 Raiane S. Rabelo
Raiane S. Rabelo Guilherme Sena
Guilherme Sena José F. Gonçalves Jr
José F. Gonçalves Jr- 1AquaRiparia/Limnology Laboratory, Department of Ecology, Institute of Biological Sciences, University of Brasília, Brasília, Brazil
- 2Tropical Water Research Alliance-TWRA, Brasília, Brazil
Factors that may accelerate decomposition are important for ecosystem functioning since plant litter decomposition is essential for carbon and nutrient cycling, but it is a generally slow process, which can take weeks up to years. In this context, studies have demonstrated that the chemical characteristics of litter mixtures can accelerate decomposition through several mechanisms. Tropical riparian forests are known for their high diversity of tree species, which leads to a wide array of litter types in tropical streams, each with distinct chemical properties. This underscores the key role of litter chemistry in significantly influencing the litter breakdown rate within of these streams. Here, we explore the interplay among litter chemistry, plant species diversity, and litter decomposition in tropical streams. We highlight the importance of litter physical and chemical characteristics for decomposition, as well as of the preservation of the natural floristic composition of tropical riparian forests. In this sense, more attention must be paid to the influence that the environment and phylogeny may have on the phytochemical characteristics of riparian forest plant species in different tropic biomes, and how the insertion of different exotic species interferes with the decomposition process. Furthermore, we emphasize the need for additional research into the consequences of the loss of rare plant species with unique functional characteristics to decomposition in tropical ecosystems.
Introduction
It is well established that terrestrial plant litter stands out as the predominant and most crucial source of carbon (C) and nutrients for forested streams (Webster and Benfield, 1986; Wallace et al., 1997; Gessner et al., 1999; Graça et al., 2015). Thus, over the years, some conceptual models on leaf dynamics and decomposition in streams have been proposed for understanding the ecological processes that occur in aquatic ecosystems, focusing on the entry, transformation, and decomposition of mainly fallen leaves. Initially the focus was on the entry of leaves into streams and the physical fragmentation of these leaves due to abrasion by mechanical action as they move with the water (Nykvist, 1959). This initial focus was followed by the idea that leaf quality, consumer activity and environmental factors (such as water temperature and dissolved oxygen) are important in determining the rate of leaf decomposition, recognizing that decomposition is a more complex process than simple physical fragmentation (Webster and Benfield, 1986; Gessner et al., 1999). More recently, it has been established that these variables, and the effects of diversity are important in determining the rate of leaf decomposition (Graça et al., 2015).
Factors that can accelerate decomposition, such as species diversity in litter mixtures, are important for ecosystem functioning since plant litter decomposition is essential for C and nutrient cycling (Wallace et al., 1997). Still, it is generally a slow process, which can take weeks up to years (e.g., Gonçalves et al., 2007). In forested streams, the riparian tree canopy decreases the solar irradiation in the stream, limiting its primary productivity, and therefore the food webs obtain most of their energy from the decomposition of terrestrial litter (Gessner et al., 1999). Low leaf litter decay rates can alter the flux of C (sink) and nutrients through various trophic levels in these ecosystems (from decomposers to predators; Gessner et al., 2010). With the acceleration of litter decomposition in forested streams, there is an increase in C metabolization (release of CO2) to the atmosphere, changing its stock, export and availability to consumers (Benstead et al., 2009). The phenomenon that increases the release of CO2 may contribute to climate change (Manning et al., 2018). Also, faster litter decomposition may load in the reduced form of nitrogen (N), phosphorus (P) and other nutrients into the ecosystem, promoting the streams' eutrophication (Graça et al., 2015).
Hundreds of experimental studies have been developed to assess the implications of diversity reduction on key ecosystem processes due to current human-driven biodiversity loss (Hector and Bagchi, 2007; Srivastava et al., 2009; Cardinale et al., 2011; Tilman et al., 2012; Mori et al., 2020). While evidence points to a consistent positive effect of plant species diversity on primary productivity (Cardinale et al., 2011; Tilman et al., 2012), studies have failed to demonstrate an apparent effect of plant species diversity on decomposition, especially in aquatic environments–since this effect is more evident in terrestrial environments (Srivastava et al., 2009; Cardinale et al., 2011; Mori et al., 2020). In this context, some studies have suggested that chemical diversity in litter mixtures rather than species diversity is a more determining factor in this process (e.g., Epps et al., 2007; Handa et al., 2014). The recognition of the importance of the chemical diversity of plant litter for decomposition is not new (e.g., Wardle et al., 1997), and more recently, this has been demonstrated empirically by studies in both aquatic and terrestrial environments (e.g., Handa et al., 2014; Boyero et al., 2021).
The chemical characteristics of litter are one of the main drivers of the activity of microbial decomposers and fragmentation by invertebrates and, consequently, the decomposition of leaf litter in forested streams (Cornwell et al., 2008; Boyero et al., 2016; Zhang et al., 2019). These litter characteristics refer to a set of intrinsic chemical attributes, commonly referred to as ‘detritus quality', in which higher quality litter (e.g., litter with higher concentrations of nutrients and lower phenolic and lignin contents) offers higher palatability and nutritional value with less toxicity for detritivore animals and microbial decomposers than lower quality litter (e.g., litter with a higher concentration of recalcitrant and phenolic compounds and lower concentrations of nutrients) (Gessner et al., 1999; Graça et al., 2015). Lignin and cellulose are essential structural constituents of plant cell walls, providing rigidity and strength to plant cell walls to withstand mechanical stress, protect against pathogens and herbivores, and maintain their structural integrity (Gessner, 2020). Consequently, litter rich in these compounds tends to be highly refractory, leading to slow decomposition (Graça et al., 2015; Zhang et al., 2019). In addition, the secondary compounds (e.g., phenolic compounds) are a group of secondary plant metabolites that are produced as a chemical defense against insects, ultraviolet radiation, and microbial infections (Lill and Marquis, 2001; Gould and Lee, 2002) and can remain in the leaf after senescence and negatively affect the colonization of plants decomposers (Bärlocher and Graça, 2005; Moretti et al., 2009).
Leaf litter also contains varying amounts of N, P and other essential macronutrients needed for plant growth (Reichardt and Timm, 2020), which also stimulate the rate of decomposition (Graça et al., 2015), as they are equally indispensable elements for the metabolism of decomposers (Madan and Thind, 1998; Gessner et al., 1999). Besides, we should consider that leaf litter’s elemental ratios (mainly C:N, C:P, and N:P) have been proposed as predictors of nutrient limitation in aquatic ecosystems, for example, N:P < 14 indicates limitation by N; N:P between 14 and 16 indicates colimitation by N and P and N:P > 16 indicates P limitation (Danger, 2020). Also, the excess of C can directly restrict other elements on consumers (Elser et al., 2012).
