 Ana Carolina Proença da Fonseca1,2,3,4*Izadora Sthephanie da Silva Assis1,2Kaio Cezar Rodrigues Salum1,2,5Lohanna Palhinha1Gabriella de Medeiros Abreu2,6Verônica Marques Zembrzuski2Mario Campos Junior2José Firmino Nogueira-Neto7Amanda Cambraia2Mauro Lucio Ferreira Souza Junior8Clarissa Menezes Maya-Monteiro1Pedro Hernán Cabello2Patrícia Torres Bozza1João Regis Ivar Carneiro5
Ana Carolina Proença da Fonseca1,2,3,4*Izadora Sthephanie da Silva Assis1,2Kaio Cezar Rodrigues Salum1,2,5Lohanna Palhinha1Gabriella de Medeiros Abreu2,6Verônica Marques Zembrzuski2Mario Campos Junior2José Firmino Nogueira-Neto7Amanda Cambraia2Mauro Lucio Ferreira Souza Junior8Clarissa Menezes Maya-Monteiro1Pedro Hernán Cabello2Patrícia Torres Bozza1João Regis Ivar Carneiro5- 1Laboratory of Immunopharmacology, Oswaldo Cruz Institute, Oswaldo Cruz Foundation, Rio de Janeiro, Brazil
- 2Human Genetics Laboratory, Oswaldo Cruz Institute, Oswaldo Cruz Foundation, Rio de Janeiro, Brazil
- 3Genetics Laboratory, Grande Rio University/AFYA, Rio de Janeiro, Brazil
- 4Postgraduate Program in Translational Biomedicine, Grande Rio University/AFYA, Rio de Janeiro, Brazil
- 5Clementino Fraga Filho University Hospital, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil
- 6Josué de Castro Nutrition Institute, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil
- 7Department of Pathology and Laboratory, Rio de Janeiro State University, Rio de Janeiro, Brazil
- 8Laboratory of Human and Medical Genetics, Institute of Biological Sciences, Federal University of Pará, Belém, Brazil
Introduction: Obesity is a multifactorial disease associated with the development of many comorbidities. This disease is associated with several metabolic alterations; however, it has been shown that some individuals with obesity do not exhibit metabolic syndrome. Adipose tissue neutralizes the detrimental effects of circulating fatty acids, ectopic deposition, and inflammation, among others, through its esterification into neutral lipids that are stored in the adipocyte. However, when the adipocyte is overloaded, i.e., its expansion capacity is exceeded, this protection is lost, resulting in fatty acid toxicity with ectopic fat accumulation in peripheral tissues and inflammation. In this line, this study aimed to investigate whether polymorphisms in genes that control adipose tissue fat storage capacity are potential biomarkers for severe obesity susceptibility and also metabolic complications.
Methods: This study enrolled 305 individuals with severe obesity (cases, BMI≥35 kg/m2) and 196 individuals with normal weight (controls, 18.5≤BMI≤24.9 kg/m2). Demographic, anthropometric, biochemical, and blood pressure variables were collected from the participants. Plasma levels of leptin, resistin, MCP1, and PAI1 were measured by Bio-Plex 200 Multiplexing Analyzer System. Genomic DNA was extracted and variants in DBC1 (rs17060940), SIRT1 (rs7895833 and rs1467568), UCP2 (rs660339), PPARG (rs1801282) and ADRB2 (rs1042713 and rs1042714) genes were genotyped by PCR allelic discrimination using TaqMan® assays.
Results: Our findings indicated that SIRT1 rs7895833 polymorphism was a risk factor for severe obesity development in the overdominant model. SIRT1 rs1467568 and UCP2 rs660339 were associated with anthropometric traits. SIRT1 rs1467568 G allele was related to lower medians of body adipose index and hip circumference, while the UCP2 rs660339 AA genotype was associate with increased body mass index. Additionally, DBC1 rs17060940 influenced glycated hemoglobin. Regarding metabolic alterations, 27% of individuals with obesity presented balanced metabolic status in our cohort. Furthermore, SIRT1 rs1467568 AG genotype increased 2.5 times the risk of developing metabolic alterations. No statistically significant results were observed with Peroxisome Proliferator-Activated Receptor Gama and ADRB2 polymorphisms.
Discussion/Conclusion: This study revealed that SIRT1 rs7895833 and rs1467568 are potential biomarkers for severe obesity susceptibility and the development of unbalanced metabolic status in obesity, respectively. UCP2 rs660339 and DBC1 rs17060940 also showed a significant role in obesity related-traits.
Introduction
Obesity continues to grow as a serious public health concern. Based on the latest data from the NCD Risk Factor Collaboration, almost 650 million adults had obesity in 2016 (defined by a body mass index [BMI] of ≥30 kg/m2) (WHO, 2022). The situation is just as alarming since it is estimated that 1 billion adults will exhibit this phenotype in 2025 (Abarca-Gómez et al., 2017). Obesity is associated with reduced life expectancy due to increased mortality from chronic diseases, such as type 2 diabetes mellitus (T2DM), hypertension, cardiovascular disease, and cancers (WHO, 2022). More recently, it has been highlighted that individuals with obesity present an increased risk for severity and mortality in COVID-19 and Influenza infections (Zhao et al., 2020).
Although obesity is commonly seen as the primary cause of metabolic syndrome and other human metabolic diseases, there is a large variation in the subject’s risk of developing these comorbidities. The effective fat storage in the adipose tissue is suggested to be a key factor in the protection against metabolic dysfunctions. When the capacity of fat storage in adipose tissue depots is exceeded, the excess fat can accumulate in peripheral tissues and cause inflammatory and metabolic dysfunctions in a process known as lipotoxicity (Listenberger et al., 2003; Wang et al., 2008; Unger and Scherer, 2010; Almandoz et al., 2013; Escande et al., 2015). In this context, several studies focus on the molecular pathways involved in adipose tissue fat storage capacity and inflammation, since adipocyte fat accumulation would delay the development of metabolic disease (Lê et al., 2011; Escande et al., 2015; Moreno-Navarrete et al., 2015).
DBC1 gene (Deleted in breast cancer 1) encodes a pleiotropic nuclear protein which induces the expression of nuclear factor kappa B (NF-kB) that regulates inflammatory cytokines through inhibition of sirtuin 1 activity (Lê et al., 2011; Escande et al., 2015; Moreno-Navarrete et al., 2015). Moreno-Navarrete and coworkers (2015) reported that DBC1 knockdown (KD) in mice resulted in a significant reduction in the expression of inflammatory genes and increased intracellular lipid accumulation. These results suggest that the DBC1 KD mice are protected against adipose tissue inflammation (Moreno-Navarrete et al., 2015). Escande and colleagues (2015) also observed that the DBC1 knockout increased the fat accumulation capacity in vitro and in vivo, resulting in a “healthy obesity” phenotype (Escande et al., 2015). In this context, the increased activation of NF-kB in adipose tissue is associated with decreased adipogenic capacity (Lê et al., 2011).
Sirtuin 1, encoded by SIRT1 gene, is a nicotinamide adenine dinucleotide (NAD+)-dependent protein deacetylase that has anti-inflammatory activities in adipose tissue by direct deacetylation of NF-kB and chromatin remodeling at inflammatory gene promoters (Kotas et al., 2013; Moreno-Navarrete et al., 2015). SIRT1 also acts as a cellular energy sensor and drives glucose and fat metabolism by regulating the expression of important genes, such as PPARG and UCP2 (Picard et al., 2004; Bordone et al., 2006; Chaudhary and Pfluger, 2009). Picard and coworkers (2014) showed that SIRT is a suppressor of adipogenesis by inhibiting the expression of genes responsible for promoting the differentiation of white adipocytes and lipid accumulation (Picard et al., 2004).
PPARG (Peroxisome Proliferator-Activated Receptor Gama) gene encodes two major isoforms (PPARγ1 and PPARγ2). PPARγ2 is highly expressed in adipose tissue and, upon activation, forms a heterodimer with retinoid X receptor, regulating the expression of numerous target genes (Spiegelman, 1998; Auwerx, 1999). PPARG plays an important role in adipose tissue formation and also regulates lipid and glucose metabolism. Therefore, PPARG is a known target in the study of T2DM and obesity (Luan et al., 2001; Ali et al., 2009; Hasan et al., 2017; Bakhashab et al., 2020). PPARG is repressed by SIRT1, which results in attenuated adipogenesis and increased lipolysis in white adipose tissue (Picard et al., 2004; Chaudhary and Pfluger, 2009; Dmitrenko et al., 2022). In addition, the reduction of PPARG also results in the improvement of insulin resistance and inhibition of glucose production (Spiegelman, 1998; Auwerx, 1999; Bakhashab et al., 2020).
UCP2 encodes an anion-carrier protein, called uncoupling protein 2. This protein is located in the mitochondrial inner membrane which participates in the energy metabolism of cells (Nicoletti et al., 2017). In humans, five UCP members were identified (UCP1—UCP5). UCP2 is expressed in a wide range of tissues, including the pancreas, central nervous system, and white adipose tissue. UCP2 plays an important role in inhibiting the formation of reactive oxygen species, negative regulation of insulin secretion by pancreatic beta cells, and fatty acid metabolism, which are mechanisms associated with metabolic disease (Pecqueur et al., 2001; Azzu and Brand, 2010; Souza et al., 2011). Interestingly, PPARγ increases the expression of UCP2, while SIRT1 inhibits the expression (Affourtit and Brand, 2008). During food deprivation, Sirt1 negatively regulates UCP2, leading to ATP production and increasing insulin secretion by pancreatic β cells (Chaudhary and Pfluger, 2009).
Lastly, the β2-adrenergic receptor is a major lipolytic receptor in human adipocytes, codified by the ADRB2 gene. This receptor stimulated by catecholamines in adipose tissue, increases lipolysis and resultsin the reduction of adipocyte size and lipid content (Lafontan et al., 2000; Jalba et al., 2008; Kurylowicz et al., 2015). The expression of ADRB2 is lower in adipose tissue of subjects with obesity when compared to normal-weight individuals. Furthermore, lower expression levels of ADRB2 were observed in visceral adipose tissue when compared to subcutaneous adipose tissue of subjects with obesity (Kurylowicz et al., 2015).
