 William A. Cunningham1,2,3*
William A. Cunningham1,2,3* Jay J. Van Bavel4
Jay J. Van Bavel4 Nathan L. Arbuckle5
Nathan L. Arbuckle5 Dominic J. Packer6
Dominic J. Packer6 Ashley S. Waggoner7
Ashley S. Waggoner7- 1Department of Psychology, University of Toronto, Toronto, ON, Canada
- 2Rotman School of Management, University of Toronto, Toronto, ON, Canada
- 3Department of Psychology, The Ohio State University, Columbus, OH, USA
- 4Department of Psychology, New York University, New York, NY, USA
- 5Mind Research Network, Albuquerque, NM, USA
- 6Department of Psychology, Lehigh University, Bethlehem, PA, USA
- 7Department of Psychology, Indiana University, Bloomington, IN, USA
Research on person categorization suggests that people automatically and inflexibly categorize others according to group memberships, such as race. Consistent with this view, research using electroencephalography (EEG) has found that White participants tend to show an early difference in processing Black versus White faces. Yet, new research has shown that these ostensibly automatic biases may not be as inevitable as once thought and that motivational influences may be able to eliminate these biases. It is unclear, however, whether motivational influences shape the initial biases or whether these biases can only be modulated by later, controlled processes. Using EEG to examine the time course of biased processing, we manipulated approach and avoidance motivational states by having participants pull or push a joystick, respectively, while viewing White or Black faces. Consistent with previous work on own-race bias, we observed a greater P100 response to White than Black faces; however, this racial bias was attenuated in the approach condition. These data suggest that rapid social perception may be flexible and can be modulated by motivational states.
Introduction
People often perceive others according to their race, gender, or other social category membership (Brewer, 1988; Fiske and Neuberg, 1990). This process of social categorization provides an efficient way to understand others and guides the direction of limited attentional and cognitive resources. In the past few decades, social psychologists have found extensive evidence that social categorization can occur rapidly and without intention, effort, or conscious control, triggering stereotypes (Devine, 1989), prejudice (Fazio et al., 1995), and ultimately, discrimination (Dovidio et al., 1997). Several dual-process models of person perception have proposed that processing others according to social category membership is the initial stage in person perception, and that only sufficiently motivated perceivers individuate targets or correct for initial categorical judgments in a later stage (e.g., Brewer, 1988; Devine, 1989; Fiske and Neuberg, 1990). However, while there is evidence that social categories influence the earliest phases of social perception, others have argued that the initial influence of social categories may not be inevitable (see Van Bavel and Cunningham, 2011 for a discussion). The current paper utilizes electroencephalography (EEG) to examine the malleability of early perceptual processes in social categorization during the first few 100 ms of face perception.
Several recent studies using event-related potentials (ERPs), which offer precise information about the timing of different cognitive processes as they unfold online, have shown that social categories can influence perceptual processing very quickly (Ito and Cacioppo, 2000; Smith et al., 2003; Ito et al., 2007). People differentially process own-race and other-race faces within a few 100 ms of stimulus presentation (see Ito and Bartholow, 2009 for a review). For instance, target race can modulate ERPs to faces as early as 122 ms after face onset (Ito and Urland, 2003). Moreover, these racial biases in perceptual processing persist even when participants attend to another dimension of social categorization (e.g., gender; Ito and Urland, 2003) or attempt to individuate the faces (Ito and Urland, 2005). Consistent with most dual-process models of person perception, these results have led some researchers to conclude that racial biases in “automatic attentional allocation cannot be inhibited” except under conditions of perceptual load (Ito et al., 2007, p. 410) or later during subsequent controlled processing (e.g., Devine, 1989).
In contrast, recent developments in the cognitive and neural sciences suggest that human information processing is better characterized in terms of dynamical system models, rather than dual-process models (see Dehaene et al., 2006; Cunningham and Zelazo, 2007; Van Bavel et al., in press for recent reviews). In a dynamical systems approach, what have generally been considered to be inevitable automatic responses may be influenced by top-down processes. For example, the Iterative Reprocessing (IR) Model (Cunningham and Zelazo, 2007; Cunningham et al., 2007), argues that a sharp distinction between automatic and controlled processes is not accurate (see also Freeman and Ambady, 2011). Instead, the IR Model suggests that goals and contextual features can shape the computations in brain regions involved in ostensibly automatic processes. As such, automatic responses—even those occurring within 100–200 ms of stimulus onset—may be shaped by the goals or motivations of the perceiver.
