 Pengpeng Liu1†
Pengpeng Liu1† Zhe Zhang2†
Zhe Zhang2† Yuruo Yin2†Shanshan Yan2
Yuruo Yin2†Shanshan Yan2 Yong Ren3Wei Sang1Hongjun Xu1Xinnian Han1Fengjuan Cui1
Yong Ren3Wei Sang1Hongjun Xu1Xinnian Han1Fengjuan Cui1 Yingbin Nie1Dezhen Kong1Wei Li1
Yingbin Nie1Dezhen Kong1Wei Li1 Caixia Lan2*
Caixia Lan2* Peiyuan Mu1*
Peiyuan Mu1*- 1Institute of Crop Research, Xinjiang Academy of Agri-Reclamation Sciences, Key Lab of Xinjiang Production and Construction Corps for Cereal Quality Research and Genetic Improvement, Shihezi, Xinjiang, China
- 2National Key Laboratory of Crop Genetic Improvement, Hubei Hongshan Laboratory, College of Plant Science and Technology, Huazhong Agricultural University, Wuhan, Hubei, China
- 3Mianyang Institute of Agricultural Science, Crop Characteristic Resources Creation and Utilization Key Laboratory of Sichuan Province, Mianyang City, Sichuan, China
In order to understand the difference of quality for Chinese and CIMMYT wheat varieties (lines), we selected 153 wheat germplasm from both China and CIMMYT to explore the contribution relationship of different allelic variation combinations to wheat quality through genotyping and phenotyping, including grain hardness, polyphenol oxidase (PPO) activity, lipoxygenase (LOX) activity, yellow pigment (YP) content and protein content. In terms of flour milling quality, Chinese wheat varieties were mainly carrying Pina-D1a/Pinb-D1b, accounting for 32.0% of the total tested varieties, while the CIMMYT wheat lines were mainly carrying Pina-D1b/Pinb-D1a with 45.8% of the total collection. The distribution frequencies of subunit 1/2* and 5 + 10 were 47.0% and 42.5%, respectively, in CIMMYT varieties, however they were only 31.4% and 13.7% respectively of the Chinese wheat tested varieties. In addition, the proportion of phytoene synthase (PSY) allele, PPO allele and LOX active allele were roughly the same between Chinese and CIMMYT varieties. Based on the present study, we found that Pina gene had a greater impact on grain hardness value than Pinb gene; The influence of PPO-A1 gene on polyphenol oxidase activity was more significant than PPO-D1 gene. The high protein content of varieties mostly containing hardness genes and 1/2*/5 + 10 subunit combinations. Based on the present study, we found that the quality gene distribution of Chinese and CIMMYT varieties was quite different, for instance, the high-quality HMW-GS subunits of Chinese varieties were lower than CIMMYT lines. It will be much useful for Chinese wheat breeders to develop good quality wheat variety by crossing with 3 good strong gluten CIMMYT wheat lines by molecular marker-assisted selection.
Introduction
Wheat is one of the most important food crops in the world. China is the largest wheat producer and consumer worldwide, accounting for about 1/6 of the global wheat yield production. Although China has high production and inventories of wheat, there is still a large gap of good quality wheat resulting in a huge import (Sun et al., 2017; Wang et al., 2019). With the improvement of people’s living standards, the demand of good quality bread continues to grow, leading to structural imbalances between supply and demand. Therefore, it is necessary to accelerate the improvement of wheat quality through molecular marker-assisted analysis of quality genes and phenotype of quality traits.
Wheat quality was mainly affected by genetics. For example, wheat grain hardness is regulated by a pair of major genes Ha, which is controlled by Pina and Pinb (Pins) genes, respectively. The loss of Pina protein or the mutation of Pinb protein gene can cause the change of wheat from soft to hard (Chen et al., 2010). Grain hardness is one of the key traits that determine wheat milling quality. In addition, it affects wheat flour yield, moisture content and processing quality, etc. (Morris, 2002; Chen et al., 2005; Liu et al., 2015). High molecular weight glutenin subunits (HMW-GS) are one of the measurement standards of processing quality. Its content in grains is low, but it has a significant impact on quality, such as whiteness, wet gluten content, water absorption and stability (Zhang et al., 2019). Wheat HMW-GS is encoded by the Glu-1 locus on the long arms of the wheat chromosomes 1A, 1B and 1D, which are Glu-A1, Glu-B1 and Glu-D1, respectively. Each site produces two subunits, named as x and y subunits (Zhang et al., 2016). The high-quality subunits, namely, Ax1 or Ax2*(1/2*), Dx5+Dy10 (5 + 10) and their combinations have a positive effect on the strength of gluten (Payne, 1987; Cooper et al., 2015; Vancini et al., 2019; Guzmán et al., 2022; Sönmez et al., 2023).
Wheat yellow pigment (YP), polyphenol oxidase (PPO) activity, and grain lipoxygenase (LOX) activity mainly affect the quality of wheat flour. In China, the color of flour is a crucial trait of wheat quality evaluation. Phytoene synthase (Psy) is a key enzyme of yellow pigment in flour, which plays a significant role in yellow pigment in flour. Previous studies showed that one of the major QTL on wheat chromosome 7A showed significant association with yellow pigment content, which can explain 60% of YP phenotypic variation (Kuchel et al., 2006). PPO activity is related to browning and impacts the whiteness of flour. High PPO activity is able to cause browning in flour processing and storage resulting in reduction the color of flour, while low PPO activity can increase white color of flour. The major genes that control the PPO activity of wheat grains were located on chromosomes 2A, 2B, and 2D (Zhang et al., 2006). Furthermore, the genes on chromosomes 1AL and 4BS have a major effect upon wheat LOX activity (Geng et al., 2012). LOX can couple and oxidize the carotenoids in wheat flour, which can make wheat flour white. Notwithstanding, excessive LOX activity will destroy the yellow pigment of wheat, making wheat flour too white and losing many nutrients (Han et al., 2015).
At present, the low-quality wheat inventory is high, while the high-quality wheat gap is large. It has become an important issue to explore the quality differences of core germplasm at home and abroad, accelerate the cultivation of high-quality wheat, and improve the structural supply-demand relationship in China. At the present study, we collected 153 wheat germplasm from Chinese and CIMMYT and genotyped with molecular makers of known genes and phenotyped them with the related traits. The objectives of this study are 1) to analyze the differences of wheat quality between Chinese and CIMMYT wheat germplasm; 2) to identify good quality wheat germplasm for wheat breeder to develop new wheat varieties; 3) to figure out the best quality gene combinations for achieving good quality wheat varieties.