Chemical diversity significantly influences the decomposition of litter mixtures within forested streams, primarily due to variations in the chemical composition of individual litter species (Epps et al., 2007). The key mechanism driving this process is the transfer of nutrients from nutrient-rich litter to those with lower nutrient content (Salamanca et al., 1998). Consequently, this nutrient transfer can spur the decomposition of lower-quality litter, thereby accelerating the overall decomposition of the mixture (Gessner et al., 2010; Handa et al., 2014; Rabelo et al., 2022). This transfer of nutrients and essential compounds occurs through fungal hyphae, which form connections between distinct litter components and facilitate leaching (McTiernan et al., 1997). However, it is important to note that secondary inhibitory compounds present in the litter can also influence the microhabitat of mixed litter decomposition. Their leaching increases acidity and toxicity, potentially hampering decomposition and limiting microbial activity within the mixed litter (McArthur et al., 1994). Additionally, alternative mechanisms come into play, such as the stimulation of microbial processing (Bardgett and Shine, 1999), alterations in habitat complexity for detritivores (Sanpera-Calbet et al., 2009), and complementarity in the use of resources–the use of complementary resources derived from different types of plant litter by consumers (Vos et al., 2013). These multifaceted interactions underscore the complex interplay of chemical diversity in shaping the decomposition dynamics of litter mixtures in forested stream ecosystems.
Tropical riparian forests are known for their high diversity of tree species (Wantzen, 2008). This leads to a wide array of litter types in tropical streams, each with distinct chemical properties (Gonçalves and Callisto, 2013; Gonçalves et al., 2014). Consequently, the litter in these streams can vary significantly, ranging from nutrient-rich to nutrient-poor and high to low concentrations of phenolic and lignin compounds. This underscores the key role of chemical diversity in significantly influencing the litter breakdown rate within tropical streams.
This work aims to review and explore the interplay among litter chemistry, plant species diversity, and litter decomposition in tropical streams. We have outlined two primary inquiries: (i) What are the key factors influencing plant litter’s diversity and chemical composition in tropical streams? (ii) How does the quality and chemical diversity of mixed litter types contribute to the decomposition process in tropical stream ecosystems?
Plant litter diversity to tropical streams
Low latitudes support a higher plant species diversity in riparian forests, surpassing other regions (Wantzen, 2008; Slik et al., 2015). In tropical zones, it is common to find an average of around 100 species of trees in 1 ha (e.g., França et al., 2009; Wittmann et al., 2010; Gonçalves et al., 2014), whereas in other parts of the world, like temperate regions, riparian forests typically host between 5 and 10 tree species (Swan and Palmer, 2004). The species composition of riparian plants and their phenotypic responses are strongly related to the heterogeneity environmental, such as natural variations of edaphic characteristics of soil pH, soil fertility and water availability, temperature, rainfall and ecological interactions in the community defining the high tropical biodiversity (Haridasan, 2008; Bambi et al., 2016; Miatto et al., 2016; Sena et al., 2022) (Figure 1).
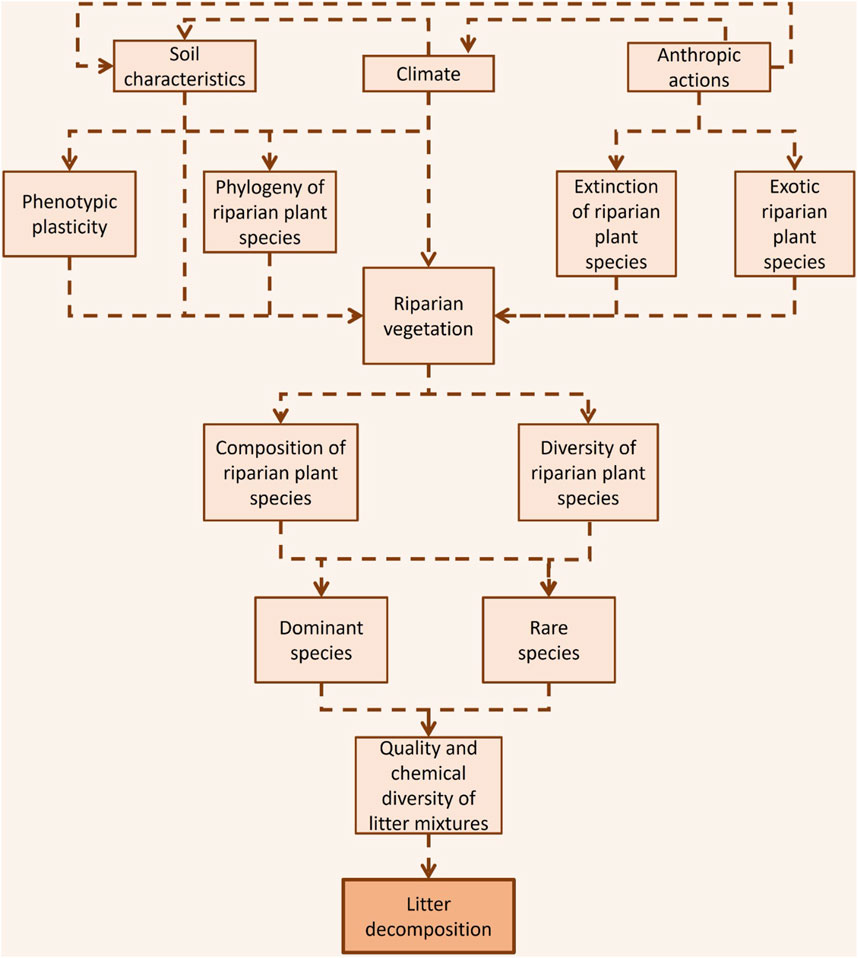
FIGURE 1. Conceptual model of the interplay among litter chemistry, plant species diversity, and litter decomposition in tropical streams. The factors that regulate the riparian vegetation (soil characteristics, climate and anthropic actions) determine the composition and diversity of dominant and rare riparian plant species, and thus regulate the quality and chemical diversity of litter mixtures. The quality and chemical diversity of litter mixtures drive decomposition in tropical streams.
