 Anthony G. Vaccaro
Anthony G. Vaccaro Brandon Scott1,2
Brandon Scott1,2 Sarah I. Gimbel
Sarah I. Gimbel Jonas T. Kaplan
Jonas T. Kaplan
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Hum. Neurosci. , 05 July 2021
Sec. Cognitive Neuroscience
Volume 15 - 2021 | https://doi.org/10.3389/fnhum.2021.665319
This article is part of the Research Topic Examining the neurodynamics of emotional experiences with naturalistic paradigms View all 6 articles
Engaging with narratives involves a complex array of cognitive and affective processes. These processes make stories persuasive in ways that standard arguments are not, though the underlying reasons for this remain unclear. Transportation theory proposes a potential explanation for this: narratives are processed in a way which makes individuals feel immersed in the world of a story, which in turn leads people to resonate emotionally with the events of the story. Recent fMRI studies have shown that the posterior medial cortex (PMC) and anterior insula (AI) play important roles in understanding the meaning of stories and experiencing the feelings they produce. In this study, we aimed to explore the AI’s and PMC’s role in narrative processing by measuring their functional connectivity with the rest of the brain during story listening, and how connectivity changes as a function of narrative transportation and the persuasiveness of the story. We analyzed data from 36 right-handed subjects who listened to two stories, obtained from podcasts, inside the fMRI scanner. After the scan, subjects were asked a series of questions, including a measure of how transported into the story they felt, how likely they would be to donate to causes related to the messages of the stories. We used searchlight multivariate pattern analysis (MVPA) to classify functional connectivity maps using seeds in both the AI and PMC and to compare these maps between participants who differed in transportation and prosocial intention. We found that connectivity to various regions successfully distinguished between high and low ratings on each of these behavioral measures with accuracies over 75%. However, only one pattern of connectivity was consistent across both stories: PMC-inferior frontal gyrus connectivity successfully distinguished high and low ratings of narrative transportation in both stories. All other findings were not consistent across stories. Instead, we found that patterns of connectivity may relate more to the specific content of the story rather than to a universal way in which narratives are processed.
Recent research suggests that messages in narrative form may be more persuasive compared with standard arguments (Deighton et al., 1989; Slater and Rouner, 2002; Bilandzic and Busselle, 2013). The underlying reasons for the persuasiveness of stories remain unclear, but likely depend upon the specific cognitive and affective mechanisms involved in story comprehension. Whereas processing a standard argument requires focusing on a sequence of facts and arguments, processing stories requires attention to characters, their intentions, and sequences of events. Furthermore, stories often evoke affect in ways standard arguments do not. One prominent hypothesis as to why stories are generally more persuasive in their messages is transportation theory (Green and Brock, 2000; Green and Sestir, 2017). Transportation theory posits that the way narratives are processed can make individuals feel as if they immersed in the world of a story (Green et al., 2004; Batat and Wohlfeil, 2009; Van Laer et al., 2014). Being immersed then causes people to react emotionally in a way that is similar to how they react to real-life events. This is thought to make the listener/reader more likely to be persuaded of the story’s message (Escalas, 2004; Green et al., 2004; Banerjee and Greene, 2012; Green and Sestir, 2017; Morris et al., 2019). Narrative transportation can facilitate persuasion where individuals’ beliefs are typically resistant to change, such as in regard to health-related behaviors (Banerjee and Greene, 2012; Green and Clark, 2013; Dillard et al., 2018), intergroup prejudice (Mazzocco et al., 2010; Caputo and Rouner, 2011; Johnson et al., 2013; Guerrero and Igartua, 2017; Moyer-Gusé et al., 2019) and prosocial decision-making (Harjusola-Webb et al., 2012; Johnson, 2012; Steinemann et al., 2017).
Narrative transportation is influenced by individual state and trait differences in affect. For example, when individuals are in an affective state that matches the emotional tone of the story they are about to experience, the reported feeling of transportation increases (Green et al., 2012). On the trait-level, individuals who actively seek out emotion-inducing situations are more likely to feel transported into a narrative and more likely to be persuaded by the narrative (Appel and Richter, 2010). Trait empathy also correlates with the tendency to be transported into narratives: transported readers experience more empathy toward characters they read about (Green and Donahue, 2009; Hall and Bracken, 2011; Weiss, 2015).
Individuals who feel transported into a story identify more with the story’s characters, displaying more of their traits and emotions (Green et al., 2004; Johnson et al., 2014; Christy and Fox, 2016; Igartua and Frutos, 2017). They also report temporary differences in their self-concept which align more with the characters (Sestir and Green, 2010; Breen et al., 2017). Some highly transportable people may be motivated to engage with narrative media in order to temporarily redefine their self-concept as more alike to a narrative world’s character (Hirschman, 1983; Greenwood and Long, 2009; Klimmt et al., 2009; Li et al., 2011; Slater et al., 2014; Christy and Fox, 2016; Kuo et al., 2016). Narratives may also trigger autobiographical memories, which are known to play an important role in defining sense of self (Dunlop et al., 2010; Prebble et al., 2013; Rasmussen et al., 2014; McDonald et al., 2015) and are organized in narrative form (Robinson and Taylor, 1998; Conway et al., 2004; Singer and Blagov, 2004; Fivush, 2011; Prebble et al., 2013). For this reason, processing the structure of narratives in media may be an effective way to prime specific autobiographical memories. This would then affect one’s current sense of self by highlighting specific feelings related to that memory (Baumgartner et al., 1992; Moore and Homer, 2008; McDonald et al., 2015; Hartmann et al., 2016).
During the last few years there has been an increase in fMRI studies focusing on narratives. Many of these studies have highlighted an important role for the default mode network (DMN), and in particular, the posterior medial cortex (PMC) (Fransson and Marrelec, 2008). PMC regions (which include the mesial precuneus, posterior cingulate cortex, and retrosplenial areas) are known to be important for self-related processes and self-concept (Seger et al., 2004; Parvizi et al., 2006; Summerfield et al., 2009; Rameson et al., 2010; Araujo et al., 2015; Feng et al., 2018). Given the PMC’s role in autobiographical self-related processes, it is no surprise that these regions would also be relevant to narrative processing. The DMN, including the PMC, appears to be involved in the process of making sense out of long form event sequences (Lerner et al., 2011; Baldassano et al., 2017, 2018). Inter-subject correlation analyses have found that the specific meanings of narratives can be decoded across subjects by the spontaneous correlation patterns within BOLD activity in the DMN and PMC regions (Hasson et al., 2008; Simony et al., 2016; Nguyen et al., 2019). Furthermore, searchlight multi-voxel pattern analysis can accurately predict shared meanings of stories presented to different subjects in their native languages, and even the higher-level moral values of these stories (Dehghani et al., 2017; Kaplan et al., 2017). Euclidian distance-based classification has shown that BOLD activity in PMC regions can predict people’s specific interpretations of stories as well, demonstrating how closely this area corresponds to the phenomenological experiences of narratives (Yeshurun et al., 2017).
Another region which may be relevant to the experience of narratives is the anterior insula. The anterior insula (AI) is known to integrate homeostatic information from the body and is foundational to the conscious subjective experience of emotions (Craig, 2009; Zaki et al., 2012; Gu et al., 2013). This integrative role has been shown to be operative while listening to narratives. In one such study, effective connectivity was used to show that the AI was integrating audio information from the narrative with heart rate variability information from the posterior insula, as well as displaying more activity in general during emotionally salient parts of a story (Nguyen et al., 2016). The insula’s role in integrating information for emotional experience may also lead its activity to reflect the perceived vividness of stories (Bruneau et al., 2013).
Importantly, AI activity has been found to correlate with differences in experiences between narrative consumers. AI activity correlates with various types of admiration for a narrative’s protagonist (Immordino-Yang et al., 2009), and AI activity has also been used to differentiate cultural influences on how narrative-induced feelings are formed. For instance, while the ventral AI (which modulates the autonomic nervous system) is associated with how admirable Chinese listeners felt the protagonists in a story were, the dorsal AI (which relates to somatosensory and visceral input) associated with how admirable Americans felt the characters were (Immordino-Yang et al., 2014). Furthermore, regardless of cultural background, the strength of the correlation between admiration and the dorsal AI was moderated by how expressive the person was in a previous interview about the narrative (Immordino-Yang et al., 2016). Culturally specific processes of feeling explain why stories written within a certain culture are more likely to influence transportability and be persuasive: the language of stories reflect these distinctions in how emotions are subjectively felt in different cultures (Stolte and Fender, 2007; Larkey and Hecht, 2010; Swanson et al., 2017).
Despite consistent findings implicating the PMC and AI in narrative processing, there has been little research into the functional connectivity of these regions while listening to narratives. The AI (Jabbi et al., 2008; Cauda et al., 2011) and PMC (Fransson and Marrelec, 2008; Zhang and Chiang-shan, 2012; Khalsa et al., 2014) function as nodes within neural networks rather than as isolated regions. By understanding how they function within broader neural networks while listening to stories, we can learn more about what cognitive and affective processes help facilitate the effects previously discussed, such as the feeling of transportation and the increased persuasive influence. Furthermore, functional connectivity analysis lends itself well to the study of complex, naturalistic stimuli such as stories.
Our first set of hypotheses concerned functional connectivity while listening to stories relating to behavioral characteristics of the individual listeners. We predicted that functional connectivity patterns would differ between those who were transported into the stories and those who weren’t. Likewise, we predicted that functional connectivity patterns would differ between subjects who were persuaded to support causes and those who weren’t.
We also hypothesized that AI and PMC connectivity to the broader DMN would be predictive of transportation and of the story-specific prosocial intentions (i.e., for a story involving cancer, the intention to donate to cancer research rather than to another cause). Increased connectivity between these two seeds and other DMN regions, such as the medial prefrontal cortex and angular gyrus, may reflect the emotional impact of the story having more influence on self-other boundaries, and thus reflect the persuasive influence of the story. Specifically, we chose to focus on the dorsal AI due to its unique role in modulating the DMN compared to other insular sub-regions (Nomi et al., 2016; Uddin et al., 2017), and its role integrating somatosensory and visceral input with other cognitive and affective functions, which we believe is particularly relevant to these stories (Deen et al., 2010; Uddin et al., 2014). Finally, we hypothesized that AI and PMC connectivity to sensorimotor areas would be additionally predictive of narrative transportation due to the individuals feeling as-if they are situated in the actions and sensations of the story.
