
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 26 May 2023
Sec. Neurogenomics
Volume 14 - 2023 | https://doi.org/10.3389/fgene.2023.1158028
 Mingxue Wang1
Mingxue Wang1 Yujia Gu2Shuhan Meng1Lixin Kang1Jing Yang3Degang Sun3Yuxing Liu4Ze Wan5Yi Shan6Dongjie Xue7Chang Su8Shufen Li9Ran Yan1Yu Liu2
Yujia Gu2Shuhan Meng1Lixin Kang1Jing Yang3Degang Sun3Yuxing Liu4Ze Wan5Yi Shan6Dongjie Xue7Chang Su8Shufen Li9Ran Yan1Yu Liu2 Yashuang Zhao1*Yonghui Pan10*
Yashuang Zhao1*Yonghui Pan10*Background: Genetic and environmental factors contribute to migraine and the comorbidities of anxiety and depression. However, the association between genetic polymorphisms in the transient receptor potential (TRP) channels and glutamatergic synapse genes with the risk of migraine and the comorbidities of anxiety and depression remain unclear.
Methods: 251 migraine patients containing 49 comorbidities with anxiety and 112 with depression and 600 controls were recruited. A customized 48-plex SNPscan kit was used for genotyping 13 SNPs of nine target genes. Logistic regression was conducted to analyze these SNPs’ association with the susceptibility of migraine and comorbidities. The generalized multifactor dimension reduction (GMDR) was applied to analyze the SNP-SNP and gene-environment interactions. The GTEx database was used to examine the effects of the significant SNPs on gene expressions.
Results: The TRPV1 rs8065080 and TRPV3 rs7217270 were associated with an increased risk of migraine in the dominant model [ORadj (95% CI): 1.75 (1.09–2.90), p = 0.025; 1.63 (1.02–2.58), p = 0.039, respectively]. GRIK2 rs2227283 was associated with migraine in the edge of significance [ORadj (95% CI) = 1.36 (0.99–1.89), p = 0.062]. In migraine patients, TRPV1 rs222741 was associated with both anxiety risk and depression risk in the recessive model [ORadj (95% CI): 2.64 (1.24–5.73), p = 0.012; 1.97 (1.02–3.85), p = 0.046, respectively]. TRPM8 rs7577262 was associated with anxiety (ORadj = 0.27, 95% CI = 0.10–0.76, p = 0.011). TRPV4 rs3742037, TRPM8 rs17862920 and SLC17A8 rs11110359 were associated with depression in dominant model [ORadj (95% CI): 2.03 (1.06–3.96), p = 0.035; 0.48 (0.23–0.96), p = 0.042; 0.42 (0.20–0.84), p = 0.016, respectively]. Significant eQTL and sQTL signals were observed for SNP rs8065080. Individuals with GRS (Genetic risk scores) of Q4 (14–17) had a higher risk of migraine and a lower risk of comorbidity anxiety than those with Genetic risk scores scores of Q1 (0–9) groups [ORadj (95% CI): 2.31 (1.39–3.86), p = 0.001; 0.28 (0.08–0.88), p = 0.034, respectively].
Conclusion: This study suggests that TRPV1 rs8065080, TRPV3 rs7217270, and GRIK2 rs2227283 polymorphism may associate with migraine risk. TRPV1 rs222741 and TRPM8 rs7577262 may associate with migraine comorbidity anxiety risk. rs222741, rs3742037, rs17862920, and rs11110359 may associate with migraine comorbidity depression risk. Higher GRS scores may increase migraine risk and decrease comorbidity anxiety risk.
Migraine is a multifactorial neurovascular disorder in which genetic factors play a relevant role in both predisposing and determining underlying mechanisms. According to the Global Burden of Disease (GBD) Survey 2019, migraine ranked the second highest specific cause of disability worldwide and the first among women aged 15–49 years (Steiner et al., 2020).
Recently, the association of genetic, environmental, and migraine and its comorbidities of anxiety and depression have attracted wide attention (Louter et al., 2015). The comorbidity of depression and migraine may involve abnormal brain development, a common genetic basis, 5-hydroxytryptamine, sex hormones, and other mechanisms (Zhang et al., 2019; Yang et al., 2022; Lv et al., 2023). The calcitonin gene-related peptide (CGRP) is recognized as a critical player in the pathophysiology of migraine, which can trigger migraine attacks (Wattiez et al., 2020; Rees et al., 2022). Meanwhile, CGRP can act on the bed nucleus of the stria terminals (BNST) to evoke anxiety-like responses, activate anxiety-related structural neurons, and modulate behavioral responses to stress in rats (Sink et al., 2011). Given the common pathogenesis of anxiety, depression, and migraine, we hypothesize that there may be specific genetic characteristics sharing between migraine and the comorbidities of anxiety and depression.
Transient receptor potential (TRP) channels are a family of cation channels expressed primarily on the cell membrane. Activation of TRP channels is well-known to promote the release of CGRP from sensory nerve endings (Shibata and Tang, 2021). They have been used extensively as probes for the function of CGRP in various processes (Russell et al., 2014). Further, TRP channels have an essential role in migraine pain and associated symptoms, such as hyperalgesia and allodynia (Spekker et al., 2022). Several studies have explored the role of TRP channels gene polymorphisms in migraine across ethnic groups but with inconsistent results (Carreño et al., 2012; Chasman et al., 2014; Chen et al., 2018; Fu et al., 2019; Gavva et al., 2019; Yakubova et al., 2021; Siokas et al., 2022). Animal studies suggest that TRPA1 and TRPV1 antagonism might be a target for the treatment of anxiety and depression (de Moura et al., 2014; Escelsior et al., 2020; Ngoc et al., 2023). TRPA1 mRNA was colocalized with Edinger-Westphal nucleus (EWcp)/urocortin 1 (UCN1) neurons, and TRPA1 may contribute to the regulation of depression-like behavior and stress-adaptative responses in mice (Kormos et al., 2022). Moreover, it is well known that TRPV1 mediates glutamate in the brain (Fawley et al., 2014) and deficiency of TRPV1 induces antidepressant and anxiolytic effects by altering the expression of serotonin, gamma-aminobutyric acid (GABA) and glutamate N-methyl-d-aspartate (NMDA) receptors (You et al., 2012). The glutamate NMDA receptors play a relevant role in allodynia to mechanical stimuli, while TRPA1 channels may associate with NMDARs to promote ascending acute and chronic pain signals and to control mu-opioid receptors antinociception (Cortés-Montero et al., 2020). While the effects of TRPV1 on anxiety and depression may be related to cannabinoid 1 (CB1) receptor interactions (Sartim et al., 2017) and regulation of the glutamate/NMDA pathway (Lisboa and Guimarães, 2012).
In addition, malfunctions of the glutamatergic system may contribute to migraine symptoms (Gasparini et al., 2016). Glutamate can initiate migraine by cortical spreading depression (CSD), which is the lynchpin of migraine aura (Charles and Baca, 2013; Eikermann-Haerter, 2021). Thus far, only a few studies have focused on the relationship between the polymorphisms in ionotropic glutamate receptor genes (GRIA1 and GRIA3) and migraine (Formicola et al., 2010; Maher et al., 2013; Fang et al., 2015). As the critical hub in neuronal metabolism, signaling, and plasticity, the role of glutamatergic synapse gene polymorphism in migraine is still not yet clarified (Frenguelli, 2022). Growing evidence supports the use of the NMDA receptor antagonist ketamine as a rapid-acting antidepressant, facilitating research into glutamate signaling modulators as depression treatment agents (Murrough et al., 2017; Hashimoto, 2019). However, the association of these genetic polymorphisms with migraine comorbidities anxiety, and depression remains unclear.
In summary, TRP is associated with hyperalgesia and allodynia, which mediates glutamate in the brain and can trigger migraine through CSD; moreover, activation of TRP channels can promote the release of CGRP, which can trigger migraine attacks. Therefore, we focused on candidate genes of TRP channels and glutamatergic synapses related to the proposed pathways mechanisms of migraine. This study aimed to examine the relationship between the polymorphisms of target 10 genes and migraine, migraine with or without aura, and anxiety and depression of migraine through a case control study in China.
Subjects in this case control study were recruited from four hospitals (the First Affiliated Hospital of Harbin Medical University and three hospitals of Agricultural Reclamation, Heilongjiang Province: Hong Xinglong Administration Hospital, Bao Quanling Central Hospital, and Bao Quanling Farm Hospital) in northern China. The case group included 251 migraine patients, of which 49 patients were combined with anxiety and 112 were combined with depression. All the migraine patients were diagnosed according to the International Classification of Headache Disorders, 3rd edition, beta version (ICHD-3β version) criteria (Headache Classification Committee of the International Headache, 2013). The control group consisted of 600 nonmigraine subjects from the physical examination center of the same hospital in the same period. Considering the effect of age and especially gender differences on the results, we further conducted a sensitive analysis by matching the cases and controls at a 1:2 frequency based on sex concordance and age within 2 years. We conducted a questionnaire that included general basic information, lifestyle, medical history, anxiety, depression, and sleep status. All subjects signed informed consent prior to the study, and the study was approved by the Ethics Committee of Harbin Medical University.
Three self-assessment scales were used to assess anxiety, depression, and sleep quality, respectively. Self-Rating Anxiety Scale (SAS), which contains a 20-item questionnaire scored on a 4-point scale was used (Zung, 1971). The SAS scores were the raw score multiplied by 1.25. Then the SAS scores were divided into no anxiety (25–49 points), mild anxiety (50–60 points), moderate anxiety (61–70 points), and severe anxiety (71–100 points). The Patient Health Questionnaire-9 (PHQ-9) was used to assess depressive symptoms (Kroenke et al., 2001). PHQ-9 scores are divided into no depression (0–4 points), mild depression (5-9 points), moderate depression (10–14 points), and moderate to severe depression (15–27 points). The Pittsburgh Sleep Quality Index (PSQI) is composed of 19 items, which are classified into 7 components weighted from 0 to 3 (Buysse et al., 1989). PSQI score <5 indicated good sleep quality.
