 Zhihan Zhou
Zhihan Zhou Jinglei Qian
Jinglei Qian Jianhui Xie
Jianhui Xie
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Genet., 02 May 2023
Sec. Evolutionary and Population Genetics
Volume 14 - 2023 | https://doi.org/10.3389/fgene.2023.1182028
The Y-chromosomal haplogroup tree, which consists of a group of Y-chromosomal loci with phylogenetic information, has been widely applied in anthropology, archaeology and population genetics. With the continuous updating of the phylogenetic structure, Y-chromosomal haplogroup tree provides more information for recalling the biogeographical origin of Y chromosomes. Generally, Y-chromosomal insertion-deletion polymorphisms (Y-InDels) are genetically stable as Y-chromosomal single nucleotide polymorphisms (Y-SNPs), and therefore carry mutations that can accumulate over generations. In this study, potential phylogenetic informative Y-InDels were filtered out in haplogroup O-M175, which is dominant in East Asia, based on population data retrieved from the 1000 Genomes Project. A group of 22 phylogenetic informative Y-InDels were identified and then assigned to their corresponding subclades of haplogroup O-M175, which provided a supplement for the update and application of Y-chromosomal markers. Especially, four Y-InDels were introduced to define subclades determined using a single Y-SNP.
The Y-chromosomal variants are widely applied in the fields of human genetic studies by virtue of its paternal inheritance (Jobling and Tyler-Smith, 1995; Hammer and Zegura, 1996; Stone et al., 1996; Kayser, 2003; Wang et al., 2014; Kayser, 2017). Among genetic markers on the Y chromosome, Y-chromosomal single nucleotide polymorphisms (Y-SNPs) are genetically stable and widely distributed, which can reveal the steady transmission of genetic material between ancestors and descendants (Xue et al., 2009). A specific set of Y-SNPs accumulates over generations and gradually forms multiple Y-chromosomal haplogroups as humans migrate and populations expand (Jobling and Tyler-Smith, 2003). On this basis, the first uniformly named phylogenetic Y-chromosomal haplogroup tree was published in 2002 and since then the worldwide Y-SNP types have been divided into 20 main Y-chromosomal haplogroups A to T (Underhill et al., 2001; Consortium, 2002).
With the development of detection technology, more validated phylogenetic informative Y-SNPs are identified, thus more refined phylogenetic structure of the Y-chromosomal haplogroups are constantly consummated (Van Geystelen et al., 2013). Recently, a batch of studies demonstrated more evidence for the update of Y-chromosomal haplogroups, resulting in a renewed understanding of the phylogenetic structure of Y-chromosomal haplogroup C, D, J, K and S (Mondal et al., 2017; Kutanan et al., 2018; Haber et al., 2019; Kanzawa-Kiriyama et al., 2019; Pinotti et al., 2019; Urasin et al., 2019). The continually updated Y-chromosomal haplogroup tree and its subclades provide a refined Y-chromosomal phylogenetic topology structure to further investigate population evolution in more recent era, which facilitates the understanding of the biogeographical origins of ethnic minorities and more specific populations in the application of molecular population anthropology. Besides, the Y-chromosomal haplogroup tree has also attracted the attention of forensic scientists. Previous studies have reported similarity or consistency in unrelated male individuals and father-son discordance during Y-chromosomal STR analysis (Jung et al., 2016; Wang et al., 2016; Zhou et al., 2018). While current Y-chromosomal STR typing cannot make a definite exclusion, the information of Y-chromosomal haplogroups/sub-haplogroups can provide a useful supplement.
Y-chromosomal insertion-deletion polymorphisms (Y-InDels) were generated during the transmission between father and son with stable inheritance (Nachman and Crowell, 2000). As Y-SNPs, such mutations have been passed down along with human evolution and migration, allowing them to be used to trace biogeographical origins. Currently, little concern was paid to phylogenetic information carried by Y-InDels. In Y-chromosomal haplogroup tree 2019–2020, Y-chromosomal haplogroups/sub-haplogroups were mainly defined using Y-SNPs, while Y-InDels accounted for only a very small fraction. In fact, Y-InDels with fragment length polymorphisms confer to convenience in practice (Mills et al., 2006).
