 Duan Sun
Duan Sun Zihao Ding
Zihao Ding Yong Hai
Yong Hai Yunzhong Cheng
Yunzhong Cheng
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Genet. , 26 July 2023
Sec. Epigenomics and Epigenetics
Volume 14 - 2023 | https://doi.org/10.3389/fgene.2023.1211376
This article is part of the Research Topic The role of epigenetics in bone metabolism View all 5 articles
Scoliosis is a three-dimensional structural deformity of the spine; more than 80% of scoliosis has no specific pathogenesis but is understood to be closely related to genetic, hormonal, and environmental factors. In recent years, the epigenetic alterations observed in scoliosis have been analyzed in numerous studies to determine the pathogenesis and progression of this condition, however, there is currently no comprehensive review of the epigenetic factors to date. We searched PubMed, Embase, and Web of Science databases for relative studies without language and date restrictions in March 2023. Twenty-five studies were included in this review and analyzed from the four main aspects of epigenetic alteration: DNA methylation, non-coding RNAs, histone modifications, and chromatin remodeling. The relationship between DNA methylation, non-coding RNAs, and scoliosis was considerably reported in the literature, and the corresponding related signaling pathways and novel biomarkers observed in scoliosis provide insights into innovative prevention and treatment strategies. However, the role of histone modifications is rarely reported in scoliosis, and few studies have investigated the relationship between scoliosis and chromatin remodeling. Therefore, these related fields need to be further explored to elucidate the overall effects of epigenetics in scoliosis.
Scoliosis is a generic term that describes a three-dimensional structural deformity of the spine (Giampietro, 2012), primarily encompassing adolescent idiopathic scoliosis (AIS) (Weinstein et al., 2008), congenital scoliosis (CS) (Hedequist and Emans, 2007; Pahys and orthopedics, 2018), and neuromuscular scoliosis (NMS) (Vialle et al., 2013; Wishart et al., 2021). The pathogenesis of scoliosis has not been established but is currently considered to be associated with a variety of factors, including genetic, hormonal, and environmental factors (Qiu, 2018). Nonetheless, in 2013, Fendri et al. (2013) identified a characteristic gene expression pattern in patients with AIS and used RT-PCR to identify 145 differentially expressed genes (DEGs). Therefore, the effects of epigenetic modifications in scoliosis have attracted the attention of numerous researchers in recent years.
Epigenetics is defined as variations in gene expression that are not caused by DNA sequence alterations and are heritable in mitosis and meiosis (Delcuve et al., 2009). Key epigenetic mechanisms include DNA methylation, histone modification, non-coding RNA, and chromatin remodeling (Delcuve et al., 2009). Epigenetic alterations can regulate gene expression at various stages, including replication, transcription, and translation. Epigenetic changes are known to be involved in the development of cancer, neurodegenerative pathologies, and autoimmune diseases (Nebbioso et al., 2018; Zhang et al., 2020). Current research on AIS has indicated that genetic variations only explain approximately 2%–3% of the causative factors (Peng et al., 2020), which implies the involvement of other factors, such as epigenetic changes may play an important role in scoliosis. In this review, we summarize the current research on the relationship between epigenetics and the development of scoliosis; additionally, based on the four main mechanisms of epigenetic variation, we categorized the current studies and extracted the key changes, as well as the related mechanism hypothesis and regulation-pathways.
In this review, the terms “scoliosis,” “epigenetics,” “DNA methylation,” “histone modification,” “chromatin remodeling,” and “non-coding RNA” were used as keywords to find relative articles in PubMed, Embase, and Web of Science databases. This search was conducted until March 2023. The inclusion criteria were as follows: 1) clinical or basic research relevant to the above keywords; 2) papers published in journals with no language restrictions. The exclusion criteria were as follows: i) papers without full-text availability; ii) research with low relevance to the topic; iii) research with an unclear description of the original text or a relatively insufficient evidence-based rating. A total of 487 relevant papers were searched, and 25 papers were finally included in this review according to the selection criteria. Ten articles were related to DNA methylation, 13 related to noncoding RNA, 2 articles introduced histone modification, and we did not find article focused on the relationship between chromatin remodeling and scoliosis. The source of samples in each article were summarized in Table 1, 2. The literature search process is detailed in Figure 1.
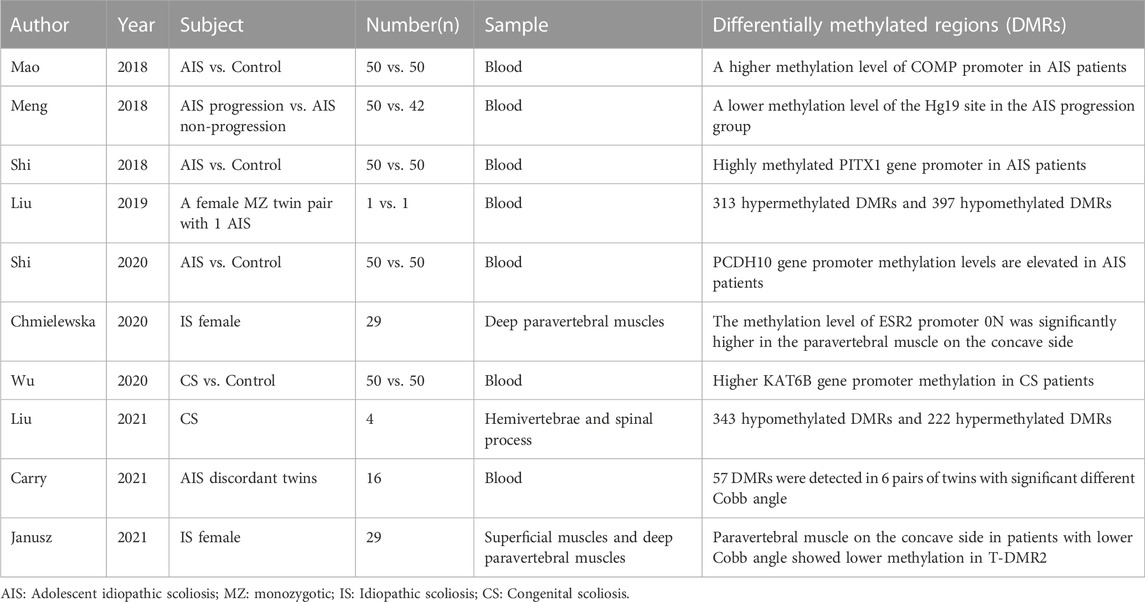
TABLE 1. The relationship between DNA methylation and scoliosis.