Based on the data from the previous literature and combined with the dbSNP database, 15 SNPs in 10 genes were identified (Table 1). The 8 tag SNPs, were selected through the dbSNPs database (http://www.ncbi.nlm.nih.gov/) with minor allele frequency (MAF) > 0.1 and R2 > 0.8 in CHB (Han Chinese in Beijing) and CHS (Han Chinese in Southern). There is no significant linkage disequilibrium (R2 < 0.8) among the candidate SNPs.
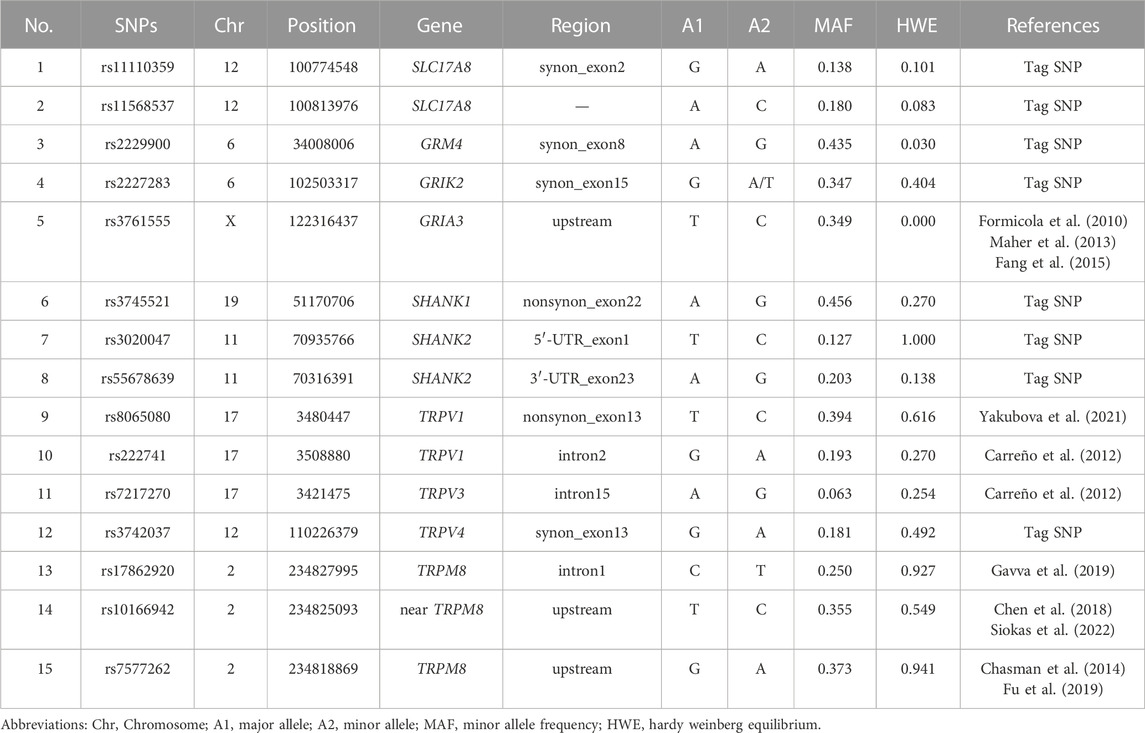
TABLE 1. Detailed information on 15 SNPs of TRP channels and glutamatergic synapse genes.
We collected 2 mL venous blood samples from all participants, separated out serum by centrifugation, and stored it at −80°C. The genomic DNA was extracted using the QIAamp® DNA Blood Mini Kit (Qiagen, Germany). The extracted DNA was diluted to a working concentration of 30 ng/μl for further genotyping. The purity and concentration of DNA samples were measured by a Nanodrop 2000 spectrophotometer (Thermo Scientific, United States). Genotyping was performed using a customized 48-plex SNPscan kit (Gene Sky Biotechnologies Inc., Shanghai, China), which used double ligation and multiplex fluorescent PCR. We randomly chose 7% (58/851) of the samples and genotyped twice to ensure the accuracy and repeatability of genotyping, which were more than 95.0% (Supplementary Table S1).
The Hardy-Weinberg equilibrium (HWE) was analyzed by Fisher’s test using the GWASExactHW package of R 4.0.3 (Wigginton et al., 2005). We conducted logistic regression to examine the association between SNPs and the number of risk alleles and migraine. The gene-gene interaction analysis was conducted by GMDR software Beta version 0.7 (http://www.ssg.uab.edu/gmdr/). Power analysis using PASS 15 software. Statistical analyses were performed using statistical software R 4.0.3. p < 0.05 was considered statistically significant. Multiple testing adjustments were controlled by Bonferroni correction (0.05/13).
To further examine the potential function of the significant SNPs on gene expression, we utilized the GTEx database (https://gtexportal.org/home/) to investigate the tissue-specific expression quantitative trait loci (eQTL) and splicing quantitative trait loci (sQTL). The Genotype-Tissue Expression (GTEx) project aims to collect and analyze multiple human tissues that are densely genotyped and analyzed for global RNA expression (Consortium, 2015). We also used the website (https://snpinfo.niehs.nih.gov/) to make functional predictions for significant SNPs (Xu and Taylor, 2009).
The basic characteristics of all participants are summarized in Table 1; Supplementary Table S8, which include characteristics such as age, gender, lifestyle, and history of the disease. We recruited 600 controls and 251 migraine patients, of which 51 (20.32%) had migraine with aura (MA) and 200 (79.68%) had migraine without aura (MO). We observed that the migraine groups had a higher proportion of women, a higher proportion of anxiety, depression, and poor sleep quality (p = 0.001, p = 0.002, p < 0.001, p < 0.001, respectively) than the controls. Moreover, migraine patients had a lower proportion of alcohol consumption and exercise habits (p = 0.015, p = 0.001, respectively). Of the 251 migraines, 49 (19.52%) were combined with anxiety, and 112 (44.62%) were combined with depression. Among 851 participants, 281 cases were anxiety or depression including 123 (43.77%) in migraines and 158 (56.23%) in controls. Compared to the controls, the features of the MA and MO groups were almost the same as migraineurs. There were no significant differences between the MA and MO groups in terms of age and sex, but PHQ-9 scores (5.28 ± 4.66 vs. 3.64 ± 4.03, p = 0.024) and PSQI scores (5.77 ± 3.11 vs. 4.56 ± 2.72, p = 0.013) differed between the two groups (Table 2).
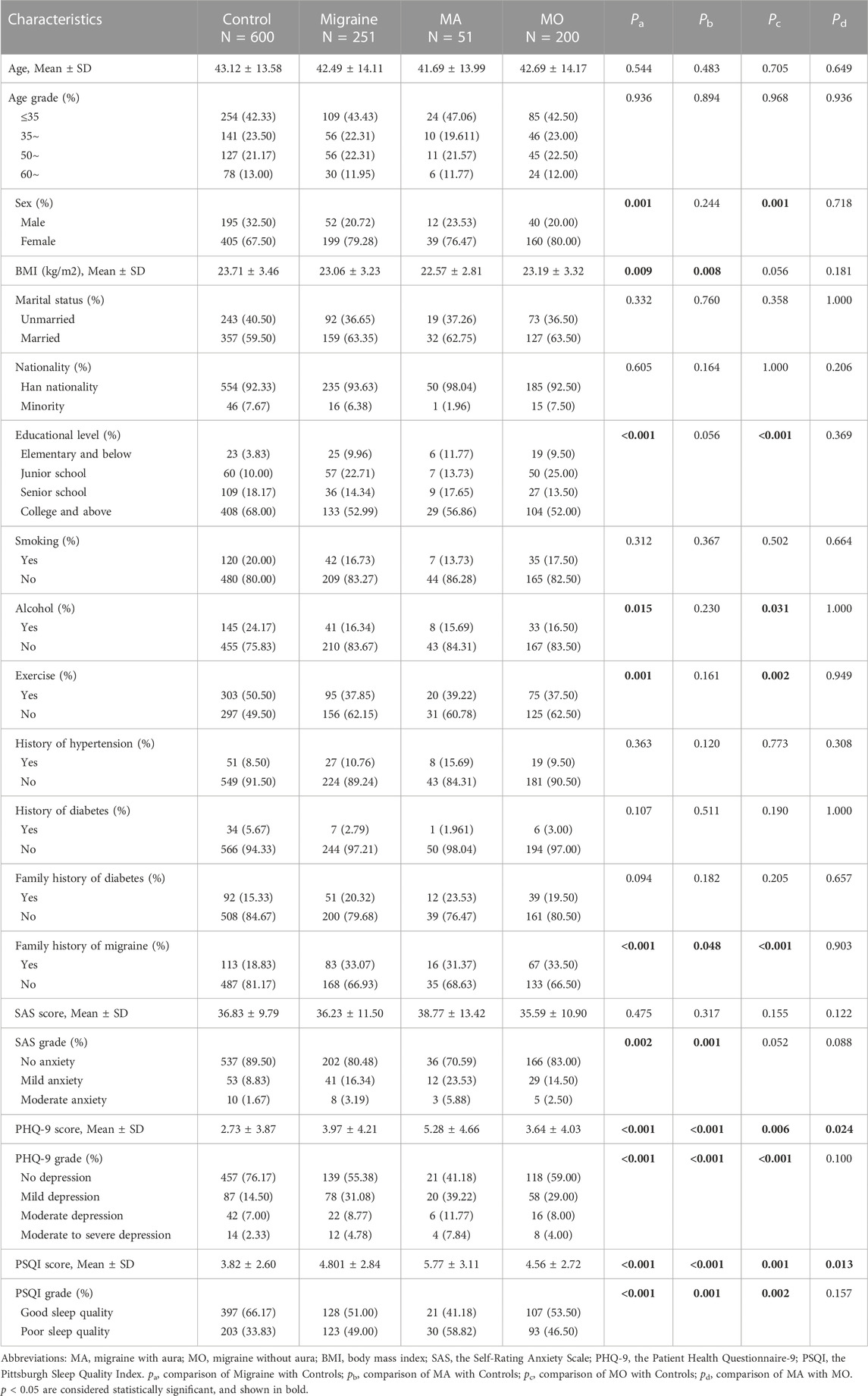
TABLE 2. Basic demographic and clinical characteristics of the migraine, MA, MO and control groups.