Plant litter entering streams in high latitudes commonly reflects the diversity and composition of the adjacent riparian community of tree species (Swan and Palmer, 2004). Nonetheless, this is often not true in tropical streams because of their higher riparian plant diversity (Gonçalves et al., 2014; Tonin et al., 2017), the phenological variation of leaf fall of species (Sakai, 2001) and the pronounced seasonality (Tonin et al., 2017). Due to this complex dynamic, tree species from the riparian forest contribute with different amounts and composition of plant litter to the aquatic ecosystem throughout the year in streams in the tropics (Gonçalves and Callisto, 2013; Gonçalves et al., 2014). This results in a plant litter pool instream with a continuous variety of litter types with diverse chemical characteristics (Covich, 1988; Tonin et al., 2021). Also, the composition of the plant litter pool in these streams varies widely throughout the year (Gonçalves and Callisto, 2013; Gonçalves et al., 2014). Therefore, the litter mixtures' decomposition may play a key role, i.e., the way to know how the energy flow and nutrient reloading for the food web of the tropical aquatic ecosystem.
Factors that determine the litter chemical composition into tropical streams
Climate emerges as a principal determinant guiding the distribution and composition of plant species within terrestrial ecosystems (Reich and Olesksyn, 2004; Wright et al., 2004) (Figure 1). This climatic influence gives rise to a latitudinal gradient in leaf litter properties worldwide, resulting in tropical riparian forest litter exhibiting heightened recalcitrance, as demonstrated by Boyero et al. (2017). Consequently, leaf litter from tropical riparian forests may have higher toughness, heightened chemical defenses (such as elevated phenolic content), and a more pronounced phosphorus limitation, characterized by higher nitrogen-to-phosphorus (N:P) ratios, in comparison to litter from riparian forests in other regions (Boyero et al., 2017).
The climate also makes riparian forests in tropical biomes exhibit natural seasonal changes in the amount of litter input streams (Gonçalves and Callisto, 2013; Gonçalves et al., 2014; Tonin et al., 2017). For example, the Amazon and the Cerrado have a marked seasonality in the dynamics of the litter input, relating to precipitation as a limiting factor (Tonin et al., 2017). These seasonal variations in the litter input also suggest a change in the chemistry of the litter budget that has been input in the streams (Gonçalves and Callisto, 2013; Tonin et al., 2017; Tonin et al., 2021), as observed for the Cerrado. The concentration of structural compounds (carbon, fibers, lignin and cellulose) and nutrients (N and P) is highest in wet and wet-dry transition seasons and decreased toward the dry season or until the beginning of the dry-wet transition in this biome (Tonin et al., 2021).
In addition to climate variations, soil characteristics strongly influence the chemical characteristics of green leaves (Reich and Olesksyn, 2004) (Figure 1), and these characteristics are often retained as leaves transition into their senescent phase (Killingbeck, 1996). However, leaf senescence allows efficient remobilization of nutrients from leaves to other parts of the plant, serving as a survival mechanism during adverse environmental conditions such as drought or water stress (Taiz et al., 2017). For example, litter from the riparian forests of Cerrado biome are more refractory (higher C:N and C:P mass ratios and tougher tissues) than those from the Amazon and Atlantic Forest biomes (Sena et al., 2022) due to the harsh climatic and edaphic factors for plants found in the Cerrado, such as low pH, low fertility and reduced water availability in some periods of the year (Kraus et al., 2004; Haridasan, 2008; Miatto et al., 2016). Also, much of the tropical riparian forest litter biomass comes from a few dominant species (about 20 species dominate litter phenological patterns and are responsible for more than 70% of annual litter biomass; França et al., 2009; Gonçalves et al., 2014; Bambi et al., 2016; Rezende et al., 2017; Tonin et al., 2021). This means that shifts in the identity of dominant plant species in these riparian forests significantly influence the litter chemistry within the streams (Tonin et al., 2021).
Although litter characteristics are primarily driven by environmental variations (Boyero et al., 2017), species phylogeny also influences the chemical characteristics of the litter budget (LeRoy et al., 2020; Sena et al., 2022) (Figure 1). Litter from closer phylogenetic species has more similar chemical characteristics than litter from distant species due to the conservation of characteristics during their evolution species lineages (LeRoy et al., 2020; Sena et al., 2022). For example, in tropical riparian forest closely related species have similar P contents in the Atlantic Forest, which suggests that phylogeny is essential for determining P content within this tropical biome (Sena et al., 2022). Furthermore, plant species in riparian forests across different tropical biomes display phenotypic plasticity, leading to distinct variations in litter chemical characteristics within the same species (Sena et al., 2022) (Figure 1). This demonstrates that in tropical riparian forests functional responses of individual traits are induced by environmental conditions (Boyero et al., 2017; Sena et al., 2022). Thus, litter budget chemical and physical variations are influenced by climate variations, edaphic factors, composition of riparian plant species, species phylogeny and phenotypic plasticity in tropical riparian ecosystems.
Litter chemistry diversity vs. plant species diversity on decomposition into tropical streams
The established correlation between plant species diversity and various ecosystem processes is well-documented, as demonstrated by studies by Hooper et al. (2005) and Hector and Bagchi (2007). This diversity significantly influences the chemical characteristics of litter, including nutrient content, phenolic compounds, and structural elements, subsequently impacting decomposer and detritivore activity and leaf decomposition in streams (Figure 1). Evidence from previous studies shows that litter species mixtures can accelerate decomposition by up to 30% compared to single species in these ecosystems (e.g., Kominoski et al., 2007; Lecerf et al., 2007; Handa et al., 2014; Oliva et al., 2019), especially at low latitudes where more generalist detritivores appear to be relatively more efficient when more varied resources are available, through a balanced diet effect (Boyero et al., 2016; Boyero et al., 2021). However, negative or no effects of litter species diversity in mixtures on decomposition in streams were also recorded (Moretti et al., 2007; Ferreira et al., 2012; Frainer et al., 2015; Mori et al., 2020; Boyero et al., 2021). In this context, it is important to note that an increase in plant species diversity does not uniformly translate to a rise in the chemical diversity of the litter stock. Some species may have functional redundancy or unique characteristics compared to others in the community (Violle et al., 2017; Dee et al., 2019). When functional redundancy exists, species extinction may not substantially affect the ecosystem, as the remaining species can compensate for their loss (Violle et al., 2017).
