
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Hum. Neurosci. , 10 November 2021
Sec. Motor Neuroscience
Volume 15 - 2021 | https://doi.org/10.3389/fnhum.2021.749017
This article is part of the Research Topic The Neural Control of Locomotion: Current Knowledge and Future Research View all 17 articles
 Seongmi Song1
Seongmi Song1 Andrew D. Nordin1,2,3*
Andrew D. Nordin1,2,3*Walking or running in real-world environments requires dynamic multisensory processing within the brain. Studying supraspinal neural pathways during human locomotion provides opportunities to better understand complex neural circuity that may become compromised due to aging, neurological disorder, or disease. Knowledge gained from studies examining human electrical brain dynamics during gait can also lay foundations for developing locomotor neurotechnologies for rehabilitation or human performance. Technical barriers have largely prohibited neuroimaging during gait, but the portability and precise temporal resolution of non-invasive electroencephalography (EEG) have expanded human neuromotor research into increasingly dynamic tasks. In this narrative mini-review, we provide a (1) brief introduction and overview of modern neuroimaging technologies and then identify considerations for (2) mobile EEG hardware, (3) and data processing, (4) including technical challenges and possible solutions. Finally, we summarize (5) knowledge gained from human locomotor control studies that have used mobile EEG, and (6) discuss future directions for real-world neuroimaging research.
Understanding human brain processes during real-world behaviors is a major neuroscience challenge. Moving cognitive and motor neuroscience studies beyond stationary, seated experiments and into complex, realistic environments is a necessary step forward to decipher real-world human brain dynamics. Because walking is a fundamental motor task that can have profound effects on quality of life and requires complex interactions throughout the nervous system, there is a need to better understand healthy human neuromotor control and to identify pathways affected by a loss of neurological function due to disease, disorder, injury, or aging (Snijders et al., 2007). Although basic locomotor control can be primarily attributed to subcortical structures and spinal central pattern generators, a growing body of evidence has shown that cortical structures directly modulate locomotion, including motor planning, execution, and error correction. Understanding cortical involvement during locomotion is, therefore, necessary to improve clinical detection and rehabilitation results.
Contemporary brain imaging technologies can measure neural dynamics by capturing a number of contrasting physiological signals. Neural electromagnetic signals can be measured using electroencephalography (EEG) and magnetoencephalography (MEG), changes in blood oxygenation can be measured using hemodynamic measurement methods such as magnetic resonance imaging (MRI) and functional near-infrared spectroscopy (fNIRS), and molecular imaging methods such as positron emission tomography (PET) and single-photon emission computed tomography (SPECT) enable scientists and clinicians to non-invasively study human brain functions. Technological limitations, however, have largely limited neuroimaging studies to motionless conditions from participants who are seated or lying supine due to the physical size of the recording equipment or because of noise introduced by participant or equipment motions that compromise signal recording quality. In recent years, a growing need for continuous brain monitoring during movement has promoted the development of mobile brain/body imaging (MOBI) approaches. The advantages and limitations of each technology must be considered in the context of the temporal and spatial resolution of each system and the associated cost and portability. Although fNIRS can portably measure human brain hemodynamics, and advancements in MEG technologies that rely on novel optically pumped magnetometers (OPM) (Boto et al., 2018; Hill et al., 2020; Tierney et al., 2020) provide promising paths forward for studying real-world human brain dynamics, the low-cost portability and precise temporal resolution of EEG has enabled the expansion of human neuromotor research into more complex and dynamic tasks (Allali et al., 2018).
A primary limitation of mobile EEG for studying real-world human brain dynamics has been the need to eliminate noise contamination from scalp EEG recordings. During unconstrained movements such as walking, electrode motions on the scalp and cable sway increase along with electrophysiological signals from the heart, eye movements, and facial and neck muscle activities. Low signal-to-noise ratio and comparatively poor spatial resolution in relation to fMRI and molecular imaging methods have been progressively addressed through advanced signal processing to isolate and localize electrocortical source activity using independent component analysis and forward head modeling techniques (Delorme and Makeig, 2004; Acar and Makeig, 2010; Vorwerk et al., 2018). Leveraging these advancements, recent locomotion studies have extended our understanding of human brain activity during treadmill (Castermans et al., 2014; Nathan and Contreras-Vidal, 2016; Bradford et al., 2016; Nordin et al., 2019a) and overground locomotion (Luu et al., 2017a) in complex virtual (Luu et al., 2016, 2017b) and real-world environments (Bruijn et al., 2015; An et al., 2019; Peterson and Ferris, 2019), and during robotically assisted gait (Wagner et al., 2012; Li et al., 2018).
Improved capabilities for measuring real-world human brain dynamics using state-of-the-art mobile EEG technologies will continue to advance the field of human cognitive and motor neuroscience. Here, we identify current mobile EEG technologies and analysis methods that have enabled groundbreaking discoveries into human neuromotor control during locomotion, and we briefly summarize some of the remaining challenges and paths forward for human mobile brain and body imaging studies.
Table 1 provides a summary of representative commercially available mobile high-density EEG system specifications. The range of electrode array density (number of channels), recording electrode type, system size, portability, and mass provide advantages and disadvantages for studying neural control of human locomotion. Here, we discuss considerations for measuring human electrical brain dynamics using contrasting mobile EEG system configurations.
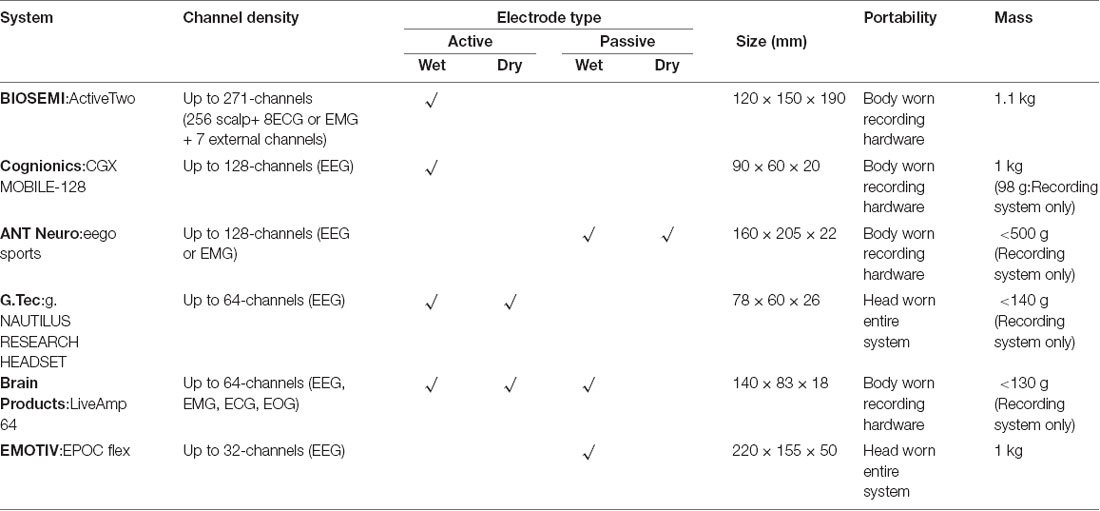
Table 1. Mobile high-density EEG systems for studying neural control of human locomotion.
Non-invasive EEG signal acquisition occurs using electrodes placed on the scalp. The recorded signal represents the summation of post-synaptic electrical potentials from the underlying and surrounding brain structures (Teplan, 2002; Sanei and Chambers, 2013), together with electrical noise from the surrounding environment, recording equipment, and ongoing electrophysiological activity from the eyes (Dement and Kleitman, 1957; Overton and Shagass, 1969; Schlögl et al., 2007), heart (Stephenson and Gibbs, 1951; Park et al., 2002), muscles (Goncharova et al., 2003; Whitham et al., 2007; Muthukumaraswamy, 2013). Electroencephalographic signals measured from the scalp show microvolt-scale fluctuations (peak-peak range: 0.5 μV–100 μV; Teplan, 2002), while noise contamination can occur at the millivolt scale (1,000× greater amplitude). The electrode characteristics and scalp-electrode interface can have large implications on EEG signal quality. Typical EEG electrodes commonly use silver/silver chloride (Ag/AgCl) that interfaces indirectly with the scalp through a conductive gel in a so-called wet electrode configuration. The Ag/AgCl wet electrodes have favorable reliability and signal integrity, with a low electrical impedance that diminishes low-frequency noise compared to high impedance electrodes recording in a warm, humid environment (Kappenman and Luck, 2010; Laszlo et al., 2014; Mathewson et al., 2017; Hinrichs et al., 2020). Any EEG setup can cause discomfort for the participant during prolonged use, but the main drawback of wet EEG electrodes is that the conductive gel dries over time, which reduces signal recording quality.
Dry contact electrodes were proposed to resolve some of the disadvantages of wet electrodes (Taheri et al., 1994; Gargiulo et al., 2008; Lopez-Gordo et al., 2014) by enabling electrical brain recordings over longer durations without a need for conductive gel (Xu et al., 2017). Additional advantages of dry contact electrodes include reduced setup time and limited inconvenience to the participant. However, dry contact electrodes show a higher impedance range and are more vulnerable to motion artifacts than wet electrodes due to the sensitive direct skin-electrode interface that is critical to mobile EEG signal recording quality (Xu et al., 2017). To overcome this, pressure is often applied to the electrodes and scalp through mechanical tension in the setup, which can lead to discomfort.
Quasi-dry electrodes, which combine advantages from both wet and dry contact electrodes, have been introduced as an intermediate solution for robust EEG signal recording (Mota et al., 2013). The quasi-dry electrode has a hydrated local skin interface with a moisturizing solution drawn from a reservoir inside the electrode. The significant advantages of quasi-dry electrodes include maintenance of lower electrode impedance similar to wet electrodes, with reduced discomfort compared to dry electrodes (Xu and Zhong, 2018). Quasi-dry electrodes also allow long-term EEG measurements due to the small amount of moisturizing solution that spreads and dries on the scalp less than typical wet electrode conductive gel (Mota et al., 2013). Quasi-dry electrodes do, however, often require additional pressure placed on the electrode to dispense the gel, which can result in non-uniform scalp pressure and discomfort.