We used searchlight-based multi-voxel pattern analysis (MVPA) to compare connectivity maps between groups of subjects. Because signal from many voxels are considered simultaneously, MVPA shows increased sensitivity compared with traditional univariate analysis (Kriegeskorte et al., 2006; Norman et al., 2006; Kriegeskorte and Bandettini, 2007). Our first set of hypotheses would be supported if our MVPA of the functional connectivity maps can accurately classify the behavioral characteristics we are measuring (high vs. low transportation and intention to support causes). The strongest outcome for the rest of our hypotheses would be if these patterns of connectivity were then consistent across both stories. A pattern of connectivity in one story but not the other may suggest a process specific to the story’s content rather than a general mechanism universal for stories.
All procedures were approved by the Institutional Review Board of University of Southern California.
Forty-five right-handed, native-English speaking subjects were recruited for this fMRI study from the Los Angeles area. However, after removing three subjects for excessive movement (movements >1.5 mm from one volume to the next), one for falling asleep during the scan, one for an incidental finding in the brain, as well as another four subjects for failing basic comprehension questions about the stories they listened to, 36 subjects remained (18 female, mean age: 22.30 ± 0.83 years) (see Table 1). Subjects had no history of neurological disorder or of psychiatric disorder (as measured by the Neuropsychiatric Inventory Questionnaire).
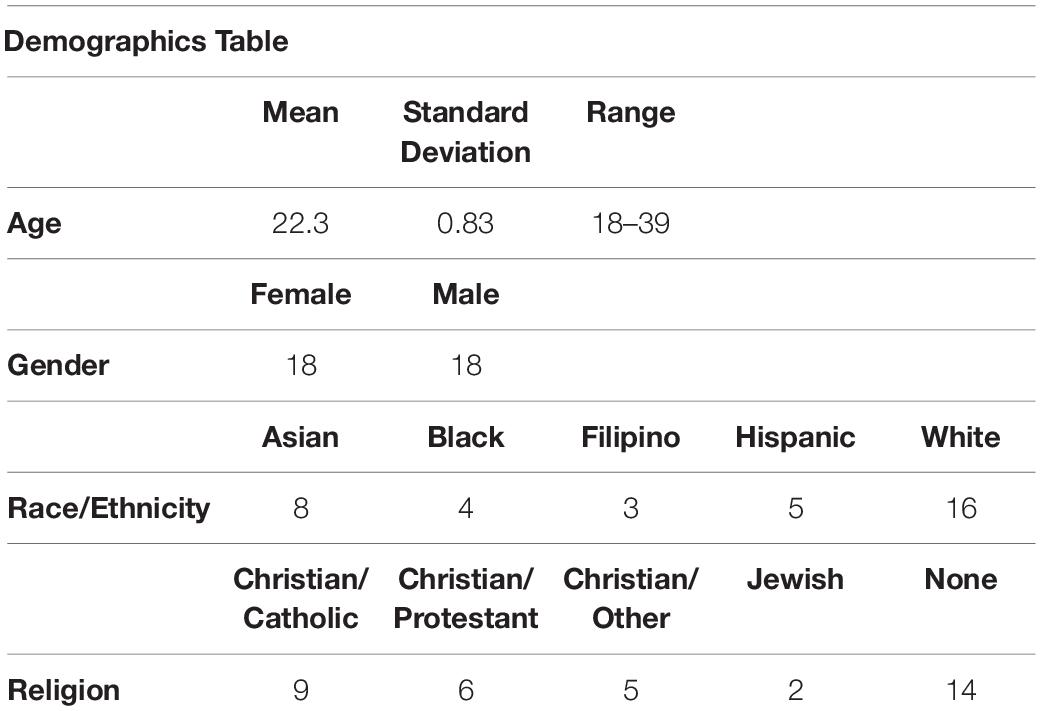
Table 1. Participant demographics.
Two stories were selected from popular podcasts. These two stories were “The Living Room” from Love + Radio podcast and “The Hitcher” from This American Life. In “The Living Room,” the protagonist describes watching a young couple over time through their open window. Eventually, one member of the couple gets cancer and the protagonist describes watching them decline over time, eventually succumbing to the disease. In “The Hitcher,” two young brothers hitchhike to get home one night. Along the way, the driver brings them to various locations and the brothers are unsure if they are being kidnapped before they eventually decide to escape. Each story was transcribed from the podcast and then read aloud by a professional voice actor. “The Living Room” was read by a female actor while “The Hitcher” was read by a male actor.
Participants were given instructions to listen to the stories with their eyes open and to lie still in the scanner. Each participant started with a resting state scan (not analyzed here), followed by the two stories, counterbalanced in order across the subjects. Each story started with 10 s of silence. The stories were “The Living Room” (11 min, 42 s), originally from Love + Radio podcast), and “The Hitcher” (10 min, 5 s), originally from This American Life). Participants listened to the stories through a set of fMRI-compatible in-ear headphones (model S14, Sensimetrics). The volume was calibrated for each subject to ensure they could hear the stories over the noise of the scanner. The participants’ eyes were monitored by the experimenter in real-time with an eye tracking camera to ensure wakefulness throughout the story scans.
After the scan, subjects answered comprehension questions to assure they were actively listening to the stories. Subjects also answered the questions of the Transportation Scale Short Form (TS-SF) relative to each story (Appel et al., 2015). Each item on this scale is rated from 1 (not at all) to 7 (very much) and averaged for a total score. Finally, subjects answered various questions about how willing they were to contribute to various causes on a scale from 0 to 10. Relevant to the current analyses, there was a question related to the theme of “The Living Room,” and a question related to the theme of “The Hitcher” as follows:
“How willing are you to donate $5 to a cancer research organization?”
“How willing are you sign a petition supporting the education of minors on the dangers of hitchhiking?”
A 3T Siemens MAGNETON Prisma system with a 32-channel head coil was used for the fMRI portion of the study (located at the Dana and David Dornsife Neuroscience Institute at University of Southern California). T1-weighted scans were obtained at 1 mm × 1 mm × 1 mm resolution using a 3D magnetization-prepared rapid acquisition gradient (MPRAGE) sequence. One-hundred seventy-six slices were obtained with a 10-degree flip angle, 256 × 256 matrix, and a phase encoding direction of right to left. TR was 2,300 ms and TE was 2.26 ms.
Functional images were obtained with a gradient-echo, echo-planar T2∗-weighted multiband pulse sequence. Forty-eight slices 3 mm × 3 mm × 3 mm resolution slices were obtained with a 90-degree flip angle, 64 × 64 matrix. TR was 1,000 ms and TE was 35 ms, in interleaved ascending order. For “The Living Room” 740 functional volumes were obtained and for “The Hitcher” 625 functional volumes were obtained. The gradient-echo field map was also obtained, to be used for later field-inhomogeneity correction, with a 90-degree flip angle and 64 × 64 matrix. TR was 1,000 ms, TE1 was 10 ms, and TE2 was 12.45 ms.
Pre-processing and seed-connectivity analyses were performed using FSL version 5.0.81. The gradient-echo field map was used to correct magnetic field inhomogeneity along with FSL’s FUGUE tool for unwarping EPI’s (anterior-posterior direction, 10% signal loss threshold). Motion-correction was performed with a rigid-body alignment to the middle volume of the scan as a point of reference. Slice-time correction was performed with Fourier-space time series phase shifting. Skull-stripping was performed using FSL’s BET brain extraction tool with the center voxel of the brain manually specified (fractional intensity threshold = 0.4). Spatial smoothing was performed with a 5 mm FWHM kernel. High-pass temporal filtering was applied with a Gaussian weighted least-squares line (sigma = 60 s) and temporal autocorrelation was accounted for with FSL’s pre-whitening algorithm. In order to remove the effects of the onset and offset of the audio stimulus, which could drive significant variability in the signal, we removed 16 volumes from the beginning (10 s of silence + 6 for hemodynamic delay) and four volumes from the end of each dataset.
Functional images were registered to their respective T1 images using FSL FLIRT’s boundary-based registration algorithm. Finally, the images were registered to a common MNI 152 space with 12 degree of freedom affine transformation and FNIRT registration (warp resolution = 10 mm).
Whole-brain functional connectivity maps for the AI and for the PMC were generated for each subject. For the AI seed we used a dorsal AI seed derived from the parcellations used in a study on functional insular subdivisions (Deen et al., 2010). The PMC seed was derived from a previous study in our group on narrative processing, defined as a cluster in the posterior medial region that was more activated by reading sacred stories compared with non-sacred stories (Kaplan et al., 2017). The time-series for each of these ROI’s were extracted for each subject. These extracted time-series were then used as regressors for a first level general linear model analysis in FSL’s FEAT. Anatomical brain images for each subject were parcellated using FAST to generate a mask of cerebrospinal fluid (CSF) and the timecourse of signal from CSF was included as a nuisance regressor in this analysis. The resulting functional connectivity maps indicated how correlated any given voxel’s time series was with the time series of the seed region. Individual unthresholded functional connectivity maps were then subjected to the cross-subject MVPA analysis. Additionally, we performed a group-level GLM to describe connectivity with the seed regions during story listening overall. This group GLM modeled only the group level mean and used a FLAME mixed-effects model to generate group level statistical maps in the standard space which were then thresholded using FSL’s cluster thresholding algorithm with a cluster-forming threshold of Z > 3.1 and a cluster size probability of p < 0.05.
Searchlight multivariate pattern analysis (MVPA) (Kriegeskorte et al., 2006) was performed on the seed-connectivity maps using difference behavioral measures as the classifying attribute. Analyses were performed using PyMVPA (Hanke et al., 2009). For each analysis, subjects were divided into “high” or “low” categories for a variable based on whether they were above or below the mean on said variable.
The seed-connectivity maps for each story were classified for high vs. low reported transportation as measured by the TS-SF. Each story’s seed connectivity map was also classified based on the story-relevant prosocial measure (i.e., for “The Hitcher” the self-reported willingness to sign the hitchhiking dangers education petition) and the story-irrelevant prosocial measure (i.e., for “The Hitcher” the self-reported willingness to donate to cancer research). The story irrelevant prosocial measure analyses were included to distinguish whether the patterns of connectivity were reflecting a process of story-specific influence vs. capturing a more generic process related to general prosocial tendencies.
In the searchlight analyses, support-vector machine (SVM) classifiers were trained on the 5 mm spherical area surrounding each voxel using a leave-one-subject-out cross-validation approach. Each voxel in the resulting searchlight map thus represents the overall accuracy of the classifier in distinguishing high vs. low scores on that variable based upon seed-connectivity to the immediate area surrounding that voxel. The resulting maps indicate how accurate the classifier was at predicting the behavioral category based on data from each spatial location.