Indeed, several behavioral studies have shown that goals or contextual factors can diminish the automatic activation of attitudes and stereotypes suggesting that automatic biases in social categorization are not inevitable (see Blair, 2002 for a review). For example, in a pair of recent studies, people who were assigned to a mixed-race team had relatively positive automatic evaluations toward in-group members on a response-window priming task, regardless of their race, whereas people who were not assigned to a mixed-race team had more positive automatic evaluations toward own-race versus other-race faces (Van Bavel and Cunningham, 2009). However, because these studies only capture the behavioral consequences of perceptual and cognitive processing, it is difficult to determine the time course of these processes. It is, therefore, unclear whether these manipulations affected initial responses to social categories, altered underlying stereotypic or evaluative associations, or produced controlled processes to correct for initial biases (see Conrey et al., 2005).
The current study was designed to determine if rapid responses to members of different social categories can be modulated by relatively transient, motivational states. Prior research has shown that a variety of motivational states, such as approach/avoidance, can affect perception and attention (e.g., Cacioppo et al., 1993a; Crites and Cacioppo, 1996; Fazio et al., 2000; Friedman and Forster, 2005). For example, non-Black participants who repeatedly used a joystick to approach (versus avoid) Black target stimuli were subsequently faster to associate the self with Blacks and showed less racial bias on the Implicit Association Task (Phills et al., 2011; see also Amodio, 2010). Therefore, in order to determine if motivational processes can modulate automatic social perception, we placed participants in an approach or avoidant frame during a person perception task while collecting scalp EEG data. Specifically, we examined whether approaching other-race faces would attenuate racial biases in early perceptual processing.
Although the influence of social categories during person perception is widely distributed in the brain (see Cunningham and and Van Bavel, 2009), social categories appear, in particular, to influence very early components of the face processing network (see Ito and Bartholow, 2009). For instance, the core and extended face processing network (Kanwisher et al., 1997), including the fusiform gyri and amygdala, respectively, have been associated with own-race (Golby et al., 2001; Lieberman et al., 2005) and own-group (Van Bavel et al., 2008, 2011) biases in social perception. Although the relationship between specific brain regions and ERP waves is not perfectly precise (see Luck, 2005), very early ERP waves, such as the P100 (Bentin et al., 1996) and N170 (Herrmann et al., 2005) appear to subserve early face processing. The P100 is the first positive going component and peaks around 100 ms following stimulus presentation with a source generated in the ventrolateral prestriate cortex (Martinez et al., 1999; Di Russo et al., 2003). The N170 is a negative going component that peaks around 170 ms after stimulus presentation with a source generated in the fusiform and inferior-temporal gyri (Halgren et al., 2000). Several studies have shown differential responses to race in the N170 (e.g., Ito and Urland, 2005; Herrmann et al., 2007). For the purposes of the present research, we were interested in examining whether motivational states might influence these very rapid responses to the race of target faces.
If the very early effects of social categories are inevitable, motivational states induced during person perception should not affect rapid biases—that is, manipulating an approach/avoidance frame should have no impact on very early ERP components, and participants should show racial bias in perceptual processing regardless of motivational state. However, if the earliest effects of social categories are not inevitable, but are sensitive to current motivational states, then biases in early ERP components may differ depending on whether people are in an approach versus avoidance frame. Specifically, we predict that race-based biases will be attenuated when participants are approaching versus avoiding social stimuli because social categories are less likely to be used in person perception in an approach-motivated state. A person that one approaches is more motivationally relevant than a person that one avoids, and prior studies have shown that altering the motivational relevance of social stimuli through manipulations of processing goals (Cunningham et al., 2008) and group membership (Van Bavel et al., 2008) affects the way that people are perceived and evaluated. In particular, increasing motivational relevance during person perception seems to increase the extent to which a target is individuated rather than processed in terms of a category membership (e.g., Neuberg and Fiske, 1987; Van Bavel et al., 2011). Thus, if motivational processes can affect initial social categorization, then putting people in the mindset of approaching others should alter the immediate processing of race and attenuate racial bias in early perceptual processing.
Materials and Methods
Participants
Fourteen White male undergraduate psychology students from the University of Toronto completed this study for partial completion of course credit or $15. All participants gave informed consent.