The Y-chromosomal haplogroup O-M175 is overwhelmingly dominant in East Asia (Jin and Su, 2000; Karafet et al., 2001; Karafet et al., 2010; Zhong et al., 2011). Recent surveys have shown that haplogroup O-M175 and its sub-haplogroups represent a high proportion in the Chinese population, reaching more than 80% (Lang et al., 2019; Song et al., 2019; Zhang et al., 2020; Wang et al., 2022; Tao et al., 2023). The dominance of the haplogroup O-M175 has brought much focus on its refined phylogenetic structure. Previous researches have investigated potential phylogenetic informative Y-SNPs and provided evidences of newly topologies of haplogroup O-M175 (Yan et al., 2011; Naitoh et al., 2013; Kwon et al., 2015; Ning et al., 2016).
In the current study, we aimed to identify a subset of phylogenetic informative Y-InDels in haplogroup O-M175. After selection of potential Y-InDels, the validation experiments were carried out. Moreover, we reviewed the topology structure of haplogroup O-M175, then assigned candidate Y-InDels to corresponding subclades.
Blood samples were collected from unrelated males in a Chinese Han population with written informed consent according to protocols approved by the Ethics Committee of the School of Basic Medical Sciences, Fudan University. DNA extractions were performed according to Chelex-100 procedure (Walsh et al., 1991).
To select potential phylogenetic informative Y-InDels in haplogroup O-M175, the variant call format (VCF) file from the 30× sequencing data of the fourth phase 1,000 Genomes Project were analyzed (Byrska-Bishop et al., 2022). All male individuals were assigned to the corresponding Y-haplogroups using yhaplo package (Poznik, 2016) based on Python 3.11.1 (https://www.python.org/), with an index manually updated according to the Y-chromosomal haplogroup tree 2019–2020. After obtaining the data of all sequenced bi-allelic Y-InDels, the allele frequencies of each Y-InDel in each Y-chromosomal haplogroup/sub-haplogroup were calculated by direct counting. The Y-InDels which were polymorphic only in haplogroup O-M175 were pending further screening.
With frequencies of candidate Y-InDels in each subclade of haplogroup O-M175, target Y-InDels were screened following two criteria: first, the candidate Y-InDel was mutant in all individuals of the corresponding sub-haplogroup and its subclade; second, the candidate Y-InDel was not mutated in all individuals except the corresponding sub-haplogroup and its subclade. The first principle ensures that the mutation status of candidate Y-InDel does not result from random mutations, and the second principle guarantees the specificity of the candidate Y-InDel.
Primer pairs were designed using the Primer Premier 5 software (PREMIER Biosoft, CA, United States), considering following criteria: amplicon size limited between 200 bp to 1,000 bp; Tm values from 55°C to 65°C; GC content from 40% to 60%; avoiding primer-dimer, hairpin loop formations and complementary 3′-ends. The specificity of DNA sequences and primer pairs was evaluated using the BLAST-Like Alignment Tool (Kent, 2002) and the Basic Local Alignment Search Tool (Altschul et al., 1990), respectively. For capillary electrophoresis analysis, one of each primer sets were modified with fluorescent dyes (FAM or HEX) at the 5′-ends. All primer sets were validated by agarose gel electrophoresis.
PCR amplification was performed in a volume of 30 μL, consisted of 15 μL of 2× Taq Master Mix (Novoprotein, China), 0.3 μL of primer and 1.5 μL of template DNA. Thermal cycling was performed in a GeneAmp PCR System 9,700 (Applied Biosystems, CA, United States), with an initial denaturation step at 94°C for 90 s, followed by 35 cycles of 94°C for 30 s, 58°C for 60 s, 72°C for 60 s, then a final extension at 72°C for 10 min, and maintained at 4°C.
