 Shijie Ma
Shijie Ma Binjie Gan
Binjie Gan
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Genet. , 22 October 2024
Sec. Genomics of Plants and the Phytoecosystem
Volume 15 - 2024 | https://doi.org/10.3389/fgene.2024.1473717
This article is part of the Research Topic Advances in crop breeding for abiotic stress tolerance View all 8 articles
Low temperature chilling is one of the major abiotic stresses affecting growth and yield of Triticum aestivum L. With global climate change, the risk of cold damage in wheat production has increased. In recent years, with the extensive research on wheat chilling resistance, especially the development of genetic engineering technology, the research on wheat chilling resistance has made great progress. This paper describes the mechanism of wheat cold damage, including cell membrane injury, cytoplasmic concentration increased as well as the imbalance of the ROS system. Mechanisms of cold resistance in wheat are summarised, including hormone signalling, transcription factor regulation, and the role of protective enzymes of the ROS system in cold resistanc. Functions of cloned wheat cold resistance genes are summarised, which will provide a reference for researchers to further understand and make use of cold resistance related genes in wheat. The current cold resistant breeding of wheat relies on the agronomic traits and observable indicators, molecular methods are lacked. A strategy for wheat cold-resistant breeding based on QTLs and gene technologies is proposed, with a view to breeding more cold-resistant varieties of wheat with the deepening of the research.
Wheat (Triticum aestivum L.) is one of the world’s top three grain crops. According to the FAO, around 200 million hectares of wheat will be planted globally with a production of about 787 million tonnes in 2023. Wheat is the calorie source for one-fifth of the global population (Pang et al., 2021), therefore, high and stable wheat production plays a crucial role in global food security. In recent decades, the likelihood of extreme weather on Earth has increased dramatically as greenhouse gases continue to be emitted (Jiang et al., 2022), low-temperature freezes occur from time to time, posing a significant threat to wheat production (Sutka, 2001; Vij and Tyagi, 2007). It occurs not only in the main temperate wheat-producing regions, but also in subtropical and Mediterranean climatic regions, covering the main wheat-producing countries such as China, the United States, Canada, Australia and some parts of South America. In the 25 years since 1990, the frequent occurrence of wheat freezes has caused severe losses over large areas in the main wheat-producing regions of central China, with the most severe reductions in yields of up to 30% (Yang and Yuan, 2014). In the central regions of the United States, statistics from 1955 to 2010 show that the yield of wheat is reduced by 210 kg per hectare due to low-temperature frosts (Holman et al., 2011). In the UK and eastern Australia, there has also been a marked increase in recorded frosts since the 1950s, causing some damage to wheat production (Whaley et al., 2004; Zheng et al., 2015). The incidence of frost damage results in millions of tonnes of food production being lost each year, with direct economic losses in excess of tens of billions of dollars. Wheat growth and development will be subjected to more cold stresses as the global climate continues to change, so improving varietal cold tolerance is very important in wheat breeding (Pang et al., 2021).
Wheat is a winter-habituated crop that must experience low temperatures to complete vernalisation (Fowler and Carles, 1979; Fowler et al., 1996), during inter-annual growth, they are exposed to the effects of multiple cold stress. The wheat is in the seedling stage before the winter, it will appear leaves dry, root damage or even dead seedlings phenomenon in case of a sudden drop in temperature and the yields will be also affected. After winter, if wheat is weakly resistant to cold the low-temperatures can damage plant tissues and inhibit growth and development (Subedi et al., 1998; Barton et al., 2014). During the period from nodulation to tasseling, wheat is in a period of parallel nutritive and reproductive growth, spikes can become hollow or underdeveloped as a result of frost or “spring cold” (Limin and Fowler, 2006), which will have a disastrous effect on wheat production. In summary, wheat is often exposed to low temperatures at critical stages of growth and development, and data shows that nearly 85% of the world’s wheat area is affected by spring cold each year (Ferrante et al., 2021), it also shows the practical importance of research on cold resistance in wheat for production. The research on the function of plant cold resistance genes has been widely carried out with the deepening of research on the mechanism of plant cold resistance and plant genetic engineering. In this review, we will summarise the achievements in the mining and utilisation of cold-resistant genes in wheat and clarify the physiological and molecular mechanisms of cold resistance, and strategies for wheat cold resistance breeding will be provided.
Low temperature affect the physiological and biochemical responses of plant cells seriously. Numerous studies have shown that the cold resistance of wheat is closely related to the mobility of cell membrane, reactive oxygen system and cellular osmoregulation. The mechanism of damages in wheat under cold stress is shown in Figure 1. The response to low-temperature involves multiple levels of regulation in wheat, including transcriptional regulation, hormonal regulation and signal transmission. These regulatory processes are quite complex and involve a large number of genes related to cold resistance.
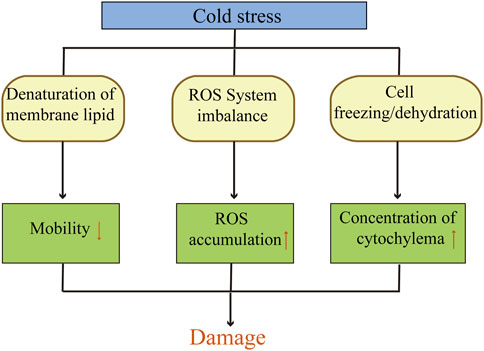
Figure 1. Mechanism of damages in wheat under cold stress.
After suffering from cold damage, wheat cell tissues can be damaged or even die, mainly due to the destruction of cell membrane structures (Lyons, 1973). The stability of cell membranes is determined by the composition of membrane lipid fatty acids. The stability of cell membrane is determined by the fatty acid composition of membrane lipids. Generally, a higher content of unsaturated fatty acids in membrane lipids correlates with an enhanced plant cold resistance. Therefore, fatty acid desaturase (FAD)plays a certain role in plant cold resistance (Ishizaki-Nishizawa et al., 1996; Kodama et al., 1995). The cell membrane also serves as a sensor and conduit for low-temperature signals, encompassing calcium signals, protein kinases, phosphatases, and transcription factors, all of which participate in the cold stress signaling pathway (Soleimani et al., 2022). Researchers indicated that the electrical conductivity method for determining the permeability of cell membranes of winter wheat leaves under indoor cold stress showed a significant positive correlation with the cold resistance of wheat in the field (Ju et al., 2012).
The reactive oxygen species (ROS) accumulation and scavenging system in plants is crucial for maintaining the normal cellular functions. Generally, the accumulation and scavenging of ROS in this system is in dynamic balance. Persistent low temperatures can disrupt this balance, accelerating ROS accumulation and producing a large amount of free radicals, which in turn cause physiological disorders within cells (Zeng et al., 2011; Bhattacharjee, 2005). SOD, per-Oxidase (POD), CAT, and ascorbate per-Oxidase (APX) are important protective enzymes in wheat (Türkan et al., 2005). They can scavenge excess ROS and free radicals generated during cold stress, maintaining the normal growth of wheat (Demiral and Türkan, 2004).
