 Andrea Ravignani
Andrea Ravignani Henkjan Honing
Henkjan Honing Sonja A. Kotz
Sonja A. Kotz- 1Veterinary and Research Department, Sealcentre Pieterburen, Pieterburen, Netherlands
- 2Language and Cognition Department, Max Planck Institute for Psycholinguistics, Nijmegen, Netherlands
- 3Artificial Intelligence Lab, Vrije Universiteit Brussel, Brussels, Belgium
- 4Music Cognition Group, Amsterdam Brain and Cognition, Institute for Logic, Language, and Computation, University of Amsterdam, Amsterdam, Netherlands
- 5Basic and Applied NeuroDynamics Lab, Faculty of Psychology and Neuroscience, Department of Neuropsychology and Psychopharmacology, Maastricht University, Maastricht, Netherlands
- 6Department of Neuropsychology, Max-Planck Institute for Human Cognitive and Brain Sciences, Leipzig, Germany
Editorial on the Research Topic
The Evolution of Rhythm Cognition: Timing in Music and Speech
Overview of This Paper
This editorial serves a number of purposes. First, it aims at summarizing and discussing 33 accepted contributions to the special issue “The evolution of rhythm cognition: Timing in music and speech.” The major focus of the issue is the cognitive neuroscience of rhythm, intended as a neurobehavioral trait undergoing an evolutionary process. Second, this editorial provides the interested reader with a guide to navigate the interdisciplinary contributions to this special issue. For this purpose, we have compiled Table 1, where methods, topics, and study species are summarized and related across contributions. Third, we also briefly highlight research relevant to the evolution of rhythm that has appeared in other journals while this special issue was compiled. Altogether, this editorial constitutes a summary of rhythm research in music and speech spanning two years, from mid-2015 until mid-2017.
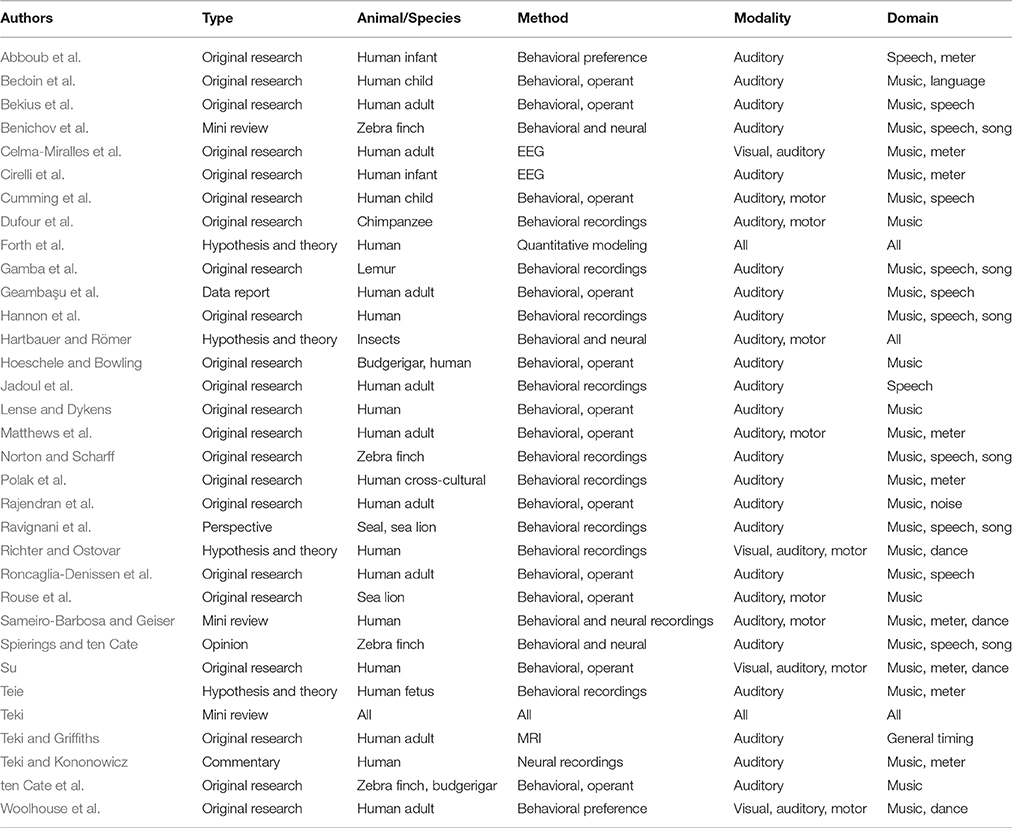
Table 1. Papers in this issue categorized along methodological and conceptual dimensions.
Timing in Music and Speech
Human speech and music differ in many respects but also share similarities. One of the main similarities lies in their temporal nature. In fact, both music and speech:
(i) develop over time and have a temporal dimension crucial to physically characterize music and speech,
(ii) rely on timing as one of their most conspicuous perceptual dimensions,
(iii) can be tokenized as sequences of temporal intervals, which are perceived as a “rhythm,”
(iv) are composed by temporal intervals that possibly differ in duration and acoustic marking by different spectral properties, generating metrical expectations.
Humans seem to be particularly rhythmic animals. Decades of research have shown that human brains are tuned-in to the fine degrees of rhythmic information in music and speech (Bolton, 1894; Fraisse, 1981, 1982, 1984; Longuet-Higgins and Lee, 1982, 1984; Povel, 1984, 1985; Essens and Povel, 1985; Povel and Essens, 1985; Shmulevich and Povel, 2000). This human propensity to perceive, produce, and process rhythm is increasingly well understood, though its evolutionary origins remain a bit of a mystery. Let's compare this to what we know about the eye. This organ has evolved in animals as a complex photoreceptor to supply the need of sensing light (Fitch, 2015a). In addition, color vision in humans and many other species appears particularly useful to assess the ripeness of food or the quality of a potential mate, hence conferring an evolutionary advantage. Unfortunately, we are still far from providing similar answers for a complex neurobehavioral trait such as rhythm. However, we firmly believe that rhythm needs to be anchored in an evolutionary perspective.