Conversely, the extinction of species with distinct sets of characteristics can significantly impact ecosystem functioning, as other species in the community cannot readily replace their unique roles (Lyons et al., 2005; Dee et al., 2019). For example, the absence of plant species with specific chemical attributes in the litter stock in tropical streams may decrease fungal biomass and cause difficulty in leaf decomposition (Rabelo et al., 2022; Rabelo et al., 2023). Thus, while plant species diversity remains a key for aquatic ecosystem functioning, as observed in tropical streams (e.g., Handa et al., 2014), the chemical compounds of the litter stock hold more significant way over the decomposition process than plant diversity alone (Handa et al., 2014; Gonçalves et al., 2017; Rabelo et al., 2022; Rabelo et al., 2023). This underscores the importance of considering plant diversity and chemical characteristics in studies of ecological systems.
Importance of chemical characteristics from dominant and rare plant species for decomposition into tropical streams
The dominant plant species in the litter (higher relative contribution) are widely used by decomposition studies evaluating diversity effects, since they contribute most to ecosystem processes according to the mass ratio hypothesis (Grime, 1998). These species are the main species determining litterfall chemistry patterns (or average quality; Tonin et al., 2021). Therefore, they may direct decomposition in litter mixtures through resource concentration effects, that is, litter mixtures with a higher concentration of nutrients and lower concentration of secondary compounds, and/or litter toughness (García-Palacios et al., 2017; Boyero et al., 2021). In general, the dominant species with a higher nutrient concentration accelerate decomposition in litter mixtures, whereas those with a higher toughness or secondary compound concentration delay this process (Graça et al., 2015), as found by several studies in tropical and worldwide streams (Gonçalves et al., 2007; Gonçalves et al., 2012; Handa et al., 2014; Gonçalves et al., 2017; Zhang et al., 2019).
Despite the low relative abundance of rare plant species, it has been shown that these species may also play a potentially high role in the decomposition of litter mixtures due to their unique functional characteristics (Mouillot et al., 2013; Dee et al., 2019). These characteristics, such as distinct chemical or physical properties within the ecosystem (different concentrations of nutrients and degrees of toughness), can have a notable effect. One study has tested the impact of the loss of rare plant species on litter decomposition (Rabelo et al., 2022) and found that the loss of these species may reduce litter decomposition by around 20% in tropical streams in Central Brazil, which may lead to a decrease in nutrient reloading in the aquatic ecosystem. This emphasizes the need to consider the broader ecological implications of both dominant and rare species in the context of decomposition dynamics, especially in tropical regions.
In theory, the loss of rare plant species (conceptualized here as those with the lowest contribution to biomass) can reduce the functional diversity of litter mixtures, that is, the variation in species characteristics relevant to ecosystem functions (Tilman, 2001), and influences the resource concentration. These two aspects–functional diversity and resource concentration–are critical in litter decomposition (García-Palacios et al., 2017). It is expected that the loss of rare plant species will have an impact mainly on the functional diversity of litter mixtures since rare species are more likely to have unique functional characteristics than other more common species (e.g., high concentration of scarce resources for the community; Mouillot et al., 2013; Dee et al., 2019). As rare species have a low representation in terms of abundance or biomass of communities (Rabinowitz, 1981), the effect of rare plant species on resource concentration is expected to be low. Previous studies suggest that litter mixtures' functional diversity can accelerate decomposition and other related processes, such as nutrient cycling (Handa et al., 2014; García-Palacios et al., 2017; Rabelo et al., 2022). This acceleration may be attributed to (i) nutrients being transferred between different litter types, facilitated by fungal mediation or leaching (Handa et al., 2014); (ii) complementarity in resource utilization allowing consumers to use a broader range of resources from various plant litter types (Vos et al., 2013); (iii) furthermore, a diverse physical environment benefits detritivore (Sanpera-Calbet et al., 2009).
The potential loss of rare plant species, particularly prevalent in high-richness tropical ecosystems (Slik et al., 2015), could significantly affect decomposition dynamics (Rabelo et al., 2022) (Figure 1). Moreover, the loss of rare plant species can reduce the fungal biomass production in tropical streams, especially if the natural plant species proportion is preserved (Rabelo et al., 2023). This is because mixtures reflecting an environmental gradient may have a higher occurrence of positive and non-additive effects and thus favour more microbial growth (Swan et al., 2009). For example, fungi can benefit from the higher resource variability via nutrient transfer by fungal hyphae, contributing to their growth and activity (Bardgett and Shine, 1999; Chapman et al., 2013).
Importance of chemical characteristics from exotic plant species for decomposition into tropical streams
Exotic species are one of the leading drivers of global biodiversity decline (IPBES, 2019) and often show an initial dominance in communities (Seabloom et al., 2003). Thus, the invasion by these species is an important factor driving changes in dominance patterns (Woitke and Dietz, 2002). When invasive exotic species have functional characteristics that are different from those of the dominant native species, they often affect the ecosystem processes differently. In contrast, when the invader is functionally similar to the dominant native species, it affects the ecosystem similarly (Finerty et al., 2016). Thus, exotic plant species functionally dissimilar to dominant native plant species can significantly alter litter decomposition via changes in mixed litter composition.
Tropical riparian forests are constantly invaded by exotic plant species (van Kleunen et al., 2015), and most studies evaluating the impacts of these species (e.g., Eucalyptus species) on streams have reported changes in the seasonality, quantity, and quality of litter entering streams (Molinero and Pozo, 2004) (Figure 1). These changes can reduce (Pelizari et al., 2022), increase (Gomes et al., 2018) or do not affect (Gonçalves et al., 2012) litter decomposition, which is related mainly to higher or lower chemical and physical compounds in exotic species (i.e., N and P content and presence of refractory compounds such as lignin and cellulose) compared to native species, and to the type of community involvement in the decomposition process (microorganisms and shredder invertebrates) (Ferreira et al., 2019). In regions where shredder invertebrates are not important decomposers and litter decomposition is microbial-driven primarily, as is often the case in tropical regions [e.g., Gonçalves et al. (2007)], the litter decomposition may be less affected by invasive exotic plant species since their effects are more potent for streams where shredders are well represented (Ferreira et al., 2016; Ferreira et al., 2019), although invasion by these species can have significant impacts on microbial colonization and decomposition (Gomes et al., 2018).
Conclusion
We seek to demonstrate insights into the intricate links among litter chemistry, plant species diversity, and litter decomposition in tropical streams. Our study highlights the importance of litter’s physical and chemical characteristics for decomposition and the importance of preservation of the natural floristic composition of tropical riparian forests.