Results
Genotypes of wheat quality related genes for a germplasm collection
Based on the previous studies, we collected most of the wheat quality related molecular markers and summarized them in the Supplementary Table S1. We used all of these molecular markers to genotype 153 wheat collection from China and CIMMYT (Figure 1; Supplementary Table S2). The result showed that the frequency of 1BL/1RS translocations was 17.6%, while it was 41.8% and 62.1% of alleles Ppo-A1b and Ppo-D1a, respectively, those were correlated with the lower PPO activity. The frequencies of Psy-A1b and Zds-A1a were 32.7% and 31.4%, respectively, and they were associated with the lower YP content. The proportions of hardness genes, Pina-D1b and Pinb-D1b, were 48.4% and 32.7%, respectively, while LOX-B1b was accounted for 78.4% that was relevant to the lower LOX activity. The frequencies of 1/2* and 5 + 10 were 78.4% and 56.2%, respectively, association with the HMW-GS subunits (Figure 2).
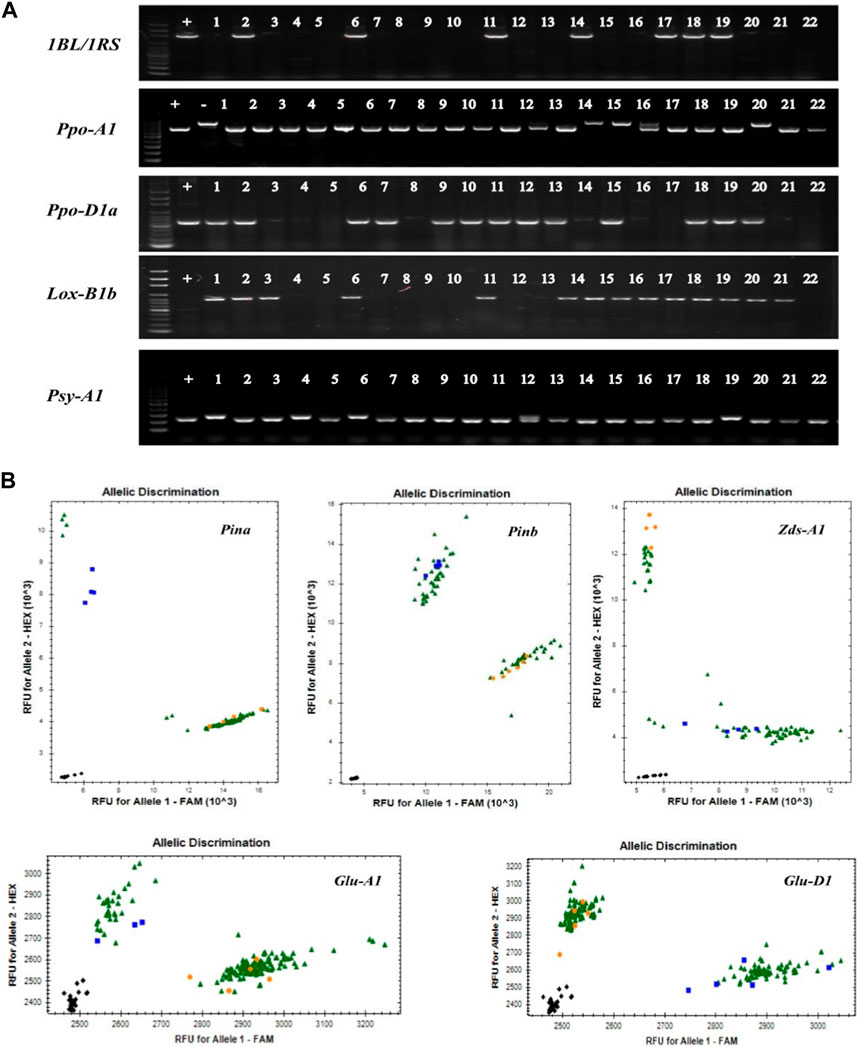
FIGURE 1. Part of PCR amplification of wheat cultivars. The detection results of partial materials with quality genes display (A) Common PCR results. +, Positive control; −, Negative control; 1–22, Test materials; (B) Kasp detection results of Glu-A1 and Glu-D1. Black, ddH2O; Blue and Orange, control varieties; Green, test material.
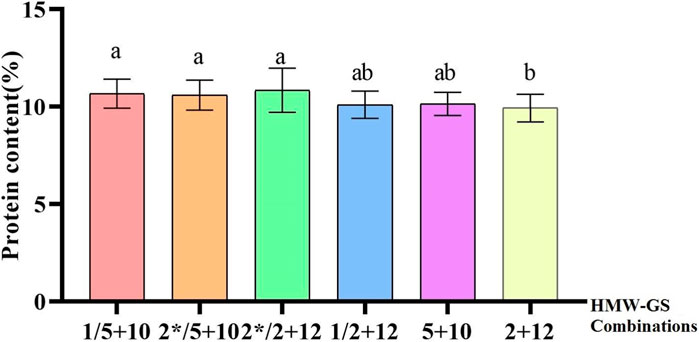
FIGURE 2. Known quality gene distribution of 153 materials based on the functional molecular markers.
Distribution of allelic variants among the collection
In terms of appearance quality, the distribution frequency of Chinese and CIMMYT wheat varieties is roughly the same among different genes or subunit combinations. There were four different types of allelic variants in the two major genes, Ppo-A1 and Ppo-D1 that control the activity of PPO. The PPO activity of the combination containing Ppo-A1a gene was markedly higher than that of other variant types. The activity of Ppo-A1b/Ppo-D1a was significantly lower than that of Ppo-A1a/Ppo-D1b, Ppo-A1a/Ppo-D1a, Ppo-A1b/Ppo-D1b. PPO activity varied greatly among different wheat varieties, and it was ranged from 114.7 to 898.0 and from 90.4 to 756.9 across 2-year data (Table 1). The variation range of yellow pigment content was 0.94–3.47 and 0.93–2.91 over 2 years, while there was no significant difference of YP content trend between different genotypes. The Psy-A1a/Zds-A1b gene combination that controls the yellow pigment content was obviously higher than that of Psy-A1b/Zds-A1b and Zds-A1a/Psy-A1b (Table 2). In addition, the LOX activity was significantly higher for wheat varieties containing LOX-B1a gene than that of LOX-B1b (Table 3).