Experimental Design
Upon arrival at the lab, participants were informed that they would be completing an experiment designed to examine the neural processing underlying social cognition. Participants were asked to respond to the presentation of faces with a joystick as quickly as possible. The experiment employed a block design in which participants approached or avoided blocks of three faces presented in succession. To simulate approach and avoidance toward the faces, participants pulled or pushed a joystick at the onset of each face (Cacioppo et al., 1993b; Kawakami et al., 2007). At the start of each block an instruction screen appeared for 2 s. During the approach blocks, this instruction screen indicated that participants should “pull each face toward” them. During the avoidance blocks, this instruction screen indicated that participants should “push each face away” from them. Twenty-four faces of Black or White college-aged males were presented randomly during the blocks (taken from Van Bavel and Cunningham, 2009). Faces were cropped to include only the faces and neck, but included hair. None of the faces had facial hair. Faces were fully counterbalanced across conditions to ensure that interactive effects with condition could not be attributed to low-level features of the stimuli. Participants saw four runs of 16 blocks of three trials each for a total of 192 trials (48 trials per condition). In each block, three faces were presented for 1 s each, and 3 s of fixation separated each face. To create and maintain a motivational mental set, two runs included approach blocks and two runs included avoid blocks (randomized within participants). To minimize blink artifacts during experimental trials, runs were separated by a 12 s rest period and participants were encouraged to blink during this period rather than during the task. Further, between each set of three faces, participants were instructed to blink if necessary during this period. Scalp electroencephalographic data were acquired with a 128-channel ANT system (Advanced Neuro Technology, Netherlands) using a 64-channel acquisition setup, sampled at 512 Hz, using an average reference, digitally filtered off-line with a 1–15 Hz bandpass filter.
Results
After deleting trials with noncephalic artifacts using BESA default settings1 (MEGIS Software, Germany), each participant's trials were aggregated based on trial type. On average, 86% of trials were retained. For each participant, trials in each the four stimulus conditions (Black-Pull, Black-Push, White-Pull, White-Push) were averaged to create grand average waveforms for each electrode, as well as a grand average aggregating across all trials. The 100 ms period prior to stimulus presentation was used as baseline for averaging. Because EEG data is not independent, a source modeling analysis was run on the average waveform in order to characterize the whole brain signal (see Smith et al., 2003), using the full time series for the epoch and all of the electrodes as input using BESA 5.2. The average waveform was used for source modeling to avoid biasing the results toward any particular condition.
The resulting PCA indicated that one latent source centered in the right occipito-temporal cortex accounted for 90.2% of the observed variance (see Figure 1A). The occipito-temporal cortex plays a key role in face processing (Kanwisher et al., 1997) and this region has been associated with own-race (Golby et al., 2001; Lieberman et al., 2005) and own-group (Van Bavel et al., 2008, 2011) biases in social perception. An additional LORETA source analysis replicated this localization to the right fusiform gyrus (see Figure 1B). Plotting the latent time courses, we found that this latent variable included both the P100 and N170 components typically found in studies of attention and face processing for each of the four experimental conditions (Figure 2). These results are consistent with previous studies that have shown the P100 (Bentin et al., 1996) and N170 may be associated with processing in the fusiform gyrus (Herrmann et al., 2005). Therefore, in order to test our hypotheses, we analyzed these waveforms as a function of target race and motivational condition.
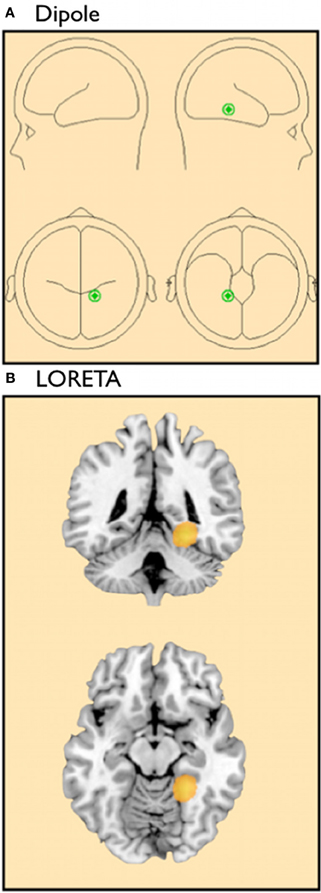
Figure 1. (A) Localization of BESA Dipole modeling for latent variable. (B) LORETA source modeling result for latent variable.