A number of critical questions spurred this special issue. When did the sensitivity for rhythm arise in human evolutionary history? How did rhythm cognition develop in human evolution? How does this evolutionary path relate to rhythm ontogeny? What is the biological function of rhythm in the millisecond to second range? Do environmental rhythms affect the evolution of brain rhythms, and how? Do speech and music share rhythm-specific neural circuits and cognitive modules? Are these circuits shared with other domains and even across species?
Rhythm: A Multidisciplinary Field
In general, the long-time scales involved in evolutionary processes prevent direct observation. Sometimes the evolutionary dynamics of simple traits can be replicated in the lab: For instance, the evolution of learning in fruit flies can be directly observed (Mery and Kawecki, 2002). Instead, the evolution of human behavior and neurobiology requires a more indirect scientific method. This is why understanding the cognitive neuroscience of rhythm and its evolution calls for a tight integration of different perspectives (Fitch, 2015b; Honing et al., 2015; Ravignani, 2017a). In particular, complementary approaches include but are not limited to:
(1) developmental studies of rhythm that are useful in understanding whether rhythm perception and production involve critical acquisition periods, or instead result mostly from enculturation during the whole lifespan (Hannon and Trehub, 2005),
(2) comparative and cross-cultural studies of rhythm that serve to explain whether musical enculturation or exposure to specific languages can affect which specific rhythmic patterns can be produced/perceived and how frequently (Greenberg et al., 1978; Rzeszutek et al., 2012),
(3) comparisons of rhythm processing in music and speech, at both behavioral and neural levels (Peretz et al., 2015) that help understanding whether common music-speech networks exist and similar behavioral patterns can be observed when humans engage in music and speech production,
(4) evidence and comparison of rhythm processing across modalities and domains that are used to understand whether, for instance, metrical expectation in speech is strictly bound to the speech domain or instead recruits the same capacities for metricality available in music, or even in dance and vision (Iversen et al., 2015),
(5) studies of rhythm in interaction and context (Yu and Tomonaga, 2015), explaining how social, affective, and other factors affect the emergence of rhythmic patterns,
(6) archaeological findings trying to reconstruct rhythm-related behavior and cognition in our early hominid ancestors (Morley, 2003),
(7) mathematical and computational models (e.g., connectionist, symbolic) of the mechanisms underlying perception and production of rhythmic behavior (Desain and Honing, 1989, 1991, 2003),
(8) mathematical and computational models of rhythmic capacities as evolved behaviors (Miranda et al., 2003) in line with a long tradition in evolutionary and theoretical biology,
(9) evidence of spontaneous rhythmic behavior in other animals (Fuhrmann et al., 2014; Ravignani et al., 2014a) showing how similar rhythmic traits can evolve via similar pressures in phylogenetically distant species,
(10) controlled experiments in non-human animals (Cook et al., 2013) probing the potential for producing/perceiving rhythm (even though these are not usually part of these species' natural behavior); these experiments can show the existence of basic, evolutionary conserved cognitive processes that may have been exapted in humans for rhythmic purposes.
The cognitive and neurobiological bases of rhythm are increasingly well understood (Honing et al., 2015; Merchant et al., 2015). While researchers in many fields are interested in rhythm, there is little awareness of how related and potentially converging their research strands are. This special issue builds a bridge across a large number of scientific disciplines; the focus lies in the cognitive neurosciences of rhythm, conceptualizing rhythm as a neurobehavioral trait undergoing an evolutionary process.
Development Evidence
A good proportion of the papers in this issue deals with developmental aspects of rhythm (Abboub et al.; Bedoin et al.; Cirelli et al.; Cumming et al.; Hannon et al.; Lense and Dykens; Teie). Among those, one theoretical contribution raised the intriguing possibility that an individual's fetal environment may already affect his future rhythmic repertoire (Teie). Two contributions tested rhythmic abilities in infants ranging from 7 to 15 months of age, with a focus either on beat perception (Cirelli et al.) or speech meter and grouping (Abboub et al.). A corpus-based approach investigated rhythmic regularities in children's songs and finds a connection between rhythms in song and non-song speech features (Hannon et al.). Two contributions focused on the interaction between musical beat and language in 9 year old children with specific language impairments (Bedoin et al.; Cumming et al.). Finally, Lense and Dykens tracked rhythmic abilities over the lifespan in a sample of 74 children and adults affected by Williams syndrome.
Cross-Cultural Evidence
The study of rhythm in speech and music is increasingly adopting a global, cross-cultural perspective (Abboub et al.; Bekius et al.; Polak et al.; Roncaglia-Denissen et al.; Teie). The field seems to be expanding beyond learners of English as first language or musically-enculturated Westerners. In speech, three contributions explored the relationship between different languages or learning a foreign language, and rhythmic capacities (Abboub et al.; Bekius et al.; Hannon et al.; Roncaglia-Denissen et al.). In music, the focus is on biologically-driven rhythmic universals (Teie) and experiments involving cross-cultural comparisons (Polak et al.).
EEG and Frequency Tagging
Along a methodological dimension, empirical papers adopted three alternative approaches: corpus analyses, behavioral experiments or brain imaging/electrophysiology. It is interesting to note that all experimental papers in this issue that employed EEG also adopted a frequency-tagging approach (Celma-Miralles et al.; Cirelli et al.; Teki and Kononowicz), rather than a grand-average ERP method (but see Henry et al., 2017 for a note of caution).