The higher diversity of environments, the wide variation climate and the higher richness of plant species in the riparian forests of tropical streams have required a great effort to increase understanding of the litter decomposition process in tropical streams. In this sense, the further steps attention should be paid to (i) the influence that the environment and phylogeny may have on the phytochemical characteristics of different plant species in riparian forests in different biomes; (ii) the impact of possible limitations on the metabolism of aquatic decomposer organisms; (iii) and how the insertion of different exotic plant species and the loss of rare plant species interfere with the decomposer community and the decomposition process. Furthermore, studies considering the current context of effects of global climate (Martins et al., 2017) and environmental changes (Campos et al., 2022) converging with the suppression or replacement of species in the riparian forests are required once these changes may maximize the negative impacts on decomposition in tropical streams.
Author contributions
RR: Conceptualization, Investigation, Writing–original draft, Writing–review and editing. GS: Conceptualization, Investigation, Writing–original draft, Writing–review and editing. JG: Conceptualization, Writing–original draft, Writing–review and editing.
Funding
The author(s) declare financial support financial support was received for the research, authorship, and/or publication of this article. JG is grateful to CNPq for the resources for the TWRA structuring project (Proc. 400439/2022-0) and the PDE grant (Proc. 200356/2022-4).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bambi, P., Rezende, R. d. S., Cruz, T. M. S., Batista, J. E. d. A., Miranda, F. G. G., Vieira dos Santos, L., et al. (2017). Diversidade da flora fanerogâmica de três matas de galeria no bioma cerrado. Heringeriana 10, 147–167. doi:10.17648/heringeriana.v10i2.224
Bardgett, R. D., and Shine, A. (1999). Linkages between plant litter diversity, soil microbial biomass and ecosystem function in temperate grasslands. Soil Biol. Biochem. 31, 317–321. doi:10.1016/S0038-0717(98)00121-7
Bärlocher, F., and Graça, M. A. S. (2005). Total phenolics. In M. A. S. Graça, F. Bärlocher, and M. O. Gessner, (Eds.), In: Methods to study litter decomposition: a practical guide (pp. 97–100). Springer.
Benstead, J. P., Rosemond, A. D., Cross, W. F., Wallace, J. B., Eggert, S. L., Suberkropp, K., et al. (2009). Nutrient enrichment alters storage and fluxes of detritus in a headwater stream ecosystem. Ecology 90 (9), 2556–2566. doi:10.1890/08-0862.1
Boyero, L., Graça, M. A. S., Tonin, A. M., Pérez, J., J Swafford, A., Ferreira, V., et al. (2017). Riparian plant litter quality increases with latitude. Sci. Rep. 7 (1), 10562–10610. doi:10.1038/s41598-017-10640-3
Boyero, L., Pearson, R. G., Hui, C., Gessner, M. O., Pérez, J., Alexandrou, M. A., et al. (2016). Biotic and abiotic variables influencing plant litter breakdown in streams: a global study. Proc. R. Soc. B Biol. Sci. 283 (1829), 20152664. doi:10.1098/rspb.2015.2664
Boyero, L., Pérez, J., López-Rojo, N., Tonin, A. M., Correa-Araneda, F., Pearson, R. G., et al. (2021). Latitude dictates plant diversity effects on instream decomposition. Sci. Adv. 7 (13), eabe7860. doi:10.1126/sciadv.abe7860
Campos, C. A., Tonin, A. M., Kennard, M. J., and Gonçalves Júnior, J. F. (2022). Setting thresholds of ecosystem structure and function to protect streams of the Brazilian savanna. Front. Environ. Sci. 10, 867905. doi:10.3389/fenvs.2022.867905
Cardinale, B. J., Matulich, K. L., Hooper, D. U., Byrnes, J. E., Duffy, E., Gamfeldt, L., et al. (2011). The functional role of producer diversity in ecosystems. Am. J. Bot. 98 (3), 572–592. doi:10.3732/ajb.1000364
Chapman, S. K., Newman, G. S., Hart, S. C., Schweitzer, J. A., and Koch, G. W. (2013). Leaf litter mixtures alter microbial community development: mechanisms for non-additive effects in litter decomposition. PLoS One 8, e62671. doi:10.1371/journal.pone.0062671
Cornwell, W. K., Cornelissen, J. H., Amatangelo, K., Dorrepaal, E., Eviner, V. T., Godoy, O., et al. (2008). Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 11 (10), 1065–1071. doi:10.1111/j.1461-0248.2008.01219.x
Covich, A. P. (1988). Geographical and historical comparisons of neotropical streams: biotic diversity and detrital processing in highly variable habitats. J. North Am. Benthol. Soc. 7, 361–386. doi:10.2307/1467297
Danger, M. (2020). Ecological stoichiometry in detritus-based ecosystems with a special focus on forested headwater streams. Limnetica 39 (1), 335–352. doi:10.23818/limn.39.22
Dee, L. E., Cowles, J., Isbell, F., Pau, S., Gaines, S. D., and Reich, P. B. (2019). When do ecosystem services depend on rare species? Trends Ecol. Evol. 34, 746–758. doi:10.1016/j.tree.2019.03.010
Elser, J. J., Loladze, I., Peace, A. L., and Kuang, Y. (2012). Lotka re-loaded: modeling trophic interactions under stoichiometric constraints. Ecol. Model. 245, 3–11. doi:10.1016/j.ecolmodel.2012.02.006
Epps, K. Y., Comerford, N. B., Reeves III, J. B., Cropper Jr, W. P., and Araujo, Q. R. (2007). Chemical diversity–highlighting a species richness and ecosystem function disconnect. Oikos 116 (11), 1831–1840. doi:10.1111/j.0030-1299.2007.15853.x
Ferreira, V., Boyero, L., Calvo, C., Correa, F., Figueroa, R., Gonçalves, J. F., et al. (2019). A global assessment of the effects of eucalyptus plantations on stream ecosystem functioning. Ecosystems 22, 629–642. doi:10.1007/s10021-018-0292-7
Ferreira, V., Encalada, A. C., and Graça, M. A. (2012). Effects of litter diversity on decomposition and biological colonization of submerged litter in temperate and tropical streams. Freshw. Sci. 31 (3), 945–962. doi:10.1899/11-062.1
Ferreira, V., Koricheva, J., Pozo, J., and Graça, M. A. (2016). A meta-analysis on the effects of changes in the composition of native forests on litter decomposition in streams. For. Ecol. Manag. 364, 27–38. doi:10.1016/j.foreco.2016.01.002
Finerty, G. E., de Bello, F., Bílá, K., Berg, M. P., Dias, A. T., Pezzatti, G. B., et al. (2016). Exotic or not, leaf trait dissimilarity modulates the effect of dominant species on mixed litter decomposition. J. Ecol. 104 (5), 1400–1409. doi:10.1111/1365-2745.12602
Frainer, A., Moretti, M. S., Xu, W., and Gessner, M. O. (2015). No evidence for leaf-trait dissimilarity effects on litter decomposition, fungal decomposers, and nutrient dynamics. Ecology 96 (2), 550–561. doi:10.1890/14-1151.1
França, J. S., Gregório, R. S., D'Arc de Paula, J., Gonçalves Júnior, J. F., Ferreira, F. A., and Callisto, M. (2009). Composition and dynamics of allochthonous organic matter inputs and benthic stock in a Brazilian stream. Mar. Freshw. Res. 60, 990–998. doi:10.1071/MF08247
García-Palacios, P., Shaw, E. A., Wall, D. H., and Hättenschwiler, S. (2017). Contrasting mass-ratio vs. niche complementarity effects on litter C and N loss during decomposition along a regional climatic gradient. J. Ecol. 105 (4), 968–978. doi:10.1111/1365-2745.12730
Gessner, M. O. (2020). “Lignin and cellulose,” in Methods to study litter decomposition: a practical guide, 179–185.
