 Xavier C. Monger1,2,3,4
Xavier C. Monger1,2,3,4 Linda Saucier1,3,4
Linda Saucier1,3,4 Frédéric Guay1
Frédéric Guay1 Annie Turcotte5Joanie Lemieux6
Annie Turcotte5Joanie Lemieux6 Eric Pouliot7
Eric Pouliot7 Sylvain Fournaise7
Sylvain Fournaise7 Antony T. Vincent1,2,3,4*
Antony T. Vincent1,2,3,4*- 1Département des Sciences Animales, Université Laval, Québec, QC, Canada
- 2Institut de Biologie Intégrative et des Systèmes, Université Laval, Québec, QC, Canada
- 3Institut sur La Nutrition et Les Aliments Fonctionnels, Université Laval, Québec, QC, Canada
- 4Centre de Recherche en Infectiologie Porcine et Avicole, Faculté de Médecine Vétérinaire, Université de Montréal, Saint-Hyacinthe, QC, Canada
- 5Département de Biologie, Microbiologie, Université de Sherbrooke, Sherbrooke, QC, Canada
- 6Département de Biochimie, Microbiologie et Bio-informatique, Université Laval, Québec, QC, Canada
- 7Olymel S.E.C./L.P., Boucherville, QC, Canada
Introduction: To consider the growing health issues caused by antibiotic resistance from a “one health” perspective, the contribution of meat production needs to be addressed. While antibiotic resistance is naturally present in microbial communities, the treatment of farm animals with antibiotics causes an increase in antibiotic resistance genes (ARG) in the gut microbiome. Pigs are among the most prevalent animals in agriculture; therefore, reducing the prevalence of antibiotic-resistant bacteria in the pig gut microbiome could reduce the spread of antibiotic resistance. Probiotics are often studied as a way to modulate the microbiome and are, therefore, an interesting way to potentially decrease antibiotic resistance.
Methods: To assess the efficacy of a probiotic to reduce the prevalence of ARGs in the pig microbiome, six pigs received either treatment with antibiotics (tylvalosin), probiotics (Pediococcus acidilactici MA18/5M; Biopower® PA), or a combination of both. Their faeces and ileal digesta were collected and DNA was extracted for whole genome shotgun sequencing. The reads were compared with taxonomy and ARG databases to identify the taxa and resistance genes in the samples.
Results: The results showed that the ARG profiles in the faeces of the antibiotic and combination treatments were similar, and both were different from the profiles of the probiotic treatment (p < 0.05). The effects of the treatments were different in the digesta and faeces. Many macrolide resistance genes were detected in a higher proportion in the microbiome of the pigs treated with antibiotics or the combination of probiotics and antibiotics. Resistance-carrying conjugative plasmids and horizontal transfer genes were also amplified in faeces samples for the antibiotic and combined treatments. There was no effect of treatment on the short chain fatty acid content in the digesta or the faeces.
Conclusion: There is no positive effect of adding probiotics to an antibiotic treatment when these treatments are administered simultaneously.
1 Introduction
Antibiotic resistance has been a major health concern worldwide for many years, and yet it is still a growing problem in human and animal health (WHO, 2020). While resistance to antibiotics predates their discovery, the selective pressure from antibiotic use, notably in medicine and agriculture, is contributing to the rise of resistant or multi-resistant bacteria (Larkin, 2023). This phenomenon is exacerbated by the capacity of bacteria to evolve rapidly and exchange genetic material between species (Khan and Rao, 2019; Qu and Chen, 2022). When resistance is acquired by pathogenic bacteria, it threatens the efficacy of antibiotic treatments against the diseases caused by the bacteria (Pan et al., 2022).
Antibiotics began to be used in agriculture shortly after their discovery, first to treat and prevent disease in animals and then as growth promoters; however, the latter purpose is no longer socially accepted and no longer used in many countries (OIE, 2018). With the growing awareness of antibiotic use on the selection of resistance, there is an increasing will to further reduce antibiotic use; however, there is still a need to treat sick animals. Animals are particularly vulnerable to infections during the post-weaning stage, and antibiotics are still commonly used to prevent mortality during this period (Canibe et al., 2022). The frequency of treatments and the large number of animals being raised makes pig production particularly interesting to investigate when it comes to the agricultural use of antibiotics (Monger et al., 2021).
Pig farms can contribute to the spread of antimicrobial resistance in many ways. The selection of resistant bacteria and the horizontal transfer of resistance genes originates in the gut microbiota of the animals after they are treated with antibiotics, with this effect being even stronger after oral treatment than after injection (Ricker et al., 2020). Microorganisms can be transferred to the environment through air, manure, wastewater, and occasionally through the meat (Monger et al., 2021).
Many alternatives to antibiotics have been explored in pig rearing, including pre- and probiotics and essential oils (Lopez-Galvez et al., 2021; Pereira et al., 2022). Probiotics are often studied to improve microbiome health in agriculture, and show potential to not only reduce the use of antibiotics in disease prevention but also as a growth promoter (Joysowal et al., 2021; Pereira et al., 2022; Pezsa et al., 2022; Guevarra et al., 2023). Although their effects on growth, feed efficiency, and resistance to infections has been studied to some extent, the mechanisms by which they achieve results and their complex interactions with the animal microbiota are still not fully understood. The interactions between probiotics and antibiotics are also not well understood, although they can reduce the population of pathogenic bacteria, by competition (Shin et al., 2019) and production of bacteriocin (Kim et al., 2023). Whether or not this effect could reduce the acquisition of resistance to antibiotics in pathogens is still unknown.
The effect of interaction between antibiotics, probiotics, and the pig gut microbiome on antibiotics resistance was investigated in this study. To accomplish this, pigs were administered probiotics, antibiotics, and a combination of both. Their faeces and digesta were collected and the DNA from those samples was extracted and sequenced. The sequences were then analyzed to give insight into how the gut microbiome of the pigs was affected by antibiotic or probiotic treatment at the genomic level.
2 Materials and methods
2.1 Animal housing and care
Animal housing conditions were described in a previously published paper (Monger et al., 2022). The experimental design was approved by the Animal Protection Committee of Université Laval prior to the experiments (2019057–1). Briefly, six Yorkshire-Landrace male pigs at 55 days of age were transported to the Université Laval animal research facility. At 66 days, they underwent surgery to implant an ileal-T canula, as previously described (Wubben et al., 2001). They were allowed a 11-day recovery period before the beginning of the treatments, as it was demonstrated that inflammation at the cannulation site was negligible 7 days after the surgery (Zhang et al., 2014). The six pigs received three different treatments: macrolide antibiotic (tylvalosin TYL; 250 g/ton of feed; Aivlosin® (17% tylvalosin), ECO Animal Health Princeton, NJ, USA), probiotic Pediococcus acidilactici MA18/5M (PA; 108 CFUs/day; Biopower® PA, Lallemand Animal Nutrition, Milwaukee, WI, USA), and a combination of both antibiotic and probiotic treatment at the same concentrations as the individual treatments. Each pig received the three treatments in a different order, and a period of recovery was given between each treatment with a control diet not supplemented, according to a crossover design (Figure 1). The periods of treatment and recovery were 3 weeks each.
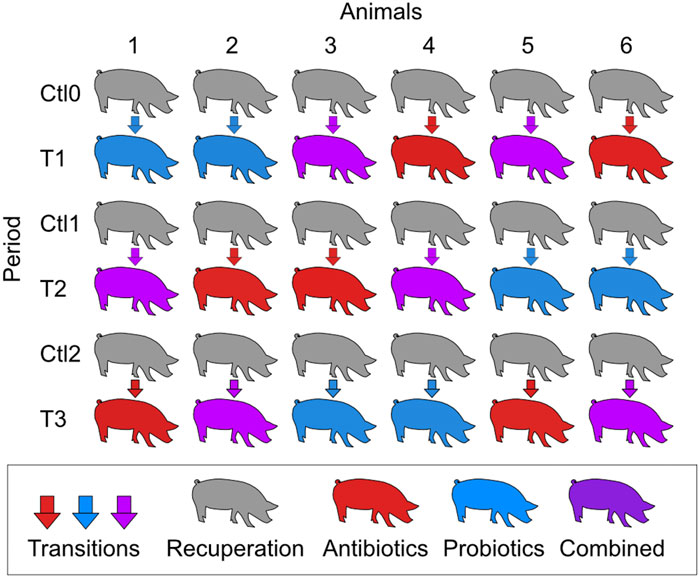
Figure 1. Schematic representation of the crossover experimental design. T1, T2, and T3 refer to the treatment periods. The treatments were a macrolide antibiotic (tylvalosin; 250 g/ton of feed), a commercial probiotic (Pediococcus acidilactici MA18/5M, 108 CFUs/day), and a combination of both treatments at the same concentrations as the individual treatments. During the control periods (Ctl0, Ctl1, and Ctl2), the animals were fed the same diet without treatment. Each period lasted 3 weeks.