Music, Speech, and Syntax
The relationship between music, language, and speech continues being of great interest in the scientific community. This continued interest is found also in the papers in this issue (Bedoin et al.; Bekius et al.; Cumming et al.; Geambaşu et al.; Norton and Scharff; Ravignani et al.; Roncaglia-Denissen et al.). In particular, one paper investigated how beat keeping and phonological patterning are related (Bekius et al.). Another study focused on recursion, a topic of great debate in linguistics and showed how human adults are sensitive to recursive structures in rhythmic patterns (Geambaşu et al.).
Modality
Modality-specificity and domain-specificity were also explored in this issue (Celma-Miralles et al.; Matthews et al.; Richter and Ostovar; Su). Findings about rhythm in vision (Celma-Miralles et al.; Su) and movement (Su) suggested that some circuits for rhythmic timing may coincide across modalities.
Rhythm in Interaction
Complementary to meticulously controlled individual experiments, rhythm can be investigated by taking a more holistic approach, and probing rhythmic behaviors in interaction (Benichov et al.; Gamba et al.; Hartbauer and Römer; Lense and Dykens; Richter and Ostovar; Woolhouse et al.). Vocal coordination behavior in groups of primates (Gamba et al.), songbirds (Benichov et al.), and insects (Hartbauer and Römer) can offer insights for human interactional timing. Connections between internal rhythms and group behaviors can be investigated in healthy adults (Richter and Ostovar; Woolhouse et al.) and individuals with specific syndromes affecting musicality and sociality (Lense and Dykens).
Dance
Three papers discussed rhythm from the perspective of dance (Richter and Ostovar; Su; Woolhouse et al.). Rhythm and dance should be thought as a tightly connected pair (Richter and Ostovar), which can be empirically investigated (Su) and shed light on other aspects of cognition (Woolhouse et al.).
Quantitative Models
Two papers in the issue were devoted to mathematical and computational modeling (Forth et al.; Jadoul et al.). These approaches are complementary. On the one hand, rhythm and timing can be investigated using top-down abstract models (Forth et al.). On the other hand, different aspects of speech timing can be statistically modeled with different degrees of precisions and assumptions made (Jadoul et al.).
Animal Research
This issue also contains ample evidence on rhythm from a comparative approach (Benichov et al.; Dufour et al.; Gamba et al.; Hartbauer and Römer; Hoeschele and Bowling; Norton and Scharff; Ravignani et al.; Rouse et al.; Spierings and ten Cate; ten Cate et al.). Songbirds continue to be a particularly often-used model species in the study of rhythm (Benichov et al.; Hoeschele and Bowling; Norton and Scharff; Spierings and ten Cate; ten Cate et al.). For instance, important advances have been made by confirming how the subjective “feeling of rhythm” experienced when listening to a songbird has a quantitative, isochronous counterpart in the animal's song (Norton and Scharff). Rhythmic behaviors in two primate species were also explored in this issue. These works examined either the closest primate to humans, the chimpanzee (Dufour et al.) or one of the phylogenetically farthest group, the lemur (Gamba et al.). This suggests that some components of human rhythmicity may be due to evolutionary homology (common descent from our last common ancestor with chimpanzees) while other traits to analogy (convergent evolution in man and singing lemurs). A taxonomic group emerging as particularly promising for future rhythm research is the pinnipeds, which features harbor seals, sea lions, and walruses (Ravignani et al.; Rouse et al.).
General Timing and Others
Other papers discussed general issues related to timing and time perception (Rajendran et al.; Sameiro-Barbosa and Geiser; Teki; Teki and Griffiths). Two contributions tested general aspects of the relationship among timing, rhythm and cognitive functions (Rajendran et al.; Teki and Griffiths). A theoretical paper reviewed neural entrainment mechanisms (Sameiro-Barbosa and Geiser). Finally, Teki provided a useful overview of timing papers since 2000, ordering them by number of citations, so to identify community trends and overall research interests.
Rhythm in Other Journals Since Late 2015
Since the launch of this Frontiers Research Topic, a number of publications on rhythm have appeared in other journals. Among those, some strands are particularly relevant to research in the evolution of rhythm. Far from attempting a comprehensive overview, we mention these papers and summarize some of them below.
Evolutionary Hypotheses for Rhythm Origins
Some review papers have properly focused on the evolutionary origins of musical rhythm and animal species showing human-like rhythmic traits (Bannan, 2016; Iversen, 2016; Wilson and Cook, 2016). Bannan (2016) provided a recount of Charles Darwin's thoughts on music and how he thought human musicality may have emerged via sexual selection. Iversen (2016) summarized and compared many evolutionary hypotheses on the origins of rhythm in humans. Wilson and Cook (2016) discussed which animal species are capable of synchronizing to a beat, either spontaneously or after being trained, and how this evidence relates to evolutionary hypotheses. Some of these evolutionary hypotheses on music and rhythm have been tested via genetics (Mosing et al., 2015), behavioral experiments (Miani, 2016), electrophysiology (Bouwer et al., 2016) or animal comparative work (ten Cate et al.; van der Aa et al., 2015).
Speech Rhythm and Comparative Anatomy of Vocal Tracts
In the evolution of speech, several studies have shown how vocal tracts in non-human primates are more flexible than previously thought. Other primates' vocal tracts are capable of producing a human-like range of vowels (Fitch et al., 2016; Boë et al., 2017) and consonants (Lameira et al., 2015, 2016, 2017). The overall conclusion is that the complexity of human speech, including its rhythmical nuances, must have neural, rather than morphological, bases (Ravignani et al., 2014b; Fitch et al., 2016; Belyk and Brown, 2017).