Obesity and comorbidities have a significant genetic component, and the disease development is dependent on the interaction between genetic background and environmental factors (da Fonseca et al., 2017). However, there are still controversies and gaps about the responsible genes, their common variants, and obesity (or metabolic complications in obesity). In the present study, we have investigated common variants in DBC1 (rs17060940), SIRT1 (rs7895833 and rs1467568), UCP2 (rs660339), PPARG (rs1801282) and ADRB2 (rs1042713 and rs1042714) (Table 1).
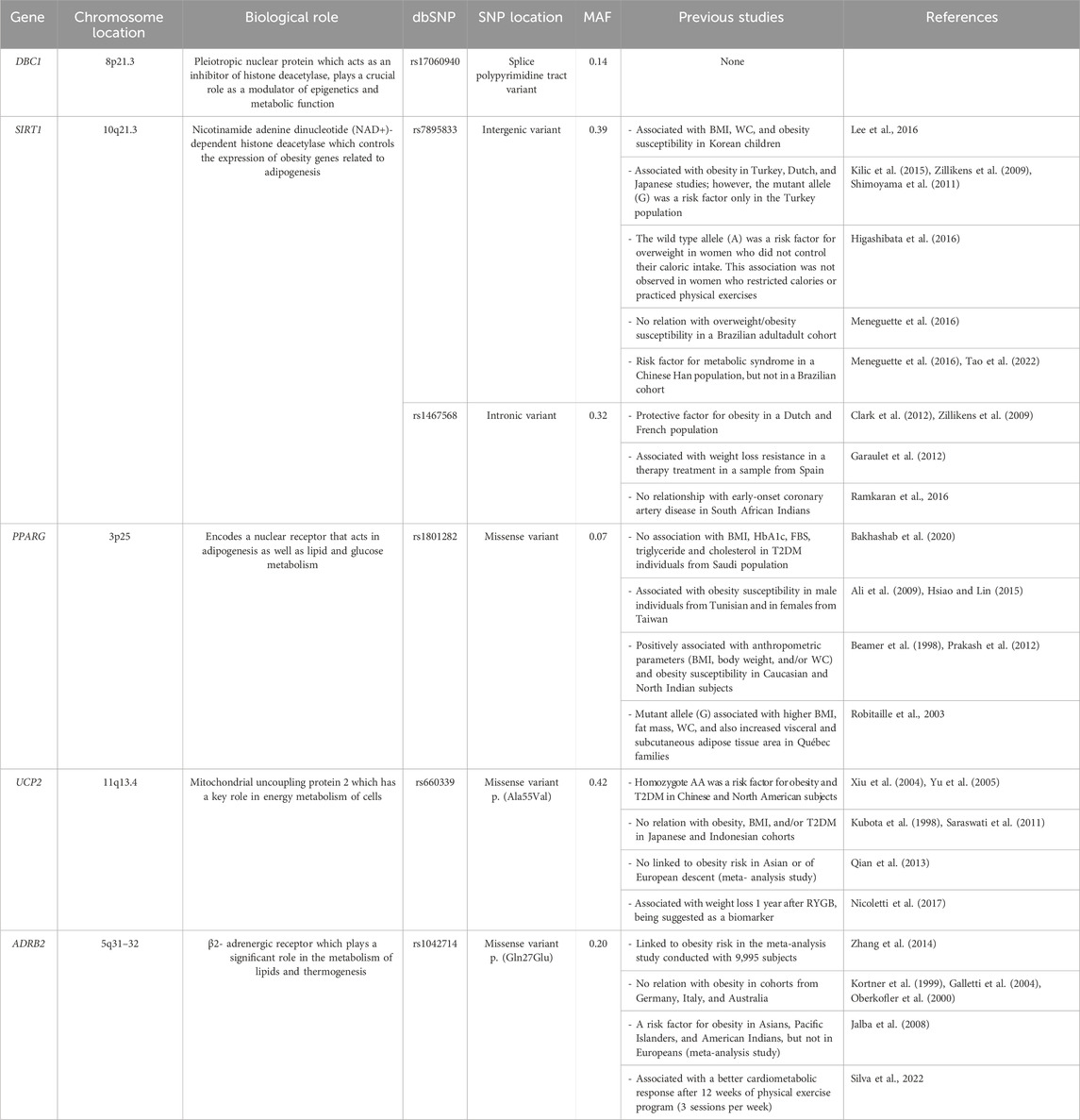
Table 1. Previous findings of the candidate genes and common variants selected in the study.
In the literature, it was observed a significant increase in the SIRT1 protein level with age, and found that older individuals carrying rs7895833 AG genotype had the highest SIRT1 levels. This suggests that the rs7895833 polymorphism may be associated with differences in SIRT1 protein expression, potentially through mechanisms that affect gene expression or protein stability (Kilic et al., 2015a). Functional studies of the PPARG rs1801282 polymorphism showed that the presence of risk allele is associated with decreased binding affinity to promoter element, which impacts on ability to transactivate its responsive promoters. This molecular impact could influence adipogenesis and glucose/lipid metabolism, contributing to lower BMI and insulin sensitivity among individuals carrying PPARG rs1801282 risk allele (Deeb et al., 1998; Rocha et al., 2015). UCP2 rs660339 variant influences the reduction of 24-hour-resting energy expenditure and also decreases the fat oxidation rate that could contribute to body weight regulation (Astrup et al., 1999). ADRB2 rs1042713 polymorphism was shown to reduce thermogenic response to its receptor stimulation, while ADRB2 rs1042714 did not exhibit the same effect (Oomen et al., 2005). Recently, a functional in vitro study of ADRB2 polymorphisms (rs1042713 and rs1042714) showed that mRNA expression was significantly decreased in mutated cells when compared to wild-type (Xie et al., 2019). Our study also chose DBC1 (rs17060940) and SIRT1 (rs1467568) polymorphisms. Despite lacking functional studies, these variants or genes have been already investigated or linked to obesity (or related traits) (Garaulet et al., 2012; Jurgens et al., 2022). Therefore, the present study aimed to investigate whether common variants in DBC1, SIRT1, PPARG, UCP2, and ADRB2 genes are potential biomarkers for severe obesity susceptibility and/or metabolic complications in a Brazilian population. These results could provide useful knowledge for clinical management and risk assessment of obesity and/or metabolic dysfunctions.
Materials and methods
Study subjects
A case-control cross-sectional study was performed with 501 adult individuals, aged 18—70 years, from Rio de Janeiro, Brazil. The inclusion criteria were participants with normal weight (controls, 18.5≤ BMI ≤24.9 kg/m2) and severe obesity (cases, BMI ≥35 kg/m2). The exclusion criteria included (1) pregnancy, (2) lactation, and (3) the use of medication to lose or gain weight directly. The case group was recruited from a non-governmental organization, called Rescue Group to Self-Esteem and Citizenship of the Obese (in Portuguese, “Grupo de Resgate à Autoestima e Cidadania do Obeso”). All case participants are candidates for bariatric surgery. The control group consisted of volunteers from public hospitals in the same city. Both cases and controls share a common geographic origin in Southeastern Brazil. All subjects have signed an informed consent form before the enrollment of the study. The protocol of the study was approved by the ethics committee of the Oswaldo Cruz Foundation (CAAE: 09225113.0.0000/Protocol No: 346.634).
Demographic and lifestyle factors
At the time of recruitment, a standardized questionnaire was applied to each participant by trained interviewers. Race/skin color was self-reported and divided according to the criteria of the Demographic Census conducted through the Brazilian Institute of Geography and Statistics. Marital status was categorized as single, married/cohabiting, separated/divorced, and widower. Smoking habits were assessed as smoking status, divided into “never smoked” or “already smoked” (current smokers and ex-smokers). Physical activity information was classified according to the activities performed in the last month (“yes” or “no”).
Anthropometric parameters, blood biochemistry, and hormone analysis
All participants were examined after an overnight fast. Body weight, waist, and hip circumference were measured as described earlier (da Fonseca et al., 2019). BMI was calculated as weight (kilograms) divided by the square of height (meters). Body adiposity index (BAI) was calculated by the formula: hip circumference/(height1.5)-18. It is used to estimate the pattern of body fat distribution (Bergman et al., 2011). Systolic and diastolic blood pressure was measured in the sitting position using a wrist blood pressure monitor.
Fasting plasma glucose, total cholesterol (TC), high-density lipoprotein cholesterol (HDL-c), and triglyceride (TG) were measured using the oxidase-peroxidase method (BioSystems). Low-density lipoprotein cholesterol (LDL-c) levels were calculated by the Friedewald formula (LDL-c = TC—HDLc—TG/5). C-reactive protein (CRP) and glycated hemoglobin levels were measured by the latex agglutination method and turbidimetric inhibition immunoassay (TINIA), respectively.
Leptin, resistin monocyte chemoattractant protein-1/CCL2 (MCP1), and plasminogen activator inhibitor-1 (PAI-1) were measured using Human Adipocyte Magnetic Bead (Millipore-Merck [cat# HADCYMAG-61k]) on Bio-Plex 200 Multiplexing Analyzer System, according to the manufacturer’s protocol.
Metabolic health status in obesity
Participants with obesity were classified as balanced metabolic obesity (BMO) and unbalanced metabolic obesity (UMO) according to the presence/absence of metabolic abnormalities. Metabolic status was obtained based on components of metabolic abnormalities: fasting glucose level (≥100 mg/dL or drug treatment for elevated glucose), triglycerides (≥150 mg/dL or drug treatment for elevated triglycerides), HDL-cholesterol (<40 mg/dL in men and <50 mg/dL in women or drug treatment for reduced HDL-cholesterol), and blood pressure (≥130/85 mmHg or drug treatment for hypertension). Participants with equal or less than 1 component of metabolic syndrome were classified as BMO, and equal or more than 2 components were defined as UMO (Grundy et al., 2004; Eckel et al., 2010; Blüher, 2020).
