
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Genome Ed. , 03 September 2021
Sec. Genome Editing in Plants
Volume 3 - 2021 | https://doi.org/10.3389/fgeed.2021.734951
This article is part of the Research Topic Utilization of Protoplasts to Facilitate Gene-Editing in Plants View all 10 articles
 Kelsey M. Reed
Kelsey M. Reed Bastiaan O. R. Bargmann*
Bastiaan O. R. Bargmann*The development of gene-editing technology holds tremendous potential for accelerating crop trait improvement to help us address the need to feed a growing global population. However, the delivery and access of gene-editing tools to the host genome and subsequent recovery of successfully edited plants form significant bottlenecks in the application of new plant breeding technologies. Moreover, the methods most suited to achieve a desired outcome vary substantially, depending on species' genotype and the targeted genetic changes. Hence, it is of importance to develop and improve multiple strategies for delivery and regeneration in order to be able to approach each application from various angles. The use of transient transformation and regeneration of plant protoplasts is one such strategy that carries unique advantages and challenges. Here, we will discuss the use of protoplast regeneration in the application of new plant breeding technologies and review pertinent literature on successful protoplast regeneration.
Since the advent of CRISPR/Cas9 and related gene-editing technology, direct modification of crop genomes has become the way of the future for advanced breeding techniques in agriculture (Zhang et al., 2019). These new plant breeding technologies (NPBT) have opened avenues of fundamental and translational research that were previously inaccessible. In contrast to transgenic approaches, NPBT can avoid costly and time-consuming regulatory hurdles and accelerate the introduction of new crop lines to the ag market (Lassoued et al., 2021).
Breeding for the introgression of new traits from a wild relative or mutagenized population into an elite crop cultivar is a lengthy procedure, requiring numerous rounds of selection to regain the characteristics of the parental strain (Figure 1A). The ability to efficiently modify crop genes can save several years over conventional breeding approaches and phenotypic recurrent selection (Bull et al., 2017). However, the current most commonly used NPBT method of inserting a transgenic CRISPR/Cas9 construct into the host genome and then crossing it out again to obtain transgene-free progeny still requires multiple rounds of selection (Figure 1B). This is especially true for highly heterozygous and/or outcrossing crops.
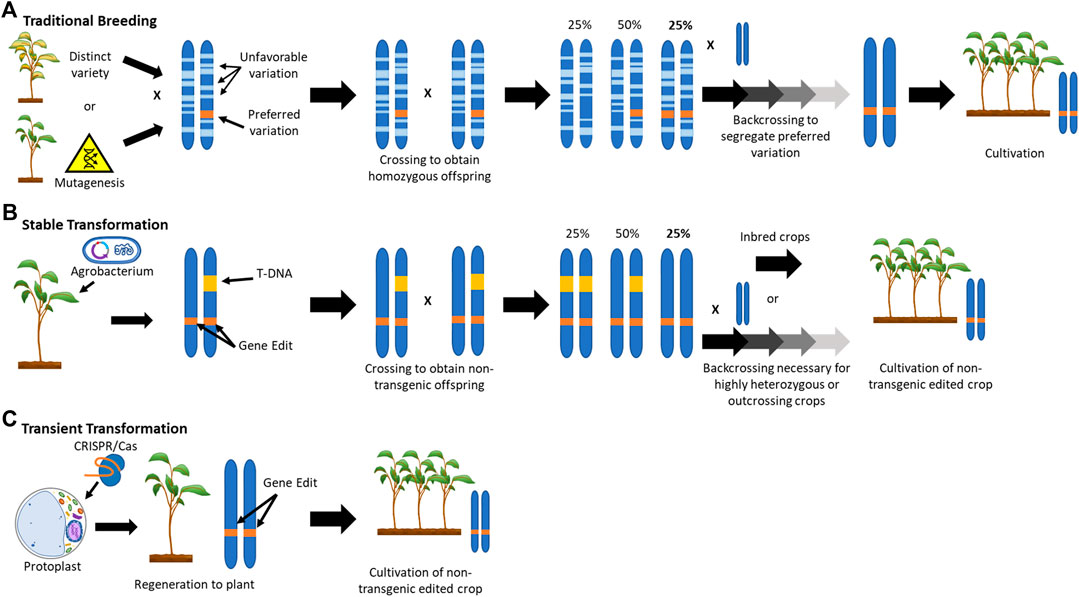
FIGURE 1. Schematic Representation of the Application of New Plant Breeding Technologies.
In contrast to conventional breeding or transgenic CRISPR/Cas9 approaches, gene editing through transient transformation and regeneration of protoplasts can achieve the desired genetic outcome within a single clonal generation by avoiding the integration of foreign DNA into the host genome (Figure 1C). Aside from the potential to speed up the application of NPBT, the use of protoplasts may have numerous other advantages.
As stated above, the use of transient transformation of protoplasts can circumvent transgenesis (the integration of genetic material from one organism into the genome of another organism). The enzymatic removal of the plant cell wall allows for the introduction of foreign DNA, RNA, or protein into protoplasts through either polyethylene glycol (PEG) treatment or electroporation. Although relatively infrequent, the use of DNA (often in the form of plasmids) does not fully preclude the random integration of transgenes (Lin et al., 2018). However, CRISPR/Cas9 can also be expressed through transformation with mRNA encoding the Cas9 enzyme along with the desired guide RNA (gRNA) (Zhang et al., 2016). Alternatively, protoplasts can be transformed with ribonucleoprotein complexes, consisting of Cas9 associated with the gRNA (Svitashev et al., 2016). The latter two approaches more effectively preclude the integration of foreign DNA, although there have been cases where DNA-template contamination in the in vitro transcribed mRNA or gRNA has led to insertions, e.g. (Andersson et al., 2018). Particle bombardment is a potential alternative for transient delivery method for DNA-free gene-editing tools, e.g. (Liang et al., 2018). However, it may suffer from limitations in transformation efficiency and the regeneration of chimeric plants (as discussed below).
If the goal of the gene-editing approach goes beyond site-specific insertions and/or deletions for the knock-out of gene function but instead aims for specific nucleotide substitutions or insertion of a specific sequence through homologous recombination, there is a need for the co-introduction of a DNA-repair template (as in oligo directed mutagenesis) or a donor sequence, respectively. Prime editing and viral replicons are potential methods to deliver such templates and donors transgenically (Čermák et al., 2015; Lin et al., 2020). However, in addition to the potential for a non-transgenic outcome, the use of protoplasts allows for more control over the amount of template or donor delivered and effect higher precision and efficiency, e.g. (Sauer et al., 2016).
In many plant species, the lack of host susceptibility to Agrobacterium transformation limits the use of transgenic NPBT approaches. This is seen in particular in monocots (Hwang et al., 2017). Host-pathogen incompatibility is also expected to be a limiting factor in the applicability of viruses for the delivery of gene-editing tools (Ma et al., 2020). In such cases, the use of protoplasts (or particle bombardment) may be a feasible alternative delivery method.
Chimerism (where only parts of the regenerated plant are descended from an edited cell) can be an issue when using conventional, tissue-culture based approaches where a callus intermediate is used, e.g. (Charrier et al., 2019). This phenomenon occurs because de novo shoots or embryos can be formed from a group of cells rather than a single antecedent. In the case of protoplasts, regenerated plants are (in most cases) derived from a single cell, thereby avoiding this potential problem. Chimerism can be a concern especially when non-selectable, non-transgenic approaches are used together with conventional tissue culture, e.g. transient transformation with Agrobacterium or particle bombardment. Additionally, such non-selectable strategies can suffer from low editing efficiency in the regenerated plants because only the cells on the surface of the tissue are potentially edited whereas regeneration can also occur from the numerous non-transformed cells. In comparison, protoplast transformation efficiencies are much higher and plants regenerated from protoplasts transiently transformed with editing tools will therefore have better chance of being successfully edited.
However, a glaring limitation in the use of protoplasts for NPBT is the challenges faced in the regeneration of plants from single cells and there appears to be no universal strategy that applies to diverse (sub)species. Plant tissue culture in general, and protoplast regeneration in particular, is often lightheartedly considered more of an artform than a science, requiring an experienced eye and instinctual decision making, as comprehensive systematic approaches are too vast in scope to be feasible. In this review, we will discuss a compilation of literature on plant regeneration from protoplasts. We will deliberate protoplast isolation, protoplast culture, and plant regeneration from protoplast culture, specifically in the light of the application of NPBT.
The tissue from which protoplasts are derived is very important for obtaining regenerable starting material. The genotype, organ or tissue, and growth conditions of the plants used can be a significant determinant in regeneration success.
Different cultivars or ecotypes can have widely varying success rates in tissue culture and protoplast regenerative capacity. Depending on the species being worked with and the end goal of the application, it is recommended to assess the regenerative capacity of multiple genotypes and select the most suitable for further use.
When comparing four different Arabidopsis (Arabidopsis thaliana) ecotypes (Col-0, Ws-2, No-0, and HR-10), all gave a similar number of protoplasts with an optimized digestion, but differed significantly when comparing optimal protoplast division media, callus induction media, and shoot induction media (Jeong et al., 2021). Ws-2 showed the highest regeneration efficiency, whereas the Col-0, No-0, and HR-10 had relatively ineffective regeneration rates, regardless of efforts to vary the composition of media and tissue culture methods.
Comparison of three different Cyclamen species (C. graecum, C. mirabile, and C. alpinum) found significant differences in protoplast culture and regeneration, including division frequencies (often referred to as plating efficiency) and morphological appearance of regenerating embryos (Prange et al., 2010a). Plants were regenerated from protoplasts derived from embryogenic callus in all three species, but had different efficiencies in microcallus formation and development of somatic embryos. Interestingly, there was no correlation between the regenerative capacity of the source embryogenic callus and the ability of the protoplasts to divide and regenerate, with C. graecum performing the worst in regeneration from callus but showing the highest protoplast division rates.
Different source materials for protoplast isolation can affect the number, size, viability, and regenerative capacity of protoplasts. There are examples of protoplast isolation and regeneration from numerous tissues, including leaves, cotyledons, roots, petioles, hypocotyls, petals, callus, and suspension cultures (Table 1).
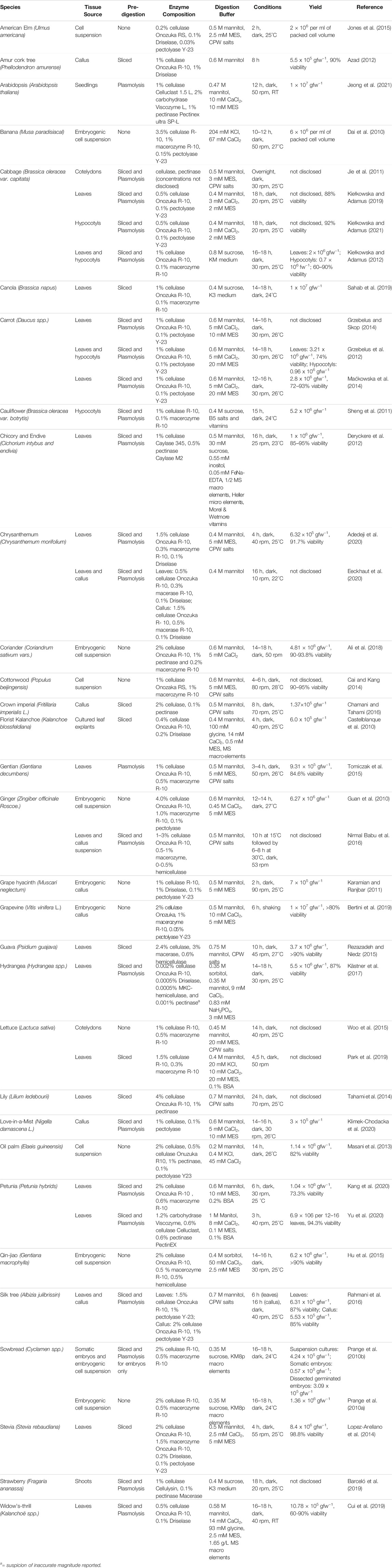
TABLE 1. Obtaining Protoplasts.