Novel electrode configurations have also been introduced, including concentric ring electrode designs (He et al., 2001; Besio et al., 2006, 2014). By simultaneously recording from multiple closely-spaced recording sites on each electrode, signal-to-noise ratio and EEG spatial resolution can improve compared to conventional recording electrodes. Tripolar concentric ring electrodes have even outperformed bipolar and quasi-bipolar electrode designs by calculating the surface Laplacian or spatial second derivative using a multi-point differential among concentric rings on each electrode (He et al., 2001; Besio et al., 2006, 2014). Flexible electrodes provide another promising opportunity for configuring mobile EEG systems by relying on compliant, lightweight materials that can improve user comfort and enable longer duration recordings (Wang et al., 2012; Debener et al., 2015; Someya and Amagai, 2019; Acar et al., 2019; Shustak et al., 2019). Non-invasive flexible EEG electrodes, such as tattoos (Kim et al., 2011; Shustak et al., 2019; Ferrari et al., 2020) and conductive textiles (Löfhede et al., 2010, 2012), are capable of measuring electrocortical signals from the scalp, but are compromised by hair underlying the recording surface (Löfhede et al., 2012; Casson, 2019) and have therefore typically been placed on the forehead or around the ears (Kim et al., 2011; Debener et al., 2015; Acar et al., 2019; Shustak et al., 2019). These approaches have remained limited in locomotion studies or restricted to motor-related brain regions (Bunge, 2004), but fully portable and wireless EEG recording hardware with soft scalp electrodes that minimally penetrate the epidermis can provide a viable alternative (Mahmood et al., 2021). Widespread adoption of these innovative technologies remains dependent on signal recording quality, the ability to record electrical brain activity across the entire scalp surface, and user comfort. Mobile EEG recording innovations will continue to emerge and improve our abilities for measuring robust electrocortical activity during human locomotion.
In addition to the skin-electrode interface, electrode configurations and system designs have been proposed to enhance EEG signal recording quality. One representative idea is to integrate miniature amplifiers on each electrode, in a so-called active electrode configuration. Because active electrodes amplify the EEG signal at the recording site, EEG signal quality can improve by minimizing noise induced by cable sway (Mathewson et al., 2017). The influence of signal pre-amplification on EEG data quality using active electrodes, however, likely depends on the overall system configuration and relative placement of the system components (Scanlon et al., 2020). Active electrode designs can also enable skin-electrode impedance monitoring periodically throughout a data recording session to ensure low scalp-electrode impedance and high signal quality are preserved over time (Patki et al., 2012). Compared to standard passive EEG electrode configurations, each active electrode amplifier is electrically powered, often requiring additional wiring (Xu and Zhong, 2018).
Measured at the scalp, EEG electrodes record electrical potentials from the brain that are a mixture of multiple source components (Hyvärinen and Oja, 2000). Blind source separation methods, specifically independent component analysis (ICA; Bell and Sejnowski, 1995; Makeig et al., 1997), can effectively decompose the channel-based electrode recordings into independent source components. Widely available through MATLAB-based open-source scripts in EEGLab (Delorme and Makeig, 2004), ICA is central to many mobile EEG studies for isolating electrocortical signals from the complex mixture of signal and noise measured from EEG channel recordings. Independent components that are isolated from the electrode channel recordings can then be categorized into brain components and other components such as noise or other physiological signals. Although numerous ICA-based algorithms exist for deriving source components from high-density EEG channel data, an adaptive mixture independent component analysis algorithm (AMICA; Palmer et al., 2012) that relies on an unsupervised learning approach has been reported to be most effective at reducing mutual information among ICA-derived source components (Delorme et al., 2012). This approach is also able to detect time-varying brain states through multiple modeling (Hsu et al., 2018), though sufficient data are needed from long-duration EEG recordings to effectively separate source signal components, which comes at a higher computational cost.
Distinguishing independent components that originate from the brain and non-brain sources is a critical step when studying human electrical brain dynamics during locomotion. In practice, identification criteria have included, but are not limited to, scalp topography, source dipole location, time series, and power spectrum. Subjective methods based on visual inspection and objective statistical criteria have each been used for selecting electrocortical source activity derived from ICA, but the effectiveness of these approaches is dependent on ICA decomposition quality and between-subject variability (Ullsperger and Debener, 2010). To increase consistency and efficiency for classifying brain and non-brain independent components, automatic classification data processing toolboxes available in EEGLab have been made available. Some of these toolboxes include ICLabel (Pion-Tonachini et al., 2019), MARA (Multiple Artifact Rejection Algorithm; Haresign et al., 2021), FASTER (Fully Automated Statistical Thresholding for EEG Artifact Rejection; Nolan et al., 2010), SASICA (Semi-Automated Selection of Independent Components of the electroencephalogram for Artifact correction; Chaumon et al., 2015), ADJUST (Automatic EEG artifact Detection based on the Joint Use of Spatial and Temporal features; Mognon et al., 2011), and IC_MARC (Frølich et al., 2015). Although these approaches can help to distinguish the brain and non-brain source components from ICA decomposition, visual inspection is still typically advisable.
Multivariate source separation techniques used in Brain-Computer Interface (BCI) applications, may also provide viable alternatives to ICA signal decomposition. Because BCI studies have fewer trials for real-time control, preprocessing is essential (Wolpaw and McFarland, 2004; Kübler et al., 2005; Blankertz et al., 2007), with data-driven supervised decomposition algorithms typically used as a spatial filter (Blankertz et al., 2007; Nikulin et al., 2011; Dähne et al., 2014; Haufe et al., 2014). Common Spatial Patterns (CSP; Müller-Gerking et al., 1999; Ramoser et al., 2000; Blankertz et al., 2007) generate spatial filters to improve BCI classification and can improve EEG signal quality by optimizing spatial filters based on predominant event-related desynchronization or synchronization (ERD: spectral power decrease and ERS: spectral power increase, respectively) within a certain frequency band compared between conditions (Blankertz et al., 2007). Source Power Comodulation (SPoC; Dähne et al., 2014) is designed to find spatial filters for extracting oscillatory signals from continuous variables, and when applied to scalp patterns from simulation data, improved ground truth source power estimation compared to ICA (Dähne et al., 2014). Spatio-spectral decomposition (SSD; Nikulin et al., 2011) has also been used to improve signal quality within specific frequency bands by estimating noise around the frequency range of interest. Because SSD assumes that noise spans a broad frequency range from a few Hz to tens of Hz, rather than white or 1/f noise (Nikulin et al., 2011), researchers should also consider noise traits specific to each dataset.
Estimating the source locations of electrical brain activity using independent components derived from scalp EEG recordings requires the solution of a so-called inverse problem. That is, determine the source signal locations required to produce the mixture of signals recorded at the scalp electrodes. It is essential for clinical and functional brain research applications to identify the brain structures involved in a task or behavior (Cuffin, 1998; Keil et al., 2014), but finding accurate spatial source locations is difficult due to the effects of volume conduction (Jung et al., 2001) as the electrical source activity propagates through cortical tissues, cerebrospinal fluid, resistive scalp layers, and the skull (Burle et al., 2015). To solve the EEG inverse problem, various techniques, such as non-parametric and parametric methods, were proposed (Grech et al., 2008). Modeled as an electrical dipole, non-parametric approaches assume that source components distributed in the whole brain maintain fixed orientations. Such methods include LORETA (Low resolution electrical tomography; Baillet, 1998), VARETA (Variable resolution electromagnetic tomography; Valdes-Sosa et al., 2000; Bosch-Bayard et al., 2001), S-MAP (Spatial regularization; Baillet, 1998; Grech et al., 2008), ST-MAP (Spatio-temporal regularization; Baillet and Garnero, 1997; Grech et al., 2008), LAURA (Local autoregressive average; de Peralta Menendez et al., 2004), SSLOFO (Standardized shrinking LORETA-FOCUSS; Liu et al., 2005), and ALF (Adaptive standardized LORETA/FOCUSS; Schimpf et al., 2005). In contrast, parametric approaches consider dipole changes in time and try to search for the best dipole positions and orientations. These methods include FINES (First Principle Vectors; Xu et al., 2004), simulated annealing (Miga et al., 2002), and computational intelligence algorithms [e.g., Neural network (Robert et al., 2002) and Artificial neural network (Van Hoey et al., 2000)]. Source localization technologies exist to improve source location estimation by co-registering the precise location of EEG electrodes on the scalp with the subject-specific head anatomy. Imaging technologies, such as 3D scanning, ultrasound, optoelectronic, or camera-based computer vision methods, therefore, provide opportunities to improve electrical source localization accuracy (Koessler et al., 2010; Baysal and Sengül, 2010; Shirazi and Huang, 2019) when combined with forward head models that incorporate subject-specific MRI scans and conductivity estimates for anatomical head structures.
The millisecond temporal resolution of EEG enables the study of precise electrocortical dynamics during rapid movements or in reactive real-world scenarios. Many EEG analysis methods have been used to record electrocortical responses elicited by external stimuli. By studying changes in electrical potentials from the brain that are tied to an event of interest, such as auditory, visual, somatosensory, or vestibular cues, event-related potential (ERP) studies have uncovered changes in electrical brain activity during cognitive and motor behaviors (Kappenman and Luck, 2010). Event-related potentials represent phase-locked neural responses that can be measured during experimental manipulations (Gutberlet et al., 2009; Nidal and Malik, 2014) by repeating an event of interest (Galambos, 1992) to study the latency, morphology, and scalp topography of positive and negative voltage peaks and deflections. These analyses can be extended to study electrocortical changes in both time and frequency. During EEG analyses, the spectral power is used to study the distribution of signal power among frequencies (Sanei and Chambers, 2013) that are grouped into frequency bands based on functional roles and characteristics in the brain, including delta (1–3 Hz), theta (4–7 Hz), alpha (8–12 Hz), beta (15–30 Hz), and gamma bands (>30 Hz). Lower frequency bands indicate a subconscious state, while higher frequency reflects a more active and aroused state (Jensen et al., 2016). In order to track the temporal changes of the frequency spectrum, time-frequency EEG analyses were proposed. Event-related spectral perturbation (ERSP) analyses can show stimulus-induced, non-phase-locked brain activity over time (Tallon-Baudry et al., 1996; Rossi et al., 2014) and can provide insight into specific frequency bands that relate to functional brain processes (Rossi et al., 2014).