Molecular analyses
Genomic DNA was extracted from peripheral blood cells using QIAamp Blood Kit (Qiagen, Valencia, CA, United States), according to the manufacturer’s protocol. All samples were genotyped for polymorphisms in DBC1 (rs17060940), SIRT1 (rs7895833 and rs1467568), UCP2 (rs660339), PPARG (rs1801282) and ADRB2 (rs1042713 and rs1042714) genes by real-time PCR using TaqMan® assays (ThermoFisher, Foster City, CA, United States) in a StepOne® Plus Real-Time PCR System (Supplemental Table S1). Allelic discrimination plots were analyzed by StepOne software v2.3.
Statistical analyses
The normality of continuous variables was analyzed by Kolmogorov–Smirnov and Shapiro–Wilk tests. All data were shown as non-normally dispersed. The distribution of continuous and categorical variables in groups was presented as median (interquartile range) and number (percentage), respectively. Comparison of clinical, anthropometric, biochemical, and plasma biomarkers between cases and controls was performed using Mann-Whitney (quantitative variables) and Chi-square tests (qualitative variables). These statistical analyses were performed using SPSS (SPSS, Chicago, IL, United States). Genotype and allele frequencies were calculated by gene counting. The balance between the genotypes was checked by the Hard-Weinberg Equilibrium (HWE) test. A Pairwise linkage disequilibrium analysis was carried out in ADRB2 and SIRT1 variants using Haploview v.4.2. The genetic association was analyzed and controlled for age, gender, skin color, physical activity, and marital status using SNPassoc package (http://CRAN.Rproject.org/package=SNPassoc). The results are shown as odds ratio (OR) with 95% confidence interval (CI). An association analysis between genetic variants and obesity susceptibility was performed under five different genetic models (codominant, dominant, recessive, overdominant and log-additive). The best genetic model was determined using Akaike Information Criteria (AIC) (González et al., 2007). Linear regression analyses were performed to test the association between studied variants and the continuous variables (logarithmically transformed before the test). Age, gender, race/skin color, physical activity, and marital status were used as covariables for body weight and BMI. These variables and BMI were used as confounders for all other continuous parameters studied. The level of significance was set at p-value < 0.008 (Bonferroni correction for multiple tests).
According to the Quanto power and sample size calculator, the genetic variants analyzed here have at least 80% power to detect effect sizes between 1.35–1.65, based on the minor allele frequency (MAF) of these alterations (https://keck.usc.edu/biostatistics/software/).
Results
Characteristics of the study population
A total of 501 individuals were enrolled in this study, stratified into participants with severe obesity (cases, n = 305) and normal weight (controls, n = 196). The background characteristics of the study subjects are shown in Table 2. As has been known and reported, individuals with severe obesity presented higher anthropometric, blood pressure, and biochemical levels when compared to control participants. In our study, the individuals with normal weight had higher values for height and HDL cholesterol. As expected, individuals with severe obesity had higher levels of leptin, PAI-1, and resistin when compared to controls.
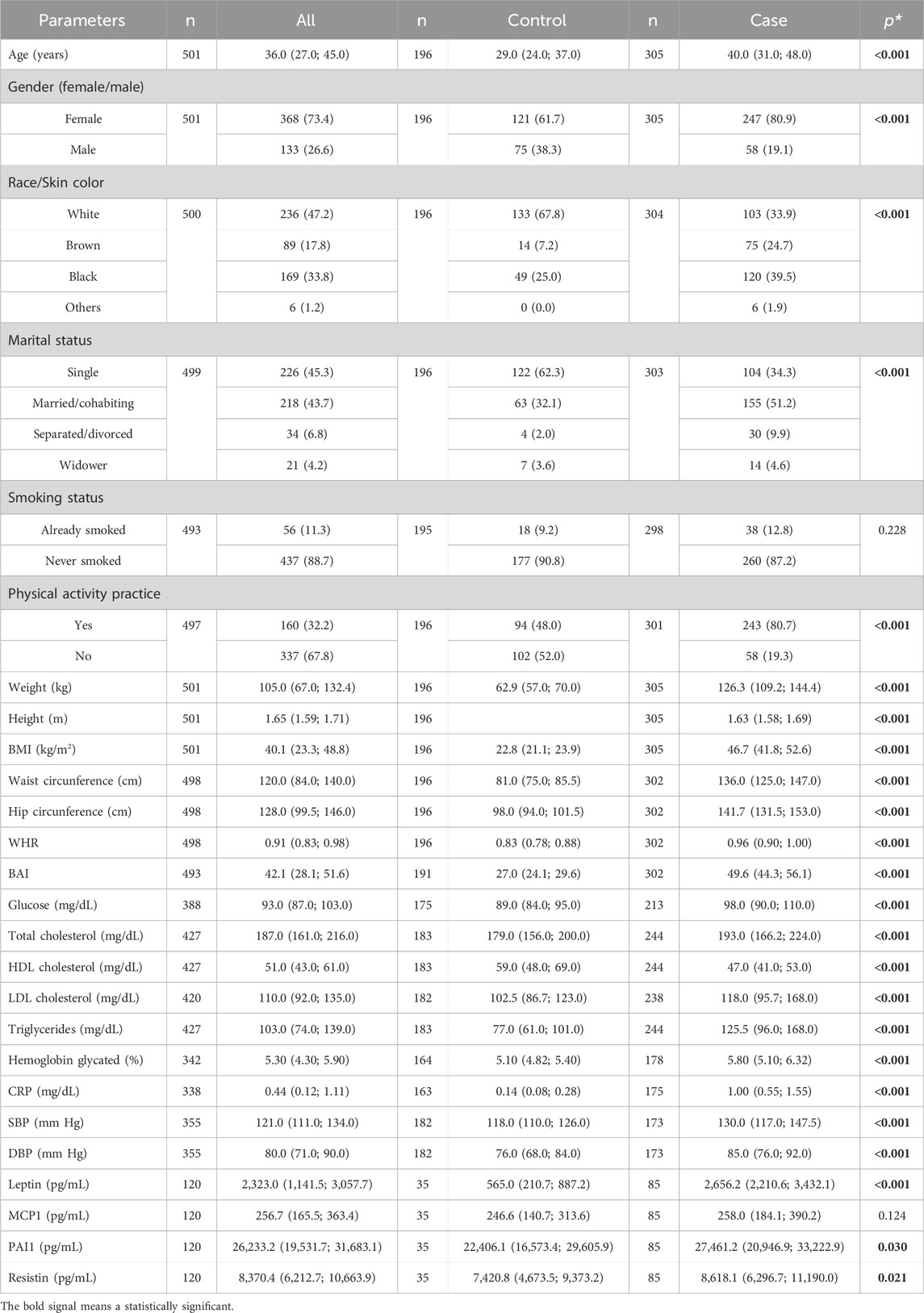
Table 2. Basic clinical characteristics of the studied population.
Furthermore, our data showed significant differences in age, gender, color skin/ethnicity, physical activity practice, and marital status between case and control groups. Thus, these five variables were further included as covariates in the logistic and the linear regression model to remove bias from confounding factors (model 2).
Association analyses of polymorphisms studied and severe obesity risk
The sample details about genotype and allele frequencies are presented in Table 3. PPARG rs1801282 polymorphism showed a low frequency in our cohort since the homozygous GG genotype was found only in one control and two case subjects. Additionally, DBC1 (rs17060940) TT genotype was only observed in the case group (16 individuals). Thus, genetic analyses of PPARG rs1801282 and DBC1 rs17060940 were performed in the dominant model and log-additive. Regarding HWE analyses, only ADRB2 rs1042713 polymorphism was deviated for controls and was excluded in the following analyses (p < 0.05) (Supplemental Table S2). Furthermore, our analysis demonstrated that SIRT1 (rs7895833 and rs1467568; D’ = 0.973; r2 = 0.244; LOD = 37.91) and ADRB2 (rs1042713 and rs1042714; D’ = 0.829; r2 = 0.223; LOD = 31.86) are in linkage disequilibrium.
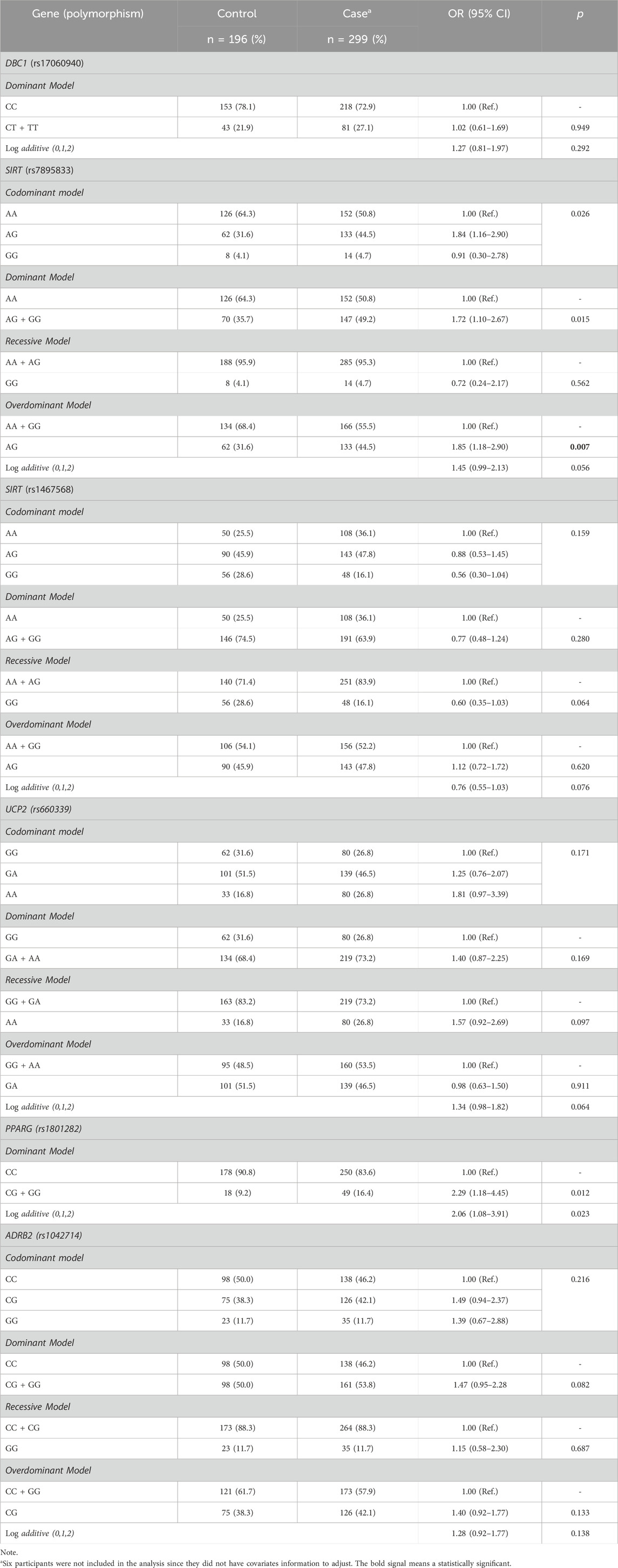
Table 3. Association of common variants and severe obesity susceptibility.