In cabbage (Brassica oleracea), it was observed that hypocotyl-derived protoplasts yielded more regenerated shoots than leaf-derived protoplasts (Kiełkowska and Adamus, 2012). In a comparison on the regeneration capacity of protoplasts derived from leaves, cotyledons, and callus from coastal medick (Medicago littoralis), leaf protoplast-derived callus was found to have the highest regeneration capacity with a frequency of 20% and cotyledon protoplast-derived callus had a regeneration frequency of 15% (Zafar et al., 1995). In this study, callus-derived protoplasts developed only a few microcolonies that were not tested for regeneration. Embryogenic callus can potentially provide improved regeneration success in cases where somatic tissues fail to produce regenerable protoplasts, e.g. in grapevine (Vitis vinifera) (Bertini et al., 2019).
The age of the source tissue can also be of importance, both for protoplast yield and viability as well as regeneration success. Generally, protoplasts derived from younger tissues perform better in culture. This has been shown for hypocotyls and leaves in cabbage (Kiełkowska and Adamus, 2012) and cell suspension cultures in oil palm (Elaeis guineensis) (Masani et al., 2013), for example.
The growth conditions of the starting material, including growth media and light, can have a significant effect on the regenerative capacity of protoplasts. An important consideration is that the material needs to be sterile (either grown under aseptic conditions or sterilized upon harvest) in order to be used for further culture of the obtained protoplasts.
In Arabidopsis, plants grown on Gamborg B5 medium and harvested 3 weeks after germination had a larger rosette with nearly twice as many leaves when compared to plant grown on Murashige and Skoog (MS) medium, resulting in twice as many protoplasts per harvested plant. However, during protoplast culture, the plants grown initially on MS media showed two to three times higher plating efficiency. And when comparing the photoperiod under which plants were grown, short day (10 h) resulted in a fourfold higher plating efficiency than long day (16 h) (Masson and Paszkowski, 1992).
Examination of cauliflower (Brassica oleracea) leaf protoplast quality of shoots grown in various vessel types found that protoplast yield, viability, division, and shoot regeneration was higher from tissue of plants grown in containers with vented lids compared to containers with closed lids (Chikkala et al., 2009).
When it comes to isolating protoplasts, it is not only about obtaining a high number of protoplasts, but also about optimizing their viability and regenerative capacity. Many factors in the enzymolysis procedure may be of influence, including the utilized pretreatment, buffer composition, cell-wall digestion enzymes, incubation conditions, and purification methods (Table 1). Although, to our knowledge, there are not studies on the effect on protoplast regeneration directly for all of the different factors described here, it seems reasonable to assume that effects on the quality (viability) of the isolated protoplasts will translate to an influence on regenerative capacity of the isolated protoplasts.
Pretreatment of tissue can be used to augment the number of viable protoplasts isolated by increasing the access of the used enzymes to the plant cell wall. This can be achieved through physical disruption of the tissue (e.g. slicing leaf tissue), vacuum infiltration of the enzyme solution, or a preplasmolysis treatment.
Slicing tissue into smaller sections or strips before moving to the enzyme solution allows for more surface area for the enzymes to work, leading to the release of more protoplasts. With rice (Oryza sativa), longitudinal cutting, parallel to the veins, before enzyme digestion resulted in over twice as many viable protoplast as leaves cut in cross section (Lin et al., 2018). Another example of physical disruption is the “Tape-Arabidopsis Sandwich” method (Wu et al., 2009). This method uses tape on both sides of a leaf to add support and allow the removal of the bottom epidermal layer. This protocol has been successfully applied to other Brassicaceae species, including B. oleracea, B. napus, Cleome spinosa, C. monophilla, and C. gynadra (Lin et al., 2018).
In addition to physical disruption, vacuum infiltration of plant tissue with the enzyme solution can be used to ensure that the enzymes are able to reach more of the cells, which could increase protoplast yield. In both apple (Malus domestica) and grapevine, vacuum infiltration was a part of the optimization of the protoplast isolation procedure to obtain the highest number of viable protoplasts per gram of fresh weight (Osakabe et al., 2018).
Preplasmolysis treatment is used to shrink the protoplasts away from the cell wall before introducing the enzyme solution. This is thought to avoid damage to the cell membrane. When comparing protoplasts isolated from birdsfoot trefoil (Lotus corniculatus) tissue with and without preplasmolysis, the pretreated protoplasts had roughly five times more cell wall formation than the nontreated after 3 days of culture. After 1 week, the viability of the nontreated protoplasts decreased significantly (Vessabutr and Grant, 1995).
The buffer for the enzyme solution is critical for optimal enzyme activity and ensuring a high number of viable protoplasts. The buffer solution typically includes KCl; CaCl2; mannitol, sorbitol, or salts as osmolytes; MES (2-(N-morpholino)ethanesulfonic acid) as pH buffer; BSA (bovine serum albumin) as an alternate target for proteases that may degrade the enzymes; and β-mercaptoethanol as a reducing agent (Table 1). Frearson et al. (1973) first formulated a combination of salts that many still use, called the cell and protoplast washing (CPW) salts. This basal salt solution is often modified with the addition of mannitol or sorbitol for osmotic pressure and different enzymes for optimal protoplast isolation (Jie et al., 2011; Jones et al., 2015; Tomiczak et al., 2015).
Proper osmolality is crucial in order to ensure the survival of the cells and provide an environment for potential cell wall formation and division, leading to regeneration. Protoplast development has been shown to be inhibited by excess osmotic pressure during isolation and culture by impairing metabolism (Ruesink, 1978) as well as division and cell wall regeneration (Pearce and Cocking, 1973).
Enzyme solutions with the same (or similar) composition as the subsequent protoplast culture medium have also been used successfully in protoplast regeneration applications. For example sugar beet (Beta vulgaris) callus protoplasts were isolated using Kao and Michayluk salts in the enzyme solution (Dovzhenko and Koop, 2003); Mango (Mangifera indica) pro-embryogenic mass-derived protoplasts were isolated using an enzyme solution containing Gamborg B5 and Murashige and Skoog salts (Ara et al., 2000); petunia (Petunia spp.) and calibrachoa (Calibrachoa spp.) leaf protoplasts were isolated with Kao and Michayluk and Gamborg B5 salts in the solution (Meyer et al., 2009).
Many commercially available cell-wall degrading enzymes (or enzyme mixtures) are used for the isolation of protoplasts. They differ in their substrates as well as the purity or combination of the enzymes in the extract. Enzymolysis is generally achieved using both cellulases and hemicellulases (e.g. beta-glucanases, xylanases, protopectinases, polygalacturonases, pectin lyases, and pectinesterases). Some of the most commonly used enzymes or enzyme mixtures are Cellulase R-10, Macerozyme R-10, and Pectolyase Y-23 (Table 1). The manufacturer/supplier of the enzymes may be a factor in the success rates (personal experience and communication with others).
The effect of different enzyme combinations and concentrations were tested on the isolation of protoplasts from stevia (Stevia rebaudiana) leaves (Lopez-Arellano et al., 2014). The optimized enzyme solution contained 2% Cellulase R-10, 1.5% Macerozyme Onozuka R-10, 0.2% Driselase, and 0.1% Pectolyase Y-23. When the Cellulase R-10 was decreased to 1% or increased to 3%, there was a significant drop in both the yield and viability of the protoplasts. There was also a lower viability when pectolyase Y-23 was not present. When isolating protoplasts from tobacco (Nicotiana tabacum) leaves, it was found that Pectolyase Y-23 was 20 times more effective than Macerozyme R-10 (Nagata and Ishii, 1979). This was determined to be due to the Pectolyase Y-23 having 50 times stronger endopolygalacturonase activity.
As the cost of lab-grade enzymes can be prohibitive, the use of food-grade cell wall degrading enzymes was investigated as a low-cost alternative for the isolation of switchgrass (Panicum virgatum) leaf protoplasts (Burris et al., 2016). It was determined that using a combination of Rohament CL with Rohapect 10 L and Rohapect UF (cellulases and pectinases commonly used in brewing and juicing) yielded up to 8.4 × 105 protoplasts per gram of leaf tissue.
Although (to our knowledge) there have been no systematic analyses of whether the combination of enzymes used may influence the division rates and regenerative capacity of the produced protoplasts, one can imagine that there could well be an effect. The enzymes themselves, the crude extracts, as well as the cell-wall degradation products they produce can all be recognized by plant cells as pathogenic elicitors, to a greater or lesser extent, depending on the sensitivity of the genotype used to the different enzymes and extracts employed. Protoplast yield and viability may well be a good measure for protoplast isolation, but it could be the case that an enzyme combination that does not necessarily give the highest yield and viability could be more suitable for subsequent regeneration of the protoplasts.
Conditions during protoplast isolation (i.e. duration, temperature, light, and agitation) can play a significant role in the subsequent yield, viability and regenerative capacity of the protoplasts.
The length of a digestion period typically ranges from 2 to 18 h (Table 1). The duration of digestion needs to be long enough to release sufficient numbers of protoplasts, but not too long as to decrease the viability due to cell damage or the lack of nutrients and growth regulators in the enzymolysis solution. For example, when comparing 4, 8, and 12 h digestion duration of crown imperial (Fritillaria imperialis) callus, the yield and viability were highest at 8 h (Chamani and Tahami, 2016).
Temperature also plays an important role in protoplast yield and viability. Room temperature is the most commonly used, although there are examples of higher temperatures being employed (Table 1). There could be effects on enzyme activity (and protoplast yield) as well as protoplast viability and regenerative capacity. Intuitively, it may be preferable to use a temperature that is close to that used for the growth of the source material and/or subsequent protoplast culture conditions, in order to minimize temperature fluctuations or shocks. Conversely, perhaps a particular temperature treatment may actually benefit regenerative capacity.
Digestion in a light or dark condition may additionally influence the protoplast isolation, with most choosing dark conditions (Table 1). This may avoid the production of free radicals and photoinhibition in cells containing chloroplasts. Although there are also examples where digestion under light performed better than in the dark. In geranium (Pelargonium x hortorum) leaf protoplast isolation, protoplast yield and viability were increased when the digestion occurred in light; in the dark, the enzymes were efficient but most of the released protoplasts had burst (Nassour and Dorion, 2002). The protoplasts isolated from the light condition were regenerated into plants, but the effect of light or dark condition during digestion on the regeneration capacity was not investigated.
Agitation of the enzymatic solution on a gyratory shaker during the protoplast digestion can increase the protoplast yields. Typically, speeds range from 0 to 90 rpm, with the average being around 40 rpm (Table 1). Alternatively, the agitation can be implemented only at the end of the digestion period to facilitate the release of protoplasts from the cell wall remnants.
Again, protoplast yield and viability may well be a good measure, but it could be the case that digestion conditions that do not necessarily give the highest yield and viability could be more suitable for subsequent regeneration of the protoplasts.
Following enzymolysis, separation of the protoplasts from undigested tissue, cell wall debris, and dead cells can be an important factor in the culture of the protoplasts. Debris and dead cells may elicit negative effects in the living protoplasts that will inhibit their division and development, e.g. in kalanchoe (Kalanchoe blossfeldiana) (Castelblanque et al., 2010). Filtration and sucrose cushions, or floatation through a density gradient, are commonly used techniques.