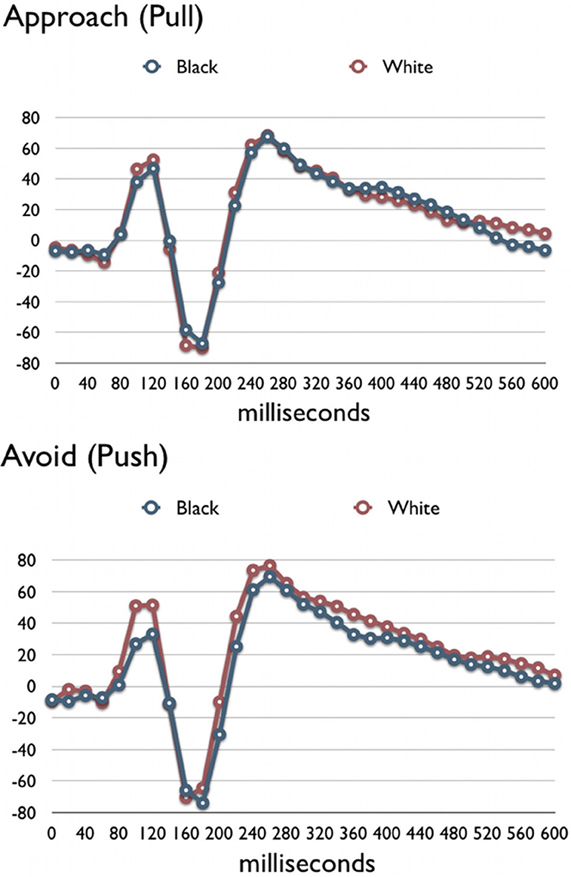
Figure 2. Latent time courses for Black and White faces in the Pull and Push conditions. Because these values are projected from latent space, values on the y axis are scaled in arbitrary units with respect to underlying microvolts.
If the early effects of social categories such as race are inevitable, there should only be a main effect of race on early ERP waveforms (Ito et al., 2007), regardless of motivational state. However, if early processes in person perception are malleable, there should be an interaction between race and motivational state, such that any pattern of racial bias in the avoidance blocks should be attenuated in the approach blocks. To examine this hypothesis, we back projected the latent variable to create individual scores for each participant for each trial type. These scores were subjected to a 2 (race: Black, White) × 2 (motivational state: approach, avoid) ANOVA. Separate analyses were conducted for the P100 (mean amplitude between 90 and 110 ms) and N170 (mean amplitude between 135 and 200 ms) components of the waveform.
Consistent with the prior evidence that people show racial bias during early perceptual process, White faces were associated with a larger P100 than Black faces, [F(1, 13) = 12.68, p < 0.01, partial η2 = 0.49] (see Figure 3). This pattern replicates previous research showing an own-race bias during very automatic facial processing (e.g., Ito et al., 2004). Importantly, this own-race effect was qualified by a race × motivational state interaction, [F(1, 13) = 4.32, p = 0.05, partial η2 = 0.25]. Simple effects indicated that the own-race effect was significant for the avoid blocks, [F(1, 13) = 12.51, p < 0.01], but not the approach blocks, [F(1, 13) = 4.07, p = 0.07]. Moreover, a contrast pitting all other conditions versus the Black-Push condition (1, 1, 1, −3) was significant, [F(1, 13) = 10.66, p = 0.01]. These results are consistent with the suggestion that approach oriented motivational states can alter the influence of social categories—even during the first 100 ms of perceptual processing. When considering the later N170 component, there was no main effect of race, [F(1, 13) = 1.25, p = 0.28, partial η2 = 0.09], nor was there a race × motivational state interaction, [F(1, 13) = 0.62, p = 0.45, partial η2 = 0.05].
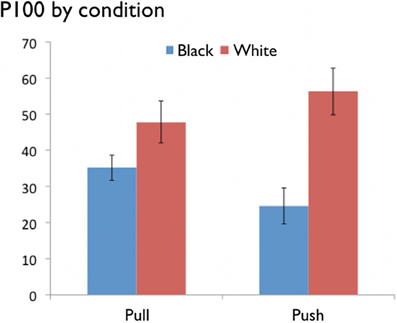
Figure 3. Mean amplitudes for the P100 for Black and White faces in the Pull and Push conditions.
