
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet., 28 October 2022
Sec. Evolutionary and Population Genetics
Volume 13 - 2022 | https://doi.org/10.3389/fgene.2022.974472
 Arunas L. Radzvilavicius1,2
Arunas L. Radzvilavicius1,2 Iain G. Johnston1,2*
Iain G. Johnston1,2*Bioenergetic organelles—mitochondria and plastids—retain their own genomes (mtDNA and ptDNA), and these organelle DNA (oDNA) molecules are vital for eukaryotic life. Like all genomes, oDNA must be able to evolve to suit new environmental challenges. However, mixed oDNA populations in cells can challenge cellular bioenergetics, providing a penalty to the appearance and adaptation of new mutations. Here we show that organelle “bottlenecks,” mechanisms increasing cell-to-cell oDNA variability during development, can overcome this mixture penalty and facilitate the adaptation of beneficial mutations. We show that oDNA heteroplasmy and bottlenecks naturally emerge in evolutionary simulations subjected to fluctuating environments, demonstrating that this evolvability is itself evolvable. Usually thought of as a mechanism to clear damaging mutations, organelle bottlenecks therefore also resolve the tension between intracellular selection for pure cellular oDNA populations and the “bet-hedging” need for evolvability and adaptation to new environments. This general theory suggests a reason for the maintenance of organelle heteroplasmy in cells, and may explain some of the observed diversity in organelle maintenance and inheritance across taxa.
Mitochondria and plastids are organelles that exist in populations in eukaryotic cells and perform vital bioenergetic, metabolic, and signalling tasks. The majority of cells in the majority of eukaryotes contain mitochondria; cells in photoautotrophs also contain plastids. Originally independent organisms, mitochondria and plastids retain their own genomes (mtDNA and ptDNA, respectively; ptDNA is also called cpDNA for chloroplast DNA) that encode vital bioenergetic apparatus. Mutations in these organelle DNA (oDNA) molecules can compromise cellular function with fatal consequences (Wallace and Chalkia, 2013).
oDNA molecules exist in dynamic populations in eukaryotic cells (Stewart and Chinnery, 2015; Johnston, 2018; Johnston and Burgstaller, 2019). Each cell may contain hundreds or thousands of oDNA molecules, and these molecules may not be genetically identical. A mixture of genetically different oDNA molecules in a cell is referred to as heteroplasmic. Recent evidence suggests that small-scale heteroplasmy, where each genome differs at a small number of loci from others, is common in cellular mtDNA populations (Morris et al., 2017; Aryaman et al., 2019). Larger-scale mtDNA diversity (differences at many loci) exist in populations of organisms, giving rise to, for example, the geographical distribution of different mitochondrial haplogroups in the human population (Mishmar et al., 2003; Ruiz-Pesini et al., 2004; Ruiz-Pesini et al., 2006; Wallace and Chalkia, 2013).
As with nuclear genomes, organelle genomes are subject to evolution in sequence and structure; for example, experimental work has highlighted how environmental pressures shape mtDNA through selection (Blier et al., 2001; Mishmar et al., 2003; Ruiz-Pesini et al., 2004; Dowling et al., 2007; Pereira et al., 2014), although the role of (particularly positive) environmental selection in human populations remains debated (Kivisild et al., 2006; Sun et al., 2007). The ability to evolve in response to environmental change is often termed evolvability (Kirschner and Gerhart, 1998; Wagner, 2013) and is an essential aspect of life. A classical question is how biological systems resolve the perceived tradeoff between two desirable characteristics: Evolvability (producing new phenotypes through mutations) and robustness (retaining the same phenotype in the face of mutations) (Masel and Meredith, 2010; Wagner, 2013). This tradeoff can be resolved, at the population level, if a “robust” subset of a population occupies regions of genetic space that are robust and an “evolvable” subset occupies other regions from which other phenotypes are mutationally accessible (Wagner, 2008). Such genetic spread can act to “hedge bets” against future environmental changes, by increasing the number of potential new solutions to uncertain future challenges (Fraser and Kaern, 2009; Tufto, 2015).