The Social Roots of Rhythm
The relationship between rhythm and sociality has seen a steady increase in research and has probably been the most investigated topic over the last 2 years (Large and Gray, 2015; Yu and Tomonaga, 2015; Ellamil et al., 2016; Gebauer et al., 2016; Greenfield et al., 2016; Moore et al., 2016; Reddish et al., 2016; Rennung and Göritz, 2016; Schirmer et al., 2016; Tunçgenç and Cohen, 2016; Wallot et al., 2016; Bishop and Goebl, 2017; Chang et al., 2017; Cirelli et al., 2017; Hannon et al., 2017; Knight et al., 2017; Mogan et al., 2017; Murphy and Schul, 2017; Rorato et al., 2017; Myers et al.). Common foci are the relationship between synchronization and prosociality (Gebauer et al., 2016; Reddish et al., 2016; Rennung and Göritz, 2016; Tunçgenç and Cohen, 2016; Cirelli et al., 2017), and different forms of rhythmic behaviors in interaction (Large and Gray, 2015; Ravignani, 2015; Yu and Tomonaga, 2015; Ellamil et al., 2016; Gebauer et al., 2016; Greenfield et al., 2016; Moore et al., 2016; Schirmer et al., 2016; Wallot et al., 2016; Murphy and Schul, 2017).
Speech, Music, and Prosody
Another topic of broad interest centers on the relationship between speech, prosody, and music (Toro and Nespor, 2015; Vanden Bosch der Nederlanden et al., 2015; Chang et al., 2016; Filippi, 2016; Frühholz et al., 2016; Kotz and Schwartze, 2016; Schwartze and Kotz, 2016; Weidema et al., 2016; Carr et al., 2017; Ding et al., 2017; Spierings et al., 2017; Toro and Hoeschele, 2017). An intriguing hypothesis is that speech prosody may be the “missing link” between music and language (Filippi, 2016) or that music and language may be preceded by musical prosody (Fitch, 2013; Honing, 2017). This may inform us on early proto-musical and proto-linguistic behaviors in our early hominid ancestors.
Cultural Evolution and Cognitive Biases
Rhythm seems to be slowly overcoming the classical nature-nurture debate that actually is built on a false dichotomy. Along these lines, recent research has focused on the cultural evolution of musical rhythm and perceptual priors (Savage et al., 2015; Trehub, 2015; Hansen et al., 2016; Le Bomin et al., 2016; Ravignani et al., 2016; Fitch, 2017; Jacoby and McDermott, 2017). Statistical universals found in musical rhythms all over the world (Savage et al., 2015) can emerge via the combined effect of human cognitive biases and cultural transmission (Ravignani et al., 2016). Interestingly, these biases seem at least partly modulated by enculturation (Jacoby and McDermott, 2017).
The Evolution of Dance
The field of musical rhythm is increasingly expanding to encompass the scientific study of dance (Ellamil et al., 2016; Fink and Shackelford, 2017; Fitch, 2016; Laland et al., 2016; Ravignani and Cook, 2016; Su, 2016). Only in 2016, three papers have introduced conceptual frameworks for the evolutionary study of dance (Fitch, 2016; Laland et al., 2016; Ravignani and Cook, 2016). We believe the field would benefit from connecting these theoretical frameworks with recent empirical findings on dance (Ellamil et al., 2016; Su, 2016).
Timing and Time Perception
The science of timing and time perception has been a major research area in the last century. After a less active period, this field is again experiencing an increase in research efforts. A whole special issue of Current Opinion in Behavioral Sciences was recently devoted to “Time in perception and action” (Meck and Ivry, 2016). In addition, a “Timing Research Forum” was established in 2016, to spur and connect research on timing and time perception across disciplines.
Measuring Rhythm
Finally, new methods to model (van der Weij et al., 2017) and measure rhythmicity have been proposed, either quantitatively from data (Daniele, 2017; Malisz et al., 2017; Ravignani, 2017b; Ravignani and Norton, 2017) or as a test battery on human participants (Dalla Bella et al., 2016).
Final Considerations
Similarly to other fields, the study of the evolution of rhythm must build on a tight integration of experiments, theory, and modeling. Ideally, empirical observations of rhythm in music and speech are first recorded in the field. Observations are then contrasted to generate testable hypotheses. Based on these hypotheses, experiments on linguistic and musical rhythm are performed. Experimental factors and variations can encompass sensory modalities, ages, and animal species, to name a few, in order to address questions about domain-specificity, development, and evolutionary phylogeny. Finally, experimental insights should be integrated via synthetic modeling. The advantage of models is that they generate predictions that are quantitatively testable. Following these predictions, new empirical observations should be collected and compared, continuing the incremental loop of scientific investigation.
This journal issue contains novel empirical findings and state of the art reviews of hot topics in each discipline. We hope it will be useful as a reference volume on the evolution of rhythm cognition. Combining well-established findings and novel results on the evolution of rhythm, it should serve as an introductory reference for newcomers, a source of novel findings for researchers more familiar with one of the areas, and an interdisciplinary overview of progress in neighboring disciplines.
All contributions discussed so far show the many sides of rhythm. From this volume, rhythm emerges not as a monolithic concept, but as a multifaceted phenomenon for research. We hope that exciting future research will be ignited by this multifaceted display of rhythm across domains and species.
Author Contributions
All authors listed, have made substantial, direct, and intellectual contribution to the work, and approved it for publication.
Funding
AR was supported by funding from the European Union's Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No 665501 with the research Foundation Flanders (FWO) (Pegasus2 Marie Curie fellowship 12N5517N awarded to AR), a visiting fellowship in Language Evolution from the Max Planck Society (awarded to AR), and ERC grant 283435 ABACUS (awarded to Bart de Boer). HH was supported by a Distinguished Lorentz fellowship granted by the Lorentz Center for the Sciences and the Netherlands Institute for Advanced Study in the Humanities and Social Sciences (NIAS), and a Horizon grant (317-70-10) of the Netherlands Organization for Scientific Research (NWO). SK was supported by a BBSRC award (BB/M009742/1), a British Academy Skills Acquisition Award (SQ140010), and a Marie Skłodowska Curie Action (MSCA: 707727) under the Seventh Framework Programme (FP7).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to the editors of Frontiers in Human Neuroscience, Frontiers in Psychology - Auditory Cognitive Neuroscience, Frontiers in Neuroscience - Auditory Cognitive Neuroscience (Isabelle Peretz, Robert J. Zatorre, Hauke R. Heekeren, and Srikantan S. Nagarajan), all members of the editorial teams, and the guest editors of some of the papers in this issue: Virginia Penhune, Mikhail Lebedev, Huan Luo, Angela D. Friederici. We would like to thank all referees who have offered their time and expertise in reviewing the papers.