2.2 Sample collection and processing
Digesta and faeces from each pig were collected at the end of each treatment period for a total of 36 faeces samples and 35 digesta samples. The digesta from pig number two at the Ctl2 period could not be collected as there was no digesta at the time of sampling. All the samples were stabilized using the PERFORMAbiome•GUT | PB-200 sampling kit according to the manufacturer’s instructions (DNAgenotek, Ottawa, Ontario, Canada) and as suggested elsewhere (Monger et al., 2022).
The DNA was extracted from the samples with the QIAamp PowerFecal Pro DNA Kit (QIAGEN, Toronto, Canada) using 250 µL of stabilized faeces or digesta. The DNA was quantified using the PicoGreen kit (Invitrogen, Waltham, Massachusetts, USA). Integrity of the DNA was assessed by electrophoresis on an agarose gel.
Short chain fatty acids (SCFAs) were quantified as previously described (Laforge et al., 2023). Briefly, samples were stored at −80°C, then thawed and homogenized in water. The homogenized samples were centrifugated and SCFAs were extracted by liquid-liquid extraction. The SCFAs were subjected to gas chromatography coupled with a flame ionization detector (GC-FID Shimadzu, Kyoto, Japan) following the protocol published by Roussel et al. (2022).
2.3 Sequencing and data analysis
Shotgun sequencing libraries were generated from 50 ng of gDNA using the NEBNext Ultra II DNA Library Prep Kit for Illumina (New England BioLabs, Whitby, Canada) as per the manufacturer’s recommendations. Sequencing was done on an Illumina NovaSeq 6000 by the Genome Quebec Centre of Expertise and Services (Montréal, Canada). Base calling was performed with Illumina’s Real Time Analysis software version 3.4.4. A minimum of 70 million paired reads was generated for each of the samples. The optical duplicates in the reads were removed with the clumpify tool from bbmap version 38.96 (Bushnell, 2014). The reads were then filtered using fastp version 0.23.1 (Chen S. et al., 2018), and the sequences were mapped on pig genome sequences using bowtie2 version 2.4.4 (Langmead and Salzberg, 2012) and samtools version 1.17 (Li et al., 2009) to keep only those of the microbiome. The reference genome used was Sscrofa11.1 (RefSeq: GCF_000003025.6). The number of reads at each step is presented in Supplementary Table S1. The whole genome shotgun sequencing data set was deposited in the NCBI Sequence Read Archive database under the BioProject ID PRJNA1049315.
The reads were then analyzed and coassembled with the SqueezeMeta pipeline, version 1.5.1, using default parameters (Tamames and Puente-Sanchez, 2019). Only contigs with a length greater than or equal to 200 bp were retained for further analysis. The SQMtools R package (Puente-Sanchez et al., 2020) was used to import the data in R. The vegan R package (Dixon, 2003) was used for alpha diversity analysis, and the phyloseq (McMurdie and Holmes, 2013) and microbial (Kai, 2021) packages were used for beta diversity and linear discriminant analysis (LDA) analysis, respectively. The cutoffs for LDA were a score greater than or equal to two and an adjusted p-value less than 0.05. The pvclust package was used for hierarchical clustering of the data with 10,000 bootstrap replications (Suzuki and Shimodaira, 2006).
The assembly was screened for plasmidic contigs using PlasForest version 1.4 (Pradier et al., 2021). The plasmid sequences were reconstructed from contigs using the MOB-suite version 3.1.4 (Robertson and Nash, 2018). ARGs were identified by mapping the reads against the Comprehensive Antibiotic Resistance Database (CARD) (Alcock et al., 2020) using MetaProtMiner with a minimum similarity of 90% and a query cover of 80% (Galiot et al., 2023).
The 16S rRNA gene was sequenced and analyzed as described elsewhere (Laforge et al., 2023). Briefly, the v3–v4 region of the 16S DNA was sequenced at the Plateforme d’analyse génomique of the Institut de Biologie Intégrative et des Systèmes (Université Laval, Quebec City, Canada). The sequences were pretreated and taxonomically assigned with the R package DADA2 (Callahan et al., 2016). During filtration, the first 17 nucleotides of the forward reads and the first 21 nucleotides of the reverse reads were trimmed to remove primers. Dereplication, sample inference, chimera identification, and merging of the paired-end reads were performed using the default parameters. Taxonomic assignment was conducted using the SILVA rRNA database (release 138.1) with the naive Bayesian classifier method (utilizing the assignTaxonomy command of the DADA2 package). The sequencing data set was deposited in the NCBI Sequence Read Archive database under the BioProject ID PRJNA1049315.
2.4 Statistical analysis
Statistical analyses were performed using the prism software version 9.2.0. For multiple comparisons, data normality was assessed using the Shapiro-Wilk test. A Kruskal–Wallis test followed by a post hoc Dunn’s test was used to analyze alpha diversity and short-chain fatty acids. A Tukey test was used to compare antibiotic resistance gene scores. Adjusted p-values were employed to evaluate the significance of the results. Statistical analysis of the beta diversity was performed with the adonis function from the vegan package, which is a function to compute permanova tests with 10,000 iterations. Correlation analysis of the SCFAs data (lactic, propionic, butyric, and acetic acids) with bacterial taxa was done with R using the ccrepe package and 10,000 iterations (Schwager et al., 2019). Only correlations with compositionality corrected p-values less than 0.05 were considered significant.
3 Results
3.1 Taxonomic profile
An antibiotic treatment (tylvalosin), a probiotic treatment (P. acidilactici), and a combination of both treatments were administered to six pigs to investigate the resilience and adaptation of their gut microbiome using metagenomic analysis of their faeces and ileal digesta samples. To assess the effect of the treatments on the microbial diversity within the samples (alpha diversity), the Shannon and Simpson diversity indexes were computed for all samples and the treatments were compared (Figure 2). Both indexes are a measure of diversity considering evenness and number of taxa; the Shannon index is more influenced by richness, and the Simpson index is more influenced by evenness of repartition and less influenced by rare species.
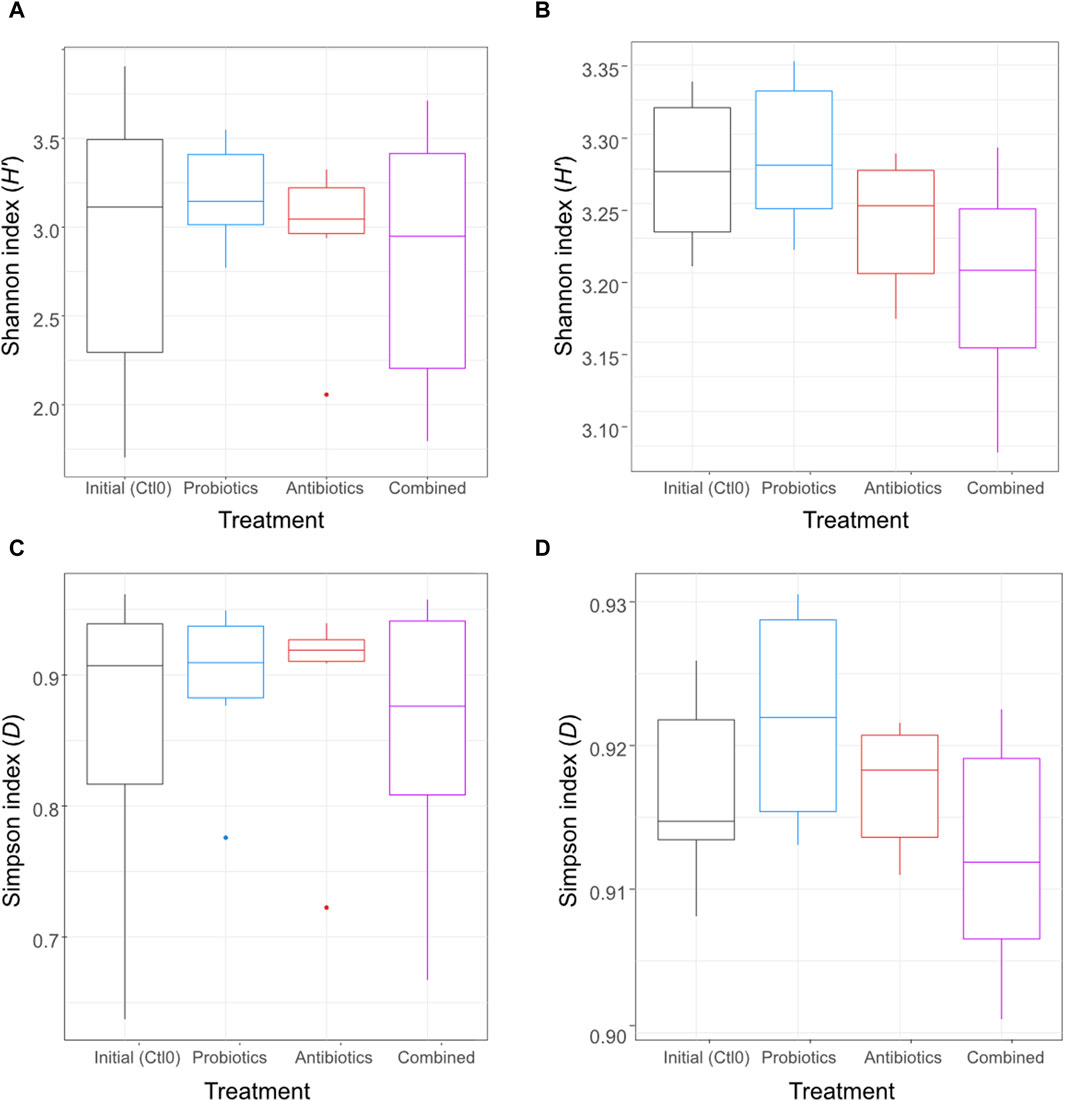
Figure 2. Alpha diversity using Shannon index of the digesta (A) and the faeces (B) and alpha diversity using the Simpson index of the digesta (C) and the faeces (D) for animals treated with the commercial probiotic Pediococcus acidilactici MA18/5M (blue), a macrolide antibiotic (tylvalosin, red), or a combination of both (purple). Data from samples taken before the animals received treatments (Ctl0) are shown in black. Box plots show means and quartiles. No comparison was significant (p > 0.05, Kruskal–Wallis test).