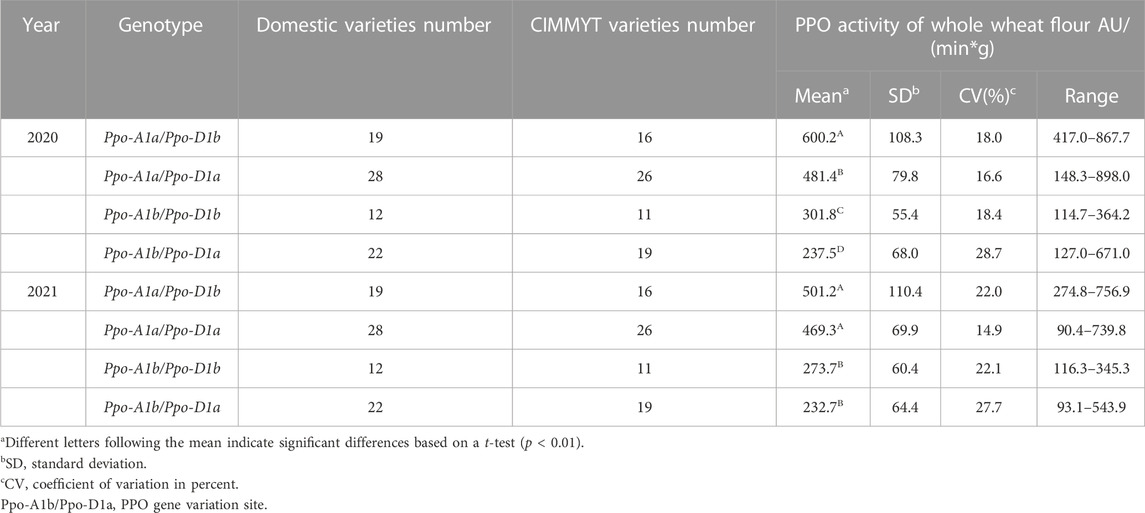
TABLE 1. Comparison the PPO activities among different genotypes.
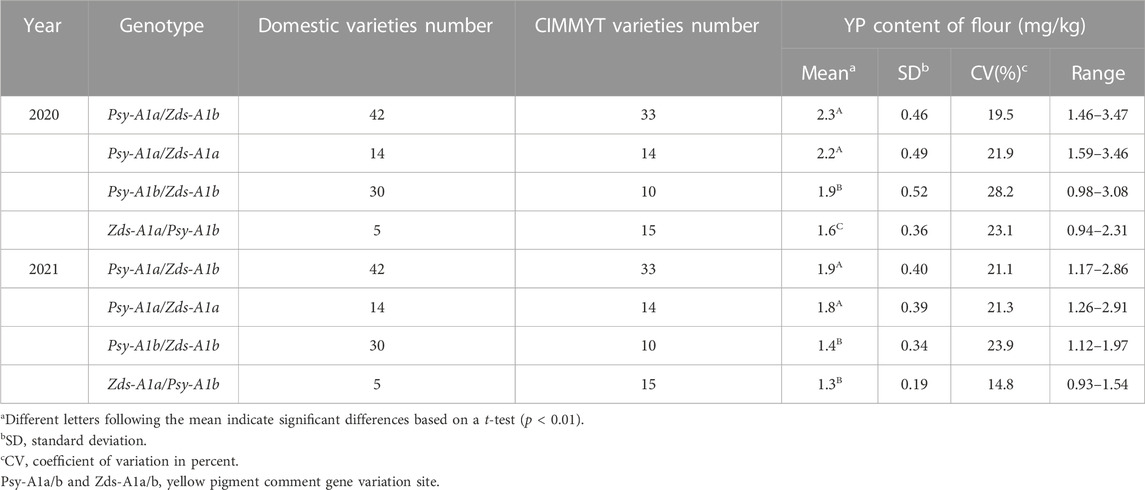
TABLE 2. Comparison the YP content among different genotypes.
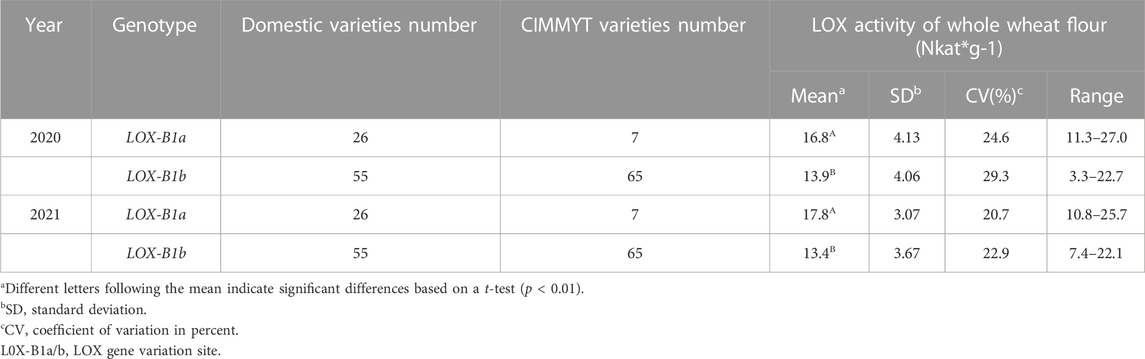
TABLE 3. Comparison the LOX activities among different genotypes.
Identification and origin of good wheat quality germplasm among the collection
In terms of grain hardness, we identified 46 hard wheat varieties with grain hardness higher than 70 and 30 soft wheat varieties with grain hardness less than 50, and they carry gene combinations of Pina-D1b/Pinb-D1a and Pina-D1a/Pinb-D1a, respectively. Among the 46 hard wheat varieties, 38 were from CIMMYT materials and 8 were from Chinese materials; 30 soft wheat germplasm were all Chinese materials (Figure 3A; Supplementary Tables S3, S4). As for protein content, we found five strong gluten wheat with protein content higher than 14% and 35 weak gluten wheat with protein content lower than 10%, while they carry high molecular weight Glutelin subunit combinations of 1Ax1/1Dx5+1Dy10 and 1Dx2+1Dy12, respectively. Among the 5 strong gluten wheat varieties, 4 were from CIMMYT and 1 was from China. Among the 35 weak gluten wheat varieties, 15 were from CIMMYT and 20 were from China (Figure 3B; Supplementary Table S5). In addition, we identified 3 Chinese wheat varieties with more than 3 μg/g of high yellow pigment content with genotype combinations of Psy-A1a/Zds-A1 (Figure 3C). In terms of polyphenol oxidase, we found 21 low polyphenol oxidase activity materials with polyphenol oxidase activity lower than 200 U/(g.min), and their genotype combinations were Ppo-A1b/Ppo-D1b or Ppo-A1b/Ppo-D1a (Figure 3D). Among the 21 accessions, 15 were from CIMMYT and 6 were from China. For lipoxygenase activity, 17 low lipoxygenase active accessions with lipoxygenase activity below 10 AU/(min g) were selected, and their genotype was Lox-B1b. Among them, 15 were from CIMMYT and 2 were from China (Figure 3E). On the part of the 1BL/1RS translocation line, a total of 28 accessions contained this translocation and all of them were Chinse accessions. The germplasm carrying this translocation showed significantly reduced grain hardness, increased yellow pigment content and lipoxygenase activity, and reduced quality (Table 4).