The results of our case-control association analyses are also shown in Table 3. The genotype frequencies of SIRT1 rs7895833 differed significantly between case and control groups, indicating that this polymorphism is associated with severe obesity risk. The frequency of SIRT1 rs7895833 AG genotype was higher in the case group when compared to the control (44.5% vs. 31.6%, respectively). To explore this association, dominant, recessive, overdominant and log-additive models were carried out. SIRT1 rs7895833 was associated with severe obesity in overdominant model. In this line, overdominant was the lowest AIC score. The SIRT1 rs7895833 AG was observed as a genetic predisposition factor for severe obesity development, increasing the risk more than 1.8 times [OR = 1.85 (1.18–2.90); p = 0.007] when compared to GG + AA genotypes. No association was observed between severe obesity and DBC1 (rs17060940), SIRT1 (rs1467568), PPARG (rs1801282), UCP2 (rs660339), and ADRB2 (rs1042714).
Comparisons of metabolic variables according to polymorphisms studied
The effect of the studied polymorphisms on metabolic variables was also investigated in our sample (Supplemental Table S3). DBC1 rs17060940 was associated with glycated hemoglobin, in which the presence of a mutated allele was associated with higher levels. SIRT1 rs1467568 was associated with BAI and HC, in which the median values decreased according to the number of risk alleles. Thus, carriers of two mutated alleles (AA) had lower median values when compared to none or one. Finally, UCP2 rs660339 influenced BMI. Interestingly, our results highlighted that the UCP2 rs660339 AA genotype had an increased BMI median when compared to GG or GA genotypes. None of the other common variants studied was seen as related to the metabolic variables. We also did not observe an association between the polymorphisms studied and the levels of adipokines and inflammatory markers.
Metabolic health status and association analyses of polymorphisms and hormones studied
Metabolic health status was obtained in 256 participants with obesity, in which 27.3% were BMO and 72.7% were UMO. Interestingly, our data indicated that individuals with UMO have increased levels of leptin when compared to BMO (2,830.5 [2,238.5; 3,812.1] vs. 2,350.6 [1,745.5; 2,656.2], respectively) (p = 0.017). However, this result did not remain after correcting for multiple tests. No significant results were observed with MCP1, PAI1, and resistin (data not shown). In addition, our results indicated that SIRT1 rs1467568 was associated with metabolic health status in our cohort (Table 4). The frequency of the SIRT1 rs1467568 AG genotype was higher in the UMO group when compared to BMO (53.8% vs. 32.9%, respectively). To deeply investigate this association, different genetic models were performed. Our results indicated that SIRT1 rs1467568 was associated with metabolic health status in overdominant models. Individuals carrying the heterozygous genotype (AG) were 2.5 times more likely to develop UMO when compared to GG + AA [OR = 2.49 (1.36—4.56); p = 0.003]. No association was observed between metabolic healthy status and DBC1 (rs17060940), SIRT (rs7895833), UCP2 (rs660339), PPARG (rs1801282), and ADRB2 (rs1042714) (data not shown).
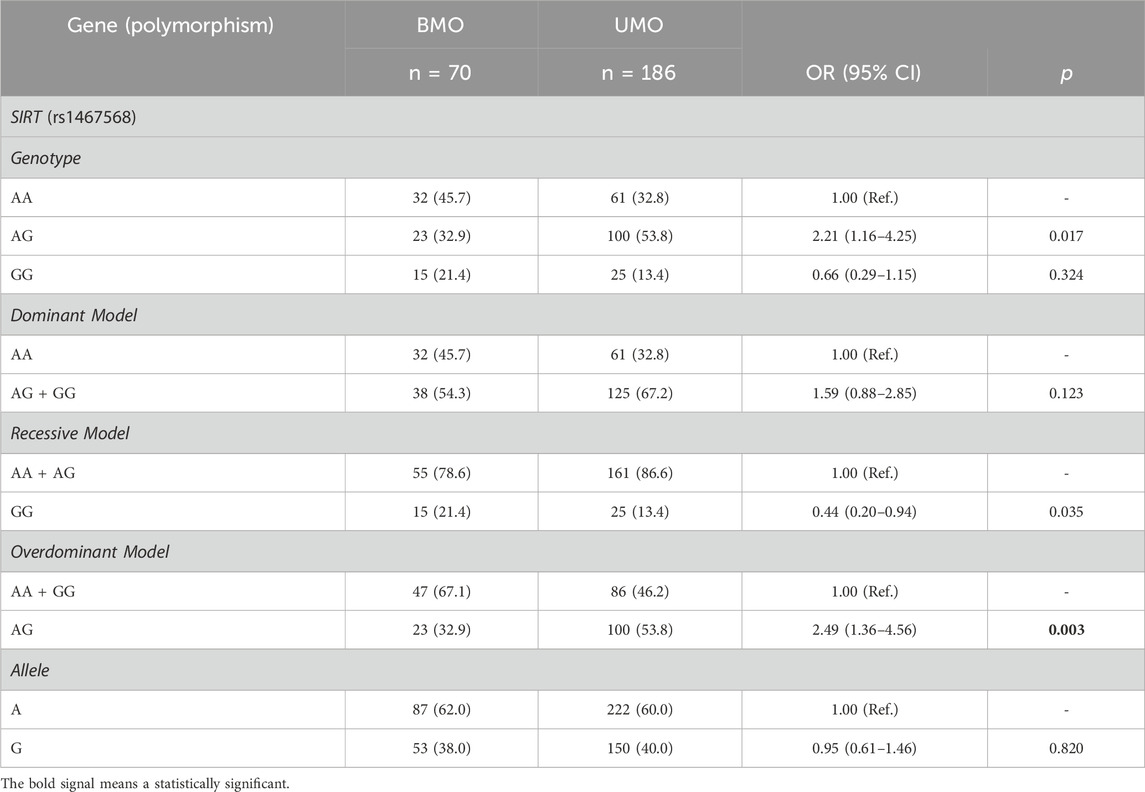
Table 4. Association of SIRT rs1467568 polymorphism and metabolic status.
Discussion
Obesity is a serious public health issue, and understanding its genetic background has gained importance in the world. Several studies have led to the identification of genes that contribute to obesity; however, only a small fraction of its genetic heritability has been elucidated. Additionally, obesity is considered a risk factor for metabolic disorders, although there is a large variation in the individual risk of developing these alterations. Our study focused on the molecular pathway involved in adipose tissue fat capacity and inflammation to investigate potential genetic biomarkers for severe obesity and metabolic health status in obesity. To this end, we performed a Brazilian case-control study and genotyped subjects with severe obesity (n = 305) and normal-weight (n = 196) for common variants in DBC1, SIRT1, UCP2, PPARG and ADRB2 genes. We also measured important adipokines and hormones classically linked to obesity, such as leptin, MCP1, PAI-1, and resistin, in order to characterize our sample as well as to evidence possible associations between the variants and the secretion of these mediators.
Despite this important role in adipogenesis and body metabolic function, information about the association between DBC1 polymorphisms and obesity is lacking in the literature. Our study investigated a splice polypyrimidine tract variant in DBC1 gene (rs17060940), which may disrupt pre-mRNA splicing. This disruption may lead to exon skipping or partial/full intron retention in the mature mRNA, altering the protein-coding sequence (Dufner-Almeida et al., 2019). Our results showed that the presence of the mutant (T) allele does not per se confer a risk for severe obesity in our population. However, the mutant genotype TT was found only in our case group (n = 16) with a frequency of 5.1%. According to the 1000 Genomes Project (Phase 3), the frequency of genotype TT varies with the population. The highest frequency is found in Africa (15.3%), followed by South Asia (0.4%) and America (0.3%). This genotype was not observed in Europe and East Asia. In the Online Archive of Brazilian Mutations (ABraOM) database, the frequency of genotype TT was 10.8% (available at http://abraom.ib.usp.br/). Unfortunately, no clinical data is available in this database. Thus, we suggest that this variant should be investigated in other studies using cohorts from different populations to examine the possible influence on obesity susceptibility.
In our analysis, the presence of mutant allele (T) in DBC1 rs17060940 was significantly associated with higher levels of glycated hemoglobin. Previous studies have suggested that DBC1 regulates the metabolic gene network and it is downregulated in cells of patients with T2DM (Sathishkumar et al., 2016; Basu et al., 2020). Briefly, DBC1 acts as a negative regulator of histone deacetylase 3 (HDAC3), which is responsible for controlling inflammation and the metabolism of glucose and insulin. It was also reported that the HDAC3 activity/HDAC3 mRNA levels are positively linked to inflammation, poor glycemic control (higher levels of fasting plasma glucose and glycated hemoglobin), and insulin resistance (Sathishkumar et al., 2016). Based on the literature, we suggest that DBC1 rs17060940 polymorphism may impact the structure of DBC1 pre-mRNA, decreasing the levels of the transcript. This reduction may affect the negative regulation of HDAC3 that impacts glycemic control, consequently, it increases the levels of glycated hemoglobin. This hypothesis is in line with previous studies, which reported that variants in DBC1 were associated with diabetes risk in the United Kingdom Biobank. Furthermore, polymorphisms near this gene were also linked to diabetes susceptibility (Jurgens et al., 2022).