No significant difference was observed (p > 0.05) between treatments for either digesta (Figures 2A, C) or faeces (Figures 2B, D) samples. The metataxonomic analysis of 16S rRNA gene sequences did not reveal any significant difference between treatments for either sample type (Supplementary Figure S1).
The dissimilarity between the samples (beta diversity) was visualized by a principal coordinate analysis (PCoA) plot of the Bray-Curtis distance (Figure 3). The treatments had no significant effect on the microbiota composition of the digesta (p > 0.05; Figure 3A). In the faeces (Figure 3B), all groups were different from the first control period (all p < 0.0096). No other groups were found to be significantly different. As expected, the digesta and the faeces were significantly different from each other (p < 0.0001, Figure 3C) and the variability between faeces samples was greater than the variability between digesta samples. Also, 16S rRNA gene metagenomics validated that the microbiota remained stable between treatments (Supplementary Figure S2).
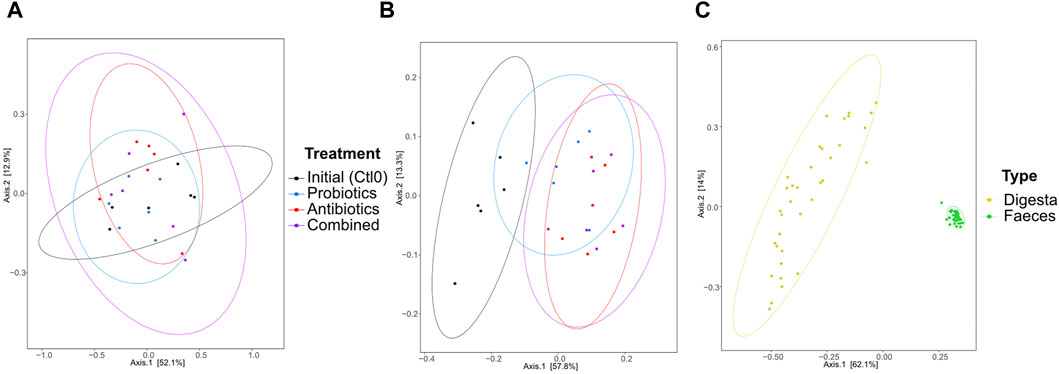
Figure 3. Beta diversity represented by a principal coordinate analysis plot using Bray-Curtis dissimilarities of bacterial taxa from the digesta (A), the faeces (B), and all samples (C) for animals treated with the commercial probiotic Pediococcus acidilactici MA18/5M (blue), a macrolide antibiotic (tylvalosin, red), or a combination of both (purple). Data from samples taken before the animals received treatments (Ctl0) are shown in black. Each point represents a sample. The distance between points reflects the difference in microbial composition between samples; closer points indicate higher similarity. The principal axes represent dimensions that maximize variance among samples. Percentages indicate the proportion of variance explained by each axis. The ellipses represent a 95% confidence interval.
The microbiota composition is similar between treatments (Supplementary Figure S3) and as expected (Holman et al., 2017). The digesta is predominantly composed of the Lactobacillaceae and Peptostreptococcaceae families, accounting for more than half of the microbial composition. Notably, there is a significant increase in Streptococcaceae observed in the samples taken after probiotic treatment. The composition of the feces is more evenly distributed among various taxa, including several families of unclassified Eubacteriales, Bacteroidales, Firmicutes, and Clostridia, as well as the Prevotellaceae family. However, as anticipated, the fecal microbiome is initially largely colonized by bacteria from the Prevotellaceae family at the beginning of the experiment, followed by a subsequent reduction.
Marker taxa were identified using LDA and some of the bacteria varied significantly (p < 0.05) between the antibiotic and probiotic groups (Supplementary Figure S4). Interestingly, the genera that were significantly different between these two groups were only detected during treatment with the probiotic. The most amplified genus in the digesta was the Methanosphaera archaea, while in the faeces it was an unclassified bacterial genus belonging to the Akkermansiaceae family.
3.2 Resistome profile
For each treatment period, the ARG score (calculated by MetaProtMiner) of the previous recuperation period was subtracted from the score calculated for each treatment period to assess only the effect of the treatment (Figures 4A, B). There was no significant difference in digesta samples between treatments (p > 0.05, Figure 4A). However, in the faecal samples, the ARG score change caused by the treatments was significantly higher for the antibiotics and combined treatments than for the probiotic treatment (Figure 4B).
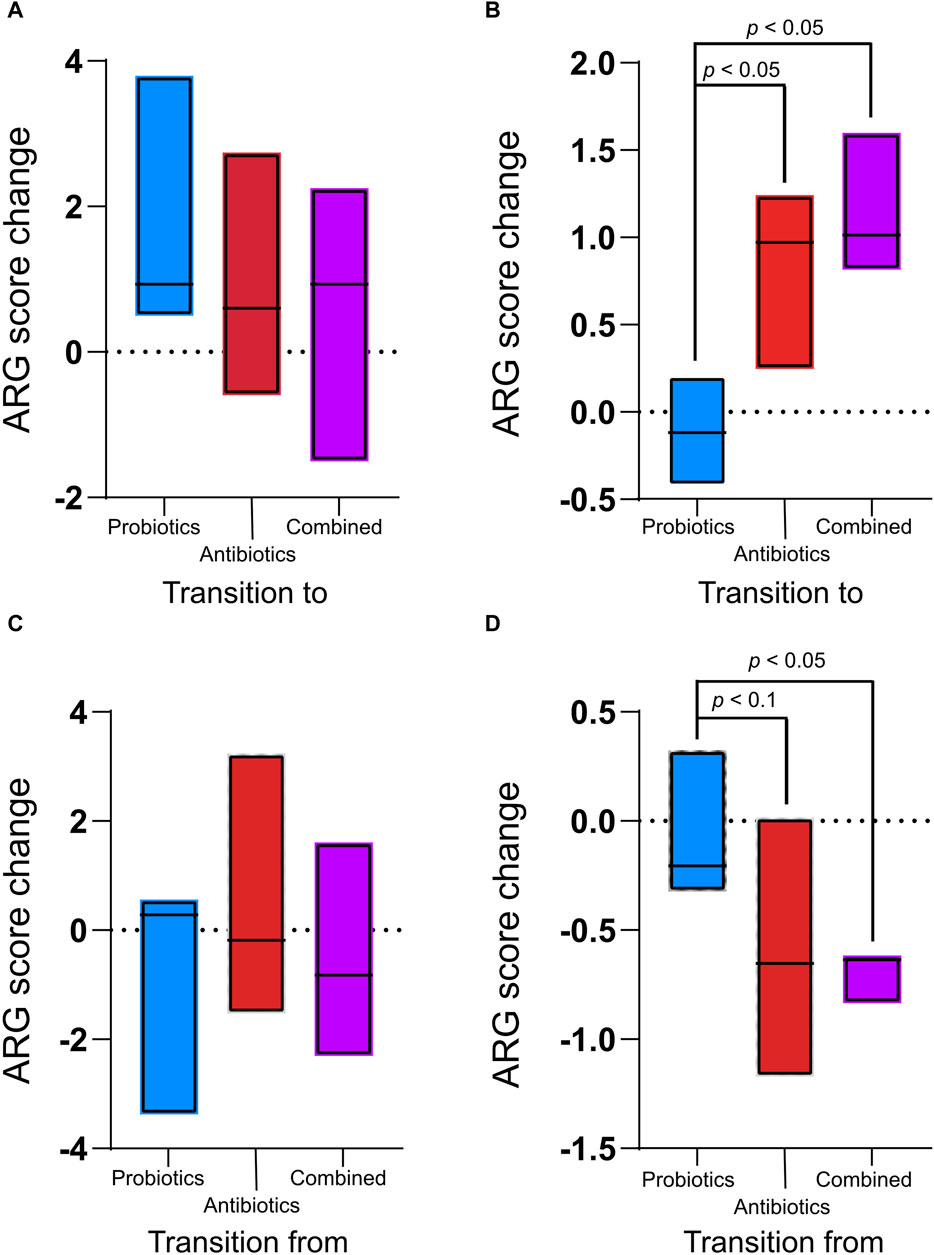
Figure 4. Antibiotic resistance gene (ARG) score change between the treatments (antibiotics, probiotics, or a combination of both) and their preceding recuperation period in the digesta (A) and the faeces (B) and ARG score change between recuperation periods and the following treatment period in the digesta (C) and the faeces (D). The ARG score, computed using MetaProtMiner (Galiot et al., 2023), for a sample corresponds to the sum of reads aligned to each antibiotic resistance gene, divided by the total gene length and the number of reads for the dataset, multiplied by one million. Only p-values less than 0.05 (Tukey test) are shown.