To detect the genotype of the candidate Y-InDels, the PCR products were first subjected to capillary electrophoresis. The samples were prepared by combination of 1 μL PCR products and 9 μL of a 19:1 mixture of deionized Hi-Di™ Formamide (Thermo Fisher Scientific, MA, United States) and GeneScan™ 500 LIZ® size standard (Thermo Fisher Scientific, MA, United States). The mixture was denatured at 95°C for 5 min and then chilled at 4°C. The products were separated by ABI PRISM 3130xL Genetic Analyzer (Applied Biosystems, CA, United States). The run data were analyzed using GeneMapper® ID software v3.2 (Applied Biosystems, CA, United States). Genomic DNA 2800M (Promega, United States) was applied to be a positive control.
To validate potential phylogenetic informative Y-InDels, male samples belonging to the haplogroup O-M175 were screened out by a previously developed 16-plex Y-SNP typing system (Zhou et al., 2020) for subsequent experiments. With genotype of candidate Y-InDels, a series of Sanger sequencing were performed (Saiheng Biotechnology Co., Shanghai, China). Both wild-type and mutant samples of each candidate Y-InDel were sequenced to confirm the alleles of loci. After that, we selected the representative Y-SNPs of the corresponding subclades of each candidate Y-InDel. Next, we sequenced those representative Y-SNPs for wild-type and mutant samples. After obtaining the sequencing results, the mutation status of the candidate Y-InDels was compared with that of the subclade-determining Y-SNPs to determine the subclades which the candidate Y-InDels belonged to. In addition, we cross-validated the haplogroup/sub-haplogroup assignment of newly identified Y-InDels and the 16-plex Y-SNP typing system (Zhou et al., 2020) using 235 male samples.
After initial processing of sequencing data from 1000 Genomes Project, a total of 23,726 bi-allelic Y-InDels on 1233 Y chromosomes were acquired. After the frequencies of Y-InDels in each Y-chromosomal haplogroup was calculated, a batch of 2,375 Y-InDels were variant only in haplogroup O-M175. Following screening criteria, a number of 614 potential phylogenetic informative Y-InDels and their corresponding subclades were identified. This batch of Y-InDels and the representative Y-InDels of the Y-chromosomal tree 2019–2020 were compared to remove identical Y-InDels, namely, M175, M111, A15721, M121, FGC12511, M134, M117, M133 and M333. A total of 605 Y-InDels were selected for further screening. It is worth noting that a number of candidate Y-InDels were also eliminated due to their high homology with sequences in other regions or difficulties in their detection. Eventually, a group of 22 Y-InDels was identified as potential phylogenetic informative Y-InDels for subsequent validation. The general information of newly phylogenetic informative Y-InDels is presented in Table 1.