When encountering low temperature cold damage, the loss of water molecules in plant cells increases the concentration of cell fluid, causing damage to the cells themselves. In order to avoid this kind of damage, the body will actively accumulate some soluble substances to increase the cell’s ability to absorb water and maintain normal metabolic functions (Lalk and Dorffling, 1985). The soluble sugar content of wheat is positively correlated with cold resistance, and sucrose and fructose play an important role in the cold resistance of wheat (Kamata and Uemura, 2004; Zeng et al., 2011). The content of proline (Pro) is positively correlated with the cold resistance of wheat. Many results show that when the temperature is low, the proline content in the wheat body rises, more water is aggregated on the protein, preventing the protein from deforming due to dehydration at low temperatures, thereby protecting wheat cells from damage (Bao et al., 2021; Chen et al., 2007).
Plant cells can respond to low temperature stress by precisely regulating the expression of transcription factors and effector genes. These signaling pathways consist of transcription, translation, and post-transcriptional and post-translational regulatory factors that induce the expression of functional genes in response to cold temperatures. Within one of the signaling pathways is a cis-element known as the C-repeat element/dehydration-responsive element (CRT/DRE), to which C-repeat binding factors (CBF) can bind. This pathway is referred to as the CBF-COR signaling pathway. Studies on a variety of plants have shown that this pathway makes plants have cold resistance by regulating the expression of downstream cold-resistant proteins (Ding et al., 2019). CBF homologous genes in wheat and barley have also been identified (Mohseni et al., 2012; Skinner et al., 2005). ICE1 (CBF expression inducer) is a MYC-type bHLH transcription factor and is the most characteristic transcription activator of CBF genes so far (Chinnusamy et al., 2003; Lee et al., 2005). Overexpression of ICE1 can increase the expression of CBFs (Fursova et al., 2009; Agarwal et al., 2006; Juan et al., 2015). The expression of ICE1 gene TaICE41 in wheat is induced by cold stress (Badawi et al., 2008; Guo et al., 2019). The ICE-CBF-COR pathway plays an important regulatory role in the cold resistance pathway of wheat (Gong et al., 2020; Caccialupi et al., 2024) and is considered to be the main cold signal transduction pathway in wheat (Figure 2).
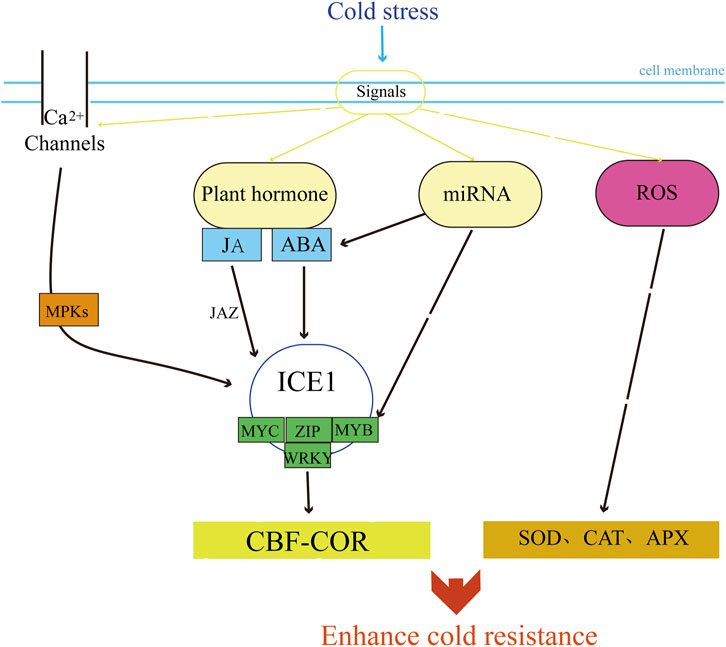
Figure 2. Gene mechanism of ICE-CBF-COR pathway in wheat under cold stress.
Plant hormones are crucial regulators in perceiving and transmitting various environmental signals and defense responses. Abscisic acid (ABA) and jasmonic acid (JA) are the most studied plant hormones in relation to wheat cold resistance. Lower temperatures can increase endogenous ABA levels in plants, participating in cold stress signal transduction and activating the expression of cold-resistance genes (Zhang et al., 2020; An et al., 2021; Liu et al., 2013). For instance, the cor gene family members such as cor6.6, cor15a, cor47, and cor78 are highly expressed due to increased ABA levels, thereby enhancing plant freeze tolerance (Hajela et al., 1990). Studies on wheat have found that endogenous ABA content increases during cold acclimation in winter wheat, with a rapid rise in ABA levels inducing the expression of stress-responsive genes and improving freeze resistance (Liu et al., 2013). Another plant hormone, JA, has also been recently discovered to play a regulatory role in plant cold stress responses. Cold stress activates the expression of JA biosynthesis-related genes (AOC, AOS1, AOS2, and LOX2), increasing JA content (Hu et al., 2017; Du et al., 2013). Accumulated JA conjugates with isoleucine to form the active JA-Ile, which is perceived by the COI1-JAZ co-receptor and promotes the ubiquitination and degradation of the repressor protein JAZ via the 26S proteasome, thereby activating the ICE-CBF pathway and inducing the expression of certain cold-responsive genes, enhancing plant cold tolerance (Hu et al., 2013).
Excessive ROS production in plant cells under stress conditions is highly reactive and toxic to proteins, lipids, and nucleic acids. ROS in plants primarily exists in the form of free radicals and non-radicals, including hydrogen peroxide (H2O2), superoxide anion (O2·-), hydroxyl radical (OH·-), and singlet oxygen (1O2), etc. Elevated ROS levels lead to various harmful cytological effects (Nadarajah, 2020), ultimately resulting in cell damage and death (Gill and Tuteja, 2010). On the other hand, the increase in ROS during stress periods is also considered a signal for the activation of stress response pathways (Baxter et al., 2014). ROS can also function as a signaling molecule, the overaccumulation of ROS at the initial stage of cold stress promotes the activation of antioxidant enzyme genes such as SOD, CAT, and APX to eliminate ROS toxicity. Notably, studies in wheat have demonstrated that the overexpression of the wheat TaCAT gene, which eliminates excessive H2O2, improves the freeze tolerance of transgenic plants (Matsumura et al., 2010).
While calcium (Ca2+) is known to enhance stress tolerance (Malko et al., 2023). In plants, low temperatures induce the accumulation of intracellular Ca2+ (Yang et al., 2010), and the calcium/calcineurin (Ca2+/CaM) regulated receptor kinase CLRK1/2 acts as a sensor for Ca2+/CaM under low temperatures. It interacts with and phosphorylates MEKK1, triggering the activation of downstream MPK4/6 and thereby positively regulating the cold response (Furuya et al., 2013). Cold stress activates MKK2, which in turn activates MPK4 and MPK6 (Ichimura et al., 2000). Additionally, cold stress activates protein kinases MPK3 and MPK6, phosphorylates ICE1 protein, and decreases its protein stability and transcriptional activity, thus negatively regulating the expression of CBF and the cold tolerance of plants (Li et al., 2017). Studies in wheat have shown that Ca2+ induces cold-responsive genes (WCOR413, WCOR410, WCOR14, and Wrab17) enhanced cold stress tolerance through maintained cellular redox homeostasis (Malko et al., 2023). Transformation with the Ta MPK6 gene significantly enhances the expression of the ICE-CBF-COR pathway genes in Arabidopsis, improving the plant’s cold tolerance (Yu et al., 2023).