The ARG score for the preceding treatment was subtracted from the score for each recuperation period to verify the resilience of the animals’ microbiome following treatment. During the recuperation periods following treatments, the ARG score change was not significantly different between treatments for the digesta (Figure 4C). However, in the faeces, ARGs decreased during the recuperation period following the antibiotic and combined treatments (Figure 4D). The change during recuperation from the combined treatment was significantly lower than during recuperation from the probiotic treatment (p = 0.04). There was a tendency (p < 0.061) for the decrease in ARGs to be smaller during recuperation after antibiotic treatment than during recuperation from probiotic treatment. In the faeces for the combined treatment, the decrease in ARGs during recuperation was smaller than the increase during treatment (p = 0.02), while the difference was not significant for the antibiotic treatment (p = 0.31).
Sample grouping based on the resistome profile was done with a PCoA plot using Bray-Curtis dissimilarity (Figure 5). In the digesta (Figure 5A), significant differences were observed between the initial microbiome (control) and the antibiotics treatment (p = 0.046), but the comparison was not significant between the initial microbiome and the combined treatment, although the p-value is close to 0.05 (p = 0.051). Differences were also found between the probiotic treatment and antibiotic treatment (p = 0.037) and the combined treatment (p = 0.005). In the faeces (Figure 5B), all groups were different (all p < 0.003) except for the combined and antibiotic treatments (p = 0.7534). The resistome profile was also different between the total samples coming from the digesta and the faeces (p < 0.0001, Figure 5C). More variability in the resistome profile was observed in the faeces samples from the probiotic treatment compared with the combined and antibiotic treatments. Interestingly, the opposite trend was observed in the digesta samples: the samples from the probiotic treatment group were less variable than the combined and antibiotic treatment groups. Globally, there was also more variability in the faeces samples than in the digesta samples (Figure 5C).
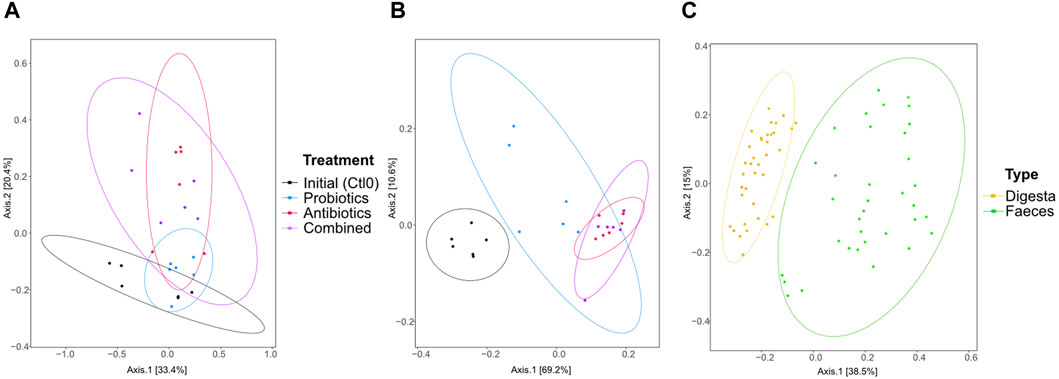
Figure 5. PCoA of the resistome based on the antibiotic resistance gene (ARG) scores during the first recuperation period (Ctl0) and the treatment periods (antibiotics, probiotics, or a combination of both) of the digesta (A), the faeces (B), and all samples (C). The ARG score, computed using MetaProtMiner (Galiot et al., 2023), for a sample corresponds to the sum of reads aligned to each antibiotic resistance gene, divided by the total gene length and the number of reads for the dataset, multiplied by one million. Each point represents a sample. The distance between points reflects the difference in resistance genes composition between samples; closer points indicate higher similarity. The principal axes represent dimensions that maximize variance among samples. Percentages indicate the proportion of variance explained by each axis. The ellipses represent a 95% confidence interval.
Marker resistance genes between the probiotic and antibiotic treatments were identified with LDA analysis (Supplementary Figure S5); no markers were found between the combined and antibiotic treatments. A heatmap of the abundance score of those genes across samples is shown in Figure 6. In both faeces and digesta, hierarchical clustering analysis of the transitions in the whole resistome between the control and treatments showed that the changes induced by the probiotic treatment clustered together, while the changes induced by the antibiotic or combined treatments clustered together (Figure 6). The ARGs that were considered markers with LDA were amplified in the transition to antibiotic and combined treatments and were stable or diminished in the transition to probiotic treatment. Five ARG markers were found in the digesta (Figure 6A), of which ermB, ermG, and ermQ were macrolide resistance genes. A total of 14 ARG markers were found for the faeces samples (Figure 6B), and five of these (ermB, ermG, ermT, ermQ, and ermF) were macrolide resistance genes. This is not surprising, as tylvalosin is an antibiotic of the macrolide class. Interestingly, all the genes identified as markers in the digesta samples were also markers in the faeces samples.
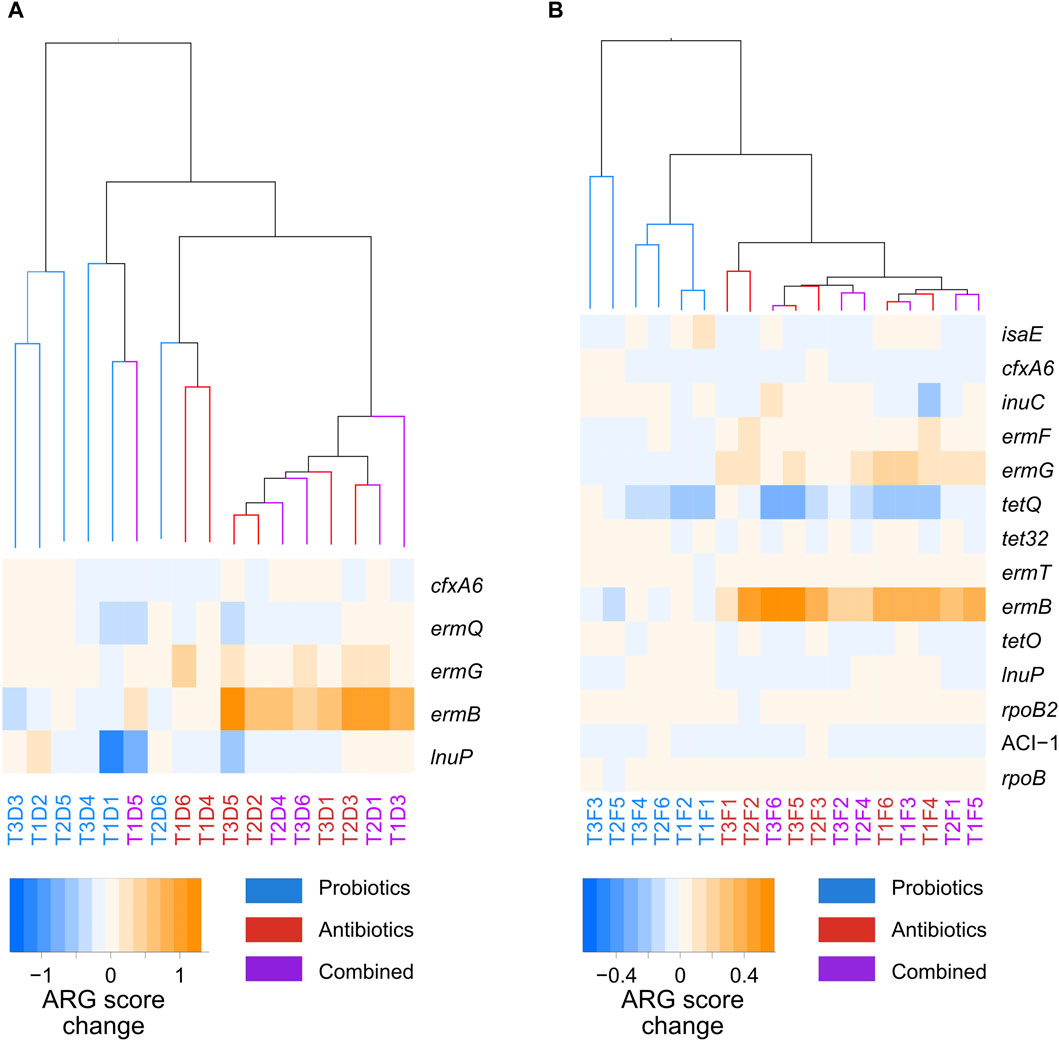
Figure 6. Hierarchical clustering of the whole-resistome changes caused by the treatments, and a heatmap of the marker genes between probiotic, antibiotic, and a combination of both treatments for samples taken from the digesta (A) and the faeces (B). The samples names correspond to the treatment period of the samples: T1, T2, or T3; the nature of the samples: faeces (F) or digesta (D); and the pig from which the sample was taken: pig 1 to 6. Blue corresponds to probiotic treatment, red to antibiotic treatment, and purple to a combination of both treatments.