SIRT1 is involved in biological processes of inflammation as well as glucose and fat metabolism by regulating the transcriptional activity of downstream genes (Picard et al., 2004; Bordone et al., 2006; Chaudhary and Pfluger, 2009; Kotas et al., 2013; Moreno-Navarrete et al., 2015). Our study has investigated two variants in SIRT1, which were associated with obesity susceptibility and/or obesity traits (Zillikens et al., 2009; Shimoyama et al., 2011; Kilic et al., 2015b; Higashibata et al., 2016; Casarotto et al., 2019). SIRT1 rs7895833 (A>G) is located in the promoter region, specifically, it lies in a W-box-like element (sequence TTGACT) (Hwang et al., 2008; Tao et al., 2022). In our research, the SIRT1 rs7895833 AG heterozygous genotype increased the risk by 1.93 times for severe obesity, when compared to the homozygous forms (AA and GG) [overdominant model].
In a previous study, the SIRT1 rs7895833 AG genotype carried a high risk of obesity in children from Turkey—as shown in our study. They also suggested that the A allele has a protective effect against obesity (Kilic et al., 2015b). According to a Dutch study, SIRT1 rs7895833 A allele was associated with higher BMI (Zillikens et al., 2009). In Japanese, the A allele increased the risk for obesity in male individuals (Shimoyama et al., 2011). Further, this allele was also a risk factor for overweight women who did not restrict their calorie intake. However, it was not correlated with BMI in women who controlled their calorie intake or practiced physical exercises (Higashibata et al., 2016). Finally, SIRT1 rs7895833 polymorphism was not associated with obesity in Brazilian adults (MENEGUETTE et al., 2016). These different findings may be explained by a study design, sample criteria selection, and genetic background.
In the literature, it has been described that the variants in the promoter region are more common than in coding regions of a gene and may affect its expression (Deihimi et al., 2012). The SIRT1 rs7895833 polymorphism lies on a W-box-like element in the promoter region, which might disturb the gene expression. Based on the literature, SIRT1 negatively controls adipogenesis by repressing genes associated with white adipocyte differentiation and fat accumulation. Moreover, SIRT1 interacts with PPARG to suppress its transcriptional activity, which results in the inhibition of adipogenesis (Picard et al., 2004). In this context, a previous study also observed that lower SIRT1 expression is associated with obesity (Clark et al., 2012). We hypothesized that the SIRT1 rs7895833 polymorphism may affect the promoter activity, decreasing the expression of the SIRT1 gene. Consequently, it may impact SIRT1 negative regulation, increasing the risk for obesity. However, functional studies are necessary to confirm this hypothesis.
Concerning the SIRT1 rs1467568 polymorphism, the presence of a minor allele (G) was associated with lower BAI and HC in our study. Our analyses showed a dose-effect relationship between the polymorphism and these anthropometric traits. This variant located in the intronic region was previously associated with obesity and weight loss resistance in a therapy treatment (Zillikens et al., 2009; Clark et al., 2012; Garaulet et al., 2012). Our findings are in line with previous studies which reported that the SIRT1 rs1467568 G allele has a protective effect against obesity in a Dutch and French population (Zillikens et al., 2009; Clark et al., 2012). Besides these results, we also observed that the SIRT1 rs1467568 AG genotype is a risk factor for UMO in individuals with severe obesity. To our knowledge, no study has investigated this association until now. However, it was reported that ∼10–30% of subjects with obesity have a lower risk for cardiometabolic abnormalities (Blüher, 2020)—similar to our findings. Phenome-wide association study (PheWAS) was used to deeply investigate the effect of SIRT1 rs1467568 on anthropometric, biochemical, and blood pressure in the literature. This variant was associated with BMI, weight, glucose, glycated hemoglobin, HOMA-IR as well as systolic and diastolic blood pressure (https://gwas.mrcieu.ac.uk/phewas/). Thus, we suggest that more studies should investigate this association to understand the genetic background of cardiometabolic abnormalities.
The PPARG plays an important role in adipogenesis, and glucose and lipid metabolism (Spiegelman, 1998; Auwerx, 1999). The PPARG rs1801282 is one of the most studied variants in this gene, which leads to the replacement of alanine with proline at codon 12 [p. (Pro12Ala)]. Previous studies reported an association of this variant with obesity risk and measures of this disease; however, others have reported contradictory results (Masud et al., 2003; Ali et al., 2009; Carlos et al., 2013; Hsiao and Lin, 2015; Hasan et al., 2017; Bakhashab et al., 2020; Dmitrenko et al., 2022). Our study showed that PPARG rs1801282 was not associated with severe obesity risk nor influenced obesity traits variables. Our findings are in line with Bakhashab and coworkers (2020) who reported no association between PPARG rs1801282 and anthropometric/biochemical parameters (BMI, HbA1c, FBS, triglyceride and cholesterol) in individuals from the Saudi population (Bakhashab et al., 2020).
Different findings were observed by Prakash and coworkers (2012), which reported that rs1801282 was associated with obesity risk (dominant model) in the North Indian population (Prakash et al., 2012). Similar results were also observed in two Caucasian populations (Beamer et al., 1998). A previous meta-analysis study observed that the mutant allele (G) was associated with higher BMI in individuals with overweight/obesity. They also found that BMI was 0.07 units higher in individuals carrying this allele (Masud et al., 2003). Ali and colleagues (2009) observed a gender-specific effect of this polymorphism with on obesity risk. They reported that the presence of mutant allele (G) was a risk factor for obesity in a male Tunisian population (Ali et al., 2009). These results are in agreement with Carlos and coworkers (2013), which did not observe an association between PPARG rs1801282 and obesity in Portuguese women (Carlos et al., 2013). However, Hsiao and colleagues (2015) reported that PPARG rs1801282 predicts overweight and higher BMI only in females from the Taiwan population (Hsiao and Lin, 2015). In a Brazilian cohort, it was also found that this polymorphism was associated with overweight and obesity only in female individuals. Despite our different results, they also observed a low frequency of PPARG rs1801282 GG genotype in the case (1.0%) and control group (0.80%) (Castro et al., 2021).
UCP2 has a crucial role in the energy metabolism of cells. UCP2 is a candidate gene for obesity and some common variants have been investigated. One of these variants is the missense polymorphism in exon 4 (rs660339), which changes an alanine to valine in codon 55 [p. (Ala55Val)] (Nicoletti et al., 2017). In a number of genetic studies, the association between UCP2 rs660339 polymorphism and obesity risk has been investigated. However, the results of these studies have been controversial: while some of them reported an association, others were unable to find any relationship between this variant and obesity susceptibility (Kubota et al., 1998; Xiu et al., 2004; Yu et al., 2005; Saraswati et al., 2011; Qian et al., 2013). Previous studies reported that the UCP2 rs660339 AA genotype increased the risk for obesity development, and it was associated with higher BMI or abdominal obesity when compared to subjects with GG and GA genotypes from China and North America (Xiu et al., 2004; Yu et al., 2005). These results are similar to our findings, in which AA genotype was associated with higher BMI. However, such results were not found in a cohort of Japanese and Indonesian individuals (Kubota et al., 1998; Saraswati et al., 2011). Similarly, a meta-analysis study also showed that this polymorphism was not associated with BMI in subjects of Asian and European descent (Qian et al., 2013). Thus, all these findings suggest that the effect of UCP2 rs660339 on obesity susceptibility is dependent on the ethnic group.
ADRB2 is involved in lipid mobilization, especially in adipocyte cells. The missense variant ADRB2 rs1042714 is one of the most common polymorphisms in this gene, leading to the alteration of glutamine to glutamic acid at codon 27 [p. (Gln27Glu)]. Due to the modification of the amino acid sequence in the extracellular N-terminus of the protein, it was suggested that this polymorphism may alter its function (Reihsaus et al., 1993; Zhang et al., 2014). In the literature, the results concerning the contribution of ADRB2 rs1042714 polymorphism toward obesity development (or obesity-traits) are contradictory. Two previous meta-analysis studies involving this polymorphism and obesity highlighted that these different results may be explained due to the divergence in the genetic background. First, Jalba and coworkers (2008) reported that ADRB2 rs1042714 is a risk factor for obesity in Asians, Pacific Islanders, and American Indians, but not in Europeans (Jalba et al., 2008). Later, Zhang and colleagues (2014) conducted a meta-analysis that involved 17 studies (n = 9,995 subjects), in which ADRB2 rs1042714 was associated with increased obesity susceptibility in the heterozygote and dominant model (Zhang et al., 2014). The results of our study showed no significant association between ADRB2 rs1042714 and obesity, which was similar to those observed in Europeans (Jalba et al., 2008). Our results supported previous results of Kortner (Kortner et al., 1999), Galletti (Galletti et al., 2004) and Oberkofler (Oberkofler et al., 2000), which reported no association in a German, Italian and Australian populations—respectively.
Regarding the adipokines and/or inflammatory markers, our results showed that individuals with obesity have higher levels of leptin, PAI-1, and resistin. These results are in accordance with previous reports in the literature. It is well-established obesity alters secretory adipose tissue function in rodents and humans, especially adipokines and pro-inflammatory mediator secretions (Codoñer-Franch and Alonso-Iglesias, 2015; Barnard et al., 2016; Obradovic et al., 2021).
PAI-1 is produced by adipose tissue and is found to increase in individuals with obesity, being suggested as a biochemical marker of this disease. Additionally, PAI-1 is a component of metabolic syndrome and it is the main inhibitor of fibrinolysis, reenforcing to be a risk factor for cardiovascular disease in individuals with obesity (Mertens et al., 2006; Barnard et al., 2016). Resistin is also an important factor in obesity-mediated inflammation and plays a key role as a trigger of metabolic complications and insulin resistance (Benomar and Taouis, 2019). Previous studies showed that resistin is a key hormone linking hypertension and insulin resistance to obesity through activating the Toll-Like Receptor (TLR4) pathway, which can promote the progressive onset of type 2 diabetes (Benomar et al., 2016; Jiang et al., 2016; Benomar and Taouis, 2019). Finally, leptin is a hormone that regulates food intake, total body fat, and proinflammatory immune responses. It is produced and secreted by white adipose tissue, and smaller quantities are detected in brown adipose tissue (MacDougald et al., 1995). Regarding the molecular effect of leptin, this adipokine promotes the expression of several inflammatory cytokines. In a feedback loop, these inflammatory makers increase leptin expression, promoting a low-grade and chronic inflammation (Dubern and Clement, 2012; Obradovic et al., 2021).