To evolve and adapt, mutational changes must be acquired by cellular oDNA populations, and these changes must be inherited from parent organism to offspring organism (Birky, 2001). There is a negligibly small probability that the same mutational change is acquired simultaneously by all oDNA molecules in a cellular population, so the acquisition of mutational change necessarily induces cellular heteroplasmy (Wallace, 2010). However, recent experimental work has shown that some instances of even limited heteroplasmy can compromise cellular and organismal function (Acton et al., 2007; Lane, 2012; Sharpley et al., 2012; Picard et al., 2014; Latorre-Pellicer et al., 2019). Genetic differences in mtDNA molecules within a cell may lead to incompatibilities between their encoded protein products, challenging bioenergetic capacity (Latorre-Pellicer et al., 2019). This challenge is multifaceted, including, for example, compromised respiratory chain activity and downstream metabolism, elevated reactive oxygen species, and impacts on autophagy and cell ultrastructure (Latorre-Pellicer et al., 2019; Lechuga-Vieco et al., 2020). This raises the question: how can potentially beneficial mutation changes be acquired by oDNA, if the corresponding heteroplasmy has a negative impact on fitness?
The inheritance of mtDNA in many organisms involves a genetic “bottleneck,” where the effective number of mtDNA molecules is reduced as the female germline develops (Xu, 2005; Wallace and Chalkia, 2013; Garone et al., 2018; Johnston, 2019). This bottleneck can arise in part, but not necessarily completely, from a physical reduction of mtDNA copy number, and from stochastic cell divisions, mtDNA turnover, and other processes (Freyer et al., 2012; Rebolledo-Jaramillo et al., 2014; Johnston et al., 2015; Li et al., 2016; Peter et al., 2018; Johnston, 2019). For heteroplasmic mothers, this bottleneck increases cell-to-cell variability in heteroplasmy level in the germ cells of the next generation; selective differences between mtDNA types will also be amplified by the reduction in effective population size, acting in parallel to change mean heteroplasmy levels (Burgstaller et al., 2018; Lieber et al., 2019; Wei et al., 2019). For example, a mother carrying 50% type A mtDNA and 50% type B mtDNA may produce egg cells with a range of 30%–70% type A. This increased variability can then be exploited by purifying selection at the cellular level to remove cells with a high proportion of deleterious oDNA, helping to limit Muller’s ratchet—the buildup of deleterious mutations over generations (Muller, 1964; Bergstrom and Pritchard, 1998; Lynch and Blanchard, 1998; Krakauer et al., 1999; Lynch et al., 2006). The bottleneck can allow rapid shifts of mtDNA makeup even between one generation and the next (Ashley et al., 1989; Johnston, 2019).
In addition to helping clear deleterious mutations, the variability generated by the bottleneck has also been suggested to help adaptation (Wallace, 2010). Previous theoretical analyses have explored the role of cell-to-cell oDNA variability in fixing new mutants within model cells and populations (James, 1992; Roze et al., 2005). However, to our knowledge, the fitness penalty associated with heteroplasmic cellular oDNA populations—and the consequent potential “barrier” or fitness valley—has not yet been considered in this literature. Conversely, studies are beginning to explore the role of heteroplasmic costs on evolutionary and cellular behaviour (Christie et al., 2015; Hoitzing et al., 2019) but to our knowledge have yet to be linked to the bottleneck’s role in adaptation and purification. Whether heteroplasmic fitness penalties pose a theoretical barrier to oDNA adaptation thus remains to be determined.
Here, we asked whether the rapid shifting in oDNA type due to a genetic “bottleneck” can help to overcome the fitness penalty associated with heteroplasmic cellular oDNA populations, and thus enhance evolvability. Our corollary question was, if bottlenecks can help evolvability in this way, is robustness necessarily sacrificed? We proceed by introducing a simple model for cellular oDNA populations and their behaviour between generations. We then analyse this model with stochastic simulations to investigate the extent to which heteroplasmy variability facilitates the evolvability of oDNA, and whether bottlenecks and heteroplasmy can naturally emerge as evolutionarily positive “strategies” where evolvability is essential.
We consider a two-scale model, describing the inheritance of oDNA at the individual level, and evolution of a population of individuals.