3.3 Plasmidome profile
After screening of the total assembly for plasmidic sequences, 38,565 of 9,444,776 contigs were identified to be putatively plasmidic. These sequences were compared with the CARD database, and 27 contigs were carrying at least one ARG and two contigs were carrying two ARGs. No contigs were found carrying more than two ARGs. Plasmid sequences were reconstructed from contigs, and those carrying ARGs are described in Table 1. Two plasmids, AA998 and AA448, were carrying multiple ARGs, and both were considered as conjugative plasmids by the tool mob_suite. The plasmid AA448 is associated with Klebsiella pneumonia and was carrying genes for resistance against the tetracycline, gentamycin, and sulfonamide classes of antibiotics. The plasmid AA998 was associated with Salmonella enterica serovar Typhi and was carrying resistance genes against the beta-lactam, tetracycline, chloramphenicol, and macrolide classes of antibiotics (Table 1).
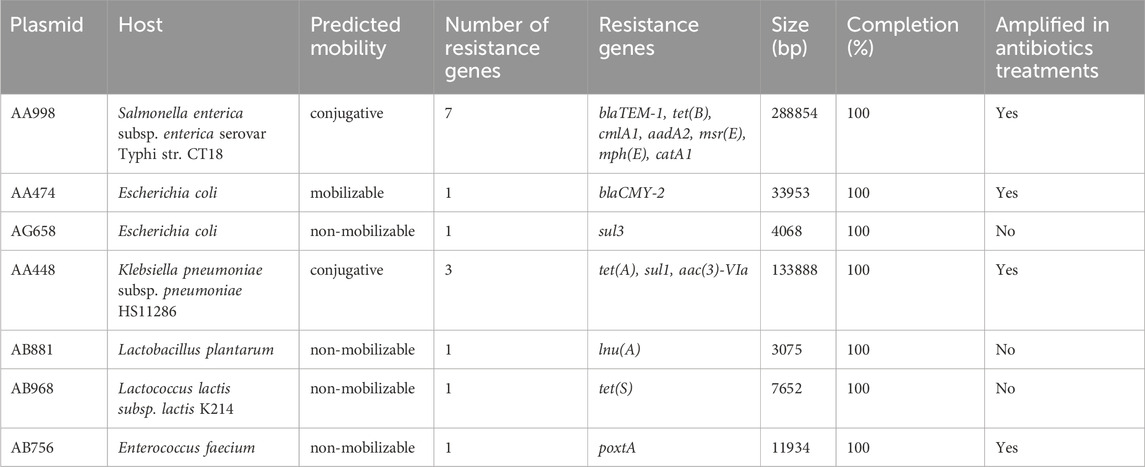
Table 1. Description of antibiotic resistance gene–carrying plasmids found in all digesta and faeces samples.
To identify marker plasmidic contigs between the antibiotic and probiotic treatments, LDA analysis was performed (Supplementary Figure S6). In the digesta, all maker plasmids were amplified in the probiotic treatment; however, none of the marker contigs were carrying ARGs (Supplementary Figure S6). In the faeces, all markers for plasmidic contigs were amplified in the antibiotic treatment. Four of the contigs that were markers in the faeces were carrying ARGs, and three of these belonged to plasmids that were either conjugative or mobilizable.
Samples were grouped based on the transitions in the whole plasmidome between the control and treatments using hierarchical clustering with heatmaps of the transition of the ARG-carrying plasmids (Supplementary Figure S7). The whole plasmidome of the samples did not cluster according to treatments.
3.4 Functional profile
To more specifically assess the role of the treatments on functional profile, the relative abundance of functions, as defined by the Pfam database (Finn et al., 2008), was established and markers were identified using LDA analysis. In the digesta, all the marker functions were more frequent in the probiotics group and no horizontal transfer functions were identified as markers (Figures 7A). In the faeces, two of the functions identified as markers for antibiotic treatments were recombinase (PF07508 [Recombinase]) and resolvase (PF00239 [Resolvase, N terminal domain]); these two functions play a role in gene exchange (Figures 7B) (Johnson and Grossman, 2015; Rice, 2015).
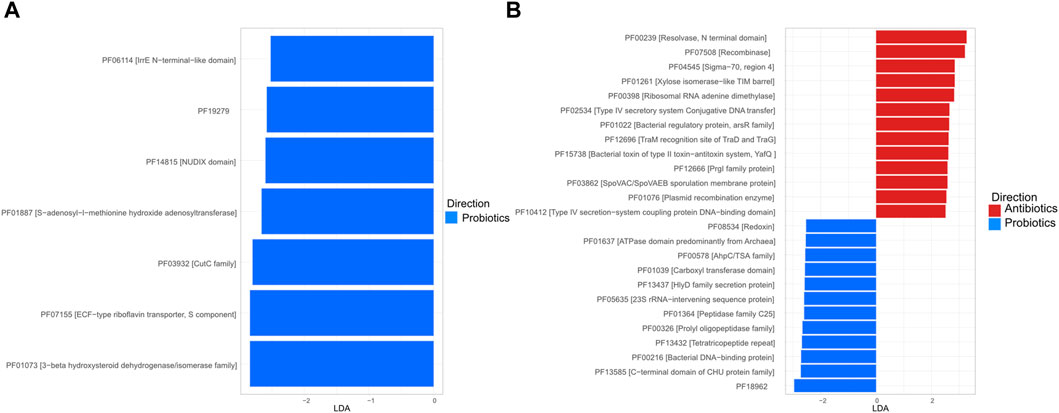
Figure 7. Linear discriminant analysis (LDA) plot of the marker function, as defined in the Pfam database, in the digesta (A) and the faeces (B). For the digesta, only samples from animals fed with the probiotic had markers (score ≥2 and a p < 0.05).
Four SCFAs were measured in the samples: acetic acid, lactic acid, propionic acid, and butyric acid; these are important metabolites for host health (Bergman, 1990; De Vadder et al., 2014; Jacobson et al., 2018; Schulthess et al., 2019). The SCFA content was compared between treatments, and there were no significant differences in the quantity of SCFAs measured between treatments in the digesta (Supplementary Figure S8) or the faeces (Supplementary Figure S9).
The individual SCFA contents were also compared between the digesta and the faeces (Figure 8). The contents of acetic, propionic, and butyric acid were higher in the faeces (p < 0.0001) while lactic acid was higher in the digesta (p < 0.0001).
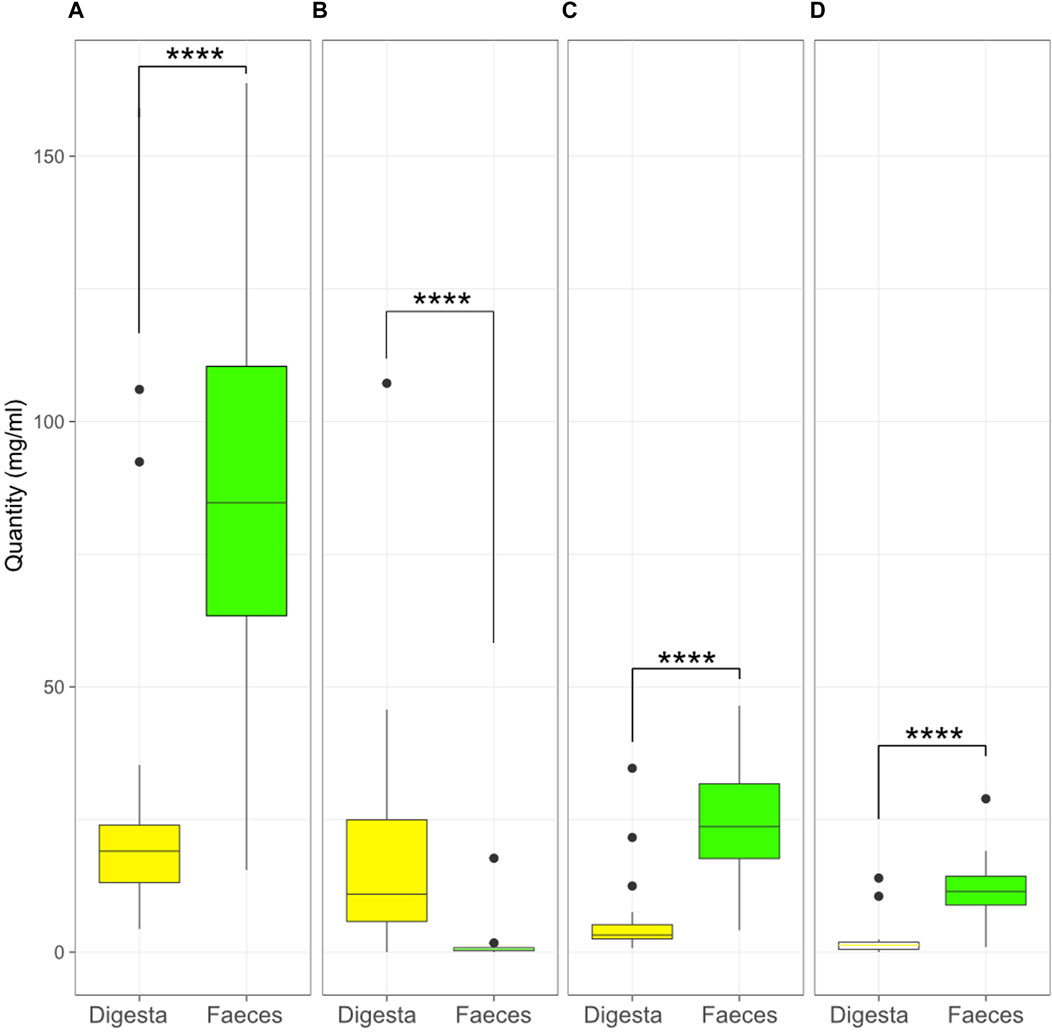
Figure 8. Quantity (mg/mL) of acetic (A), lactic (B), propionic (C), and butyric (D) acid in the digesta and the faeces (****, p < 0.0001, Kruskal–Wallis test). Box plots show means and quartiles.