Our study has limitations that have to be considered. (1) It was a cross-sectional study: we were not able to eliminate fluctuations in clinical, anthropometric and biochemical parameters. (2) different variables were included as confounding factors; however, we cannot exclude the complete role of environmental variables, which could influence the severe obesity risk. (3) It was not possible to confirm that the studied polymorphisms affect gene expression since we do not have data on mRNA or protein expression.
In conclusion, our data present putative genetic biomarkers for severe obesity and metabolic complications susceptibility. Our findings showed that SIRT1 rs7895833 (AG) increased the risk for severe obesity, and SIRT1 rs1467568 (AG) was associated with UMO development. We also observed that SIRT1 rs1467568, DBC1 rs17060940, and UCP2 rs660339 influenced obesity-related traits in the studied Brazilian population. Therefore, our results indicated that SIRT1 rs7895833 and SIRT1 rs1467568 may be genetic biomarkers for severe obesity and/or unbalanced metabolic status.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving humans were approved by the ethics committee of the Oswaldo Cruz Foundation (CAAE: 09225113.0.0000/Protocol No: 346.634). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
ACPF: Conceptualization, Formal Analysis, Investigation, Methodology, Writing–original draft, Writing–review and editing. ISSA: Methodology, Writing–review and editing, Validation. KCRS: Formal Analysis, Methodology, Writing–review and editing. LP: Methodology, Writing–review and editing. GMA: Methodology, Validation, Writing–review and editing. VMZ: Funding acquisition, Visualization, Writing–review and editing. MCJ: Funding acquisition, Resources, Writing–review and editing. JFNN: Methodology, Writing–review and editing. AC: Methodology, Validation, Writing–review and editing. MLFSJ: Methodology, Writing–review and editing, Formal Analysis, Validation. CMMM: Conceptualization, Investigation, Resources, Writing–review and editing. PHC: Formal Analysis, Writing–review and editing. PTB: Conceptualization, Writing–review and editing, Funding acquisition, Investigation, Resources, Supervision. JRIC: Conceptualization, Funding acquisition, Resources, Supervision, Writing–review and editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Oswaldo Cruz Foundation (Fiocruz, Brazil) and Carlos Chagas Filho Foundation for Research Support in the State of Rio de Janeiro (FAPERJ) [GRANT: E-26/210.663/2021 and E-26/202.291/2019].
Acknowledgments
The authors would like to thank Edson Fernandes and Nereida Proença da Fonseca for their great technical assistance and the “Programa de Desenvolvimento Tecnologico em Insumos para Saude” (PDTIS) platform for multiplex analysis. We are grateful to the patients who kindly agreed to participate in this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2024.1363417/full#supplementary-material
References
Abarca-Gómez, L., Abdeen, Z. A., Hamid, Z. A., Abu-Rmeileh, N. M., Acosta-Cazares, B., Acuin, C., et al. (2017). Worldwide trends in body-mass index, underweight, overweight, and obesity from 1975 to 2016: a pooled analysis of 2416 population-based measurement studies in 128·9 million children, adolescents, and adults. Lancet 390 (10113), 2627–2642. doi:10.1016/S0140-6736(17)32129-3
Affourtit, C., and Brand, M. D. (2008). On the role of uncoupling protein-2 in pancreatic beta cells. Biochim. Biophys. Acta - Bioenerg. 1777 (7), 973–979. doi:10.1016/j.bbabio.2008.03.022
Ali, S. B., Yahia, F. B., Sediri, Y., Kallel, A., Ftouhi, B., Feki, M., et al. (2009). Gender-specific effect of Pro12Ala polymorphism in peroxisome proliferator-activated receptor γ-2 gene on obesity risk and leptin levels in a Tunisian population. Clin. Biochem. 42 (16–17), 1642–1647. doi:10.1016/j.clinbiochem.2009.08.019
Almandoz, J. P., Singh, E., Howell, L. A., Grothe, K., Vlazny, D. T., Smailovic, A., et al. (2013). Spillover of fatty acids during dietary fat storage in type 2 diabetes: relationship to body fat depots and effects of weight loss. Diabetes 62 (6), 1897–1903. doi:10.2337/db12-1407
Astrup, A., Toubro, S., Dalgaard, L. T., Urhammer, S. A., Sørensen, T. I. A., and Pedersen, O. (1999). Impact of the v/v 55 polymorphism of the uncoupling protein 2 gene on 24-h energy expenditure and substrate oxidation. Int. J. Obes. 23 (10), 1030–1034. doi:10.1038/sj.ijo.0801040
Auwerx, J. (1999). PPARgamma, the ultimate thrifty gene. Diabetologia 42 (9), 1033–1049. doi:10.1007/s001250051268
Azzu, V., and Brand, M. D. (2010). The on-off switches of the mitochondrial uncoupling proteins. Trends Biochem. Sci. 35 (5), 298–307. doi:10.1016/j.tibs.2009.11.001
Bakhashab, S., Filimban, N., Altall, R. M., Nassir, R., Qusti, S. Y., Alqahtani, M. H., et al. (2020). The effect sizes of PPARγ rs1801282, FTO rs9939609, and MC4R rs2229616 variants on type 2 diabetes mellitus risk among the western Saudi population: a cross-sectional prospective study. Genes (Basel) 11 (1), 98. doi:10.3390/genes11010098
Barnard, S. A., Pieters, M., and De Lange, Z. (2016). The contribution of different adipose tissue depots to plasma plasminogen activator inhibitor-1 (PAI-1) levels. Blood Rev. 30 (6), 421–429. doi:10.1016/j.blre.2016.05.002
Basu, S., Barad, M., Yadav, D., Nandy, A., Mukherjee, B., Sarkar, J., et al. (2020). DBC1, p300, HDAC3, and Siah1 coordinately regulate ELL stability and function for expression of its target genes. Proc. Natl. Acad. Sci. 117 (12), 6509–6520. doi:10.1073/pnas.1912375117
Beamer, B. A., Yen, C.-J., Andersen, R. E., Muller, D., Elahi, D., Cheskin, L. J., et al. (1998). Association of the Pro12Ala variant in the peroxisome proliferator-activated receptor-gamma2 gene with obesity in two Caucasian populations. Diabetes 47 (11), 1806–1808. doi:10.2337/diabetes.47.11.1806
Benomar, Y., Amine, H., Crépin, D., Al Rifai, S., Riffault, L., Gertler, A., et al. (2016). Central resistin/TLR4 impairs adiponectin signaling, contributing to insulin and FGF21 resistance. Diabetes 65 (4), 913–926. doi:10.2337/db15-1029
Benomar, Y., and Taouis, M. (2019). Molecular mechanisms underlying obesity-induced hypothalamic inflammation and insulin resistance: pivotal role of resistin/TLR4 pathways. Front. Endocrinol. (Lausanne) 10, 140. doi:10.3389/fendo.2019.00140
Bergman, R. N., Stefanovski, D., Buchanan, T. A., Sumner, A. E., Reynolds, J. C., Sebring, N. G., et al. (2011). A better index of body adiposity. Obesity 19 (5), 1083–1089. doi:10.1038/oby.2011.38
Blüher, M. (2020). Metabolically healthy obesity. Endocr. Rev. 41 (3), bnaa004. doi:10.1210/endrev/bnaa004
Bordone, L., Motta, M. C., Picard, F., Robinson, A., Jhala, U. S., Apfeld, J., et al. (2006). Sirt1 regulates insulin secretion by repressing UCP2 in pancreatic beta cells. PLoS Biol. 4 (2), e31. doi:10.1371/journal.pbio.0040031
Carlos, F. F., Silva-Nunes, J., Flores, O., Brito, M., Doria, G., Veiga, L., et al. (2013). Association of FTO and PPARG polymorphisms with obesity in Portuguese women. Diabetes, Metab. Syndr. Obes. targets Ther. 6, 241–245. doi:10.2147/DMSO.S45779
Casarotto, A. A. F., Galera, B. B., Sumiyoshi, L. M., and Floôr, T. M. (2019). Polymorphism rs7895833 in the SIRT1 gene and its association with dyslipidaemia in the elderly. Rev. Esp. Geriatr. Gerontol. 54 (4), 214–219. doi:10.1016/j.regg.2019.01.008
Castro, G. V., Latorre, A. F. S., Korndorfer, F. P., de Carlos Back, L. K., and Lofgren, S. E. (2021). The impact of variants in four genes: MC4R, FTO, PPARG and PPARGC1A in overweight and obesity in a large sample of the Brazilian population. Biochem. Genet. 59 (6), 1666–1679. doi:10.1007/s10528-021-10079-2
Chaudhary, N., and Pfluger, P. T. (2009). Metabolic benefits from Sirt1 and Sirt1 activators. Curr. Opin. Clin. Nutr. Metab. Care 12 (4), 431–437. doi:10.1097/MCO.0b013e32832cdaae
Clark, S. J., Falchi, M., Olsson, B., Jacobson, P., Cauchi, S., Balkau, B., et al. (2012). Association of sirtuin 1 (SIRT1) gene SNPs and transcript expression levels with severe obesity. Obesity 20 (1), 178–185. doi:10.1038/oby.2011.200
Codoñer-Franch, P., and Alonso-Iglesias, E. (2015). Resistin: insulin resistance to malignancy. Clin. Chim. acta. 438, 46–54. doi:10.1016/j.cca.2014.07.043
da Fonseca, A. C. P., Abreu, G. M., Zembrzuski, V. M., Campos Junior, M., Carneiro, J. R. I., Neto, J. F. N., et al. (2019). The association of the fat mass and obesity-associated gene (FTO) rs9939609 polymorphism and the severe obesity in a Brazilian population. Diabetes, Metab. Syndr. Obes. Targets Ther. 12, 667–684. doi:10.2147/DMSO.S199542
da Fonseca, A. C. P., Mastronardi, C., Johar, A., Arcos-Burgos, M., and Paz-Filho, G. (2017). Genetics of non-syndromic childhood obesity and the use of high-throughput DNA sequencing technologies. J. Diabetes Complicat. 31 (10), 1549–1561. doi:10.1016/j.jdiacomp.2017.04.026
Deeb, S. S., Fajas, L., Nemoto, M., Pihlajamäki, J., Mykkänen, L., Kuusisto, J., et al. (1998). A Pro12Ala substitution in PPARgamma2 associated with decreased receptor activity, lower body mass index and improved insulin sensitivity. Nat. Genet. 20 (3), 284–287. doi:10.1038/3099
Deihimi, T., Niazi, A., Ebrahimi, M., Kajbaf, K., Fanaee, S., Bakhtiarizadeh, M. R., et al. (2012). Finding the undiscovered roles of genes: an approach using mutual ranking of coexpressed genes and promoter architecture-case study: dual roles of thaumatin like proteins in biotic and abiotic stresses. Springerplus 1 (1), 30–10. doi:10.1186/2193-1801-1-30
Dmitrenko, O. P., Karpova, N. S., and Nurbekov, M. K. (2022). ACE, PPARG, SIRT1 gene polymorphisms but not PPARGC1A polymorphism are risk factors for gestational diabetes in the Russian population. J. Genet. Genomic Sci. 7 (035), 2. doi:10.24966/GGS-2485/100035
Dubern, B., and Clement, K. (2012). Leptin and leptin receptor-related monogenic obesity. Biochimie 94 (10), 2111–2115. doi:10.1016/j.biochi.2012.05.010
Dufner-Almeida, L. G., do Carmo, R. T., Masotti, C., and Haddad, L. A. (2019). in Chapter Two - understanding human DNA variants affecting pre-mRNA splicing in the NGS era (United States: Academic Press), 39–90.