The results showed that except for the GRM4-rs2229900 and GRIA3-rs3761555 loci which did not conform to the HWE (p < 0.05), the remaining 13 SNPs were all agreed with the HWE (p > 0.05) and were included in the subsequent analysis (Table 1).
As shown in Table 3, TRPV1 rs8065080 was associated with an increased risk of migraine in the dominant model (CC + TC vs. TT: ORadj = 1.78, 95% CI = 1.09–2.90, p = 0.025). TRPV1 rs8065080 genotype TT decreased migraine risk compared to wild type CC genotype (ORadj = 0.55, 95% CI = 0.32–0.92, p = 0.025). TRPV3 rs7217270 was associated with increased migraine risk based on the dominant model (AA + AG vs. GG: ORadj = 1.63, 95% CI = 1.02–2.58, p = 0.039). However, none of the associations was significant after Bonferroni correction for multiple tests with p < 0.0038 (0.05/13). GRIK2 rs2227283 was associated with migraine in the edge of significance [ORadj (95% CI) = 1.36 (0.99–1.89), p = 0.062]. In univariate analysis, TRPM8 rs7577262 and TRPM8 rs17862920 were associated with migraine [OR (95% CI): 0.68 (0.49–0.93), p = 0.016; 0.71 (0.52–0.97), p = 0.031, respectively]. However, after adjusting for confounders these associations narrowly avoided significance with p of 0.120 and 0.141. Additional nonsignificant results are in the Appendix (Supplementary Table S2).
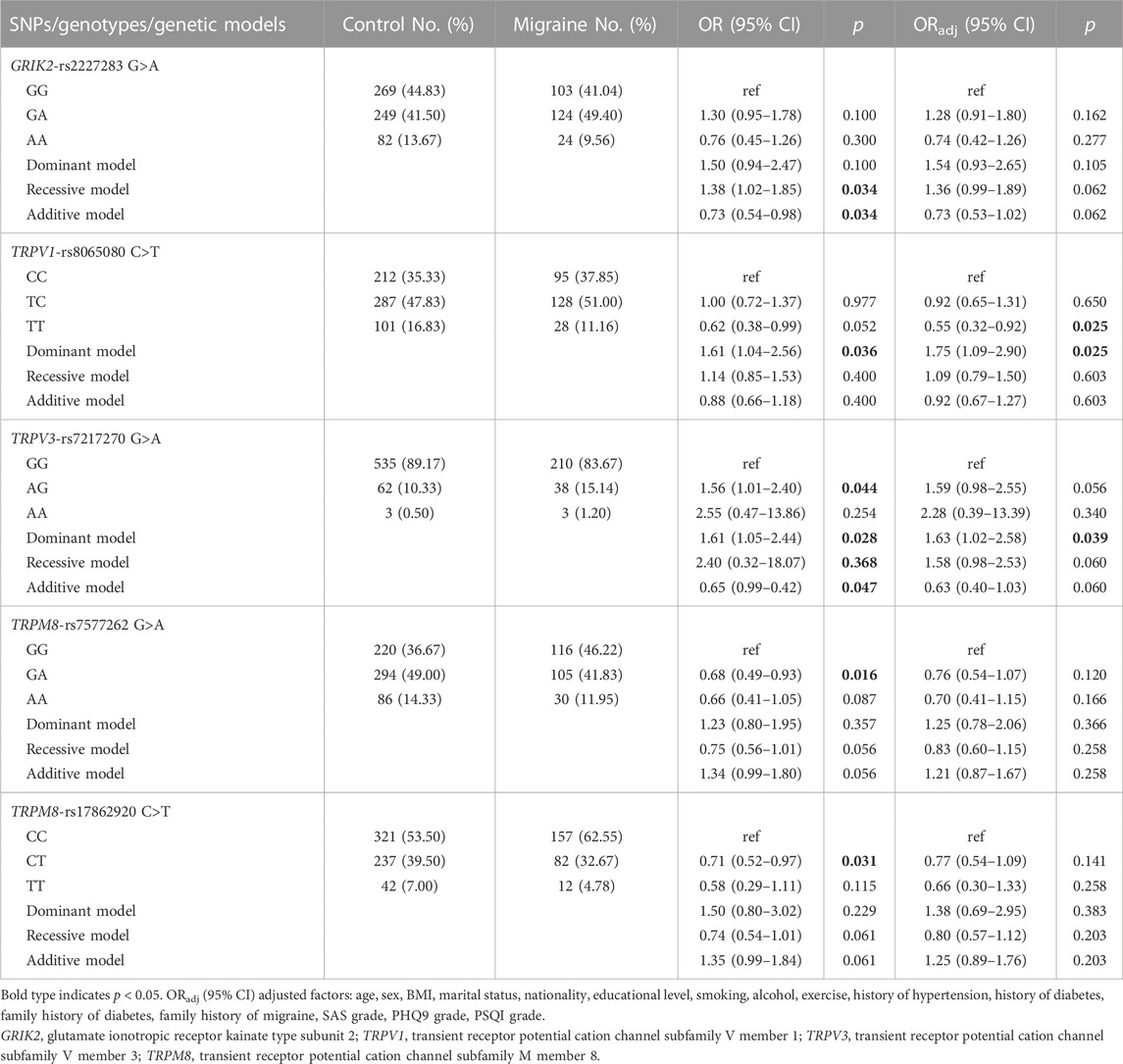
TABLE 3. Associations between the GRIK2, TRPV1, TRPV3 and TRPM8 gene polymorphisms and the risk of migraine.
Furthermore, we conducted a sensitivity analysis by matching the cases and controls at a 1:2 frequency based on sex and age within 2 years. Consistently, we observed only three of the five SNPs (TRPV3 rs7217270, TRPM8 rs7577262, and TRPM8 rs17862920) in association with migraine and in the same direction as the unmatched data results, but possibly marginally significant after multifactorial adjustment with p of 0.058, 0.110 and 0.105, respectively. Differently, the associations of TRPV1 rs8065080 and GRIK2 rs2227283 with migraine were not significant due to the small sample size in matched data (Supplementary Table S8).
In addition, the results of subgroup analysis showed that the frequency of GA genotype of SLC17A8 rs11110359 was marked higher in the MO group compared to the control (ORadj = 1.56, 95% CI = 1.03–2.34, p = 0.034) and showed a significant risk for MO patients in three models (Dominant: ORadj = 1.51, 95% CI = 1.02–2.24, p = 0.040; Recessive: ORadj = 1.55, 95% CI = 1.03–2.33, p = 0.036; Additive: ORadj = 0.65, 95% CI = 0.43–0.98, p = 0.036, respectively). The CC genotype of SLC17A8 rs11568537 was associated with decreased MO risk compared to the AA genotype (ORadj = 0.22, 95% CI = 0.05–0.70, p = 0.021) and increased MO risk in the dominant model (ORadj = 4.40, 95% CI = 1.42–19.57, p = 0.023). The genotype TT of TRPV1 rs8065080 was associated with decreased MO risk (ORadj = 0.55, 95% CI = 0.31–0.97, p = 0.043). rs8065080 increased MO risk in the dominant model (CC + TC vs. TT: ORadj = 1.82, 95% CI = 1.09–3.17, p = 0.023). TRPV3 rs7217270 and TRPM8 rs17862920 were associated with MO risk in a very slight trend toward significance with p of 0.056 and 0.169. SHANK1 rs3745521 and TRPM8 rs7577262 were associated with MA risk and tended to approach significance with p of 0.070 and 0.081 (Table 4). However, all the p-values did not withstand Bonferroni correction.