Gessner, M. O., Chauvet, E., and Dobson, M. (1999). A perspective on leaf litter breakdown in streams. Oikos 85, 377–384. doi:10.2307/3546505
Gessner, M. O., Swan, C. M., Dang, C. K., McKie, B. G., Bardgett, R. D., Wall, D. H., et al. (2010). Diversity meets decomposition. Trends Ecol. Evol. 25 (6), 372–380. doi:10.1016/j.tree.2010.01.010
Gomes, P. P., Ferreira, V., Tonin, A. M., Medeiros, A. O., and Júnior, J. F. G. (2018). Combined effects of dissolved nutrients and oxygen on plant litter decomposition and associated fungal communities. Microb. Ecol. 75, 854–862. doi:10.1007/s00248-017-1099-3
Gonçalves, J. F., Couceiro, S. R., Rezende, R. S., Martins, R. T., Ottoni-Boldrini, B. M., Campos, C. M., et al. (2017). Factors controlling leaf litter breakdown in Amazonian streams. Hydrobiologia 792, 195–207. doi:10.1007/s10750-016-3056-4
Gonçalves, J. F., Rezende, R. d. S., França, J., and Callisto, M. (2012). Invertebrate colonisation during leaf processing of native, exotic and artificial detritus in a tropical stream. Mar. Freshw. Res. 63 (5), 428–439. doi:10.1071/MF11172
Gonçalves, J. F. J., and Callisto, M. (2013). Organic-matter dynamics in the riparian zone of a tropical headwater stream in Southern Brasil. Aquat. Bot. 109, 8–13. doi:10.1016/j.aquabot.2013.03.005
Gonçalves, J. F. J., de Souza Rezende, R., Gregório, R. S., and Valentin, G. C. (2014). Relationship between dynamics of litterfall and riparian plant species in a tropical stream. Limnologica 44, 40–48. doi:10.1016/j.limno.2013.05.010
Gonçalves, J. F. J., Graça, M. A. S., and Callisto, M. (2007). Litter decomposition in a Cerrado savannah stream is retarded by leaf toughness, low dissolved nutrients and a low density of shredders. Freshw. Biol. 52 (8), 1440–1451. doi:10.1111/j.1365-2427.2007.01769.x
K. S. Gould, and D. W. Lee (2002). “Anthocyanins and leaves. The function of anthocyanins in vegetative organs,” Advances in ecological research (London: Academic), 37. doi:10.1016/S0065-2296(02)37040-X
Graça, M. A., Ferreira, V., Canhoto, C., Encalada, A. C., Guerrero-Bolaño, F., Wantzen, K. M., et al. (2015). A conceptual model of litter breakdown in low order streams. Int. Rev. Hydrobiology 100 (1), 1–12. doi:10.1002/iroh.201401757
Grime, J. P. (1998). Benefits of plant diversity to ecosystems: immediate, filter and founder effects. J. Ecol. 86, 902–910. doi:10.1046/j.1365-2745.1998.00306.x
Handa, I. T., Aerts, R., Berendse, F., Berg, M. P., Bruder, A., Butenschoen, O., et al. (2014). Consequences of biodiversity loss for litter decomposition across biomes. Nature 509 (7499), 218–221. doi:10.1038/nature13247
Haridasan, M. (2008). Nutritional adaptations of native plants of the cerrado biome in acid soils. Braz. J. Plant Physiol. 20 (3), 183–195. doi:10.1590/S1677-04202008000300003
Hector, A., and Bagchi, R. (2007). Biodiversity and ecosystem multifunctionality. Nature 448 (7150), 188–190. doi:10.1038/nature05947
Hooper, D. U., Chapin, F. S., Ewel, J. J., Hector, A., Inchausti, P., Lavorel, S., et al. (2005). Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol. Monogr. 75 (1), 3–35. doi:10.1890/04-0922
Ipbes, W. (2019). “Intergovernmental science-policy platform on biodiversity and ecosystem services,” in Summary for policy makers of the global assessment report on biodiversity and ecosystem services of the intergovernmental science-policy platform on biodiversity and ecosystem services (Bonn, Germany: IPBES Secretariat).
Killingbeck, K. T. (1996). Nutrients in senesced leaves: keys to the search for potential resorption and resorption proficiency. Ecology 77 (6), 1716–1727. doi:10.2307/2265777
Kominoski, J. S., Pringle, C. M., Ball, B. A., Bradford, M. A., Coleman, D. C., Hall, D. B., et al. (2007). Nonadditive effects of leaf litter species diversity on breakdown dynamics in a detritus-based stream. Ecology 88 (5), 1167–1176. doi:10.1890/06-0674
Kraus, T. E. C., Zasoski, R. J., Dahlgren, R. A., Horwath, W. R., and Preston, C. M. (2004). Carbon and nitrogen dynamics in a forest soil amended with purified tannins from different plant species. Soil Biol. Biochem. 36, 309–321. doi:10.1016/j.soilbio.2003.10.006
Lecerf, A., Risnoveanu, G., Popescu, C., Gessner, M. O., and Chauvet, E. (2007). Decomposition of diverse litter mixtures in streams. Ecology 88 (1), 219–227. doi:10.1890/0012-9658(2007)88[219:DODLMI]2.0.CO;2
LeRoy, C. J., Hipp, A. L., Lueders, K., Follstad Shah, J. J., Kominoski, J. S., Ardón, M., et al. (2020). Plant phylogenetic history explains instream decomposition at a global scale. J. Ecol. 108 (1), 17–35. doi:10.1111/1365-2745.13262
Lill, J. T., and Marquis, R. J. (2001). The effects of leaf quality on herbivore performance and attack from natural enemies. Oecologia 126, 418–428. doi:10.1007/s004420000557
Lyons, K. G., Brigham, C. A., Traut, B. H., and Schwartz, M. W. (2005). Rare species and ecosystem functioning. Conserv. Biol. 19, 1019–1024. doi:10.1111/j.1523-1739.2005.00106.x
Madan, M., and Thind, K. S. (1998). “Growth”. in Physiology of fungi. Editor S. B. Nangia, (Darya Ganj, New Delhi, India: APH Publishing), 31–40.