MicroRNA (miRNA) is a class of non-coding small RNA molecules derived from the genomes of eukaryotic organisms, mostly ranging from 21 to 25 nt in length. Their main mode of action is post-transcriptional regulation of target gene expression, which also plays an important role in plant stress response (Naya et al., 2014; Tiwari et al., 2020). Using deep sequencing and bioinformatics prediction methods, miRNAs responding to cold stress have been identified in various crops and model species (Lv et al., 2010; Cao et al., 2014; Sun et al., 2015). In wheat, overexpression of miR399 can positively regulate the protein levels of ICE1 and the expression of key genes CORs in the CBF signaling pathway of plants (Peng et al., 2021).
At present, many studies have explained the physiological and molecular mechanisms of cold resistance in wheat from different perspectives, and the research on the genes related to cold resistance in wheat has also progressed rapidly. Cold resistant genes are a class of induced genes that can be activated and expressed in large quantities to produce corresponding cold-resistant substances only at specific periods and conditions. Cold resistance is a complex trait involving many physiological mechanisms, which is the result of the combined action of multiple genes. With the rapid development of molecular biology technology and its wide application in the field of plant breeding, several quantitative trait loci (QTL) related to cold resistance have been mapped by linkage analysis.
Many studies have confirmed that due to the huge genetic background of wheat, wheat cold resistance is controlled and played a role by multiple minor genes. Researchers have mapped allelic loci related to wheat cold resistance on more than 10 pairs of chromosomes such as 1B, 1D, 2B, 2D, 4D, 5A, 5D, and 7A of wheat. The DH population of wheat was used to map QTL for the semi lethal temperature, an index related to cold resistance traits of wheat, and found a total of five major QTL loci controlling the semi lethal temperature, which were distributed on chromosomes 2A, 5A, 1D and 6D of wheat (Båga et al., 2007). Some studies have also found that there are some alleles on chromosomes 5A and 5D that regulate the cold resistance of wheat. At present, QTL mapping loci cover almost all wheat chromosomes, among which 5A and 5D chromosomes are most closely related to cold resistance in wheat (Sutka, 1994; Limin et al.,. 1997), As well as a major-effect QTL related to frost tolerance was reported located on chromosomes 4A recently (Bolouri et al., 2023). The vrnl FRL segment of 5A chromosome in wheat regulates cold resistance in wheat. In common wheat, the major loci controlling cold resistance (FR-1 and FR-2) have been mapped to the long arm of chromosome 5 (Galiba et al., 1995; Tóth et al., 2003). FR-2 coincides with the CBFs gene cluster in wheat and barley (Miller et al., 2006; Francia et al., 2007) and directly induces downstream cor/lea gene expression during cold adaptation (Takumi et al., 2008). A large number of deletions in the CBF cluster of fr-b2 significantly reduced the cold tolerance of tetraploid and hexaploid wheat (Pearce et al., 2013). A recent study reported that two genes named Wcr-3 and Wcr-4 control cold resistance in wheat, loci on 2B and 2D chromosomes (Lei et al., 2023).
In order to further explore the molecular mechanism of winter wheat response to low temperature stress, several cold resistance genes in wheat were transferred into model plants and proved to affect its cold tolerance. Genes with cold resistance function were cloned from wheat as shown in Table 1. Studies have shown that overexpression of TabZIP1 reduces the MDA content and electrical conductivity of Arabidopsis at 4°C, and improves the cold tolerance of Arabidopsis (Lv et al., 2018); Overexpression of TaMYB56-B transgenic Arabidopsis has a higher survival rate at 4°C, and the expression of CBF3 and COR15a was strongly induced (Zhang et al., 2012). A recent study showed that overexpression of TaMYB4 enhanced the freezing tolerance of transgenic Arabidopsis, AtCBF1, AtCBF2, AtCBF3, AtCOR15A, AtCOR47, AtKIN1 and AtRD29A in transgenc lines was significantly upregulated (Tian et al., 2023). TabZIP6 was found as a negative regulator of cold resistance in wheat. The cold tolerance of Arabidopsis thaliana overexpressing TabZIP6 at 4 °C was significantly decreased, and the expression of multiple genes in the CBF-COR cold signaling pathway was downregulated (Cai et al., 2018). Wheat TaICE41 and TaICE87 are homologous to Arabidopsis AtICE1 genes (Badawi et al., 2008). TaCBF14 and TaCBF15 play a role in cold tolerance of spring barley, and transgenic TaCBF14 and TaCBF15 genes improve the cold tolerance of spring barley (Soltész et al., 2013). Researchers found that TaMYC2 interacted with TaICE41, activated the downstream CBF-COR signaling pathway, and positively regulated the freezing resistance of Arabidopsis thaliana (Wang et al., 2022). TaMPK3 is also involved in regulating the ICE-CBF-COR cold resistance module through its interaction with TaICE41, improving freezing tolerance in wheat (Wang et al., 2024). TaFAD2.8 genes high expression in response to temperature stress (Hajiahmadi et al., 2020). Transgenic TaMPK6 gene significantly reduced the production and accumulation ofROS in Arabidopsis plants. The expression levels of genes encoding superoxide dismutase (SOD) and catalase (CAT) and the enzyme activities of SOD and CAT were significantly increased. The CAT genes were upregulated by cold stress (Ghorbel et al., 2023). Overexpression of TaCAT increased the CAT activity of transgenic rice, eliminated excessive accumulation of H2O2, and improved the cold resistance of transgenic rice (Matsumura et al., 2010). The soluble sugar content of winter wheat seedlings was significantly accumulated (Yu et al., 2008), and the expression of genes encoding key enzymes in glucose metabolism was significantly upregulated. The ectopic expression of wheat TaFBA-A10 gene was showed in Arabidopsis thaliana could increase the content of soluble sugar by affecting the rate of glycolysis and Calvin cycle, thereby improving the cold resistance of Arabidopsis thaliana (Zeng et al., 2011); ectopic expression of TaTPS11, a gene encoding wheat trehalose-6-phosphate synthase, in Arabidopsis can increase the frost resistance of Arabidopsis by catalyzing the expression level of SnRK1 in sucrose (Peng et al., 2022), and downregulates the expression of this gene with significantly lower cold resistance than Arabidopsis wild-types (Lu et al., 2024). Overexpressing the key enzyme genes TaG6PDH and Ta6PGDH in the PPP pathway of wheat could improve its cold resistance by scavenging ROS generated at low temperature (Liu et al., 2019; Tian et al., 2021). The mechanism of action of some cold-resistant genes in wheat depends on hormone signal transduction. Lv found that exogenous MeJA treatment can significantly increase the expression of transcription factors (TabZIP1, TaWABI5, TaMYB80, TaNAC2, TaWRKY80) in winter wheat under low temperature stress, thereby regulating the CBF signaling pathway to improve cold resistance (Lv et al., 2017; Lv et al., 2018). Chu identified a SA methyltransferase, TaSAMT1 that converts SA to methyl SA (MeSA) and confers cold tolerance in wheat (Chu et al., 2024). For another example, under low temperature, exogenous ABA increased the expression of bZIP1, NAC2 and several key enzyme genes of sugar metabolism in wheat (Lv et al., 2018; Liu et al., 2013).