To determine an appropriate statistical thresholding, permutation testing was performed by repeatedly shuffling the map labels inside a single sphere and classifying the shuffled data to produce a distribution of accuracies under the null hypothesis. We then computed the number of resolution elements (resels) in the image by dividing the total volume of the brain by the volume of one sphere to yield the number of non-overlapping spheres. An alpha level of 0.05 was divided by the number of resels to determine a corrected alpha (Kaplan and Meyer, 2012). From this process, a threshold emerged of 75% accuracy for significance, which is the corrected p-value derived from the permutation test.
The average score on the TS-SF relative to the cancer story (“The Living Room”) was 5.75 (SD = 1.07; ranging from 2.2 to 7.0) and relative to the hitchhiking story (“The Hitcher”) was 5.2 (SD = 1.28; ranging from 2.0 to 7.0). The transportation scores for the two stories were highly correlated within individuals (r = 0.511; p < 0.001), though subjects felt significantly more transported into the “Living Room” story compared to “The Hitcher” [t(36) = 2.82, p = 0.0078].
The average willingness to donate to cancer research after listening to the stories was 6.55 (SD = 3.04; ranging from 0.0–10.0) out of 10. The average willingness to sign a petition to educate kids on the dangers of hitchhiking was 6.55 (SD = 2.66; ranging from 0.1 to 10.0). These reported pro-social measures were significantly correlated within individuals (r = 0.417; p = 0.011); those who were more likely to support one measure were also more likely to support the other.
Narrative transportation in the cancer story was highly correlated with reported willingness to donate to cancer research (r = 0.425, p = 0.009). Narrative transportation in the cancer story was also correlated with willingness to sign the petition on the dangers of hitchhiking (r = 0.384, p = 0.02) (see Table 2). Narrative transportation in the hitchhiking story was not significantly correlated with either prosocial measure. This may be due to individuals feeling overall less transported into that story.

Table 2. Correlations between narrative transportation and pro-social intentions.
Results of the group GLM of seed region connectivity are show in Figure 1. AI showed significant connectivity with large regions of the medial surface of the brain, including most of the cingulate gyrus and parts of the medial prefrontal cortex, the cuneus, the inferior parietal lobes, the precentral gyrus, the middle and inferior frontal gyri, and the cerebellum. PMC showed significant connectivity with DMN nodes, including medial prefrontal cortex, inferior parietal cortex, much of the temporal lobes. For both seeds, connectivity maps showed substantial overlap between the two stories.
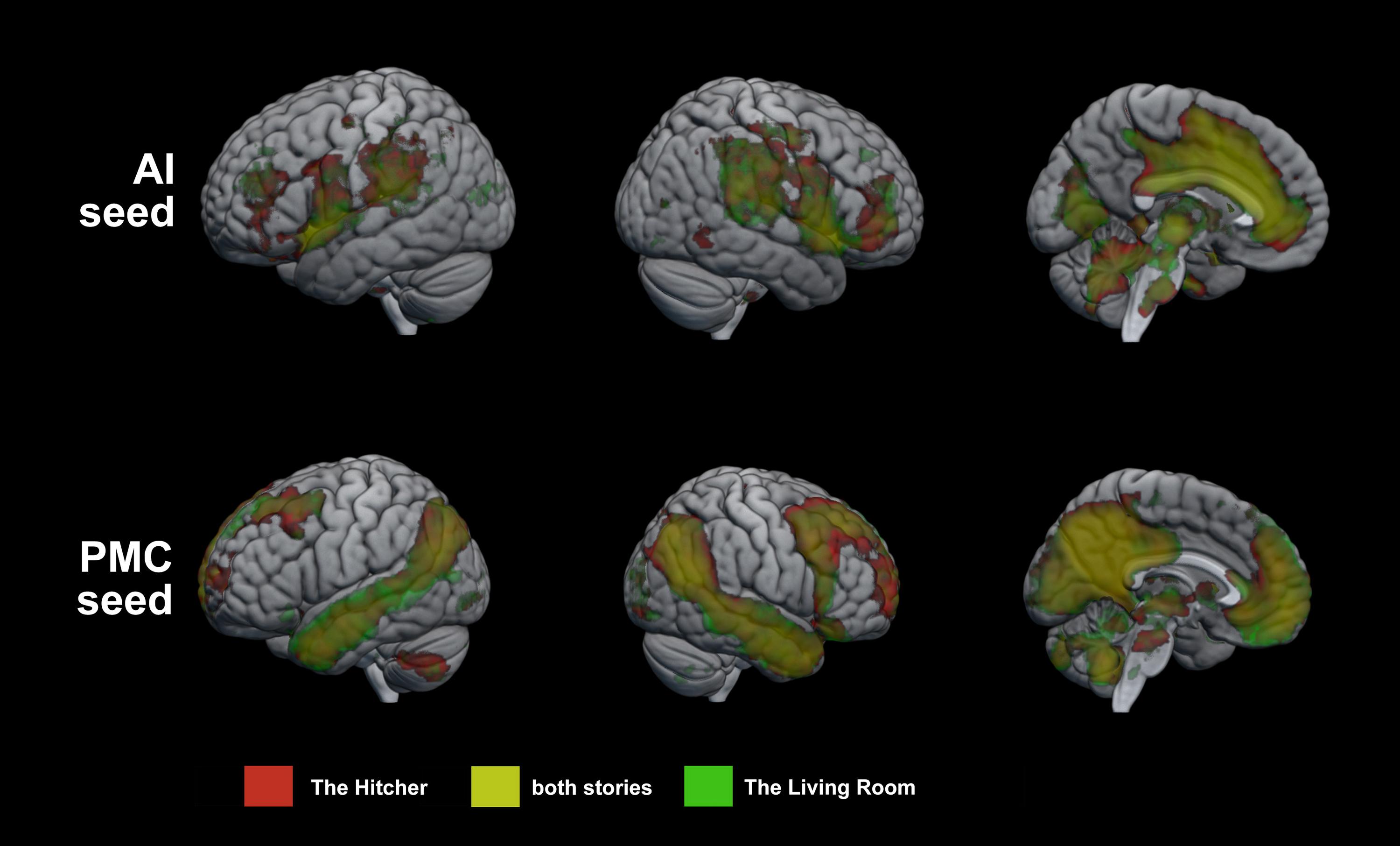
Figure 1. Group GLM of seed connectivity with AI and PMC. Regions of significant connectivity with the seed regions during story listening. Red = The Hitcher, Green = The Living Room, Yellow = connectivity with the seed during both stories.
For the cancer story, the mean split lead to 16 participants being classified as “low” in narrative transportation, and 20 being classified as “high.” For the hitchhiking story, the two groups were equally split with 18 participants each.
No clusters of AI connectivity significantly predicted transportation during the cancer story. Transportation during the hitchhiking story was significantly predicted by AI connectivity to a large cluster in the left precentral gyrus (−54, −12, and 50; 86.1%) as well as to smaller clusters in left postcentral gyrus (−41,−38, and 56; 80.5%) the left thalamus (−12, −17, and 16; 83.3%), and the left putamen (−30, −15, and −5; 83.3%) (see Figure 2).
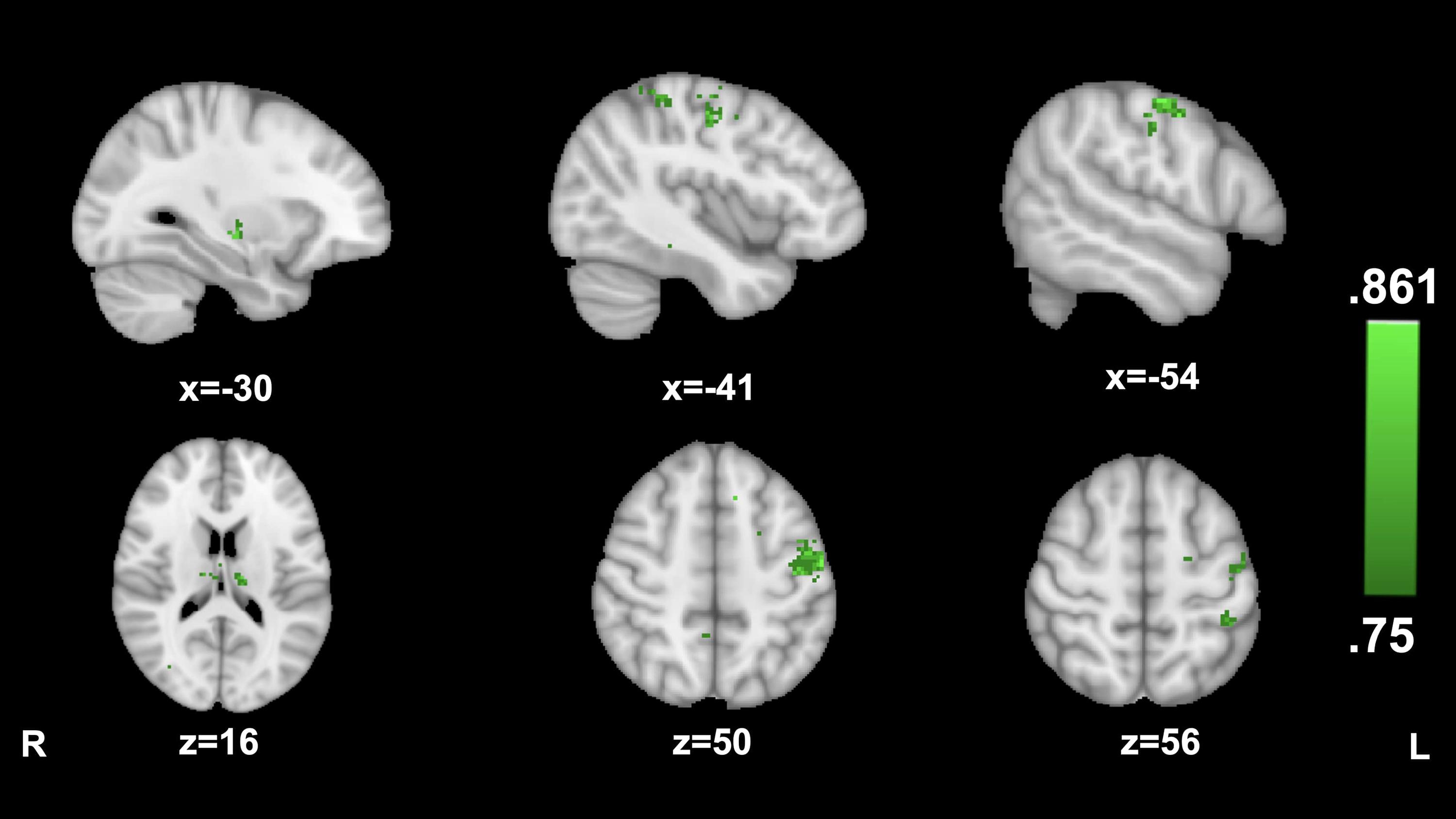
Figure 2. Anterior insula connectivity and narrative transportation during The Hitcher. MVPA searchlight showing above chance classification of AI connectivity maps for high vs. low transportation during the Hitchhiking story.