Beyond quantifying spatial, temporal, and spectral dynamics of electrical brain activity, the use of mobile EEG for studying the neural control of human locomotion can improve our understanding of functional interactions between brain structures and between brain and muscle during locomotor control. When significant temporal or spatial correlations are observed between neurophysiological processes, this phenomenon is often referred to as functional connectivity (Fingelkurts et al., 2005; Sakkalis, 2011). Coherence or correlation strength is considered directly proportional to the degree of functional connectivity between neuroanatomical structures when comparing electrophysiological signals (Thatcher et al., 1986; Im, 2018). To quantify signal interdependence without consideration for directional causation (Bullock et al., 1995; Kaplan et al., 1997), non-directed functional connectivity metrics such as correlation, coherence, mutual information, phase locking value, and pairwise phase consistency have been used (Bastos and Schoffelen, 2016). To quantify directed functional connectivity with consideration for causation, metrics such as cross-correlation, phase slope index, Granger causality, and transfer entropy, have been applied (Granger, 1969; Bastos and Schoffelen, 2016). These metrics can also be categorized based on considerations for signal amplitude or phase. Signal amplitude comparisons are conducted using correlation, mutual information, cross-correlation, Granger’s causality, partial directed coherence, transfer entropy, and dynamic causal modeling metrics (Im, 2018). Phase domain analyses include coherence, phase locking, pairwise phase consistency, and phase slope index (Im, 2018). Phase comparisons can also be assessed using Granger causality methods that include both parametric and non-parametric approaches (Geweke, 1982). Non-parametric Granger causality is calculated using autoregression and does not require model order to be determined (Bastos and Schoffelen, 2016). Parametric Granger causality is based on Fourier or wavelet-based methods, which require less data than non-parametric equivalents (Bastos and Schoffelen, 2016). For single-trial data and when model order is known, parametric Granger causality methods have shown greater sensitivity for quantifying neural functional connectivity compared to non-parametric Granger causality methods (Richter et al., 2015; Bastos and Schoffelen, 2016).
Electrophysiological signals not limited to electrical brain activity are detectable from scalp EEG measurements. The heart rhythm, eye movements, and electrical muscle activity can influence each obscure electrocortical source activity and may also present contrasting signal characteristics that require specific noise removal strategies beyond independent component analysis. Cardiac activity is detectable from EEG measurements when the electrode is placed on or near a blood vessel (Goncharova et al., 2003). The repetitive low-frequency (~1.2 Hz) signal characteristics of the heart rhythm are distinguishable from cerebral activity, and removal can be assisted by relying on a reference waveform or an electrocardiogram (ECG; Jiang et al., 2019).
Ocular artifacts caused by eye blinks and saccades present large amplitude and low-frequency voltage fluctuations compared to EEG signals. Because of the different signal characteristics between brain activity and eye movements, electrooculographic (EOG) recordings from electrodes placed on the skin around the eye can help to parse EOG from EEG signals using ICA or regression methods (Jung et al., 1998; Li et al., 2006; Schlögl et al., 2007; Winkler et al., 2015). Alternative statistical decomposition methods that can be implemented in a sliding window to remove transient large amplitude artifacts have proven to be particularly effective. Artifact subspace reconstruction (ASR; Kothe and Jung, 2016) is a component-based artifact removal method that can be used to clean large-variance signal components based on thresholds compared to clean baseline data and subsequent reconstruction of EEG channel data. By pre-conditioning EEG channel data and removing eye movement and muscle artifacts ahead of ICA, it is possible to improve ICA decomposition quality (Chang et al., 2019).
Myoelectric artifacts can appear in EEG signals due to muscular contractions from the scalp, face, and neck. Electromyographic (EMG) recordings can present broad spectral distributions, including low and high frequencies (>200 Hz; Shackman et al., 2009; Urigüen and Garcia-Zapirain, 2015) but are usually more localized in higher frequency bands above 14 Hz (Narasimhan and Dutt, 1996). Conventional low pass filtering approaches can remove high-frequency signal content but may also eliminate electrocortical signals in beta (13–30 Hz) or gamma bands (>30 Hz) depending on the selected filter cutoff. To minimize the risk of undesirable signal loss, canonical correlation analysis (CCA) has been used to remove muscle artifacts from EEG data (De Clercq et al., 2006; Raghavendra and Dutt, 2011; Jiang et al., 2019). CCA measures the linear relationship between two datasets to derive signal components based on correlation or autocorrelation when derived from a single dataset, such as EEG channel recordings. Due to the high-frequency spectral characteristics of electrical muscle activity, canonical components with low autocorrelation tend to show high-frequency spectral content indicative of electrical muscle activity. Component removal or filtering can therefore help to eliminate myoelectric EEG signal contamination.
In addition to the mixture of electrophysiological signals captured by EEG recording electrodes, external noise sources can contaminate mobile EEG data. External artifacts can include alternating current power line noise, electromagnetic interference from electronic devices in the surrounding environment, and movement-related artifacts introduced by the movement of mobile EEG system components, such as cables and electrodes. Alternating current power line noise predominantly occurs at either 50 Hz or 60 Hz, depending on the country, and can largely be eliminated using a notch filter at the respective frequency band (Leske and Dalal, 2019). However, notch filtering can eliminate electrocortical target signals in gamma band (>30 Hz) depending on notch filter width. Alternative methods for power line noise removal have been implemented, such as Discrete Fourier Transform (DFT) filter (Oostenveld et al., 2011), frequency-domain regression (Bigdely-Shamlo et al., 2015), and spectrum interpolation (Leske and Dalal, 2019).
Inherent to the study of neural control of locomotion using mobile EEG, gait-related movement artifacts have posed non-trivial challenges to researchers. Small electrode motions on the scalp, cable sway (Symeonidou et al., 2018), and system component vibrations introduce signal contamination during each step of the gait cycle, causing voltage fluctuations in the EEG signal that exceed electrical brain activity. Signal fluctuations occur at the step frequency but can also extend into higher frequency bands at harmonics of the step cycle, with a non-uniform influence of noise among recording electrodes across the scalp (Kline et al., 2015; Snyder et al., 2015). Cable bundling and more effectively securing electrodes and system components to the participant can minimize motion artifact causes (Nathan and Contreras-Vidal, 2016), but it remains difficult to completely eliminate motion-induced noise through ICA decomposition methods alone. High pass filtering mobile EEG data provides a partial solution (Winkler et al., 2015), with a 1–2 Hz high pass filter improving subsequent ICA decomposition results, but a number of alternative signal processing solutions have been implemented in mobile EEG studies. Adaptive filtering (Kilicarslan et al., 2016), template regression (Gwin et al., 2010), and component-based statistical decomposition methods, including artifact subspace reconstruction (Chang et al., 2018, 2019), have been used to eliminate motion artifacts from mobile EEG data at relatively slow gait speeds (<1.0 m/s; Gwin et al., 2010; Wagner et al., 2012, 2016; Bradford et al., 2016, 2019; Oliveira et al., 2017a,b; Bradford et al., 2019), but gait speeds closer to, and in excess of, preferred human walking speed (1.4 m/s; Bohannon, 1997) have remained challenging and have required novel solutions.
Many EEG preprocessing procedures have been developed and incorporated into EEGLab toolboxes to provide standardized data analysis pipelines for improving rigor and reproducibility among mobile EEG studies (Bigdely-Shamlo et al., 2015; Gabard-Durnam et al., 2018; Pedroni et al., 2019). The PREP pipeline (Bigdely-Shamlo et al., 2015), AUTOMAGIC (Pedroni et al., 2019), and HAPPE (Harvard Automated Processing Pipeline for EEG) have each introduced functions for analyzing EEG data. These toolboxes include methods for identifying and removing unusually noisy channel data (findNoisyChannels; Bigdely-Shamlo et al., 2015; Pedroni et al., 2019), a multi-stage robust referencing scheme that eliminates noise from recorded EEG signals prior to computing a common average reference (Bigdely-Shamlo et al., 2015), alternating current powerline noise removal (cleanLineNoise and ZapLine; de Cheveigné, 2020), and artifact corrections for eye movements (Pedroni et al., 2019). Automatic open-source data processing toolboxes can improve consistency among mobile EEG analyses, but it remains important to ensure that mobile EEG hardware is configured to eliminate as many possible signal contaminating noise sources ahead of EEG data recording.
A common approach for mobile EEG artifact removal is to rely on simultaneously recorded reference signals from sources known to exist in the complex mixture of signals captured in EEG recordings (e.g., EOG, EMG, or ECG). Similar approaches using isolated motion and/or electrical noise recordings have been developed and applied. Using information about the participant’s head motions during gait is one possible solution for quantifying the causes of motion artifacts in mobile EEG. Optoelectronic motion capture, accelerometry, or inertial measurement units can be used for this purpose (Casson, 2019). Adopted from solutions to overcome significant signal contamination introduced by gradient artifacts during simultaneous MRI and EEG (Chowdhury et al., 2014), isolated noise recordings have recently been used to eliminate motion artifacts from mobile EEG during human locomotion using dual-layer EEG (Nordin et al., 2018). In this configuration, one layer of EEG electrodes measured a mixture of physiological signals and motion artifacts from the scalp, but the second layer of electrodes measured only electrical noise and motion artifacts from mechanically coupled but electrically isolated secondary electrodes. Noise-only electrodes were referenced to an overlaid conductive fabric cap that served as an artificial skin circuit but also more effectively secured the recording electrodes to the participant’s head. By conducting benchmark tests using a robotically controlled motion platform that reproduced human head motions during walking and an electronic head phantom device that generated ground truth artificial brain signals (Nordin et al., 2018; Richer et al., 2020), the ability of dual-layer mobile EEG for motion artifact removal was validated. These methods were subsequently applied during human treadmill locomotion at a range of gait speeds (Nordin et al., 2019a,b), including while navigating over unexpected obstacles on a treadmill belt (Nordin et al., 2019c).
Recent mobile EEG studies have expanded our understanding of human supraspinal locomotor control, revealing electrocortical spectral power fluctuations tied to each step in the gait cycle (Wagner et al., 2012, 2016; Bradford et al., 2016, 2019; Luu et al., 2017a; Nordin et al., 2019a,c). Broadly distributed electrocortical network dynamics further show activations from the frontal cortex (Sipp et al., 2013; Bulea et al., 2015; Wagner et al., 2016), anterior cingulate cortex (Bulea et al., 2015; Bradford et al., 2016; Wagner et al., 2016; Luu et al., 2017a; Yokoyama et al., 2020), sensorimotor cortex (Wagner et al., 2012; Sipp et al., 2013; Bradford et al., 2016; Luu et al., 2017a; Nordin et al., 2019a; Yokoyama et al., 2020), auditory cortex (Wagner et al., 2016; Nordin et al., 2019a,b), supplementary motor area (Nordin et al., 2019c), premotor cortex (Nordin et al., 2019c), motor cortex (Wagner et al., 2012; Bulea et al., 2015), somatosensory cortex (Yokoyama et al., 2020), and the parietal cortex (Bulea et al., 2015; Bradford et al., 2016; Wagner et al., 2016; Luu et al., 2017a) during locomotor tasks ranging from steady treadmill gait to navigating over complex terrain or walking with robotic assistance. Knowledge gained from these studies provides the basis for understanding human electrocortical dynamics during balance and gait control that can be used for the development of assistive devices and neuroprostheses for rehabilitation, and to restore locomotor function for individuals with neurological disorders and disease.