Protoplast culture media are central to protoplast division and plant regeneration. The appropriate macro-, micro-nutrients, and additives, such as plant growth regulators, osmotic stabilizers, medium solidifiers, and supplements, are essential in protoplast culture.
Optimal protoplast culture media vary widely, depending on the genotype and source tissue used (Table 2). Common medium formulations (such as MS (Murashige and Skoog, 1962), Gamborg (B5) (Gamborg et al., 1968), Kao and Michayluk (KM (Kao and Michayluk, 1975)), Y3 (Eeuwens, 1976), or Nitsch (Nitsch and Nitsch, 1969)), or slight modification thereof, are often used in protoplast culture. Although there are also examples of custom formulations, e.g. TM2G for tomato (Solanum lycopersicon) protoplast culture (Shahin, 1985). This is also a case where the manufacturer/supplier of the premixed media may be a factor in the success rates (personal experience and communication with others). When establishing and optimizing a protoplast culture procedure, it is prudent to assay an array of medium formulations for suitability.
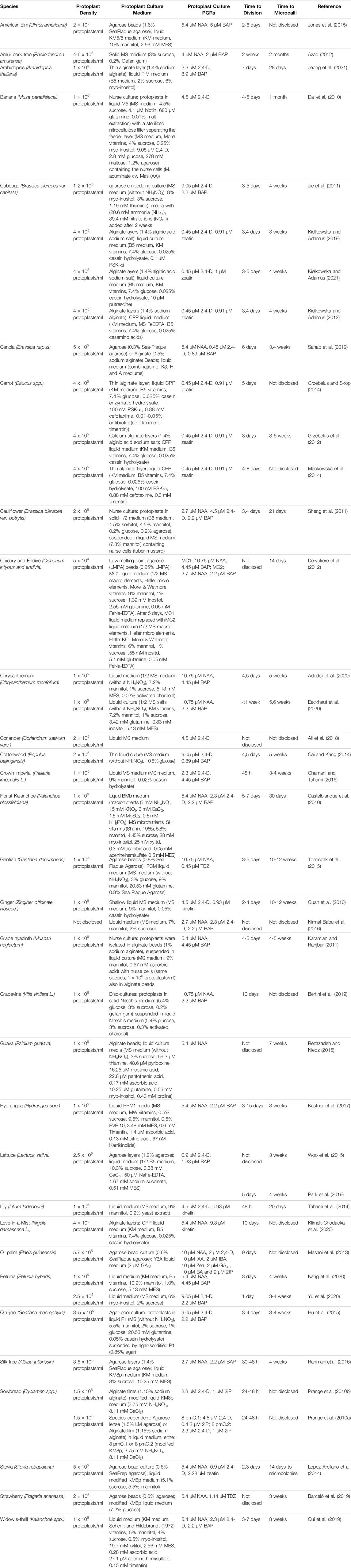
TABLE 2. Protoplast Culture.
In a comparison of 14 formulations based on MS, KM, and Y3 media for oil palm cell suspension-derived protoplast division, Y3-based medium gave the fastest cell wall formation, quickest division, and highest division frequency (Masani et al., 2013). Amur cork tree (Phellodendron amurense) stem protoplasts were cultured in MS, half-strength MS, and Woody Plant Medium (WPM), and culture in full-strength MS medium resulted in the highest colony formation rate (Azad, 2012).
Protoplast cultures also need a carbon source for energy metabolism, typically sucrose or glucose and to a lesser degree mannitol or sorbitol (Table 2). Comparing the effect of 1 and 2% of either glucose or sucrose as the carbon source for chrysanthemum (Chrysanthemum morifolium) leaf protoplast culture, 1% sucrose performed best (Adedeji et al., 2020). Although 2% sucrose resulted in the highest division rate, there was no subsequent colony formation. Only 1% sucrose and 2% glucose led to microcallus formation, with 1% sucrose more rapidly producing larger microcalli. For Arabidopsis seedling protoplast culture, three different variations of supplements with B5 medium and vitamins were tested for protoplast proliferation (Jeong et al., 2021). Myo-inositol as the primary carbon source along with sucrose resulted in the highest proliferation rate across four the different Arabidopsis ecotypes. A simplification of KM8p medium with the removal of all of the sugars (fructose, ribose, xylose, mannose, rhamnose, cellobiose, sorbitol and mannitol) except glucose still resulted in protoplast division that led to callus and embryo formation from carrot (Daucus carota) leaf protoplasts (Grzebelus et al., 2012).
Osmotic pressure is an important aspect of protoplast culture media. Generally, mannitol, sorbitol, sucrose, glucose, myo-inositol or a combination of these components is used to ensure the proper osmolarity. Determining the proper solute concentration is critical for the protoplast survival and division rates. Generally, the concentration of the major osmoticum used in the initial protoplast culture medium varies from 0.1 to 0.8 M (Table 2). Intuitively, it seems that having a comparable osmolarity between enzymolysis and initial culture conditions would expose the protoplasts to less osmotic shock upon transfer to culture medium and benefit their viability and vigor.
For cabbage cotyledon protoplasts, myo-inositol was a better osmotic regulator than mannitol (Jie et al., 2011). It is theorized that myo-inositol may be advantageous to both carbohydrate metabolism in cell walls and inositol metabolism in cell membranes in protoplast culture. However, whether these advantages are gained with a small addition of myo-inositol with a different primary osmoticum or if a large quantity of myo-inositol is needed has yet to be determined.
Osmolarity is commonly decreased gradually as the protoplast reform their cell walls and begin to divide. For example, gradually reducing the osmolarity for oil palm cell suspension protoplast cultures doubled the number of microcalli (Masani et al., 2013). In gentian (Gentiana decumbens) leaf protoplast culture, the osmolarity of the liquid medium around agarose beads was decreased by reducing the mannitol concentration from 0.5 to 0.33 M during the fifth and sixth week of culture, followed by another decrease to 0.17 M mannitol in the seventh and eighth week, and no mannitol for the subsequent weeks (Tomiczak et al., 2015). In chrysanthemum protoplast culture, after the first week in liquid culture medium, myo-inositol was omitted from the refresh medium and mannitol concentrations were dropped from the initial 0.4 M to 0.32, 0.21, and 0.11 M for weeks 2, 3, and 4, respectively (Eeckhaut et al., 2020).
Plant growth regulators, particularly cytokinins and auxins, are essential for the growth of microcalli from protoplasts. Additionally, gibberellic acid (GA3) has been shown to be beneficial in some cases. The most common cytokinins are 6-benzylaminopurine (BAP), zeatin, kinetin, isopentenyl adenine (2iP), and thidiazuron (TDZ). The most common auxins are indole-3-acetic acid (IAA), indole-3-butyric acid (IBA), 2,4-dichlorophenoxyacetic acid (2,4-D), and naphthalene acetic acid (NAA). Optimal concentrations, combinations, and ratios vary widely, depending on the genotype and source tissue of the protoplasts (Table 2).
A ratio of a relatively higher concentration of auxin with a lower concentration of cytokinins was effective for microcallus formation from populus (Populus beijingensis) cell suspension protoplasts (Cai and Kang, 2014). Conversely, in kalanchoe leaf protoplast culture, a higher cytokinin to auxin ratio resulted in better proliferation and microcallus formation; having cytokinin exclusively resulted in slow growth and the microcalli eventually died (Castelblanque et al., 2010).
Coconut water is a natural source of plant growth regulators, both auxin (IAA) and cytokinins (various) as well as other phytohormones, such as gibberellins, and other supplements, such as vitamins and minerals, that have been found to be beneficial in plant tissue culture (Yong et al., 2009). As a supplement in corn (Zea mays) embryogenic callus protoplast culture, coconut water led to a high efficiency of microcallus formation, with a 2% coconut water addition producing the most microcalli (Imbrie-Milligan et al., 1987). Coconut water was also found to increase protoplast cell division in orchid (Phalaenopsis spp.) callus protoplasts (Kobayashi et al., 1993).
Additional supplements, such as polyvinylpyrrolidone, antioxidants, activated charcoal, silver nitrate, antibiotics, complex organics, amino acids, polyamines, conditioned medium, and peptide growth factors, can be added to the media to support protoplast division and microcallus formation (Table 2).
Antioxidants, such as ascorbic acid, citric acid, reduced glutathione, and L-cysteine, can be used to mitigate the inhibitory effects of reactive oxygen species. In oil palm protoplast regeneration, it was found that 200 mg/L ascorbic acid gave the greatest indication of further cell growth and development with the microcalli turning yellow and developing into embryogenic calli (Masani et al., 2013). With this supplementation, two types of embryogenic callus were observed, compact and friable embryogenic callus, which were both able to further develop into somatic embryos and regenerate into plantlets.
Polyvinylpyrrolidone (PVP) is used to adsorb phenolics. While phenolics may be beneficial for plant defense (Bhattacharya et al., 2010), an accumulation during protoplast culture has been found to lead to oxidative browning of the culture medium, inhibiting protoplast growth and division (Reustle and Natter, 1994; Prakash et al., 1997). There has also been reports of PVP suppressing tissue browning and improving callus formation in peony (Paeonia lactiflora) petal explant tissue culture (Cai et al., 2020). Polyvinylpolypyrrolidone (PVPP), a highly cross-linked version of PVP, has also been found to inhibit tissue necrosis in Virginia pine (Pinus virginiana) callus culture (Tang et al., 2004), as well as preventing browning better than PVP in guar (Cyamopsis tetragonoloba) cotyledon protoplast culture (Saxena and Gill, 1986). When PVP was added to the PVPP culture of guar cotyledon protoplasts, not only was it found to enhance the necrosis inhibition, but it also improved the protoplast division frequency. Another compound known to decrease tissue browning is 2-aminoindane-2-phosphonic acid (AIP), which is a reversible inhibitor of phenylalanine ammonia lyase (PAL), an enzyme necessary for polyphenol production (Appert et al., 2003). While the inhibition of PAL was able to increase the cell wall digestibility and facilitate sustained cell division in American elm (Ulmus americana), extended inhibition results in decreased shoot growth in tissue culture (Jones et al., 2012). This decrease in plant growth due to PAL inhibition from AIP has also been seen in birch (Betula pubescens) (Nybakken et al., 2007) and St. John’s wort (Hypericum spp.) (Klejdus et al., 2013). It could be hypothesized that an early addition of AIP will increase the likelihood of protoplast survival, but it should not be used for an extended period as to disrupt the callus and shoot growth, as described for American elm protoplast regeneration (Jones et al., 2015).
Activated charcoal is a commonly used additive employed for its ability to adsorb inhibitory elements, such as phenolics and reactive oxygen species, that can impede protoplast division. Adedeji et al. (2020) found that the ideal concentration of activated charcoal for chrysanthemum leaf protoplast regeneration was 0.02% (w/v) and adding a higher concentration of 0.1% resulted in agglutination of the protoplasts, causing them to die before entering the microcolony stage. In primrose (Primula spp.) cell suspension-derived protoplast culture, the addition of 0.1% PVP did not induce callus formation; however, the addition of activated charcoal did (Mizuhiro et al., 2001).
Silver nitrate (AgNO3), an inhibitor of ethylene action, has been shown in some cases to increase callus formation and regeneration efficiency as well as effect protoplast isolation efficiency. The culture of hypocotyl protoplasts from several Brassica species was markedly improved by the addition of silver nitrate in the culture medium (Pauk et al., 1991; Hu et al., 1999). With rice (Oryza sativa) suspension cultures, the addition of silver nitrate during protoplast isolation reduced protoplast yield but increased the frequency of colony formation (Ishii, 1988).