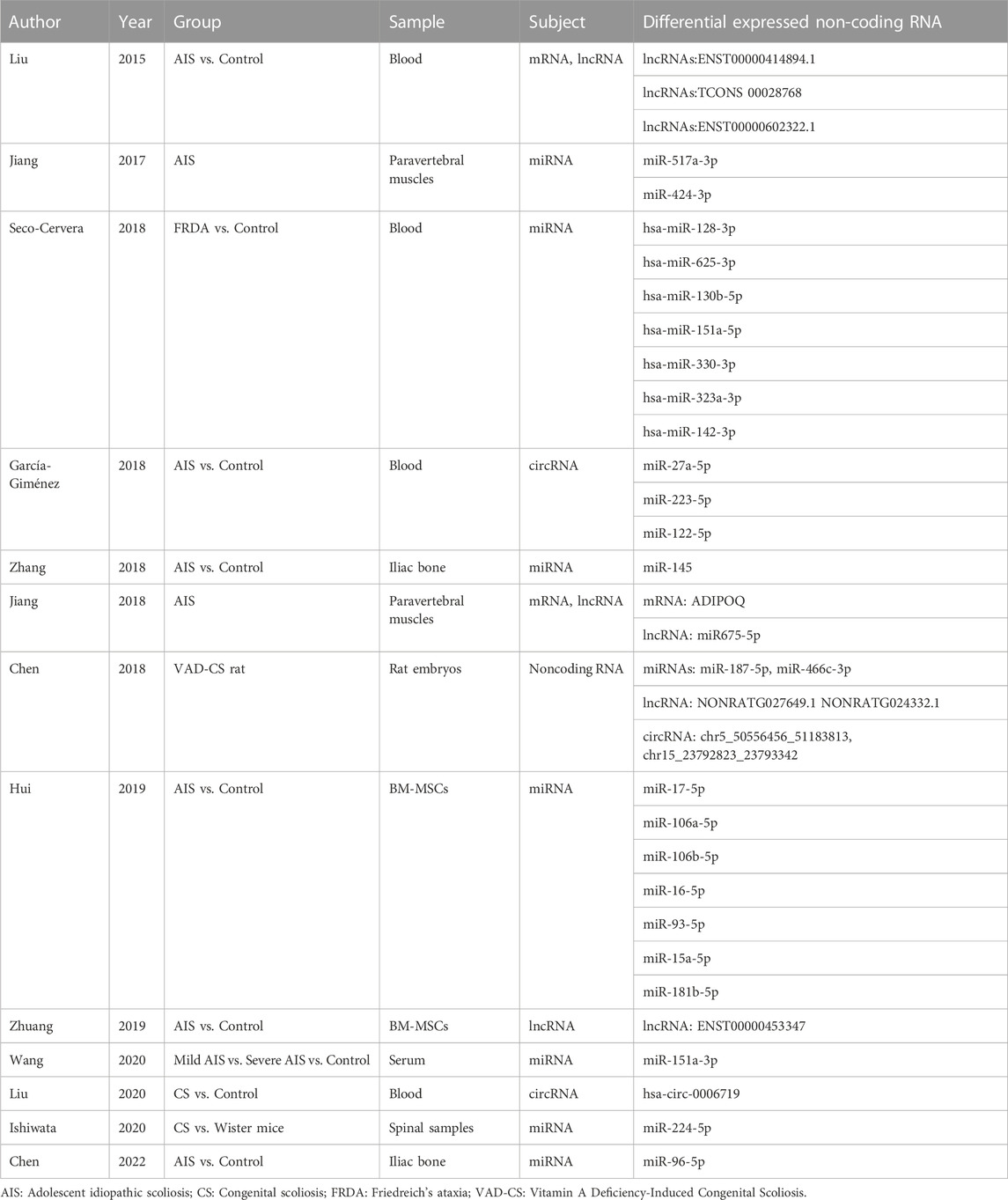
TABLE 2. Non-coding RNA and scoliosis.
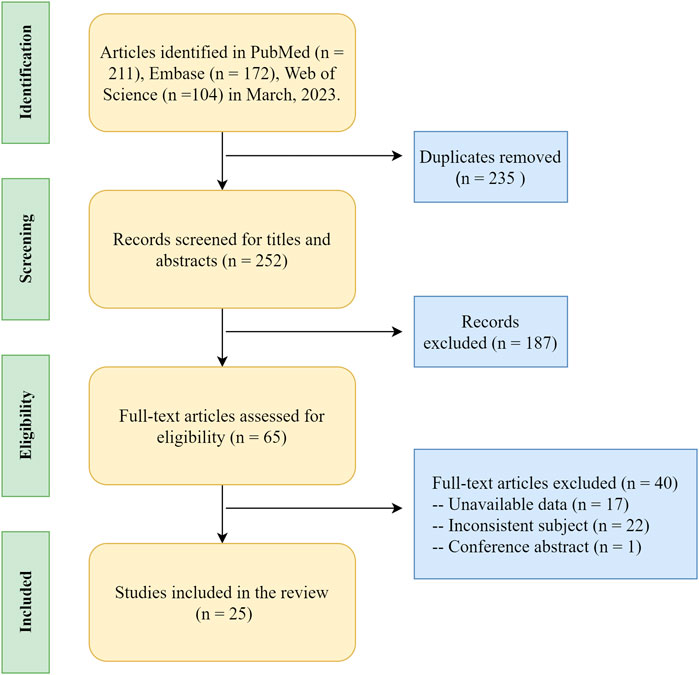
FIGURE 1. The literature search process on epigenetic research of scoliosis.
DNA methylation within the human genome typically occurs on the cytosine of the CpG dinucleotide, which is often located at the starting point of gene transcription (Delcuve et al., 2009). Under typical conditions, DNA methylation presents a dynamic equilibrium (Szyf, 2007), and silencing of selected genes according to physiological needs assists in controlling gene expression homeostasis (Miranda and Jones, 2007). In pathological states, abnormal DNA methylation can lead to dysregulated gene expression, which can cause abnormal expression of critical downstream products, triggering abnormal cell proliferation or apoptosis, thereby resulting in the development of scoliosis.
Deficiencies in critical components or enzymes of spinal growth and development can lead to spinal dysplasia and corresponding development or progression of scoliosis. Cartilage oligomeric matrix protein (COMP) is an extracellular matrix component essential for cartilage growth. Some studies have demonstrated that the secretion of COMP is significantly lower in patients with AIS than in normal controls (Gerdhem et al., 2015). Mao et al. examined the methylation and expression levels of COMP at five CpG loci within the COMP gene from patients with AIS and healthy controls (Mao et al., 2018). The level of COMP promoter methylation was significantly higher in patients with AIS than that in controls, whereas COMP expression was lower in patients with AIS than that in controls; additionally, the level of this abnormal methylation exhibited a positive correlation with an early onset of AIS and a larger Cobb angle. Overall, this revealed the relationship between abnormal DNA methylation and decreased COMP expression.
In 2018, Shi et al. established that the promoter of the PITX1 gene was hypermethylated, and the expression levels of downstream products were markedly reduced in patients with AIS compared to those in controls (Shi et al., 2018). The methylation levels at the six CpG sites on the PITX1 promoter region exhibited a correlation with the severity of scoliosis. PITX1 is a member of the RIEG/PITX homologous frame of transcription factors (Logan and Tabin, 1999), which act as a transcriptional regulator that participates in basal level regulation of prolactin, is involved in the process of hormonally regulated changes in prolactin activity; the abnormal expression of this gene has been found in many skeletal-related disorders (Alvarado et al., 2011; Pandey et al., 2012). Wu et al. found high levels of KAT6B gene promoter methylation and significantly reduced gene expression of KAT6B in patients with CS (Wu Y. et al., 2020). Correlation analysis identified a positive correlation between increased KAT6B methylation and an increased Cobb angle in these patients. Related research confirmed that the KAT6B gene was responsible for encoding part of the histone acetyltransferase and MOZ/MORF protein complexes (Tajul-Arifin et al., 2003). The MOZ/MORF protein plays a critical role in early skeletal and neuronal cell metabolism (Desh et al., 2014); therefore, aberrant DNA methylation at this site contributes to the development of CS. Taken together, this research demonstrated that the aberrant methylation of genes that play a crucial role in bone formation and development is an essential contributor to the development and progression of scoliosis.