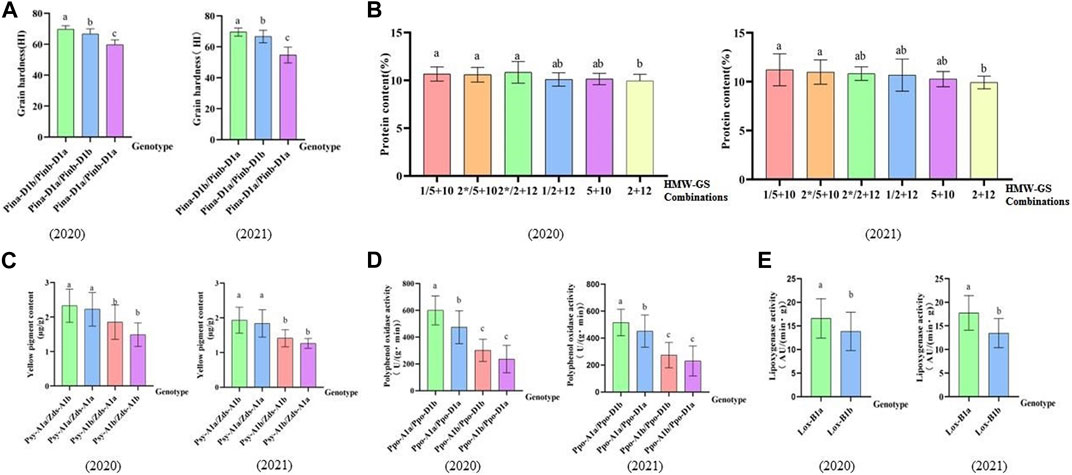
FIGURE 3. Effects of different genotypes on related quality traits. (A–E) Bar plots display different genotypes on quality through analyzing 2020, 2021data. (A) Bar plot shows the effect of different genotype combinations on grain hardness. (B) Bar plot indicates the influence of different HMW-GS combinations on protein content. (C) Bar plot demonstrates the effect of different genotypes on the yellow pigment content. (D) Bar plot presents the effect of different genotypes on the activity of polyphenol oxidase. (E) Bar plot reveals the effect of different genotypes on the activity of polyphenol oxidase.
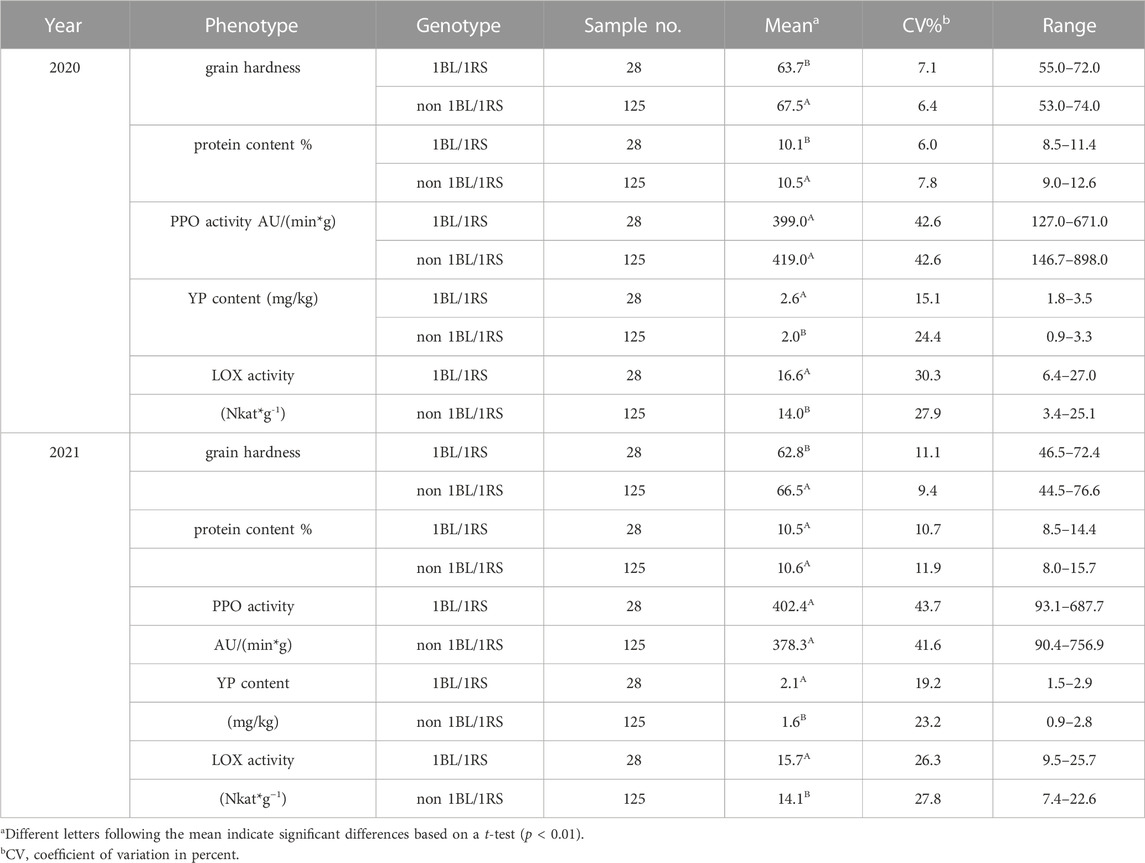
TABLE 4. Comparison of 1BL/1RS translocation and quality traits.
Discussion
Effects of grain hardness genes on quality traits
Wheat with different grain hardness has different uses for flour. Hard wheat is suitable for making bread and noodles, and soft wheat is good for making biscuits and cakes. The hardness index (HI) values have been used to define the hard wheat (>62) and soft wheat (<46) (Sun et al., 2008). Through the measurement of grain hardness, it was found that there were 123 hard wheats in this study, accounting for 80.4% of the tested varieties. Hard wheat in both Chinese and CIMMYT materials were mainly Pinb-D1b mutation type. This also verified by the previous studies (Xia et al., 2005). The Chinese wheat varieties were mainly Pina-D1a/Pinb-D1b and Pina-D1a/Pinb-D1a variant types. CIMMYT wheat varieties were primarily Pina-D1b/Pinb-D1a, and the hardness genotype was relatively narrow. By comparing the hardness values among different variant combinations, we found that the grain hardness of wheat with Pina-D1b genotype was significant higher than that of wheat with Pinb-D1a genotype. From this, we knew the effect of Pinb gene on grain hardness is minor than that of Pina gene. The research results were consistent with previous studies (Chen et al., 2007; Yang et al., 2017; Sang et al., 2020). Grain hardness was positively correlated with crude protein content (Li et al., 2001). This research showed that the protein content of Pina-D1b gene was the highest, which was outstandingly higher than that of other variation combinations.