Antibiotics may be used to avoid endogenous or exogenous contamination, however they can either inhibit or stimulate explant growth and development with the direct causation not yet understood (Qin et al., 2011). A study analyzing the effects of three β-lactam antibiotics (cefotaxime, carbenicillin, and timentin) at different concentrations on carrot seedling protoplasts found that, while plating efficiencies decreased in all antibiotic concentrations higher than 100 mg/L, cefotaxime and timentin in the range of 100–500 mg/L increased regeneration efficiency (Grzebelus and Skop, 2014). Timentin was used with Hydrangea leaf protoplasts to limit the endophytes and it was observed that in antibiotic-free medium, the protoplasts rebuilt the cell wall faster and divided earlier, but callus was only formed in medium with antibiotics (Kästner et al., 2017).
The exact composition of complex organics, such as casein hydrolysate, casamino acids, coconut water, and yeast extract, is typically undefined and varies depending on the manufacturer/supplier and potentially the batch. However, the amino acids, hormones, vitamins, fatty acids, carbohydrates, and other growth supplements they provide may enhance growth and regeneration of plants (Bhatia, 2015). The addition of casein hydrolysate was initially shown to give a more consistent high rate of microcallus formation from tobacco (Nicotiana tabacum) protoplasts (Galun and Raveh, 1975), and is currently an addition to protoplast culture media regularly (Table 2).
Polyamines can regulate plant growth and stress responses through many means, including increasing antioxidant activity and regulating oxidative stresses (Chen et al., 2019). In a comparison of the exogenous addition of the polyamines putrescine, spermidine, and spermine on sugar beet (Beta vulgaris) cell suspension-derived protoplasts, spermine resulted in the highest plating efficiency, likely due to its stronger inhibitory effect on ethylene production (Majewska-Sawka et al., 1997). Polyamines exogenously applied in different concentrations on cabbage hypocotyl protoplast culture obtained the highest frequency of shoot organogenesis from protoplasts treated with putrescine (Kiełkowska and Adamus, 2021). However, the addition of putrescine had no effect on the culture or regeneration of Love-in-a-Mist (Nigella damascena) callus protoplasts (Klimek-Chodacka et al., 2020).
Conditioned medium (spent liquid medium used for cell-suspension cultures that is filtered and subsequently used as a supplement for protoplast culture) may contain compounds that encourage growth and mitotic activity. Fresh conditioned medium from cell-suspension cultures significantly increased the plating efficiency in chrysanthemum leaf protoplast culture (Zhou et al., 2005).
Phytosulfokine (PSK), specifically PSK-α, is a peptide that was originally detected secreted in conditioned medium, but was later found in whole plants (Yang et al., 1999). It was found to promote cell growth, enhance callus growth as well as adventitious root and bud formation, and improve somatic embryogenesis in multiple species, and has also been shown to enhance protoplast regeneration in carrot (Maćkowska et al., 2014) and cabbage (Kiełkowska and Adamus, 2019). With carrot leaf protoplasts, application of PSK-α during the initial culture resulted in a four-fold increase in regenerated plants (Maćkowska et al., 2014). PSK-α was shown to be both genotype- and dose-dependent and did not require a constant presence to maintain cell divisions in cabbage leaf protoplasts (Kiełkowska and Adamus, 2019). Not only was the PSK-α found to promote cell proliferation, but it also increased differentiation and organogenesis in five of the six cabbage accessions tested.
Protoplast culture conditions, such as the use of liquid or semi-solid medium, temperature and light, cell density, or the presence of nurse cultures, can have a significant effect on the division and microcallus formation potential of protoplasts.
When it comes to determining the solidity of the media to use with protoplast culturing, there are multiple factors to consider, including imaging potential, media refreshing, toxin accumulation, and cell aggregation.
Liquid medium is the most straightforward to make since it requires no agar manipulation. However, it faces a multitude of challenges. With imaging, unless each cell is in a separate space, it is impossible to track the growth of an individual cell. There is also the potential for aggregation of cells to form a non-homogeneous callus, possibly resulting in chimerism of the regenerated plants. Aggregation can also cause a local accumulation of toxic substances released from dying cells that may inhibit the growth of neighboring cells (Deryckere et al., 2012).
To avoid cell agglutination, embedding the protoplasts in semi-solid medium can ensure physical separation of cells. The embedding medium will typically contain agar, agarose, or alginate as a solidifier. Alginate is favorable for heat-sensitive protoplasts because the gelling is induced by exposure to calcium ions rather than the need to heat the agar or agarose solutions above the melting point.
In a comparison between thin alginate layers and extra thin alginate films on carrot shoot protoplast culture, thin alginate layers resulted in nearly a 20% increase in plating efficiency in every accession tested (Maćkowska et al., 2014). Sterilizing the alginate solution through filter-sterilization was also found to give over a 10% increase in plating efficiency over autoclave-sterilization in several of the accessions used.
The amount of liquid medium surrounding alginate beads can affect the protoplast proliferation capability. In American elm (Ulmus americana) cell suspension-derived protoplast alginate bead culture, cultures that contained less than 2 ml or more than 3 ml of liquid medium failed to develop beyond the first cell division; whereas cultures that contained 2 or 3 ml of liquid medium continued to proliferate (Jones et al., 2015).
The temperature and light conditions used during protoplast culture vary widely (Table 2) and have both been shown to be of effect in regeneration success. Cabbage leaf protoplast cultures were greatly affected by light and temperature, with very few divisions occurring in cultures moved from dark at 25°C to light at 23°C after 7 days of culture, compared to those kept in the dark conditions for all 15 days (Kaur et al., 2006). Using lettuce (Lactuca saligna) leaf protoplasts, dark culture led to sustained division while light bleached and killed the protoplasts in 3 days (Brown et al., 1987). However, Arabidopsis cotyledon protoplasts did not show a significant variation in either the plating density or growth rates whether cultured in the light or dark (Dovzhenko et al., 2003).
The protoplast plating density can range from single cells up to a few million protoplasts per milliliter, but typically range from 5 × 104–1 × 106 protoplasts/ml (Table 2). In a comparison of plating densities of petunia (Petunia hybrida) leaf protoplast culture, 1 × 106 protoplasts/ml produced a significantly higher division frequency and number of calli than 5 × 104 protoplasts/ml (Kang et al., 2020). However, the microcolony viability decreased with the plating density increasing to 1.5 × 106 protoplasts/ml, potentially due to high phenolics accumulation. Over-crowding the protoplasts can also result in a lower viability due to a lack of available nutrients (Kiełkowska and Adamus, 2012). In contrast, a lower density may also be desired to track an individual protoplast after transformation or fusion (Bhojwani and Dantu, 2013). However, a lower protoplast density can be more costly and time consuming. Additionally, protoplasts can release growth factors which can stimulate mitotic division non-cell-autonomously. This is also the basis for nurse cultures.
Nurse cultures are the culture of target protoplasts with additional actively dividing protoplasts or suspension cells, either from the same species (e.g. in crocus (Crocus cancellatus) embryogenic calli-derived protoplast culture (Karamian and Ebrahimzadeh, 2001)) or from another, often closely related species (e.g. in desert banana (Musa paradisiacal) embryonic cell suspension protoplast culture (Dai et al., 2010) and cauliflower (Brassica oleracea var. botrytis) hypocotyl protoplast culture (Sheng et al., 2011)). There are many nurse culture techniques, one example is feeder layer-cultures, which can be embedding the target protoplasts in agar layers with the nurse cells in a liquid surrounding the layers (Sheng et al., 2011), or the target protoplasts in liquid culture with the nurse cells embedded in agarose (Dai et al., 2010). Alginate bead cultures, which can be performed by embedding the target protoplasts in alginate beads and having the nurse cells in liquid medium (e.g. with rice (Oryza sativa) suspension culture protoplasts (Kyozuka et al., 1987)). An alternate method for ensuring a separation of the nurse cells and the target protoplasts is using a nitrocellulose filter which allows growth factors, signaling molecules, and nutrients to pass through, but not cells (Dai et al., 2010).
From microcalli, regeneration could come from organogenesis or embryogenesis. Organogenesis-oriented microcalli can be moved to a callus proliferation medium to increase the callus size, whereas embryogenesis-oriented microcalli can be moved to embryo formation medium; however, either could also proliferate callus or form embryos on the microcallus medium, depending on the genotype, source tissue, and medium composition.
Organogenesis typically relies on moving callus to a medium containing both a cytokinin and auxin or a shooting medium followed by a rooting medium. When it comes to the timeframe for regeneration, it is difficult to directly compare organogenesis and embryogenesis between different species and source tissues (Table 3). Intuitively, embryogenesis should take less time than organogenesis due to the extended time the callus needs to shoot and then root versus an embryo’s ability to grow and differentiate both organs at the same time.
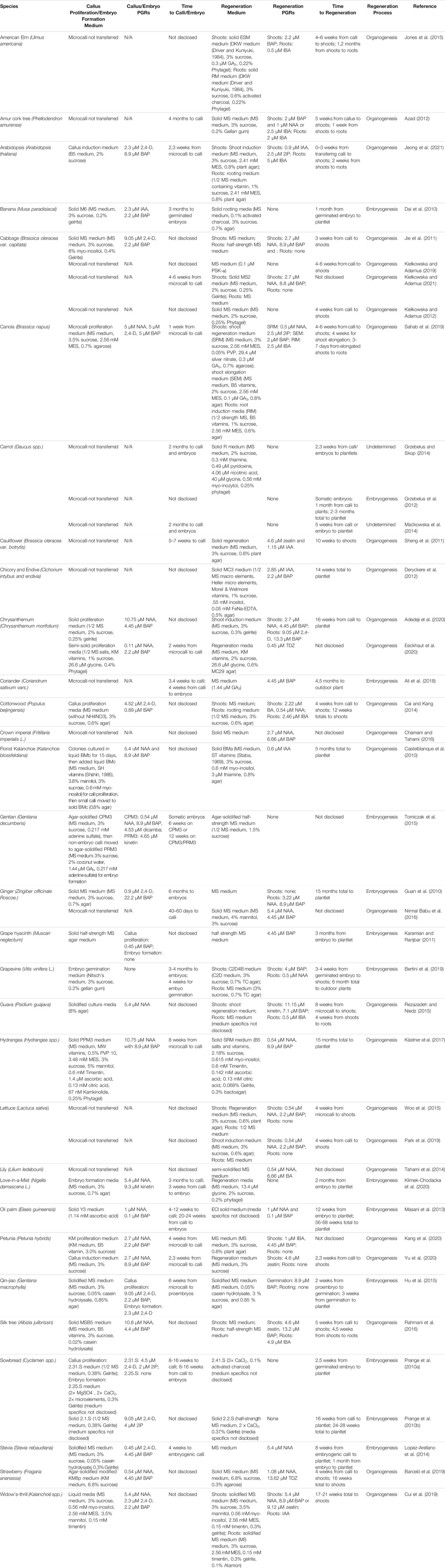
TABLE 3. Regeneration from Protoplast Culture.
Embryogenic callus formation can be from somatic protoplasts (somatic embryogenesis) or from embryogenic callus-derived protoplasts (secondary embryogenesis). Embryogenesis relies on cells within the microcalli presenting embryogenic properties, i.e. isodiametric, cytoplasm-rich cells (Dai et al., 2010). The embryogenic microcalli can then proliferate into embryogenic callus or form embryos directly. Embryos that form from the (micro)callus can be moved to media for germination and plantlet maturation.