Manning, D. W., Rosemond, A. D., Gulis, V., Benstead, J. P., and Kominoski, J. S. (2018). Nutrients and temperature additively increase stream microbial respiration. Glob. Change Biol. 24 (1), e233–e247. doi:10.1111/gcb.13906
Martins, R. T., Rezende, R. S., Gonçalves, J. F., Lopes, A., Piedade, M. T. F., Cavalcante, H. L., et al. (2017). Effects of increasing temperature and CO2 on quality of litter, shredders, and microorganisms in Amazonian aquatic systems. PLoS One 12, e01887. doi:10.1371/journal.pone.0188791
McArthur, J. V., Aho, J. M., Rader, R. B., and Mills, G. L. (1994). Interspecific leaf interactions during decomposition in aquatic and floodplain ecosystems. J. North Am. Benthol. Soc. 13, 57–67. doi:10.2307/1467265
McTiernan, K. B., Ineson, P., and Coward, P. A. (1997). Respiration and nutrient release from tree leaf litter mixtures. Oikos 78, 527–538. doi:10.2307/3545614
Miatto, R. C., Wright, I. J., and Batalha, M. A. (2016). Relationships between soil nutrient status and nutrient-related leaf traits in Brazilian cerrado and seasonal forest communities. Plant Soil 404, 13–33. doi:10.1007/s11104-016-2796-2
Molinero, J., and Pozo, J. (2004). Impact of a eucalyptus (Eucalyptus globulus Labill.) plantation on the nutrient content and dynamics of coarse particulate organic matter (CPOM) in a small stream. Hydrobiologia 528, 143–165. doi:10.1007/s10750-004-2338-4
Moretti, M., Goncalves, J. F., and Callisto, M. (2007). Leaf breakdown in two tropical streams: differences between single and mixed species packs. Limnologica 37 (3), 250–258. doi:10.1016/j.limno.2007.01.003
Moretti, M. S., Loyola, R. D., Becker, B., and Callisto, M. (2009). Leaf abundance and phenolic concentrations codetermine the selection of case-building materials by Phylloicus sp. (Trichoptera, Calamoceratidae). Hydrobiologia 630, 199–206. doi:10.1007/s10750-009-9792-y
Mori, A. S., Cornelissen, J. H. C., Fujii, S., Okada, K. I., and Isbell, F. (2020). A meta-analysis on decomposition quantifies afterlife effects of plant diversity as a global change driver. Nat. Commun. 11, 4547. doi:10.1038/s41467-020-18296-w
Mouillot, D., Bellwood, D. R., Baraloto, C., Chave, J., Galzin, R., Harmelin-Vivien, M., et al. (2013). Rare species support vulnerable functions in high-diversity ecosystems. PLoS Biol. 11, 1001569. doi:10.1371/journal.pbio.1001569
Nykvist, N. (1959). Leaching and decomposition of litter I. Experiments on leaf litter of Fraxinus excelsior. Oikos 10, 190–211. doi:10.2307/3565146
Oliva, R. L., Beltran, L. A. D. G., and Okamoto, M. (2019). Influence of leaf functional diversity on leaf breakdown in a tropical stream. Pan-Am. J. Aquat. Sci. 14 (1), 34–41.
Pelizari, G. P., Vaz, A. A., Butturi-Gomes, D., and Smith, W. S. (2022). Leaf breakdown in a tropical stream: comparison between the exotic Eucalyptus grandis and two native species. Acta Limnol. Bras. 34, e12. doi:10.1590/S2179-975X2321
Rabelo, R. S., Tonin, A. M., Boyero, L., Miranda, F. G., Gomes, P. P., Bambi, P., et al. (2022). Plant litter from rare species increases functional diversity and decomposition of species mixtures. Ecosystems 26, 42–54. doi:10.1007/s10021-022-00740-1
Rabelo, R. S., Tonin, A. M., Medeiros, A. O., Boyero, L., and Gonçalves Júnior, J. F. (2023). Evenness modulates effects of the loss of plant litter from rare species on fungal decomposers in tropical streams. Fungal Ecol. 65, 101275. doi:10.1016/j.funeco.2023.101275
Rabinowitz, D. (1981). “Seven forms of rarity,” in Biological aspects of rare plant conservation. Editor H. Synge (UK: John Wiley), 205–217. Chichester.
Reich, P. B., and Oleksyn, J. (2004). Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. 101 (30), 11001–11006. doi:10.1073/pnas.0403588101
Reichardt, K., and Timm, L. C. (2020). “How plants absorb nutrients from the soil,” in Soil, plant and atmosphere (Cham: Springer), 313–330.