A growing body of evidence shows dynamic cortical activations during the initiation, maintenance, and modification of human gait (Choi and Bastian, 2007; Grillner et al., 2008; Wagner et al., 2012; Castermans et al., 2014; Bradford et al., 2016; Nordin et al., 2019c). During each step of the gait cycle, sensorimotor electrocortical spectral power modulations occur in alpha (8–12 Hz) and beta bands (13–30 Hz). Multiple studies have shown alpha and beta band spectral power increases during double support and decreases during limb swing of continuous gait (Gwin et al., 2010; Wagner et al., 2012; Bulea et al., 2015; Bradford et al., 2016, 2019; Oliveira et al., 2017b; Nordin et al., 2018, 2019a,b). Detectable changes in electrocortical spectral power have also been identified between uphill, downhill, and level treadmill gait (Bradford et al., 2016), walking with eyes closed compared to eyes open (Oliveira et al., 2017b), during transitions in gait speed (Wagner et al., 2016), and when navigating over complex terrain (Luu et al., 2017a). Compared to level and downhill walking (Bradford et al., 2016), during incline walking spectral power from the anterior cingulate cortex, sensorimotor cortex, and the posterior parietal cortex increased in theta band (4–7 Hz) and decreased in gamma band (>30 Hz). Walking with restricted vision induced desynchronization from theta to beta bands during the transition to single support for the somatosensory cortex (Oliveira et al., 2017b), suggesting that restricted vision increases sensory processing and integration compared to visually guided walking. Although changes in gait speed can be largely controlled subcortically, alpha and beta band sensorimotor electrocortical spectral power were shown to decrease at faster gait speeds (2.0 m/s) compared to slower walking (0.5 m/s; Luu et al., 2017a). While navigating complex overground terrain that included level ground, ramps, and stairs, participants showed reduced alpha and beta band spectral power from sensorimotor cortex during ramp and stair ascent compared to level-ground walking (Luu et al., 2017a). Beta and gamma-band spectral power also increased from the sensorimotor cortex during initial limb swing while ascending stairs. Collectively, these findings uncover a distributed network of cortical activity involved in movement control and sensory processing during gait, leaving considerable work to uncover dynamic interactions among brain structures and how these locomotor network dynamics differ among populations.
During bipedal gait, dynamic balance maintains upright posture by supporting body weight. The ability to maintain and recover from the loss of balance is critical to healthy gait function. The unexpected loss of balance due to external perturbations has been associated with electrocortical activations from primary sensory and motor cortices, supplementary motor area, premotor cortex, anterior cingulate cortex, prefrontal cortex, temporal cortex, parietal cortex, and visual cortex (Massion et al., 1999; Slobounov et al., 2009; Sipp et al., 2013; Marlin et al., 2014; Varghese et al., 2019). Loss of balance while walking on a balance beam has also shown greater theta band power and reduced beta power from the sensorimotor cortex compared to steady treadmill walking (Sipp et al., 2013). Divergent electrical brain dynamics also emerge during a loss of balance due to physical or visual perturbations (Peterson and Ferris, 2018). While walking on a balance beam, participants who experienced a physical pull at the waist, compared to a visual rotation of the environment using a virtual reality headset, showed increased spectral power from the sensorimotor cortex in the theta band and decreased beta power after perturbation onset (Peterson and Ferris, 2018). Compared to the loss of balance due to a physical pull at the waist, however, visual perturbations elicited more prominent responses from the parieto-occipital areas. During recovery from loss of balance during unexpected slips, compared to steady walking, spectral power from sensorimotor cortex similarly increased in the theta band and decreased in the alpha band, while alpha and beta band spectral power decreased from the parietal cortex (An et al., 2019). In response to unexpected obstacles that appeared on a treadmill belt during walking and running, event-related spectral power fluctuations from time-frequency analysis further identified spectral power increases from the premotor cortex, supplementary motor area, and the parietal cortex in the delta, theta, and alpha bands. The timing of electrocortical activation onset varied with locomotion speed, initiating two steps before stepping over the obstacle to enable foot placement planning around the obstacle (Nordin et al., 2019c). The ability to detect perturbation onset in advance of motor responses could provide bio signals for developing brain-machine interface technologies to properly assist in counteracting the loss of balance due to perturbations or changes in the environment during standing balance and gait.
Robotic-assistive devices are widely used for rehabilitation purposes to provide bodyweight support or to guide locomotor limb movements. To better understand the influence of robotic assistance on human locomotor control, researchers have studied changes in electrocortical spectral dynamics using mobile EEG. By comparing electrical brain activity during active treadmill gait to walking with assistive forces applied to the limbs or passive limb motions with bodyweight support, changes in sensorimotor processing have been uncovered. During robotically-assisted gait that provides bodyweight support and limb guidance, spectral power from premotor cortex and sensorimotor areas increased in alpha, beta, and gamma bands, compared to active treadmill walking (Wagner et al., 2012; Knaepen et al., 2014; Seeber et al., 2014). Recent mobile EEG studies that used a unilateral lower-limb exoskeleton to generate assistive joint torque outside the laboratory also showed hemispherical effects on parietooccipital regions in beta band compared to walking without robotic assistance (Li et al., 2018). As assistive robotic technologies for rehabilitation continue to develop, it becomes more important to better understand healthy human brain dynamics during locomotion. This knowledge not only informs how changes in electrocortical activity influence movement control, but also provides possible biomarkers for measuring adaptation to assistive devices or tracking rehabilitative progress and provides the mechanism for identifying electrocortical control signals that can be used for brain-machine interface technologies.
Brain-machine interfaces have shown increasingly promising applications for controlling output devices using direct communication with the human brain (Millán et al., 2004; Lebedev and Nicolelis, 2017; Tariq et al., 2018). Non-invasive EEG-based brain-machine interface systems can provide effective closed-loop strategies for deciphering user intentions while controlling physical or virtual machines, including multi-directional brain-actuated wheelchairs (Vanacker et al., 2007; Galán et al., 2008) or lower-limb exoskeletons (Noda et al., 2012; Contreras-Vidal and Grossman, 2013; Sczesny-Kaiser et al., 2015). Recent brain-computer interface demonstrations have allowed users to control a walking avatar in virtual reality using scalp EEG signals. In this application, alpha band spectral power from the posterior parietal cortex and inferior parietal lobe decreased along with increased gamma band spectral power from the anterior cingulate cortex, which has been attributed to error monitoring during walking (Luu et al., 2016, 2017b). Continued innovations using non-invasive EEG-based brain-machine interfaces can therefore advance current capabilities and lead to more intuitive assistive devices for rehabilitation, injury prevention, or human performance enhancement using the user’s own neurophysiological control signals.
In this review, we discussed mobile EEG technologies, including advancements in hardware and signal processing technologies for mobile applications. We identified different hardware strategies for improving signal recording quality and contemporary analytical methods for effectively extracting electrocortical source signals that can be localized to specific cortical structures with progressively better spatial resolution. We also identified the benefits of these fast timescale recordings for studying changes in voltages and spectral power that can be used to better understand how electrical brain activity changes during dynamic behaviors. Current state-of-the-art mobile EEG methods have led to considerable discoveries in the neural control of human locomotion, and through continued mobile hardware and signal processing innovations, new discoveries will continue to emerge that will enable studies in more realistic tasks and environments. Recent locomotion studies have revealed complex electrocortical dynamics that can be measured in time, space, and frequency. Effectively extracting and decoding these electrical brain signals will enable the development of robust, non-invasive brain-computer interface technologies for use in restoring, maintaining, and improving human gait.
Future improvements in mobile EEG technologies that enhance system usability will lead to more widespread adoption of these methods, including the use of compact, lightweight, and wireless system designs that can be entirely worn on the head and require reduced preparation time, but are also comfortable for the user to wear and remove to enable robust long-term recordings (Hairston et al., 2014; Izdebski et al., 2016; Oliveira et al., 2016; Bateson et al., 2017; Athavale and Krishnan, 2017). Analytical methods that cssan be realistically implemented in real-time for closed-loop applications will also enable the development of next-generation neurotechnologies. In addition to advancements in mobile EEG recording and analysis methods, unified approaches for simultaneously recording biosignals, such as eye gaze, electromyography, and biomechanical measures for quantifying whole-body human movement outside of conventional laboratory environments, will continue to expand the study of neural control of human locomotion into real-world scenarios.
SS and AN drafted and edited the manuscript. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acar, Z. A., and Makeig, S. (2010). Neuroelectromagnetic forward head modeling toolbox. J. Neurosci. Methods 190, 258–270. doi: 10.1016/j.jneumeth.2010.04.031
Acar, G., Ozturk, O., Golparvar, A. J., Elboshra, T. A., Böhringer, K., and Yapici, M. K. (2019). Wearable and flexible textile electrodes for biopotential signal monitoring: a review. Electronics 8:479. doi: 10.3390/electronics8050479
Allali, G., Blumen, H. M., Devanne, H., Pirondini, E., Delval, A., and Van De Ville, D. (2018). Brain imaging of locomotion in neurological conditions. Neurophysiol. Clin. 48, 337–359. doi: 10.1016/j.neucli.2018.10.004
An, J., Yoo, D., and Lee, B.-C. (2019). “Electrocortical activity changes in response to unpredictable trip perturbations induced by a split-belt treadmill,” in Proceedings of the 2019 41st Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC) (Berlin, Germany: IEEE), 110–113.
Athavale, Y., and Krishnan, S. (2017). Biosignal monitoring using wearables: observations and opportunities. Biomed. Signal Process. Control 38, 22–33. doi: 10.1016/j.bspc.2017.03.011
Baillet, S. (1998). Toward Functional Brain Imaging of Cortical Electrophysiology Markovian Models for Magneto and Electroencephalogram Source Estimation and Experimental Assessments. Orsay, France: University of Paris XI.