Transportation during the cancer story was significantly predicted by PMC connectivity to clusters in left inferior frontal gyrus (−35, 15, and 20; 86.1%), right temporal pole (20, −19, and −32; 83.3%), the left thalamus (−8, −13, and −2; 86.1%), and left postcentral gyrus (−8, −36, and 75; 83.3%).
Transportation during the hitchhiking story was significantly predicted by PMC connectivity with a cluster in the right caudate (9, 6, −3; 80.5%), as well as several smaller clusters in left inferior frontal gyrus (−44, 16, and 16; 80.5%), left inferior temporal cortex (−44, −26, and −20; 80.5%), and the left visual-association area (−30, −90, and 14; 83.3%) (see Figure 3).

Figure 3. PMC connectivity predictive of narrative transportation. MVPA searchlights showing above chance classification of PMC connectivity maps for high vs. low transportation during The Hitcher (green), and The Living Room (red).
For willingness to donate to cancer research, the mean split lead to 20 subjects being classified as “low” willingness and 16 as “high.” For willingness to donate to hitchhiking dangers awareness, 15 were classified as “low” and 21 were classified as “high.”
Reported willingness to donate to cancer research was predicted by insula connectivity with small clusters in the precuneus (4, −47, and 50; 83.3%) and in the left posterior fusiform gyrus (−40, −32, and −27; 86.1%) while listening to the cancer story. The reported willingness to sign the petition on hitchhiking dangers awareness was predicted by insula connectivity with a cluster in the right anterior prefrontal cortex (23, 50, and 14; 83.3%) (see Figure 4).
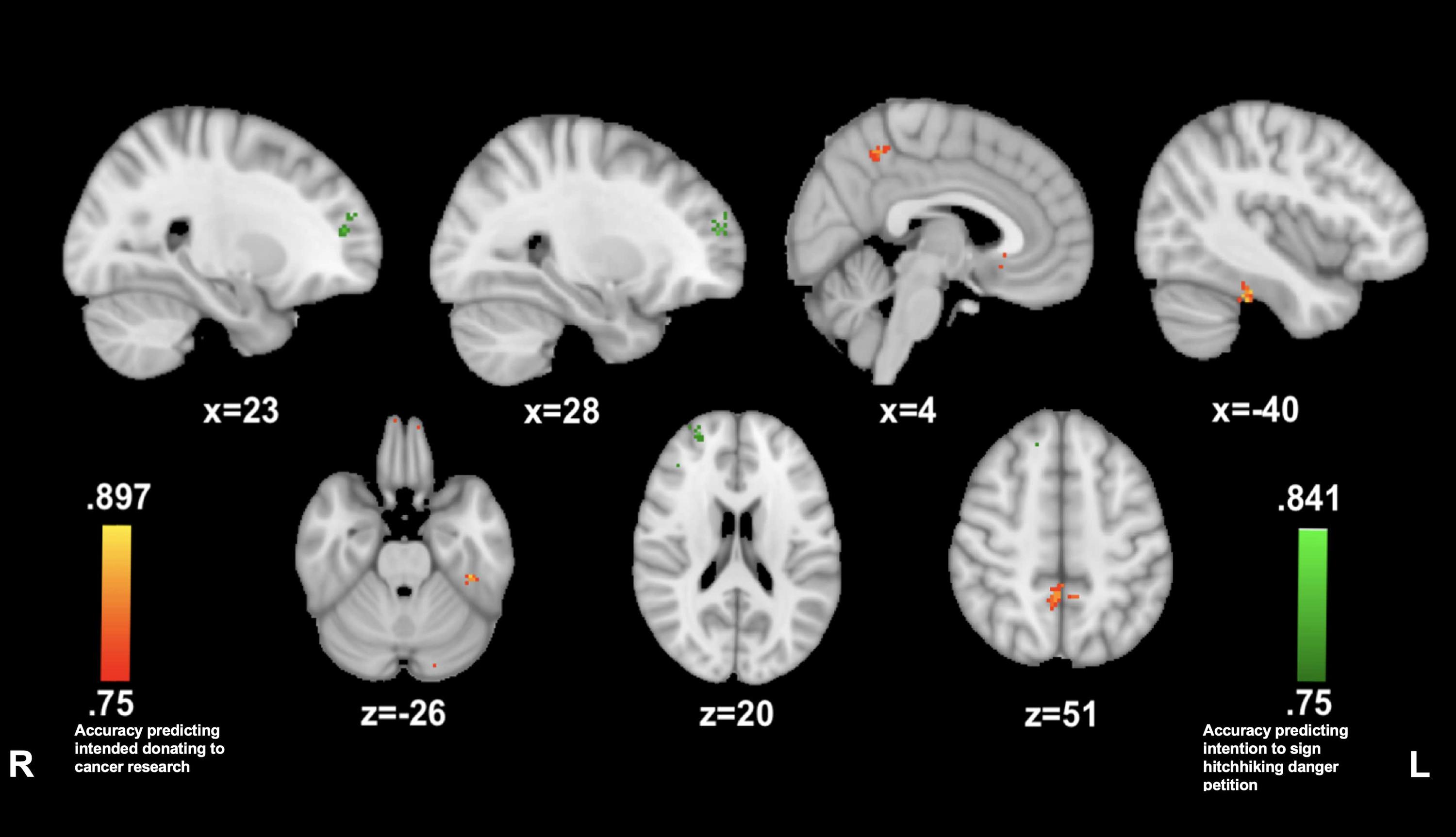
Figure 4. Anterior insula connectivity predictive of prosocial intention during The Living Room. MVPA searchlights showing above chance classification of AI connectivity maps for high vs. low prosocial intention for related (red) vs. unrelated (green) issues during The Living Room.
Reported willingness to sign the petition on hitchhiking dangers awareness was predicted by a prominent cluster of insula connectivity to the right anterior prefrontal cortex (29, 45, and 20; 86.1%) while listening to the hitchhiking story. The reported willingness to donate to cancer research was predicted by insula connectivity to clusters in left precentral gyrus (−12, −10, and 62; 91.6%) precuneus (13, −66, and 28; 88.8%) and right superior temporal gyrus (43, −48, and −9; 83.3%) (see Figure 5).
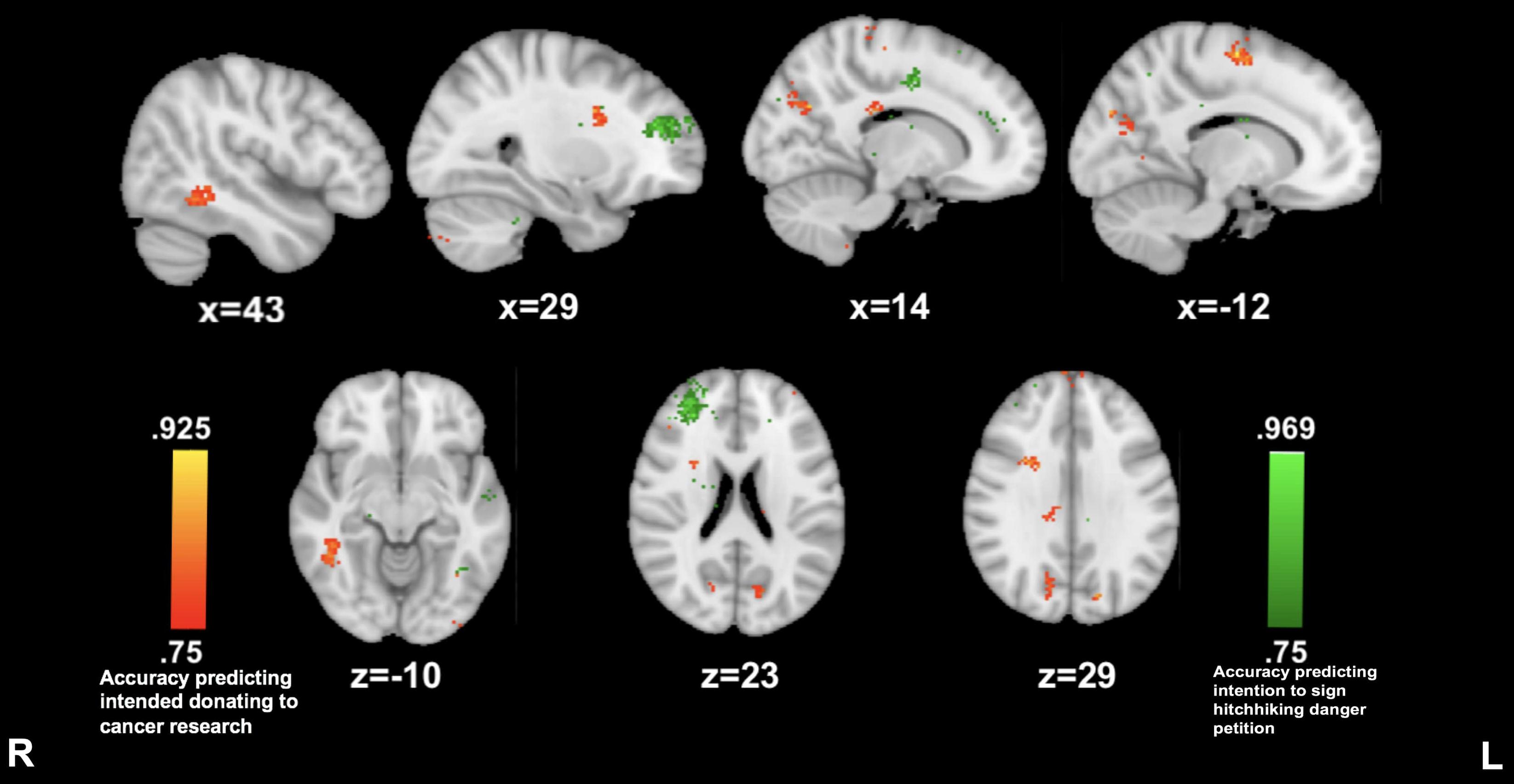
Figure 5. Anterior insula connectivity predictive of prosocial intention during The Hitcher. MVPA searchlights showing above chance classification of AI connectivity maps for high vs. low prosocial intention for related (red) vs. unrelated (green) issues during The Hitcher.