The level of DNA methylation at key loci can also be applied as biomarkers to predict the progression of scoliosis. In 2018, Meng et al. determined that the methylation level at the cg01374129 locus was significantly lower in the AIS progression group than in the non-progression group (Meng et al., 2018). Through regression analysis, hypomethylation at this locus could be used as an independent prognostic factor for scoliosis exacerbation. Specifically, by assessing the degree of cg01374129 methylation, a sensitivity of 76.4% and specificity of 85.6% was achieved when differentiating between patients in progressive and non-progressive scoliosis groups, thereby suggesting that DNA methylation status had strong potential for use as a novel biomarker to determine prognosis. This locus is close to the gene encoding hyaluronan synthase 2 (HAS2), which plays an essential role in promoting intervertebral disc and vertebral body development in developmental rat models. Therefore, Roughley et al. suggested that abnormal methylation at the cg01374129 locus affects the progression of scoliosis via the disrupted development of the vertebral body and disc in these patients (Roughley et al., 2011).
Scoliosis progression can also be influenced by a variety of critical products of macroscopic pathways. Shi et al. found elevated PCDH10 gene promoter methylation and downregulated PCDH10 expression in patients with AIS compared to those in controls (Shi et al., 2020); additionally, these increased methylation levels were positively correlated with larger Cobb angles. PCDH10 is the target protein of p53, which regulates cell migration, but is not directly involved in cartilage development (Shi et al., 2015). However, as PCDH10 is involved in immunomodulatory-related roles in the Wnt-tyrosine kinase pathway, it has been postulated that in addition to directly affecting the development of bone growth, abnormal DNA methylation may also have an impact on the progression of scoliosis via the immune pathway. Overall, the studies discussed above evaluated the relationship between DNA methylation and scoliosis at a global level by analyzing peripheral blood specimens from patients with scoliosis.
Alternatively, other studies focused on DNA methylation alterations that are specific to skeletal muscle tissue around the spine of patients with scoliosis. In 2020, Chmielewska et al. examined DNA methylation levels in deep paravertebral muscle samples taken from the convex and concave sides of patients with AIS (Chmielewska et al., 2020). The methylation of estrogen receptor 2 (ESR2) promoter 0N in the paravertebral muscle was significantly higher on the concave side of the patient than that on the convex side. Corresponding correlation analysis revealed that ESR2 promoter 0N methylation variation was strongly associated with the development of AIS but was not significantly associated with the degree of curvature. This study was the first to elucidate a possible mechanism for the development of scoliosis from the perspective of aberrant local tissue DNA methylation. Janusz et al. (Janusz et al., 2021)assessed the deep paravertebral and superficial dorsal muscles of patients with idiopathic scoliosis to analyze the regulation of differentially methylated regions (T-DMRs) on the estrogen receptor 1 (ESR1) gene. ESR1 expression was determined to correlate with the methylation of CpGs in the T-DMR2 of the concave paravertebral muscle. The methylation of the four CpG islands in the T-DMR2 of the concave paravertebral muscle was markedly low in patients with less severe scoliosis (Cobb’s angle <70°). These results establish a correlation between aberrant local tissue methylation at key loci and the progression of scoliosis.
Differentially methylated regions (DMRs) at a range of loci often differ between patients with scoliosis and normal controls; therefore, in addition to aberrant methylation at key loci, attention has been provided to the impact of aberrant methylation sites across the genome. Since monozygotic (MZ) twins are genotypically identical but phenotypically different (Simony et al., 2016), the correlation between abnormal DNA methylation and the onset and progression of scoliosis can be more clearly explored by measuring DMRs in these twins. In 2019, Liu et al. utilized a pair of MZ twins with AIS within a study to find their DMRs, and further validate the role of these DMRs in 20 patients with AIS and controls (Liu et al., 2019). A total of 313 and 397 hypermethylated and hypomethylated DMRs, respectively, were identified in this study. The main regulation of the expression of these genes is attributed to the MAPK/PI3K-Akt pathway. Previous studies identified that the MAPK/PI3K-Akt signaling pathway is primarily associated with osteoblast differentiation and bone formation (Zhang et al., 2014; Iezaki et al., 2018). Additionally, Liu et al. reported the role of MAPK and other pathways in patients with CS (Liu et al., 2021). These DMRs control the downstream production of proteins predominantly found in the MAPK and calmodulin pathways, which are involved in the regulation of cytogenesis. Additionally, the calmodulin pathway directly regulates the osteogenesis process and affects the development of vertebral bodies in patients with scoliosis. In a study conducted by Carry et al. (Carry et al., 2021), eight pairs of MZ twins who exhibited scoliosis were enrolled. In six of these pairs of twins, large differences in Cobb angles were observed; 57 DMRs were detected within these twins, of which 28 DMRs exhibited a correlation with the degree of scoliosis. Of the DMRs, cg02477677 (within the vicinity of the RARA gene on chromosome 17), cg08826461 (corresponding to an unknown gene on chromosome 2), cg12922161 (within the LOC150622 gene on chromosome 2), and cg16382077 (corresponding to an unknown gene on chromosome 7) were the four most highly correlated DMRs observed in this study. Most of the corresponding gene products were enriched in the Wnt signaling pathway, which is associated with the production of neuropeptide Y (NPY). The Wnt signaling pathway has been previously reported to be important for bone formation and remodeling (Vink et al., 2014); additionally, NPY regulates bone and energy homeostasis (Wu J. Q. et al., 2020). Aberrant DNA methylation can thus contribute to the progression of scoliosis by affecting normal skeletal growth metabolism (Table 1).
In recent years, an increasing number of studies have demonstrated that non-coding RNAs play an essential role in the regulation of gene expression in the development of various diseases (Vondervoort et al., 2013; Jusic et al., 2020). The most common non-coding RNAs are long non-coding RNAs (lncRNAs), microRNAs (miRNAs), and circular RNAs (circRNAs). lncRNA strands are >200 nucleotides in length and are involved in transcriptional regulation, which has been observed to affect gene expression and participate in organismal regulation (Bridges et al., 2021). miRNAs are endogenously expressed non-coding transcripts that bind and silence corresponding mRNAs, thereby regulating protein expression (Hansen et al., 2013). circRNAs are non-coding RNAs with a stable circular structure. Studies found that circRNAs are rich in attachment sites to miRNAs and can affect gene expression by regulating the level of miRNAs (Hansen et al., 2013). The stability of the circular structure in circRNAs also enables them to be used as a stable biomarker that can be readily detected in the blood. (Garcia-Gimenez et al., 2018; Zhang et al., 2018).
Liu et al. revealed a series of 546 mRNAs with 139 lncRNAs differentially expressed in AIS children and controls (Liu et al., 2015). In particularly, ENST00000440778.1 was significantly under-expressed, and ENST00000414894.1, TCONS 00028768, and ENST00000602322.1 were significantly over-expressed in patients with AIS. The comparison between groups indicated that children with a high expression of ENST00000602322.1 had an early onset of AIS, while patients with low expression of ENST00000440778.1 had a high grade of Risser’s sign, and a high expression of ENST00000414894.1 conferred a Cobb angle >40°. Therefore, this study demonstrated the potential of lncRNAs as a predictor of disease.
Abnormal mesenchymal stem cell differentiation is a possible etiology for the formation of scoliosis. In 2019, Zhuang et al. identified a series of 1,483 lncRNAs that were differentially expressed in the bone marrow mesenchymal stem cells (BM-MSCs) of patients with AIS and normal controls (Zhuang et al., 2019). Specifically, ENST00000453347 was significantly under-expressed in patients with AIS. This lncRNA was named lncAIS, and the corresponding difference in expression was further verified by RT-PCR analysis of samples from patients and normal subjects. BM-MSCs with lncAIS knockdown exhibited impaired differentiation in in vitro cytological assays; moreover, this fraction of knocked-down cells failed to effectively form bone. This study further established that in normal humans, lncAIS interacted with NF90 to promote HOXD8 mRNA stability, thereby enhancing RUNX2 transcription in BM-MSCs and safeguarding the osteogenic differentiation of normal BM-MSCs. Within the AIS population, a significantly low expression of lncAIS causes impaired osteogenesis of the differentiated MSCs, resulting in the development of scoliosis (Zhuang et al., 2019).