We model cells—which may be individuals, or female germline cells in a multicellular organism—as containing a population of n oDNA molecules which can generally include a mixture of oDNA types. Each generation consists of N mother cells. A mother cell in one generation gives rise to Noff daughter cells in the next generation. Our model does not assume that a generation involves a particular number of cell divisions [for example, around 25 in some mammalian germlines (Drost and Lee, 1995), or one in the case of unicellular organisms]: the contribution of these divisions and other segregating processes are pooled in our segregation parameter. We rather consider cells at identical developmental points, one generation to the next. Our default parameterisation uses N = 20 and Noff = 2 with n = 100; we verified that changes to this parameterisation had only limited influence on the structure of our results (Supplementary Figures S1, S2). Each cell has a mutant proportion h, ranging between 0 and 1. Each daughter cell inherits an oDNA complement from its mother. Modelling the accumulation of mutations and the action of segregation during development, this complement is transformed by two operators describing the action of these processes:
The functions gμ and gσ respectively describe changes in heteroplasmy due to mutation and segregation. For mutation, we use
So that the mutant proportion in a cell is changed over a generation by a normal mutation kernel [N(a, b) denotes the normal distribution with mean a and standard deviation b], and constrained to lie between 0 and 1. The mean and variance of this kernel are derived from considering how mutational processes change the copy number of the different oDNA types during development (see Supplementary Material). Our mutation rate μ is not directly equated with the population-level mutation rate that is often considered when tracking oDNA mutations in populations of organisms through evolutionary history. Rather, it describes the rate of mutation of individual oDNA molecules in the cellular population, hence setting a characteristic scale of heteroplasmy change within a cell during a generation. For segregation, we use
Following the well-known result from statistical genetics (Wright, 1942; Johnston, 2019), with final mutant proportion again constrained to lie between 0 and 1. The σ parameter in our model, controlling the strength of segregation (the inverse of an effective “bottleneck” size) is then roughly comparable to the ‘normalised heteroplasmy variance’ V′(h) = V(h)/(h(1 − h)) commonly observed in genetic studies. For heteroplasmy distributions that approach normality, σ → V′(h); departures from normality make this equivalence weaker. These normal kernels are used for computational speed in the rather demanding simulations of the model; they have been shown to approximate the true, discrete behaviour of cellular oDNA population well across a range of values in previous work (Johnston et al., 2015).
In the population level of our model, each cell has a fitness value with contributions from 1) the performance of its constituent oDNA molecules in the current environment, and 2) an admixture penalty if the cell contains a mixture of different oDNA types. Each oDNA type provides a fitness contribution that is a function of environment, with different oDNA types performing better in different environments. The fitness function is sigmoidal, taking values between 0 and 1:
The performance part fE(h) = −h in environment A and h in environment B, so that the oDNA wildtype is favoured in environment A and the mutant is favoured in environment B. The admixture part is by default set to fϵ(h) = 0 for homoplasmic h = 0 and h = 1, and − ϵ otherwise, following the observation that even limited heteroplasmy can challenge cell performance due to (Latorre-Pellicer et al., 2019). We also investigated a range of alternative fitness functions, including where the admixture contribution was a smooth function of mutant proportion
Following assignment of fitness values, a new population of size N is selected from the N × Noff daughter cells, with cells selected (with replacement) with probability proportional to their fitness (roulette wheel selection). Fitness and genetic content is tracked through tmax generations (tmax = 500 by default), and different regimes of environmental change, described for each experiment, are applied through generations. Adaptation to a new oDNA type is defined as the first time when the mutant proportion of that type exceeds 75%; changing this criterion simply rescales adaptation time results, leaving the qualitative behaviour intact.
The default picture above considers fitness to be governed by a single locus, so that mutant and wildtype oDNA molecules are the only types involved in the model. For generality we also consider a case where a third, dysfunctional, type of oDNA is included, and mutations can transform both mutant and wildtype oDNA molecules into this dysfunctional state. This models a picture where mutation leads to deleterious effects at other oDNA loci. In this case we explicitly track different oDNA types in the cell, with copy numbers w (wildtype), m (mutant), and d (dysfunctional). We define mutant proportion h = m/(w + m) (hence, the proportion of mutant oDNAs among the cellular population of functional oDNAs), and introduce the dysfunctional proportion hD = d/(w + m + d), reporting the proportion of dysfunctional oDNAs in a cell. Cells are now described by {h, hD}, and the update rule now becomes
We first asked to what extent a fitness penalty associated with heteroplasmy may challenge organelle adaptation to new environments. We initialised a model population of cells with internal homoplasmic organelle populations, all of the variant optimal for environment A. At time t = 0, we impose environment B, and track how organelle populations within the cell population subsequently evolve. We varied mutation rate μ and the magnitude of the fitness penalty ϵ experienced by cells with heteroplasmic populations.
We found that adaptation to the new environment was impossible in some circumstances, specifically for low mutation rates and high fitness penalties (Figures 1A,C). Similar behaviour was observed for the range of alternative fitness functions we considered (Supplementary Figure S4). In these regions, the selective penalty associated with the appearance of new mutations (and hence the appearance of heteroplasmy) prevents such mutations from ever propagating sufficiently to provide a fitness benefit from environmental matching. The penalty constitutes a “fitness valley” that the system must overcome in order to fix new mutations. At higher mutation rates, the heteroplasmy level can increase rapidly enough—crossing the “valley”—to allow this advantage. However, this high-mutation case may well not be a viable evolutionary strategy: although higher mutation rates may help to cross the heteroplasmic fitness valley for a single locus, a higher mutation rate will likely lead to many other potentially deleterious changes through other sites in the oDNA. Such a mutational meltdown across the genome should clearly be avoided.