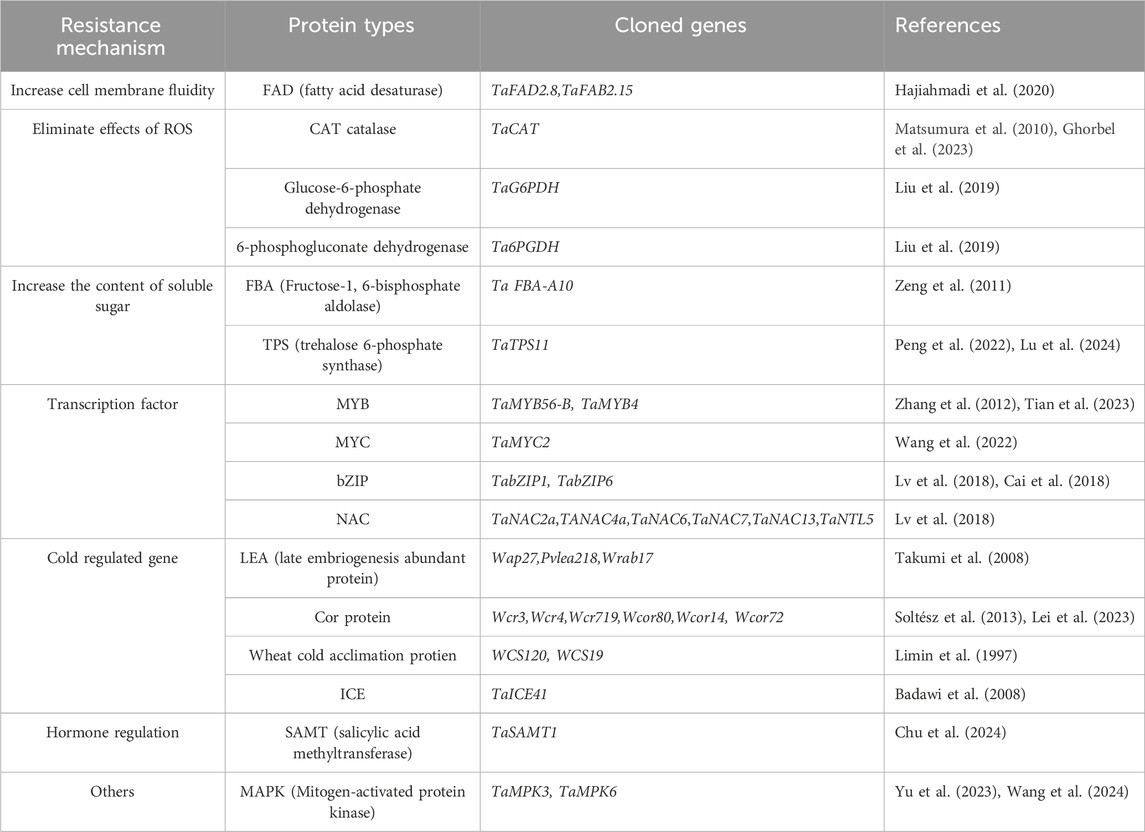
Table 1. Summary of cold resistance genes in wheat.
Breeding cold-resistant wheat varieties to protect wheat from low-temperature damage and maintain high and stable yields is of significant importance in production. However, breeding for cold resistance in wheat is still at the level of phenotypic selection, the cloned genes were also only functionally characterized in model plants. To make more rational and effective use of cold resistance genes, the following cold resistance breeding strategies for wheat are proposed (Figure 3).
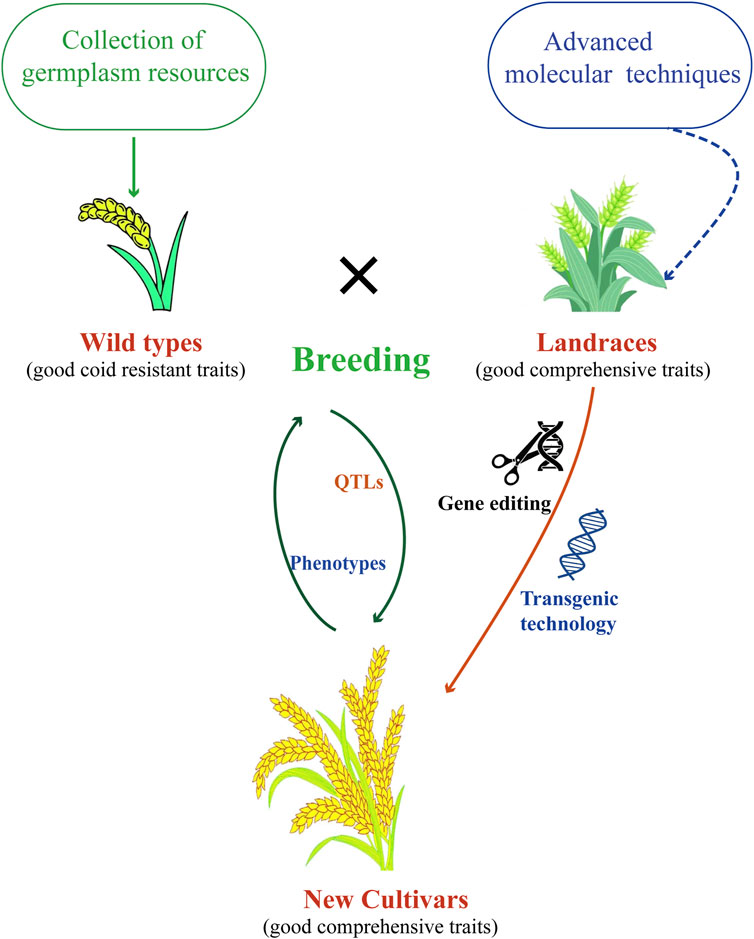
Figure 3. Strategies used for breeding wheat with cold resistance.
Agronomic traits are key factors in wheat cultivars breeding (Tshikunde et al., 2019). Try harder to collect, identify, and study the cold resistance of various germplasm resources (commercial varieties, breeding lines, local varieties, wild relatives, and distant species) is very necessary, and clarify their genetic characteristics and traits of resistance. Only by collecting enough germplasm resources can we explore new diversity of cold resistance genes for cloning, functional verification and aggregation.
There are a number of methods for evaluating and characterising cold resistance criteria in wheat (Valluru et al., 2012; Ou and Wang, 2019). To develop an accurate, fast and scalable method for identifying cold resistance indexes of wheat, so as to judge the cold resistance of breeding materials and germplasm resources through relevant indexes in the field.
QTLs have great potential for accelerating traditional breeding processes (Jha et al., 2017). Fine mapping and cloning of cold resistance genes/QTLs, studying the mechanism of cold resistance in wheat, and analyzing the regulatory network. Effectively use the existing cold-resistant genes with identification functions, use polymerization hybridization and backcross breeding, combined with molecular marker-assisted selection and generation-adding breeding techniques, accumulate different cold-resistant genes to high-yield varieties with excellent comprehensive traits, and create new materials for high-yield and multi-resistance breeding. On this basis, different hybrid combinations were combined by conventional breeding methods to cultivate new varieties with high yield and durable multi-resistance. Construct various genetic populations and natural populations, and use current wheat genome sequencing and molecular marker chips to carry out the mapping of cold resistance gene QTLs. Develop simple, accurate, and reliable molecular markers and detection technology systems that can be applied in practical breeding.