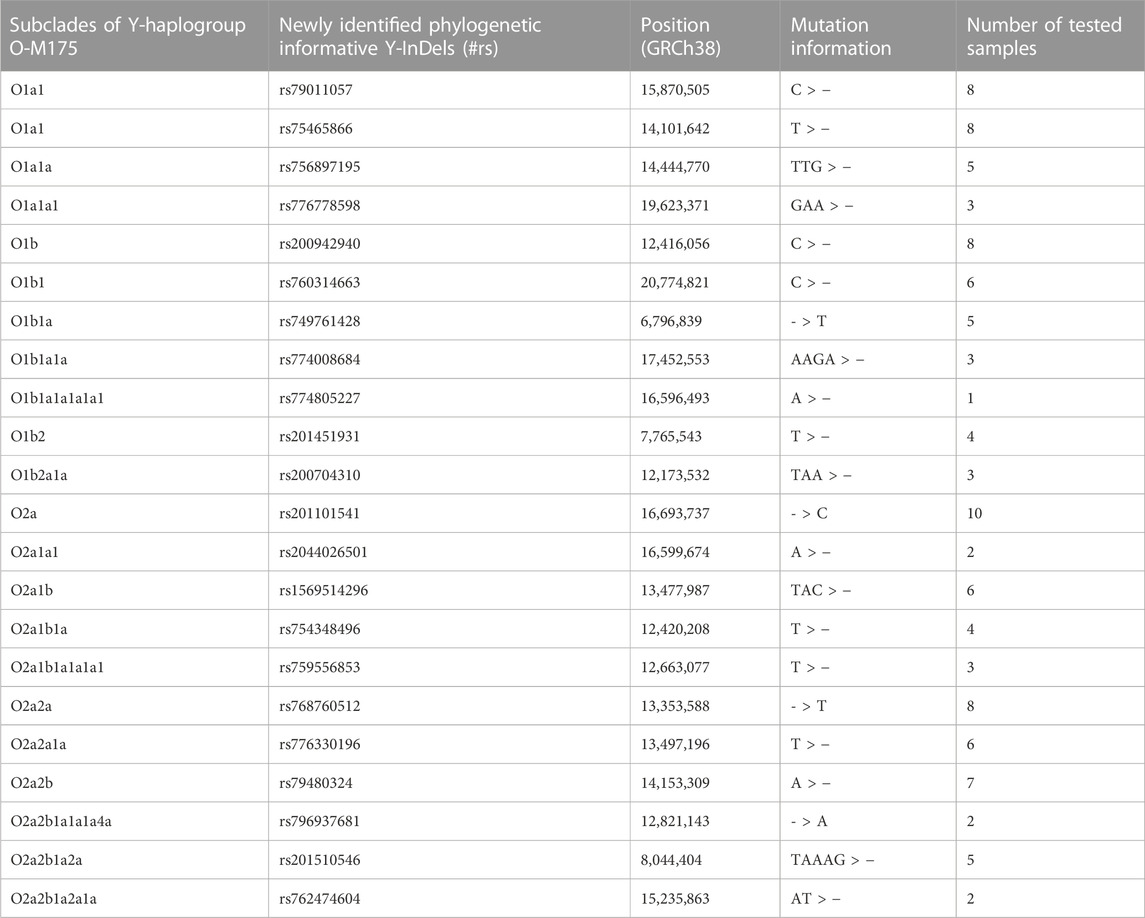
TABLE 1. The summary of phylogenetic informative Y-InDels and corresponding subclades in haplogroup O-M175.
The ancestral and derived status of each candidate Y-InDel was first validated using samples. The results from capillary electrophoresis analysis showed the presence of ancestral and derived status of each candidate Y-InDel, which confirms the diversity of candidate Y-InDels in the population. Variants at rs79011057 and rs75465866 are most likely synchronized, as suggested in both data screening and validation experiments.
In order to assign the subclade of candidate Y-InDels, corresponding representative Y-SNPs were sequenced in individuals that may belong to the corresponding subclade. The selected representative Y-SNPs and their corresponding subclades is visually displayed in Figure 1. In each subclade, the mutation status of candidate Y-InDels was validated by sequencing to be consistent with representative Y-SNPs and occurred synchronously in the same samples (Table 1). Thereby, candidate Y-InDels could be assigned to the corresponding subclade of haplogroup O-M175 while tested samples were assigned into corresponding haplogroups/sub-haplogroups. Importantly, five Y-InDels (rs79011057, rs75465866, rs749761428, rs754348496 and rs768760512) could be assigned, respectively, to haplogroup O1a1, O1b1a, O2a1b1a and O2a2a which were defined only using a single Y-SNP in Y-haplogroup tree 2019–2020.

FIGURE 1. A phylogenetic tree constructed by phylogenetic informative Y-InDels with representative Y-SNPs of corresponding subclades. The dotted lines indicate haplogroups/sub-haplogroups which is not included in the current study. The bold fonts indicate Y-InDel loci.
In attempt to provide additional evidence, we performed a previously developed 16-plex system on investigated samples (Zhou et al., 2020). The assignment of Y-haplogroups/sub-haplogroups from the 16-plex system and the newly identified Y-InDels were compared. Although the two sets of loci did not cover the identical Y-chromosomal haplogroups/sub-haplogroups, there were no conflicting assignment in all tested samples. The 16-plex system contains only the main subclades of the haplogroup O-M175, whereas the 22 newly identified Y-InDels can further assign samples to further downstream sub-haplogroups. For example, the 16-plex system contains two subclades, O1b and O1b1a2, and lacks representative sites of other downstream branches of haplogroup O1b. The addition of newly identified Y-InDels could identify haplogroup O1b1 (rs760314663), O1b1a (rs749761428), O1b1a1a (rs774008684), O1b1a1a1a1a1 (rs774805227), O1b2 (rs201451931) and O1b2a1a (rs200704310) (Figure 1), thus providing more detailed Y-chromosomal haplogroup typing results. Furthermore, these 6 Y-InDels showed ancestral status in samples with haplogroup O1b1a2.