The effect of HMW-GS on quality traits
HMW-GS accounts for 10% of the storage protein content, but it is a key factor affecting wheat processing quality (Payne et al., 1987; Zhang et al., 2009). Previous studies have proved that the Glu-D1 site has the greatest effect, while Glu-B1 and Glu-A1 have less influence. The different subunits at different sites showed different effects on the processing quality of wheat flour. 1, 2*, 5 + 10 are high-quality subunits for improving the quality of gluten (Liu et al., 2005; Lu et al., 2017). Previous research showed that 2 + 12 subunits of Glu-D1 site associated with gluten titers were the most common subunit in 123 regional varieties and modern bread wheat varieties, accounting for 63.4%, and 5 + 10 subunits associated with strong gluten bread wheat were found in 22 wheat varieties, accounting for 17.8%. Apart from these two subunits, 2.1 + 12, 2 + 12′, and 2 + 12* subunits were also found among the varieties (Sönmez et al., 2023). A previous study found Glu-B1 and Glu-B3 had the highest effect on the variations for gluten, dough and end-use quality traits, whereas Glu-A1 and Glu-D3 had the lowest impact on average. The Glu-D1 locus had a strong impact on gluten strength but its contribution to either SDS-Sedimentation volume, gluten extensibility and bread loaf volume was minimal (Guzmán et al., 2022). However, this study revealed that there were some differences in HMW-GS composition of Chinese and CIMMYT wheat varieties. The frequencies of 1/2* and 5 + 10 subunits in CIMMYT varieties accounted for 47.1% and 42.5% of the tested varieties, respectively. For Chinese wheat varieties, the proportions of 1/2* and 5 + 10 subunits were 31.4% and 13.7%, respectively, while 1 and 5 + 10 had a positive impact on protein content (Xie et al., 2016). The protein content of combinations containing 1/2* and 5 + 10 subunits was significant higher than that of combinations with NULL and 2 + 12 subunits. Among the detected varieties, there were 11 varieties containing both high quality subunit combinations 1 + 2* and 5 + 10, all of which were CIMMYT varieties. There were 26 inferior subunit combinations of Null/2 + 12 varieties, and they all were Chinese varieties. This may also be one of the reasons for the poor quality of wheat in China.
The influence of PPO activity, LOX activity and YP content on quality traits
PPO activity, LOX activity and YP content will affect the color of flour. High PPO activity will cause browning during flour processing and reduce the color of flour, while low PPO activity will make the flour white and bright (Fuerst et al., 2006; Leenhardt et al., 2006). In the present study, the trends of PPO alleles PPO-A1 and PPO-D1 in Chinese and CIMMYT wheat varieties were roughly the same, while the two major genes control PPO activity of wheat. In addition, the effect of Ppo-A1 on PPO activity of wheat was greater than that of Ppo-D1, which squared with the results of previous studies (Brian and Skinner, 2011). The wide range of PPO activity between different variant combinations indicated that the selection potential was large, and low PPO activity could be selected through breeding to improve the color browning of wheat products.
Some studies suggest that carotenoid content is significantly positively correlated with flour yellowness (Hu et al., 2004). Two-year variance analysis showed that Psy-A1 was the main gene controlling yellow pigment content. The gene combination containing Psy-A1a was significant higher than other combinations, as reported in the previous studies (Zhou et al., 2014).
The results showed that Lox-B1b and Lox-B1a genes accounted for 78.4% and 21.6% of the tested varieties, respectively. The activity range of 153 tested varieties in the 2 years was 3.3–27.0 Nkat*g−1 and 7.4–25.7 Nkat*g−1. The order of the difference in activity was basically the same, but the activity value of the same species changed obviously in different years. It was depend on the weather and Fusarium head blight PPO, YP, and LOX had a wide range of variations. Based on various breeding requirements, different alleles can be selected to improve the color of wheat flour.
The effect of 1BL/1RS translocation line on quality traits
In the 1990s, the translocation line was widely used in Chinese wheat breeding because the wheat-rye 1BL/1RS translocation line had good high yield and disease resistance (Liu et al., 2020). Later studies found that wheat varieties containing 1BL/1RS had poor quality, and the utilization rate of 1BL/1RS translocated lines significantly decreased (Yu et al., 2011). A total of 27 varieties incorporated 1BL/1RS translocations, with frequencies of 32.1% in Chinese varieties and 1.4% in CIMMYT wheat varieties, respectively. The wheat varieties encompassing 1BL/1RS translocation lines had higher LOX activity, YP content and lower grain hardness. The existence of 1BL/1RS translocation line significantly affected the YP content, as previous researches mentioned (He et al., 2008). The relationship between LOX activity and the 1BL/1RS translocation line has not been studied before. In this study, the LOX of the varieties containing the 1BL/1RS translocation line and the varieties without the 1BL/1RS translocation line were quietly different. The reason may be that most varieties containing 1BL/1RS translocation line in this test also included LOX-B1b gene. Compared with varieties without 1BL/1RS translocation line, the protein content of varieties with 1BL/1RS translocation lines were lower in 2020, but there was no significant difference in 2021, possibly because the protein content was susceptible to the environmental impact of that year. What’s more, we found that spring wheat had high protein content. Among the 153 varieties, CIMMYT varieties were all spring wheat varieties and basically did not contain translocation line, which may be one of the reasons for the high protein content of non-1BL/1RS translocation line. Therefore, whether 1BL/1RS translocation lines have an impact on protein content still needs further study. The introduction of 1BL/1RS translocation line will lead to the deterioration of some quality traits, which is not conducive to the cultivation of high-quality wheat varieties.
Materials and methods
Plant varieties and field trials
A total of 153 germplasm resources were tested, including 81 from domestic varieties, and 72 from CIMMYT varieties (Supplementary Table S2). It was tested in Xiangyang Test Base for 2 years from 2019 to 2021. Each material was sown 3 g and planted in a single row with a length of 1 m, row spacing of 0.3 m. A total of 3 replicates have been used with randomized block design. The field management is the same as the local field production. After normal maturity, each material was harvested separately. When all of grains moisture were lower than 13%, phenotype of quality was initiated by using standard methods with 3 replicates per material.