Secondary analyses were run using the raw data rather than the latent variable. For these analyses, we averaged data from the right and left occipital electrodes to create two waveforms of interest2. The P100 and the N170 mean amplitudes were extracted from the subject level grand averages using the same time windows as above and were subjected to a 2 (race: Black, White) × 2 (motivational state: approach, avoid) × 2 (laterality: right, left occipital electrodes) ANOVA. Replicating the results from the latent variable analysis, we found a main effect of race, [F(1, 13) = 9.20, p < 0.01], and a race × motivational state interaction, [F(1, 13) = 9.00, p < 0.01], for the P100 component. Specifically, the P100 difference between Black and White faces was larger for the avoid blocks (MWhite = 2.68; MBlack = 1.22) than the approach blocks (MWhite = 2.04; MBlack = 1.58). There were no significant laterality interactions. Unlike the latent variable analysis, we did find a race × motivational state for the N170 [F(1, 13) = 5.91, p < 0.03]. This analysis suggested that there was a larger N170 response to Black faces than White faces (MWhite = −1.07; MWhite = −0.45) for the avoid blocks, but not the approach blocks (MBlack = −0.76; MWhite = −0.75). However, this effect was not significant when controlling for the P100 effects [F(1, 13) = 1.55, p = ns]. No significant differences in latency were found for either component.
Discussion
The current research is consistent with the idea that the earliest aspects of social perception are flexible and sensitive to motivational frames. Putting people in the mindset of approaching others attenuated racial bias in very early perceptual processing. Specifically, when people pushed a joystick away from themselves—an experimental manipulation designed to induce an avoidance motivation—they showed an own-race bias, such that early perceptual processing (as indexed by ERP activity around 100 ms after stimulus onset) was stronger to own-race than other-race faces. However, this own-race bias was modulated by motivational state, such that approaching faces by pulling a joystick toward oneself reduced the bias in neural activity.
This finding challenges theories suggesting that early biases associated with social categorization are inevitable and only interrupted by controlled processing (e.g., Devine, 1989) or perceptual load (e.g., Ito et al., 2007). These models are difficult to reconcile with the current results since the P100 occurs much faster than downstream corrective processes (Amodio et al., 2008), and there is no reason to believe that the two conditions in the current study (approach versus avoid) differ in terms of perceptual demands.
Instead, this research adds to a growing literature demonstrating that motivational processes can influence the most automatic aspects of social perception and evaluation (e.g., Cunningham et al., 2005; Cunningham and Zelazo, 2007; Amodio, 2010). Additionally, this research adds to the literature suggesting that motivational relevance can determine whether top-down processes will override automatic, bottom-up perceptual and attentional effects (Cunningham et al., 2008; Van Bavel et al., 2008). By showing that very early processes in person perception are sensitive to motivational states, this research demonstrates that processes once thought to be inevitable may in fact be malleable. As such, dual-process models of social perception may be unable to account for the flexibility of automatic social perception and evaluation (see Cunningham et al., 2007).
Addressing the question of the inevitability of attentional biases to social categories requires measures that are highly temporally sensitive. Whereas other research has shown that biases associated with social categorization can be eliminated in a number of ways, such as by getting people to focus on a different dimension of categorization (e.g., Van Bavel and Cunningham, 2009), the behavioral methods typically used to measure bias do not provide sufficient information to determine if these biases are circumvented during early perceptual processing or rapidly corrected after the fact. By using a measure with exquisite temporal resolution (i.e., EEG), we were able to determine that very rapid aspects of social categorization are not inevitable and can be altered by modifying motivational states.
It is important to note that Ito et al. (2007) conclusion regarding the inevitability of early attentional biases in social categorization was based on the N170 waveform, while our results were found for the P100 waveform. Although they likely differ in important ways, both waveforms are related to early attentional processing (Bentin et al., 1996; Clark and Hilyard, 1996) as well as face processing (e.g., Liu et al., 2002; Herrmann et al., 2005). Moreover, a recent study found that people with greater left alpha asymmetries in the prefrontal cortex—a correlate of approach motivation—had different ERP responses to Black (versus White) faces on the P2, which peaked approximately 170 ms following stimulus presentation (Amodio, 2010). In that study, P2 responses to Black faces were also associated with decreased racial bias on an implicit measure of racial stereotypes. Together with the current research, these results suggest that motivation may help one have less prejudiced responses by modulating perception (see Balcetis and Dunning, 2006).