Eckel, R. H., Alberti, KGMM, Grundy, S. M., and Zimmet, P. Z. (2010). The metabolic syndrome. Lancet 375 (9710), 181–183. doi:10.1016/S0140-6736(09)61794-3
Escande, C., Nin, V., Pirtskhalava, T., Chini, C. C. S., Tchkonia, T., Kirkland, J. L., et al. (2015). Deleted in breast cancer 1 limits adipose tissue fat accumulation and plays a key role in the development of metabolic syndrome phenotype. Diabetes 64 (1), 12–22. doi:10.2337/db14-0192
Galletti, F., Iacone, R., Ragone, E., Russo, O., Valle, E. D., Siani, A., et al. (2004). Lack of association between polymorphism in the beta2-adrenergic receptor gene, hypertension, and obesity in the Olivetti heart study. Am. J. Hypertens. 17 (8), 718–720. doi:10.1016/j.amjhyper.2004.04.012
Garaulet, M., Esteban Tardido, A., Lee, Y. C., Smith, C. E., Parnell, L. D., and Ordovas, J. M. (2012). SIRT1 and CLOCK 3111T> C combined genotype is associated with evening preference and weight loss resistance in a behavioral therapy treatment for obesity. Int. J. Obes. 36 (11), 1436–1441. doi:10.1038/ijo.2011.270
González, J. R., Armengol, L., Solé, X., Guinó, E., Mercader, J. M., Estivill, X., et al. (2007). SNPassoc: an R package to perform whole genome association studies. Bioinformatics 23 (5), 644–645. doi:10.1093/bioinformatics/btm025
Grundy, S. M., Brewer, H. B., Cleeman, J. I., Smith, S. C., Lenfant, C., American Heart Association, , et al. (2004). Definition of metabolic syndrome: report of the national heart, lung, and blood Institute/American heart association conference on scientific issues related to definition. Circulation 109 (3), 433–438. doi:10.1161/01.CIR.0000111245.75752.C6
Hasan, N. S., Kamel, S. A., Hamed, M., Awadallah, E., Rahman, A. H. A., Musa, N. I., et al. (2017). Peroxisome proliferator-activated receptor-γ polymorphism (rs1801282) is associated with obesity in Egyptian patients with coronary artery disease and type 2 diabetes mellitus. J. Genet. Eng. Biotechnol. 15 (2), 409–414. doi:10.1016/j.jgeb.2017.08.002
Higashibata, T., Wakai, K., Naito, M., Morita, E., Hishida, A., Hamajima, N., et al. (2016). Effects of self-reported calorie restriction on correlations between SIRT1 polymorphisms and body mass index and long-term weight change. Gene 594 (1), 16–22. doi:10.1016/j.gene.2016.08.051
Hsiao, T.-J., and Lin, E. (2015). The Pro12Ala polymorphism in the peroxisome proliferator-activated receptor gamma (PPARG) gene in relation to obesity and metabolic phenotypes in a Taiwanese population. Endocrine 48 (3), 786–793. doi:10.1007/s12020-014-0407-7
Hwang, S.-H., Lee, I. A., Yie, S. W., and Hwang, D.-J. (2008). Identification of an OsPR10a promoter region responsive to salicylic acid. Planta 227 (5), 1141–1150. doi:10.1007/s00425-007-0687-8
Jalba, M. S., Rhoads, G. G., and Demissie, K. (2008). Association of codon 16 and codon 27 beta 2-adrenergic receptor gene polymorphisms with obesity: a meta-analysis. Obesity 16 (9), 2096–2106. doi:10.1038/oby.2008.327
Jiang, Y., Lu, L., Hu, Y., Li, Q., An, C., Yu, X., et al. (2016). Resistin induces hypertension and insulin resistance in mice via a TLR4-dependent pathway. Sci. Rep. 6 (1), 22193. doi:10.1038/srep22193
Jurgens, S. J., Choi, S. H., Morrill, V. N., Chaffin, M., Pirruccello, J. P., Halford, J. L., et al. (2022). Analysis of rare genetic variation underlying cardiometabolic diseases and traits among 200,000 individuals in the UK Biobank. Nat. Genet. 54 (3), 240–250. doi:10.1038/s41588-021-01011-w
Kilic, U., Gok, O., Elibol-Can, B., Ozgen, I. T., Erenberk, U., Uysal, O., et al. (2015b). SIRT1 gene variants are related to risk of childhood obesity. Eur. J. Pediatr. 174 (4), 473–479. doi:10.1007/s00431-014-2424-1
Kilic, U., Gok, O., Erenberk, U., Dundaroz, M. R., Torun, E., Kucukardali, Y., et al. (2015a). A remarkable age-related increase in SIRT1 protein expression against oxidative stress in elderly: SIRT1 gene variants and longevity in human. PLoS One 10 (3), e0117954. doi:10.1371/journal.pone.0117954
Kortner, B., Wolf, A., Wendt, D., Beisiegel, U., and Evans, D. (1999). Lack of association between a human beta-2 adrenoceptor gene polymorphism (gln27glu) and morbid obesity. Int. J. Obes. 23 (10), 1099–1100. doi:10.1038/sj.ijo.0801063
Kotas, M. E., Gorecki, M. C., and Gillum, M. P. (2013). Sirtuin-1 is a nutrient-dependent modulator of inflammation. Adipocyte 2 (2), 113–118. doi:10.4161/adip.23437
Kubota, T., Mori, H., Tamori, Y., Okazawa, H., Fukuda, T., Miki, M., et al. (1998). Molecular screening of uncoupling protein 2 gene in patients with noninsulin-dependent diabetes mellitus or obesity. J. Clin. Endocrinol. Metab. 83 (8), 2800–2804. doi:10.1210/jcem.83.8.4994
Kurylowicz, A., Jonas, M., Lisik, W., Jonas, M., Wicik, Z. A., Wierzbicki, Z., et al. (2015). Obesity is associated with a decrease in expression but not with the hypermethylation of thermogenesis-related genes in adipose tissues. J. Transl. Med. 13 (1), 31. doi:10.1186/s12967-015-0395-2
Lafontan, M., Sengenes, C., Galitzky, J., Berlan, M., De Glisezinski, I., Crampes, F., et al. (2000). Recent developments on lipolysis regulation in humans and discovery of a new lipolytic pathway. Int. J. Obes. 24 (4), S47–S52. doi:10.1038/sj.ijo.0801505
Lê, K.-A., Mahurkar, S., Alderete, T. L., Hasson, R. E., Adam, T. C., Kim, J. S., et al. (2011). Subcutaneous adipose tissue macrophage infiltration is associated with hepatic and visceral fat deposition, hyperinsulinemia, and stimulation of NF-κB stress pathway. Diabetes 60 (11), 2802–2809. doi:10.2337/db10-1263
Lee, M., Choi, S., Lee, Y., and Oh, H. (2016). The gender association of the SIRT1 rs7895833 polymorphism with pediatric obesity: a 3-year panel study. Lifestyle Genom. 9 (5–6), 265–75.