DNA extraction and identification of quality genes with molecular markers
Genomic DNA of 153 wheat varieties was isolated from their leaf using the modified cetyltrimethyl ammonium bromide (CTAB) method (Dreisigacker et al., 2016). 153 wheat varieties (Lines) were screened out by using the functional/KASP markers of quality trait genes (PPO, Psy, Lox, Pina, Pinb, 1BL/1RS, Glu-D, and Glu-A1). A standard polymerase chain reaction (PCR) was performed by Dreisigacker et al. (2016). A 10 μL system was used for PCR amplification, including 1 μL DNA, 5 μL 2×Taq PCR Mix (Vazyme P112-03, China) and 1 μmol·L−1 primer, adding ddH2O to 10 μL. The PCR amplification process was as follows: predenaturation at 94°C for 5 min; denaturation at 94°C for 1 min; annealing at 50°C–66°C for 1 min; extension at 72°C for 2 min. And it was 30–35 cycles in total. Then the PCR amplification was prolonged for 10 min at 72°C and stored at 4°C. The PCR products were detected by 1.2% agarose gel electrophoresis. KASP markers were assayed by RT-PCR (CFX Connect, BIO RAD). PCR system could be summarized as follows: DNA 1 μL, KASP Master Mix (KBS-1016-001, KASP V4.0 2×Master Mix96/384, Standard Rox) 2.5 μL, primer 0.25 μL and ddH2O 1.25 μL, 95°C pre-denaturation for 15 min, 94°C denaturation for 15 s, annealing at 65°C for 60 s, then each cycle decreased by 0.8°C, a total of 10 cycles, finally reduced to 57°C, 94°C denaturation for 20 s, 57°C annealing for 60 s, a total of 40 cycles. The primers used were synthesized by Beijing Tsingke Biotech Co.
Quality traits determination method
Grain hardness
Use wheat hardness tester JYDX100 × 40 to determine the hardness value HI of wheat samples according to the national standard GB/T21304 (Luo et al., 2018).
Flour protein content
Use a near-infrared instrument (NIR) 2600XT-R to determine the protein content of wheat in terms of AACC39-25.
LOX activity
The LOX activity was determined using the method reported by Zheng et al. (2011).
PPO activity
The PPO activity was determined using the method reported by Si et al. (2007).
YP content
AACC 14–50 has been slightly modified (Approved Methods of AACC. Method 14–50.); Pour 0.5 g of flour into a 10 mL centrifuge tube; add 2 mL of water-saturated n-butanol extract (volume ratio 5:1); stir well and shake for 30 min; centrifuge with Eppendorf at 4,000 r/min for 10 min; 200 μL supernatant is taken from pipette and poured into a 96-well microtiter plate; use a water-saturated nbutanol solution as a control; use a microplate reader to measure the absorbance of the supernatant at 436.5 nm·A, Yellow pigment content = 30.1*A (American Association of Cereal Chemists International, 2000).
Statistical analysis
The phenotype data and genotype test results were processed and analyzed using data processing software SAS9.2 (https://www.sas.com/zh_cn/home.html) and Microsoft Excel, incorporating descriptive statistics, phenotypic correlation analysis and analysis of variance (ANOVA).
Conclusion
Wheat is one of the most important food crops in China. Although the supply of wheat is sufficient, it still cannot meet people’s demand for good quality wheat. Therefore, screening germplasm resources to identify good-quality wheat is of great significance to improve the supply and demand structure of wheat in China. In this study, we found that there were some differences in quality of wheat varieties among Chinese and CIMMYT wheat varieties in hardness, high molecular weight wheat grain subunit protein, PPO, LOX and YP and all of them controlled by genotypes, which could be screened and introduced by marker-assisted selection. The genetic complexity of wheat quality characteristics was discovered on the basis of the detection of genotype and phenotypic traits. Numerous studies in recent years proved that molecular marker-assisted selection had sufficient accuracy in wheat quality breeding and could effectively select superior genotypes for wheat quality traits. In the future, excellent germplasm resources can be chosen by combining genotype and phenotypic traits to meet the breeding needs and speed up the process of wheat quality breeding. In this study, we identified two good strong gluten CIMMTY wheat germplasm, T. DICOCCONCI9309/AE.SQUARROSA (409)//2*PANDORA/5/WAXWING/3 /BL1496/MILAN//PI610750/4/FRNCLN/6/KACHU/BECARD//WBLL1*2/BRAMBLING and KACHU*2/3/ND643//2*PRL/2*PASTOR/4/KACHU/DANPHE as well as three higher weak gluten Chinese wheat varieties, such as EEN 1, Luo 6010 and Emai 9721. These five wheat germplasm will be used for wheat breeders to develop wheat varieties of better quality through marker-assisted selection.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
PM and CL initiated the project, designed experiment. PL and ZZ contributed to phenot-ype data, designed the experiment and finished the manuscript. YY assisted in data analysis and participated in the drafting of the manuscript. YR, SY, and WS prepared samples, extracted DNA. HX and FC contributed to the section on phenotype data in the field. Lastly, all authors contributed to the article and approved the submitted version.
Funding
This work was supported by development and application of new wheat varieties with good quality and high yield for the Middle and Lower Reaches of the Yangtze River (NK2022060601), Key Research and Development Program of Hubei Province (2022BBA0035), HZAU-AGIS Co-operation Fund (SZYJY2022035), Xinjiang Bingtuan Science and Technology Program (2019AB021, 2016AC027, 2022DB013, and 2022CA006), The National Natural Science Foundation of China (U1178306 and 31560397).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2023.1198835/full#supplementary-material
References
American Association of Cereal Chemists International, (2000). Approved methods of AACC. Method 14-50. St. Paul, MN: AACCI, 14–50.
Brian, B., and Skinner, D. (2011). Molecular cloning and expression analysis of multiple polyphenol oxidase genes in developing wheat (Triticum aestivum) kernels. J. Cereal Sci. 53, 371–378. doi:10.1016/j.jcs.2011.01.015
Chen, F., Beecher, B., and Morris, C. F. (2010). Physical mapping and a new variant of Puroindolineb 2 genes in wheat. Theor. Appl. Genet. 120, 745–751. doi:10.1007/s00122-009-1195-y
Chen, F., He, Z., Chen, D., Zhang, C., and Xia, X. (2007). Allelic variation of puroindoline genes inChinese spring wheats. Sci. Agric. Sin. 02, 217–224. (In Chinese with English abstract). doi:10.3321/j.issn:0578-1752.2007.02.001
Chen, F., He, Z., Morten, L., and Xia, X. (2005). Detection of allelic variation for grain hardness in CIMMYT common wheat. Acta Agron. Sin. 31, 1277–1283. (In Chinese with English abstract). doi:10.3321/j.issn:0496-3490.2005.10.006
Cheng, L., Yang, Z., Li, G., Zeng, Z., Zhang, Y., Zhou, J., et al. (2008). Isolation of a new repetitive DNA sequence from Secale africanum enables targeting of Secale chromatin in wheat background. Euphytica 159, 249–258. doi:10.1007/s10681-007-9484-5
Cooper, J., Stromberger, J., Morris, C., Bai, G., and Haley, S. (2015). End-use quality and agronomic characteristics associated with the Glu-B1al High-Molecular-Weight glutenin allele in U.S. Hard winter wheat. Crop Sci. 56, 2348–2353. doi:10.2135/cropsci2015.10.0610
Dreisigacker, S., Sehgal, D., Luna, G., Reyes, A., and Mollins, J. (2016). CIMMYT wheat molecular genetic-s laboratory protocols and applications to wheat breeding. Mexico: CIMMYT, D.F.