Conclusion
The current research suggests that very early effects of social categorization can be modulated before they impact subsequent perceptions, evaluations, or behavior. Given that post-categorization control often has negative side effects, like rebound (Macrae et al., 1994) and depletion (Richeson and Shelton, 2003; Gordijn et al., 2004), altering initial processing of social categories by changing motivational states provides a powerful alternative for changing these biases. While preventing the effects of attentional biases associated with social categorization is not necessarily positive (Trawalter et al., 2008), it does provide an opportunity to overcome ostensibly automatic stereotyping and prejudice. Knowing which steps in the processing sequence are malleable is important for understanding what types of interventions will be successful in preventing the downstream consequences of social categorization.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This research was presented at the Association for Psychology Science (2006), Society for Personality and Social Psychology (2007) and the Society for Social and Affective Neuroscience (2008). The authors would like to thank Peter Lenkic for help with data collection. This research was supported by grants from the SSHRC to William Cunningham, Jay Van Bavel and Dominic Packer and the NSF (BCS-0819250) to William Cunningham.
Footnotes
References
Amodio, D. M. (2010). Coordinated roles of motivation and perception in the regulation of intergroup responses: frontal cortical asymmetry effects on the P2 event-related potential and behavior. J. Cogn. Neurosci. 22, 2609–2617.
Amodio, D. M., Devine, P. G., and Harmon-Jones, E. (2008). Individual differences in the regulation of intergroup bias: the role of conflict monitoring and neural signals for control. J. Pers. Soc. Psychol. 94, 60–74.
Balcetis, E., and Dunning, D. (2006). See what you want to see: motivational influences on visual perception. J. Pers. Soc. Psychol. 91, 612–625.
Bentin, S., Allison, T., Puce, A., Perez, E., and McCarthy, G. (1996). Electrophysiological studies of face perception in humans. J. Cogn. Neurosci. 8, 551–565.
Blair, I. V. (2002). The malleability of automatic stereotypes and prejudice. Pers. Soc. Psychol. Rev. 6, 242–261.
Brewer, M. B. (1988). “A dual process model of impression formation,” in Advances in Social Cognition, Vol. 1, eds R. S. Wyer and T. K. Srull (Hillsdale, NJ: Erlbaum), 1–36.
Cacioppo, J. T., Crites, S. L. Jr., Berntson, G. G., and Coles, M. G. H. (1993a). If attitudes affect how stimuli are processed, should they not affect the event-related brain potential. Psychol. Sci. 4, 108–112.
Cacioppo, J. T., Priester, J. R., and Berntson, G. G. (1993b). Rudimentary determinants of attitudes. II: arm flexion and extension have differential effects on attitudes. J. Pers. Soc. Psychol. 65, 5–17.
Clark, V., and Hilyard, S. A. (1996). Spatial selective attention affects early extrastriate but not striate components of the visual evoked potential. J. Cogn. Neurosci. 8, 387–402.
Conrey, F. R., Sherman, J. W., Gawronski, B., Hugenberg, K., and Groom, C. (2005). Separating multiple processes in implicit social cognition: the quad-model of implicit task performance. J. Pers. Soc. Psychol. 89, 469–487.
Crites, S. L. Jr., and Cacioppo, J. T. (1996). Electrocortical differentiation of evaluative and nonevaluative categorizations. Psychol. Sci. 7, 318–321.
Cunningham, W. A., and Van Bavel, J. J. (2009). “A neural analysis of intergroup perception and evaluation,” in Handbook of Neuroscience for the Behavioral Sciences, Vol. 1, eds G. G. Berntson and J. T. Cacioppo (Hoboken, NJ: John Wiley and Sons, Inc.), 975–984.
Cunningham, W. A., Van Bavel, J. J., and Johnsen, I. R. (2008). Affective flexibility: evaluative processing goals shape amygdala activity. Psychol. Sci. 19, 152–160.
Cunningham, W. A., and Zelazo, P. D. (2007). Attitudes and evaluations: a social cognitive neuroscience perspective. Trends Cogn. Sci. 11, 97–104.
Cunningham, W. A., Raye, C. L., and Johnson, M. K. (2005). Neural correlates of evaluation associated with promotion and prevention regulatory focus. Cogn. Affect. Behav. Neurosci. 5, 202–211.
Cunningham, W. A., Zelazo, P. D., Packer, D. J., and Van Bavel, J. J. (2007). The iterative reprocessing model: a multi-level framework for attitudes and evaluation. Soc. Cogn. 25, 736–760.