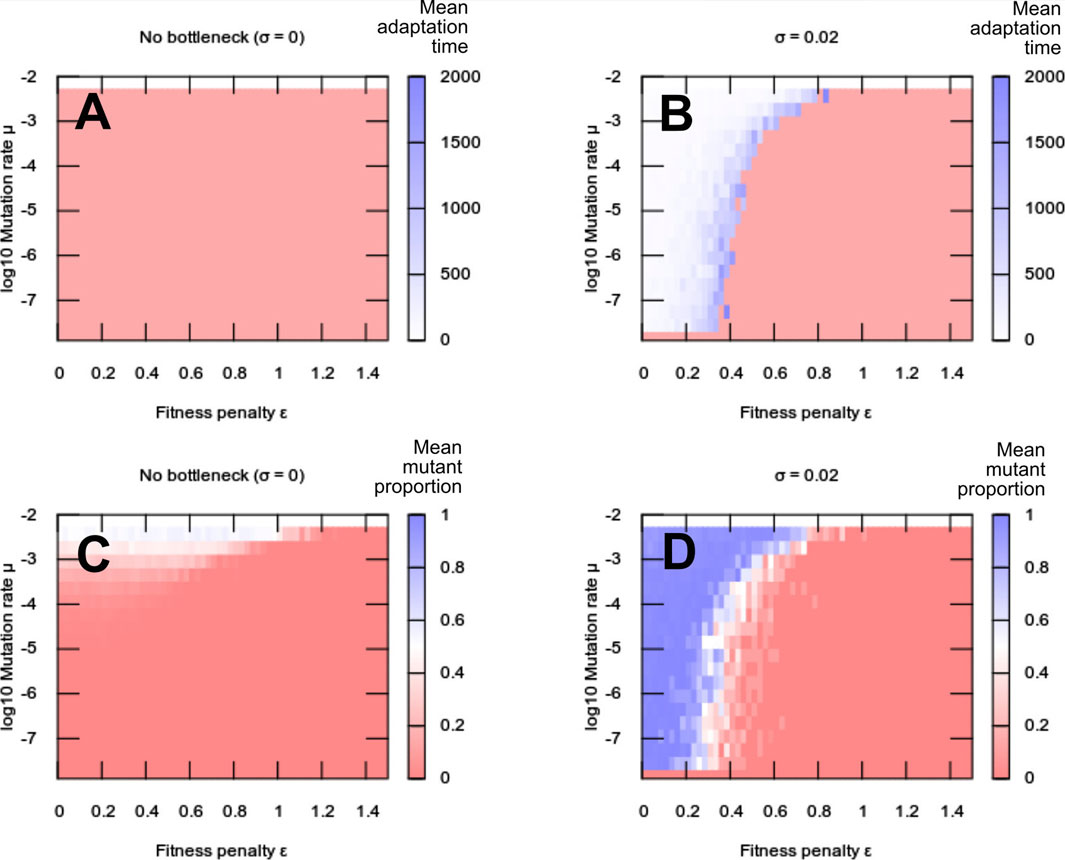
FIGURE 1. Adaptation to a new environment. (A,B) Adaptation times (number of generations until a new mutant dominates) and (C,D) mean mutant frequency in a population after a change of environment, with heteroplasmy fitness penalty ϵ and mutation rate μ. (A,C) No between-generation segregation of mutations (σ = 0); (B,D) nonzero segregation (σ = 0.02). Without segregation, the “fitness valley” prevents adaptation to the new environment, with the population either remaining adapted to the previous state (low mutation rate, high penalty) or reaching an intermediate state driven by mutation. Except in the case of zero mutation rate, segregation allows traversal of the fitness valley and facilitates adaptation. Each pixel value is the average over 10 simulation instances; the bottom row of pixels in each plot gives the μ = 0 (no adaptation) cases.
We next asked whether between-generation segregation of organelle mutations, via a genetic bottleneck, could help traverse this fitness valley and increase evolvability without requiring such an elevated mutation rate. We introduced this segregation via a new process in the model, where the mutant proportion of daughter cells differs from that of the mother cell by a normal random variate. This is not a perfect model of, for example, the mtDNA bottleneck, but captures the essential increase in variance in an intuitively simple way.