When it comes to regenerating plants from protoplast-derived callus, either embryogenic or somatic callus, the media composition can determine the efficiency of the regeneration. A majority of methods use solid MS media supplemented with auxin and cytokinin (Table 3). Typically, shooting is the primary goal with rooting coming shortly after, then planting in soil for maturation. It is generally easier to get roots from shoots than shoots from roots.
In cabbage leaf protoplast shoot regeneration, MS versus Gamborg B5 based media supplemented with PSK-α and with or without plant growth regulators was compared (Kiełkowska and Adamus, 2019). Microcolonies were freed from alginate layers and, after transferring to regeneration medium, the callus would turn green, remain white, or begin to brown. The browning callus was considered dead, the white callus grew slightly but did not form shoots, and the green callus led to shoot regeneration roughly 4–6 weeks after transfer. It was found that the highest shoot regeneration came from callus placed on MS media with PSK-α and without PGRs across a majority of the genotypes tested.
When determining the effect of cytokinin on shoot induction from guava (Psidium guajava) leaf protoplast-derived callus, BAP and kinetin concentrations were investigated (Rezazadeh and Niedz, 2015). Concentrations of 7.1 μM BAP and 11.15 μM kinetin were optimal for shoot production; a higher concentration did not significantly increase the number of shoots. It was also found that changing the kinetin level was more effective than BAP.
Some methods involve the addition of other supplements to the regeneration medium to assist the callus growth and differentiation. Activated charcoal is a common addition, with its ability to prevent browning of callus by adsorbing growth inhibitors (Prange et al., 2010a; Masani et al., 2013). Masani et al. (2013) also examined the effects of ascorbic acid to reduce discoloration and promote embryogenesis. They found that ascorbic acid increased the number of embryogenic calli which subsequentially improved the regeneration efficiency of oil palm embryogenic cell suspension-derived protoplasts.
Somaclonal variation is the genetic or phenotypic variation that occurs in plants from tissue culture. A phenotypic change can be explained by either a genetic or epigenetic modification. Somaclonal variation can influence the fertility of the regenerant as well as the potential for changing the ploidy level, which is crucial for breeding.
Somaclonal variation is a potential occurrence in protoplast regeneration that can reveal itself in morphological or ploidy variation (Prange et al., 2010b; Sheng et al., 2011; Grzebelus et al., 2012; Tomiczak et al., 2015; Barceló et al., 2019). In strawberry (Fragaria ananassa), morphological differences between the control and regenerated protoclones were observed (including plant size and leaflets per leaf) that were not explained by ploidy level changes but rather genetic variation detected by microsatellite markers (Barceló et al., 2019). Prange et al. (2010b) and Tomiczak et al. (2015) both collected regenerated plants that were tetraploid from protoplasts that were initially diploid. In Cyclamen coum, it was observed that a single callus would give rise to both tetraploid and diploid regenerants which was reasoned could be a result from either the chromosomes doubling during callus culture or an error in separation of callus during culturing (Prange et al., 2010b). With Gentiana decumbens, there was no morphological difference in the regenerants, besides wider leaf blades (Tomiczak et al., 2015), yet 100% of the regenerated plants were tetraploid.
When considering the culture method’s role in this somaclonal variation, one hypothesis is that if genome duplication occurred during protoplast culture, it is most likely due to the possibility that tetraploid protoplasts divide faster than diploid protoplasts, as shown in tobacco (Nicotiana plumbaginifolia) (Magnien et al., 1982) and rapeseed (Brassica napus) (Magnien et al., 1982; Chen et al., 1994). If the polyploidization occurred during callus formation, the hypothesis is endoreduplication (amplification of DNA without mitosis) in callus cells, shown previously in pea (Pisum sativum) (Ochatt et al., 2000) and barrelclover (Medicago truncatula) (Elmaghrabi and Ochatt, 2006) and would explain this increase of DNA content. It has also been shown that plant growth regulators typically added to protoplast culture media have an effect on endoreduplication frequency in sugar beet (Beta vulgaris) (Lukaszewska et al., 2012).
Time in tissue culture increases chances of somaclonal variation. Isolating protoplast from plant tissue may therefore be favorable over isolating from callus tissue in order to avoid somaclonal variation due to the additional in vitro step that is required to obtain callus. This additional step has the potential to introduce genetic variation and effect the protoplast regeneration efficiency.
While somaclonal variation is undesirable in commercial crop production, it does have the benefit of creating phenotypic variability with a large number of regenerants that can be obtained through protoplast regeneration. This gives the potential for the identification of mutations that could be beneficial for a variety of uses, such as biotic resistance (Grzebelus et al., 2013), abiotic resistance (Kiełkowska et al., 2019), or create a desirable ornamental property.
Electroporation as a method for protoplast transformation is not as popular as PEG-mediated transformation. With electroporation, there are more factors to consider that potentially have effects on transfection efficiency and cell survival: pulse voltage, pulse length, pulse number, cell number, DNA concentration, and electroporation buffer composition (Lee et al., 2020). However, when optimized, electroporation can be very efficient. Lee et al. (2020) found that when electroporation transformation was optimized for cabbage protoplasts, the transformation efficiency was nearly double that of PEG-mediated delivery, although both transformation rates were low (3.4 and 1.8%, respectively). Wójcik and Rybczyński (2015) studied the effect of electroporation the culture of embryogenic cell suspension-derived protoplasts from gentian (Gentiana kurroo). A high electric field voltage over 1 kV/cm significantly decreased protoplast survival and division. A single pulse had nine-fold higher protoplast viability than two pulses. Comparing the effect of length of the electric pulse on protoplast viability, it was found that 5 ms completely killed the protoplasts and 40 μs was too long and resulted in no division of the protoplast. A 20 μs pulse had the highest protoplast viability and division, 70 and 44.5% respectively. Significantly higher protoplast viability was obtained with an electroporation buffer with KCl, higher MgCl2 and pH, and lower MES (Wójcik and Rybczyński, 2015).
The more common PEG-mediated transformation requires less materials than electroporation but does require chemicals that could potentially damage the protoplasts. The main factors to consider with regards to transformation efficiency and cell survival are PEG concentration, transfection time, DNA concentration, and cell number (which has previously been shown to influence the results (Burris et al., 2016)). Transformation with PEG can reach a high transformation rate, such as 90% in petunia leaf protoplasts (Subburaj et al., 2016) and 80% in both wheat leaf protoplasts and rice sheath protoplasts (Shan et al., 2013). Although, a high transformation rate does not translate to a large number of transformed regenerants. For example, petunia leaf protoplasts transiently transformed with PEG for CRISPR/Cas9 ribonuclear protein multiplexing of two genes had a 55% transfection efficiency, but only eight of the 67 regenerated plants (11.9%) had indel mutations (Yu et al., 2020). PEG-mediated transformation of potato (Solanum tuberosum) leaf protoplasts resulted in more callus formation when treated with 12.5% PEG than 20% PEG; however, even the 12.5% PEG treatment resulted in a ten-fold decrease in callus formation compared to the untreated control (Craig et al., 2005).
In our opinion, the use of protoplast regeneration in NPBT has a promising future. It has been used in numerous applications of gene-editing for crop trait improvement; e.g. the knock-out of the BRASSINOSTEROID INSENSITIVE 2 (BIN2) gene in lettuce (Lactuca sativa) (Woo et al., 2015) and the granule bound starch synthase (GBSS) gene in potato (Andersson et al., 2018) or the oligo-directed mutagenesis of the 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) gene in flax (Linum usitatissimum) (Sauer et al., 2016). We expect to see many more examples of its successful application in the coming years.
Nonetheless, there are obstacles that need to be addressed in order to overcome some of the challenges associated with protoplast regeneration. It is a process that demands specialized tissue culture expertise, requires complex manipulation, and can be time-consuming. Overall, current methods for protoplast regeneration are very genotype-specific and need to be made more universal for increased applicability and success.
One potential approach for making protoplast regeneration universally available is to gain fundamental knowledge of the transcriptional regulation of the regeneration process via transcriptomic analysis. While transcriptomic analysis of protoplast culture (e.g. in moss (Physcomitrella patens) protonema protoplasts for the initial 72 h of culture (Xiao et al., 2012)) has previously been investigated, there is a lack of and difficulty in knowing the transcriptional activity of solely protoplasts destined for regeneration. Single-cell transcriptome profiling has been demonstrated (Shulse et al., 2019), but the question remains on how to differentiate between protoplasts with regeneration capability and the larger, doomed protoplast population. Additional challenges arise when taking the cell-type composition of the source organ as well as the genotype into account.
Another process that can potentially improve universal application of protoplast regeneration technologies is through ectopic expression of embryogenic or morphogenic factors. Theoretically, if an ample number of protoplasts can directly develop into embryos, the regeneration frequency would multiply, resulting in a large number of regenerated plantlets. The direct development of protoplasts into embryos could also decrease the time in tissue culture, reducing the potential of somaclonal variation. The embryogenic or morphogenic transcription factors would need be to be transiently expressed in order to avoid any developmental effects that constitutive expression may cause (e.g. ectopic expression of BABY BOOM causing embryogenic growth on vegetative tissue (Boutilier et al., 2002)). Identification of appropriate embryogenic or morphogenic transcription factors, which could function individually or as a collective, as well as the timing of expression would need to be investigated. Recently, a study using Arabidopsis mesophyll protoplasts showed that timed transcriptional activation of auxin biosynthesis can significantly enhance regeneration success (Sakamoto et al., 2021). It will be interesting to see whether this approach is applicable to divergent species.
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
Research in the Bargmann lab is supported in part by the USDA National Institute of Food and Agriculture, Hatch project VA-160133, Multistate S-009 project VA-136377, the Virginia Tech School of Plant and Environmental Sciences, and the Virginia Tech Open Access Subvention Fund.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Adedeji, O. S., Naing, A. H., and Kim, C. K. (2020). Protoplast Isolation and Shoot Regeneration from Protoplast-Derived Calli of Chrysanthemum Cv. White ND. Plant Cel. Tiss. Organ. Cult 141, 571–581. doi:10.1007/s11240-020-01816-3
Ali, M., Mujib, A., Zafar, N., and Tonk, D. (2018). Protoplast Isolation and Plant Regeneration in Two Cultivated Coriander Varieties, Co-1 and RS. bta 99, 345–355. doi:10.5114/bta.2018.79965
Andersson, M., Turesson, H., Olsson, N., Fält, A.-S., Ohlsson, P., Gonzalez, M. N., et al. (2018). Genome Editing in Potato via CRISPR-Cas9 Ribonucleoprotein Delivery. Physiol. Plantarum 164, 378–384. doi:10.1111/ppl.12731
Appert, C., Zoń, J., and Amrhein, N. (2003). Kinetic Analysis of the Inhibition of Phenylalanine Ammonia-Lyase by 2-Aminoindan-2-Phosphonic Acid and Other Phenylalanine Analogues. Phytochemistry 62, 415–422. doi:10.1016/s0031-9422(02)00561-7
Ara, H., Jaiswal, U., and Jaiswal, V. S. (2000). Plant Regeneration from Protoplasts of Mango ( Mangifera Indica L.) through Somatic Embryogenesis. Plant Cel. Rep. 19, 622–627. doi:10.1007/s002990050783
Azad, M. A. K. (2012). Plant Regeneration through Callus-Derived Protoplasts of Phellodendron Amurense Rupr. BioTechnology 6, 317–326.