In-depth study of key functional genes is a method to improve the yield and quality of crops such as wheat. Genome editing methods, such as CRISPR/Cas9, allowed to manipulate wheat genome to improve agronomic traits (Kim et al., 2018). Transgenic and genome editing was carried out to directly create new varieties of high-yield and high-quality cold-resistant wheat on the basis of the original excellent large varieties. Some technologies in non-biological fields also canbe applied to cold resistance in wheat, such as nanotechnology (Venzhik et al., 2024).
SM: Conceptualization, Investigation, Software, Writing–original draft. XH: Writing–review and editing. XZ: Methodology, Writing–review and editing. LL: Software, Writing–review and editing. LZ: Methodology, Software, Writing–review and editing. BG: Conceptualization, Investigation, Writing–review and editing, Writing–original draft.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The work was supported by Open Project of Key Laboratory of Crop Quality improvement of Anhui Province (2024ZW005) and Joint research project on Wheat Breeding of Anhui Province (2021–2025).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Agarwal, M., Hao, Y., Kapoor, A., Dong, C. H., Fujii, H., Zheng, X., et al. (2006). A R2R3 type MYB transcription factor is involved in the cold regulation of CBF genes and in acquired freezing tolerance. J. Biol. Chem. 281, 37636–37645. doi:10.1074/jbc.M605895200
An, J. P., Wang, X. F., Zhang, X. W., You, C. X., and Hao, Y. J. (2021). Apple B-box protein BBX37 regulates jasmonic acid mediated cold tolerance through the JAZ-BBX37-ICE1-CBF pathway and undergoes MIEL1-mediated ubiquitination and degradation. New Phytol. 229 (5), 2707–2729. doi:10.1111/nph.17050
Badawi, M., Reddy, Y. V., Agharbaoui, Z., Tominaga, Y., Danyluk, J., Sarhan, F., et al. (2008). Structure and functional analysis of wheat ICE (inducer of CBF expression) genes. Plant Cell Physiol. 49 (8), 1237–1249. doi:10.1093/pcp/pcn100
Båga, M., Chodaparambil, S. V., Limin, A. E., Pecar, M., Fowler, D. B., and Chibbar, R. N. (2007). Identification of quantitative trait loci and associated candidate genes for low-temperature tolerance in cold-hardy winter wheat. Funct. and Integr. Genomics 7 (1), 53–68. doi:10.1007/s10142-006-0030-7
Bao, Y., Yang, N., Meng, J., Wang, D., Fu, L., Wang, J., et al. (2021). Adaptability of winter wheat Dongnongdongmai 1 (Triticum aestivum L.) to overwintering in alpine regions. Plant Biol. 23 (3), 445–455. doi:10.1111/plb.13200
Barton, D. A., Cantrill, L. C., Law, A. M., Phillips, C. G., Sutton, B. G., and Overall, R. L. (2014). Chilling to zero degrees disrupts pollen formation but not meiotic microtubule arrays in Triticum aestivumL. Plant Cell and Environ. 37 (12), 2781–2794. doi:10.1111/pce.12358
Baxter, A., Mittler, R., and Suzuki, N. (2014). ROS as key players in plant stress signalling. J. Exp. Bot. 65 (5), 1229–1240. doi:10.1093/jxb/ert375
Bhattacharjee, S. (2005). Reactive oxygen species and oxidative burst: roles in stress, senescence and signal transduction in plants. Currnt Sci. 89 (7). doi:10.1073/pnas.0506897102
Bolouri, P., Haliloğlu, K., Mohammadi, S. A., Türkoğlu, A., İlhan, E., Niedbała, G., et al. (2023). Identification of novel QTLs associated with frost tolerance in winter wheat (Triticum aestivum L.). Plants (Basel) 12 (8), 1641. doi:10.3390/plants12081641
Caccialupi, G., Milc, J., Caradonia, F., Nasar, M. F., and Francia, E. (2023). The triticeae CBF gene cluster-to frost resistance and beyond. Cells 12 (22), 2606. doi:10.3390/cells12222606
Cai, W., Yang, Y., Wang, W., Guo, G., Liu, W., and Bi, C. (2018). Overexpression of a wheat (Triticum aestivum L.) bZIP transcription factor gene, TabZIP6, decreased the freezing tolerance of transgenic Arabidopsis seedlings by down-regulating the expression of CBFs. Plant physiology Biochem. 124, 100–111. doi:10.1016/j.plaphy.2018.01.008
Cao, X., Wu, Z., Jiang, F., Zhou, R., and Yang, Z. (2014). Identification of chilling stress-responsive tomato microRNAs and their target genes by high-throughput sequencing and degradome analysis. BMC Genomics 15 (1), 1130. doi:10.1186/1471-2164-15-1130
Chen, X., Li, J., Ma, L., and Zhang, F. (2007). Effect of low temperature stress on change of free proline content in the leaves of spring and winter wheat. Xinjiang Agric. Sci. 4 (5), 553–556. doi:10.3969/j.issn.1001-4330.2007.05.002
Chinnusamy, V., Ohta, M., Kanrar, S., Lee, B. H., Hong, X., Agarwal, M., et al. (2003). ICE1: a regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis. Genes and Dev. 17 (8), 1043–1054. doi:10.1101/gad.1077503
Chu, W., Chang, S., Lin, J., Zhang, C., Li, J., Liu, X., et al. (2024). Methyltransferase TaSAMT1 mediates wheat freezing tolerance by integrating brassinosteroid and salicylic acid signaling. Plant Cell 36 (7), 2607–2628. doi:10.1093/plcell/koae100
Demiral, T., and Türkan, I. (2004). Does exogenous glycinebetaine affect antioxidative system of rice seedlings under NaCl treatment? J. Plant Physiology 161 (10), 1089–1100. doi:10.1016/j.jplph.2004.03.009
Ding, Y., Shi, Y., and Yang, S. (2019). Advances and challenges in uncovering cold tolerance regulatory mechanisms in plants. New Phytol. 222 (4), 1690–1704. doi:10.1111/nph.15696
Du, H., Liu, H., and Xiong, L. (2013). Endogenous auxin and jasmonic acid levels are differentially modulated by abiotic stresses in rice. Front. Plant Sci. 4 (397), 397. doi:10.3389/fpls.2013.00397
Ferrante, A., Cullis, B. R., Smith, A. B., and Able, J. A. (2021). A multi-environment trial analysis of frost susceptibility in wheat and barley under Australian frost-prone field conditions. Front. Plant Sci. 12, 722637. doi:10.3389/fpls.2021.722637
Fowler, D. B., and Carles, R. J. (1979). Growth, development, and cold tolerence of fall-acclimated cereal grains. Crop Sci. 19 (6), 915–922. doi:10.2135/cropsci1979.0011183X001900060040x