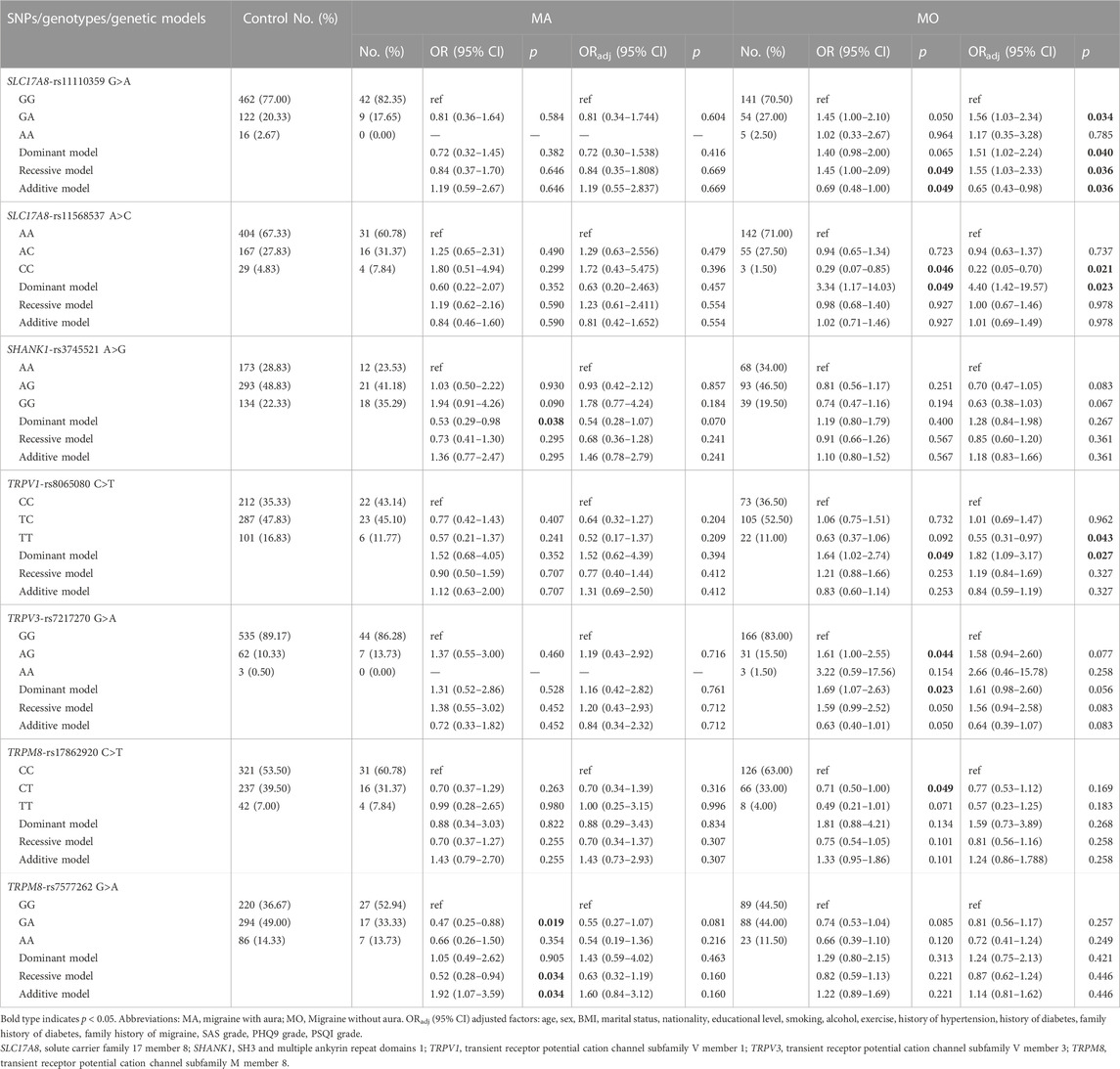
TABLE 4. Associations between SLC17A8, SHANK1, TRPV1, TRPV3 and TRPM8 gene polymorphisms and the risk of migraine by aura.
Consistent with the above results, the results of matched subjects showed that three SNPs (SLC17A8 rs11110359, SLC17A8 rs11568537, and TRPV3 rs7217270) were associated with MO, and TRPM8 rs7577262 was associated with MA. Differently, TRPV1 rs8065080 was associated with MA rather than MO. And GRIK2 rs2227283 was associated with MA rather than migraine (Supplementary Table S9).
The relationship between the gene polymorphisms of TRP channels and glutamatergic synapse genes and migraine comorbidities risk was demonstrated by stratification analysis, as shown in Table 5; Supplementary Tables S4–S6. In migraine patients, the TRPM8 rs7577262 was associated with decreased anxiety risk in a dominant model (ORadj = 0.27, 95% CI = 0.10–0.76, p = 0.011). The genotype GA of TRPV1 rs222741 was associated with increased anxiety risk as compared to the AA genotype (ORadj = 2.72, 95% CI = 1.26–5.99, p = 0.011). TRPV1 rs222741 was associated with both anxiety risk and depression risk in the recessive model (ORadj = 2.64, 95% CI = 1.24–5.73, p = 0.012; ORadj = 1.97, 95% CI = 1.02–3.85, p = 0.046, respectively) and additive model (ORadj = 0.38, 95% CI = 0.17–0.81, p = 0.012; ORadj = 0.51, 95% CI = 0.26–0.98, p = 0.046, respectively). SLC17A8 rs11110359, TRPV4 rs3742037 and TRPM8 rs17862920 were associated with depression in dominant model (ORadj = 0.42, 95% CI = 0.20–0.84, p = 0.016; ORadj = 2.03, 95% CI = 1.06–3.96, p = 0.035; ORadj = 0.48, 95% CI = 0.23–0.96, p = 0.042, respectively). TRPM8-rs10166942 was associated with depression risk in recessive model (ORadj = 0.51, 95% CI = 0.27–0.94, p = 0.034) and additive model (ORadj = 1.98, 95% CI = 1.06–3.77, p = 0.034) (Table 5). Other not significant results were shown in Supplementary Table S4. In nonmigraine subjects, only SHANK2 rs55678639 was associated with depression (Supplementary Table S5). In anxiety or depression patients, TRPM8 rs7577262 was still associated with migraine (Supplementary Table S6).
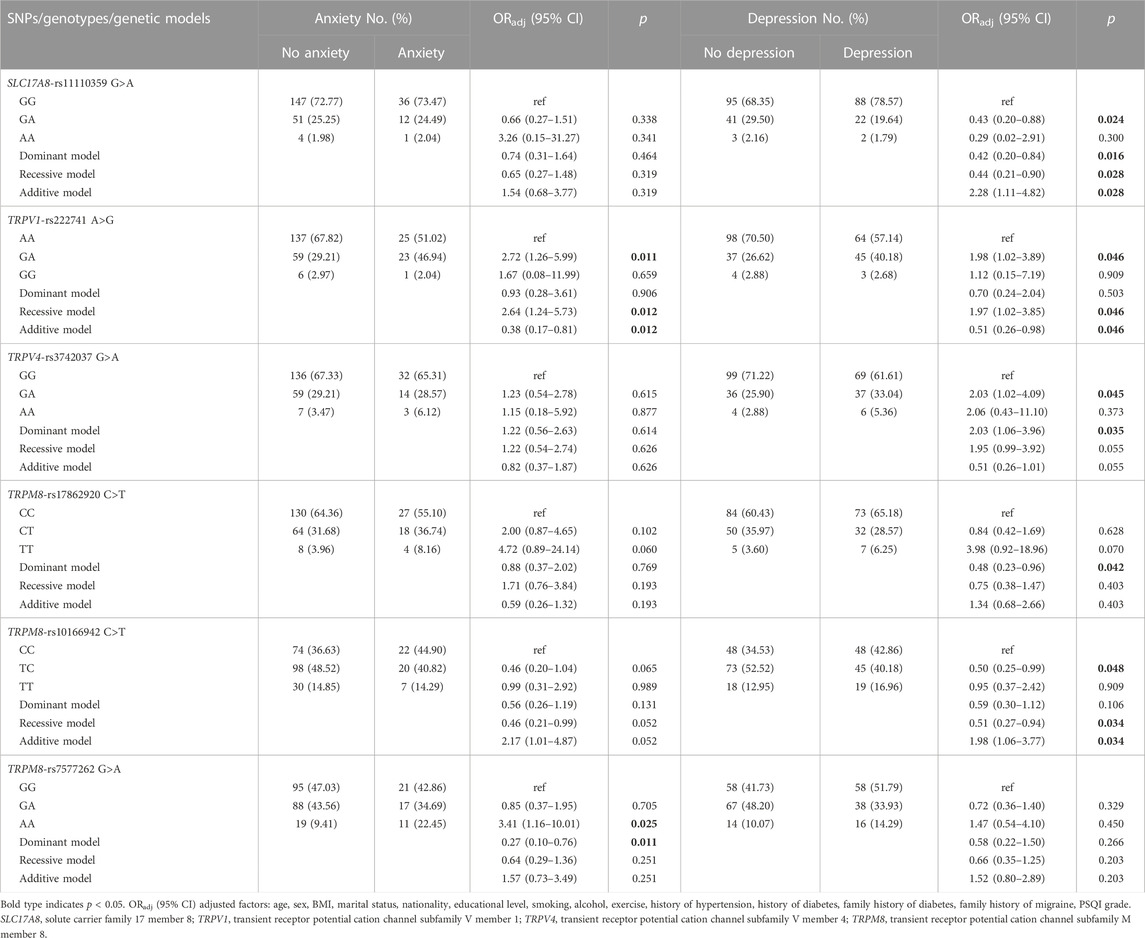
TABLE 5. Associations between SLC17A8, TRPV1, TRPV4 and TRPM8 gene polymorphisms and anxiety and depression in migraine patients.
The GRS was significantly higher in the migraine group compared with the control group (mean GRS: 11.47 ± 2.45 vs. 10.91 ± 2.42, p = 0.002) and there was a positive trend (p trend = 0.002). Compared to the GRS scores of the Q1 (0–9) group, individuals with GRS scores of Q4 (14–17) had a higher risk of migraine (ORadj = 2.31, 95% CI = 1.39–3.86, p = 0.001) but not significant in GRS scores of Q2 (10–11) groups (ORadj = 1.41, 95% CI = 0.90–2.21, p = 0.136) or Q3 (12–13) groups (ORadj = 1.41, 95% CI = 0.89–2.24, p = 0.149) (Table 6). However, GRS scores of Q3 and Q4 groups had a lower risk of the comorbidity of anxiety than GRS scores of Q1 group (ORadj = 0.31, 95% CI = 0.11–0.89, p = 0.032; ORadj = 0.28, 95% CI = 0.08–0.89, p = 0.034, respectively) and the p trend was 0.020 (Table 7). A significant GRS score risk trend for comorbid depression was found (p trend = 0.025) though not significant in three GRS groups (p = 0.939, p = 0.137, p = 0.068, respectively) (Table 7).
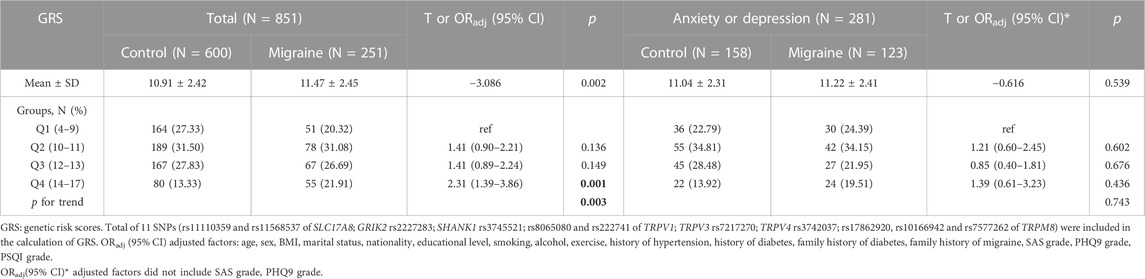
TABLE 6. The relationship between the GRS and migraine susceptibility.