Barceló, M., Wallin, A., Medina, J. J., Gil-Ariza, D. J., López-Casado, G., Juarez, J., et al. (2019). Isolation and Culture of Strawberry Protoplasts and Field Evaluation of Regenerated Plants. Scientia Horticulturae 256, 108552. doi:10.1016/j.scienta.2019.108552
Bertini, E., Tornielli, G. B., Pezzotti, M., and Zenoni, S. (2019). Regeneration of Plants from Embryogenic Callus-Derived Protoplasts of Garganega and Sangiovese grapevine (Vitis vinifera L.) Cultivars. Plant Cel. Tiss. Organ. Cult. 138, 239–246. doi:10.1007/s11240-019-01619-1
Bhatia, S. (2015). “Plant Tissue Culture,” in Modern Applications of Plant Biotechnology in Pharmaceutical Sciences. Editors S. Bhatia, K. Sharma, R. Dahiya, and T. Bera (Boston: Academic Press), 31–107. doi:10.1016/b978-0-12-802221-4.00002-9
Bhattacharya, A., Sood, P., and Citovsky, V. (2010). The Roles of Plant Phenolics in Defence and Communication during Agrobacterium and Rhizobium Infection. Mol. Plant Pathol. 11, 705–719. doi:10.1111/j.1364-3703.2010.00625.x
Bhojwani, S. S., and Dantu, P. K. (2013). “Parasexual Hybridization,” in Plant Tissue Culture: An Introductory Text. Editors S. S. Bhojwani, and P. K. Dantu (India: Springer), 173–198. doi:10.1007/978-81-322-1026-9_14
Boutilier, K., Offringa, R., Sharma, V. K., Kieft, H., Ouellet, T., Zhang, L., et al. (2002). Ectopic Expression of BABY BOOM Triggers a Conversion from Vegetative to Embryonic Growth. Plant Cell 14, 1737–1749. doi:10.1105/tpc.001941
Brown, C., Lucas, J. A., and Power, J. B. (1987). Plant Regeneration from Protoplasts of a Wild Lettuce Species (Lactuca Saligna L.). Plant Cel. Rep. 6, 180–182. doi:10.1007/bf00268472
Bull, S. E., Alder, A., Barsan, C., Kohler, M., Hennig, L., Gruissem, W., et al. (2017). FLOWERING LOCUS T Triggers Early and Fertile Flowering in Glasshouse Cassava (Manihot Esculenta Crantz). Plants (Basel) 6. doi:10.3390/plants6020022
Burris, K. P., Dlugosz, E. M., Collins, A. G., Stewart, C. N., and Lenaghan, S. C. (2016). Development of a Rapid, Low-Cost Protoplast Transfection System for Switchgrass (Panicum Virgatum L.). Plant Cel. Rep. 35, 693–704. doi:10.1007/s00299-015-1913-7
Cai, X., Wei, H., Liu, C., Ren, X., Thi, L. T., and Jeong, B. R. (2020). Synergistic Effect of NaCl Pretreatment and PVP on Browning Suppression and Callus Induction from Petal Explants of Paeonia Lactiflora Pall. ‘Festival Maxima'. Plants (Basel) 9, 13. doi:10.3390/plants9030346
Cai, X., and Kang, X.-Y. (2014). Plant Regeneration from Cell Suspension-Derived Protoplasts of Populus × Beijingensis. In Vitro Cell.Dev.Biol.-Plant 50, 92–98. doi:10.1007/s11627-013-9540-x
Castelblanque, L., García-Sogo, B., Pineda, B., and Moreno, V. (2010). Efficient Plant Regeneration from Protoplasts of Kalanchoe Blossfeldiana via Organogenesis. Plant Cel. Tiss. Organ. Cult. 100, 107–112. doi:10.1007/s11240-009-9617-8
Čermák, T., Baltes, N. J., Čegan, R., Zhang, Y., and Voytas, D. F. (2015). High-frequency, Precise Modification of the Tomato Genome. Genome Biol. 16, 232.
Chamani, E., and Tahami, S. k. (2016). Efficient Protocol for Protoplast Isolation and Plant Regeneration of Fritillaria Imperialis L. J. Agric. Sci. Technol. 18, 467–482.
Charrier, A., Vergne, E., Dousset, N., Richer, A., Petiteau, A., and Chevreau, E. (2019). Efficient Targeted Mutagenesis in Apple and First Time Edition of Pear Using the CRISPR-Cas9 System. Front. Plant Sci. 10, 40. doi:10.3389/fpls.2019.00040
Chen, D., Shao, Q., Yin, L., Younis, A., and Zheng, B. (2019). Polyamine Function in Plants: Metabolism, Regulation on Development, and Roles in Abiotic Stress Responses. Front. Plant Sci. 9, 1945. doi:10.3389/fpls.2018.01945
Chen, Z., Hsiao, K.-C., and Bornman, C. H. (1994). Ploidy and Division Efficiency of Petiolar Protoplasts of Brassica Napus. Hereditas 120, 41–46.
Chikkala, V. R. N., Nugent, G. D., Dix, P. J., and Stevenson, T. W. (2009). Regeneration from Leaf Explants and Protoplasts of Brassica oleracea Var. Botrytis (Cauliflower). Scientia Horticulturae 119, 330–334. doi:10.1016/j.scienta.2008.07.036
Craig, W., Gargano, D., Scotti, N., Nguyen, T. T., Lao, N. T., Kavanagh, T. A., et al. (2005). Direct Gene Transfer in Potato: A Comparison of Particle Bombardment of Leaf Explants and PEG-Mediated Transformation of Protoplasts. Plant Cel. Rep. 24, 603–611. doi:10.1007/s00299-005-0018-0
Cui, J., Kuligowska Mackenzie, K., Eeckhaut, T., Müller, R., and Lütken, H. (2019). Protoplast Isolation and Culture from Kalanchoë Species: Optimization of Plant Growth Regulator Concentration for Efficient Callus Production. Plant Cel. Tiss. Organ. Cult. 138, 287–297. doi:10.1007/s11240-019-01624-4
Dai, X.-M., Xiao, W., Huang, X., Zhao, J.-T., Chen, Y.-F., and Huang, X.-L. (2010). Plant Regeneration from Embryogenic Cell Suspensions and Protoplasts of Dessert Banana Cv. 'Da Jiao' (Musa Paradisiacal ABB Linn.) via Somatic Embryogenesis. In Vitro Cell.Dev.Biol.-Plant 46, 403–410. doi:10.1007/s11627-010-9314-7
Deryckere, D., Eeckhaut, T., Van Huylenbroeck, J., and Van Bockstaele, E. (2012). Low Melting point Agarose Beads as a Standard Method for Plantlet Regeneration from Protoplasts within the Cichorium Genus. Plant Cel. Rep. 31, 2261–2269. doi:10.1007/s00299-012-1335-8
Dovzhenko, A., Dal Bosco, C., Meurer, J., and Koop, H. U. (2003). Efficient Regeneration from Cotyledon Protoplasts in Arabidopsis thaliana. Protoplasma 222, 107–111. doi:10.1007/s00709-003-0011-9
Dovzhenko, A., and Koop, H.-U. (2003). Sugarbeet ( Beta Vulgaris L.): Shoot Regeneration from Callus and Callus Protoplasts. Planta 217, 374–381. doi:10.1007/s00425-003-1006-7
Driver, J. A., and Kuniyuki, A. H. (1984). In Vitro propagation of Paradox walnut Rootstock [Juglans Hindsii X Juglans Regia, Tissue Culture]. Hort. Sci. 19, 507–509.
Eeckhaut, T., Van Houtven, W., Bruznican, S., Leus, L., and Van Huylenbroeck, J. (2020). Somaclonal Variation in Chrysanthemum × Morifolium Protoplast Regenerants. Front. Plant Sci. 11, 607171. doi:10.3389/fpls.2020.607171
Eeuwens, C. J. (1976). Mineral Requirements for Growth and Callus Initiation of Tissue Explants Excised from Mature Coconut Palms (Cocos Nucifera) and Cultured In Vitro. Physiol. Plant 36, 23–28. doi:10.1111/j.1399-3054.1976.tb05022.x
Elmaghrabi, A., and Ochatt, S. (2006). Isoenzymes and Flow Cytometry for the Assessment of True-To-Typeness of Calluses and Cell Suspensions of Barrel Medic Prior to Regeneration. Plant Cel. Tiss. Organ. Cult 85, 31–43. doi:10.1007/s11240-005-9046-2
Frearson, E. M., Power, J. B., and Cocking, E. C. (1973). The Isolation, Culture and Regeneration of Petunia Leaf Protoplasts. Developmental Biol. 33, 130–137. doi:10.1016/0012-1606(73)90169-3
Galun, E., and Raveh, D. (1975). In Vitro culture of Tobacco Protoplasts: Survival of Haploid and Diploid Protoplasts Exposed to X-ray Radiation at Different Times after Isolation. Radiat. Bot. 15, 79–82. doi:10.1016/s0033-7560(75)80017-2
Gamborg, O. L., Miller, R. A., and Ojima, K. (1968). Nutrient Requirements of Suspension Cultures of Soybean Root Cells. Exp. Cel. Res. 50, 151–158. doi:10.1016/0014-4827(68)90403-5
Grzebelus, E., Kruk, M., Macko-Podgórni, A., and Grzebelus, D. (2013). Response of Carrot Protoplasts and Protoplast-Derived Aggregates to Selection Using a Fungal Culture Filtrate of Alternaria Radicina. Plant Cel. Tiss. Organ. Cult. 115, 209–222. doi:10.1007/s11240-013-0353-8
Grzebelus, E., and Skop, L. (2014). Effect of β-lactam Antibiotics on Plant Regeneration in Carrot Protoplast Cultures. In Vitro Cell.Dev.Biol.-Plant 50, 568–575. doi:10.1007/s11627-014-9626-0
Grzebelus, E., Szklarczyk, M., and Baranski, R. (2012). An Improved Protocol for Plant Regeneration from Leaf- and Hypocotyl-Derived Protoplasts of Carrot. Plant Cel. Tiss. Organ. Cult. 109, 101–109. doi:10.1007/s11240-011-0078-5
Guan, Q., Guo, Y., Wei, Y., Meng, F., and Zhang, Z. (2010). Regeneration of Somatic Hybrids of Ginger via Chemical Protoplast Fusion. Plant Cel. Tiss. Organ. Cult. 102, 279–284. doi:10.1007/s11240-010-9730-8
Hu, Q., Andersen, S. B., and Hansen, L. N. (1999). Plant Regeneration Capacity of Mesophyll Protoplasts from Brassica Napus and Related Species. Plant Cel. Tissue Organ. Cult. 59, 189–196. doi:10.1023/a:1006417530587
Hu, X., Yin, Y., and He, T. (2015). Plant Regeneration from Protoplasts of Gentiana Macrophylla Pall. Using agar-pool Culture. Plant Cel. Tiss. Organ. Cult. 121, 345–351. doi:10.1007/s11240-014-0705-z
Hwang, H.-H., Yu, M., and Lai, E.-M. (2017). Agrobacterium-mediated Plant Transformation: Biology and Applications. Arab 15. doi:10.1199/tab.0186
Imbrie-Milligan, C., Kamo, K. K., and Hodges, T. K. (1987). Microcallus Growth from maize Protoplasts. Planta 171, 58–64. doi:10.1007/bf00395067
Ishii, S. (1988). Factors Influencing Protoplast Viability of Suspension-Cultured Rice Cells during Isolation Process. Plant Physiol. 88, 26–29. doi:10.1104/pp.88.1.26
Jeong, Y. Y., Lee, H.-Y., Kim, S. W., Noh, Y.-S., and Seo, P. J. (2021). Optimization of Protoplast Regeneration in the Model Plant Arabidopsis thaliana. Plant Methods 17, 21. doi:10.1186/s13007-021-00720-x
Jie, E.-Y., Kim, S.-W., Jang, H.-R., In, D.-S., and Liu, J.-R. (2011). Myo-inositol Increases the Plating Efficiency of Protoplast Derived from Cotyledon of Cabbage (Brassica oleracea Var. Capitata). J. Plant Biotechnol. 38, 69–76. doi:10.5010/jpb.2011.38.1.069
Jones, A. M. P., Chattopadhyay, A., Shukla, M., Zoń, J., and Saxena, P. K. (2012). Inhibition of Phenylpropanoid Biosynthesis Increases Cell wall Digestibility, Protoplast Isolation, and Facilitates Sustained Cell Division in American Elm (Ulmus Americana). BMC Plant Biol. 12, 75. doi:10.1186/1471-2229-12-75
Jones, A. M. P., Shukla, M. R., Biswas, G. C. G., and Saxena, P. K. (2015). Protoplast-to-plant Regeneration of American Elm (Ulmus Americana). Protoplasma 252, 925–931. doi:10.1007/s00709-014-0724-y
Kang, H. H., Naing, A. H., and Kim, C. K. (2020). Protoplast Isolation and Shoot Regeneration from Protoplast-Derived Callus of Petunia Hybrida Cv. Mirage Rose. Biology 9, 228. doi:10.3390/biology9080228
Kao, K. N., and Michayluk, M. R. (1975). Nutritional Requirements for Growth of Vicia Hajastana Cells and Protoplasts at a Very Low Population Density in Liquid media. Planta 126, 105–110. doi:10.1007/bf00380613
Karamian, R., and Ebrahimzadeh, H. (2001). Plantlet Regeneration from Protoplast-Derived Embryogenic Calli of Crocus Cancellatus. Plant Cel. Tissue Organ. Cult. 65, 115–121. doi:10.1023/a:1010661620753
Karamian, R., and Ranjbar, M. (2011). Somatic Embryogenesis and Plantlet Regeneration from Protoplast Culture of Muscari Neglectum Guss. Afr. J. Biotechnol. 10, 4602–4607.