Reported willingness to donate to cancer was predicted by PMC connectivity with left postcentral gyrus (−43, −22, and 20; 88.8%), the anterior cingulate (−2, 3, 32; 88.8%), right supramarginal gyrus (53, −34, and 32; 86.1%), and right supplementary motor area (36, −4, and 53; 86.1%). Reported willingness to sign the petition on hitchhiking dangers awareness was predicted by PMC connectivity to the left premotor area (−16, −20, and 49; 88.8%) and to the left angular gyrus (−62, −44, and 27; 83.3%) (see Figure 6).
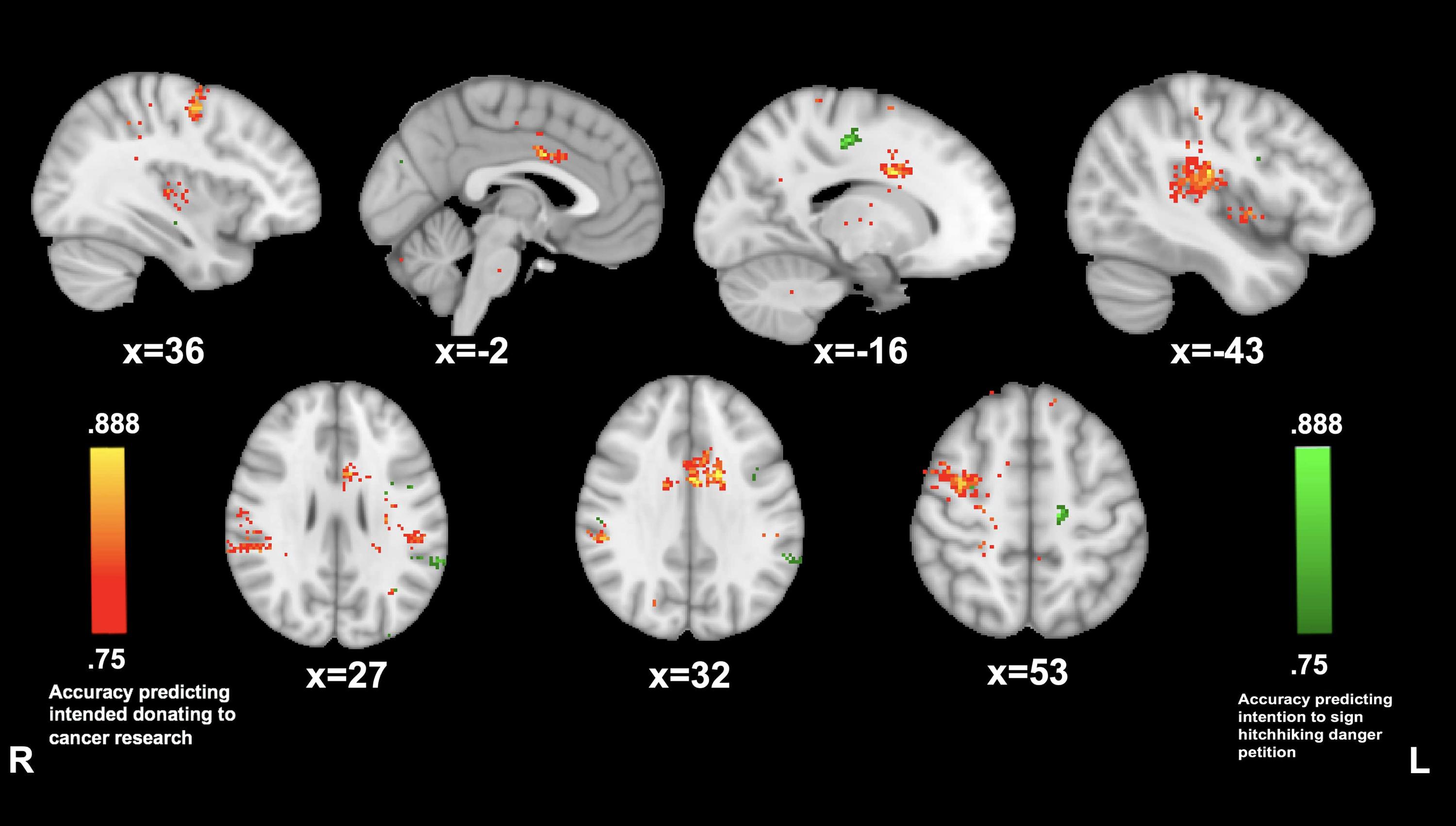
Figure 6. Posterior medial cortex connectivity predictive of prosocial intention during The Living Room. MVPA searchlights showing above chance classification of PMC connectivity maps for high vs. low prosocial intention for related (red) vs. unrelated (green) issues during The Living Room.
PMC connectivity while listening to the hitchhiking story was neither significantly predictive of willingness to donate to cancer research nor willingness to sign a hitchhiking dangers petition.
Our hypothesis that patterns of functional connectivity would be predictive of behavioral characteristics was largely supported. Patterns of AI connectivity significantly predicted the intention to support to both causes during both stories. AI connectivity, however, was only predictive of narrative transportation during the hitchhiking story. Patterns of PMC functional connectivity were only predictive of the intention to support causes during the cancer story. However, PMC functional connectivity was predictive of narrative transportation for both stories. These data demonstrate that there are different patterns of functional connectivity while listening to stories between those who feel transported and those who do not. Likewise, there are different patterns of functional connectivity between those who intend to support a cause and those who do not. The predictive patterns of connectivity in these analyses did correspond to some DMN and sensorimotor regions as predicted (such as angular gyrus, the precuneus, precentral gyrus, and postcentral gyrus).
More ambitiously, we hypothesized that patterns of connectivity to DMN and sensorimotor areas would be consistent across both stories. This consistency would demonstrate that the underlying processes were general to narratives rather than specific to one story or the other. For the analyses involving narrative transportation, this hypothesis would have been borne out if the seed regions were significantly connected with the same clusters in both stories. For the most part this was not supported by the data. Despite similar patterns of functional connectivity while listening to both stories, there was only one area of possible overlap for the two stories in predicting narrative transportation: PMC connectivity with left inferior frontal gyrus (IFG). This region was not one we had predicted, and will be further discussed.
For the analyses predicting prosocial intentions, we hypothesized that connectivity from the seed region would be the same for the cancer story predicting intention to donate to cancer research as it would be for the hitchhiking story predicting intention to sign the hitchhiking danger petition. In short, this would demonstrate functional connectivity consistently corresponding to a story-specific influence. This was not the case. Furthermore, we found that functional connectivity during stories were predictive of the intention to support causes that were irrelevant to its content. The lack of a story-specific effect suggests that these patterns of functional connectivity were not directly related to the stories’ influence persuading people on specific causes.
While these results did not suggest a unified process across narrative transportation and persuasion, these data may show that these processes are specific to story content. The specific content of a story may influence which processes play a role in transportation or in eliciting empathetic responses. For example: anecdotal reports from listeners suggest that the hitchhiking story was less emotionally compelling throughout most of its length. Long periods without emotionally gripping content may make the story simply a worse stimulus for studying narratives’ effects on persuading prosocial responses. Furthermore, unique elements of the hitchhiking story, such as more physical action, may explain why its AI connectivity patterns could successfully classify subjects based on narrative transportation. Notably, subjects reported being more transported into the Living Room story. This difference in transportation between the two stories may contribute to differences in how connectivity patterns are predictive of prosocial causes.
The differences in the functional connectivity patterns underlying transportation and intention to support causes between the two stories highlights a challenge for fMRI studies that employ naturalistic stimuli. There is a necessary tradeoff in studies using this design between ecological validity and experimental control. Because stories, movies, and musical pieces that aren’t designed by experimenters will necessarily differ from each other in content, emotional dynamics, and many other characteristics, results obtained with a single stimulus are less likely to generalize to others. The same qualities that make these media compelling as stimuli make them unique. One lesson we take from our results, therefore, is to stress the need for analyzing multiple different naturalistic stimuli in order to draw generalizable conclusions.
The one consistent pattern of connectivity found between both stories was connectivity between the PMC and the IFG predicting high vs. low transportation. The left IFG and PMC have been shown to work in tandem for a variety of tasks. For example, when listening to music the left IFG is believe to play a role in perceiving emotion in the musical structure whereas the PMC is involved in feeling those emotions (Tabei, 2015). The left IFG and PMC also work together for recognition memory of newly encoded musical stimuli (Watanabe et al., 2008) and pictures (Lundstrom et al., 2005). There are obvious parallels between these processes and those believed to be at play in narrative processing. One of the cognitive processes which makes narrative transportation possible is perceiving emotions and contexts within stimuli which relate to autobiographical memories (Baumgartner et al., 1992; Hartmann et al., 2016). The perception of these elements in narratives may trigger an involuntary retrieval of autobiographical memories (Moore and Homer, 2008; McDonald et al., 2015). Like with music and pictures, the IFG may parse the emotional and content-related cues in narratives before PMC regions relate them to memory traces. The linguistic nature of stories may be further evidence of this pathways role. The left IFG is known to play a role in parsing the semantic units of sentences, such as where the verb or subject is located within a sentence. The precuneus may then “store” these units to construct an overall meaning (Chou et al., 2009; Obleser and Kotz, 2010; Meyer et al., 2012). The use of this pathway in the parsing semantic meaning coupled with the PMC’s role in autobiographical memory and the self may explain this connection between narrative comprehension and the self (Regev et al., 2013). In fact, the left IFG has been found to be active in 55.9% of all studies focusing on self-referential processes (Morin and Michaud, 2007). It is believed this represents the crucial role of inner-speech in self-referential processes. This idea mirrors the theory that autobiographical knowledge is organized in a narrative manner, demonstrating how linked these linguistic forms of semantic ordering are to the self (Robinson and Taylor, 1998; Singer and Bluck, 2001; Conway et al., 2004).
For the hitchhiking story, AI connectivity was found to be predictive of narrative transportation. The regions where AI connectivity and was predictive of narrative transportation (precentral gyrus, postcentral gyrus, the thalamus, and the putamen) appear to tell a unified story. These regions may suggest a link with sensory and motor imagery (Szameitat et al., 2007; Belardinelli et al., 2009). Interestingly, it is the posterior insula not the AI which is normally associated with functional connectivity to sensorimotor regions (Cauda et al., 2011). However, some data suggests that the AI may play a role in integrating sensorimotor information with affective information (Kurth et al., 2010; Kleber et al., 2017). The hitchhiking story at its climax involves the protagonists making a run away from their potential captors: an element of physicality not present in the cancer story. It may be this sensorimotor imagery which creates the strongest link for immersion into the hitchhiking story. Those who can more effectively simulate these actions have more functional connectivity between the AI and sensorimotor regions, and are more likely to feel as if they are truly within the story. This would be a different cognitive process facilitating narrative transportation for this story as compared to the cancer story.