Fuerst, E., Anderson, J., and Morris, C. (2006). Delineating the role of polyphenol oxidase in the darkening of alkaline wheat noodles. J. Agric. Food Chem. 54, 2378–2384. doi:10.1021/jf0526386
Geng, H., He, Z., Zhang, L., Qu, Y., and He, Z. (2012). Development of functional markers for a lipoxygenase GeneTaLox-B1on chromosome 4BS in common wheat. Crop Sci. 52, 568–576. doi:10.2135/cropsci2011.07.0365
Guzmán, C., Crossa, J., Mondal, S., Govindan, V., Huerta, J., Crespo-Herrera, L., et al. (2022). Effects of glutenins (Glu-1 and Glu-3) allelic variation on dough properties and bread-making quality of CIM-MYT bread wheat breeding lines. Field Crops Res. 284, 108585. doi:10.1016/J.FCR.2022.108585
Han, B., Wang, C., Ren, Y., Zhao, X., Dong, Y., Li, Y., et al. (2015). Advances on the wheat molecular breeding - functional genes of wheat grain quality. J. Shanxi Agric. Sci. 43, 233–236. (In Chinese with English abstract). doi:10.3969/j.issn.1002-2481.2015.02.29
He, X., He, Z., Zhang, L., Sun, D., Morris, C., Fuerst, E., et al. (2007). Allelic variation of polyphenol oxidase (PPO) genes located on chromosomes 2A and 2D and development of functional markers for the PPO genes in common wheat. Theor. Appl. Genet. 115, 47–58. doi:10.1007/s00122-007-0539-8
He, X., Zhang, Y., He, Z., Wu, Y., Xiao, Y., Ma, C., et al. (2008). Characterization of a phytoene synthase 1 gene (Psy1) located on common wheat chromosome 7A and development of a functional marker. Theor. Appl. Genet. 116, 213–221. doi:10.1007/s00122-007-0660-8
Hu, R., Tian, J., and Lü, J. (2004). Stability analysis of carotenoid content and its coefficient correlation with yellow alkaline noodle color characteristics in Common wheat (Triticum aestivum L.). Acta Agron. Sin. 6, 597–601. (In Chinese with English abstract). doi:10.3321/j.issn:0496-3490.2004.06.014
Kuchel, H., Langridge, P., Mosionek, L., Williams, K., and Jefferies, S. (2006). The genetic control of milling yield, dough rheology and baking quality of wheat. Theor. Appl. Genet. 112, 1487–1495. doi:10.1007/s00122-006-0252-z
Leenhardt, F., Lyan, B., Rock, E., Boussard, A., Potus, J., Chanliaud, E., et al. (2006). Genetic variability carotenoid concentration and lipoxygenase and peroxidase activities among cultivated wheat sp-ecies and bread wheat varieties. Eur. J. Agron. 25, 170–176. doi:10.1016/j.eja.2006.04.010
Li, S., Shan, M., Wang, Y., Li, B., and Zhang, S. (2001). Evaluation of wheat quality for wet noodle making. Acta Agron. Sin. 03, 334–338. (In Chinese with English abstract). doi:10.3321/j.issn:0496-3490.2001.03.011
Liu, C., Han, R., Wang, X., Gong, W., Cheng, D., Cao, X., et al. (2020). Research progress of wheat wild hybridization, disease resistance genes transfer and utilization. Sci. Agric. Sin. 53, 1287–1308. (In Chinese with English abstract). doi:10.3864/j.issn.0578-1752.2020.07.001
Liu, D., Chen, W., Zhang, B., Liu, D., Liu, B., and Zhang, H. (2015). Identification and analysis of new puroi-ndoline pina-d1o. Acta Agric. Boreali-occidentalis Sin. 24, 52–53. (In Chinese with English abstract). doi:10.1016/j.jpurol.2015.02.002
Liu, L., He, Z., Yan, J., Zhang, Y., Xia, X., and Pena, R. (2005). Allelic variation at the Glu-1 and Glu-3 loci, presence of the 1B.1R translocation, and their effects on mixographic properties in Chinese bread wheats. Euphytica 142, 197–204. doi:10.1007/s10681-005-1682-4
Lu, J., Pang, L., and Chai, S. (2017). Effects of HMW-GS on quality properties of spring wheat and e-valuation of subunit score system. J. Nucl. Agric. Sci. 31, 80–87. (In Chinese with English abstract). doi:10.11869/j.issn.100-8551.2017.01.0080
Luo, X., Yang, Z., and Ye, G. (2018). Study on operation of JYDB100×40 wheat hardness index tester. Grain Sci. Technol. Econ. 43, 42–43. (In Chinese with English abstract). doi:10.16465/j.gste.cn431252ts.20180408
Morris, C. (2002). Puroindolines: The molecular genetic basis of wheat grain hardness. Plant Mol. Biol. 48, 633–647. doi:10.1023/a:1014837431178
Payne, P. (1987). Genetics of wheat storage proteins and the effect of allelic variation on bread-making quality. Annu. Rev. Plant Physiology 38, 141–153. doi:10.1146/annurev.pp.38.060187.001041
Payne, P., Nightingale, M., Krattiger, A., and Holt, L. (1987). The relationship between HMW glutenin subunit composition and the bread-making quality of British-grown wheat varieties. J. Sci. Food Agric. 40, 51–65. doi:10.1002/jsfa.2740400108
Rasheed, A., Wen, W., Gao, F., Zhai, S., Jin, H., Liu, J., et al. (2016). Development and validation of KASP assays for genes underpinning key economic traits in bread wheat. Theor. Appl. Genet. 129, 1843–1860. doi:10.1007/s00122-016-2743-x
Sang, W., Liu, P., Nie, Y., Cui, F., Han, X., Kong, D., et al. (2020). Influence of Pins gene on milling quality and Xinjiang hand-stretched noodle processing quality. J. Triticeae Crops 12, 1–9. (In Chinese with English abstract). doi:10.7606/j.issn.1009-1041.2020.12.03