References
Bannan, N. (2016). Darwin, music and evolution: new insights from family correspondence on The Descent of Man. Mus. Sci. 21, 3–25. doi: 10.1177/1029864916631794
Belyk, M., and Brown, S. (2017). The origins of the vocal brain in humans. Neurosci. Biobehav. Rev. 77, 177–193. doi: 10.1016/j.neubiorev.2017.03.014
Bishop, L., and Goebl, W. (2017). Beating time: How ensemble musicians' cueing gestures communicate beat position and tempo. Psychol. Music doi: 10.1177/0305735617702971
Boë, L.-J., Berthommier, F., Legou, T., Captier, G., Kemp, C., Sawallis, T. R., et al. (2017). Evidence of a vocalic proto-system in the baboon (Papio papio) suggests pre-hominin speech precursors. PLoS ONE 12:e0169321. doi: 10.1371/journal.pone.0169321
Bouwer, F. L., Werner, C. M., Knetemann, M., and Honing, H. (2016). Disentangling beat perception from statistical learning using ERPs: the role of attention and musical training. Neuropsychologia 85, 80–90. doi: 10.1016/j.neuropsychologia.2016.02.018
Carr, K. W., Fitzroy, A. B., Tierney, A., White-Schwoch, T., and Kraus, N. (2017). Incorporation of feedback during beat synchronization is an index of neural maturation and reading skills. Brain Lang. 164, 43–52. doi: 10.1016/j.bandl.2016.09.005
Chang, A., Livingstone, S. R., Bosnyak, D. J., and Trainor, L. J. (2017). Body sway reflects leadership in joint music performance. Proc. Natl. Acad. Sci. U.S.A. 114, E4134–E4141. doi: 10.1073/pnas.1617657114
Chang, S.-E., Chow, H. M., Wieland, E. A., and McAuley, J. D. (2016). Relation between functional connectivity and rhythm discrimination in children who do and do not stutter. NeuroImage 12, 442–450. doi: 10.1016/j.nicl.2016.08.021
Cirelli, L. K., Wan, S. J., Spinelli, C., and Trainor, L. J. (2017). Effects of Interpersonal Movement Synchrony on Infant Helping Behaviors. Music Percept. 34, 319–326. doi: 10.1525/mp.2017.34.3.319
Cook, P., Rouse, A., Wilson, M., and Reichmuth, C. J. (2013). A california sea lion (Zalophus californianus) can keep the beat: motor entrainment to rhythmic auditory stimuli in a non vocal mimic. J. Comp. Psychol. 127, 1–16. doi: 10.1037/a0032345
Dalla Bella, S., Farrugia, N., Benoit, C.-E., Begel, V., Verga, L., Harding, E., et al. (2016). BAASTA: Battery for the assessment of auditory sensorimotor and timing abilities. Behav. Res. Methods 49, 1128–1145. doi: 10.3758/s13428-016-0773-6
Daniele, J. R. (2017). The “Rhythmic Fingerprint”: an extension of the nPVI to quantify rhythmic influence. Emp. Musicol. Rev. 11, 243–260. doi: 10.18061/emr.v11i2.5426
Desain, P., and Honing, H. (1989). The quantization of musical time: a connectionist approach. Comput. Music J. 13, 56–66. doi: 10.2307/3680012
Desain, P., and Honing, H. (1991). “Quantization of musical time: a connectionist approach,” in Music and Connectionism, eds P. M. Todd and G. Loy (Cambridge, MA: MIT Press), 150–167.
Desain, P., and Honing, H. (2003). The formation of rhythmic categories and metric priming. Perception 32, 341–365. doi: 10.1068/p3370
Ding, N., Patel, A. D., Chen, L., Butler, H., Luo, C., and Poeppel, D. (2017). Temporal modulations in speech and music. Neurosci. Biobehav. Rev. doi: 10.1016/j.neubiorev.2017.02.011. [Epub ahead of print].
Ellamil, M., Berson, J., Wong, J., Buckley, L., and Margulies, D. S. (2016). One in the dance: musical correlates of group synchrony in a real-world club environment. PLoS ONE 11:e0164783. doi: 10.1371/journal.pone.0164783
Essens, P. J., and Povel, D.-J. (1985). Metrical and nonmetrical representations of temporal patterns. Attent. Percept. Psychophys. 37, 1–7. doi: 10.3758/BF03207132
Filippi, P. (2016). Emotional and interactional prosody across animal communication systems: a comparative approach to the emergence of language. Front. Psychol. 7:1393. doi: 10.3389/fpsyg.2016.01393
Fink, B., and Shackelford, T. K. (2017). Why did dance evolve? a comment on Laland, Wilkins, and Clayton (2016). Evol. Psychol. Sci. 3, 147–148. doi: 10.1007/s40806-016-0075-3
Fitch, W. T. (2013). “Musical protolanguage: Darwin's theory of language evolution revisited,” in Birdsong, Speech and Language: Exploring the Evolution of Mind and Brain, eds J. J. Bolhuis and M. B. H. Everaert (Cambridge, MA: MIT Press).
Fitch, W. T. (2015a). “The biology and evolution of musical rhythm: an update,” in Structures in the Mind: Essays on Language, Music, and Cognition in Honor of Ray Jackendoff, eds D. I. Toivonen, P. Csúri, and E. van der Zee (Cambridge, MA: MIT Press), 293–324.