Fowler, D. B., Chauvin, L. P., Limin, A. E., and Sarhan, F. (1996). The regulatory role of vernalization in the expression of low-temperature-induced genes in wheat and rye. Theor. and Appl. Genet. 93 (4), 554–559. doi:10.1007/BF00417947
Francia, E., Barabaschi, D., Tondelli, A., Laidò, G., Rizza, F., Stanca, A. M., et al. (2007). Fine mapping of a HvCBF gene cluster at the frost resistance locus Fr-H2 in barley. Theor. and Appl. Genet. 115 (8), 1083–1091. doi:10.1007/s00122-007-0634-x
Fursova, O. V., Pogorelko, G. V., and Tarasov, V. A. (2009). Identification of ICE2, a gene involved in cold acclimation which determines freezing tolerance in Arabidopsis thaliana. Gene 429 (1-2), 98–103. doi:10.1016/j.gene.2008.10.016
Furuya, T., Matsuoka, D., and Nanmori, T. (2013). Phosphorylation of Arabidopsis thaliana MEKK1 via Ca(2+) signaling as a part of the cold stress response. J. Plant Res. 126 (6), 833–840. doi:10.1007/s10265-013-0576-0
Galiba, G., Quarrie, S. A., Sutka, J., Morgounov, A., and Snape, J. W. (1995). RFLP mapping of the vernalization (Vrn1) and frost resistance (Fr1) genes on chromosome 5A of wheat. Theor. and Appl. Genet. 90 (7-8), 1174–1179. doi:10.1007/BF00222940
Ghorbel, M., Zribi, I., Haddaji, N., Siddiqui, A. J., Bouali, N., and Brini, F. (2023). Genome-wide identification and expression analysis of catalase gene families in triticeae. Plants (Basel) 13 (1), 11. doi:10.3390/plants13010011
Gill, S. S., and Tuteja, N. (2010). Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiology Biochem. 48 (12), 909–930. doi:10.1016/j.plaphy.2010.08.016
Gong, Z., Xiong, L., Shi, H., Yang, S., Herrera-Estrella, L. R., Xu, G., et al. (2020). Plant abiotic stress response and nutrient use efficiency. Sci. China. Life Sci. 63 (5), 635–674. doi:10.1007/s11427-020-1683-x
Guo, J., Ren, Y., Tang, Z., Shi, W., and Zhou, M. (2019). Characterization and expression profiling of the ICE-CBF-COR genes in wheat. PeerJ 7 (1–2), e8190. doi:10.7717/peerj.8190
Hajela, R. K., Horvath, D. P., Gilmour, S. J., and Thomashow, M. F. (1990). Molecular cloning and expression of cor (Cold-Regulated) genes in Arabidopsis thaliana. Plant Physiol. 93 (3), 1246–1252. doi:10.1104/pp.93.3.1246
Hajiahmadi, Z., Abedi, A., Wei, H., Sun, W., Ruan, H., Zhuge, Q., et al. (2020). Identification, evolution, expression, and docking studies of fatty acid desaturase genes in wheat (Triticum aestivum L.). BMC Genomics 21 (1), 778. doi:10.1186/s12864-020-07199-1
Holman, J. D., Schlegel, A. J., Thompson, C. R., and Lingenfelser, J. E. (2011). Influence of precipitation, temperature, and 56 years on winter wheat yields in western Kansas. Crop Manag. 10, 1–10. doi:10.1094/CM-2011-1229-01-RS
Hu, Y., Jiang, L., Wang, F., and Yu, D. (2013). Jasmonate regulates the inducer of cbf expression-C-repeat binding factor/DRE binding factor1 cascade and freezing tolerance in Arabidopsis. Plant Cell 25 (8), 2907–2924. doi:10.1105/tpc.113.112631
Hu, Y., Jiang, Y., Han, X., Wang, H., Pan, J., and Yu, D. (2017). Jasmonate regulates leaf senescence and tolerance to cold stress: crosstalk with other phytohormones. J. Exp. Bot. 68 (6), 1361–1369. doi:10.1093/jxb/erx004
Ichimura, K., Mizoguchi, T., Yoshida, R., Yuasa, T., and Shinozaki, K. (2000). Various abiotic stresses rapidly activate Arabidopsis MAP kinases ATMPK4 and ATMPK6. Plant J. 24 (5), 655–665. doi:10.1046/j.1365-313x.2000.00913.x
Ishizaki-Nishizawa, O., Fujii, T., Azuma, M., Sekiguchi, K., Murata, N., Ohtani, T., et al. (1996). Low-temperature resistance of higher plants is significantly enhanced by a nonspecific cyanobacterial desaturase. Nat. Biotechnol. 14 (8), 1003–1006. doi:10.1038/nbt0896-1003
Jha, U. C., Bohra, A., and Jha, R. (2017). Breeding approaches and genomics technologies to increase crop yield under low-temperature stress. Plant Cell Rep. 36 (1), 1–35. doi:10.1007/s00299-016-2073-0
Jiang, T., Zhai, J. Q., Luo, Y., Su, B. D., Chao, Q. C., Wang, Y. J., et al. (2022). Understandings of assessment reports on climate change impacts, adaptation and vulnerability:progress from IPCC AR5 to IPCC AR6. Trans. Atmos. Sci. 45 (4), 502–511. doi:10.13878/j.cnki.dqkxxb.20220529013
Ju, W., Yang, C. F., Zhang, S. H., Tian, J. C., Hai, Y., and Yang, X. J. (2012). Mapping QTL for cell membrane permeability of leaf treated by low temperature in winter wheat. Crop J. 38 (7), 1247–1252. doi:10.3724/SP.J.1006.2012.01247
Juan, J. X., Yu, X. H., Jiang, X. M., Gao, Z., Zhang, Y., Li, W., et al. (2015). Agrobacterium-mediated transformation of tomato with the ICE1 transcription factor gene. Genet. and Mol. Res. 14 (1), 597–608. doi:10.4238/2015.January.30.1
Kamata, T., and Uemura, M. (2004). Solute accumulation in heat seedlings during cold acclimation: contribution to increased freezing tolerance. Cryoletters 25 (5), 311–322. doi:10.1163/1568540042781748
Kim, D., Alptekin, B., and Budak, H. (2018). CRISPR/Cas9 genome editing in wheat. Funct. and Integr. Genomics 18, 31–41. doi:10.1007/s10142-017-0572-x
Kodama, H., Horiguchi, G., Nishiuchi, T., Nishimura, M., and Iba, K. (1995). Fatty acid desaturation during chilling acclimation is one of the factors involved in conferring low-temperature tolerance to young tobacco leaves. Plant Physiol. 107 (4), 1177–1185. doi:10.1104/pp.107.4.1177
Lalk, I., and Dorffling, K. (1985). Hardening, abscisic acid, proline and freezing resistance in two winter wheat varieties. Physiol. Plant. 63 (3), 287–292. doi:10.1111/j.1399-3054.1985.tb04267.x
Lee, B. H., Henderson, D. A., and Zhu, J. K. (2005). The Arabidopsis cold-responsive transcriptome and its regulation by ICE1. Plant Cell 17 (11), 3155–3175. doi:10.1105/tpc.105.035568
Lei, C., Li, M., Chen, Z., He, W., Liu, B., Liu, S., et al. (2023). Molecular mapping of two novel cold resistance genes in common wheat by 660K SNP array. Mol. Breed. 43 (12), 83. doi:10.1007/s11032-023-01425-w
Li, H., Ding, Y., Shi, Y., Zhang, X., Zhang, S., Gong, Z., et al. (2017). MPK3- and MPK6-mediated ICE1 phosphorylation negatively regulates ICE1 stability and freezing tolerance in Arabidopsis. Dev. Cell 43 (5), 630–642. doi:10.1016/j.devcel.2017.09.025