Further, the RNA changes within the paravertebral muscle tissue around the spine are of equal concern. In 2018, Jiang et al. collected paired paravertebral muscle tissue specimens from patients with AIS on the convex and concave sides of the spine and identified a total of 40 DEGs on both sides of the paravertebral muscle (Jiang et al., 2018). Specifically, these genes were enriched in the peroxisome proliferator-activated receptor (PPAR) signaling pathway, with three mRNAs, ADIPOQ, FABP4, and MSTN, being markedly differentially expressed. Additionally, the lncRNA H19, encoded by the H19 gene, was observed to be significantly under-expressed in the concave paravertebral muscle tissue of patients with AIS, implying that this lncRNA plays an important role in the regenerative differentiation of skeletal muscle (Dey et al., 2014; Gao et al., 2014). Further, correlation analysis demonstrated that differential expression of ADIPOQ and H19 mRNA in both paravertebral muscles was associated with the early onset of AIS and high lateral convexity angle.
Studies utilizing animal models have also provided new insights. Chen et al. explored the expression and mechanism of action of the lncRNA SULT1C2A within a rat embryo model of vitamin A deficiency–induced scoliosis (VAD-CS) (Chen et al., 2019). SULT1C2A expression was determined to be downregulated, rno-miR-466c-5p expression was upregulated, and Foxo4, Pax1, Nkx3-2, and Sox9 somatic cell–associated genes were downregulated in VAD-CS embryos. Specifically, rno-miR-466c-5p could bind to the 3′-end untranslated region (3′-UTR) of Foxo4, thereby downregulating Foxo4 levels. Luciferase and siRNA assays revealed that SULT1C2A downregulates rno-miR-466c-5p expression through the PI3K-ATK signaling pathway, thereby increasing Foxo4 expression. Foxo4 is a forkhead box (Fox) transcription factor that participates in the development of cartilage and skeletal muscle during embryonic development (Matsuzaki et al., 2018). Therefore, it was suggested that increasing the expression of lncRNA SULT1C2A may be a possible treatment option for VAD-CS.
miRNA is a small type of RNA that can bind to mRNA to promote its expression or breakdown. Overall, miRNA contributes to the development of scoliosis through the action of signaling pathways. In 2017, Jiang Heng et al. selected muscle tissue from the convex and concave sides of the paravertebral vertebrae of three female patients with AIS (Jiang, 2017); corresponding RNA sequencing analysis identified a total of 18 differentially expressed miRNAs, with the convex side significantly overexpressing miR-517a-3p and the concave side overexpressing miR-424-3p. These two miRNAs are located on the mammalian miRNA gene cluster Gtl2-Dio3 and regulated by the transcription factor MEF2A. Therefore, it was concluded that these miRNAs act primarily through the Wnt signaling pathway and are associated with skeletal muscle regeneration, fibrosis, and cartilage development, which, in turn, contribute to paravertebral muscle imbalance and influence the onset and progression of AIS. Hui et al. also analyzed the differential miRNA expression observed in the BMCs of patients with AIS and normal subjects (Hui et al., 2019); a total of 54 previously unreported differential miRNAs were identified, with key functions in small GTPase-mediated signal transduction, DNA-dependent transcription, cytoplasmic division, cell adhesion, transmembrane transport, and hypoxic responses. Pathway analysis demonstrated that differentially expressed miRNAs were primarily concentrated in the MAPK, PI3K-Akt, calcium, Notch, and ubiquitin-mediated protein hydrolysis pathways. These pathways play an integral role in osteogenesis and adipose differentiation (Kanno et al., 2007; Baker et al., 2015), which are involved in the development of AIS. In this study, the seven differentially expressed miRNAs, including miR-17-5p, miR-106a-5p, miR-106b-5p, miR-16-5p, miR-93-5p, miR-15a-5p, and miR-181b-5p, were most correlated with the development of AIS and significantly associated with concomitant bone loss.
The differences of non-coding RNA changes in the bone tissue samples could better illustrate scoliosis pathogenesis. Zhang et al. analyzed miRNA microarrays in iliac bone tissue from patients with AIS and normal controls and identified miR-145 as a miRNA associated with AIS (Zhang et al., 2018). RT-qPCR confirmed that miR-145 was aberrantly upregulated in patients with AIS; additionally, aberrantly high expression of this miRNA was also found in bone cells from patients with AIS cultured in vitro. miR-145 expression was positively correlated with the expression of CTNNB1, which encodes β-catenin; further, continued activation of β-catenin may impair osteoblast function in patients with AIS. After knocking down the miR-145 gene in AIS osteoblasts, miR-145 expression was downregulated, the formation of an active β-catenin/Tcf4 complex was significantly inhibited, and normal expression of osteoblast markers, including osteoprotegerin (OPG), transmembrane glycoprotein E11, and sclerostin secretion (SOST), was restored. Additional specific disorders can accompany scoliosis. In 2018, Seco-Cervera et al. analyzed miRNA differences between patients with Friedreich’s ataxia (FRDA) and controls (Seco-Cervera et al., 2018). FRDA is a rare genetic disorder that causes progressive neurological impairment and motor problems; patients with FRDA often have scoliosis, heart disease, and diabetic complications (Filla et al., 1996). hsa-miR-128-3p, hsa-miR-625-3p, hsa-miR-130b-5p, hsa-miR-151a-5p, hsa-miR-330-3p, hsa-miR-323a-3p, and hsa-miR-142-3p were the most significant differentially expressed miRNAs, with these correlation results being validated by RT-qPCR. Of these, hsa-miR323a-3p could be used as a biomarker to discriminate patients with or without cardiac disease. However, this study did not further explore the interactions between these miRNAs and the corresponding mRNAs or the possible mechanisms influencing disease onset or progression.
The potential of miRNAs as biomarkers for identifying scoliosis is extensive. In 2020, Wang et al. evaluated five patients with mild AIS (Cobb angle >40°), five patients with severe AIS (Cobb angle >50°), and five healthy controls and sequenced their serum samples for miRNA (Wang et al., 2020). Corresponding results revealed that miR-941, miR-151a-3p, and miR-148b-5p were significantly elevated in the serum of the patients with AIS. When evaluating patients in the validation cohort, miR-151a-3p was determined to be significantly elevated in the sera of patients with severe scoliosis. After analysis of miRNA target genes, they ultimately established that GREM1 expression levels were significantly downregulated in osteoblasts from patients with severe AIS, thus indicating that miR-151a-3p may affect bone homeostasis and contribute to the development of AIS by binding to GREM1. miR-151a-3p could be used as a biomarker to predict severe AIS. In 2022, Chen et al. evaluated four patients with severe AIS and four age-matched controls (Chen et al., 2022); in this study, they extracted intraoperative iliac spine bone tissue for RNA microarray analysis. miR-96-5p expression was significantly higher in patients with AIS. An AIS prediction model combining serum miR-96-5p levels, patient age, age at menarche, and body weight was developed; an area under the ROC curve of 0.752 without the need for a bone mass scan demonstrated the corresponding potential of miR-96-5p as a biomarker for AIS.