The patterns of PMC connectivity which predicted the intention to donate to cancer research while listening to the story may relate to listeners’ empathy with the characters in this emotionally evocative story. The anterior cingulate, supplementary motor area, postcentral gyrus, and supramarginal gyrus have all been found to associate with empathy for pain (Lloyd et al., 2004; Lawrence et al., 2006; Morelli et al., 2014). Furthermore, the activity and structural integrity of these regions have been found to associate with reported intention for, and real-world, prosocial behaviors (Hayashi et al., 2014; Morelli et al., 2014). The cancer story involved vivid discussions of watching somebody get sicker and sicker. It is likely that more vividly somebody experiences this character’s suffering the more they empathize, and thus the more the narrative influences their intention to donate to cancer research. The right supramarginal gyrus may play an important role in this process. This region is believed to be involved in determining the emotional “reference point” through which judgments are made; simply put: whether or not the person of reference is the self or another person (Hayashi et al., 2014; Hoffmann et al., 2016). While the supramarginal gyrus is under normal circumstances more active for self-reference than for referencing others (Vogeley and Fink, 2003; van der Heiden et al., 2013), it has been found that heightened activity in the region associates with using a third-party as the emotional point of reference (Silani et al., 2013; Steinbeis and Singer, 2014; Riva et al., 2016). PMC connectivity to this region may represent this “switching” mechanism, with the emotional information processed from the story now being utilized as the new reference point. This would also explain why motor and sensory regions were also experiencing connectivity with the PMC, simulating the physical experience of the character.
While we hypothesized that pro-social intentions would only be predicted in a story-specific manner (i.e., the cancer story would predict willingness to donate to cancer and the hitchhiking story would predict willingness to donate to hitchhiking danger awareness), this was not the case. AI connectivity was predictive of pro-social intentions for both causes during both stories, and PMC connectivity was predictive of pro-social intentions for both causes during The Living Room. Functional connectivity during different stories therefore does not appear to be able to disentangle personal characteristics from story-specific influence in this case. It is possible that these patterns of connectivity are related to the manner in which individuals engage with all narratives regardless of the specific context. Pro-social giving was significantly correlated across questions; in other words, people who indicated prosocial intent for one scenario were more likely to indicate high prosocial intent for the other as well. The trait underlying this willingness to give may be marked by specific patterns of functional connectivity. For example, AI connectivity to the right anterior prefrontal cortex was significantly predictive of willingness to donate to the hitchhiking dangers petition during both stories. Connectivity between the AI and prefrontal cortex has been previously implicated in the regulation of empathic responses (Pino et al., 2016; Yao et al., 2016). This type of individual ability could be evident during narrative processing and relate to the tendency to give regardless of the specific context of the narrative.
We found that individual differences in the subjective immersion into a narrative world, and the intention to contribute to a cause related to the narrative can be accurately predicted from functional connectivity while listening to the story. Our more specific hypotheses on what these patterns of predictive connectivity would look like focused on what processes may be alike between the two stories. The results instead reveal a more valuable question: how does a story’s specific content influence the cognitive and affective functions used to process it? With the diversity of cognitive processes thought to be involved narrative transportation (empathy, autobiographical memory, perspective taking, mental imagery etc.) different stories are bound to be facilitated differentially by different processes. This underscores the necessity of employing multiple naturalistic stimuli that differ in content when studying generalized processes like transportation and empathy, to support generalization of results.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The studies involving human participants were reviewed and approved by USC Institutional Review Board. The patients/participants provided their written informed consent to participate in this study.
SG helped to design the study and collect the data. BS participated in the data analysis. AV participated in the data analysis and writing the manuscript. JK participated in the study design, data collection, analysis, and manuscript preparation. All authors contributed to the article and approved the submitted version.
This work was supported by the Institute for New Economic Thinking (award #INO15-00037) and the Templeton World Charity Foundation (award #TWCF-0334).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Appel, M., Gnambs, T., Richter, T., and Green, M. C. (2015). The transportation scale–short form (TS–SF). Media Psychol. 18, 243–266. doi: 10.1080/15213269.2014.987400
Appel, M., and Richter, T. (2010). Transportation and need for affect in narrative persuasion: a mediated moderation model. Media Psychol. 13, 101–135. doi: 10.1080/15213261003799847
Araujo, H. F., Kaplan, J., Damasio, H., and Damasio, A. (2015). Neural correlates of different self domains. Brain Behav. 5:e00409. doi: 10.1002/brb3.409
Baldassano, C., Chen, J., Zadbood, A., Pillow, J. W., Hasson, U., and Norman, K. A. (2017). Discovering event structure in continuous narrative perception and memory. Neuron 95, 709–721.e5. doi: 10.1016/j.neuron.2017.06.041
Baldassano, C., Hasson, U., and Norman, K. A. (2018). Representation of real-world event schemas during narrative perception. J. Neurosci. 38, 9689–9699. doi: 10.1523/JNEUROSCI.0251-18.2018
Banerjee, S. C., and Greene, K. (2012). Role of transportation in the persuasion process: cognitive and affective responses to Antidrug narratives. J. Health Commun. 17, 564–581. doi: 10.1080/10810730.2011.635779
Batat, W., and Wohlfeil, M. (2009). Getting lost ‘Into the Wild’: understanding consumers’ movie enjoyment through a narrative transportation approach. Adv. Consum. Res. 36, 372–377.
Baumgartner, H., Sujan, M., and Bettman, J. R. (1992). Autobiographical memories, affect, and consumer information processing. J. Consum. Psychol. 1, 53–82. doi: 10.1207/s15327663jcp0101_04
Belardinelli, M. O., Palmiero, M., Sestieri, C., Nardo, D., Di Matteo, R., Londei, A., et al. (2009). An fMRI investigation on image generation in different sensory modalities: the influence of vividness. Acta Psychol. 132, 190–200. doi: 10.1016/j.actpsy.2009.06.009
Bilandzic, H., and Busselle, R. (2013). “Narrative persuasion,” in The Sage Handbook of Persuasion: Developments in Theory and Practice, 2nd Edn, eds J. P. Dillard and L. Shen (Thousand Oaks, CA: Sage), 200–219. doi: 10.4135/9781452218410.n13
Breen, A. V., McLean, K. C., Cairney, K., and McAdams, D. P. (2017). Movies, books, and identity: exploring the narrative ecology of the self. Qual. Psychol. 4, 243–259. doi: 10.1037/qup0000059
Bruneau, E., Dufour, N., and Saxe, R. (2013). How we know it hurts: item analysis of written narratives reveals distinct neural responses to others’ physical pain and emotional suffering. PLoS One 8:e63085. doi: 10.1371/journal.pone.0063085
Caputo, N. M., and Rouner, D. (2011). Narrative processing of entertainment media and mental illness stigma. Health Commun. 26, 595–604. doi: 10.1080/10410236.2011.560787
Cauda, F., D’Agata, F., Sacco, K., Duca, S., Geminiani, G., and Vercelli, A. (2011). Functional connectivity of the insula in the resting brain. Neuroimage 55, 8–23. doi: 10.1016/j.neuroimage.2010.11.049
Chou, T.-L., Chen, C.-W., Wu, M.-Y., and Booth, J. R. (2009). The role of inferior frontal gyrus and inferior parietal lobule in semantic processing of Chinese characters. Exp. Brain Res. 198, 465–475. doi: 10.1007/s00221-009-1942-y
Christy, K. R., and Fox, J. (2016). Transportability and presence as predictors of avatar identification within narrative video games. Cyberpsychol. Behav. Soc. Netw. 19, 283–287. doi: 10.1089/cyber.2015.0474
Conway, M. A., Singer, J. A., and Tagini, A. (2004). The self and autobiographical memory: correspondence and coherence. Soc. Cogn. 22, 491–529. doi: 10.1521/soco.22.5.491.50768
Craig, A. D. (2009). How do you feel–now? The anterior insula and human awareness. Nat. Rev. Neurosci. 10, 59–70. doi: 10.1038/nrn2555
Deen, B., Pitskel, N. B., and Pelphrey, K. A. (2010). Three systems of insular functional connectivity identified with cluster analysis. Cereb. Cortex 21, 1498–1506. doi: 10.1093/cercor/bhq186
Dehghani, M., Boghrati, R., Man, K., Hoover, J., Gimbel, S. I., Vaswani, A., et al. (2017). Decoding the neural representation of story meanings across languages. Hum. Brain Mapp. 38, 6096–6106. doi: 10.1002/hbm.23814
Deighton, J., Romer, D., and McQueen, J. (1989). Using drama to persuade. J. Consum. Res. 16, 335–343. doi: 10.1086/209219
Dillard, A. J., Ferrer, R. A., and Welch, J. D. (2018). Associations between narrative transportation, risk perception and behaviour intentions following narrative messages about skin cancer. Psychol. Health 33, 573–593. doi: 10.1080/08870446.2017.1380811
Dunlop, S. M., Wakefield, M., and Kashima, Y. (2010). Pathways to persuasion: cognitive and experiential responses to health-promoting mass media messages. Commun. Res. 37, 133–164. doi: 10.1177/0093650209351912
Escalas, J. E. (2004). Imagine yourself in the product: mental simulation, narrative transportation, and persuasion. J. Advert. 33, 37–48. doi: 10.1080/00913367.2004.10639163
Feng, C., Yan, X., Huang, W., Han, S., and Ma, Y. (2018). Neural representations of the multidimensional self in the cortical midline structures. Neuroimage 183, 291–299. doi: 10.1016/j.neuroimage.2018.08.018
Fransson, P., and Marrelec, G. (2008). The precuneus/posterior cingulate cortex plays a pivotal role in the default mode network: evidence from a partial correlation network analysis. Neuroimage 42, 1178–1184. doi: 10.1016/j.neuroimage.2008.05.059
Green, M. C., and Brock, T. C. (2000). The role of transportation in the persuasiveness of public narratives. J. Pers. Soc. Psychol. 79, 701–721. doi: 10.1037/0022-3514.79.5.701
Green, M. C., Brock, T. C., and Kaufman, G. F. (2004). Understanding media enjoyment: the role of transportation into narrative worlds. Commun. Theory 14, 311–327. doi: 10.1111/j.1468-2885.2004.tb00317.x
Green, M. C., Chatham, C., and Sestir, M. A. (2012). Emotion and transportation into fact and fiction. Sci. Study Lit. 2, 37–59. doi: 10.1075/ssol.2.1.03gre
Green, M. C., and Clark, J. L. (2013). Transportation into narrative worlds: implications for entertainment media influences on tobacco use. Addiction 108, 477–484. doi: 10.1111/j.1360-0443.2012.04088.x
Green, M. C., and Donahue, J. K. (2009). “Simulated worlds: transportation into narratives,” in Handbook of Imagination and Mental Simulation, eds K. Markman, W. M. Klein, and J. A. Suhr (New York, NY: Psychology Press) 241–256.