In the current study, a number of candidate phylogenetic informative Y-InDels in haplogroup O-M175 were screened by data analysis. Phylogenetic information of candidate Y-InDels was identified by a series of validation experiments, then distributed to the corresponding subclades of haplogroup O-M175. The development of Y-InDel markers can complement the phylogenetic informative markers to enrich the definition and to provide the typing of Y-haplogroup/sub-haplogroup in a convenient manner. In fact, it is difficult to identify new subclades using Y-InDels due to the wide investigation in the definition of subclades of Y-haplogroups using Y-SNPs. However, as a type of genetically stable markers with length polymorphism, Y-InDels has the advantage of convenient detection, which introduces a prospect for its further application.
In this study, the characteristics and advantages of Y-InDels were fully considered, so as to use them as Y-chromosomal phylogenetic informative markers. In haplogroup O-M175 from the Y-chromosomal haplogroup tree 2019–2020, a total of 9 haplogroup/sub-haplogroups can be determined using Y-InDels including haplogroup O-M175, O1b1a1a1a1a1-M111, O1b1a1a1a1a1a1a1a-A15721, O2a1a1a1a1-M121, O2a2b1-M134, O2a1b1a-FGC12511, O2a2b1a1-M117, O2a2b1a1a-M133 and O2a4-M333. The newly phylogenetic informative Y-InDels provided candidates in haplogroup O-M175, especially for the subclades with only one representative Y-SNP in Y-haplogroup tree 2019–2020, such as haplogroup O1a1-B384/Z23193, O1b1a-M1470, O2a1b1a-FGC12511, and O2a2a-M188. The newly phylogenetic informative Y-InDels involved in subclades of several levels. For instance, the rs796937681 corresponding to the 12th level subclade O2a2b1a1a1a4a-CTS4658 of haplogroup O-M175, indicating that the newly phylogenetic informative Y-InDels had the ability to subdivide the haplogroup O-M175. The detailed division of Y-haplogroups can provide more information of biogeographical origins, thus making phylogenetic informative Y-InDels a practical supplement in various researches.
The information of paternal biogeographical origin contained in Y-haplogroup tree has always been the focus of anthropological and archaeological research to trace the history of human evolution and expansion. When the emergence of human out of Africa was confirmed by Y-haplogroups (Underhill et al., 2000), the biogeographical origins of populations have been revealed, with the continuous refinement and renewal of Y-haplogroups/sub-haplogroups (Lall et al., 2021; Platt et al., 2021). Using the newly identified phylogenetic informative Y-InDels from this study, the subdivision of Y-haplogroup targeting the haplogroup O-M175 can be performed in a more convenient form. On the other hand, smaller amplicon size at Y-InDels also facilitate the detection of archaeological samples and ancient DNA. The subdivision of Y-haplogroups provides the possibility for further investigation of population evolution in more recent era, which may improve the understanding of the biogeographical origins of ethnic minorities and more specific populations.
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
The studies involving human participants were reviewed and approved by the Ethics Committee of the School of Basic Medical Sciences, Fudan University. The patients/participants provided their written informed consent to participate in this study.
ZZ contributed to conceptualization, investigation, methodology, data curation and analysis, validation, visualization, and writing of original draft. ZL contributed to investigation and data curation. YY and QJ contributed to resources. JQ contributed to investigation. CS contributed to resources and investigation. JX contributed to conceptualization, supervision, writing, project administration and funding acquisition.