Kästner, U., Klocke, E., and Abel, S. (2017). Regeneration of Protoplasts after Somatic Hybridisation of Hydrangea. Plant Cel. Tiss. Organ. Cult 129, 359–373. doi:10.1007/s11240-017-1183-x
Kaur, N. D., Vyvadilová, M., Klíma, M., and Bechyně, M. (2006). A Simple Procedure for Mesophyll Protoplast Culture and Plant Regeneration in Brassica oleracea L. And Brassica Napus L. Czech. J. Genet. Plant Breed. 42, 103–110.
Kiełkowska, A., and Adamus, A. (2012). An Alginate-Layer Technique for Culture of Brassica oleracea L. protoplastsIn Vitro. Cel. Dev. Biol. Plant. 48, 265–273. doi:10.1007/s11627-012-9431-6
Kiełkowska, A., and Adamus, A. (2021). Exogenously Applied Polyamines Reduce Reactive Oxygen Species, Enhancing Cell Division and the Shoot Regeneration from Brassica oleracea L. Var. Capitata Protoplasts. Agronomy 11, 735.
Kiełkowska, A., and Adamus, A. (2019). Peptide Growth Factor Phytosulfokine-α Stimulates Cell Divisions and Enhances Regeneration from B. oleracea Var. Capitata L. Protoplast Culture. J. Plant Growth Regul. 38, 931–944.
Kiełkowska, A., Grzebelus, E., Lis-Krzyścin, A., and Maćkowska, K. (2019). Application of the Salt Stress to the Protoplast Cultures of the Carrot (Daucus Carota L.) and Evaluation of the Response of Regenerants to Soil Salinity. Plant Cell Tissue Organ Cult. PCTOC 137, 379–395.
Klejdus, B., Kováčik, J., and Babula, P. (2013). PAL Inhibitor Evokes Different Responses in Two Hypericum Species. Plant Physiol. Biochem. 63, 82–88. doi:10.1016/j.plaphy.2012.11.019
Klimek-Chodacka, M., Kadluczka, D., Lukasiewicz, A., Malec-Pala, A., Baranski, R., and Grzebelus, E. (2020). Effective Callus Induction and Plant Regeneration in Callus and Protoplast Cultures of Nigella Damascena L. Plant Cel. Tiss. Organ. Cult 143, 693–707. doi:10.1007/s11240-020-01953-9
Kobayashi, S., Kameya, T., and Ichihashi, S. (1993). Plant Regeneration from Protoplasts Derived from Callus of Phalaenopsis. Plant Tissue Cult. Lett. 10, 267–270. doi:10.5511/plantbiotechnology1984.10.267
Kyozuka, J., Hayashi, Y., and Shimamoto, K. (1987). High Frequency Plant Regeneration from rice Protoplasts by Novel Nurse Culture Methods. Mol. Gen. Genet. 206, 408–413. doi:10.1007/bf00428879
Lassoued, R., Phillips, P. W. B., Macall, D. M., Hesseln, H., and Smyth, S. J. (2021). Expert Opinions on the Regulation of Plant Genome Editing. Plant Biotechnol. J. doi:10.1111/pbi.13597
Lee, M. H., Lee, J., Choi, S. A., Kim, Y.-S., Koo, O., Choi, S. H., et al. (2020). Efficient Genome Editing Using CRISPR-Cas9 RNP Delivery into Cabbage Protoplasts via Electro-Transfection. Plant Biotechnol. Rep. 14, 695–702. doi:10.1007/s11816-020-00645-2
Liang, Z., Chen, K., Zhang, Y., Liu, J., Yin, K., Qiu, J.-L., et al. (2018). Genome Editing of Bread Wheat Using Biolistic Delivery of CRISPR/Cas9 In Vitro Transcripts or Ribonucleoproteins. Nat. Protoc. 13, 413–430. doi:10.1038/nprot.2017.145
Lin, C.-S., Hsu, C.-T., Yang, L.-H., Lee, L.-Y., Fu, J.-Y., Cheng, Q.-W., et al. (2018). Application of Protoplast Technology to CRISPR/Cas9 Mutagenesis: from Single-Cell Mutation Detection to Mutant Plant Regeneration. Plant Biotechnol. J. 16, 1295–1310. doi:10.1111/pbi.12870
Lin, Q., Zong, Y., Xue, C., Wang, S., Jin, S., Zhu, Z., et al. (2020). Prime Genome Editing in rice and Wheat. Nat. Biotechnol. 38, 582–585. doi:10.1038/s41587-020-0455-x
Lopez-Arellano, M., Dhir, S., Albino, C. N., Santiago, A., Morris, T., and Dhir, S. K. (2014). Somatic Embryogenesis and Plantlet Regeneration from Protoplast Culture of Stevia rebaudiana. Br. Biotechnol. J. 5, 1–12.
Lukaszewska, E., Virden, R., and Sliwinska, E. (2012). Hormonal Control of Endoreduplication in Sugar Beet (Beta Vulgaris L.) Seedlings Growing In Vitro. Plant Biol. (Stuttg) 14, 216–222. doi:10.1111/j.1438-8677.2011.00477.x
Ma, X., Zhang, X., Liu, H., and Li, Z. (2020). Highly Efficient DNA-free Plant Genome Editing Using Virally Delivered CRISPR-Cas9. Nat. Plants 6, 773–779. doi:10.1038/s41477-020-0704-5
Maćkowska, K., Jarosz, A., and Grzebelus, E. (2014). Plant Regeneration from Leaf-Derived Protoplasts within the Daucus Genus: Effect of Different Conditions in Alginate Embedding and Phytosulfokine Application. Plant Cell Tissue Organ Cult. PCTOC 117, 241–252.
Magnien, E., Dalschaert, X., and Faraoni-Sciamanna, P. (1982). Transmission of a Cytological Heterogeneity from the Leaf to the Protoplasts in Culture. Plant Sci. Lett. 25, 291–303. doi:10.1016/0304-4211(82)90159-6
Majewska-Sawka, A., Niklas, A., and Jażdżewska, E. (1997). The Effect of Polyamines on the Development of Sugar Beet Protoplasts. Biol. Plant. 39, 561–567. doi:10.1023/a:1000926714622
Masani, M. Y. A., Noll, G., Parveez, G. K. A., Sambanthamurthi, R., and Prüfer, D. (2013). Regeneration of Viable Oil palm Plants from Protoplasts by Optimizing media Components, Growth Regulators and Cultivation Procedures. Plant Sci. 210, 118–127. doi:10.1016/j.plantsci.2013.05.021
Masson, J., and Paszkowski, J. (1992). The Culture Response ofArabidopsis Thalianaprotoplasts Is Determined by the Growth Conditions of Donor Plants. Plant J. 2, 829–833. doi:10.1111/j.1365-313x.1992.tb00153.x
Meyer, L., Serek, M., and Winkelmann, T. (2009). Protoplast Isolation and Plant Regeneration of Different Genotypes of Petunia and Calibrachoa. Plant Cel. Tiss. Organ. Cult. 99, 27–34. doi:10.1007/s11240-009-9572-4
Mizuhiro, M., Kenichi, Y., Ito, K., Kadowaki, S., Ohashi, H., and Mii, M. (2001). Plant Regeneration from Cell Suspension-Derived Protoplasts of Primula Malacoides and Primula Obconica. Plant Sci. 160, 1221–1228. doi:10.1016/s0168-9452(01)00390-9
Murashige, T., and Skoog, F. (1962). A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant 15, 473–497. doi:10.1111/j.1399-3054.1962.tb08052.x
Nagata, T., and Ishii, S. (1979). A Rapid Method for Isolation of Mesophyll Protoplasts. Can. J. Bot. 57, 4. doi:10.1139/b79-226
Nassour, M., and Dorion, N. (2002). Plant Regeneration from Protoplasts of Micropropagated Pelargonium X Hortorum ‘Alain': Effect of Some Environmental and Medium Factors on Protoplast System Efficiency. Plant Sci. 163, 169–176. doi:10.1016/s0168-9452(02)00093-6
Nirmal Babu, K., Samsudeen, K., Divakaran, M., Pillai, G. S., Sumathi, V., Praveen, K., et al. (2016). “Protocols for In Vitro Propagation, Conservation, Synthetic Seed Production, Embryo Rescue, Microrhizome Production, Molecular Profiling, and Genetic Transformation in Ginger (Zingiber Officinale Roscoe.),” in Protocols for in Vitro Cultures and Secondary Metabolite Analysis of Aromatic and Medicinal Plants. Methods in Molecular Biology. Editor S. M. Jain Second Edition (New York, NY: Springer), 403–426. doi:10.1007/978-1-4939-3332-7_28
Nitsch, J. P., and Nitsch, C. (1969). Haploid Plants from Pollen Grains. Science 163, 85–87. doi:10.1126/science.163.3862.85
Nybakken, L., Keski-Saari, S., Falck, M. A., and Julkunen-Tiitto, R. (2007). Restoration of Secondary Metabolism in Birch Seedlings Relieved from PAL-Inhibitor. Trees 21, 273–281. doi:10.1007/s00468-006-0120-0
Ochatt, S. J., Mousset-Déclas, C., and Rancillac, M. (2000). Fertile Pea Plants Regenerate from Protoplasts when Calluses Have Not Undergone Endoreduplication. Plant Sci. 156, 177–183. doi:10.1016/s0168-9452(00)00250-8
Osakabe, Y., Liang, Z., Ren, C., Nishitani, C., Osakabe, K., Wada, M., et al. (2018). CRISPR-Cas9-mediated Genome Editing in Apple and grapevine. Nat. Protoc. 13, 2844–2863. doi:10.1038/s41596-018-0067-9
Park, J., Choi, S., Park, S., Yoon, J., Park, A. Y., and Choe, S. (2019). “DNA-free Genome Editing via Ribonucleoprotein (RNP) Delivery of CRISPR/Cas in Lettuce,” in Plant Genome Editing with CRISPR Systems: Methods and Protocols. Methods in Molecular Biology. Editor Y. Qi (New York, NY: Springer), 337–354. doi:10.1007/978-1-4939-8991-1_25
Pauk, J., Fekete, S., Vilkki, J., and Pulli, S. (1991). Protoplast Culture and Plant Regeneration of Different Agronomically Important Brassica Species and Varieties. AFSci 63, 371–378. doi:10.23986/afsci.72416
Pearce, R. S., and Cocking, E. C. (1973). Behaviour in Culture of Isolated Protoplasts from .Paul's Scarlet. Rose Suspension Culture Cells. Protoplasma 77, 165–180. doi:10.1007/bf01276755
Prakash, A. H., Rao, K. S., and Kumar, M. U. (1997). Plant Regeneration from Protoplasts ofCapsicum Annuum L. Cv. California Wonder. J. Biosci. 22, 339–344. doi:10.1007/bf02703236
Prange, A. N. S., Bartsch, M., Serek, M., and Winkelmann, T. (2010a). Regeneration of Different Cyclamen Species via Somatic Embryogenesis from Callus, Suspension Cultures and Protoplasts. Scientia Horticulturae 125, 442–450. doi:10.1016/j.scienta.2010.04.018
Prange, A. N. S., Serek, M., Bartsch, M., and Winkelmann, T. (2010b). Efficient and Stable Regeneration from Protoplasts of Cyclamen Coum Miller via Somatic Embryogenesis. Plant Cel. Tiss. Organ. Cult. 101, 171–182. doi:10.1007/s11240-010-9674-z
Qin, Y. H., Teixeira da Silva, J. A., Bi, J. H., Zhang, S. L., and Hu, G. B. (2011). Response of In Vitro Strawberry to Antibiotics. Plant Growth Regul. 65, 183–193. doi:10.1007/s10725-011-9587-9
Rahmani, M.-S., Pijut, P. M., and Shabanian, N. (2016). Protoplast Isolation and Genetically True-to-type Plant Regeneration from Leaf- and Callus-Derived Protoplasts of Albizia Julibrissin. Plant Cel. Tiss. Organ. Cult. 127, 475–488. doi:10.1007/s11240-016-1072-8
Reustle, G., and Natter, I. (1994). Effect of Polyvinylpyrrolidone and Activated Charcoal on Formation of Microcallus from grapevine Protoplasts (Vitis sp.). VITIS - J. Grapevine Res. 33, 117–121.