Green, M. C., and Sestir, M. (2017). Transportation Theory. The International Encyclopedia of Media Effects (Hoboken, NJ: John Wiley and Sons, Inc), 1–14. doi: 10.1002/9781118783764.wbieme0083
Greenwood, D. N., and Long, C. R. (2009). Psychological predictors of media involvement: solitude experiences and the need to belong. Commun. Res. 36, 637–654. doi: 10.1177/0093650209338906
Gu, X., Hof, P. R., Friston, K. J., and Fan, J. (2013). Anterior insular cortex and emotional awareness. J. Comp. Neurol. 521, 3371–3388. doi: 10.1002/cne.23368
Guerrero, I., and Igartua, J. J. (2017). “The role of the narrator point of view and the similarity with the protagonist in narratives designed to reduce prejudice towards stigmatized immigrants,” in Proceedings of the 5th International Conference on Technological Ecosystems for Enhancing Multiculturality, Vol. 94, (New York, NY), 1–5. doi: 10.1145/3144826.3145441
Hall, A. E., and Bracken, C. C. (2011). I really liked that movie. Testing the relationship between trait empathy, transportation, perceived realism, and movie enjoyment. J. Media Psychol. 23, 90–99. doi: 10.1027/1864-1105/a000036
Hanke, M., Halchenko, Y. O., Sederberg, P. B., Hanson, S. J., Haxby, J. V., and Pollmann, S. (2009). PyMVPA: a python toolbox for multivariate pattern analysis of fMRI data. Neuroinformatics 7, 37–53. doi: 10.1007/s12021-008-9041-y
Harjusola-Webb, S., Hubbell, S. P., and Bedesem, P. (2012). Increasing prosocial behaviors of young children with disabilities in inclusive classrooms using a combination of peer-mediated intervention and social narratives. Beyond Behav. 21, 29–36.
Hartmann, P., Apaolaza, V., and Eisend, M. (2016). Nature imagery in non-green advertising: the effects of emotion, autobiographical memory, and consumer’s green traits. J. Advert. 45, 427–440. doi: 10.1080/00913367.2016.1190259
Hasson, U., Furman, O., Clark, D., Dudai, Y., and Davachi, L. (2008). Enhanced intersubject correlations during movie viewing correlate with successful episodic encoding. Neuron 57, 452–462. doi: 10.1016/j.neuron.2007.12.009
Hayashi, A., Abe, N., Fujii, T., Ito, A., Ueno, A., Koseki, Y., et al. (2014). Dissociable neural systems for moral judgment of anti-and pro-social lying. Brain Res. 1556, 46–56. doi: 10.1016/j.brainres.2014.02.011
Hirschman, E. C. (1983). Predictors of self-projection, fantasy fulfillment, and escapism. J. Soc. Psychol. 120, 63–76. doi: 10.1080/00224545.1983.9712011
Hoffmann, F., Koehne, S., Steinbeis, N., Dziobek, I., and Singer, T. (2016). Preserved self-other distinction during empathy in autism is linked to network integrity of right supramarginal gyrus. J. Autism. Dev. Disord. 46, 637–648. doi: 10.1007/s10803-015-2609-0
Igartua, J.-J., and Frutos, F. J. (2017). Enhancing attitudes toward stigmatized groups with movies: mediating and moderating processes of narrative persuasion. Int. J. Commun. 11, 158–177.
Immordino-Yang, M. H., McColl, A., Damasio, H., and Damasio, A. (2009). Neural correlates of admiration and compassion. Proc. Natl. Acad. Sci. U.S.A. 106, 8021–8026. doi: 10.1073/pnas.0810363106
Immordino-Yang, M. H., Yang, X.-F., and Damasio, H. (2014). Correlations between social-emotional feelings and anterior insula activity are independent from visceral states but influenced by culture. Front. Hum. Neurosci. 8:728. doi: 10.3389/fnhum.2014.00728
Immordino-Yang, M. H., Yang, X.-F., and Damasio, H. (2016). Cultural modes of expressing emotions influence how emotions are experienced. Emotion 16, 1033–1039. doi: 10.1037/emo0000201
Jabbi, M., Bastiaansen, J., and Keysers, C. (2008). A common anterior insula representation of disgust observation, experience and imagination shows divergent functional connectivity pathways. PLoS One 3:e2939. doi: 10.1371/journal.pone.0002939
Johnson, D. R. (2012). Transportation into a story increases empathy, prosocial behavior, and perceptual bias toward fearful expressions. Pers. Individ. Dif. 52, 150–155. doi: 10.1016/j.paid.2011.10.005
Johnson, D. R., Huffman, B. L., and Jasper, D. M. (2014). Changing race boundary perception by reading narrative fiction. Basic Appl. Soc. Psychol. 36, 83–90. doi: 10.1080/01973533.2013.856791
Johnson, D. R., Jasper, D. M., Griffin, S., and Huffman, B. L. (2013). Reading narrative fiction reduces Arab-Muslim prejudice and offers a safe haven from intergroup anxiety. Soc. Cogn. 31, 578–598. doi: 10.1521/soco.2013.31.5.578
Kaplan, J., Gimbel, S. I., Dehghani, M., Immordino-Yang, M. H., Sagae, K., Wong, J. D., et al. (2017). Processing narratives concerning protected values: a cross-cultural investigation of neural correlates. Cereb. Cortex 27, 1428–1438.
Kaplan, J., and Meyer, K. (2012). Multivariate pattern analysis reveals common neural patterns across individuals during touch observation. Neuroimage 60, 204–212. doi: 10.1016/j.neuroimage.2011.12.059
Khalsa, S., Mayhew, S. D., Chechlacz, M., Bagary, M., and Bagshaw, A. P. (2014). The structural and functional connectivity of the posterior cingulate cortex: comparison between deterministic and probabilistic tractography for the investigation of structure–function relationships. Neuroimage 102, 118–127. doi: 10.1016/j.neuroimage.2013.12.022
Kleber, B., Friberg, A., Zeitouni, A., and Zatorre, R. (2017). Experience-dependent modulation of right anterior insula and sensorimotor regions as a function of noise-masked auditory feedback in singers and nonsingers. Neuroimage 147, 97–110. doi: 10.1016/j.neuroimage.2016.11.059
Klimmt, C., Hefner, D., and Vorderer, P. (2009). The video game experience as “true” identification: a theory of enjoyable alterations of players’ self-perception. Commun. Theory 19, 351–373. doi: 10.1111/j.1468-2885.2009.01347.x
Kriegeskorte, N., and Bandettini, P. (2007). Analyzing for information, not activation, to exploit high-resolution fMRI. Neuroimage 38, 649–662. doi: 10.1016/j.neuroimage.2007.02.022
Kriegeskorte, N., Goebel, R., and Bandettini, P. (2006). Information-based functional brain mapping. Proc. Natl. Acad. Sci. U.S.A. 103, 3863–3868. doi: 10.1073/pnas.0600244103
Kuo, A., Lutz, R. J., and Hiler, J. L. (2016). Brave new World of Warcraft: a conceptual framework for active escapism. J. Consum. Mark. 33, 498–506. doi: 10.1108/jcm-04-2016-1775
Kurth, F., Zilles, K., Fox, P. T., Laird, A. R., and Eickhoff, S. B. (2010). A link between the systems: functional differentiation and integration within the human insula revealed by meta-analysis. Brain Struct. Funct. 214, 519–534. doi: 10.1007/s00429-010-0255-z
Larkey, L. K., and Hecht, M. (2010). A model of effects of narrative as culture-centric health promotion. J. Health Commun. 15, 114–135. doi: 10.1080/10810730903528017
Lawrence, E. J., Shaw, P., Giampietro, V., Surguladze, S., Brammer, M. J., and David, A. S. (2006). The role of ‘shared representations’ in social perception and empathy: an fMRI study. Neuroimage 29, 1173–1184. doi: 10.1016/j.neuroimage.2005.09.001
Lerner, Y., Honey, C. J., Silbert, L. J., and Hasson, U. (2011). Topographic mapping of a hierarchy of temporal receptive windows using a narrated story. J. Neurosci. 31, 2906–2915. doi: 10.1523/JNEUROSCI.3684-10.2011
Li, D., Liau, A., and Khoo, A. (2011). Examining the influence of actual-ideal self-discrepancies, depression, and escapism, on pathological gaming among massively multiplayer online adolescent gamers. Cyberpsychol. Behav. Soc. Netw. 14, 535–539. doi: 10.1089/cyber.2010.0463
Lloyd, D., Di Pellegrino, G., and Roberts, N. (2004). Vicarious responses to pain in anterior cingulate cortex: is empathy a multisensory issue? Cogn. Affect. Behav. Neurosci. 4, 270–278. doi: 10.3758/cabn.4.2.270
Lundstrom, B. N., Ingvar, M., and Petersson, K. M. (2005). The role of precuneus and left inferior frontal cortex during source memory episodic retrieval. Neuroimage 27, 824–834. doi: 10.1016/j.neuroimage.2005.05.008
Mazzocco, P. J., Green, M. C., Sasota, J. A., and Jones, N. W. (2010). This story is not for everyone: transportability and narrative persuasion. Soc. Psychol. Pers. Sci. 1, 361–368. doi: 10.1177/1948550610376600
McDonald, D. G., Sarge, M. A., Lin, S.-F., Collier, J. G., and Potocki, B. (2015). A role for the self: media content as triggers for involuntary autobiographical memories. Commun. Res. 42, 3–29. doi: 10.1177/0093650212464771
Meyer, L., Obleser, J., Anwander, A., and Friederici, A. D. (2012). Linking ordering in Broca’s area to storage in left temporo-parietal regions: the case of sentence processing. Neuroimage 62, 1987–1998. doi: 10.1016/j.neuroimage.2012.05.052
Moore, D. J., and Homer, P. M. (2008). Self-brand connections: the role of attitude strength and autobiographical memory primes. J. Bus. Res. 61, 707–714. doi: 10.1016/j.jbusres.2007.09.002
Morelli, S. A., Rameson, L. T., and Lieberman, M. D. (2014). The neural components of empathy: predicting daily prosocial behavior. Soc. Cogn. Affect. Neurosci. 9, 39–47. doi: 10.1093/scan/nss088
Morin, A., and Michaud, J. (2007). Self-awareness and the left inferior frontal gyrus: inner speech use during self-related processing. Brain Res. Bull. 74, 387–396. doi: 10.1016/j.brainresbull.2007.06.013