Limin, A. E., Danyluk, J., Chauvin, L. P., Fowler, D. B., and Sarhan, F. (1997). Chromosome mapping of low-temperature induced Wcs120 family genes and regulation of cold-tolerance expression in wheat. Mol. General Genet. 253 (6), 720–727. doi:10.1007/s004380050376
Limin, A. E., and Fowler, D. B. (2006). Low-temperature tolerance and genetic potential in wheat (Triticum aestivum L.): response to photoperiod, vernalization, and plant development. Planta 224 (2), 360–366. doi:10.1007/s00425-006-0219-y
Liu, L., Cang, J., Yu, J., Wang, X., Huang, R., Wang, J., et al. (2013). Effects of exogenous abscisic acid on carbohydrate metabolism and the expression levels of correlative key enzymes in winter wheat under low temperature. Biosci. Biotechnol. Biochem. 77 (3), 516–525. doi:10.1271/bbb.120752
Liu, X., Fu, L., Qin, P., Sun, Y., Liu, J., and Wang, X. (2019). Overexpression of the wheat trehalose 6-phosphate synthase 11 gene enhances cold tolerance in Arabidopsis thaliana. Gene 710, 210–217. doi:10.1016/j.gene.2019.06.006
Lu, X., Zhang, F., Zhang, C., Li, G., Du, Y., Zhao, C., et al. (2024). TaTPS11 enhances wheat cold resistance by regulating source-sink factor. Plant physiology Biochem. 211, 108695. doi:10.1016/j.plaphy.2024.108695
Lv, D. K., Bai, X., Li, Y., Ding, X. D., Ge, Y., Cai, H., et al. (2010). Profiling of cold-stress-responsive miRNAs in rice by microarrays. Gene 459 (1-2), 39–47. doi:10.1016/j.gene.2010.03.011
Lv, Y., Cang, J., Lu, Q., Yang, N., Feng, M. F., Meng, D. Y., et al. (2017). Bioinformatic analysis and effect of low temperatureand ABA on TabZIP1 expression pattern of winter wheat dongnongdongmai 1. J. Triticeae Crops 37 (1), 22–29. doi:10.7606/j.issn.1009-1041.2017.01.04
Lv, Y., Song, C. H., Lu, Q. W., Tian, Y., Li, H. D., Zhang, D., et al. (2018). The expression characteristics of transcription factors regulated by exogenous ABA in winter wheat (Triticum aestivum) under cold stress. Russ. J. Plant Physiology 65 (6), 842–848. doi:10.1134/s1021443718060134
Lyons, J. M. (1973). Chilling injury in plants. Annu. Rev. Plant Physiology 24 (1), 445–466. doi:10.1146/annurev.pp.24.060173.002305
Malko, M. M., Peng, X., Gao, X., Cai, J., Zhou, Q., Wang, X., et al. (2023). Effect of exogenous calcium on tolerance of winter wheat to cold stress during stem elongation stage. Plants (Basel). 12 (21), 3784. doi:10.3390/plants12213784
Matsumura, T., Tabayashi, N., Kamagata, Y., Souma, C., and Saruyama, H. (2010). Wheat catalase expressed in transgenic rice can improve tolerance against low temperature stress. Physiol. Plant. 116 (3), 317–327. doi:10.1034/j.1399-3054.2002.1160306.x
Miller, A. K., Galiba, G., and Dubcovsky, J. (2006). A cluster of 11 CBF transcription factors is located at the frost tolerance locus Fr-Am2 in Triticum monococcum. Mol. General Genet. 275 (2), 193–203. doi:10.1007/s00438-005-0076-6
Mohseni, S., Che, H., Djillali, Z., Dumont, E., Nankeu, J., and Danyluk, J. (2012). Wheat CBF gene family: identification of polymorphisms in the CBF coding sequence. Genome 55 (12), 865–881. doi:10.1139/gen-2012-0112
Nadarajah, K. K. (2020). ROS homeostasis in abiotic stress tolerance in plants. Int. J. Mol. Sci. 21 (15), 5208. doi:10.3390/ijms21155208
Naya, L., Paul, S., Oswaldo, V.-L., Mendoza-Soto, A. B., Nova-Franco, B., Sosa-Valencia, G., et al. (2014). Regulation of copper homeostasis and biotic interactions by MicroRNA 398b in common bean. Plos One 9, e84416. doi:10.1371/journal.pone.0084416
Ou, X. Q., and Wang, Y. (2019). Preliminary study on wheat breeding for late spring coldness tolerance in south of Huanghuai region. J. Triticeae Crops 39, 560–566. doi:10.7606/j.issn.1009-1041.2019.05.07
Pang, Y., Wu, Y., Liu, C., Li, W., St Amand, P., Bernardo, A., et al. (2021). High-resolution genome-wide association study and genomic prediction for disease resistance and cold tolerance in wheat. Theor. Appl. Genet. 134 (9), 2857–2873. doi:10.1007/s00122-021-03863-6
Pearce, S., Zhu, J., Boldizsár, Á., Vágújfalvi, A., Burke, A., Garland-Campbell, K., et al. (2013). Large deletions in the CBF gene cluster at the Fr-B2 locus are associated with reduced frost tolerance in wheat. Theor. Appl. Genet. 126 (11), 2683–2697. doi:10.1007/s00122-013-2165-y
Peng, K., Tian, Y., Cang, J., Yu, J., Wang, D., He, F., et al. (2022). Overexpression of TaFBA-A10 from winter wheat enhances freezing tolerance in Arabidopsis thaliana. J. Plant Growth Regul. 41 (1), 314–326. doi:10.1007/s00344-021-10304-7
Peng, K., Tian, Y., Sun, X., Song, C., Ren, Z., Bao, Y., et al. (2021). tae-miR399-UBC24 module enhances freezing tolerance in winter wheat via a CBF signaling pathway. J. Agric. food Chem. 69 (45), 13398–13415. doi:10.1021/acs.jafc.1c04316
Skinner, J. S., von Zitzewitz, J., Szucs, P., Marquez-Cedillo, L., Filichkin, T., Amundsen, K., et al. (2005). Structural, functional, and phylogenetic characterization of a large CBF gene family in barley. Plant Mol. Biol. 59 (4), 533–551. doi:10.1007/s11103-005-2498-2
Soleimani, B., Lehnert, H., Babben, S., Keilwagen, J., Koch, M., Arana-Ceballos, F. A., et al. (2022). Genome wide association study of frost tolerance in wheat. Sci. Rep. 12 (1), 5275. doi:10.1038/s41598-022-08706-y
Soltész, A., Smedley, M., Vashegyi, I., Galiba, G., Harwood, W., and Vágújfalvi, A. (2013). Transgenic barley lines prove the involvement of TaCBF14 and TaCBF15 in the cold acclimation process and in frost tolerance. J. Exp. Bot. 64 (7), 1849–1862. doi:10.1093/jxb/ert050
Subedi, K. D., Gregory, P. J., Summerfield, R. J., and Gooding, M. J. (1998). Cold temperatures and boron deficiency caused grain set failure in spring wheat (Triticum aestivum L.). Field Crops Res. 57 (3), 277–288. doi:10.1016/S0378-4290(97)00148-2
Sun, X., Fan, G., Su, L., Wang, W., Liang, Z., Li, S., et al. (2015). Identification of cold-inducible microRNAs in grapevine. Front. plant Sci. 6, 595. doi:10.3389/fpls.2015.00595
