the alpha6 integrin diffusion mode. J Cell Sci (2012) 125:1478–87. doi:10.1242/jcs.093963
133. Gutierrez-Lopez MD, Ovalle S, Yanez-Mo M, Sanchez-Sanchez N, Rubinstein E, Olmo N, et al. A functionally relevant conformational epitope on the CD9 tetraspanin depends on the association with activated beta1 integrin. J Biol Chem (2003) 278:208–18. doi:10.1074/jbc.M207805200
134. Tadokoro S, Shattil SJ, Eto K, Tai V, Liddington RC, De Pereda JM, et al. Talin binding to integrin beta tails: a final common step in integrin activation. Science (2003) 302:103–6. doi:10.1126/science.1086652
135. Simonson WT, Franco SJ, Huttenlocher A. Talin1 regulates TCR-mediated LFA-1 function. J Immunol (2006) 177:7707–14. doi:10.4049/jimmunol.177.11.7707
136. Collins TL, Kassner PD, Bierer BE, Burakoff SJ. Adhesion receptors in lymphocyte activation. Curr Opin Immunol (1994) 6:385–93. doi:10.1016/0952-7915(94)90117-1
137. Wingren AG, Parra E, Varga M, Kalland T, Sjogren HO, Hedlund G, et al. T cell activation pathways: B7, LFA-3, and ICAM-1 shape unique T cell profiles. Crit Rev Immunol (1995) 15:235–53. doi:10.1615/CritRevImmunol.v15.i3-4.30
138. Udagawa T, Woodside DG, McIntyre BW. Alpha 4 beta 1 (CD49d/CD29) integrin costimulation of human T cells enhances transcription factor and cytokine induction in the absence of altered sensitivity to anti-CD3 stimulation. J Immunol (1996) 157:1965–72.
139. Perez OD, Mitchell D, Jager GC, South S, Murriel C, McBride J, et al. Leukocyte functional antigen 1 lowers T cell activation thresholds and signaling through cytohesin-1 and Jun-activating binding protein 1. Nat Immunol (2003) 4:1083–92. doi:10.1038/ni984
140. Suzuki J, Yamasaki S, Wu J, Koretzky GA, Saito T. The actin cloud induced by LFA-1-mediated outside-in signals lowers the threshold for T-cell activation. Blood (2007) 109:168–75. doi:10.1182/blood-2005-12-020164
141. Sato T, Tachibana K, Nojima Y, D’avirro N, Morimoto C. Role of the VLA-4 molecule in T cell costimulation. Identification of the tyrosine phosphorylation pattern induced by the ligation of VLA-4. J Immunol (1995) 155:2938–47.
142. Rose DM, Liu S, Woodside DG, Han J, Schlaepfer DD, Ginsberg MH. Paxillin binding to the alpha 4 integrin subunit stimulates LFA-1 (integrin alpha L beta 2)-dependent T cell migration by augmenting the activation of focal adhesion kinase/proline-rich tyrosine kinase-2. J Immunol (2003) 170:5912–8. doi:10.4049/jimmunol.170.12.5912
143. Mor A, Dustin ML, Philips MR. Small GTPases and LFA-1 reciprocally modulate adhesion and signaling. Immunol Rev (2007) 218:114–25. doi:10.1111/j.1600-065X.2007.00538.x
144. Yamada M, Sumida Y, Fujibayashi A, Fukaguchi K, Sanzen N, Nishiuchi R, et al. The tetraspanin CD151 regulates cell morphology and intracellular signaling on laminin-511. FEBS J (2008) 275:3335–51. doi:10.1111/j.1742-4658.2008.06481.x
145. Sawada S, Yoshimoto M, Odintsova E, Hotchin NA, Berditchevski F. The tetraspanin CD151 functions as a negative regulator in the adhesion-dependent activation of Ras. J Biol Chem (2003) 278:26323–6. doi:10.1074/jbc.C300210200
146. Hong IK, Jeoung DI, Ha KS, Kim YM, Lee H. Tetraspanin CD151 stimulates adhesion-dependent activation of Ras, Rac, and Cdc42 by facilitating molecular association between beta1 integrins and small GTPases. J Biol Chem (2012) 287:32027–39. doi:10.1074/jbc.M111.314443
147. Iwasaki T, Takeda Y, Maruyama K, Yokosaki Y, Tsujino K, Tetsumoto S, et al. Deletion of tetraspanin CD9 diminishes lymphangiogenesis in vivo and in vitro. J Biol Chem (2013) 288:2118–31. doi:10.1074/jbc.M112.424291
148. Sala-Valdes M, Ursa A, Charrin S, Rubinstein E, Hemler ME, Sanchez-Madrid F, et al. EWI-2 and EWI-F link the tetraspanin web to the actin cytoskeleton through their direct association with ezrin-radixin-moesin proteins. J Biol Chem (2006) 281:19665–75. doi:10.1074/jbc.M602116200
149. Gordon-Alonso M, Sala-Valdes M, Rocha-Perugini V, Perez-Hernandez D, Lopez-Martin S, Ursa A, et al. EWI-2 association with alpha-actinin regulates T cell immune synapses and HIV viral infection. J Immunol (2012) 189:689–700. doi:10.4049/jimmunol.1103708
150. Johnson JL, Winterwood N, Demali KA, Stipp CS. Tetraspanin CD151 regulates RhoA activation and the dynamic stability of carcinoma cell-cell contacts. J Cell Sci (2009) 122:2263–73. doi:10.1242/jcs.045997
151. Zhang F, Michaelson JE, Moshiach S, Sachs N, Zhao W, Sun Y, et al. Tetraspanin CD151 maintains vascular stability by balancing the forces of cell adhesion and cytoskeletal tension. Blood (2011) 118:4274–84. doi:10.1182/blood-2011-03-339531
152. Liu WM, Zhang F, Moshiach S, Zhou B, Huang C, Srinivasan K, et al. Tetraspanin CD82 inhibits protrusion and retraction in cell movement by attenuating the plasma membrane-dependent actin organization. PLoS One (2012) 7:e51797. doi:10.1371/journal.pone.0051797
Keywords: tetraspanins, tetraspanin-enriched microdomains, adhesion receptors, immunological synapse, T-cell activation
Citation: Rocha-Perugini V, Sánchez-Madrid F and Martínez del Hoyo G (2016) Function and Dynamics of Tetraspanins during Antigen Recognition and Immunological Synapse Formation. Front. Immunol. 6:653. doi: 10.3389/fimmu.2015.00653
Received: 17 September 2015; Accepted: 18 December 2015;
Published: 11 January 2016
Edited by:
Pedro Roda-Navarro, University Complutense of Madrid, SpainReviewed by:
Tomasz Zal, University of Texas MD Anderson Cancer Center, USAAlessandra Cambi, Radboud University Medical Centre, Netherlands
Silvia Deaglio, University of Turin, Italy
Copyright: © 2016 Rocha-Perugini, Sánchez-Madrid and Martínez del Hoyo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Francisco Sánchez-Madrid, ZnNtYWRyaWRAc2FsdWQubWFkcmlkLm9yZw==