Morris, B. S., Chrysochou, P., Christensen, J. D., Orquin, J. L., Barraza, J., Zak, P. J., et al. (2019). Stories vs. facts: triggering emotion and action-taking on climate change. Clim. Change 154, 19–36. doi: 10.1007/s10584-019-02425-6
Moyer-Gusé, E., Dale, K. R., and Ortiz, M. (2019). Reducing prejudice through narratives: an examination of the mechanisms of vicarious intergroup contact. J. Media Psychol. 31, 185–195. doi: 10.1027/1864-1105/a000249
Nguyen, M., Vanderwal, T., and Hasson, U. (2019). Shared understanding of narratives is correlated with shared neural responses. Neuroimage 184, 161–170. doi: 10.1016/j.neuroimage.2018.09.010
Nguyen, V. T., Breakspear, M., Hu, X., and Guo, C. C. (2016). The integration of the internal and external milieu in the insula during dynamic emotional experiences. Neuroimage 124, 455–463. doi: 10.1016/j.neuroimage.2015.08.078
Nomi, J. S., Farrant, K., Damaraju, E., Rachakonda, S., Calhoun, V. D., and Uddin, L. Q. (2016). Dynamic functional network connectivity reveals unique and overlapping profiles of insula subdivisions. Hum. Brain Mapp. 37, 1770–1787. doi: 10.1002/hbm.23135
Norman, K. A., Polyn, S. M., Detre, G. J., and Haxby, J. V. (2006). Beyond mind-reading: multi-voxel pattern analysis of fMRI data. Trends Cogn. Sci. 10, 424–430. doi: 10.1016/j.tics.2006.07.005
Obleser, J., and Kotz, S. A. (2010). Expectancy constraints in degraded speech modulate the language comprehension network. Cereb. Cortex 20, 633–640. doi: 10.1093/cercor/bhp128
Parvizi, J., Van Hoesen, G. W., Buckwalter, J., and Damasio, A. (2006). Neural connections of the posteromedial cortex in the macaque. Proc. Natl. Acad. Sci. U.S.A. 103, 1563–1568. doi: 10.1073/pnas.0507729103
Pino, M. C., Tempesta, D., Catalucci, A., Anselmi, M., Nigri, A., Iaria, G., et al. (2016). Altered cortico-limbic functional connectivity during an empathy task in subjects with post-traumatic stress disorder. J. Psychopathol. Behav. Assess. 38, 398–405. doi: 10.1007/s10862-016-9538-x
Prebble, S. C., Addis, D. R., and Tippett, L. J. (2013). Autobiographical memory and sense of self. Psychol. Bull. 139, 815–840. doi: 10.1037/a0030146
Rameson, L. T., Satpute, A. B., and Lieberman, M. D. (2010). The neural correlates of implicit and explicit self-relevant processing. Neuroimage 50, 701–708. doi: 10.1016/j.neuroimage.2009.12.098
Rasmussen, A. S., Johannessen, K. B., and Berntsen, D. (2014). Ways of sampling voluntary and involuntary autobiographical memories in daily life. Conscious. Cogn. 30, 156–168. doi: 10.1016/j.concog.2014.09.008
Regev, M., Honey, C. J., Simony, E., and Hasson, U. (2013). Selective and invariant neural responses to spoken and written narratives. J. Neurosci. 33, 15978–15988. doi: 10.1523/jneurosci.1580-13.2013
Riva, F., Triscoli, C., Lamm, C., Carnaghi, A., and Silani, G. (2016). Emotional egocentricity bias across the life-span. Front. Aging Neurosci. 8:74. doi: 10.3389/fnagi.2016.00074
Robinson, J. A., and Taylor, L. R. (1998). “Autobiographical memory and self-narratives: a tale of two stories,” in Autobiographical Memory, eds C. P. Thompson D. J. Herrmann D. Bruce J. D. Read D. G. Payne and M. P. Toglia (Mahwah, NJ: Erlbaum), 125–143.
Seger, C. A., Stone, M., and Keenan, J. P. (2004). Cortical activations during judgments about the self and an other person. Neuropsychologia 42, 1168–1177. doi: 10.1016/j.neuropsychologia.2004.02.003
Sestir, M., and Green, M. C. (2010). You are who you watch: identification and transportation effects on temporary self-concept. Soc. Influ. 5, 272–288. doi: 10.1080/15534510.2010.490672
Silani, G., Lamm, C., Ruff, C. C., and Singer, T. (2013). Right supramarginal gyrus is crucial to overcome emotional egocentricity bias in social judgments. J. Neurosci. 33, 15466–15476. doi: 10.1523/jneurosci.1488-13.2013
Simony, E., Honey, C. J., Chen, J., Lositsky, O., Yeshurun, Y., Wiesel, A., et al. (2016). Dynamic reconfiguration of the default mode network during narrative comprehension. Nat. Commun. 7:12141.
Singer, J. A., and Blagov, P. (2004). “The integrative function of narrative processing: autobiographical memory, self-defining memories, and the life story of identity,” in Studies in Self and Identity. The Self and Memory, eds D. R. Beike, J. M. Lampinen, and D. A. Behrend (New York, NY: Psychology Press), 117–138.
Singer, J. A., and Bluck, S. (2001). New perspectives on autobiographical memory: the integration of narrative processing and autobiographical reasoning. Rev. Gen. Psychol. 5, 91–99. doi: 10.1037/1089-2680.5.2.91
Slater, M. D., Johnson, B. K., Cohen, J., Comello, M. L. G., and Ewoldsen, D. R. (2014). Temporarily expanding the boundaries of the self: motivations for entering the story world and implications for narrative effects. J. Commun. 64, 439–455. doi: 10.1111/jcom.12100
Slater, M. D., and Rouner, D. (2002). Entertainment—education and elaboration likelihood: understanding the processing of narrative persuasion. Commun. Theory 12, 173–191. doi: 10.1093/ct/12.2.173
Steinbeis, N., and Singer, T. (2014). Projecting my envy onto you: neurocognitive mechanisms of an offline emotional egocentricity bias. NeuroImage, 102, 370–380. doi: 10.1016/j.neuroimage.2014.08.007
Steinemann, S. T., Iten, G. H., Opwis, K., Forde, S. F., Frasseck, L., and Mekler, E. D. (2017). Interactive narratives affecting social change. J. Media Psychol. 29, 54–66. doi: 10.1027/1864-1105/a000211
Stolte, J. F., and Fender, S. (2007). Framing social values: an experimental study of culture and cognition. Soc. Psychol. Q. 70, 59–69. doi: 10.1177/019027250707000107
Summerfield, J. J., Hassabis, D., and Maguire, E. A. (2009). Cortical midline involvement in autobiographical memory. Neuroimage 44, 1188–1200. doi: 10.1016/j.neuroimage.2008.09.033
Swanson, R. W., Gordon, A. S., Khooshabeh, P., Sagae, K., Huskey, R., Mangus, M., et al. (2017). An empirical analysis of subjectivity and narrative levels in Weblog storytelling across cultures. Dialogue Discourse 8, 105–128. doi: 10.5087/dad.2017.205
Szameitat, A. J., Shen, S., and Sterr, A. (2007). Motor imagery of complex everyday movements. An fMRI study. Neuroimage 34, 702–713. doi: 10.1016/j.neuroimage.2006.09.033
Tabei, K.-I. (2015). Inferior frontal gyrus activation underlies the perception of emotions, while precuneus activation underlies the feeling of emotions during music listening. Behav. Neurol. 2015, 529043.
Uddin, L. Q., Kinnison, J., Pessoa, L., and Anderson, M. L. (2014). Beyond the tripartite cognition-emotion-interoception model of the human insular cortex. J. Cogn. Neurosci. 26, 16–27. doi: 10.1162/jocn_a_00462
Uddin, L. Q., Nomi, J. S., Hébert-Seropian, B., Ghaziri, J., and Boucher, O. (2017). Structure and function of the human insula. J. Clin. Neurophysiol. 34, 300–306. doi: 10.1097/wnp.0000000000000377
van der Heiden, L., Scherpiet, S., Konicar, L., Birbaumer, N., and Veit, R. (2013). Inter-individual differences in successful perspective taking during pain perception mediates emotional responsiveness in self and others: an fMRI study. Neuroimage 65, 387–394. doi: 10.1016/j.neuroimage.2012.10.003
Van Laer, T., De Ruyter, K., Visconti, L. M., and Wetzels, M. (2014). The extended transportation-imagery model: a meta-analysis of the antecedents and consequences of consumers’ narrative transportation. J. Consum. Res. 40, 797–817. doi: 10.1086/673383
Vogeley, K., and Fink, G. R. (2003). Neural correlates of the first-person-perspective. Trends Cogn. Sci. 7, 38–42. doi: 10.1016/s1364-661300003-7
Watanabe, T., Yagishita, S., and Kikyo, H. (2008). Memory of music: roles of right hippocampus and left inferior frontal gyrus. Neuroimage 39, 483–491. doi: 10.1016/j.neuroimage.2007.08.024
Weiss, J. K. (2015). Maybe Medium Does Matter: Considering Differences in Individual’s Trait Empathy and the Effect on Narrative Transportation. Buffalo, NY: State University of New York.
Yao, S., Becker, B., Geng, Y., Zhao, Z., Xu, X., Zhao, W., et al. (2016). Voluntary control of anterior insula and its functional connections is feedback-independent and increases pain empathy. Neuroimage 130, 230–240. doi: 10.1016/j.neuroimage.2016.02.035
Yeshurun, Y., Swanson, S., Simony, E., Chen, J., Lazaridi, C., Honey, C. J., et al. (2017). Same story, different story: the neural representation of interpretive frameworks. Psychol. Sci. 28, 307–319. doi: 10.1177/0956797616682029
Zaki, J., Davis, J. I., and Ochsner, K. N. (2012). Overlapping activity in anterior insula during interoception and emotional experience. Neuroimage 62, 493–499. doi: 10.1016/j.neuroimage.2012.05.012
Keywords: fMRI, narrative, narrative transportation, naturalistic stimuli, functional connectivity, MVPA
Citation: Vaccaro AG, Scott B, Gimbel SI and Kaplan JT (2021) Functional Brain Connectivity During Narrative Processing Relates to Transportation and Story Influence. Front. Hum. Neurosci. 15:665319. doi: 10.3389/fnhum.2021.665319
Received: 07 February 2021; Accepted: 01 June 2021;
Published: 05 July 2021.
Edited by:
Amy Belfi, Missouri University of Science and Technology, United StatesCopyright © 2021 Vaccaro, Scott, Gimbel and Kaplan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jonas T. Kaplan, anRrYXBsYW5AdXNjLmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.