Baillet, S., and Garnero, L. (1997). A Bayesian approach to introducing anatomo-functional priors in the EEG/MEG inverse problem. IEEE Trans. Biomed. Eng. 44, 374–385. doi: 10.1109/10.568913
Bastos, A. M., and Schoffelen, J.-M. (2016). A tutorial review of functional connectivity analysis methods and their interpretational pitfalls. Front. Syst. Neurosci. 9:175. doi: 10.3389/fnsys.2015.00175
Bateson, A. D., Baseler, H. A., Paulson, K. S., Ahmed, F., and Asghar, A. U. (2017). Categorisation of mobile EEG: a researcher’s perspective. Biomed. Res. Int. 2017:5496196. doi: 10.1155/2017/5496196
Baysal, U., and Sengül, G. (2010). Single camera photogrammetry system for EEG electrode identification and localization. Ann. Biomed. Eng. 38, 1539–1547. doi: 10.1007/s10439-010-9950-4
Bell, A. J., and Sejnowski, T. J. (1995). An information-maximization approach to blind separation and blind deconvolution. Neural Comput. 7, 1129–1159. doi: 10.1162/neco.1995.7.6.1129
Besio, G., Koka, K., Aakula, R., and Dai, W. (2006). Tri-polar concentric ring electrode development for Laplacian electroencephalography. IEEE Trans. Biomed. Eng. 53, 926–933. doi: 10.1109/TBME.2005.863887
Besio, W. G., Martínez-Juárez, I. E., Makeyev, O., Gaitanis, J. N., Blum, A. S., Fisher, R. S., et al. (2014). High-frequency oscillations recorded on the scalp of patients with epilepsy using tripolar concentric ring electrodes. IEEE J. Transl. Eng. Health Med. 2:2000111. doi: 10.1109/JTEHM.2014.2332994
Bigdely-Shamlo, N., Mullen, T., Kothe, C., Su, K.-M., and Robbins, K. A. (2015). The PREP pipeline: standardized preprocessing for large-scale EEG analysis. Front. Neuroinform. 9:16. doi: 10.3389/fninf.2015.00016
Blankertz, B., Tomioka, R., Lemm, S., Kawanabe, M., and Muller, K.-R. (2007). Optimizing spatial filters for robust EEG single-trial analysis. IEEE Signal Process. Mag. 25, 41–56. doi: 10.1109/msp.2008.4408441
Bohannon, R. W. (1997). Comfortable and maximum walking speed of adults aged 20—79 years: reference values and determinants. Age Ageing 26, 15–19. doi: 10.1093/ageing/26.1.15
Bosch-Bayard, J., Valdés-Sosa, P., Virues-Alba, T., Aubert-Vázquez, E., John, E. R., Harmony, T., et al. (2001). 3D statistical parametric mapping of EEG source spectra by means of variable resolution electromagnetic tomography (VARETA). Clin. Electroencephalogr. 32, 47–61. doi: 10.1177/155005940103200203
Boto, E., Holmes, N., Leggett, J., Roberts, G., Shah, V., Meyer, S. S., et al. (2018). Moving magnetoencephalography towards real-world applications with a wearable system. Nature 555, 657–661. doi: 10.1038/nature26147
Bradford, J. C., Lukos, J. R., and Ferris, D. P. (2016). Electrocortical activity distinguishes between uphill and level walking in humans. J. Neurophysiol. 115, 958–966. doi: 10.1152/jn.00089.2015
Bradford, J. C., Lukos, J. R., Passaro, A., Ries, A., and Ferris, D. P. (2019). Effect of locomotor demands on cognitive processing. Sci. Rep. 9:9234. doi: 10.1038/s41598-019-45396-5
Bruijn, S. M., Van Dieën, J. H., and Daffertshofer, A. (2015). Beta activity in the premotor cortex is increased during stabilized as compared to normal walking. Front. Hum. Neurosci. 9:593. doi: 10.3389/fnhum.2015.00593
Bulea, T. C., Kim, J., Damiano, D. L., Stanley, C. J., and Park, H.-S. (2015). Prefrontal, posterior parietal and sensorimotor network activity underlying speed control during walking. Front. Hum. Neurosci. 9:247. doi: 10.3389/fnhum.2015.00247
Bullock, T., McClune, M., Achimowicz, J., Iragui-Madoz, V., Duckrow, R., and Spencer, S. (1995). EEG coherence has structure in the millimeter domain: subdural and hippocampal recordings from epileptic patients. Electroencephalogr. Clin. Neurophysiol. 95, 161–177. doi: 10.1016/0013-4694(95)93347-a
Bunge, S. A. (2004). How we use rules to select actions: a review of evidence from cognitive neuroscience. Cogn. Affect. Behav. Neurosci. 4, 564–579. doi: 10.3758/cabn.4.4.564
Burle, B., Spieser, L., Roger, C., Casini, L., Hasbroucq, T., and Vidal, F. (2015). Spatial and temporal resolutions of EEG: is it really black and white? A scalp current density view. Int. J. Psychophysiol. 97, 210–220. doi: 10.1016/j.ijpsycho.2015.05.004
Casson, A. J. (2019). Wearable EEG and beyond. Biomed. Eng. Lett. 9, 53–71. doi: 10.1007/s13534-018-00093-6
Castermans, T., Duvinage, M., Cheron, G., and Dutoit, T. (2014). About the cortical origin of the low-delta and high-gamma rhythms observed in EEG signals during treadmill walking. Neurosci. Lett. 561, 166–170. doi: 10.1016/j.neulet.2013.12.059
Chang, C.-Y., Hsu, S.-H., Pion-Tonachini, L., and Jung, T.-P. (2018). “Evaluation of artifact subspace reconstruction for automatic EEG artifact removal,” in Proceedings of the 2018 40th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC) (Honolulu, HI, USA: IEEE), 1242–1245.
Chang, C.-Y., Hsu, S.-H., Pion-Tonachini, L., and Jung, T.-P. (2019). Evaluation of artifact subspace reconstruction for automatic artifact components removal in multi-channel EEG recordings. IEEE Trans. Biomed. Eng. 67, 1114–1121. doi: 10.1109/TBME.2019.2930186
Chaumon, M., Bishop, D. V., and Busch, N. A. (2015). A practical guide to the selection of independent components of the electroencephalogram for artifact correction. J. Neurosci. Methods 250, 47–63. doi: 10.1016/j.jneumeth.2015.02.025
Choi, J. T., and Bastian, A. J. (2007). Adaptation reveals independent control networks for human walking. Nat. Neurosci. 10, 1055–1062. doi: 10.1038/nn1930
Chowdhury, M. E., Mullinger, K. J., Glover, P., and Bowtell, R. (2014). Reference layer artefact subtraction (RLAS): a novel method of minimizing EEG artefacts during simultaneous fMRI. NeuroImage 84, 307–319. doi: 10.1016/j.neuroimage.2013.08.039
Contreras-Vidal, J. L., and Grossman, R. G. (2013). “NeuroRex: a clinical neural interface roadmap for EEG-based brain machine interfaces to a lower body robotic exoskeleton,” in Proceedings of the 2013 35th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (Osaka, Japan: EMBC), 1579–1582.
Cuffin, B. N. (1998). EEG dipole source localization. IEEE Eng. Med. Biol. Mag. 17, 118–122. doi: 10.1109/51.715495
Dähne, S., Meinecke, F. C., Haufe, S., Höhne, J., Tangermann, M., Müller, K.-R., et al. (2014). SPoC: a novel framework for relating the amplitude of neuronal oscillations to behaviorally relevant parameters. NeuroImage 86, 111–122. doi: 10.1016/j.neuroimage.2013.07.079
Debener, S., Emkes, R., De Vos, M., and Bleichner, M. (2015). Unobtrusive ambulatory EEG using a smartphone and flexible printed electrodes around the ear. Sci. Rep. 5:16743. doi: 10.1038/srep16743
de Cheveigné, A. (2020). ZapLine: a simple and effective method to remove power line artifacts. NeuroImage 207:116356. doi: 10.1016/j.neuroimage.2019.116356
De Clercq, W., Vergult, A., Vanrumste, B., Van Paesschen, W., and Van Huffel, S. (2006). Canonical correlation analysis applied to remove muscle artifacts from the electroencephalogram. IEEE Trans. Biomed. Eng. 53, 2583–2587. doi: 10.1109/TBME.2006.879459
Delorme, A., and Makeig, S. (2004). EEGLAB: an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 134, 9–21. doi: 10.1016/j.jneumeth.2003.10.009
Delorme, A., Palmer, J., Onton, J., Oostenveld, R., and Makeig, S. (2012). Independent EEG sources are dipolar. PLoS One 7:e30135. doi: 10.1371/journal.pone.0030135
Dement, W., and Kleitman, N. (1957). Cyclic variations in EEG during sleep and their relation to eye movements, body motility, and dreaming. Electroencephalogr. Clin. Neurophysiol. 9, 673–690. doi: 10.1016/0013-4694(57)90088-3
de Peralta Menendez, R. G., Murray, M. M., Michel, C. M., Martuzzi, R., and Andino, S. L. G. (2004). Electrical neuroimaging based on biophysical constraints. NeuroImage 21, 527–539. doi: 10.1016/j.neuroimage.2003.09.051
Ferrari, L. M., Ismailov, U., Badier, J.-M., Greco, F., and Ismailova, E. (2020). Conducting polymer tattoo electrodes in clinical electro-and magneto-encephalography. NPJ Flexible Electronics 4:4. doi: 10.1038/s41528-020-0067-z
Fingelkurts, A. A., Fingelkurts, A. A., and Kähkönen, S. (2005). Functional connectivity in the brain—is it an elusive concept? Neurosci. Biobehav. Rev. 28, 827–836. doi: 10.1016/j.neubiorev.2004.10.009
Frølich, L., Andersen, T. S., and Mørup, M. (2015). Classification of independent components of EEG into multiple artifact classes. Psychophysiology 52, 32–45. doi: 10.1111/psyp.12290
Gabard-Durnam, L. J., Mendez Leal, A. S., Wilkinson, C. L., and Levin, A. R. (2018). The Harvard Automated Processing Pipeline for Electroencephalography (HAPPE): standardized processing software for developmental and high-artifact data. Front. Neurosci. 12:97. doi: 10.3389/fnins.2018.00097
Galambos, R. (1992). “A comparison of certain gamma band (40-Hz) brain rhythms in cat and man,” in Induced Rhythms in the Brain, eds E. Basar and T. H. Bullock (Boston: Springer), 201–216.
Galán, F., Nuttin, M., Lew, E., Ferrez, P. W., Vanacker, G., Philips, J., et al. (2008). A brain-actuated wheelchair: asynchronous and non-invasive brain-computer interfaces for continuous control of robots. Clin. Neurophysiol. 119, 2159–2169. doi: 10.1016/j.clinph.2008.06.001
Gargiulo, G., Bifulco, P., Calvo, R. A., Cesarelli, M., Jin, C., and Van Schaik, A. (2008). “A mobile EEG system with dry electrodes,” in Proceedings of the 2008 IEEE Biomedical Circuits and Systems Conference (Baltimore, MD, USA: IEEE), 20–22.