Si, H., Ma, C., He, K., Zhou, N., and Hu, N. (2007). Comparative analysis on detection methods for activity of polyphenol oxidase in wheat. Cereal Feed Industry 0, 8–10. (In Chinese with English abstract). doi:10.3969/j.issn.1003-6202.2007.07.004
Sönmez, M. E., Güleç, T., Demir, B., Bayraç, C., Çakmak, M., and Aydin, N. (2023). Molecular screening of the landraces from Turkey and modern bread wheat (Triticum aestivum L) cultivars for HMW-GS, wbm, waxy genes and Lr34 gene. Genet. Resour. Crop Evol. 70, 775–788. doi:10.1007/s10722-022-01460-0
Sun, D., He, Z., Xia, X., Zhang, L., Morris, C., Appels, R., et al. (2005). A novel STS marker for polyphe-nol oxidase activity in bread wheat. Mol. Breed. 16, 209–218. doi:10.1007/s11032-005-6618-0
Sun, H., Ouyang, S. H., and Duan, X. (2017). Wheat quality in China-status and challenge. Sci. Technol. Cereals, Oils Foods 25, 1–4. (In Chinese with English abstract). doi:10.16210/j.cnki.1007-7561.2017.02.001
Sun, H., Wu, C., Yang, Z., Jiang, W., Tang, H., and Zhou, Z. (2008). Hardness situation and classification of Chinese wheat. J. Chin. Cereals Oils Assoc. 23, 38–42. doi:10.3724/SP.J.1005.2008.01083
Vancini, C., Torres, G., Miranda, M., Consoli, L., and Grando, M. (2019). Impact of high molecular weight glutenin alleles on wheat technological quality. Pesqui. Agropecuária Bras. 54. doi:10.1590/S1678-3921.pab2019.v54.00639
Wang, J., Zhang, Z., Qi, S., Zhao, Y., Huang, J., Yu, L., et al. (2019). Quality analysis of wheat variaties (Lines) in South Huanghuai region. Tianjin Agric. Sci. 25, 34–37. (In Chinese with English abstract). doi:10.7606/j.issn.1009-1041.2016.05.09
Wang, X., Ma, C., He, K., Si, H., and Zhang, Y. (2008). Development and application of a STS Marker for grain PPO gene located on chromosome 2D in common wheat. Sci. Agric. Sin. 41, 1583–1590. (In Chinese with English abstract). doi:10.3864/j.issn.0578-1752.2008.06.003
Xia, L., Chen, F., He, Z., Chen, X., and Morris, C. (2005). Occurrence of puroindoline alleles in Chinese winter wheats. cereal Chem. 82, 38–43. doi:10.1094/cc-82-0038
Xie, K., Zhu, B., Sun, J., Liu, D., Chen, S., Ren, Y., et al. (2016). Composition of high molecular weigh-t glutenin subunits of wheat varieties(lines) in huanghuai southern region and their relations-hip with quality. J. Triticeae Crops 36, 595–602. (In Chinese with English abstract). doi:10.7606/j.issn.1009-1041.2016.05.09
Yang, Z., Gu, Z., Zhou, Y., Wang, A., Gao, P., Xiong, Z., et al. (2017). Detection of allelic variation for grain hardness in Huaibei region of Jiangsu province by KASP markers. J. Triticeae Crops 02, 153–161. (In Chinese with English abstract). doi:10.7606/j.issn.1009-1041.2017.02.02
Yu, L., He, F., Chen, G., Cui, F., Qi, X., Wang, H., et al. (2011). Identification of 1BL·1RS wheat rye C-hromosome translocations via 1RS specific molecular markers and genomic in situ hybridizati-on. Acta Agron. Sin. 37, 563–569. (In Chinese with English abstract). doi:10.3724/SP.J.1006.2011.00563
Zhang, D., He, T., Liang, H., Huang, L., Su, Y., Li, Y., et al. (2016). Flour quality and related molecular characterization of high molecular weight gluten in subunit genes from wild emmer wheat accession-n TD-256. J. Agric. Food Chem. 64, 5128–5136. doi:10.1021/acs.jafc.6b01547
Zhang, L., Yan, J., Xia, X., and He, Z. (2006). Possible role of potassium channel, big K in etiology of schizophrenia. Acta Agron. Sin. 32, 41–43. (In Chinese with English abstract). doi:10.1016/j.mehy.2005.09.055
Zhang, Y., Tang, J., Yan, J., Zhang, Y., Zhang, Y., Xia, X., et al. (2009). The gluten protein and interactions between components determine mixograph properties in an F6 recombinant inbred line population in bread wheat. J. Cereal Sci. 50, 219–226. doi:10.1016/j.jcs.2009.05.005
Zhang, Z., Jiang, X., Wang, Z., Zhu, Q., Liu, M., and Ru, Z. (2019). Genetic diversity of HMW-GS and its relationship with quality of wheat germplasms from different regions. Acta Agron. Sin. 34, 75–81. (In Chinese with English abstract). doi:10.7668/hbnxb.201751750
Zheng, W., Wang, H., Cui, W., Zhang, W., and Yao, D. (2011). Lipoxygenase activity in 104 wheat varieti-es. Sci. Agric. Sin. 44, 1798–1805. (In Chinese with English abstract). doi:10.3864/j.issn.0578-1752.2011.09.005
Keywords: wheat, quality property, quality related genes, molecular markers, KASP marker
Citation: Liu P, Zhang Z, Yin Y, Yan S, Ren Y, Sang W, Xu H, Han X, Cui F, Nie Y, Kong D, Li W, Lan C and Mu P (2023) Quality traits analysis of 153 wheat lines derived from CIMMYT and China. Front. Genet. 14:1198835. doi: 10.3389/fgene.2023.1198835
Received: 02 April 2023; Accepted: 20 July 2023;
Published: 02 August 2023.
Edited by:
Yuanfeng Hao, Chinese Academy of Agricultural Sciences, ChinaReviewed by:
Balpreet Kaur Dhatt, Bayer Crop Science, United StatesMing Li, Chinese Academy of Agricultural Sciences, China
Copyright © 2023 Liu, Zhang, Yin, Yan, Ren, Sang, Xu, Han, Cui, Nie, Kong, Li, Lan and Mu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Caixia Lan, Y3hsYW5AbWFpbC5oemF1LmVkdS5jbg==; Peiyuan Mu, bXVweUAxNjMuY29t
†These authors have contributed equally to this work