This work was supported by National Natural Science Foundation of China (82171875).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215 (3), 403–410. doi:10.1016/S0022-2836(05)80360-2
Byrska-Bishop, M., Evani, U. S., Zhao, X., Basile, A. O., Abel, H. J., Regier, A. A., et al. (2022). High-coverage whole-genome sequencing of the expanded 1000 Genomes Project cohort including 602 trios. Cell 185 (18), 3426–3440.e19. doi:10.1016/j.cell.2022.08.004
Consortium, Y. C. (2002). A nomenclature system for the tree of human Y-chromosomal binary haplogroups. Genome Res. 12 (2), 339–348. doi:10.1101/gr.217602
Haber, M., Jones, A. L., Connell, B. A., AsanArciero, E., Yang, H., et al. (2019). A rare deep-rooting D0 african Y-chromosomal haplogroup and its implications for the expansion of modern humans out of Africa. Genetics 212 (4), 1421–1428. doi:10.1534/genetics.119.302368
Hammer, M. F., and Zegura, S. L. (1996). The role of the Y chromosome in human evolutionary studies. Evol. Anthropol. Issues, News, Rev. 5 (4), 116–134. doi:10.1002/(SICI)1520-6505(1996)5:4<116:AID-EVAN2>3.0.CO;2-E
Jin, L., and Su, B. (2000). Natives or immigrants: Modern human origin in East Asia. Nat. Rev. Genet. 1 (2), 126–133. doi:10.1038/35038565
Jobling, M. A., and Tyler-Smith, C. (1995). Fathers and sons: the Y chromosome and human evolution. Trends Genet. 11 (11), 449–456. doi:10.1016/S0168-9525(00)89144-1
Jobling, M. A., and Tyler-Smith, C. (2003). The human Y chromosome: An evolutionary marker comes of age. Nat. Rev. Genet. 4 (8), 598–612. doi:10.1038/nrg1124
Jung, J. Y., Park, J. H., Oh, Y. L., Kwon, H. S., Park, H. C., Park, K. H., et al. (2016). Forensic genetic study of 29 Y-STRs in Korean population. Leg. Med. (Tokyo) 23, 17–20. doi:10.1016/j.legalmed.2016.09.001
Kanzawa-Kiriyama, H., Jinam, T., Kawai, Y., Sato, T., Hosomichi, K., Tajima, A., et al. (2019). Late Jomon male and female genome sequences from the Funadomari site in Hokkaido, Japan. Anthropol. Sci. 127, 83–108. doi:10.1537/ase.190415
Karafet, T. M., Hallmark, B., Cox, M. P., Sudoyo, H., Downey, S., Lansing, J. S., et al. (2010). Major east-west division underlies Y chromosome stratification across Indonesia. Mol. Biol. Evol. 27 (8), 1833–1844. doi:10.1093/molbev/msq063
Karafet, T., Xu, L., Du, R., Wang, W., Feng, S., Wells, R. S., et al. (2001). Paternal population history of East Asia: Sources, patterns, and microevolutionary processes. Am. J. Hum. Genet. 69 (3), 615–628. doi:10.1086/323299
Kayser, M. (2017). Forensic use of Y-chromosome DNA: A general overview. Hum. Genet. 136 (5), 621–635. doi:10.1007/s00439-017-1776-9
Kayser, M. (2003). The human Y-chromosome - introduction into genetics and applications. Forensic Sci. Rev. 15 (2), 77–89.