The correlations between the SCFA content and bacterial species were also tested in the faeces and the digesta. In the digesta, various bacteria, including many Clostridium species, were negatively correlated with lactic acid and positively correlated with the other SCFAs. In contrast, lactobacilli were positively correlated with lactic acid, and negatively or not correlated with other SCFAs (Figure 9A). In the faeces, lactic acid did not seem to follow any trend, but there was a clear separation between bacteria that were positively correlated with acetic acid and negatively with butyric and propionic acids, and bacteria that were positively correlated with butyric and propionic acid and negatively correlated with acetic acid (Figure 9B).
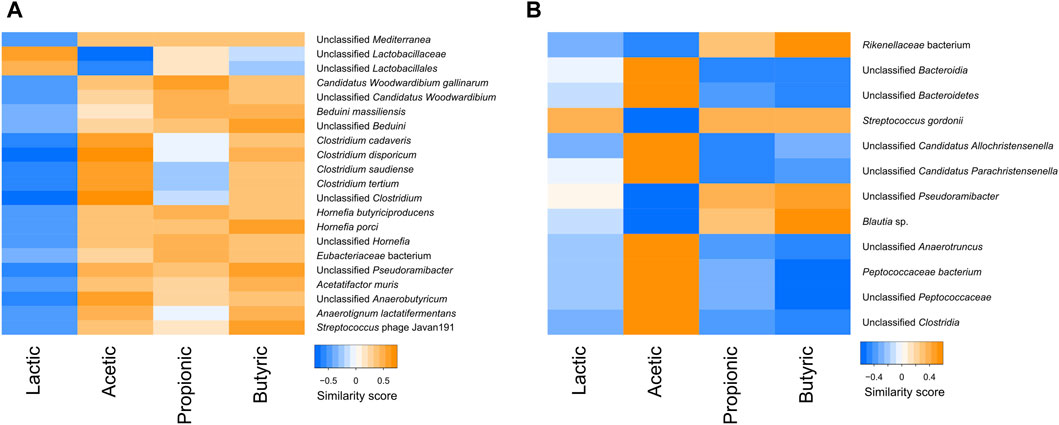
Figure 9. Heatmap of the similarity score between correlated short chain fatty acids and taxa in the digesta (A) and the faeces (B).
4 Discussion
The aim of the study was to determine whether the administration of a probiotic (P. acidilactici) can mitigate the effects of an antibiotic (tylvalosin) on the microbial community, both taxonomically and functionally, in swine. Although no significant differences in alpha diversity were observed between the treatments, a few tendencies are interesting. The combined treatment samples had the lowest diversity in the faeces, which might be the result of the combined effect of the tylvalosin and the antimicrobial compounds (e.g., bacteriocin and SCFA) produced by the probiotic P. acidilactici. Both of these antimicrobials target Gram-positive bacteria (Pharmgate, 2021), and Pediococcus bacteriocin was shown to target some Gram-negative enterobacteria when the cells were already sub-lethally injured (Osmanagaoglu, 2005). This effect was not observed in the digesta, which could be due to the composition of its microbiota. In fact, the predominant bacterial genus in digesta samples was Lactobacillus, and lactobacilli strains were reported to have an inactivating effect on some macrolide antibiotics in chicken microbiota (Devriese and Dutta, 1984). Whether or not this was the case in the present study would need to be investigated specifically with pigs and tylvalosin.
The lack of significance for beta diversity changes between the treatments could partially be explained by the low number of samples. Because there is a high variability between the microbiota of individual pigs, small changes resulting from treatment are not always distinguishable from individual variations (Chen C. et al., 2018). Comparisons between 16S metagenomics data and shotgun sequencing reveal similar conclusions regarding microbial diversity, suggesting that amplicon sequencing alone may suffice to address taxonomic questions.
While the resistome profiles of samples were different between the probiotic treatment and the antibiotic and combined treatments in both the digesta and the faeces, the total ARG score was only higher in the faeces of antibiotic-treated animals. This means that a change in the resistome profile and enrichment of specific genes does not necessarily imply an increase in the total ARG content of a sample. Therefore, simply measuring the quantity of ARGs would be insufficient to properly monitor resistance. There were also fewer increased ARGs in the digesta during antibiotic treatment compared with the faeces, further demonstrating that the digesta and faeces resistomes respond differently to antibiotic treatment. It is important to note that exposure time to in-feed antibiotics could be shorter in the digesta, as the flow of digesta is faster in the ileum than in the colon (Wilfart et al., 2007).
The recuperation period allowed for a decrease in ARGs, although it was lower than the increase occurring during the antibiotic and combined treatments. This means that the microbiome can recover from such treatments, although 3 weeks might not be enough to completely return to an untreated state. Further studies investigating the recovery of the microbiome over a longer period would be necessary to pinpoint the reversal of the treatment effects.
The increase in resolvase and recombinase in the antibiotic-treated faeces samples could suggest higher rates of plasmid transfer. It is not possible to establish what proportion of this difference is explained by higher rates of transfer or by selection of bacteria carrying plasmids with both horizontal transfer genes and ARGs. The fact that two conjugative multi-resistance–carrying plasmids were more frequently detected in samples from the antibiotic treatments, and that Pfam resolvase and recombinase functions were more frequently detected in those samples, indicates a higher risk of horizontal transfer. Other methods, such as epigenomics grouping or culturomics would be necessary to confirm the significance of horizontal transfers between the treatments (Dib et al., 2015). All ARG-carrying plasmids and horizontal gene transfer–related genes that were more present in the antibiotic-treated samples were observed in the faeces, which suggests that the potential for horizontal transfer is higher in the faeces than in the digesta. This represents a problem considering that the faeces are responsible for most of the environmental contamination by pig gut microbiota, notably in fertilized fields (Gao et al., 2020) and pig carcasses (Laforge et al., 2023).
The transferable plasmids that were amplified in the antibiotic and combined treatment samples were originally from E. coli and Klebsiella pneumoniae, bacteria that were naturally not susceptible to tylvalosin (Pharmgate, 2021); this could explain why the plasmids from E. coli and K. pneumoniae were more prevalent in antibiotic-treated samples even though they were not carrying macrolide resistance genes. Therefore, naturally non-susceptible bacteria should not be ignored in the selection and transfer of ARGs during resistome studies using antibiotic treatment.
The treatments also did not impact the quantity of SCFAs in the gut microbiome, even for the probiotic-supplemented group; although P. acidilactici is known to produce lactic (Klupsaite et al., 2019), butyric, and propionic acid, as well as low levels of acetic acid (Hoseinifar et al., 2017). Analysis of the correlations between SCFAs and bacterial taxa abundance showed that some bacteria were strongly associated with SCFAs, and those correlations varied between digesta and the faeces. The difference in the quantity of SCFAs in the digesta and faeces demonstrate a highly different SCFA profile of the digesta and the faeces, suggesting that the fermentation patterns in the ileum and colon are different in many ways (Urriola and Stein, 2010; Zhang et al., 2018).