Rezende, R. S., Sales, M. A., Hurbath, F., Roque, N., Goncalves, J. F., and Medeiros, A. O. (2017). Effect of plant richness on the dynamics of coarse particulate organic matter in a Brazilian Savannah stream. Limnologica 63, 57–64. doi:10.1016/j.limno.2017.02.002
Sakai, S. (2001). Phenological diversity in tropical forests. Popul. Ecol. 43 (1), 77–86. doi:10.1007/PL00012018
Salamanca, E. F., Kaneko, N., and Katagiri, S. (1998). Effects of leaf litter mixtures on the decomposition of Quercus serrata and Pinus densiflora using field and laboratory microcosm methods. Ecol. Eng. 10, 53–73. doi:10.1016/S0925-8574(97)10020-9
Sanpera-Calbet, I. S. I. S., Lecerf, A., and Chauvet, E. (2009). Leaf diversity influences in-stream litter decomposition through effects on shredders. Freshw. Biol. 54, 1671–1682. doi:10.1111/j.1365-2427.2009.02216.x
Seabloom, E. W., Harpole, W. S., Reichman, O. J., and Tilman, D. (2003). Invasion, competitive dominance, and resource use by exotic and native California grassland species. Proc. Natl. Acad. Sci. U.S.A. 100 (23), 13384–13389. doi:10.1073/pnas.1835728100
Sena, G., Tonin, A. M., Caliman, A., Callisto, M., Hamada, N., Hepp, L. U., et al. (2022). Divergent litter traits of riparian plant species between humid and drier biomes within the tropics. Ecography 2023, e06310. doi:10.1111/ecog.06310
Slik, J. W. F., Arroyo-Rodríguez, V., Aiba, S. I., Alvarez-Loayza, P., Alves, L. F., Ashton, P., et al. (2015). An estimate of the number of tropical tree species. Proc. Natl. Acad. Sci. 112, 7472–7477. doi:10.1073/pnas.1423147112
Srivastava, D. S., Cardinale, B. J., Downing, A. L., Duffy, J. E., Jouseau, C., Sankaran, M., et al. (2009). Diversity has stronger top-down than bottom-up effects on decomposition. Ecology 90 (4), 1073–1083. doi:10.1890/08-0439.1
Swan, C. M., Gluth, M. A., and Horne, C. L. (2009). Leaf litter species evenness influences nonadditive breakdown in a headwater stream. Ecology 90, 1650–1658. doi:10.1890/08-0329.1
Swan, C. M., and Palmer, M. A. (2004). Leaf diversity alters litter breakdown in a Piedmont stream. J. North Am. Benthol. Soc. 23, 15–28. doi:10.1899/0887-3593(2004)023<0015:ldalbi>2.0.co;2
Taiz, L., Zeiger, E., Møller, I. M., and Murphy, A. (2017) “Senescência vegetal e morte celular”. in Fisiologia e desenvolvimento vegetal. Editor L. B. Limar (Porto Alegre, Rio Grande do Sul, Brazil: Artmed Editora), 665–693.
Tilman, D. (2001). Functional diversity. Encycl. Biodivers. 3, 109–120. doi:10.1016/b0-12-226865-2/00132-2
Tilman, D., Reich, P. B., and Isbell, F. (2012). Biodiversity impacts ecosystem productivity as much as resources, disturbance, or herbivory. Proc. Natl. Acad. Sci. 109 (26), 10394–10397. doi:10.1073/pnas.1208240109
Tonin, A. M., Goncalves, J. F., Bambi, P., Couceiro, S. R., Feitoza, L. A., Fontana, L. E., et al. (2017). Plant litter dynamics in the forest-stream interface: precipitation is a major control across tropical biomes. Sci. Rep. 7, 10799. doi:10.1038/s41598-017-10576-8
Tonin, A. M., Lima, L. S., Bambi, P., Figueiredo, M. L., Rezende, R. S., and Gonçalves, J. F. (2021). Litterfall chemistry is modulated by wet-dry seasonality and leaf phenology of dominant species in the tropics. Front. glob. change. 4, 71. doi:10.3389/ffgc.2021.666116
Van Kleunen, M., Dawson, W., Essl, F., Pergl, J., Winter, M., Weber, E., et al. (2015). Global exchange and accumulation of non-native plants. Nature 525 (7567), 100–103. doi:10.1038/nature14910
Violle, C., Thuiller, W., Mouquet, N., Munoz, F., Kraft, N. J., Cadotte, M. W., et al. (2017). Functional rarity: the ecology of outliers. Trends Ecol. Evol. 32, 356–367. doi:10.1016/j.tree.2017.02.002
Vos, V. C. A., van Ruijven, J., Berg, M. P., Peeters, E. T. H. M., and Berendse, F. (2013). Leaf litter quality drives litter mixing effects through complementary resource use among detritivores. Oecologia 173, 269–280. doi:10.1007/s00442-012-2588-1
Wallace, J. B., Eggert, S. L., Meyer, J. L., and Webster, J. R. (1997). Multiple trophic levels of a forest stream linked to terrestrial litter inputs. Science 277 (5322), 102–104. doi:10.1126/science.277.5322.102
Wantzen, K. M. (2008). “Riparian wetlands of tropical streams,” in Tropical stream ecology (Academic Press), 199–217. doi:10.1016/B978-012088449-0.50009-1
Wardle, D. A., Bonner, K. I., and Nicholson, K. S. (1997). Biodiversity and plant litter: experimental evidence which does not support the view that enhanced species richness improves ecosystem function. Oikos 79, 247–258. doi:10.2307/3546010
Webster, J. R., and Benfield, E. F. (1986). Vascular plant breakdown in freshwater ecosystems. Annu. Rev. Ecol. Syst. 17 (1), 567–594. doi:10.1146/annurev.es.17.110186.003031
Wittmann, F., Schöngart, J., and Junk, W. J. (2010). “Phytogeography, species diversity, community structure and dynamics of Amazonian várzea forests,” in Ecology and management of Amazonian floodplain forests. Ecological Series. Editor W. J. Junk, (Berlin: Springer Verlag). doi:10.1007/978-90-481-8725-6_4
Woitke, M., and Dietz, H. (2002). Shifts in dominance of native and invasive plants in experimental patches of vegetation. Perspect. Plant Ecol. Evol. Syst. 5 (3), 165–184. doi:10.1078/1433-8319-00032
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., et al. (2004). The worldwide leaf economics spectrum. Nature 428, 821–827. doi:10.1038/nature02403
Keywords: organic matter, riparian forest, rare species, chemical diversity, nutrient cycling, ecosystem functioning
Citation: Rabelo RS, Sena G and Gonçalves JF Jr (2024) Interrelationships among litter chemistry, plant species diversity, and litter decomposition in tropical stream environments: a review. Front. Geochem. 2:1346457. doi: 10.3389/fgeoc.2024.1346457
Received: 29 November 2023; Accepted: 19 January 2024;
Published: 06 February 2024.
Edited by:
André Megali Amado, Juiz de Fora Federal University, BrazilReviewed by:
Richard Wilkin, U.S. Environmental Protection Agency, United StatesPedro Pato Martins, University of Aveiro, Portugal
Copyright © 2024 Rabelo, Sena and Gonçalves. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Raiane S. Rabelo, cmFpYW5lLnNlcmVqb0B5YWhvby5jb20=; Guilherme Sena, Z3NlbmE5MkBnbWFpbC5jb20=
†These authors have contributed equally to this work and share first authorship