Some studies have also used animal models to explore the role of miRNAs in scoliosis. Ishiwata et al. determined that miR-224-5p expression was greatly upregulated in the lumbar spine of a CS model (Ishibashi rats) compared to that in normal 42-day-old rats (Ishiwata et al., 2020). Further, a concomitant upregulation of miR-224-5p-regulated fibrinogen inhibitor-1 expression and an increase in type I collagen expression were also observed in this CS model. Overall, this finding indicated high osteoblast differentiation in the CS rat model. Further target gene analysis of miR-224-5p was not performed in this study; nonetheless, these results show that miR-224-5p can bind to target genes and contribute to the development of CS by affecting osteoblast differentiation. There are various effects of air pollution on embryonic congenital development (Choe et al., 2018), with the occurrence of CS being potentially closely linked to air pollution (Li et al., 2015). In 2019, Li et al. divided pregnant rats into randomized experimental and control groups, which were placed into environments with differing air qualities, indicated by PM2.5 (particulate matter with particles less than 2.5 μm in diameter) concentrations (Li Z. et al., 2019); specifically, experimental rats were placed in an environment with a PM2.5 > 200 μg/m³ and control rats were housed in an environment with a PM2.5 < 50 μg/m³. Embryos were removed for RNA sequencing on day 9 of pregnancy, and a total of 291 differentially expressed miRNAs were identified between the two groups, of which 204 miRNAs were upregulated and 87 miRNAs were downregulated in the experimental group. Convergent analysis suggested that these miRNAs were primarily involved in mitotic spindle organization, cellular respiration, ethanoic acid metabolism, and the proteasome, ultimately contributing to the development of CS.
In recent years, the possibility of utilizing specific circRNAs in the serum as diagnostic biomarkers for scoliosis-related diseases has been reported. García-Giménez et al. reported differences in the distribution of circRNA abundance between patients with AIS and normal controls (Garcia-Gimenez et al., 2018). They found that the abundance of three circRNAs, miR-122-5p, miR-27a-5p, and miR-223-5p, was significantly higher in patients with AIS than those in the normal population. These three circRNAs, together with miR-1306-3p, could be used as biomarkers to distinguish patients with AIS from normal controls. This method was validated in 30 patients with AIS compared to 13 normal subjects, with a corresponding area under the ROC curve of 0.95 and a high sensitivity, demonstrating the potential of using circRNA in combination with miRNA for AIS diagnosis. Further, Liu et al. identified a total of 22 differentially expressed circRNAs in patients with CS compared to controls (Liu et al., 2020); further, seven of these circRNAs in the validation group were confirmed by qPCR. Among them, hsa_circ_0006719 levels were highly elevated in the peripheral blood of patients with CS, and a prediction model based on this revealed an area under the ROC curve of 0.73, indicating a strong potential for the use of hsa_circ_0006719 as a diagnostic biomarker for CS.
Some studies have also focused on the overall variation of the three types of non-coding RNAs in patients with scoliosis. In 2018, Chen et al. sequenced non-coding RNAs with mRNAs in VAD-CS rat embryos (Chen et al., 2018). A total of 56 miRNAs, 70 circRNAs, and 685 lncRNAs were observed to be differentially expressed in VAD-CS and normal rat embryos. Some non-coding RNAs were further validated by RT-qPCR (miRNAs: miR-187-5p, miR-466c-3p; lncRNAs: NONRATG027649.1, NONRATG024332.1; circRNAs: chr5_50556456_51183813, chr15_23792823_23793342). Additionally, enrichment analysis demonstrated that these differentially expressed RNAs functioned in Foxo, PI3K-Akt, mTOR, EGFR, Wnt, and other signaling pathways.
In 2020, Li et al. searched PubMed, EMBASE, and GEO databases for literature that compared differences in gene, miRNA, and lncRNA expression between patients with AIS and normal control MSCs (Li et al., 2020); a computerized pooled analysis was then performed to establish a network of mRNA–miRNA–lncRNA interactions. A total of 1,027 DEGs (551 upregulated, 476 downregulated), 54 differentially expressed miRNAs (42 upregulated, 12 downregulated), and 658 differentially expressed lncRNAs (345 upregulated, 313 downregulated) were included in this review. Overall, a total of 6 mRNA–miRNA–lncRNA action networks were observed: RAP2C–AS1–miR-4419b–WDTC1, TSPEAR–AS2–miR-16-5p–CYCS/KDR/PDIA6/TGOLN2/HSPA5, TSPEAR–AS2/HCG18–miR-93-3p–CYCS/HSPA5, TSPEAR–AS2–AS2/HCG18–miR-93-5p–CYCS/TGOLN2, TSPEAR–AS2/HCG18–miR-615-3p–CKAP4 and TSPEAR–AS2/HCG18–miR-125a-3p–PGK1. These networks suggest that the non-coding RNAs may form their own reticulated regulatory system; however, these axes of action have only been analyzed in this study, and their accuracy needs to be confirmed by further experiments (Table 2).
Histone modification such as methylation and acetylation is an epigenetic modification that affects transcriptional activity (Delcuve et al., 2009). Mao et al. genotyped 500 patients with AIS and 494 age-matched controls using PCR-based Invader analysis (Mao et al., 2013). These results demonstrated that rs12459350, which regulates histone lysine 79 (H3K79) methylation, was strongly correlated with AIS susceptibility; however, this study did not further explore the specific mechanisms in this process.
In 2019, Li et al. performed histological and genetic testing of articular cartilage from 11 patients with IS compared to 10 matched controls (Li J. et al., 2019). Light microscopy of hematoxylin and eosin-stained samples revealed a significant increase in particular chondrocyte proliferation but a reduced articular cartilage thickness in the IS patient samples compared to those of the controls, suggesting that abnormal hyperplasia occurred in the samples taken from the patients with IS. Further examination revealed that the patients with IS exhibited increased expression of type II collagen and Bcl2 protein, which promote chondrocyte proliferation; therefore, it was concluded that the increased expression of Bcl2 was directly responsible for the abnormal cartilage proliferation. Examination of miRNA expression in articular cartilage tissue showed that patients had downregulated expression of a miRNA (miR-15a) that inhibits Bcl2, increased expression of the methyltransferase SUV39H1, and increased trimethylation of the miR-15a promoter region with histone H3K9. Thus, histone methylation may cause abnormal proliferation of chondrocytes through the miR-15a/Bcl2 signal axis, ultimately inducing abnormal spinal growth and leading to the development of scoliosis.
Chromosomes consist of nucleosomal units of DNA bound to histones, which enables the genome to be highly condensed (Clapier and Cairns, 2009). Chromatin remodeling complexes (remodelers) reorganize or disrupt nucleosomes through the energy released by ATP and are required for transcriptional expression, DNA replication, and DNA repair; further chromatin remodeling is largely dependent on these processes conferred by remodelers (Clapier and Cairns, 2009). This mechanism protects genes when important gene fragments are not functioning and controls the appropriate length of gene fragment exposure during replication and transcription (Clapier and Cairns, 2009). Chromatin reformulation is one of the key epigenetic mechanisms that control gene expression and has been implicated in a number of diseases, such as cerebro-oculo-facial-skeletal syndrome (Citterio et al., 2000) and Williams-Beuren syndrome (Kitagawa et al., 2003), alongside tumorigenesis and cell invasion behaviors in cancer (Denslow and Wade, 2007). However, no prior studies have reported the impact of chromatin remodeling on the pathogenesis of scoliosis; therefore, we hope that future studies explore this area further to provide a new understanding of epigenetics in scoliosis.