Kent, W. J. (2002). BLAT-the BLAST-like alignment tool. Genome Res. 12 (4), 656–664. doi:10.1101/gr.229202
Kutanan, W., Kampuansai, J., Changmai, P., Flegontov, P., Schroder, R., Macholdt, E., et al. (2018). Contrasting maternal and paternal genetic variation of hunter-gatherer groups in Thailand. Sci. Rep. 8 (1), 1536. doi:10.1038/s41598-018-20020-0
Kwon, S. Y., Lee, H. Y., Lee, E. Y., Yang, W. I., and Shin, K. J. (2015). Confirmation of Y haplogroup tree topologies with newly suggested Y-SNPs for the C2, O2b and O3a subhaplogroups. Forensic Sci. Int. Genet. 19, 42–46. doi:10.1016/j.fsigen.2015.06.003
Lall, G. M., Larmuseau, M. H. D., Wetton, J. H., Batini, C., Hallast, P., Huszar, T. I., et al. (2021). Subdividing Y-chromosome haplogroup R1a1 reveals Norse Viking dispersal lineages in Britain. Eur. J. Hum. Genet. 29 (3), 512–523. doi:10.1038/s41431-020-00747-z
Lang, M., Liu, H., Song, F., Qiao, X., Ye, Y., Ren, H., et al. (2019). Forensic characteristics and genetic analysis of both 27 Y-STRs and 143 Y-SNPs in Eastern Han Chinese population. Forensic Sci. Int. Genet. 42, e13–e20. doi:10.1016/j.fsigen.2019.07.011
Mills, R. E., Luttig, C. T., Larkins, C. E., Beauchamp, A., Tsui, C., Pittard, W. S., et al. (2006). An initial map of insertion and deletion (INDEL) variation in the human genome. Genome Res. 16 (9), 1182–1190. doi:10.1101/gr.4565806
Mondal, M., Bergstrom, A., Xue, Y., Calafell, F., Laayouni, H., Casals, F., et al. (2017). Y-chromosomal sequences of diverse Indian populations and the ancestry of the Andamanese. Hum. Genet. 136 (5), 499–510. doi:10.1007/s00439-017-1800-0
Nachman, M. W., and Crowell, S. L. (2000). Estimate of the mutation rate per nucleotide in humans. Genetics 156 (1), 297–304. doi:10.1093/genetics/156.1.297
Naitoh, S., Kasahara-Nonaka, I., Minaguchi, K., and Nambiar, P. (2013). Assignment of Y-chromosomal SNPs found in Japanese population to Y-chromosomal haplogroup tree. J. Hum. Genet. 58 (4), 195–201. doi:10.1038/jhg.2012.159
Ning, C., Yan, S., Hu, K., Cui, Y. Q., and Jin, L. (2016). Refined phylogenetic structure of an abundant East Asian Y-chromosomal haplogroup O*-M134. Eur. J. Hum. Genet. 24 (2), 307–309. doi:10.1038/ejhg.2015.183
Pinotti, T., Bergstrom, A., Geppert, M., Bawn, M., Ohasi, D., Shi, W., et al. (2019). Y chromosome sequences reveal a short beringian standstill, rapid expansion, and early population structure of native American founders. Curr. Biol. 29 (1), 149–157. doi:10.1016/j.cub.2018.11.029
Platt, D. E., Artinian, H., Mouzaya, F., Khalil, W., Kamar, F. G., Matisoo-Smith, E., et al. (2021). Autosomal genetics and Y-chromosome haplogroup L1b-M317 reveal Mount Lebanon Maronites as a persistently non-emigrating population. Eur. J. Hum. Genet. 29 (4), 581–592. doi:10.1038/s41431-020-00765-x
Poznik, G. D. (2016), Identifying Y-chromosome haplogroups in arbitrarily large samples of sequenced or genotyped men. Available at: https://www.researchgate.net/publication/345167690_Identifying_Y-chromosome_haplogroups_in_arbitrarily_large_samples_of_sequenced_or_genotyped_men.[Accessed 11 19 2016].
Song, M., Wang, Z., Zhang, Y., Zhao, C., Lang, M., Xie, M., et al. (2019). Forensic characteristics and phylogenetic analysis of both Y-STR and Y-SNP in the Li and Han ethnic groups from Hainan Island of China. Forensic Sci. Int. Genet. 39, e14–e20. doi:10.1016/j.fsigen.2018.11.016
Stone, A. C., Milner, G. R., Paabo, S., and Stoneking, M. (1996). Sex determination of ancient human skeletons using DNA. Am. J. Phys. Anthropol. 99 (2), 231–238. doi:10.1002/(SICI)1096-8644(199602)99:2<231:AID-AJPA1>3.0.CO;2-1
Tao, R., Li, M., Chai, S., Xia, R., Qu, Y., Yuan, C., et al. (2023). Developmental validation of a 381 Y-chromosome SNP panel for haplogroup analysis in the Chinese populations. Forensic Sci. Int. Genet. 62, 102803. doi:10.1016/j.fsigen.2022.102803
Underhill, P. A., Passarino, G., Lin, A. A., Shen, P., Mirazon Lahr, M., Foley, R. A., et al. (2001). The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations. Ann. Hum. Genet. 65 (1), 43–62. doi:10.1046/j.1469-1809.2001.6510043.x
Underhill, P. A., Shen, P., Lin, A. A., Jin, L., Passarino, G., Yang, W. H., et al. (2000). Y chromosome sequence variation and the history of human populations. Nat. Genet. 26 (3), 358–361. doi:10.1038/81685
Urasin, V., Waas, M., Kull, L., Nogueiro, I., Yacobi, D., and Penninx, W. (2019). Haplogroup J-z640: Genetic insight into the levantine bronze age.