Dehaene, S., Changeux, J. P., Naccache, L., Sackur, J., and Sergent, C. (2006). Conscious, preconscious, and subliminal processing: a testable taxonomy. Trends Cogn. Sci. 10, 204–211.
Devine, P. G. (1989). Stereotypes and prejudice: their automatic and controlled components. J. Pers. Soc. Psychol. 56, 5–18.
Di Russo, F., Martinez, A., and Hillyard, S. A. (2003). Source analysis of event-related cortical activity during visuo-spatial attention. Cereb. Cortex 13, 486–499.
Dovidio, J. F., Kawakami, K., Johnson, C., Johnson, B., and Howard, A. (1997). The nature of prejudice: automatic and controlled processes. J. Exp. Soc. Psychol. 33, 510–540.
Fazio, R. H., Jackson, J. R., Dunton, B. C., and Williams, C. J. (1995). Variability in automatic activation as an unobtrusive measure of racial attitudes: a bona fide pipeline? J. Pers. Soc. Psychol. 69, 1013–1027.
Fazio, R. H., Ledbetter, J. E., and Towles-Schwen, T. (2000). On the costs of accessible attitudes: detecting that the attitude object has changed. J. Pers. Soc. Psychol. 78, 197–210.
Fiske, S. T., and Neuberg, S. L. (1990). A continuum model of impression formation: from category-based to individuating processes as a function of information, motivation, and attention. Adv. Exp. Soc. Psychol. 23, 1–74.
Freeman, J. B., and Ambady, N. (2011). A dynamic interactive theory of person construal. Psychol. Rev. 118, 247–279.
Friedman, R. S., and Forster, J. (2005). Effects of motivational cues on perceptual asymmetry: implications for creativity and analytical problem solving. J. Pers. Soc. Psychol. 88, 263–275.
Golby, A. J., Gabrieli, J. D. E., Chiao, J. Y., and Eberhardt, J. L. (2001). Differential responses in the fusiform region to same-race and other-race faces. Nat. Neurosci. 4, 845–850.
Gordijn, E. H., Hindriks, I., Koomen, W., Dijksterhuis, A., and van Knippenberg, A. (2004). Consequences of stereotype suppression and internal suppression motivation: a self-regulation approach. Pers. Soc. Psychol. Bull. 30, 212–224.
Halgren, E., Raij, T., Marinkovic, K., Jousmaki, V., and Hari, R. (2000). Cognitive response profile of the human fusiform face area as determined by MEG. Cereb. Cortex 10, 69–81.
Herrmann, M., Ehlis, A., Muehlberger, A., and Fallgatter, A. (2005). Source localization of early stages of face processing. Brain Topogr. 18, 77–85.
Herrmann, M. J., Schreppel, T., Jäger, D., Koehler, S., Ehlis, A. C., and Fallgatter, A. J. (2007). The other-race effect for face perception: an event-related potential study. J. Neural Transm. 114, 951–957.
Ito, T. A., and Bartholow, B. D. (2009). The neural correlates of race. Trends Cogn. Sci. 13, 524–531.
Ito, T. A., and Cacioppo, J. T. (2000). Electrophysiological evidence of implicit and explicit categorization processes. J. Exp. Soc. Psychol. 36, 660–676.
Ito, T. A., Thompson, E., and Cacioppo, J. T. (2004). Tracking the timecourse of social perception: the effects of racial cues on event-related brain potentials. Pers. Soc. Psychol. Bull. 30, 1267–1280.
Ito, T. A., and Urland, G. R. (2003). Race and gender on the brain: electrocortical measures of attention to the race and gender of multiply categorizable individuals. J. Pers. Soc. Psychol. 85, 616–626.
Ito, T. A., and Urland, G. R. (2005). The influence of processing objectives on the perception of faces: an ERP study of race and gender perception. Cogn. Affect. Behav. Neurosci. 5, 21–36.
Ito, T. A., Willadsen-Jensen, E. C., and Correll, J. (2007). “Social neuroscience and social perception: new perspectives on categorization, prejudice, and stereotyping,” in Social Neuroscience: Integrating Biological and Psychological Explanations of Social Behavior, eds E. Harmon-Jones and P. Winkielman (New York, NY: Guilford), 401–421.