Epigenetic regulation plays an essential role in the pathogenesis of scoliosis. Consequently, recent studies have explored the relationship between scoliosis and epigenetics, with primary focus on two mechanisms: DNA methylation and non-coding RNA regulation. These mechanisms have specifically been evaluated by analyzing the changes in the expression of key products and their corresponding influence on signaling pathways, leading to a deeper understanding of the pathogenesis of scoliosis. However, the small number of studies that can be cross-examined may affect the corresponding depth of understanding within this review. Most studies analyzing epigenetics in scoliosis have not yet adopted methods such as downstream protein analysis to validate their proposed theories of pathway effects; therefore, there remains the requirement for further studies to deepen the understanding of DNA methylation and non-coding RNA regulation in scoliosis by testing the validity of current studies and further evaluating the mechanisms involved in this pathogenesis. Further, there are few studies on histone modification and chromatin remodeling, and the effects of chromatin remodeling on scoliosis are yet to be elucidated. Because the process of chromatin remodeling can be affected by a range of cellular behaviors, such as replication, transcription, and repair, the corresponding research is less reliable; therefore, developing an understanding of chromosome remodeling patterns specific to patients with scoliosis will likely be challenging. To conclude, epigenetic research remains an emerging field in scoliosis, and further studies are required to improve our current understanding of the pathogenesis of scoliosis and provide new insights into the future clinical diagnosis and treatment of this disorder.
All authors contributed to the review conception and design. Material preparation, data collection, and analysis were performed by DS and ZD. The first draft of the manuscript was written by DS and the work was critically revised by YH and YC. All authors contributed to the article and approved the submitted version.
This research is funded by Beijing Postdoctoral Work Funding Project (2022-ZZ-030).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Alvarado, D. M., Mccall, K., Aferol, H., Silva, M. J., Garbow, J. R., Spees, W. M., et al. (2011). Pitx1 haploinsufficiency causes clubfoot in humans and a clubfoot-like phenotype in mice. Hum. Mol. Genet. 20, 3943–3952. doi:10.1093/hmg/ddr313
Baker, N., Sohn, J., and Tuan, R. S. (2015). Promotion of human mesenchymal stem cell osteogenesis by PI3-kinase/Akt signaling, and the influence of caveolin-1/cholesterol homeostasis. Stem Cell Res. Ther. 6, 238. doi:10.1186/s13287-015-0225-8
Bridges, M. C., Daulagala, A. C., and Kourtidis, A. (2021). LNCcation: lncRNA localization and function. J. Cell Biol. 220, e202009045. doi:10.1083/jcb.202009045
Carry, P. M., Terhune, E. A., Trahan, G. D., Vanderlinden, L. A., Wethey, C. I., Ebrahimi, P., et al. (2021). Severity of idiopathic scoliosis is associated with differential methylation: An epigenome-wide association study of monozygotic twins with idiopathic scoliosis. Genes (Basel) 12, 1191. doi:10.3390/genes12081191
Chen, C., Tan, H., Bi, J., Li, L., Rong, T., Lin, Y., et al. (2019). LncRNA-SULT1C2A regulates Foxo4 in congenital scoliosis by targeting rno-miR-466c-5p through PI3K-ATK signalling. J. Cell Mol. Med. 23, 4582–4591. doi:10.1111/jcmm.14355
Chen, C., Tan, H., Bi, J., Li, Z., Rong, T., Lin, Y., et al. (2018). Identification of competing endogenous RNA regulatory networks in vitamin A deficiency-induced congenital scoliosis by transcriptome sequencing analysis. Cell Physiol. Biochem. 48, 2134–2146. doi:10.1159/000492556
Chen, H., Yang, K. G., Zhang, J., Cheuk, K. Y., Nepotchatykh, E., Wang, Y., et al. (2022). Upregulation of microRNA-96-5p is associated with adolescent idiopathic scoliosis and low bone mass phenotype. Sci. Rep. 12, 9705. doi:10.1038/s41598-022-12938-3
Chmielewska, M., Janusz, P., Andrusiewicz, M., Kotwicki, T., and Kotwicka, M. (2020). Methylation of estrogen receptor 2 (ESR2) in deep paravertebral muscles and its association with idiopathic scoliosis. Sci. Rep. 10, 22331. doi:10.1038/s41598-020-78454-4
Choe, S. A., Kauderer, S., Eliot, M. N., Glazer, K. B., Kingsley, S. L., Carlson, L., et al. (2018). Air pollution, land use, and complications of pregnancy. Sci. Total Environ. 645, 1057–1064. doi:10.1016/j.scitotenv.2018.07.237
Citterio, E., Van Den Boom, V., Schnitzler, G., Kanaar, R., Bonte, E., Kingston, R. E., et al. (2000). ATP-dependent chromatin remodeling by the Cockayne syndrome B DNA repair-transcription-coupling factor. Mol. Cell Biol. 20, 7643–7653. doi:10.1128/MCB.20.20.7643-7653.2000
Clapier, C. R., and Cairns, B. R. (2009). The biology of chromatin remodeling complexes. Annu. Rev. Biochem. 78, 273–304. doi:10.1146/annurev.biochem.77.062706.153223
Delcuve, G. P., Rastegar, M., and Davie, J. R. (2009). Epigenetic control. J. Cell Physiol. 219, 243–250. doi:10.1002/jcp.21678
Denslow, S. A., and Wade, P. A. (2007). The human Mi-2/NuRD complex and gene regulation. Oncogene 26, 5433–5438. doi:10.1038/sj.onc.1210611
Desh, H., Gray, S. L., Horton, M. J., Raoul, G., Rowlerson, A. M., Ferri, J., et al. (2014). Molecular motor MYO1C, acetyltransferase KAT6B and osteogenetic transcription factor RUNX2 expression in human masseter muscle contributes to development of malocclusion. Arch. Oral Biol. 59, 601–607. doi:10.1016/j.archoralbio.2014.03.005
Dey, B. K., Pfeifer, K., and Dutta, A. (2014). The H19 long noncoding RNA gives rise to microRNAs miR-675-3p and miR-675-5p to promote skeletal muscle differentiation and regeneration. Genes Dev. 28, 491–501. doi:10.1101/gad.234419.113
Fendri, K., Patten, S. A., Kaufman, G. N., Zaouter, C., Parent, S., Grimard, G., et al. (2013). Microarray expression profiling identifies genes with altered expression in Adolescent Idiopathic Scoliosis. Eur. Spine J. 22, 1300–1311. doi:10.1007/s00586-013-2728-2
Gao, Y., Wu, F., Zhou, J., Yan, L., Jurczak, M. J., Lee, H. Y., et al. (2014). The H19/let-7 double-negative feedback loop contributes to glucose metabolism in muscle cells. Nucleic Acids Res. 42, 13799–13811. doi:10.1093/nar/gku1160