Fitch, W. T. (2015b). Four principles of bio-musicology. Philos. Trans. R. Soc. B Biol. Sci. 370:20140091. doi: 10.1098/rstb.2014.0091
Fitch, W. T. (2016). Dance, music, meter and groove: a forgotten partnership. Front. Hum. Neurosci. 10:64. doi: 10.3389/fnhum.2016.00064
Fitch, W. T. (2017). Cultural evolution: Lab-cultured musical universals. Nat. Hum. Behav. 1, 1–2. doi: 10.1038/s41562-016-0018
Fitch, W. T., de Boer, B., Mathur, N., and Ghazanfar, A. A. (2016). Monkey vocal tracts are speech-ready. Sci. Adv. 2:e1600723. doi: 10.1126/sciadv.1600723
Fraisse, P. (1981). “Multisensory aspects of rhythm,” in Intersensory Perception and Sensory Integration (New York, NY: Plenum Press; Springer), 217–248. doi: 10.1007/978-1-4615-9197-9_7
Fraisse, P. (1982). Rhythm and tempo. Psychol. Music 1, 149–180. doi: 10.1016/B978-0-12-213562-0.50010-3
Fraisse, P. (1984). Perception and estimation of time. Annu. Rev. Psychol. 35, 1–37. doi: 10.1146/annurev.ps.35.020184.000245
Frühholz, S., Trost, W., and Kotz, S. A. (2016). The sound of emotions—Towards a unifying neural network perspective of affective sound processing. Neurosci. Biobehav. Rev. 68, 96–110. doi: 10.1016/j.neubiorev.2016.05.002
Fuhrmann, D., Ravignani, A., Marshall-Pescini, S., and Whiten, A. (2014). Synchrony and motor mimicking in chimpanzee observational learning. Sci. Rep. 4:5283. doi: 10.1038/srep05283
Gebauer, L., Witek, M., Hansen, N., Thomas, J., Konvalinka, I., and Vuust, P. (2016). Oxytocin improves synchronisation in leader-follower interaction. Sci. Rep. 6:38416. doi: 10.1038/srep38416
Greenberg, J. H., Ferguson, C. A., and Moravcsik, E. A. (1978). Universals of Human Language: Phonology. Stanford University Press.
Greenfield, M. D., Esquer-Garrigos, Y., Streiff, R., and Party, V. (2016). Animal choruses emerge from receiver psychology. Sci. Rep. 6:34369. doi: 10.1038/srep34369
Hannon, E. E., Schachner, A., and Nave-Blodgett, J. E. (2017). Babies know bad dancing when they see it: older but not younger infants discriminate between synchronous and asynchronous audiovisual musical displays. J. Exp. Child Psychol. 159, 159–174. doi: 10.1016/j.jecp.2017.01.006
Hannon, E. E., and Trehub, S. E. (2005). Tuning in to musical rhythms: Infants learn more readily than adults. Proc. Natl. Acad. Sci. U.S.A. 102, 12639–12643. doi: 10.1073/pnas.0504254102
Hansen, N. C., Sadakata, M., and Pearce, M. (2016). Nonlinear changes in the rhythm of european art music. Music Percept. 33, 414–431. doi: 10.1525/mp.2016.33.4.414
Henry, M. J., Herrmann, B., and Grahn, J. A. (2017). What can we learn about beat perception by comparing brain signals and stimulus envelopes? PLoS ONE 12:e0172454. doi: 10.1371/journal.pone.0172454
Honing, H. (2017). “Musicality as an upbeat to music: introduction and research agenda,” in The Origins of Musicality, ed H. Honing (Cambridge, MA: MIT Press).
Honing, H., ten Cate, C., Peretz, I., and Trehub, S. E. (2015). Without it no music: cognition, biology, and evolution of musicality. Philos. Trans. R. Soc. B Biol. Sci. 370:20140088. doi: 10.1098/rstb.2014.0088
Iversen, J. R. (2016). “In the beginning was the beat: Evolutionary origins of musical rhythm in humans,” in The Cambridge Companion to Percussion, ed R. Hartenberger (Cambridge: Cambridge University Press). doi: 10.1017/CBO9781316145074.022
Iversen, J. R., Patel, A. D., Nicodemus, B., and Emmorey, K. (2015). Synchronization to auditory and visual rhythms in hearing and deaf individuals. Cognition 134, 232–244. doi: 10.1016/j.cognition.2014.10.018
Jacoby, N., and McDermott, J. H. (2017). Integer ratio priors on musical rhythm revealed cross-culturally by iterated reproduction. Curr. Biol. 27, 359–370. doi: 10.1016/j.cub.2016.12.031
Knight, S., Spiro, N., and Cross, I. (2017). Look, listen and learn: Exploring effects of passive entrainment on social judgements of observed others. Psychol. Music 45, 99–115. doi: 10.1177/0305735616648008
Kotz, S. A., and Schwartze, M. (2016). “Motor-Timing and Sequencing in Speech Production: A General-Purpose Framework,” in Neurobiology of Language, eds G. Hickok and S. L. Small (San Diego: Academic Press), 717–724. doi: 10.1016/B978-0-12-407794-2.00057-2