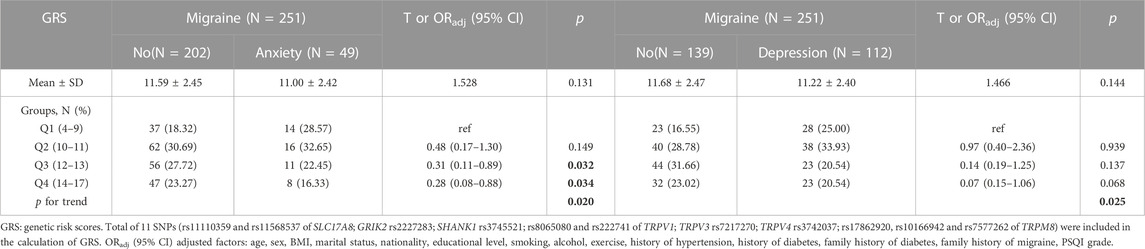
TABLE 7. The relationship between the GRS and migraine comorbidities of anxiety and depression susceptibility.
GMDR model was used to evaluate the effect of SNP-SNP and gene-environment interaction on migraine risk. As shown in Table 8, we found a three-locus SNP-SNP interaction model (rs11568537× rs17862920× rs2227283) contributed to the best model with the most excellent cross-validation consistency of 9/10 (p = 0.0010) (Supplementary Figure S1A). For gene-depression interaction, an important four-locus model (rs10166942× rs3745521× rs8065080× Depression) contributed to the best model (p = 0.0010) (Supplementary Figure S1B). We further assessed the combined effect of the six risk SNPs and depression on migraine risk. Compared with subjects with no depression and zero to six risk alleles, migraine patients with depression and seven to twelve risk alleles had the highest migraine risk (ORadj = 4.06, 95% CI = 2.48–6.81, p < 0.001) (Figure 1).
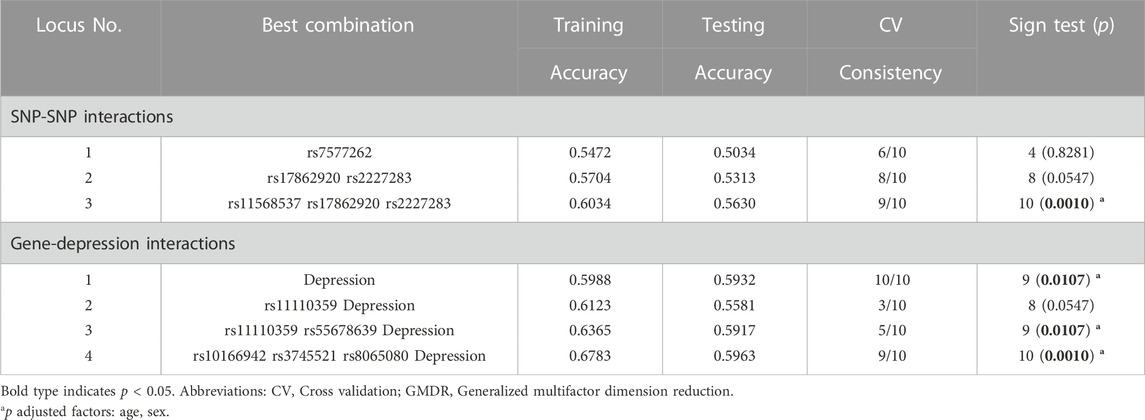
TABLE 8. GMDR analysis for the best SNP-SNP and gene-depression interaction models.
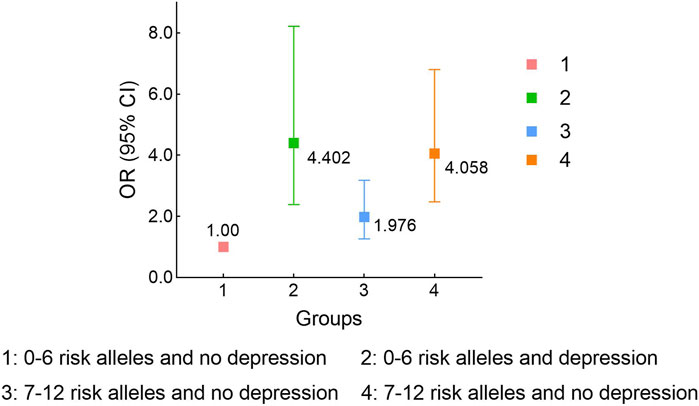
FIGURE 1. Stratified analysis for rs11568537, rs17862920, rs2227283, rs10166942, rs3745521, rs8065080 and depression interaction on migraine risk using logistic regression. Adjusted for age and sex.
We identified seven significant sQTL signals for rs8065080 on TRPV1 (Table 9). The C allele of rs8065080 was related to the increased level of alternative splicing of the pre-mRNA for gene TRPV1 from multiple types of human tissues. In addition, significant eQTL signals were identified for SNPs rs2227283, rs7577262, rs8065080 and rs7217270 (Supplementary Tables S10–S13). The threshold of p values was 0.005 (0.05/10) or 0.001 (0.05/46 or 0.05/49). Four significant signals (4/49) were identified for SNP rs8065080 and gene expression levels of TRPV1 (Supplementary Table S1). The T allele of rs8065080 was significantly associated with an increased gene expression level of TRPV1 in the skin-sun exposed (lower leg) (Supplementary Figure S2A, p = 2.30 × 10−5) and in the skin-not Sun exposed (suprapubic) (SSupplementary Figure S2B, p = 5.20 × 10−5). However, no statistically significant was found between TRPV1 gene expression and neural or brain tissues.
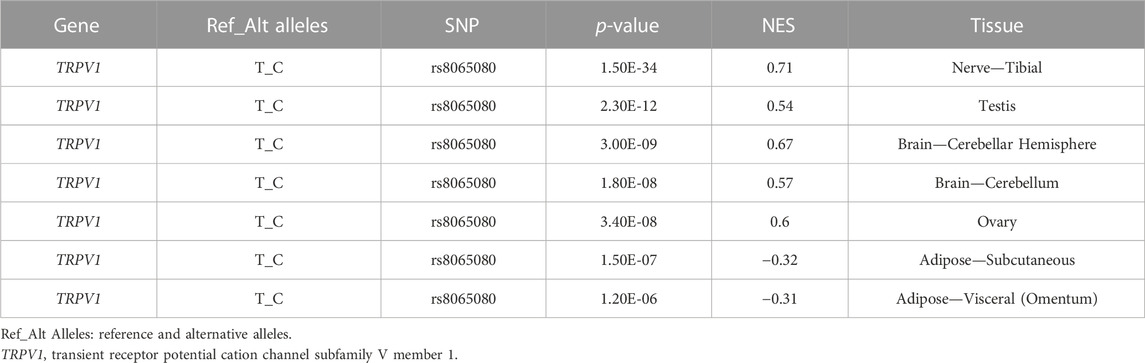
TABLE 9. Significant sQTL signals of SNP rs8065080 on TRPV1.
In addition, we found that SLC17A8 rs11110359 and TRPV1 rs8065080 are located at exonic splicing enhancer (ESE) or exonic splicing silencer (ESS) -binding sites (Supplementary Table S1). We found several microRNA binding sites For SLC17A8 rs11568537 and SHANK2 rs55678639 (Supplementary Table S5).
We calculated the power of the study for 13 SNPs using odds ratios (OR) of 1.8, 2.0, and 3.0 to assess whether the sample size of this study was sufficient to be biologically meaningful. Except for SNP rs7217270, the power analysis indicated that our sample size achieved sufficient power (>0.831) to conclude with OR 1.8 or higher (Table 10). As for subgroup analysis, except for SNP rs7217270 and SNP rs11110359, the power was higher than 80% in groups of MO vs. Control and not approaching 80% in groups of MA vs. Control with OR 1.8 or higher (Table 10).
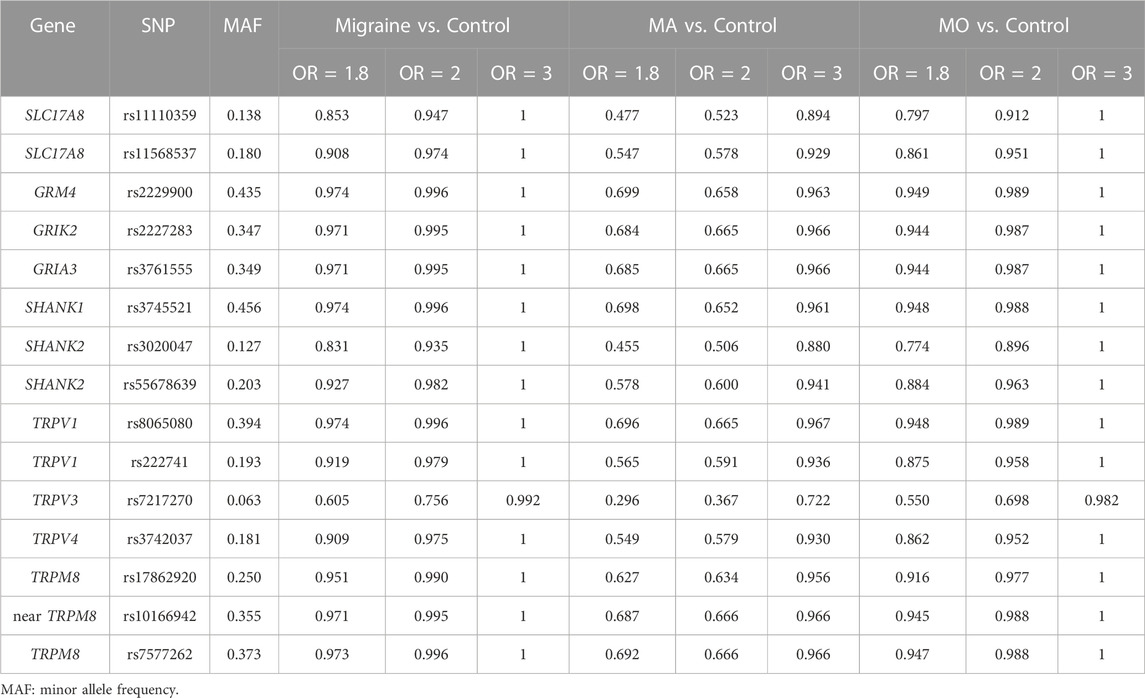
TABLE 10. Power of the study with different odds ratios (OR) in an allelic model.