Listenberger, L. L., Han, X., Lewis, S. E., Cases, S., Farese, Jr R. V., Ory, D. S., et al. (2003). Triglyceride accumulation protects against fatty acid-induced lipotoxicity. Proc. Natl. Acad. Sci. 100 (6), 3077–3082. doi:10.1073/pnas.0630588100
Luan, J., Browne, P. O., Harding, A.-H., Halsall, D. J., O’Rahilly, S., Chatterjee, V. K. K., et al. (2001). Evidence for gene-nutrient interaction at the PPARgamma locus. Diabetes 50 (3), 686–689. doi:10.2337/diabetes.50.3.686
MacDougald, O. A., Hwang, C.-S., Fan, H., and Lane, M. D. (1995). Regulated expression of the obese gene product (leptin) in white adipose tissue and 3T3-L1 adipocytes. Proc. Natl. Acad. Sci. 92 (20), 9034–9037. doi:10.1073/pnas.92.20.9034
Masud, S., and Ye, S.SAS Group (2003). Effect of the peroxisome proliferator activated receptor-γ gene Pro12Ala variant on body mass index: a meta-analysis. J. Med. Genet. 40 (10), 773–780. doi:10.1136/jmg.40.10.773
Meneguette, M. V. de O., Oliveira, C. A. de, Lima, M. H. de M., Pina, K. N., and Amaral, M. E. C. do (2016). Polymorphism in the SIRT1 gene and parameters of metabolic syndrome in a sample of the adult Brazilian population. Rev. Nutr. 29, 1–10. doi:10.1590/1678-98652016000100001
Mertens, I., Verrijken, A., Michiels, J. J., Van der Planken, M., Ruige, J. B., and Van Gaal, L. F. (2006). Among inflammation and coagulation markers, PAI-1 is a true component of the metabolic syndrome. Int. J. Obes. 30 (8), 1308–1314. doi:10.1038/sj.ijo.0803189
Moreno-Navarrete, J. M., Moreno, M., Vidal, M., Ortega, F., Ricart, W., and Fernández-Real, J. M. (2015). DBC1 is involved in adipocyte inflammation and is a possible marker of human adipose tissue senescence. Obesity 23 (3), 519–522. doi:10.1002/oby.20999
Nicoletti, C. F., de Oliveira, APRP, Brochado, M. J. F., Pinhel, M. A. S., de Oliveira, B. A. P., Marchini, J. S., et al. (2017). The Ala55Val and-866G> A polymorphisms of the UCP2 gene could be biomarkers for weight loss in patients who had Roux-en-Y gastric bypass. Nutrition 33, 326–330. doi:10.1016/j.nut.2016.07.020
Oberkofler, H., Esterbauer, H., Hell, E., Krempler, F., and Patsch, W. (2000). The Gln27Glu polymorphism in the beta2-adrenergic receptor gene is not associated with morbid obesity in Austrian women. Int. J. Obes. 24 (3), 388–390. doi:10.1038/sj.ijo.0801180
Obradovic, M., Sudar-Milovanovic, E., Soskic, S., Essack, M., Arya, S., Stewart, A. J., et al. (2021). Leptin and obesity: role and clinical implication. Front. Endocrinol. (Lausanne) 12, 585887. doi:10.3389/fendo.2021.585887
Oomen, J. M., van Rossum, C. T. M., Hoebee, B., Saris, W. H. M., and van Baak, M. A. (2005). beta2-adrenergic receptor polymorphisms and salbutamol-stimulated energy expenditure. J. Clin. Endocrinol. Metab. 90 (4), 2301–2307. doi:10.1210/jc.2004-1356
Pecqueur, C., Alves-Guerra, M.-C., Gelly, C., Lévi-Meyrueis, C., Couplan, E., Collins, S., et al. (2001). Uncoupling protein 2, in vivo distribution, induction upon oxidative stress, and evidence for translational regulation. J. Biol. Chem. 276 (12), 8705–8712. doi:10.1074/jbc.M006938200
Picard, F., Kurtev, M., Chung, N., Topark-Ngarm, A., Senawong, T., Machado de Oliveira, R., et al. (2004). Sirt1 promotes fat mobilization in white adipocytes by repressing PPAR-gamma. Nature 429 (6993), 771–776. doi:10.1038/nature02583
Prakash, J., Srivastava, N., Awasthi, S., Agarwal, C., Natu, S., Rajpal, N., et al. (2012). Association of PPAR-γ gene polymorphisms with obesity and obesity-associated phenotypes in north indian population. Am. J. Hum. Biol. 24 (4), 454–459. doi:10.1002/ajhb.22245
Qian, L., Xu, K., Xu, X., Gu, R., Liu, X., Shan, S., et al. (2013). UCP2-866G/A, Ala55Val and UCP3-55C/T polymorphisms in association with obesity susceptibility—a meta-analysis study. PLoS One 8 (4), e58939. doi:10.1371/journal.pone.0058939
Ramkaran, P., Khan, S., Moodley, D., Chuturgoon, A. A., and Phulukdaree, A. (2016). Sirtuin 1 rs1467568 and rs7895833 in South African Indians with early-onset coronary artery disease. Cardiovasc. J. Afr. 27 (4), 213–7.
Reihsaus, E., Innis, M., MacIntyre, N., and Liggett, S. B. (1993). Mutations in the gene encoding for the beta 2-adrenergic receptor in normal and asthmatic subjects. Am. J. Respir. Cell Mol. Biol. 8 (3), 334–339. doi:10.1165/ajrcmb/8.3.334
Robitaille, J., Després, J., Perusse, L., and Vohl, M. (2003). The PPAR-gamma P12A polymorphism modulates the relationship between dietary fat intake and components of the metabolic syndrome: results from the Québec Family Study. Clin. Genet. 63 (2), 109–16.
Rocha, R. M., Barra, G. B., Rosa, ÉCCC, Garcia, É. C., Amato, A. A., and Azevedo, M. F. (2015). Prevalence of the rs1801282 single nucleotide polymorphism of the PPARG gene in patients with metabolic syndrome. Arch. Endocrinol. Metab. 59, 297–302. doi:10.1590/2359-3997000000086
Saraswati, M. R., Suastika, K., Malik, S. G., and Sudoyo, H. (2011). The uncoupling protein 2 Ala55val polymorphism is associated with diabetes mellitus in a Balinese population. J. ASEAN Fed. Endocr. Soc. 26 (1), 39–43. doi:10.15605/jafes.026.01.08
Sathishkumar, C., Prabu, P., Balakumar, M., Lenin, R., Prabhu, D., Anjana, R. M., et al. (2016). Augmentation of histone deacetylase 3 (HDAC3) epigenetic signature at the interface of proinflammation and insulin resistance in patients with type 2 diabetes. Clin. Epigenetics 8 (1), 125–212. doi:10.1186/s13148-016-0293-3
Shimoyama, Y., Suzuki, K., Hamajima, N., and Niwa, T. (2011). Sirtuin 1 gene polymorphisms are associated with body fat and blood pressure in Japanese. Transl. Res. 157 (6), 339–347. doi:10.1016/j.trsl.2011.02.004
Silva, S. de S., Leite, N., Furtado-Alle, L., de Souza, R. L. R., Corazza, P. R. P., Tradiotto, M. C., et al. (2022). ADRB2 gene influences responsiveness to physical exercise programs: a longitudinal study applied to overweight or obese Brazilian children and adolescents. Gene. 820, 146296.
Souza, B. M. de, Assmann, T. S., Kliemann, L. M., Gross, J. L., Canani, L. H., and Crispim, D. (2011). The role of uncoupling protein 2 (UCP2) on the development of type 2 diabetes mellitus and its chronic complications. Arq. Bras. Endocrinol. Metabol. 55, 239–248. doi:10.1590/s0004-27302011000400001
Spiegelman, B. M. (1998). PPAR-gamma: adipogenic regulator and thiazolidinedione receptor. Diabetes 47 (4), 507–514. doi:10.2337/diabetes.47.4.507
Tao, T. T., Lin, X. H., Tang, S. J., Gui, W. W., Zhu, W. F., and Li, H. (2022). Association of genetic variants in the Sirt1 and Nrf2 genes with the risk of metabolic syndrome in a Chinese Han population. BMC Endocr. Disord. 22 (1), 84–88. doi:10.1186/s12902-022-00965-0
Unger, R. H., and Scherer, P. E. (2010). Gluttony, sloth and the metabolic syndrome: a roadmap to lipotoxicity. Trends Endocrinol. Metab. 21 (6), 345–352. doi:10.1016/j.tem.2010.01.009
Wang, M.-Y., Grayburn, P., Chen, S., Ravazzola, M., Orci, L., and Unger, R. H. (2008). Adipogenic capacity and the susceptibility to type 2 diabetes and metabolic syndrome. Proc. Natl. Acad. Sci. 105 (16), 6139–6144. doi:10.1073/pnas.0801981105
Xie, W., He, R., Zhang, J., He, Y., Wan, Z., Zhou, C., et al. (2019). β‑blockers inhibit the viability of breast cancer cells by regulating the ERK/COX‑2 signaling pathway and the drug response is affected by ADRB2 single‑nucleotide polymorphisms. Oncol. Rep. 41 (1), 341–350. doi:10.3892/or.2018.6830
Xiu, L. L., Weng, J. P., Sui, Y., Wang, J., Yan, J. H., and Huang, Z. M. (2004). Common variants in beta 3-adrenergic-receptor and uncoupling protein-2 genes are associated with type 2 diabetes and obesity. Zhonghua Yi Xue Za Zhi 84 (5), 375–379.
Yu, X., Jacobs, Jr D. R., Schreiner, P. J., Gross, M. D., Steffes, M. W., and Fornage, M. (2005). The uncoupling protein 2 Ala55Val polymorphism is associated with diabetes mellitus: the CARDIA study. Clin. Chem. 51 (8), 1451–1456. doi:10.1373/clinchem.2004.044859
Zhang, H., Wu, J., and Yu, L. (2014). Association of Gln27Glu and Arg16Gly polymorphisms in Beta2-adrenergic receptor gene with obesity susceptibility: a meta-analysis. PLoS One 9 (6), e100489. doi:10.1371/journal.pone.0100489
Zhao, X., Gang, X., He, G., Li, Z., Lv, Y., Han, Q., et al. (2020). Obesity increases the severity and mortality of influenza and COVID-19: a systematic review and meta-analysis. Front. Endocrinol. (Lausanne) 11, 595109. doi:10.3389/fendo.2020.595109
Keywords: DBC1, SIRT1, biomarkers, severe obesity, metabolic complications
Citation: da Fonseca ACP, Assis ISdS, Salum KCR, Palhinha L, Abreu GdM, Zembrzuski VM, Campos Junior M, Nogueira-Neto JF, Cambraia A, Souza Junior MLF, Maya-Monteiro CM, Cabello PH, Bozza PT and Carneiro JRI (2024) Genetic variants in DBC1, SIRT1, UCP2 and ADRB2 as potential biomarkers for severe obesity and metabolic complications. Front. Genet. 15:1363417. doi: 10.3389/fgene.2024.1363417
Received: 04 January 2024; Accepted: 01 May 2024;
Published: 22 May 2024.
Edited by:
Jing Hua Zhao, University of Cambridge, United KingdomReviewed by:
Ivana Joksic, Gynecology and Obstetrics Clinic Narodni front, SerbiaJosé María Moreno-Navarrete, CIBER Fisiopatología Obesidad y Nutrición (CIBEROBN), Spain
Copyright © 2024 da Fonseca, Assis, Salum, Palhinha, Abreu, Zembrzuski, Campos Junior, Nogueira-Neto, Cambraia, Souza Junior, Maya-Monteiro, Cabello, Bozza and Carneiro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana Carolina Proença da Fonseca, ana.proenca@ioc.fiocruz.br, ana_carol_pf2@hotmail.com