Garcia-Gimenez, J. L., Rubio-Belmar, P. A., Peiro-Chova, L., Hervas, D., Gonzalez-Rodriguez, D., Ibanez-Cabellos, J. S., et al. (2018). Circulating miRNAs as diagnostic biomarkers for adolescent idiopathic scoliosis. Sci. Rep. 8, 2646. doi:10.1038/s41598-018-21146-x
Gerdhem, P., Topalis, C., Grauers, A., Stubendorff, J., Ohlin, A., and Karlsson, K. M. (2015). Serum level of cartilage oligomeric matrix protein is lower in children with idiopathic scoliosis than in non-scoliotic controls. Eur. Spine J. 24, 256–261. doi:10.1007/s00586-014-3691-2
Giampietro, P. F. (2012). Genetic aspects of congenital and idiopathic scoliosis. Sci. (Cairo) 2012, 152365. doi:10.6064/2012/152365
Hansen, T. B., Jensen, T. I., Clausen, B. H., Bramsen, J. B., Finsen, B., Damgaard, C. K., et al. (2013). Natural RNA circles function as efficient microRNA sponges. Nature 495, 384–388. doi:10.1038/nature11993
Hedequist, D., and Emans, J. (2007). Congenital scoliosis: A review and update. J. Pediatr. Orthop. 27, 106–116. doi:10.1097/BPO.0b013e31802b4993
Hui, S., Yang, Y., Li, J., Li, N., Xu, P., Li, H., et al. (2019). Differential miRNAs profile and bioinformatics analyses in bone marrow mesenchymal stem cells from adolescent idiopathic scoliosis patients. Spine J. 19, 1584–1596. doi:10.1016/j.spinee.2019.05.003
Iezaki, T., Fukasawa, K., Horie, T., Park, G., Robinson, S., Nakaya, M., et al. (2018). The MAPK Erk5 is necessary for proper skeletogenesis involving a Smurf-Smad-Sox9 molecular axis. Development 145, dev164004. doi:10.1242/dev.164004
Ishiwata, S., Iizuka, H., Sonoda, H., Tsunoda, D., Tajika, Y., Chikuda, H., et al. (2020). Upregulated miR-224-5p suppresses osteoblast differentiation by increasing the expression of Pai-1 in the lumbar spine of a rat model of congenital kyphoscoliosis. Mol. Cell Biochem. 475, 53–62. doi:10.1007/s11010-020-03859-8
Janusz, P., Chmielewska, M., Andrusiewicz, M., Kotwicki, T., and Kotwicka, M. (2021). Methylation level of the regulatory regions of the estrogen receptor type 1 gene in paravertebral muscles of girls with idiopathic scoliosis. Stud. health Technol. Inf. 280, 254. doi:10.3233/SHTI210484
Jiang, H. (2017). Study on the imbalance of paravertebral muscles on both sides of adolescent idiopathic scoliosis and its formation mechanism. Shanghai, China: Chinese People's Liberation Army Naval Medical University. doi:10.7666/d.Y3247105
Jiang, H., Yang, F., Lin, T., Shao, W., Meng, Y., Ma, J., et al. (2018). Asymmetric expression of H19 and ADIPOQ in concave/convex paravertebral muscles is associated with severe adolescent idiopathic scoliosis. Mol. Med. 24, 48. doi:10.1186/s10020-018-0049-y
Jusic, A., Devaux, Y., and Cardiology, E. (2020). Mitochondrial noncoding RNA-regulatory network in cardiovascular disease. Basic Res. Cardiol. 115, 23. doi:10.1007/s00395-020-0783-5
Kanno, T., Takahashi, T., Tsujisawa, T., Ariyoshi, W., and Nishihara, T. (2007). Mechanical stress-mediated Runx2 activation is dependent on Ras/ERK1/2 MAPK signaling in osteoblasts. J. Cell Biochem. 101, 1266–1277. doi:10.1002/jcb.21249
Kitagawa, H., Fujiki, R., Yoshimura, K., Mezaki, Y., Uematsu, Y., Matsui, D., et al. (2003). Retracted: The chromatin-remodeling complex WINAC targets a nuclear receptor to promoters and is impaired in Williams syndrome. Cell 113, 905–917. doi:10.1016/s0092-8674(03)00436-7
Li, H. M., Liu, Y., Ding, J. Y., Zhang, R., Liu, X. Y., and Shen, C. L. (2020). In silico analysis excavates A novel competing endogenous RNA subnetwork in adolescent idiopathic scoliosis. Front. Med. (Lausanne) 7, 583243. doi:10.3389/fmed.2020.583243
Li, J., Yang, G., Liu, S., Wang, L., Liang, Z., and Zhang, H. (2019a). Suv39h1 promotes facet joint chondrocyte proliferation by targeting miR-15a/Bcl2 in idiopathic scoliosis patients. Clin. Epigenetics 11, 107. doi:10.1186/s13148-019-0706-1
Li, Z., Ma, J., Bi, J., Guo, H., Chan, M. T. V., Wu, W. K. K., et al. (2019b). MicroRNA signature of air pollution exposure-induced congenital defects. J. Cell Physiol. 234, 17896–17904. doi:10.1002/jcp.28422
Li, Z., Yu, X., and Shen, J. (2015). Environmental aspects of congenital scoliosis. Environ. Sci. Pollut. Res. Int. 22, 5751–5755. doi:10.1007/s11356-015-4144-0
Liu, G., Shen, J., Chen, C., Jiao, Y., Li, Z., Tan, H., et al. (2020). Genome-Wide Analysis of circular RNAs and validation of hsa_circ_0006719 as a potential novel diagnostic biomarker in congenital scoliosis patients. J. Cell Mol. Med. 24, 7015–7022. doi:10.1111/jcmm.15370
Liu, G., Wang, L., Wang, X., Yan, Z., Yang, X., Lin, M., et al. (2019). Whole-genome methylation analysis of phenotype discordant monozygotic twins reveals novel epigenetic perturbation contributing to the pathogenesis of adolescent idiopathic scoliosis. Front. Bioeng. Biotechnol. 7, 364. doi:10.3389/fbioe.2019.00364
Liu, G., Zhao, H., Yan, Z., Zhao, S., Niu, Y., Li, X., et al. (2021). Whole-genome methylation analysis reveals novel epigenetic perturbations of congenital scoliosis. Mol. Ther. Nucleic Acids 23, 1281–1287. doi:10.1016/j.omtn.2021.02.002
Liu, X. Y., Wang, L., Yu, B., Zhuang, Q. Y., and Wang, Y. P. (2015). Expression signatures of long noncoding RNAs in adolescent idiopathic scoliosis. Biomed. Res. Int. 2015, 276049. doi:10.1155/2015/276049
Logan, M., and Tabin, C. J. (1999). Role of Pitx1 upstream of Tbx4 in specification of hindlimb identity. Science 283, 1736–1739. doi:10.1126/science.283.5408.1736
Mao, S. H., Qian, B. P., Shi, B., Zhu, Z. Z., and Qiu, Y. (2018). Quantitative evaluation of the relationship between COMP promoter methylation and the susceptibility and curve progression of adolescent idiopathic scoliosis. Eur. Spine J. 27, 272–277. doi:10.1007/s00586-017-5309-y
Mao, S., Xu, L., Zhu, Z., Qian, B., Qiao, J., Yi, L., et al. (2013). Association between genetic determinants of peak height velocity during puberty and predisposition to adolescent idiopathic scoliosis. Spine (Phila Pa 1976) 38, 1034–1039. doi:10.1097/BRS.0b013e318287fcfd
Matsuzaki, T., Alvarez-Garcia, O., Mokuda, S., Nagira, K., Olmer, M., Gamini, R., et al. (2018). FoxO transcription factors modulate autophagy and proteoglycan 4 in cartilage homeostasis and osteoarthritis. Sci. Transl. Med. 10, eaan0746. doi:10.1126/scitranslmed.aan0746