We asked how the magnitude of this segregation effect (the “tightness” of the genetic bottleneck) influenced the system’s ability to adapt to new environments. The simulation setup was the same as in the previous section, with new variable σ describing segregation strength (Figures 1A,C thus correspond to σ = 0). We found that a nonzero segregation strength substantially enhances evolvability, allowing the propagation of beneficial mutations in regimes (particular those with low mutation rate and high fitness penalty) that posed the greatest challenge to adaptation without segregation (Figures 1B,D). This behaviour was observed consistently across demographic and cellular parameterisations of our models, and when “off-target” mutations inducing oDNA dysfunctionality was accounted for (Supplementary Figures S1–S3).
Figures 1B,D show that an organelle bottleneck increases organelle evolvability. A parallel feature of populations is robustness—the ability to retain functionality in the face of mutational pressure. Traditionally, robustness and evolvability were viewed as opposites, but they can be reconciled at the population level (Wagner, 2008).
In our model system, robustness can be pictured as related to the level of homoplasmy in the population of individuals: the higher the proportion homoplasmic for the environmentally-optimal organelle type, the more mutation can be “absorbed” without compromising the population. We found that the increased evolvability facilitated by organelle bottlenecks did not compromise system robustness: adaptation to new environments was achieved while long-term behaviour retained high levels of homoplasmy (Figure 2). Because bottlenecks allowed adaptation at lower mutation rates, the bottleneck-adapted population was able to retain a more homoplasmic state than was possible when the population was forced to adapt through mutation alone. When dysfunctional mutations were likely, only low mutation rates were possible without losing population viability, but even small bottleneck sizes were sufficient to allow adaptation even at these low mutation rates (Supplementary Figure S3).
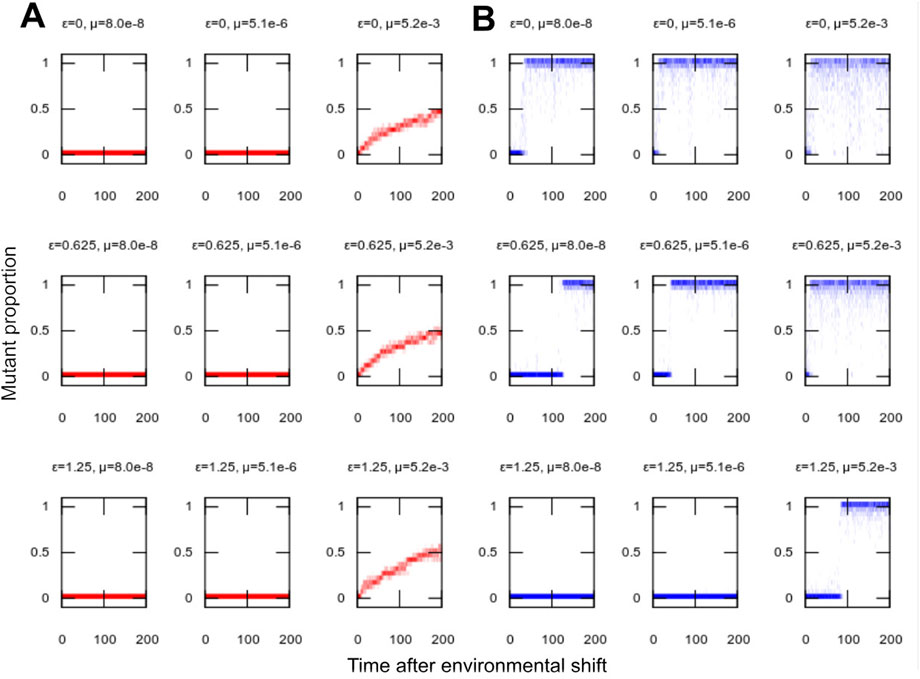
FIGURE 2. Population structure in adaptation to new environments. Population distribution of mutant proportion h with time in simulations adapting to a new environment (as in Figure 1). (A) No segregation σ = 0; (B) nonzero segregation σ = 0.2. Distributions of h are presented for single example simulations with the given parameterisations. Lower mutation rates allow more homoplasmic populations (tighter h distributions), but, in the absence of segregation, challenge adaptation to the new environment (slow or no progress towards h = 1). Higher mutation rates facilitate adaptation but lead to more heterogeneous populations (wider h distributions). Segregation allows both rapid adaptation and high levels of homoplasmy.