In this case control study, the GRIK2 rs2227283 was the first time studied and found to be associated with migraine risk in the edge of significance. More importantly, we first found that TRPV1 rs222741 was associated with migraine comorbidities both anxiety and depression. TRPM8 rs7577262 was associated with migraine comorbidity anxiety. TRPV4 rs3742037, TRPM8 rs17862920, TRPM8 rs10466942 and SLC17A8 rs11110359 were associated with migraine comorbidity depression risk. In anxiety or depression patients, rs7577262 was still related to the susceptibility of migraine. However, in controls, only SHANK2 rs55678639 was associated with depression. These results further indicate that migraine and its comorbidities of anxiety and depression may share common genetic traits.
Further, we found one novel SHANK1 rs3745521 was associated with migraine with aura risk, and two novel SNPs (rs11110359 and rs11568537 of SLC17A8 gene) were associated with the susceptibility of migraine without aura risk. The present study showed that TRPV1 rs8065080, TRPV3 rs7217270, and TRPM8 rs17862920 were significantly associated with migraine or MO risk. TRPM8 rs7577262 was associated with migraine or MA risk. These findings confirm the results of previous studies (Carreño et al., 2012; Chasman et al., 2014; Fu et al., 2019; Gavva et al., 2019). However, the associations between MA and MO may not be trustworthy enough for the sample size in the subgroups was not so large. There are no common SNPs associated with both MA and MO in these two pathways, suggesting that different pathways may be involved in the MA/MO susceptibility.
TRPV1 and TRPV4 are abundantly expressed in primary sensory neurons and play a primary role in migraine pain (Iannone et al., 2022). TRPM8 encodes for a receptor-activated non-selective cation channel activated by cold environmental temperatures and is related to pain sensor channels (Dussor and Cao, 2016). As the main members of the TRP family, the polymorphisms of TRPV1, TRPV4 and TRPM8 genes may play an important role in the occurrence and development of migraine. rs222741 (TRPV1) and rs7577262 (TRPM8) were associated with migraine overall (Carreño et al., 2012; Chasman et al., 2014). We also found that TRPM8 rs7577262 was associated with migraine risk. However, rs7577262 was not a risk factor for migraines in a small sample of the She ethnic population in China (Fu et al., 2019). Anxiety and depression are the most frequent psychiatric comorbidities in migraine (Bergman-Bock, 2018). We found that the TRP gene polymorphisms in migraine comorbidity of anxiety and depression were different except for TRPV1 rs222741. It may be that the mechanisms underlying these two comorbidities are not quite identical. One study suggested that similar clinical phenotypes of anxiety and depression rely on prefrontal alterations, whereas frontotemporal and parietal abnormalities may represent unique features of the two (Maggioni et al., 2019).
GRIK2 rs2227283 was associated with migraine risk in unmatched data and associated with MA risk in matched data. The GRIK2 gene is located in the 6q16.3 region and encodes for kainate receptors (KARs) class ionotropic glutamate receptor 6 (GluR6). GRIK2 contributes to inhibitory transmission, regulates excitatory responses and glucose homeostasis, and plays an important role in synaptic physiology (Barbon et al., 2001; Abarkan et al., 2019). Previous studies showed that the GRIK2 gene was associated with somatic anxiety, recurrent epileptic seizures, autism, and cognitive abilities (Myung et al., 2012; Yuan et al., 2015; Chandra et al., 2019; Henley et al., 2021). A recent study provided further evidence that the rs2227283 polymorphism of the GRIK2 gene was significantly related to aggressive behaviors in bipolar manic patients (Ma et al., 2019). Therefore, we hypothesize that polymorphisms in this gene contribute to migraine onset by affecting neurodevelopment.
This study showed that the polymorphisms of SLC17A8 may increase MO risk. SLC17A8 encodes the vesicular glutamate transporter 3 (VGLUT3), expressed in discrete populations of glutamatergic, cholinergic, serotonergic, and even GABAergic neurons (Favier et al., 2021). Genetic deletion of VGLUT3 triggers deficits in acute and persistent mechanical pain and inflammatory pain (Seal et al., 2009; Draxler et al., 2014). Chronic pain decreases VGLUT3 levels (Tukey et al., 2013). Moreover, Brain-derived neurotrophic factor (BDNF) might regulate neuropathic pain through the upregulation of VGLUT3 by activation of the phospholipase C-gamma (PLC-γ) signaling pathway (Liu et al., 2014). VGLUT-3 expression was upregulated in the conditions of diabetes complicated by depression (Liu et al., 2021). Interestingly, we found that SLC17A8 rs11110359 polymorphisms were associated with migraine combined depression. The metabotropic glutamate receptors (mGluRs) involved in the regulation of synaptic transmission and neuronal excitability throughout the central nervous system and the targeted drug has therapeutic effect on depression (Niswender and Conn, 2010). Our findings may provide genetic evidence for this mechanism.
Our findings suggested that polymorphisms of SHANK1 (rs3745521) might be significantly correlated with MA susceptibility. SH3 and multiple ankyrin repeat domains proteins SHANK1, SHANK2, and SHANK3 encode a family of postsynaptic scaffolding proteins present at glutamatergic synapses and play a crucial role in synaptogenesis (Liu et al., 2022). The expression and loss of allodynia and hyperalgesia are closely associated with changes in Homer1b/c and Shank1a levels in the spinal dorsal horn (Miletic et al., 2005). Moreover, SHANK2 knock-out mice showed reduced tactile perception and analgesia to chronic pain in autism spectrum disorder (Ko et al., 2016). The SHANK genes have been extensively studied in neuropsychiatric disorders such as autism and schizophrenia, but the mechanism of the role of SHANK in migraine is unclear (Schmeisser, 2015; Wan et al., 2022).
Moreover, our GRS analysis also revealed that the higher GRS scores increased migraine risk and decreased comorbidity anxiety risk. A significant GRS score risk trend for comorbid depression was found though nonsignificant between GRS groups, which was due to the small sample size of our cases. Furthermore, we identified one significant SNP-SNP interaction model (rs11568537× rs17862920× rs2227283) and a significant gene-depression interaction model (rs10166942× rs3745521× rs8065080× Depression) which contributed to the best model. We found that more risk loci and depression increased migraine risk, suggesting that genetic factors and migraine comorbidity depression combine to increase migraine risk. In addition, we investigated the potential functional consequences of the SNPs GRIK2 rs2227283, TRPV1 rs8065080, TRPV3 rs7217270 and TRPM8 rs7577262 through eQTL and sQTL data obtained from the GTEx database. However, these data should be interpreted with caution because samples collected by the GTEx database are from healthy individuals. We found that SLC17A8 rs11110359 and TRPV1 rs8065080 might disrupt mRNA splicing and severely affect protein function. SLC17A8 rs11568537 and SHANK2 rs55678639 might affect miRNA binding site activity.
Although none of the SNPs are significant after the Bonferroni correction, the positive SNP results are still meaningful in migraine genetic studies. Some limitations of the present study should be considered. First, the sample size in this study was relatively small, especially in the subgroup analysis. Second, the migraine comorbidities in this study were not diagnosed by a physician and were only assessed by a simple scale. Finally, the biological mechanism of these gene polymorphisms associated with migraine patients remains unclear, although this study found that some SNPs are associated with migraine. Therefore, further studies are necessary to determine better the biological mechanism of the polymorphisms of TRP channels genes and glutamatergic synapse genes associated with migraine.
In summary, this study suggested that the polymorphisms of TRP channels and glutamatergic synapse genes may increase the risk of migraine and the risk of the comorbidities of anxiety and depression in China. Higher GRS scores may increase migraine risk and decrease comorbidity anxiety risk. Further genetic studies with large samples and mechanistic studies are needed.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
The studies involving human participants were reviewed and approved by the Ethics Committee of Harbin Medical University (No. HMUIRB2022004). Written informed consent to participate in this study was provided by the participants' legal guardian/next of kin.
YZ, MW, and YG were responsible for the organizing and management of the project and the final edition of the paper. MW analyzed the data and wrote the entire manuscript. All authors had an essential role in collecting peripheral blood samples and questionnaires and performing the experiments. All authors contributed to the article and approved the submitted version.
This work was supported by the National Natural Science Foundation of China (No. 81773526).
The authors highly appreciate all the people for their participation in this study. The facilities provided by the hospital of the First Affiliated Hospital of Harbin Medical University and three hospitals of Heilongjiang Province Agricultural Reclamation (Hong Xinglong Administration Hospital, Bao Quanling Central Hospital, and Bao Quanling Farm Hospital) are gratefully acknowledged. We thank all the partners and staffs who help us in the process of this study.