In conclusion, the samples collected during antibiotic or combined treatment had very similar profiles of taxonomy, resistance, and plasmidome. This observation would suggest that P. acidilactici given simultaneously with tylvalosin does not reduce the resistance selection effect of antibiotic treatment on the resistome. Therefore, there is little to no advantage to this practice. The digesta and faeces responded differently to the treatments in their microbial composition, resistome, and plasmidome; therefore, inferences made from the digesta to faeces, or vice versa, are not applicable to the other for microbiome studies. It is important to mention that the pigs in this study were housed separately, and in larger pens there might be a higher transfer of faecal bacteria to the ileum through a faecal-mouth-ileum pathway. Although studies regarding digesta are still important for specific pathogens that colonize the ileum, the faeces seem more important for further investigation of the effects of antibiotics on the pig gut microbiome with respect to meat safety, as faeces are more problematic in term of antibiotics resistance selection, horizontal transfer, and contamination of the food chain. Finally, the present study uses a cannulated pig model to infer the effect of an antibiotic and a probiotic on ileal and fecal microbiota. Although this model is known to enable exploration of the microbiome efficiently (Gao et al., 2018; Lee et al., 2018; Pi et al., 2019), the results should be validated in a commercial setting representing real-life husbandry practices.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Ethics statement
The animal study was approved by the Animal Protection Committee of Université Laval. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
XM: Writing–original draft, Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Writing–review and editing. LS: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Writing–original draft, Writing–review and editing, Funding acquisition, Project administration, Resources, Supervision, Validation. FG: Conceptualization, Funding acquisition, Writing–original draft, Writing–review and editing. AT: Conceptualization, Writing–original draft, Writing–review and editing. JL: Conceptualization, Writing–original draft, Writing–review and editing. EP: Conceptualization, Writing–original draft, Writing–review and editing. SF: Conceptualization, Funding acquisition, Writing–original draft, Writing–review and editing. AV: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing–original draft, Writing–review and editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Funding was provided by A.T.V.’s start-up fund (Université Laval), a Discovery grant of the Natural Sciences and Engineering Research Council of Canada (grant No. RGPIN-2022-03321) from A.T.V, and the Canadian Food Innovation Network (CFIN) science cluster and funded in part by the Government of Canada under the Canadian Agricultural Partnership’s AgriScience Program, a federal, provincial, territorial initiative of LS.
Conflict of interest
Authors EP and SF were employed by Olymel S.E.C./L.P.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2024.1355134/full#supplementary-material
References
Alcock, B. P., Raphenya, A. R., Lau, T. T. Y., Tsang, K. K., Bouchard, M., Edalatmand, A., et al. (2020). CARD 2020: antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 48, D517–D525. doi:10.1093/nar/gkz935
Bergman, E. N. (1990). Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 70, 567–590. doi:10.1152/physrev.1990.70.2.567
Bushnell, B. (2014). BBmap. Available at: https://sourceforge.net/projects/bbmap/(Accessed October 13, 2021).
Callahan, B. J., Mcmurdie, P. J., Rosen, M. J., Han, A. W., Johnson, A. J. A., and Holmes, S. P. (2016). Dada2: high-resolution sample inference from illumina amplicon data. Nat. Methods 13, 581–583. doi:10.1038/nmeth.3869
Canibe, N., Hojberg, O., Kongsted, H., Vodolazska, D., Lauridsen, C., Nielsen, T. S., et al. (2022). Review on preventive measures to reduce post-weaning diarrhoea in piglets. Animals 12, 2585. doi:10.3390/ani12192585
Chen, C., Huang, X., Fang, S., Yang, H., He, M., Zhao, Y., et al. (2018b). Contribution of host genetics to the variation of microbial composition of cecum lumen and feces in pigs. Front. Microbiol. 9, 2626. doi:10.3389/fmicb.2018.02626
Chen, S., Zhou, Y., Chen, Y., and Gu, J. (2018a). fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 34, i884–i890. doi:10.1093/bioinformatics/bty560
De Vadder, F., Kovatcheva-Datchary, P., Goncalves, D., Vinera, J., Zitoun, C., Duchampt, A., et al. (2014). Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 156, 84–96. doi:10.1016/j.cell.2013.12.016
Devriese, L. A., and Dutta, G. N. (1984). Effects of erythromycin-inactivating lactobacillus crop flora on blood-levels of erythromycin given orally to chicks. J. Vet. Pharmacol. Ther. 7, 49–53. doi:10.1111/j.1365-2885.1984.tb00878.x
Dib, J. R., Wagenknecht, M., Farías, M. E., and Meinhardt, F. (2015). Strategies and approaches in plasmidome studies uncovering plasmid diversity disregarding of linear elements? Front. Microbiol. 6, 463. doi:10.3389/fmicb.2015.00463
Dixon, P. (2003). Vegan, a package of r functions for community ecology. J. Veg. Sci. 14, 927–930. doi:10.1111/j.1654-1103.2003.tb02228.x
Finn, R. D., Tate, J., Mistry, J., Coggill, P. C., Sammut, S. J., Hotz, H. R., et al. (2008). The pfam protein families database. Nucleic Acids Res. 36, D281–D288. doi:10.1093/nar/gkm960
Galiot, L., Monger, X. C., and Vincent, A. T. (2023). Studying the association between antibiotic resistance genes and insertion sequences in metagenomes: challenges and pitfalls. Antibiotics 12, 175. doi:10.3390/antibiotics12010175
Gao, F. Z., He, L. Y., He, L. X., Zou, H. Y., Zhang, M., Wu, D. L., et al. (2020). Untreated swine wastes changed antibiotic resistance and microbial community in the soils and impacted abundances of antibiotic resistance genes in the vegetables. Sci. Total Environ. 741, 140482. doi:10.1016/j.scitotenv.2020.140482
Gao, K., Pi, Y., Peng, Y., Mu, C.-L., and Zhu, W.-Y. (2018). Time-course responses of ileal and fecal microbiota and metabolite profiles to antibiotics in cannulated pigs. Appl. Microbiol. Biotechnol. 102, 2289–2299. doi:10.1007/s00253-018-8774-2
Guevarra, R. B., Kim, E. S., Cho, J. H., Song, M., Cho, J. H., Lee, J. H., et al. (2023). Gut microbial shifts by synbiotic combination of Pediococcus acidilactici and lactulose in weaned piglets challenged with shiga toxin-producing Escherichia coli. Front. Vet. Sci. 9, 1101869. doi:10.3389/fvets.2022.1101869
Holman, D. B., Brunelle, B. W., Trachsel, J., and Allen, H. K. (2017). Meta-analysis to define a core microbiota in the swine gut. mSystems 2, e00004-17. doi:10.1128/msystems.00004-17
Hoseinifar, S. H., Mirvaghefi, A., Amoozegar, M. A., Merrifield, D. L., and Ringo, E. (2017). In vitro selection of a synbiotic and in vivo evaluation on intestinal microbiota, performance and physiological response of rainbow trout (oncorhynchus mykiss) fingerlings. Aquac. Nutr. 23, 111–118. doi:10.1111/anu.12373
Jacobson, A., Lam, L., Rajendram, M., Tamburini, F., Honeycutt, J., Pham, T., et al. (2018). A gut commensal-produced metabolite mediates colonization resistance to Salmonella infection. Cell Host Microbe 24, 296–307. doi:10.1016/j.chom.2018.07.002
Johnson, C. M., and Grossman, A. D. (2015). Integrative and conjugative elements (ices): what they do and how they work. Annu. Rev. Genet. 49, 577–601. doi:10.1146/annurev-genet-112414-055018
Joysowal, M., Saikia, B. N., Tamuly, S., and Kalita, D. (2021). Comparative effect of probiotics (swine and dairy origin) on growth performance, nutrient digestibility, blood biochemical profile, immune status and faecal microbial count of early-weaned grower-finisher crossbred pigs. Indian J. Anim. Res. 55, 1308–1314. doi:10.18805/ijar.B-4137
Kai, G. (2021). Microbial. Available at: https://github.com/guokai8/microbial (Accessed October 23, 2023).
Khan, A., and Rao, T. S. (2019). Molecular evolution of xenobiotic degrading genes and mobile DNA elements in soil bacteria. London: Academic Press Ltd-Elsevier Science Ltd.