Following the observation that nonzero segregation enhances evolvability (without compromising robustness), we next asked how this enhancement depends on the magnitude of segregation strength (the “size” of the genetic bottleneck). To this end we varied σ, the magnitude of segregation strength, and ϵ, the fitness penalty associated with heteroplasmy, for different values of μ, mutation rate (Figure 3). Intuitively, for zero mutation rate, no adaptation was possible, because there was no generation of any genetic diversity at any time (Figure 3A). However, for all nonzero mutation rates, almost any nonzero segregation immediately allowed a substantial enhancement of evolvability, with this enhancement then increasing more slowly as bottleneck size decrease further (Figures 3B–D).
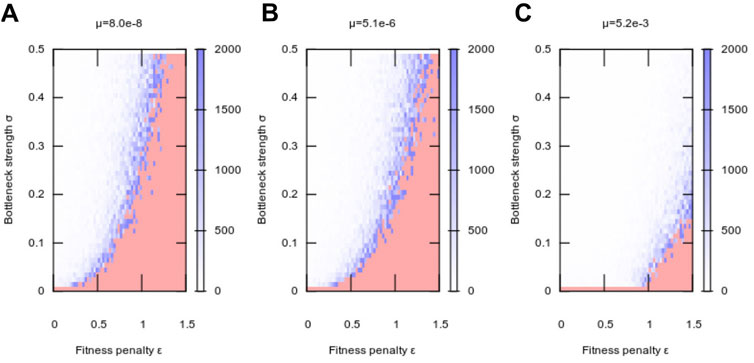
FIGURE 3. Adaptation to a new environment as a function of segregation strength. Plots show, as in Figure 1, the mean number of generations taken for a population to adapt following a change of environment, at (A) low, (B) intermediate, and (C) high mutation rate. Increasing segregation strength σ facilitates adaptation at higher fitness penalties, with higher mutation rates μ amplifying this effect. At μ = 0, no adaptation was observed in our simulations. Each pixel value is the average over 10 simulation instances.
These results suggest that a combination of mutation and a ‘genetic bottleneck’ enhance evolvability in the sense of adaptation to new environments. To see if such a strategy could emerge in an evolving system, we next adapted our simulation so that mutation rate and “bottleneck size” were themselves evolvable parameters within a population of cells. Specifically, we assign each cell in our model population a mutation rate μi and a segregation strength σi. These parameters are inherited from mother cell to daughter cells. This inheritance is imperfect, and a daughter’s mutation and segregation parameters can themselves mutate to be different from the mother. This “parameter mutation rate” (different from the organelle mutation rate μi) is labelled δ.
As the population of cells evolves, those cellular parameters that constitute evolutionarily successful strategies may thus come to dominate the population, while those parameters that constitute unsuccessful strategies will disappear. We simulated this system under different conditions. First, the new but then constant environment above (changing from environment A to environment B at time t = 0). Second, oscillating environments, changing from A to B with a period of τ generations. Lower values of τ correspond to faster environmental change; higher values of τ mean that environmental flips are infrequent. In each of these conditions, we observed the values of mutation rate and segregation strength that came to dominate the population.
We found that nonzero mutation rates and bottleneck strengths emerged in all conditions we consider (Figures 4A–C). For static environments, their evolution was dependent on the magnitude of the fitness penalty. Lower fitness penalties allowed higher mutation rates and weaker bottlenecks; higher fitness penalties led to lower mutation rates and stronger bottlenecks.