Geweke, J. (1982). Measurement of linear dependence and feedback between multiple time series. J. Am. Stat. Assoc. 77, 304–313. doi: 10.1080/01621459.1982.10477803
Goncharova, I. I., McFarland, D. J., Vaughan, T. M., and Wolpaw, J. R. (2003). EMG contamination of EEG: spectral and topographical characteristics. Clin. Neurophysiol. 114, 1580–1593. doi: 10.1016/s1388-2457(03)00093-2
Granger, C. (1969). Investigating casual relations by econometric models and cross-spektral methods. Econometrica 37, 424–438. doi: 10.2307/1912791
Grech, R., Cassar, T., Muscat, J., Camilleri, K. P., Fabri, S. G., Zervakis, M., et al. (2008). Review on solving the inverse problem in EEG source analysis. J. Neuroeng. Rehabil. 5:25. doi: 10.1186/1743-0003-5-25
Grillner, S., Wallén, P., Saitoh, K., Kozlov, A., and Robertson, B. (2008). Neural bases of goal-directed locomotion in vertebrates—an overview. Brain Res. Rev. 57, 2–12. doi: 10.1016/j.brainresrev.2007.06.027
Gutberlet, I., Debener, S., Jung, T., and Makeig, S. (2009). “Techniques of EEG recording and preprocessing,” in Quantative EEG Analysis Methods and Clinical Applications, eds S. Tong and N. V. Thakor (Norwood, MA: ARTECH HOUSE), 23–49.
Gwin, J. T., Gramann, K., Makeig, S., and Ferris, D. P. (2010). Removal of movement artifact from high-density EEG recorded during walking and running. J. Neurophysiol. 103, 3526–3534. doi: 10.1152/jn.00105.2010
Hairston, W. D., Whitaker, K. W., Ries, A. J., Vettel, J. M., Bradford, J. C., Kerick, S. E., et al. (2014). Usability of four commercially-oriented EEG systems. J. Neural Eng. 11:046018. doi: 10.1088/1741-2560/11/4/046018
Haresign, I. M., Phillips, E., Whitehorn, M., Noreika, V., Jones, E., Leong, V., et al. (2021). Automatic classification of ICA components from infant EEG using MARA. bioRxiv [Preprint]. doi: 10.1101/2021.01.22.427809
Haufe, S., Dähne, S., and Nikulin, V. V. (2014). Dimensionality reduction for the analysis of brain oscillations. NeuroImage 101, 583–597. doi: 10.1016/j.neuroimage.2014.06.073
He, B., Lian, J., and Li, G. (2001). High-resolution EEG: a new realistic geometry spline Laplacian estimation technique. Clin. Neurophysiol. 112, 845–852. doi: 10.1016/s1388-2457(00)00546-0
Hill, R. M., Boto, E., Rea, M., Holmes, N., Leggett, J., Coles, L. A., et al. (2020). Multi-channel whole-head OPM-MEG: helmet design and a comparison with a conventional system. NeuroImage 219:116995. doi: 10.1016/j.neuroimage.2020.116995
Hinrichs, H., Scholz, M., Baum, A. K., Kam, J. W., Knight, R. T., and Heinze, H.-J. (2020). Comparison between a wireless dry electrode EEG system with a conventional wired wet electrode EEG system for clinical applications. Sci. Rep. 10:5218. doi: 10.1038/s41598-020-62154-0
Hsu, S.-H., Pion-Tonachini, L., Palmer, J., Miyakoshi, M., Makeig, S., and Jung, T.-P. (2018). Modeling brain dynamic state changes with adaptive mixture independent component analysis. NeuroImage 183, 47–61. doi: 10.1016/j.neuroimage.2018.08.001
Hyvärinen, A., and Oja, E. (2000). Independent component analysis: algorithms and applications. Neural Netw. 13, 411–430. doi: 10.1016/s0893-6080(00)00026-5
Izdebski, K., Oliveira, A. S., Schlink, B. R., Legkov, P., Kärcher, S., Hairston, W. D., et al. (2016). “Usability of EEG systems: user experience study,” in Proceedings of the 9th ACM International Conference on PErvasive Technologies Related to Assistive Environments (Corfu Island, Greece: ACM), 34.
Jensen, B. R., Hovgaard-Hansen, L., and Cappelen, K. L. (2016). Muscle activation and estimated relative joint force during running with weight support on a lower-body positive-pressure treadmill. J. Appl. Biomech. 32, 335–341. doi: 10.1123/jab.2015-0075
Jiang, X., Bian, G.-B., and Tian, Z. (2019). Removal of artifacts from EEG signals: a review. Sensors 19:987. doi: 10.3390/s19050987
Jung, T.-P., Humphries, C., Lee, T.-W., Makeig, S., McKeown, M. J., Iragui, V., et al. (1998). Extended ICA removes artifacts from electroencephalographic recordings. Adv. Neural Inform. Process. Systems, 894–900.
Jung, T. P., Makeig, S., Westerfield, M., Townsend, J., Courchesne, E., and Sejnowski, T. J. (2001). Analysis and visualization of single-trial event-related potentials. Hum. Brain Mapp. 14, 166–185. doi: 10.1002/hbm.1050
Kübler, A., Nijboer, F., Mellinger, J., Vaughan, T. M., Pawelzik, H., Schalk, G., et al. (2005). Patients with ALS can use sensorimotor rhythms to operate a brain-computer interface. Neurology 64, 1775–1777. doi: 10.1212/01.WNL.0000158616.43002.6D
Kaplan, A. Y., Fingelkurts, A. A., Fingelkurts, A. A., Ivashko, R., and Darkhovsky, B. (1997). Topological mapping of sharp reorganization synchrony in multichannel EEG. Am. J. Electroneurodiagnostic Technol. 37, 265–275. doi: 10.1080/1086508X.1997.11079198
Kappenman, E. S., and Luck, S. J. (2010). The effects of electrode impedance on data quality and statistical significance in ERP recordings. Psychophysiology 47, 888–904. doi: 10.1111/j.1469-8986.2010.01009.x
Keil, A., Debener, S., Gratton, G., Junghöfer, M., Kappenman, E. S., Luck, S. J., et al. (2014). Committee report: publication guidelines and recommendations for studies using electroencephalography and magnetoencephalography. Psychophysiology 51, 1–21. doi: 10.1111/psyp.12147
Kilicarslan, A., Grossman, R. G., and Contreras-Vidal, J. L. (2016). A robust adaptive denoising framework for real-time artifact removal in scalp EEG measurements. J. Neural Eng. 13:026013. doi: 10.1088/1741-2560/13/2/026013
Kim, D.-H., Lu, N., Ma, R., Kim, Y.-S., Kim, R.-H., Wang, S., et al. (2011). Epidermal electronics. Science 333, 838–843. doi: 10.1126/science.1206157
Kline, J. E., Huang, H. J., Snyder, K. L., and Ferris, D. P. (2015). Isolating gait-related movement artifacts in electroencephalography during human walking. J. Neural Eng. 12:046022. doi: 10.1088/1741-2560/12/4/046022
Knaepen, K., Beyl, P., Duerinck, S., Hagman, F., Lefeber, D., and Meeusen, R. (2014). Human-robot interaction: kinematics and muscle activity inside a powered compliant knee exoskeleton. IEEE Trans. Neural Syst. Rehabil. Eng. 22, 1128–1137. doi: 10.1109/TNSRE.2014.2324153
Koessler, L., Cecchin, T., Ternisien, E., and Maillard, L. (2010). “3D handheld laser scanner based approach for automatic identification and localization of EEG sensors,” in Proceedings of the 2010 Annual International Conference of the IEEE Engineering in Medicine and Biology (Buenos Aires, Argentina: IEEE), 3707–3710.
Kothe, C. A. E., and Jung, T.-P. (2016). Artifact Removal Techniques with Signal Reconstruction. Google Patents. United States Patent Office (2016) US Patent Application No. 14.895,440.
Löfhede, J., Seoane, F., and Thordstein, M. (2010). “Soft textile electrodes for EEG monitoring,” in Proceedings of the 10th IEEE International Conference on Information Technology and Applications in Biomedicine (Corfu, Greece: IEEE), 1–4.
Löfhede, J., Seoane, F., and Thordstein, M. (2012). Textile electrodes for EEG recording—a pilot study. Sensors 12, 16907–16919. doi: 10.3390/s121216907
Laszlo, S., Ruiz-Blondet, M., Khalifian, N., Chu, F., and Jin, Z. (2014). A direct comparison of active and passive amplification electrodes in the same amplifier system. J. Neurosci. Methods 235, 298–307. doi: 10.1016/j.jneumeth.2014.05.012
Lebedev, M. A., and Nicolelis, M. A. (2017). Brain-machine interfaces: from basic science to neuroprostheses and neurorehabilitation. Physiol. Rev. 97, 767–837. doi: 10.1152/physrev.00027.2016
Leske, S., and Dalal, S. S. (2019). Reducing power line noise in EEG and MEG data via spectrum interpolation. NeuroImage 189, 763–776. doi: 10.1016/j.neuroimage.2019.01.026
Li, Y., Ma, Z., Lu, W., and Li, Y. (2006). Automatic removal of the eye blink artifact from EEG using an ICA-based template matching approach. Physiol. Meas. 27, 425–436. doi: 10.1088/0967-3334/27/4/008
Li, J., Thakor, N., and Bezerianos, A. (2018). Unilateral exoskeleton imposes significantly different hemispherical effect in parietooccipital region, but not in other regions. Sci. Rep. 8:13470. doi: 10.1038/s41598-018-31828-1
Liu, H., Schimpf, P. H., Dong, G., Gao, X., Yang, F., and Gao, S. (2005). Standardized shrinking LORETA-FOCUSS (SSLOFO): a new algorithm for spatio-temporal EEG source reconstruction. IEEE Trans. Biomed. Eng. 52, 1681–1691. doi: 10.1109/TBME.2005.855720
Lopez-Gordo, M. A., Sanchez-Morillo, D., and Valle, F. P. (2014). Dry EEG electrodes. Sensors 14, 12847–12870. doi: 10.3390/s140712847
Luu, T. P., Brantley, J. A., Nakagome, S., Zhu, F., and Contreras-Vidal, J. L. (2017a). Electrocortical correlates of human level-ground, slope, and stair walking. PLoS One 12:e0188500. doi: 10.1371/journal.pone.0188500
Luu, T. P., Nakagome, S., He, Y., and Contreras-Vidal, J. L. (2017b). Real-time EEG-based brain-computer interface to a virtual avatar enhances cortical involvement in human treadmill walking. Sci. Rep. 7:8895. doi: 10.1038/s41598-017-09187-0
Luu, T. P., He, Y., Brown, S., Nakagome, S., and Contreras-Vidal, J. L. (2016). Gait adaptation to visual kinematic perturbations using a real-time closed-loop brain-computer interface to a virtual reality avatar. J. Neural Eng. 13:036006. doi: 10.1088/1741-2560/13/3/036006
Müller-Gerking, J., Pfurtscheller, G., and Flyvbjerg, H. (1999). Designing optimal spatial filters for single-trial EEG classification in a movement task. Clin. Neurophysiol. 110, 787–798. doi: 10.1016/s1388-2457(98)00038-8
Mahmood, M., Kwon, S., Kim, Y.-S., Siriaraya, P., Choi, J., Boris, O., et al. (2021). Wireless soft scalp electronics and virtual reality system for motor imagery-based brain-machine interfaces. Adv. Sci. doi: 10.1002/advs.202101129 [Epub ahead of print].