Laland, K., Wilkins, C., and Clayton, N. (2016). The evolution of dance. Curr. Biol. 26, R5–R9. doi: 10.1016/j.cub.2015.11.031
Lameira, A. R., Hardus, M. E., Bartlett, A. M., Shumaker, R. W., Wich, S. A., and Menken, S. B. (2015). Speech-Like Rhythm in a Voiced and Voiceless Orangutan Call. PLoS ONE 10:e116136. doi: 10.1371/journal.pone.0116136
Lameira, A. R., Hardus, M. E., Mielke, A., Wich, S. A., and Shumaker, R. W. (2016). Vocal fold control beyond the species-specific repertoire in an orang-utan. Sci. Rep. 6:30315. doi: 10.1038/srep30315
Lameira, A. R., Vicente, R., Alexandre, A., Campbell-Smith, G., Knott, C., Wich, S., et al. (2017). Proto-consonants were information-dense via identical bioacoustic tags to proto-vowels. Nat. Hum. Behav. 1:0044. doi: 10.1038/s41562-017-0044
Large, E. W., and Gray, P. M. (2015). Spontaneous Tempo and Rhythmic Entrainment in a Bonobo (Pan Paniscus). J. Comp. Psychol. 129, 317–328. doi: 10.1037/com0000011
Le Bomin, S., Lecointre, G., and Heyer, E. (2016). The evolution of musical diversity: the key role of vertical transmission. PLoS ONE 11:e0151570. doi: 10.1371/journal.pone.0151570
Longuet-Higgins, H. C., and Lee, C. S. (1982). The perception of musical rhythms. Perception 11, 115–128. doi: 10.1068/p110115
Longuet-Higgins, H. C., and Lee, C. S. (1984). The rhythmic interpretation of monophonic music. Music Percept. 1, 424–441. doi: 10.2307/40285271
Malisz, Z., O'Dell, M., Nieminen, T., and Wagner, P. (2017). Perspectives on speech timing: coupled oscillator modeling of polish and finnish. Phonetica 73, 229–255. doi: 10.1159/000450829
Meck, W. H., and Ivry, R. B. (2016). Editorial overview: time in perception and action. Curr. Opin. Behav. Sci. 8, vi–x. doi: 10.1016/j.cobeha.2016.03.001
Merchant, H., Grahn, J., Trainor, L., Rohrmeier, M., and Fitch, W. T. (2015). Finding the beat: a neural perspective across humans and non-human primates. Phil. Trans. R. Soc. B 370:20140093. doi: 10.1098/rstb.2014.0093
Mery, F., and Kawecki, T. J. (2002). Experimental evolution of learning ability in fruit flies. Proc. Natl. Acad. Sci. U.S.A. 99, 14274–14279. doi: 10.1073/pnas.222371199
Miani, A. (2016). Sexual arousal and rhythmic synchronization: a possible effect of vasopressin. Med. Hypotheses 93, 122–125. doi: 10.1016/j.mehy.2016.05.030
Miranda, E. R., Kirby, S., and Todd, P. (2003). On computational models of the evolution of music: from the origins of musical taste to the emergence of grammars. Contemp. Music Rev. 22, 91–111. doi: 10.1080/0749446032000150915
Mogan, R., Fischer, R., and Bulbulia, J. A. (2017). To be in synchrony or not? A meta-analysis of synchrony's effects on behavior, perception, cognition and affect. J. Exp. Soc. Psychol. 72, 13–20. doi: 10.1016/j.jesp.2017.03.009
Moore, R. K., Marxer, R., and Thill, S. (2016). Vocal interactivity in-and-between humans, animals, and robots. Front. Robot. 3:61. doi: 10.3389/frobt.2016.00061
Morley, I. (2003). The Evolutionary Origins and Archaeology of Music. Cambridge, UK: Darwin College, Cambridge University.
Mosing, M. A., Verweij, K. J., Madison, G., Pedersen, N. L., Zietsch, B. P., and Ullén, F. (2015). Did sexual selection shape human music? Testing predictions from the sexual selection hypothesis of music evolution using a large genetically informative sample of over 10,000 twins. Evol. Hum. Behav. 36, 359–366. doi: 10.1016/j.evolhumbehav.2015.02.004
Murphy, M. A., and Schul, J. (2017). Does leadership indicate male quality in Neoconocephalus katydids? Behav. Ecol. Sociobiol. 71:22. doi: 10.1007/s00265-016-2253-5
Myers, A. J., Herzing, D. L. and Bjorklund, D. F. (2017) Synchrony during aggression in adult male Atlantic spotted dolphins (Stenella frontalis). Acta Ethol. 20:175. doi: 10.1007/s10211-017-0262-7
Peretz, I., Vuvan, D., Lagrois, M-É., and Armony, J. L. (2015). Neural overlap in processing music and speech. Philos. Trans. R. Soc. Lond. B Biol. Sci. 370:20140090. doi: 10.1098/rstb.2014.0090
Povel, D.-J. (1984). A theoretical framework for rhythm perception. Psychol. Res. 45, 315–337. doi: 10.1007/BF00309709
Povel, D.-J. (1985). “Time, rhythms and tension: in search of the determinants of rhythmicity,” in Time, Mind, and Behavior (Berlin; Heidelberg: Springer), 215–225.