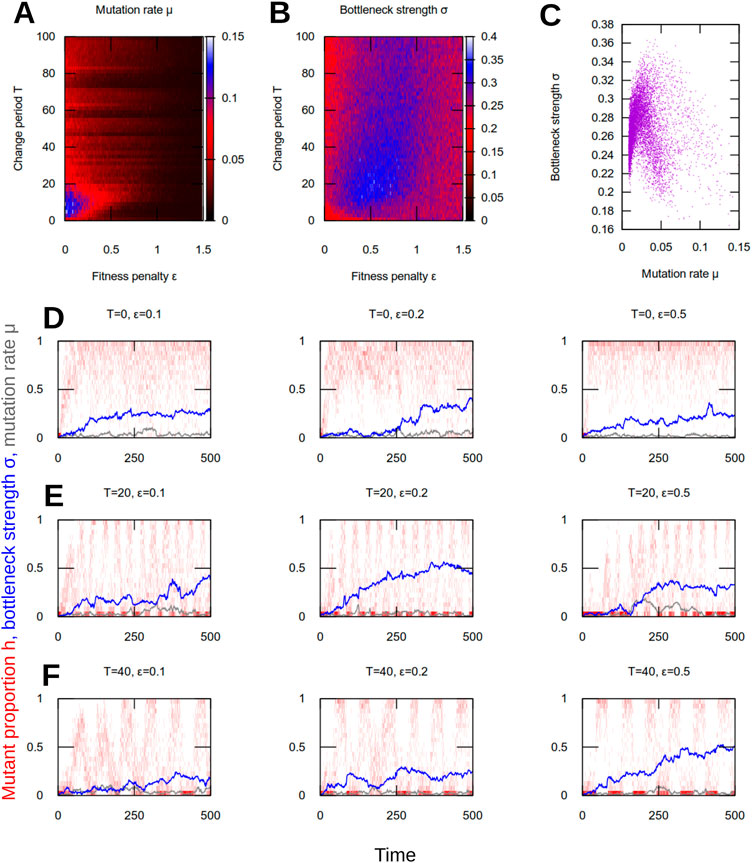
FIGURE 4. Evolution of mutation rate and segregation in dynamic environments. (A) Mean mutation rate μ, (B) mean segregation strength σ, and (C) joint mean μ and σ after tmax = 500 generations in evolutionary simulations (see text). Each pixel value is the average over 100 simulation instances. Each point in (C) is from a particular parameterisation [i.e., a particular pixel in (A) and (B)]. (D–F) Population histograms of mutant proportion h (red), plotted with traces of evolving mean mutation rate μ (grey) and segregation strength σ (blue), for different parameterisations and environmental change regimes: (D) no change; (E) rapid change; (F) slower change. Results are from single, representative simulations, many of which are averaged to give the results in (A–C). When environments change, mutation and segregation evolve together to facilitate adaptation (rapid shifting with environment) and robustness (high proportions of heteroplasmy). With higher fitness penalties ϵ this behaviour takes longer to evolve but still emerges.
For dynamic environments, substantially higher mutation rates, and stronger bottlenecks, evolved. Both mutation rate and bottleneck strength peaked at intermediate fitness penalties (Figures 4A,B). Evolved bottleneck strength showed less dependence on the period of environmental change, although for very rapid environmental change bottleneck strength was marginally weaker at both low and high fitness penalties (for example, different values of ϵ for T = 100 in Figure 4B). At lower evolved mutation rates, there was a weak positive correlation between evolved mutation rate and evolved segregation strength (Figure 4C). This correlation weakened further at high mutation rates, with evolved segregation strength plateauing around a maximum value (Figure 4C).
The effect of the evolved nonzero mutation rates in these experiments was to maintain a low level of heteroplasmy in cells and in the population [as observed experimentally (Guo et al., 2013; Morris et al., 2017; Aryaman et al., 2019); see Discussion], allowing more rapid adaptation when environmental change occurs. This genetic variability can be viewed as a bet-hedging mechanism, sacrificing perfect adaptation in the current state for increased flexibility in an uncertaint future (Fraser and Kaern, 2009; Tufto, 2015).
We also performed evolutionary simulations under fluctuating environments where we fixed mutation rate μ to a constant value and allowed segregation strength σ to evolve (Supplementary Figure S5). We found a similar picture, with evolved segregation strength peaking at intermediate fitness penalties, and observed that stronger segregation evolved to compensate for lower mutation rates when the latter was fixed at a low value.
The evolutionary dynamics of these parameters under different environmental regimes are shown in Figures 4D–F, along with example heteroplasmy distributions in a single population of cells. In all cases, as mutation rate and segregation strength increase in concert from their zero initial state, the population becomes progressively more able to adapt to the changing environments, eventually switching readily and rapidly from one environment to the other. The systematic increases observed in Figures 4A–C demonstrate that the increase of these parameters from their zero initial state is not just a consequence of random drift from their initial conditions; rather, a beneficial value is being consistently identified across simulation instances for a given environment. In parallel, the population structure remains robust as before, with a high proportion of homoplasmy, even compared to the case without fluctuating environmental conditions.