Sutka, J. (1994). “Genetic control of frost tolerance in wheat (Triticum aestivum L),” in Breeding fodder crops for marginal conditions. Developments in plant breeding. Editors O. A. Rognli, E. Solberg, and I. Schjelderup (Dordrecht: Springer). doi:10.1007/978-94-011-0966-6_42
Sutka, J. (2001). Genes for frost resistance in wheat. Euphytica 119 (1-2), 169–177. doi:10.1023/a:1017520720183
Takumi, S., Shimamura, C., and Kobayashi, F. (2008). Increased freezing tolerance through up-regulation of downstream genes via the wheat CBF gene in transgenic tobacco. Plant Physiology Biochem. 46 (2), 205–211. doi:10.1016/j.plaphy.2007.10.019
Tian, Y., Peng, K., Bao, Y., Zhang, D., Meng, J., Wang, D., et al. (2021). Glucose-6-phosphate dehydrogenase and 6-phosphogluconate dehydrogenase genes of winter wheat enhance the cold tolerance of transgenic Arabidopsis. Plant physiology Biochem. 161, 86–97. doi:10.1016/j.plaphy.2021.02.005
Tian, Y., Peng, K., Ma, X., Ren, Z., Lou, G., Jiang, Y., et al. (2023). Overexpression of TaMYB4 confers freezing tolerance in Arabidopsis thaliana. Int. J. Mol. Sci. 24 (13), 11090. doi:10.3390/ijms241311090
Tiwari, B., Habermann, K., Arif, M. A., Weil, H. L., Garcia-Molina, A., Kleine, T., et al. (2020). Identification of small RNAs during cold acclimation in Arabidopsis thaliana. BMC Plant Biol. 20 (1), 298. doi:10.1186/s12870-020-02511-3
Tóth, B., Galiba, G., Fehér, E., Sutka, J., and Snape, J. W. (2003). Mapping genes affecting flowering time and frost resistance on chromosome 5B of wheat. Theor. Appl. Genet. 107 (3), 509–514. doi:10.1007/s00122-003-1275-3
Tshikunde, N. M., Mashilo, J., Shimelis, H., and Odindo, A. (2019). Agronomic and physiological traits, and associated quantitative trait loci (QTL) affecting yield response in wheat (Triticum aestivum L.): a review. Front. plant Sci. 10, 1428. doi:10.3389/fpls.2019.01428
Türkan, I., Bor, M., Izdemir, F., and Koca, H. (2005). Differential responses of lipid peroxidation and antioxidants in the leaves of drought-tolerant P. acutifolius Gray and drought-sensitive P. vulgaris L. subjected to polyethylene glycol mediated water stress. Plant Sci. 168 (1), 223–231. doi:10.1016/j.plantsci.2004.07.032
Valluru, R., Link, J., and Claupein, W. (2012). Consequences of early chilling stress in two Triticum species: plastic responses and adaptive significance. Plant Biol. Stuttg. Ger. 14 (4), 641–651. doi:10.1111/j.1438-8677.2011.00540.x
Venzhik, Y., Deryabin, A., and Zhukova, K. (2024). Au-based nanoparticles enhance low temperature tolerance in wheat by regulating some physiological parameters and gene expression. Plants (Basel) 13 (9), 1261. doi:10.3390/plants13091261
Vij, S., and Tyagi, A. K. (2007). Emerging trends in the functional genomics of the abiotic stress response in crop plants. Plant Biotechnol. J. 5 (3), 361–380. doi:10.1111/j.1467-7652.2007.00239.x
Wang, R., Yu, M., Xia, J., Xing, J., Fan, X., Xu, Q., et al. (2022). Overexpression of TaMYC2 confers freeze tolerance by ICE-CBF-COR module in Arabidopsis thaliana. Front. plant Sci. 13, 1042889. doi:10.3389/fpls.2022.1042889
Wang, R., Yu, M., Zhao, X., Xia, J., Cang, J., and Zhang, D. (2024). Overexpression of TaMPK3 enhances freezing tolerance by increasing the expression of ICE-CBF-COR related genes in the Arabidopsis thaliana. Funct. plant Biol. 51, FP23144. doi:10.1071/FP23144
Whaley, J. M., Kirby, E. J. M., Spink, J. H., Foulkes, M. J., and Sparkes, D. L. (2004). Frost damage to winter wheat in the UK: the effect of plant population density. Eur. J. Agron. 21 (1), 105–115. doi:10.1016/S1161-0301(03)00090-X
Yang, C., and Yuan, P. (2014). The occurrence of wheat frost damage and preventive measures in the China Huanghuai region. Mod. Agric. Sci. Technol. 2 (2). doi:10.3969/j.issn.1007-5739.2014.02.060
Yang, T., Chaudhuri, S., Yang, L., Du, L., and Poovaiah, B. W. (2010). A calcium/calmodulin-regulated member of the receptor-like kinase family confers cold tolerance in plants. J. Biol. Chem. 285 (10), 7119–7126. doi:10.1074/jbc.M109.035659
Yu, J., Zhang, L., Cui, H., Zhang, Y. X., Cang, J., Hao, Z. B., et al. (2008). Physiological and biochemical characteristics of dongnongdongmai 1. Crop J. 34 (11), 2019–2025. doi:10.3724/SP.J.1006.2008.02019
Yu, M. M., Wang, R., Xia, J. Q., Li, C., Xu, Q. H., Cang, J., et al. (2023). JA-induced TaMPK6 enhanced the freeze tolerance of Arabidopsis thaliana through regulation of ICE-CBF-COR module and antioxidant enzyme system. Plant Sci. 329, 111621. doi:10.1016/j.plantsci.2023.111621
Zeng, Y., Yu, J., Cang, J., Liu, L., Mu, Y., Wang, J., et al. (2011). Detection of sugar accumulation and expression levels of correlative key enzymes in winter wheat (Triticum aestivum) at low temperatures. J. Agric. Chem. Soc. Jpn. 75 (4), 681–687. doi:10.1271/bbb.100813
Zhang, L., Zhao, G., Xia, C., Jia, J., Liu, X., and Kong, X. (2012). Overexpression of a wheat MYB transcription factor gene TaMYB56-B enhances tolerances to freezing and salt stresses in transgenic Arabidopsis. Gene 505 (1), 100–107. doi:10.1016/j.gene.2012.05.033
Zhang, N., Li, L., Zhang, L., Li, J., Fang, Y., Zhao, L., et al. (2020). Abscisic acid enhances tolerance to spring freeze stress and regulates the expression of ascorbate–glutathione biosynthesis-related genes and stress-responsive genes in common wheat. Mol. Breed. 40 (11), 108–118. doi:10.1007/s11032-020-01187-9
Keywords: Triticum aestivum L., cold resistance, gene, mechanism, breeding strategy
Citation: Ma S, Huang X, Zhao X, Liu L, Zhang L and Gan B (2024) Current status for utilization of cold resistance genes and strategies in wheat breeding program. Front. Genet. 15:1473717. doi: 10.3389/fgene.2024.1473717
Received: 31 July 2024; Accepted: 10 October 2024;
Published: 22 October 2024.
Edited by:
Yang Yang, Shanxi Agricultural University, ChinaReviewed by:
Qinghua Yang, Northwest A & F University Hospital, ChinaCopyright © 2024 Ma, Huang, Zhao, Liu, Zhang and Gan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Binjie Gan, Z2FuYmluamllQDE2My5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.