Povel, D.-J., and Essens, P. (1985). Perception of temporal patterns. Music Percept. 2, 411–440. doi: 10.2307/40285311
Ravignani, A. (2015). Evolving perceptual biases for antisynchrony: a form of temporal coordination beyond synchrony. Front. Neurosci. 9:339. doi: 10.3389/fnins.2015.00339
Ravignani, A. (2017a). Interdisciplinary debate: agree on definitions of synchrony. Nature 545:158. doi: 10.1038/545158c
Ravignani, A. (2017b). Visualizing and interpreting rhythmic patterns using phase space plots. Music Percept. 34, 557–568. doi: 10.1525/mp.2017.34.5.557
Ravignani, A., Bowling, D. L., and Fitch, W. T. (2014a). Chorusing, synchrony and the evolutionary functions of rhythm. Front. Psychol. 5:1118. doi: 10.3389/fpsyg.2014.01118
Ravignani, A., and Cook, P. (2016). The evolutionary biology of dance without frills. Curr. Biol. 26, R878–R879. doi: 10.1016/j.cub.2016.07.076
Ravignani, A., Delgado, T., and Kirby, S. (2016). Musical evolution in the lab exhibits rhythmic universals. Nat. Hum. Behav. 1:0007. doi: 10.1038/s41562-016-0007
Ravignani, A., Martins, M., and Fitch, W. (2014b). Vocal learning, prosody, and basal ganglia: don't underestimate their complexity. Behav. Brain Sci. 37, 570–571. doi: 10.1017/S0140525X13004184
Ravignani, A., and Norton, P. (2017). Measuring rhythmic complexity: a primer to quantify and compare temporal structure in speech, movement, and animal vocalizations. J. Lang. Evol. doi: 10.1093/jole/lzx002
Reddish, P., Tong, E. M., Jong, J., Lanman, J. A., and Whitehouse, H. (2016). Collective synchrony increases prosociality towards non-performers and outgroup members. Br. J. Soc. Psychol. 55, 722–738. doi: 10.1111/bjso.12165
Rennung, M., and Göritz, A. S. (2016). Prosocial consequences of interpersonal synchrony. Zeitschrift für Psychologie 224, 168–189. doi: 10.1027/2151-2604/a000252
Rorato, A. C., Araujo, S. B., Perez, D. M., and Pie, M. R. (2017). Social cues affect synchronization of male waving displays in a fiddler crab (Crustacea: Ocypodidae). Anim. Behav. 126, 293–300. doi: 10.1016/j.anbehav.2017.02.014
Rzeszutek, T., Savage, P. E., and Brown, S. (2012). The structure of cross-cultural musical diversity. Proc. R. Soc. Lond. B Biol. Sci. 279, 1606–1612. doi: 10.1098/rspb.2011.1750
Savage, P. E., Brown, S., Sakai, E., and Currie, T. E. (2015). Statistical universals reveal the structures and functions of human music. Proc. Natl. Acad. Sci. U.S.A. 112, 8987–8992. doi: 10.1073/pnas.1414495112
Schirmer, A., Meck, W. H., and Penney, T. B. (2016). The socio-temporal brain: Connecting people in time. Trends Cogn. Sci. 20, 760–772. doi: 10.1016/j.tics.2016.08.002
Schwartze, M., and Kotz, S. A. (2016). Contributions of cerebellar event-based temporal processing and preparatory function to speech perception. Brain Lang. 161, 28–32. doi: 10.1016/j.bandl.2015.08.005
Shmulevich, I., and Povel, D.-J. (2000). Measures of temporal pattern complexity. J. New Music Res. 29, 61–69. doi: 10.1076/0929-8215(200003)29:01;1-P;FT061
Spierings, M., Hubert, J., and ten Cate, C. (2017). Selective auditory grouping by zebra finches: testing the iambic–trochaic law. Anim. Cogn. doi: 10.1007/s10071-017-1089-3
Su, Y.-H. (2016). Visual enhancement of illusory phenomenal accents in non-isochronous auditory rhythms. PLoS ONE 11:e0166880. doi: 10.1371/journal.pone.0166880
Toro, J. M., and Hoeschele, M. (2017). Generalizing prosodic patterns by a non-vocal learning mammal. Anim. Cogn. 20:179. doi: 10.1007/s10071-016-1036-8
Toro, J. M., and Nespor, M. (2015). Experience-dependent emergence of a grouping bias. Biol. Lett. 11:20150374. doi: 10.1098/rsbl.2015.0374
Trehub, S. E. (2015). Cross-cultural convergence of musical features. Proc. Natl. Acad. Sci. U.S.A. 112, 8809–8810. doi: 10.1073/pnas.1510724112
Tunçgenç, B., and Cohen, E. (2016). Interpersonal movement synchrony facilitates pro-social behavior in children's peer-play. Dev. Sci. doi: 10.1111/desc.12505. [Epub ahead of print].
van der Aa, J., Honing, H., and ten Cate, C. (2015). The perception of regularity in an isochronous stimulus in zebra finches (Taeniopygia guttata) and humans. Behav. Process. 115, 37–45. doi: 10.1016/j.beproc.2015.02.018
van der Weij, B., Pearce, M., and Honing, H. (2017). A probabilistic model of meter perception: Simulating enculturation. Front. Psychol. 8:824. doi: 10.3389/fpsyg.2017.00824
Vanden Bosch der Nederlanden, C. M., Hannon, E. E., and Snyder, J. S. (2015). Finding the music of speech: Musical knowledge influences pitch processing in speech. Cognition 143, 135–140. doi: 10.1016/j.cognition.2015.06.015
Wallot, S., Mitkidis, P., McGraw, J. J., and Roepstorff, A. (2016). Beyond synchrony: joint action in a complex production task reveals beneficial effects of decreased interpersonal synchrony. PLoS ONE 11:e0168306. doi: 10.1371/journal.pone.0168306
Weidema, J. L., Roncaglia-Denissen, M., and Honing, H. (2016). Top–down modulation on the perception and categorization of identical pitch contours in speech and music. Front. Psychol. 7:817. doi: 10.3389/fpsyg.2016.00817
Wilson, M., and Cook, P. F. (2016). Rhythmic entrainment: why humans want to, fireflies can't help it, pet birds try, and sea lions have to be bribed. Psychon. Bull. Rev. 23:1647. doi: 10.3758/s13423-016-1013-x
Keywords: rhythm, meter, synchrony, interval timing, time perception, beat perception, evolution of speech, evolution of cognition
Citation: Ravignani A, Honing H and Kotz SA (2017) Editorial: The Evolution of Rhythm Cognition: Timing in Music and Speech. Front. Hum. Neurosci. 11:303. doi: 10.3389/fnhum.2017.00303
Received: 29 April 2017; Accepted: 26 May 2017;
Published: 13 June 2017.
Edited and reviewed by: Carol Seger, Colorado State University, United States
Copyright © 2017 Ravignani, Honing and Kotz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrea Ravignani, YW5kcmVhLnJhdmlnbmFuaUBnbWFpbC5jb20=
Henkjan Honing, aG9uaW5nQHV2YS5ubA==
Sonja A. Kotz, c29uamEua290ekBtYWFzdHJpY2h0dW5pdmVyc2l0eS5ubA==