Makeig, S., Jung, T.-P., Bell, A. J., Ghahremani, D., and Sejnowski, T. J. (1997). Blind separation of auditory event-related brain responses into independent components. Proc. Natl. Acad. Sci. U S A 94, 10979–10984. doi: 10.1073/pnas.94.20.10979
Marlin, A., Mochizuki, G., Staines, W. R., and McIlroy, W. E. (2014). Localizing evoked cortical activity associated with balance reactions: does the anterior cingulate play a role? J. Neurophysiol. 111, 2634–2643. doi: 10.1152/jn.00511.2013
Massion, J., Ioffe, M., Schmitz, C., Viallet, F., and Gantcheva, R. (1999). Acquisition of anticipatory postural adjustments in a bimanual load-lifting task: normal and pathological aspects. Exp. Brain Res. 128, 229–235. doi: 10.1007/s002210050842
Mathewson, K. E., Harrison, T. J., and Kizuk, S. A. (2017). High and dry? Comparing active dry EEG electrodes to active and passive wet electrodes. Psychophysiology 54, 74–82. doi: 10.1111/psyp.12536
Miga, M. I., Kerner, T. E., and Darcey, T. M. (2002). Source localization using a current-density minimization approach. IEEE Trans. Biomed. Eng. 49, 743–745. doi: 10.1109/TBME.2002.1010860
Millán, J. R., Renkens, F., Mouriño, J., and Gerstner, W. (2004). Noninvasive brain-actuated control of a mobile robot by human EEG. IEEE Trans. Biomed. Eng. 51, 1026–1033. doi: 10.1109/TBME.2004.827086
Mognon, A., Jovicich, J., Bruzzone, L., and Buiatti, M. (2011). ADJUST: an automatic EEG artifact detector based on the joint use of spatial and temporal features. Psychophysiology 48, 229–240. doi: 10.1111/j.1469-8986.2010.01061.x
Mota, A. R., Duarte, L., Rodrigues, D., Martins, A., Machado, A., Vaz, F., et al. (2013). Development of a quasi-dry electrode for EEG recording. Sensors and Actuators A: Physical 199, 310–317. doi: 10.1016/j.sna.2013.06.013
Muthukumaraswamy, S. (2013). High-frequency brain activity and muscle artifacts in MEG/EEG: a review and recommendations. Front. Hum. Neurosci. 7:138. doi: 10.3389/fnhum.2013.00138
Narasimhan, S., and Dutt, D. N. (1996). Application of LMS adaptive predictive filtering for muscle artifact (noise) cancellation from EEG signals. Comput. Electrical Eng. 22, 13–30.
Nathan, K., and Contreras-Vidal, J. L. (2016). Negligible motion artifacts in scalp electroencephalography (EEG) during treadmill walking. Front. Hum. Neurosci. 9:708. doi: 10.3389/fnhum.2015.00708
Nidal, K., and Malik, A. S. Eds. (2014). EEG/ERP Analysis: Methods and Applications. New York, NY: CRC Press.
Nikulin, V. V., Nolte, G., and Curio, G. (2011). A novel method for reliable and fast extraction of neuronal EEG/MEG oscillations on the basis of spatio-spectral decomposition. NeuroImage 55, 1528–1535. doi: 10.1016/j.neuroimage.2011.01.057
Noda, T., Sugimoto, N., Furukawa, J., Sato, M.-A., Hyon, S.-H., and Morimoto, J. (2012). “Brain-controlled exoskeleton robot for BMI rehabilitation,” in Proceedings of the 2012 12th IEEE-RAS International Conference on Humanoid Robots (Humanoids 2012) (Osaka, Japan: IEEE), 21–27.
Nolan, H., Whelan, R., and Reilly, R. B. (2010). FASTER: fully automated statistical thresholding for EEG artifact rejection. J. Neurosci. Methods 192, 152–162. doi: 10.1016/j.jneumeth.2010.07.015
Nordin, A. D., Hairston, W. D., and Ferris, D. P. (2018). Dual-electrode motion artifact cancellation for mobile electroencephalography. J. Neural Eng. 15:056024. doi: 10.1088/1741-2552/aad7d7
Nordin, A. D., Hairston, W. D., and Ferris, D. P. (2019a). Faster gait speeds reduce alpha and beta EEG spectral power from human sensorimotor cortex. IEEE Trans. Biomed. Eng. 67, 842–853. doi: 10.1109/TBME.2019.2921766
Nordin, A. D., Hairston, W. D., and Ferris, D. P. (2019b). “Faster gait speeds suppress human auditory electrocortical responses,” in Proceedings of the 2019 IEEE International Conference on Systems, Man and Cybernetics (SMC) (IEEE), 235–240.
Nordin, A. D., Hairston, W. D., and Ferris, D. P. (2019c). Human electrocortical dynamics while stepping over obstacles. Sci. Rep. 9:4693. doi: 10.1038/s41598-019-41131-2
Oliveira, A. S., Schlink, B. R., Hairston, W. D., König, P., and Ferris, D. P. (2017a). A channel rejection method for attenuating motion-related artifacts in EEG recordings during walking. Front Neurosci. 11:225. doi: 10.3389/fnins.2017.00225
Oliveira, A. S., Schlink, B. R., Hairston, W. D., König, P., and Ferris, D. P. (2017b). Restricted vision increases sensorimotor cortex involvement in human walking. J. Neurophysiol. 118, 1943–1951. doi: 10.1152/jn.00926.2016
Oliveira, A. S., Schlink, B. R., Hairston, W. D., König, P., and Ferris, D. P. (2016). Proposing metrics for benchmarking novel EEG technologies towards real-world measurements. Front. Hum. Neurosci. 10:188. doi: 10.3389/fnhum.2016.00188
Oostenveld, R., Fries, P., Maris, E., and Schoffelen, J.-M. (2011). FieldTrip: open source software for advanced analysis of MEG, EEG and invasive electrophysiological data. Comput. Intell. Neurosci. 2011:156869. doi: 10.1155/2011/156869
Overton, D., and Shagass, C. (1969). Distribution of eye movement and eyeblink potentials over the scalp. Electroencephalogr. Clin. Neurophysiol. 27:546.
Palmer, J. A., Kreutz-Delgado, K., and Makeig, S. (2012). AMICA: an adaptive mixture of independent component analyzers with shared components. Swartz Center for Computatonal Neuroscience, University of California San Diego, Technical Report.
Park, H.-J., Jeong, D.-U., and Park, K.-S. (2002). Automated detection and elimination of periodic ECG artifacts in EEG using the energy interval histogram method. IEEE Trans. Biomed. Eng. 49, 1526–1533. doi: 10.1109/TBME.2002.805482
Patki, S., Grundlehner, B., Verwegen, A., Mitra, S., Xu, J., Matsumoto, A., et al. (2012). “Wireless EEG system with real time impedance monitoring and active electrodes,” in Proceedings of the 2012 IEEE Biomedical Circuits and Systems Conference (BioCAS) (Hsinchu, Taiwan: IEEE), 108–111.
Pedroni, A., Bahreini, A., and Langer, N. (2019). Automagic: standardized preprocessing of big EEG data. NeuroImage 200, 460–473. doi: 10.1016/j.neuroimage.2019.06.046
Peterson, S. M., and Ferris, D. P. (2018). Differentiation in theta and beta electrocortical activity between visual and physical perturbations to walking and standing balance. eNeuro 5:ENEURO.0207–18.2018. doi: 10.1523/ENEURO.0207-18.2018
Peterson, S. M., and Ferris, D. P. (2019). Group-level cortical and muscular connectivity during perturbations to walking and standing balance. NeuroImage 198, 93–103. doi: 10.1016/j.neuroimage.2019.05.038
Pion-Tonachini, L., Kreutz-Delgado, K., and Makeig, S. (2019). ICLabel: an automated electroencephalographic independent component classifier, dataset and website. NeuroImage 198, 181–197. doi: 10.1016/j.neuroimage.2019.05.026
Raghavendra, B., and Dutt, D. N. (2011). Wavelet enhanced CCA for minimization of ocular and muscle artifacts in EEG. World Acad. Sci. Eng. Technol. 57, 1027–1032. doi: 10.5281/zenodo.1075224
Ramoser, H., Muller-Gerking, J., and Pfurtscheller, G. (2000). Optimal spatial filtering of single trial EEG during imagined hand movement. IEEE Trans. Rehabil. Eng. 8, 441–446. doi: 10.1109/86.895946
Richer, N., Downey, R. J., Hairston, W. D., Ferris, D. P., and Nordin, A. D. (2020). Motion and muscle artifact removal validation using an electrical head phantom, robotic motion platform and dual layer mobile EEG. IEEE Trans. Neural Syst. Rehabil. Eng. 28, 1825–1835. doi: 10.1109/TNSRE.2020.3000971
Richter, C. G., Thompson, W. H., Bosman, C. A., and Fries, P. (2015). A jackknife approach to quantifying single-trial correlation between covariance-based metrics undefined on a single-trial basis. NeuroImage 114, 57–70. doi: 10.1016/j.neuroimage.2015.04.040
Robert, C., Gaudy, J.-F., and Limoge, A. (2002). Electroencephalogram processing using neural networks. Clin. Neurophysiol. 113, 694–701. doi: 10.1016/s1388-2457(02)00033-0
Rossi, A., Parada, F. J., Kolchinsky, A., and Puce, A. (2014). Neural correlates of apparent motion perception of impoverished facial stimuli: a comparison of ERP and ERSP activity. NeuroImage 98, 442–459. doi: 10.1016/j.neuroimage.2014.04.029
Sakkalis, V. (2011). Review of advanced techniques for the estimation of brain connectivity measured with EEG/MEG. Comput. Biol. Med. 41, 1110–1117. doi: 10.1016/j.compbiomed.2011.06.020
Sanei, S., and Chambers, J. A. (2013). EEG Signal Processing. Chichester, England; Hoboken, NJ: John Wiley and Sons.