Kanwisher, N. G., McDermott, J., and Chun, M. M. (1997). The fusiform face area: a module in human extrastriate cortex specialized for face perception. J. Neurosci. 17, 4302–4311.
Kawakami, K., Phills, C. E., Steele, J. R., and Dovidio, J. F. (2007). (Close) Distance makes the heart grow fonder: improving implicit racial attitudes and interracial interactions through approach behaviors. J. Pers. Soc. Psychol. 92, 957–971.
Lieberman, M. D., Hariri, A., Jarcho, J. M., Eisenberger, N. I., and Bookheimer, S. Y. (2005). An fMRI investigation of race-related amygdala activity in african-american and caucasian-american individuals. Nat. Neurosci. 8, 720–722.
Liu, J., Harris, A., and Kanwisher, N. (2002). Stages of processing in face perception: a MEG study. Nat. Neurosci. 5, 910–916.
Luck, S. J. (2005). An Introduction to the Event-Related Potential Technique. Cambridge, MA: MIT Press.
Macrae, C. N., Bodenhausen, G. V., Milne, A. B., and Jetten, J. (1994). Out of mind but back in sight: stereotypes on the rebound. J. Pers. Soc. Psychol. 67, 808–817.
Martinez, A., Anllo-Vento, L., Sereno, M. I., Frank, L. R., Buxton, R. B., Dubowitz, D. J., Wong, E. C., Hinrichs, H., Heinze, H. J., and Hillyard, S. A. (1999). Involvement of striate and extrastriate visual cortical areas in spatial attention. Nat. Neurosci. 2, 364–369.
Neuberg, S. L., and Fiske, S. T. (1987). Motivational influences on impression formation: outcome dependency, accuracy-driven attention, and individuating processes. J. Pers. Soc. Psychol. 53, 431–444.
Phills, C. E., Kawakami, K., Tabi, E., Nadolny, D., and Inzlicht, M. (2011). Mind the gap: increasing associations between the self and blacks with approach behaviors. J. Pers. Soc. Psychol. 100, 197–210.
Richeson, J. A., and Shelton, J. N. (2003). When prejudice does not pay: effects of interracial contact on executive function. Psychol. Sci. 14, 287–290.
Smith, N. K., Cacioppo, J. T., Larsen, J. T., and Chartrand, T. L. (2003). May I have your attention please: electrocortical responses to positive and negative stimuli. Neuropsychologia 41, 171–183.
Trawalter, S., Todd, A., Baird, A. A., and Richeson, J. A. (2008). Attending to threat: race-based patterns of selective attention. J. Exp. Soc. Psychol. 44, 1322–1327.
Van Bavel, J. J., and Cunningham, W. A. (2009). Self-categorization with a novel mixed-race group moderates automatic social and racial biases. Pers. Soc. Psychol. Bull. 35, 321–335.
Van Bavel, J. J., and Cunningham, W. A. (2011). A social neuroscience approach to self and social categorisation: a new look at an old issue. Eur. Rev. Soc. Psychol. 21, 237–284.
Van Bavel, J. J., Packer, D. J., and Cunningham, W. A. (2008). The neural substrates of in-group bias: a functional magnetic resonance imaging investigation. Psychol. Sci. 19, 1131–1139.
Keywords: race, ERP, P100, social perception, face perception, motivation, approach
Citation: Cunningham WA, Van Bavel JJ, Arbuckle NL, Packer DJ and Waggoner AS (2012) Rapid social perception is flexible: approach and avoidance motivational states shape P100 responses to other-race faces. Front. Hum. Neurosci. 6:140. doi: 10.3389/fnhum.2012.00140
Received: 30 December 2011; Accepted: 30 April 2012;
Published online: 24 May 2012.
Edited by:
Chad E. Forbes, University of Delaware, USAReviewed by:
Jonathan Freeman, Tufts University, USAJamie Morris, University of Virginia, USA
Elliot Berkman, University of Oregon, USA
Copyright: © 2012 Cunningham, Van Bavel, Arbuckle, Packer and Waggoner. This is an open-access article distributed under the terms of the Creative Commons Attribution Non Commercial License, which permits non-commercial use, distribution, and reproduction in other forums, provided the original authors and source are credited.
*Correspondence: William A. Cunningham, Department of Psychology, University of Toronto, 100 St. George Street, Toronto, ON M5S 3G3, Canada. e-mail:Y3VubmluZ2hhbUBwc3ljaC51dG9yb250by5jYQ==