Rezazadeh, R., and Niedz, R. P. (2015). Protoplast Isolation and Plant Regeneration of Guava (Psidium Guajava L.) Using Experiments in Mixture-Amount Design. Plant Cel. Tiss. Organ. Cult. 122, 585–604. doi:10.1007/s11240-015-0790-7
Ruesink, A. W. (1978). Leucine Uptake and Incorporation by Convolvulus Tissue Culture Cells and Protoplasts under Severe Osmotic Stress. Physiol. Plant 44, 48–56. doi:10.1111/j.1399-3054.1978.tb01612.x
Sahab, S., Hayden, M. J., Mason, J., and Spangenberg, G. (2019). “Mesophyll Protoplasts and PEG-Mediated Transfections: Transient Assays and Generation of Stable Transgenic Canola Plants,” in Transgenic Plants: Methods and Protocols. Methods in Molecular Biology. Editors S. Kumar, P. Barone, and M. Smith (New York, NY: Springer), 131–152. doi:10.1007/978-1-4939-8778-8_10
Sakamoto, Y., Kawamura, A., Suzuki, T., Segami, S., Maeshima, M., Polyn, S., et al. (2021). Transcriptional Activation of Auxin Biosynthesis Drives Developmental Reprogramming of Differentiated Cells. bioRxiv. doi:10.1101/2021.06.26.450054
Sauer, N. J., Narváez-Vásquez, J., Mozoruk, J., Miller, R. B., Warburg, Z. J., Woodward, M. J., et al. (2016). Oligonucleotide-Mediated Genome Editing Provides Precision and Function to Engineered Nucleases and Antibiotics in Plants. Plant Physiol. 170, 1917–1928. doi:10.1104/pp.15.01696
Saxena, P. K., and Gill, R. (1986). Removal of browning and Growth Enhancement by Polyvinylpolypyrrolidone in Protoplast Cultures of Cyamopsis Tetragonoloba L. Biol. Plant 28, 313–315. doi:10.1007/bf02902302
Shahin, E. A. (1985). Totipotency of Tomato Protoplasts. Theoret. Appl. Genet. 69, 235–240. doi:10.1007/bf00662431
Shan, Q., Wang, Y., Li, J., Zhang, Y., Chen, K., Liang, Z., et al. (2013). Targeted Genome Modification of Crop Plants Using a CRISPR-Cas System. Nat. Biotechnol. 31, 686–688. doi:10.1038/nbt.2650
Sheng, X., Zhao, Z., Yu, H., Wang, J., Xiaohui, Z., and Gu, H. (2011). Protoplast Isolation and Plant Regeneration of Different Doubled Haploid Lines of Cauliflower (Brassica oleracea Var. Botrytis). Plant Cel. Tiss. Organ. Cult. 107, 513–520. doi:10.1007/s11240-011-0002-z
Shulse, C. N., Cole, B. J., Ciobanu, D., Lin, J., Yoshinaga, Y., Gouran, M., et al. (2019). High-Throughput Single-Cell Transcriptome Profiling of Plant Cell Types. Cel. Rep. 27, 2241–2247. doi:10.1016/j.celrep.2019.04.054
Staba, E. J. (1969). Plant Tissue Culture as a Technique for the Phytochemist. Recent Adv. Phytochem. 2, 75–105.
Subburaj, S., Chung, S. J., Lee, C., Ryu, S.-M., Kim, D. H., Kim, J.-S., et al. (2016). Site-directed Mutagenesis in Petunia × Hybrida Protoplast System Using Direct Delivery of Purified Recombinant Cas9 Ribonucleoproteins. Plant Cel. Rep. 35, 1535–1544. doi:10.1007/s00299-016-1937-7
Svitashev, S., Schwartz, C., Lenderts, B., Young, J. K., and Mark Cigan, A. (2016). Genome Editing in maize Directed by CRISPR-Cas9 Ribonucleoprotein Complexes. Nat. Commun. 7, 13274. doi:10.1038/ncomms13274
Tahami, S. K., Chamani, E., and Zare, N. (2014). Plant Regeneration from Protoplasts of Lilium Ledebourii (Baker) Boiss. J. Agric. Sci. Technology 16, 1133–1144.
Tang, W., Harris, L. C., Outhavong, V., and Newton, R. J. (2004). Antioxidants Enhance In Vitro Plant Regeneration by Inhibiting the Accumulation of Peroxidase in Virginia pine (Pinus Virginiana Mill.). Plant Cel. Rep. 22, 871–877. doi:10.1007/s00299-004-0781-3
Tomiczak, K., Mikuła, A., Sliwinska, E., and Rybczyński, J. J. (2015). Autotetraploid Plant Regeneration by Indirect Somatic Embryogenesis from Leaf Mesophyll Protoplasts of Diploid Gentiana Decumbens L.F. In Vitro Cell.Dev.Biol.-Plant 51, 350–359. doi:10.1007/s11627-015-9674-0
Vessabutr, S., and Grant, W. F. (1995). Isolation, Culture and Regeneration of Protoplasts from Birdsfoot Trefoil (Lotus corniculatus). Plant Cel. Tiss. Organ. Cult. 41, 9–15. doi:10.1007/bf00124081
Wójcik, A., and Rybczyński, J. J. (2015). Electroporation and Morphogenic Potential of Gentiana Kurroo (Royle) Embryogenic Cell Suspension Protoplasts. bta 1, 19–29. doi:10.5114/bta.2015.54170
Woo, J. W., Kim, J., Kwon, S. I., Corvalán, C., Cho, S. W., Kim, H., et al. (2015). DNA-free Genome Editing in Plants with Preassembled CRISPR-Cas9 Ribonucleoproteins. Nat. Biotechnol. 33, 1162–1164. doi:10.1038/nbt.3389
Wu, F.-H., Shen, S.-C., Lee, L.-Y., Lee, S.-H., Chan, M.-T., and Lin, C.-S. (2009). Tape-Arabidopsis Sandwich - a Simpler Arabidopsis Protoplast Isolation Method. Plant Methods 5, 16. doi:10.1186/1746-4811-5-16
Xiao, L., Zhang, L., Yang, G., Zhu, H., and He, Y. (2012). Transcriptome of Protoplasts Reprogrammed into Stem Cells in Physcomitrella Patens. PLoS ONE 7, e35961. doi:10.1371/journal.pone.0035961
Yang, H., Matsubayashi, Y., Nakamura, K., and Sakagami, Y. (1999). Oryza Sativa PSK Gene Encodes a Precursor of Phytosulfokine-Alpha, a Sulfated Peptide Growth Factor Found in Plants. Proc. Natl. Acad. Sci. 96, 13560–13565. doi:10.1073/pnas.96.23.13560
Yong, J. W., Ge, L., Ng, Y. F., and Tan, S. N. (2009). The Chemical Composition and Biological Properties of Coconut (Cocos Nucifera L.) Water. Molecules 14, 5144–5164. doi:10.3390/molecules14125144
Yu, J., Tu, L., Subburaj, S., Bae, S., and Lee, G.-J. (2020). Simultaneous Targeting of Duplicated Genes in Petunia Protoplasts for Flower Color Modification via CRISPR-Cas9 Ribonucleoproteins. Plant Cel. Rep. doi:10.1007/s00299-020-02593-1
Zafar, Y., Nenz, E., Damiani, F., Pupilli, F., and Arcioni, S. (1995). Plant regeneration from explant and protoplast derived calluses of Medicago littoralis. Plant Cell Tissue Organ Cult. 41: 41–48.
Zhang, Y., Liang, Z., Zong, Y., Wang, Y., Liu, J., Chen, K., et al. (2016). Efficient and Transgene-free Genome Editing in Wheat through Transient Expression of CRISPR/Cas9 DNA or RNA. Nat. Commun. 7, 12617. doi:10.1038/ncomms12617
Zhang, Y., Malzahn, A. A., Sretenovic, S., and Qi, Y. (2019). The Emerging and Uncultivated Potential of CRISPR Technology in Plant Science. Nat. Plants 5, 778–794. doi:10.1038/s41477-019-0461-5
Keywords: protoplast, regeneration, gene editing, crop improvement, tissue culture
Citation: Reed KM and Bargmann BOR (2021) Protoplast Regeneration and Its Use in New Plant Breeding Technologies. Front.Genome Ed. 3:734951. doi: 10.3389/fgeed.2021.734951
Received: 01 July 2021; Accepted: 11 August 2021;
Published: 03 September 2021.
Edited by:
Matthew R. Willmann, Pairwise, United StatesReviewed by:
Yanpeng Wang, Institute of Genetics and Developmental Biology (CAS), ChinaCopyright © 2021 Reed and Bargmann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bastiaan O. R. Bargmann, YmFzdGlhYW5AdnQuZWR1
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.