All the authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2023.1158028/full#supplementary-material
Abarkan, M., Gaitan, J., Lebreton, F., Perrier, R., Jaffredo, M., Mulle, C., et al. (2019). The glutamate receptor GluK2 contributes to the regulation of glucose homeostasis and its deterioration during aging. Mol. Metab. 30, 152–160. doi:10.1016/j.molmet.2019.09.011
Barbon, A., Vallini, I., and Barlati, S. (2001). Genomic organization of the human GRIK2 gene and evidence for multiple splicing variants. Gene 274 (1-2), 187–197. doi:10.1016/s0378-1119(01)00611-4
Bergman-Bock, S. (2018). Associations between migraine and the most common psychiatric Co-morbidities. Headache 58 (2), 346–353. doi:10.1111/head.13146
Buysse, D. J., Reynolds, C. F., Berman, S. R., and Kupfer, D. J. (1989). The Pittsburgh sleep quality Index: A new instrument for psychiatric practice and research. Psychiatry Res. 28 (2), 193–213. doi:10.1016/0165-1781(89)90047-4
Carreño, O., Corominas, R., Fernández-Morales, J., Camiña, M., Sobrido, M.-J., Fernández-Fernández, J. M., et al. (2012). SNP variants within the vanilloid TRPV1 and TRPV3 receptor genes are associated with migraine in the Spanish population. Am. J. Med. Genet. Part B, Neuropsychiatric Genet. Official Publ. Int. Soc. Psychiatric Genet. 159B (1), 94–103. doi:10.1002/ajmg.b.32007
Chandra, N., Awasthi, R., Ozdogan, T., Johenning, F. W., Imbrosci, B., Morris, G., et al. (2019). A cellular mechanism underlying enhanced capability for complex olfactory discrimination learning. ENeuro 6 (1), ENEURO.0198–18.2019. doi:10.1523/ENEURO.0198-18.2019
Charles, A. C., and Baca, S. M. (2013). Cortical spreading depression and migraine. Nat. Rev. Neurol. 9 (11), 637–644. doi:10.1038/nrneurol.2013.192
Chasman, D. I., Anttila, V., Buring, J. E., Ridker, P. M., Schürks, M., Kurth, T., et al. (2014). Selectivity in genetic association with sub-classified migraine in women. PLoS Genet. 10 (5), e1004366. doi:10.1371/journal.pgen.1004366
Chen, S. P., Fuh, J. L., Chung, M. Y., Lin, Y. C., Liao, Y. C., Wang, Y. F., et al. (2018). Genome-wide association study identifies novel susceptibility loci for migraine in Han Chinese resided in Taiwan. Cephalalgia 38 (3), 466–475. doi:10.1177/0333102417695105
Consortium, G. T. (2015). Human genomics. The genotype-tissue expression (GTEx) pilot analysis: Multitissue gene regulation in humans. Science 348 (6235), 648–660. doi:10.1126/science.1262110
Cortés-Montero, E., Rodríguez-Muñoz, M., Ruiz-Cantero, M. D. C., Cobos, E. J., Sánchez-Blázquez, P., and Garzón-Niño, J. (2020). Calmodulin supports TRPA1 channel association with opioid receptors and glutamate NMDA receptors in the nervous tissue. Int. J. Mol. Sci. 22 (1), 229. doi:10.3390/ijms22010229
de Moura, J. C., Noroes, M. M., Rachetti, V. P. S., Soares, B. L., Preti, D., Nassini, R., et al. (2014). The blockade of transient receptor potential ankirin 1 (TRPA1) signalling mediates antidepressant- and anxiolytic-like actions in mice. Br. J. Pharmacol. 171 (18), 4289–4299. doi:10.1111/bph.12786
Draxler, P., Honsek, S. D., Forsthuber, L., Hadschieff, V., and Sandkühler, J. (2014). VGluT3⁺ primary afferents play distinct roles in mechanical and cold hypersensitivity depending on pain etiology. J. Neurosci. Official J. Soc. For Neurosci. 34 (36), 12015–12028. doi:10.1523/JNEUROSCI.2157-14.2014
Dussor, G., and Cao, Y.-Q. (2016). TRPM8 and migraine. Headache 56 (9), 1406–1417. doi:10.1111/head.12948
Eikermann-Haerter, K. (2021). Neuronal plumes initiate spreading depolarization, the electrophysiologic event driving migraine and stroke. Neuron 109 (4), 563–565. doi:10.1016/j.neuron.2021.01.024
Escelsior, A., Sterlini, B., Belvederi Murri, M., Valente, P., Amerio, A., di Brozolo, M. R., et al. (2020). Transient receptor potential vanilloid 1 antagonism in neuroinflammation, neuroprotection and epigenetic regulation: Potential therapeutic implications for severe psychiatric disorders treatment. Psychiatr. Genet. 30 (2), 39–48. doi:10.1097/YPG.0000000000000249
Fang, J., An, X., Chen, S., Yu, Z., Ma, Q., and Qu, H. (2015). Case-control study of GRIA1 and GRIA3 gene variants in migraine. J. Headache Pain 17, 2. doi:10.1186/s10194-016-0592-2
Favier, M., Pietrancosta, N., El Mestikawy, S., and Gangarossa, G. (2021). Leveraging VGLUT3 functions to untangle brain dysfunctions. Trends Pharmacol. Sci. 42 (6), 475–490. doi:10.1016/j.tips.2021.03.003
Fawley, J. A., Hofmann, M. E., and Andresen, M. C. (2014). Cannabinoid 1 and transient receptor potential vanilloid 1 receptors discretely modulate evoked glutamate separately from spontaneous glutamate transmission. J. Neurosci. Official J. Soc. For Neurosci. 34 (24), 8324–8332. doi:10.1523/JNEUROSCI.0315-14.2014
Formicola, D., Aloia, A., Sampaolo, S., Farina, O., Diodato, D., Griffiths, L. R., et al. (2010). Common variants in the regulative regions of GRIA1 and GRIA3 receptor genes are associated with migraine susceptibility. BMC Med. Genet. 11, 103. doi:10.1186/1471-2350-11-103
Frenguelli, B. G. (2022). The glutamatergic synapse - a key hub in neuronal metabolism, signalling and plasticity. Neuropharmacology 207, 108945. doi:10.1016/j.neuropharm.2022.108945
Fu, X., Yang, J., Wu, X., Lin, Q., Zeng, Y., Xia, Q., et al. (2019). Association between PRDM16, MEF2D, TRPM8, LRP1 gene polymorphisms and migraine susceptibility in the She ethnic population in China. Clin. Investigative Med. Med. Clinique Exp. 42 (1), E21–E30. doi:10.25011/cim.v42i1.32389
Gasparini, C. F., Smith, R. A., and Griffiths, L. R. (2016). Genetic insights into migraine and glutamate: A protagonist driving the headache. J. Neurological Sci. 367, 258–268. doi:10.1016/j.jns.2016.06.016
Gavva, N. R., Sandrock, R., Arnold, G. E., Davis, M., Lamas, E., Lindvay, C., et al. (2019). Reduced TRPM8 expression underpins reduced migraine risk and attenuated cold pain sensation in humans. Sci. Rep. 9 (1), 19655. doi:10.1038/s41598-019-56295-0
Hashimoto, K. (2019). Rapid-acting antidepressant ketamine, its metabolites and other candidates: A historical overview and future perspective. Psychiatry Clin. Neurosci. 73 (10), 613–627. doi:10.1111/pcn.12902
Headache Classification Committee of the International Headache, S. (2013). The international classification of headache disorders, 3rd edition (beta version). Cephalalgia 33 (9), 629–808. doi:10.1177/0333102413485658
Henley, J. M., Nair, J. D., Seager, R., Yucel, B. P., Woodhall, G., Henley, B. S., et al. (2021). Kainate and AMPA receptors in epilepsy: Cell biology, signalling pathways and possible crosstalk. Neuropharmacology 195, 108569. doi:10.1016/j.neuropharm.2021.108569
Iannone, L. F., De Logu, F., Geppetti, P., and De Cesaris, F. (2022). The role of TRP ion channels in migraine and headache. Neurosci. Lett. 768, 136380. doi:10.1016/j.neulet.2021.136380
Ko, H.-G., Oh, S.-B., Zhuo, M., and Kaang, B.-K. (2016). Reduced acute nociception and chronic pain in Shank2-/- mice. Mol. Pain 12, 1744806916647056. doi:10.1177/1744806916647056
Kormos, V., Kecskés, A., Farkas, J., Gaszner, T., Csernus, V., Alomari, A., et al. (2022). Peptidergic neurons of the Edinger-Westphal nucleus express TRPA1 ion channel that is downregulated both upon chronic variable mild stress in male mice and in humans who died by suicide. J. Psychiatry & Neurosci. JPN 47 (3), E162–E175. doi:10.1503/jpn.210187
Kroenke, K., Spitzer, R. L., and Williams, J. B. (2001). The PHQ-9: Validity of a brief depression severity measure. J. Gen. Intern Med. 16 (9), 606–613. doi:10.1046/j.1525-1497.2001.016009606.x
Lisboa, S. F., and Guimarães, F. S. (2012). Differential role of CB1 and TRPV1 receptors on anandamide modulation of defensive responses induced by nitric oxide in the dorsolateral periaqueductal gray. Neuropharmacology 62 (8), 2455–2462. doi:10.1016/j.neuropharm.2012.02.008
Liu, D., Bi, Y., Liu, Z., Liu, H., and Li, Z. (2014). The expression of vesicular glutamate transporter 3 and vesicular monoamine transporter 2 induced by brain-derived neurotrophic factor in dorsal root ganglion neurons in vitro. Brain Res. Bull. 100, 93–106. doi:10.1016/j.brainresbull.2013.11.007
Liu, J., Han, Y.-S., Liu, L., Tang, L., Yang, H., Meng, P., et al. (2021). Abnormal Glu/mGluR2/3/PI3K pathway in the hippocampal neurovascular unit leads to diabetes-related depression. Neural Regen. Res. 16 (4), 727–733. doi:10.4103/1673-5374.296418