Scanlon, J. E., Jacobsen, N. S. J., Maack, M. C., and Debener, S. (2020). Does the electrode amplification style matter? a comparison of active and passive EEG system configurations during standing and walking. Eur. J. Neurosci. doi: 10.1111/ejn.15037 [Online ahead of Print].
Schimpf, P. H., Liu, H., Ramon, C., and Haueisen, J. (2005). Efficient electromagnetic source imaging with adaptive standardized LORETA/FOCUSS. IEEE Trans. Biomed. Eng. 52, 901–908. doi: 10.1109/TBME.2005.845365
Schlögl, A., Keinrath, C., Zimmermann, D., Scherer, R., Leeb, R., and Pfurtscheller, G. (2007). A fully automated correction method of EOG artifacts in EEG recordings. Clin. Neurophysiol. 118, 98–104. doi: 10.1016/j.clinph.2006.09.003
Sczesny-Kaiser, M., Höffken, O., Aach, M., Cruciger, O., Grasmücke, D., Meindl, R., et al. (2015). HA® exoskeleton training improves walking parameters and normalizes cortical excitability in primary somatosensory cortex in spinal cord injury patients. J. Neuroeng. Rehabil. 12:68. doi: 10.1186/s12984-015-0058-9
Seeber, M., Scherer, R., Wagner, J., Solis-Escalante, T., and Müller-Putz, G. R. (2014). EEG beta suppression and low gamma modulation are different elements of human upright walking. Front. Hum. Neurosci. 8:485. doi: 10.3389/fnhum.2014.00485
Shackman, A. J., McMenamin, B. W., Slagter, H. A., Maxwell, J. S., Greischar, L. L., and Davidson, R. J. (2009). Electromyogenic artifacts and electroencephalographic inferences. Brain Topogr. 22, 7–12. doi: 10.1007/s10548-009-0079-4
Shirazi, S. Y., and Huang, H. J. (2019). More reliable EEG electrode digitizing methods can reduce source estimation uncertainty, but current methods already accurately identify brodmann areas. Front. Neurosci. 13:1159. doi: 10.3389/fnins.2019.01159
Shustak, S., Inzelberg, L., Steinberg, S., Rand, D., Pur, M. D., Hillel, I., et al. (2019). Home monitoring of sleep with a temporary-tattoo EEG, EOG and EMG electrode array: a feasibility study. J. Neural Eng. 16:026024. doi: 10.1088/1741-2552/aafa05
Sipp, A. R., Gwin, J. T., Makeig, S., and Ferris, D. P. (2013). Loss of balance during balance beam walking elicits a multifocal theta band electrocortical response. J. Neurophysiol. 110, 2050–2060. doi: 10.1152/jn.00744.2012
Slobounov, S., Cao, C., Jaiswal, N., and Newell, K. M. (2009). Neural basis of postural instability identified by VTC and EEG. Exp. Brain Res. 199, 1–16. doi: 10.1007/s00221-009-1956-5
Snijders, A. H., Van De Warrenburg, B. P., Giladi, N., and Bloem, B. R. (2007). Neurological gait disorders in elderly people: clinical approach and classification. The Lancet Neurol. 6, 63–74. doi: 10.1016/S1474-4422(06)70678-0
Snyder, K. L., Kline, J. E., Huang, H. J., and Ferris, D. P. (2015). Independent component analysis of gait-related movement artifact recorded using EEG electrodes during treadmill walking. Front. Hum. Neurosci. 9:639. doi: 10.3389/fnhum.2015.00639
Someya, T., and Amagai, M. (2019). Toward a new generation of smart skins. Nat. Biotechnol. 37, 382–388. doi: 10.1038/s41587-019-0079-1
Stephenson, W., and Gibbs, F. A. (1951). A balanced non-cephalic reference electrode. Electroencephalogr. Clin. Neurophysiol. 3, 237–240. doi: 10.1016/0013-4694(51)90017-x
Symeonidou, E.-R., Nordin, A. D., Hairston, W. D., and Ferris, D. P. (2018). Effects of cable sway, electrode surface area and electrode mass on electroencephalography signal quality during motion. Sensors 18:1073. doi: 10.3390/s18041073
Taheri, B. A., Knight, R. T., and Smith, R. L. (1994). A dry electrode for EEG recording. Electroencephalogr. Clin. Neurophysiol. 90, 376–383. doi: 10.1016/0013-4694(94)90053-1
Tallon-Baudry, C., Bertrand, O., Delpuech, C., and Pernier, J. (1996). Stimulus specificity of phase-locked and non-phase-locked 40 Hz visual responses in human. J. Neurosci. 16, 4240–4249. doi: 10.1523/JNEUROSCI.16-13-04240.1996
Tariq, M., Trivailo, P. M., and Simic, M. (2018). EEG-based BCI control schemes for lower-limb assistive-robots. Front. Hum. Neurosci. 12:312. doi: 10.3389/fnhum.2018.00312
Teplan, M. (2002). Fundamentals of EEG measurement. Measurement Sci. Rev. 2, 1–11. Available online at: http://www.measurement.sk/2002/S2/Teplan.pdf
Thatcher, R., Krause, P., and Hrybyk, M. (1986). Cortico-cortical associations and EEG coherence: a two-compartmental model. Electroencephalogr. Clin. Neurophysiol. 64, 123–143. doi: 10.1016/0013-4694(86)90107-0
Tierney, T. M., Mellor, S., O’Neill, G. C., Holmes, N., Boto, E., Roberts, G., et al. (2020). Pragmatic spatial sampling for wearable MEG arrays. Sci. Rep. 10:21609. doi: 10.1038/s41598-020-77589-8
Ullsperger, M., and Debener, S. (2010). Simultaneous EEG and fMRI: Recording, Analysis and Application. New York; Oxford: Oxford University Press.
Urigüen, J. A., and Garcia-Zapirain, B. (2015). EEG artifact removal—state-of-the-art and guidelines. J. Neural Eng. 12:031001. doi: 10.1088/1741-2560/12/3/031001
Valdes-Sosa, P., Marti, F., Garcia, F., and Casanova, R. (2000). “Variable resolution electric-magnetic tomography,” in Biomag 96, eds C. J. Aine, G. Stroink, C. C. Wood, Y. Okada and S. J. Swithenby (New York, NY: Springer), 373–376. doi: 10.1007/978-1-4612-1260-7_91
Van Hoey, G., De Clercq, J., Vanrumste, B., Van de Walle, R., Lemahieu, I., D’Havé, M., et al. (2000). EEG dipole source localization using artificial neural networks. Phys. Med. Biol. 45, 997–1011. doi: 10.1088/0031-9155/45/4/314
Vanacker, G., Millán, J. D. R., Lew, E., Ferrez, P. W., Moles, F. G., Philips, J., et al. (2007). Context-based filtering for assisted brain-actuated wheelchair driving. Comput. Intell. Neurosci. 2007:25130. doi: 10.1155/2007/25130
Varghese, J. P., Staines, W. R., and McIlroy, W. E. (2019). Activity in functional cortical networks temporally associated with postural instability. Neuroscience 401, 43–58. doi: 10.1016/j.neuroscience.2019.01.008
Vorwerk, J., Oostenveld, R., Piastra, M. C., Magyari, L., and Wolters, C. H. (2018). The FieldTrip-SimBio pipeline for EEG forward solutions. Biomed. Eng. Online 17:37. doi: 10.1186/s12938-018-0463-y
Wagner, J., Makeig, S., Gola, M., Neuper, C., and Müller-Putz, G. (2016). Distinct β band oscillatory networks subserving motor and cognitive control during gait adaptation. J. Neurosci. 36, 2212–2226. doi: 10.1523/JNEUROSCI.3543-15.2016
Wagner, J., Solis-Escalante, T., Grieshofer, P., Neuper, C., Müller-Putz, G., and Scherer, R. (2012). Level of participation in robotic-assisted treadmill walking modulates midline sensorimotor EEG rhythms in able-bodied subjects. NeuroImage 63, 1203–1211. doi: 10.1016/j.neuroimage.2012.08.019
Wang, L.-F., Liu, J.-Q., Yang, B., and Yang, C.-S. (2012). PDMS-based low cost flexible dry electrode for long-term EEG measurement. IEEE Sensors J., 12, 2898–2904. doi: 10.1109/JSEN.2012.2204339
Whitham, E. M., Pope, K. J., Fitzgibbon, S. P., Lewis, T., Clark, C. R., Loveless, S., et al. (2007). Scalp electrical recording during paralysis: quantitative evidence that EEG frequencies above 20 Hz are contaminated by EMG. Clin. Neurophysiol. 118, 1877–1888. doi: 10.1016/j.clinph.2007.04.027
Winkler, I., Debener, S., Müller, K.-R., and Tangermann, M. (2015). On the influence of high-pass filtering on ICA-based artifact reduction in EEG-ERP. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2015, 4101–4105. doi: 10.1109/EMBC.2015.7319296
Wolpaw, J. R., and McFarland, D. J. (2004). Control of a two-dimensional movement signal by a noninvasive brain-computer interface in humans. Proc. Natl. Acad. Sci. U S A 101, 17849–17854. doi: 10.1073/pnas.0403504101
Xu, J., Mitra, S., Van Hoof, C., Yazicioglu, R. F., and Makinwa, K. A. (2017). Active electrodes for wearable EEG acquisition: review and electronics design methodology. IEEE Rev. Biomed. Eng. 10, 187–198. doi: 10.1109/RBME.2017.2656388
Xu, X.-L., Xu, B., and He, B. (2004). An alternative subspace approach to EEG dipole source localization. Phys. Med. Biol. 49, 327–343. doi: 10.1088/0031-9155/49/2/010
Xu, J., and Zhong, B. (2018). Review on portable EEG technology in educational research. Comput. Hum. Behav. 81, 340–349. doi: 10.1016/j.chb.2017.12.037
Keywords: EEG signal processing, motor neuroscience, neuroimaging, locomotion, mobile EEG, electroencephalography (EEG), EEG hardware
Citation: Song S and Nordin AD (2021) Mobile Electroencephalography for Studying Neural Control of Human Locomotion. Front. Hum. Neurosci. 15:749017. doi: 10.3389/fnhum.2021.749017
Received: 28 July 2021; Accepted: 05 October 2021;
Published: 10 November 2021.
Edited by:
Ioannis Ugo Isaias, Julius Maximilian University of Würzburg, GermanyReviewed by:
Teodorico Caporaso, University of Naples Federico II, ItalyCopyright © 2021 Song and Nordin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrew D. Nordin, bm9yZGluYUB0YW11LmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.