Kim, J. H., Ahn, H., Lee, D., Lee, H., and Kim, W. J. (2023). Antibiofilm activity of crude bacteriocin jm01 produced by Pediococcus acidilactici against methicillin-resistant Staphylococcus aureus (MRSA). Int. J. Food Sci. Technol. 58, 2580–2589. doi:10.1111/ijfs.16407
Klupsaite, D., Juodeikiene, G., Arbones, E., Quintáns, A. P., Zadeike, D., Bartkiene, E., et al. (2019). A comparison study on the production and recovery of lactic acid by fermenting dairy by-products with P acidilactici and lb delbrueckii spp bulgaricus. Waste Biomass Valor 10, 1519–1528. doi:10.1007/s12649-017-0171-z
Laforge, P., Vincent, A. T., Duchaine, C., Feutry, P., Dion-Fortier, A., Plante, P.-L., et al. (2023). Contribution of farms to the microbiota in the swine value chain. Front. Sys. Biol. 3. doi:10.3389/fsysb.2023.1183868
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with bowtie 2. Nat. Methods 9, 357–359. doi:10.1038/nmeth.1923
Larkin, H. (2023). Increasing antimicrobial resistance poses global threat, who says. Jama 329, 200. doi:10.1001/jama.2022.23552
Lee, K. Y., Balasubramanian, B., Kim, J. K., and Kim, I. H. (2018). Dietary inclusion of xylanase improves growth performance, apparent total tract nutrient digestibility, apparent ileal digestibility of nutrients and amino acids and alters gut microbiota in growing pigs. Anim. Feed Sci. Technol. 235, 105–109. doi:10.1016/j.anifeedsci.2017.11.015
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., et al. (2009). The sequence alignment/map format and samtools. Bioinformatics 25, 2078–2079. doi:10.1093/bioinformatics/btp352
Lopez-Galvez, G., Lopez-Alonso, M., Pechova, A., Mayo, B., Dierick, N., and Gropp, J. (2021). Alternatives to antibiotics and trace elements (copper and zinc) to improve gut health and zootechnical parameters in piglets: a review. Anim. Feed Sci. Technol. 271, 114727. doi:10.1016/j.anifeedsci.2020.114727
Mcmurdie, P. J., and Holmes, S. (2013). Phyloseq: an r package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 8, e61217. doi:10.1371/journal.pone.0061217
Monger, X. C., Gilbert, A.-A., Saucier, L., and Vincent, A. T. (2021). Antibiotic resistance: from pig to meat. Antibiotics 10, 1209. doi:10.3390/antibiotics10101209
Monger, X. C., Saucier, L., Gilbert, A.-A., and Vincent, A. T. (2022). Stabilization of swine faecal samples influences taxonomic and functional results in microbiome analyses. MethodsX 9, 101716. doi:10.1016/j.mex.2022.101716
OIE (2018). in Annual report on antimicrobial agents intended for use in animals (Paris, France: W.O.F.A. Health).
Osmanagaoglu, O. (2005). Sensitivity of sublethally injured gram-negative bacteria to pediocin P. J. Food. Saf. 25, 266–275. doi:10.1111/j.1745-4565.2005.00022.x
Pan, X. X., Zhou, Z. Y., Liu, B. W., and Wu, Z. W. (2022). A novel therapeutic concern: antibiotic resistance genes in common chronic diseases. Front. Microbiol. 13, 1037389. doi:10.3389/fmicb.2022.1037389
Pereira, W. A., Franco, S. M., Reis, I. L., Mendonca, C. M. N., Piazentin, A. C. M., Azevedo, P. O. S., et al. (2022). Beneficial effects of probiotics on the pig production cycle: an overview of clinical impacts and performance. Vet. Microbiol. 269, 109431. doi:10.1016/j.vetmic.2022.109431
Pezsa, N. P., Kovacs, D., Racz, B., and Farkas, O. (2022). Effects of Bacillus licheniformis and Bacillus subtilis on gut barrier function, proinflammatory response, ros production and pathogen inhibition properties in ipec-j2-Escherichia coli/Salmonella typhimurium co-culture. Microorganisms 10, 936. doi:10.3390/microorganisms10050936
Pharmgate (2021). Aivlosin product profile. Available at: https://pharmgate.com/canada/wp-content/uploads/2021/05/PAH_PP_Aivlosin-WSG_French-English_FINAL_050721.pdf (Accessed July 12, 2023).
Pi, Y., Gao, K., Peng, Y., Mu, C. L., and Zhu, W. Y. (2019). Antibiotic-induced alterations of the gut microbiota and microbial fermentation in protein parallel the changes in host nitrogen metabolism of growing pigs. animal 13, 262–272. doi:10.1017/S1751731118001416
Pradier, L., Tissot, T., Fiston-Lavier, A.-S., and Bedhomme, S. (2021). Plasforest: a homology-based random forest classifier for plasmid detection in genomic datasets. BMC Bioinforma. 22, 349. doi:10.1186/s12859-021-04270-w
Puente-Sanchez, F., Garcia-Garcia, N., and Tamames, J. (2020). Sqmtools: automated processing and visual analysis of 'omics data with r and anvi'o. BMC Bioinforma. 21, 358. doi:10.1186/s12859-020-03703-2
Qu, L. L., and Chen, Z. A. (2022). A mathematical model of plasmid-carried antibiotic resistance transmission in two types of cells. Appl. Math. Nonlinear Sci. 8, 2331–2344. doi:10.2478/amns.2021.2.00178
Rice, P. A. (2015). Serine resolvases. Microbiol. Spectr. 3, MDNA3–M0045. doi:10.1128/microbiolspec.MDNA3-0045-2014
Ricker, N., Trachsel, J., Colgan, P., Jones, J., Choi, J., Lee, J., et al. (2020). Toward antibiotic stewardship: route of antibiotic administration impacts the microbiota and resistance gene diversity in swine feces. Front. Vet. Sci. 7, 255. doi:10.3389/fvets.2020.00255
Robertson, J., and Nash, J. H. E. (2018). Mob-suite: software tools for clustering, reconstruction and typing of plasmids from draft assemblies. Microb. Genom. 4, e000206. doi:10.1099/mgen.0.000206
Roussel, C., Chabaud, S., Lessard-Lord, J., Cattero, V., Pellerin, F.-A., Feutry, P., et al. (2022). UPEC colonic-virulence and urovirulence are blunted by proanthocyanidins-rich cranberry extract microbial metabolites in a gut model and a 3D tissue-engineered urothelium. Microbiol. Spectr. 10, e0243221. doi:10.1128/spectrum.02432-21
Schulthess, J., Pandey, S., Capitani, M., Rue-Albrecht, K. C., Arnold, I., Franchini, F., et al. (2019). The short chain fatty acid butyrate imprints an antimicrobial program in macrophages. Immunity 50, 432–445. doi:10.1016/j.immuni.2018.12.018
Schwager, E., Bielski, C., and Weingart, G. (2019). Ccrepe_and_nc. Score. r package version 1.18.1. Available at: https://github.com/biobakery/biobakery (Accessed May 24, 2023).
Shin, D., Chang, S. Y., Bogere, P., Won, K., Choi, J.-Y., Choi, Y.-J., et al. (2019). Beneficial roles of probiotics on the modulation of gut microbiota and immune response in pigs. PLoS One 14, e0220843. doi:10.1371/journal.pone.0220843
Suzuki, R., and Shimodaira, H. (2006). Pvclust: an r package for assessing the uncertainty in hierarchical clustering. Bioinformatics 22, 1540–1542. doi:10.1093/bioinformatics/btl117
Tamames, J., and Puente-Sanchez, F. (2019). Squeezemeta, a highly portable, fully automatic metagenomic analysis pipeline. Front. Microbiol. 9, 3349. doi:10.3389/fmicb.2018.03349
Urriola, P. E., and Stein, H. H. (2010). Effects of distillers dried grains with solubles on amino acid, energy, and fiber digestibility and on hindgut fermentation of dietary fiber in a corn-soybean meal diet fed to growing pigs. J. Anim. Sci. 88, 1454–1462. doi:10.2527/jas.2009-2162
WHO (2020). Global antimicrobial resistance and use surveillance system (glass) report: early implementation 2020.
Wilfart, A., Montagne, L., Simmins, H., Noblet, J., and Van Milgen, J. (2007). Digesta transit in different segments of the gastrointestinal tract of pigs as affected by insoluble fibre supplied by wheat bran. Br. J. Nutr. 98, 54–62. doi:10.1017/s0007114507682981
Wubben, J. E., Smiricky, M. R., Albin, D. M., and Gabert, V. M. (2001). Improved procedure and cannula design for simple-t cannulation at the distal ileum in growing pigs. Contemp. Top. Lab. Anim. Sci. 40, 27–31.
Zhang, S. X., Wang, H. B., Zhang, H., Liu, H. F., Zhou, Z. F., Zhang, J. T., et al. (2014). Pilot laparoscopic ileal cannulation in pigs. Can. J. Anim. Sci. 94, 371–377. doi:10.4141/cjas2013-171
Zhang, Y. J., Liu, Q., Zhang, W. M., Zhang, Z. J., Wang, W. L., and Zhuang, S. (2018). Gastrointestinal microbial diversity and short-chain fatty acid production in pigs fed different fibrous diets with or without cell wall-degrading enzyme supplementation. Livest. Sci. 207, 105–116. doi:10.1016/j.livsci.2017.11.017
Keywords: antibiotics resistance, digesta, faeces, gut microbiome, metagenomics, plasmids, swine
Citation: Monger XC, Saucier L, Guay F, Turcotte A, Lemieux J, Pouliot E, Fournaise S and Vincent AT (2024) Effect of a probiotic and an antibiotic on the mobilome of the porcine microbiota. Front. Genet. 15:1355134. doi: 10.3389/fgene.2024.1355134
Received: 13 December 2023; Accepted: 19 March 2024;
Published: 28 March 2024.
Edited by:
Junbo Ma, University of North Carolina at Chapel Hill, United StatesReviewed by:
Mohamed Zeineldin, Animal and Plant Health Inspection Service (USDA), United StatesXianglilan Zhang, Beijing Institute of Microbiology and Epidemiology, China
Loïc Deblais, The Ohio State University, United States
Copyright © 2024 Monger, Saucier, Guay, Turcotte, Lemieux, Pouliot, Fournaise and Vincent. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Antony T. Vincent, YW50b255LnZpbmNlbnRAZnNhYS51bGF2YWwuY2E=