Liu, X., Yuan, M., Lau, B. W.-M., and Li, Y. (2022). SHANK family on stem cell fate and development. Cell Death Dis. 13 (10), 880. doi:10.1038/s41419-022-05325-3
Louter, M. A., Pijpers, J. A., Wardenaar, K. J., van Zwet, E. W., van Hemert, A. M., Zitman, F. G., et al. (2015). Symptom dimensions of affective disorders in migraine patients. J. Psychosom. Res. 79 (5), 458–463. doi:10.1016/j.jpsychores.2015.09.014
Lv, X., Xu, B., Tang, X., Liu, S., Qian, J.-H., Guo, J., et al. (2023). The relationship between major depression and migraine: A bidirectional two-sample mendelian randomization study. Front. Neurology 14, 1143060. doi:10.3389/fneur.2023.1143060
Ma, H., Xun, G., Zhang, R., Yang, X., and Cao, Y. (2019). Correlation between GRIK2 rs6922753, rs2227283 polymorphism and aggressive behaviors with Bipolar Mania in the Chinese Han population. Brain Behav. 9 (11), e01449. doi:10.1002/brb3.1449
Maggioni, E., Delvecchio, G., Grottaroli, M., Garzitto, M., Piccin, S., Bonivento, C., et al. (2019). Common and different neural markers in major depression and anxiety disorders: A pilot structural magnetic resonance imaging study. Psychiatry Res. Neuroimaging 290, 42–50. doi:10.1016/j.pscychresns.2019.06.006
Maher, B. H., Lea, R. A., Follett, J., Cox, H. C., Fernandez, F., Esposito, T., et al. (2013). Association of a GRIA3 gene polymorphism with migraine in an Australian case-control cohort. Headache 53 (8), 1245–1249. doi:10.1111/head.12151
Miletic, G., Miyabe, T., Gebhardt, K. J., and Miletic, V. (2005). Increased levels of Homer1b/c and Shank1a in the post-synaptic density of spinal dorsal horn neurons are associated with neuropathic pain in rats. Neurosci. Lett. 386 (3), 189–193. doi:10.1016/j.neulet.2005.06.007
Murrough, J. W., Abdallah, C. G., and Mathew, S. J. (2017). Targeting glutamate signalling in depression: Progress and prospects. Nat. Rev. Drug Discov. 16 (7), 472–486. doi:10.1038/nrd.2017.16
Myung, W., Song, J., Lim, S.-W., Won, H.-H., Kim, S., Lee, Y., et al. (2012). Genetic association study of individual symptoms in depression. Psychiatry Res. 198 (3), 400–406. doi:10.1016/j.psychres.2011.12.037
Ngoc, K. H., Kecskés, A., Kepe, E., Nabi, L., Keeble, J., Borbély, É., et al. (2023). Expression of the Transient Receptor Potential Vanilloid 1 ion channel in the supramammillary nucleus and the antidepressant effects of its antagonist AMG9810 in mice. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 73, 96–107. doi:10.1016/j.euroneuro.2023.04.017
Niswender, C. M., and Conn, P. J. (2010). Metabotropic glutamate receptors: Physiology, pharmacology, and disease. Annu. Rev. Pharmacol. Toxicol. 50, 295–322. doi:10.1146/annurev.pharmtox.011008.145533
Rees, T. A., Hendrikse, E. R., Hay, D. L., and Walker, C. S. (2022). Beyond CGRP: The calcitonin peptide family as targets for migraine and pain. Br. J. Pharmacol. 179 (3), 381–399. doi:10.1111/bph.15605
Russell, F. A., King, R., Smillie, S. J., Kodji, X., and Brain, S. D. (2014). Calcitonin gene-related peptide: Physiology and pathophysiology. Physiol. Rev. 94 (4), 1099–1142. doi:10.1152/physrev.00034.2013
Sartim, A. G., Moreira, F. A., and Joca, S. R. L. (2017). Involvement of CB1 and TRPV1 receptors located in the ventral medial prefrontal cortex in the modulation of stress coping behavior. Neuroscience 340, 126–134. doi:10.1016/j.neuroscience.2016.10.031
Schmeisser, M. J. (2015). Translational neurobiology in Shank mutant mice--model systems for neuropsychiatric disorders. Ann. Anat. = Anatomischer Anzeiger Official Organ Anatomische Gesellschaft 200, 115–117. doi:10.1016/j.aanat.2015.03.006
Seal, R. P., Wang, X., Guan, Y., Raja, S. N., Woodbury, C. J., Basbaum, A. I., et al. (2009). Injury-induced mechanical hypersensitivity requires C-low threshold mechanoreceptors. Nature 462 (7273), 651–655. doi:10.1038/nature08505
Shibata, M., and Tang, C. (2021). Implications of transient receptor potential cation channels in migraine pathophysiology. Neurosci. Bull. 37 (1), 103–116. doi:10.1007/s12264-020-00569-5
Sink, K. S., Walker, D. L., Yang, Y., and Davis, M. (2011). Calcitonin gene-related peptide in the bed nucleus of the stria terminalis produces an anxiety-like pattern of behavior and increases neural activation in anxiety-related structures. J. Neurosci. 31 (5), 1802–1810. doi:10.1523/JNEUROSCI.5274-10.2011
Siokas, V., Liampas, I., Aloizou, A.-M., Papasavva, M., Bakirtzis, C., Lavdas, E., et al. (2022). Deciphering the role of the rs2651899, rs10166942, and rs11172113 polymorphisms in migraine: A meta-analysis. Med. Kaunas. Lith. 58 (4), 491. doi:10.3390/medicina58040491
Spekker, E., Körtési, T., and Vécsei, L. (2022). TRP channels: Recent development in translational research and potential therapeutic targets in migraine. Int. J. Mol. Sci. 24 (1), 700. doi:10.3390/ijms24010700
Steiner, T. J., Stovner, L. J., Jensen, R., Uluduz, D., and Katsarava, Z.Lifting The Burden: the Global Campaign against Headache (2020). Migraine remains second among the world's causes of disability, and first among young women: Findings from GBD2019. J. Headache Pain 21 (1), 137. doi:10.1186/s10194-020-01208-0
Tukey, D. S., Lee, M., Xu, D., Eberle, S. E., Goffer, Y., Manders, T. R., et al. (2013). Differential effects of natural rewards and pain on vesicular glutamate transporter expression in the nucleus accumbens. Mol. Brain 6, 32. doi:10.1186/1756-6606-6-32
Wan, L., Liu, D., Xiao, W.-B., Zhang, B.-X., Yan, X.-X., Luo, Z.-H., et al. (2022). Association of SHANK family with neuropsychiatric disorders: An update on genetic and animal model discoveries. Cell. Mol. Neurobiol. 42 (6), 1623–1643. doi:10.1007/s10571-021-01054-x
Wattiez, A.-S., Sowers, L. P., and Russo, A. F. (2020). Calcitonin gene-related peptide (CGRP): Role in migraine pathophysiology and therapeutic targeting. Expert Opin. Ther. Targets 24 (2), 91–100. doi:10.1080/14728222.2020.1724285
Wigginton, J. E., Cutler, D. J., and Abecasis, G. R. (2005). A note on exact tests of Hardy-Weinberg equilibrium. Am. J. Hum. Genet. 76 (5), 887–893. doi:10.1086/429864
Xu, Z., and Taylor, J. A. (2009). SNPinfo: Integrating GWAS and candidate gene information into functional SNP selection for genetic association studies. Nucleic Acids Res. 37, W600–W605. doi:10.1093/nar/gkp290
Yakubova, A., Davidyuk, Y., Tohka, J., Khayrutdinova, O., Kudryavtsev, I., Nurkhametova, D., et al. (2021). Searching for predictors of migraine chronification: A pilot study of 1911A>G polymorphism of TRPV1 gene in episodic versus chronic migraine. J. Mol. Neurosci. MN 71 (3), 618–624. doi:10.1007/s12031-020-01683-9
Yang, Y., Wei, K., Zhang, H., Hu, H., Yan, L., Gui, W., et al. (2022). Identifying functional brain abnormalities in migraine and depression comorbidity. Quantitative Imaging Med. Surg. 12 (4), 2288–2302. doi:10.21037/qims-21-667
You, I.-J., Jung, Y.-H., Kim, M.-J., Kwon, S.-H., Hong, S.-I., Lee, S.-Y., et al. (2012). Alterations in the emotional and memory behavioral phenotypes of transient receptor potential vanilloid type 1-deficient mice are mediated by changes in expression of 5-HT-A, GABA(A), and NMDA receptors. Neuropharmacology 62 (2), 1034–1043. doi:10.1016/j.neuropharm.2011.10.013
Yuan, H., Low, C.-M., Moody, O. A., Jenkins, A., and Traynelis, S. F. (2015). Ionotropic GABA and glutamate receptor mutations and human neurologic diseases. Mol. Pharmacol. 88 (1), 203–217. doi:10.1124/mol.115.097998
Zhang, Q., Shao, A., Jiang, Z., Tsai, H., and Liu, W. (2019). The exploration of mechanisms of comorbidity between migraine and depression. J. Cell. Mol. Med. 23 (7), 4505–4513. doi:10.1111/jcmm.14390
Keywords: migraine, gene polymorphisms, TRP channels, glutamate pathway, glutamatergic synapse
Citation: Wang M, Gu Y, Meng S, Kang L, Yang J, Sun D, Liu Y, Wan Z, Shan Y, Xue D, Su C, Li S, Yan R, Liu Y, Zhao Y and Pan Y (2023) Association between TRP channels and glutamatergic synapse gene polymorphisms and migraine and the comorbidities anxiety and depression in a Chinese population. Front. Genet. 14:1158028. doi: 10.3389/fgene.2023.1158028
Received: 03 February 2023; Accepted: 17 May 2023;
Published: 26 May 2023.
Edited by:
Lucia Carboni, University of Bologna, ItalyReviewed by:
Carolina Lemos, University of Porto, PortugalCopyright © 2023 Wang, Gu, Meng, Kang, Yang, Sun, Liu, Wan, Shan, Xue, Su, Li, Yan, Liu, Zhao and Pan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yashuang Zhao, emhhb195YXNodWFuZ0AyNjMubmV0; Yonghui Pan, YWlndWkxOTkzQDEyNi5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.