Meng, Y., Lin, T., Liang, S., Gao, R., Jiang, H., Shao, W., et al. (2018). Value of DNA methylation in predicting curve progression in patients with adolescent idiopathic scoliosis. EBioMedicine 36, 489–496. doi:10.1016/j.ebiom.2018.09.014
Miranda, T. B., and Jones, P. A. (2007). DNA methylation: The nuts and bolts of repression. J. Cell Physiol. 213, 384–390. doi:10.1002/jcp.21224
Nebbioso, A., Tambaro, F. P., Dell'aversana, C., and Genetics, L. (2018). Cancer epigenetics. Mov. Forw. 14, e1007362. doi:10.1371/journal.pgen.1007362
Pahys, J. M., and Guille, J. (2018). What's new in congenital scoliosis? J. Pediatr. Orthop. 38, e172–e179. doi:10.1097/bpo.0000000000000922
Pandey, S. N., Cabotage, J., Shi, R., Dixit, M., Sutherland, M., Liu, J., et al. (2012). Conditional over-expression of PITX1 causes skeletal muscle dystrophy in mice. Biol. Open 1, 629–639. doi:10.1242/bio.20121305
Peng, Y., Wang, S. R., Qiu, G. X., Zhang, J. G., and Journal, Q. (2020). Research progress on the etiology and pathogenesis of adolescent idiopathic scoliosis. Chin. Med. J. Engl. 133, 483–493. doi:10.1097/cm9.0000000000000652
Qiu, Y. (2018). Current status and prospects of genetic research and clinical treatment of adolescent idiopathic scoliosis. Zhonghua Wai Ke Za Zhi 56, 578–582. doi:10.3760/cma.j.issn.0529-5815.2018.08.005
Roughley, P. J., Lamplugh, L., Lee, E. R., Matsumoto, K., and Yamaguchi, Y. (2011). The role of hyaluronan produced by Has2 gene expression in development of the spine. Spine (Phila Pa 1976) 36, E914–E920. doi:10.1097/BRS.0b013e3181f1e84f
Seco-Cervera, M., Gonzalez-Rodriguez, D., Ibanez-Cabellos, J. S., Peiro-Chova, L., Pallardo, F. V., and Garcia-Gimenez, J. L. (2018). Small RNA-seq analysis of circulating miRNAs to identify phenotypic variability in Friedreich's ataxia patients. Sci. Data 5, 180021. doi:10.1038/sdata.2018.21
Shi, B., Mao, S., Xu, L., Li, Y., Sun, X., Liu, Z., et al. (2020). Quantitation analysis of PCDH10 methylation in adolescent idiopathic scoliosis using pyrosequencing study. Spine (Phila Pa 1976) 45, E373–E378. doi:10.1097/BRS.0000000000003292
Shi, B., Xu, L., Mao, S., Xu, L., Liu, Z., Sun, X., et al. (2018). Abnormal PITX1 gene methylation in adolescent idiopathic scoliosis: A pilot study. BMC Musculoskelet. Disord. 19, 138. doi:10.1186/s12891-018-2054-2
Shi, D., Murty, V. V., and Gu, W. (2015). PCDH10, a novel p53 transcriptional target in regulating cell migration. Cell Cycle 14, 857–866. doi:10.1080/15384101.2015.1004935
Simony, A., Carreon, L. Y., K, H. J., Kyvik, K. O., and Andersen, M. O. (2016). Concordance rates of adolescent idiopathic scoliosis in a Danish twin population. Spine (Phila Pa 1976) 41, 1503–1507. doi:10.1097/BRS.0000000000001681
Szyf, M. (2007). The dynamic epigenome and its implications in toxicology. Toxicol. Sci. 100, 7–23. doi:10.1093/toxsci/kfm177
Tajul-Arifin, K., Teasdale, R., Ravasi, T., Hume, D. A., Mattick, J. S., Group, R. G., et al. (2003). Identification and analysis of chromodomain-containing proteins encoded in the mouse transcriptome. Genome Res. 13, 1416–1429. doi:10.1101/gr.1015703
Vialle, R., Thevenin-Lemoine, C., and Mary, P. (2013). Neuromuscular scoliosis. Orthop. Traumatol. Surg. Res. 99, S124–S139. doi:10.1016/j.otsr.2012.11.002
Vink, C. P., Ockeloen, C. W., Ten Kate, S., Koolen, D. A., Ploos Van Amstel, J. K., Kuijpers-Jagtman, A. M., et al. (2014). Variability in dentofacial phenotypes in four families with WNT10A mutations. Eur. J. Hum. Genet. 22, 1063–1070. doi:10.1038/ejhg.2013.300
Vondervoort, I., Gordebeke, P. M., Khoshab, N., Tiesinga, P. H. E., Buitelaar, J. K., Kozicz, T., et al. (2013). Long non-coding RNAs in neurodevelopmental disorders. Front. Mol. Neurosci. 6, 53. doi:10.3389/fnmol.2013.00053
Wang, Y., Zhang, H., Yang, G., Xiao, L., Li, J., and Guo, C. (2020). Dysregulated bone metabolism is related to high expression of miR-151a-3p in severe adolescent idiopathic scoliosis. Biomed. Res. Int. 2020, 4243015. doi:10.1155/2020/4243015
Weinstein, S. L., Dolan, L. A., Cheng, J. C., Danielsson, A., and Morcuende, J. A. (2008). Adolescent idiopathic scoliosis. Lancet 371, 1527–1537. doi:10.1016/S0140-6736(08)60658-3
Wishart, B. D., Medicine, E. K. J. P., and America, R. C. O. N. (2021). Neuromuscular scoliosis: When, who, why and outcomes. Phys. Med. Rehabil. Clin. N. Am. 32, 547–556. doi:10.1016/j.pmr.2021.02.007
Wu, J. Q., Jiang, N., and Yu, B. (2020a). Mechanisms of action of neuropeptide Y on stem cells and its potential applications in orthopaedic disorders. World J. Stem Cells 12, 986–1000. doi:10.4252/wjsc.v12.i9.986
Wu, Y., Zhang, H., Tang, M., Guo, C., Deng, A., Li, J., et al. (2020b). High methylation of lysine acetyltransferase 6B is associated with the Cobb angle in patients with congenital scoliosis. J. Transl. Med. 18, 210. doi:10.1186/s12967-020-02367-z
Zhang, J., Chen, H., Leung, R. K. K., Choy, K. W., Lam, T. P., Ng, B. K. W., et al. (2018). Aberrant miR-145-5p/beta-catenin signal impairs osteocyte function in adolescent idiopathic scoliosis. FASEB J. 32, fj201800281. doi:10.1096/fj.201800281
Zhang, L., Lu, Q., and Chang, C. (2020). Epigenetics in Health and disease. Adv. Exp. Med. Biol. 1253, 3–55. doi:10.1007/978-981-15-3449-2_1
Zhang, Y., Pizzute, T., and Pei, M. (2014). A review of crosstalk between MAPK and Wnt signals and its impact on cartilage regeneration. Cell Tissue Res. 358, 633–649. doi:10.1007/s00441-014-2010-x
Keywords: epigenetics, biomarkers, signaling pathways, adolescent idiopathic scoliosis, congenital scoliosis
Citation: Sun D, Ding Z, Hai Y and Cheng Y (2023) Advances in epigenetic research of adolescent idiopathic scoliosis and congenital scoliosis. Front. Genet. 14:1211376. doi: 10.3389/fgene.2023.1211376
Received: 24 April 2023; Accepted: 30 June 2023;
Published: 26 July 2023.
Edited by:
Chenyu Sun, AMITA Health, United StatesReviewed by:
Peikai Chen, The University of Hong Kong, ChinaCopyright © 2023 Sun, Ding, Hai and Cheng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yong Hai, eW9uZy5oYWlAY2NtdS5lZHUuY24=
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.