Van Geystelen, A., Decorte, R., and Larmuseau, M. H. (2013). Updating the Y-chromosomal phylogenetic tree for forensic applications based on whole genome SNPs. Forensic Sci. Int. Genet. 7 (6), 573–580. doi:10.1016/j.fsigen.2013.03.010
Walsh, P. S., Metzger, D. A., and Higuchi, R. (1991). Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques 10 (4), 506–513.
Wang, C. C., Jin, L., and Li, H. (2014). Natural selection on human Y chromosomes. J. Genet. Genomics 41 (2), 47–52. doi:10.1016/j.jgg.2014.01.006
Wang, F., Song, F., Song, M., Luo, H., and Hou, Y. (2022). Genetic structure and paternal admixture of the modern Chinese Zhuang population based on 37 Y-STRs and 233 Y-SNPs. Forensic Sci. Int. Genet. 58, 102681. doi:10.1016/j.fsigen.2022.102681
Wang, Y., Zhang, Y. J., Zhang, C. C., Li, R., Yang, Y., Ou, X. L., et al. (2016). Genetic polymorphisms and mutation rates of 27 Y-chromosomal STRs in a han population from guangdong province, southern China. Forensic Sci. Int. Genet. 21, 5–9. doi:10.1016/j.fsigen.2015.09.013
Xue, Y., Wang, Q., Long, Q., Ng, B. L., Swerdlow, H., Burton, J., et al. (2009). Human Y chromosome base-substitution mutation rate measured by direct sequencing in a deep-rooting pedigree. Curr. Biol. 19 (17), 1453–1457. doi:10.1016/j.cub.2009.07.032
Yan, S., Wang, C. C., Li, H., Li, S. L., Jin, L., and Genographic, C. (2011). An updated tree of Y-chromosome Haplogroup O and revised phylogenetic positions of mutations P164 and PK4. Eur. J. Hum. Genet. 19 (9), 1013–1015. doi:10.1038/ejhg.2011.64
Zhang, Y., Zhang, R., Li, M., Luo, L., Zhang, J., Ding, J., et al. (2020). Genetic polymorphism of both 29 Y-STRs and 213 Y-SNPs in han populations from shandong province, China. Leg. Med. (Tokyo) 47, 101738. doi:10.1016/j.legalmed.2020.101738
Zhong, H., Shi, H., Qi, X. B., Duan, Z. Y., Tan, P. P., Jin, L., et al. (2011). Extended Y chromosome investigation suggests postglacial migrations of modern humans into East Asia via the northern route. Mol. Biol. Evol. 28 (1), 717–727. doi:10.1093/molbev/msq247
Zhou, Y., Shao, C., Li, L., Zhang, Y., Liu, B., Yang, Q., et al. (2018). Genetic analysis of 29 Y-STR loci in the Chinese Han population from Shanghai. Forensic Sci. Int. Genet. 32, e1–e4. doi:10.1016/j.fsigen.2017.11.003
Keywords: human genetics, chromosome Y, InDel, haplogroup, O-M175
Citation: Zhou Z, Li Z, Yao Y, Qian J, Ji Q, Shao C and Xie J (2023) Validation of phylogenetic informative Y-InDels in Y-chromosomal haplogroup O-M175. Front. Genet. 14:1182028. doi: 10.3389/fgene.2023.1182028
Received: 08 March 2023; Accepted: 17 April 2023;
Published: 02 May 2023.
Edited by:
Guanglin He, Sichuan University, ChinaReviewed by:
Le Wang, Ministry of Public Security, ChinaCopyright © 2023 Zhou, Li, Yao, Qian, Ji, Shao and Xie. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jianhui Xie, amh4aWVAZnVkYW4uZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.