Taken together, these results suggest that organelle heteroplasmy and bottlenecks allow an intracellular resolution to the robustness-evolvability “paradox” (Wagner, 2008) (Figure 5). Specifically, a combination of intermediate mutation rate and segregation through a bottleneck allow organelle populations to resolve the tradeoff between robustness to mutational pressure and the ability to adapt to new environments. Low mutation rates alone allow a robust population but are insufficient to allow adaptation; high mutation rates alone allow (eventual) adaptation but lead to a less homoplasmic, less robust population. Intermediate mutation rates with segregation allow both rapid adaptation and the maintenance of robust homoplasmy in the population. We found that this combined “strategy” emerges under a wide range of fluctuating environmental conditions, and that a limited level of natural heteroplasmy emerges as a consequence in evolving populations. This emergent heteroplasmy can be interpreted as a mechanism for “hedging bets” against future environmental challenges (Fraser and Kaern, 2009; Tufto, 2015)—increasing evolvability—while its limited nature confers a degree of robustness to mutational challenge in the current environment. Heteroplasmy retains some within-cell genetic variance that can be rapidly exploited by a bottleneck in the case of an environmental shift.
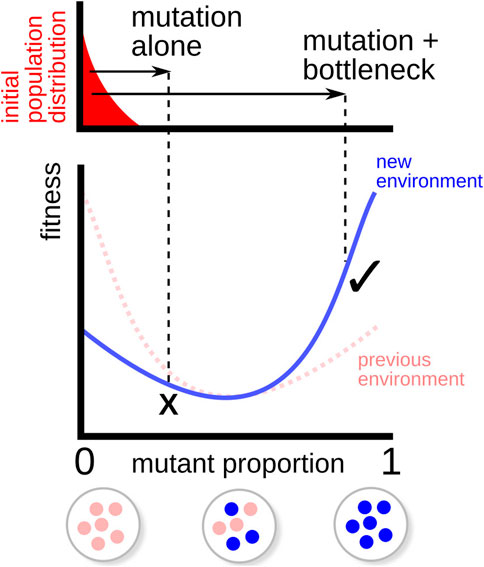
FIGURE 5. Illustration of fitness valley traversal. A population (top, red) is initially adapted to a particular environment (fitness function in dashed line), favouring a particular complement of organelle genomes (bottom). When the environment changes (fitness function in solid line), the optimal complement changes too, but the fitness penalty associated with mixed organelle populations prevents “hill-climbing” adaptation to the new optimum through mutation alone (×). When combined with a genetic bottleneck, the moderate heteroplasmy arising from mutation allows some offspring to cross this fitness valley and adapt more readily to the new environment (✓).
Rather than solely being an error associated with imperfect replication and mutagen action, these results suggest that a limited amount of organelle heteroplasmy can be an evolutionarily beneficial property for systems in fluctuating conditions. We have recently suggested analogous results for paternal leakage of organelle DNA (Radzvilavicius and Johnston, 2020), potentially providing another source of genetic variability to facilitate environmental adaptation. In both cases, the machinery governing inheritance and maintenance of organelle genomes is itself ultimately subject to evolutonary pressures. Neglecting fundamental physical limits on the fidelity of copying and transmission (Lestas et al., 2010), it may therefore be surprising if either feature was retained if it provided an exclusively negative evolutionary influence. These results suggest a possible positive influence contributing to the maintenance of these features. If biologically representative, this theory then jointly explains the presence of otherwise detrimental heteroplasmy (Wallace and Chalkia, 2013) and provide a mechanism by which evolvability is itself evolvable in this system (Pigliucci, 2008).
While it is often challenging to experimentally test evolutionary hypotheses, several observations are compatible with the predictions of our theory. The magnitudes of normalised heteroplasmy level variance V′(h) observed in experiments vary between genotypes and systems [with a range around 0.005–0.4 (Johnston, 2019)], but are not incompatible with the σ ≃ 0.2–0.4 values that emerge from our evolutionary simulation. V′(h) values around 0.2 are found, for example, in the inheritance of the disease-causing 3,243 mutation in humans (Pallotti et al., 2014; Otten et al., 2016; Wilson et al., 2016), for other human genotypes (E Bendall et al., 1996) [higher values are found for the 8,993 mutation (Otten et al., 2016; Wilson et al., 2016)] and for some observations in mice (Jenuth et al., 1996). Values of V′(h) around 0.1 are observed more commonly, for other human genotypes and species including insects, mice, primates, and humans (Rand and Harrison, 1986; Neil Howell et al., 1992; Marchington et al., 1997; Brown et al., 2001; Li et al., 2016; Burgstaller et al., 2018). Of course, in biology, this variance must be generated by physical mechanisms (including subsampling, turnover, and cell divisions), which may limit the ability of a system to achieve high segregation strengths.
Our model represents mutation and segregation processes through coarse-grained perturbation kernels. In reality, mutation and segregation can occur through many specific microscopic mechanisms, which previous and ongoing work has characterised in quantitative detail [reviewed in (Garone et al., 2018; Johnston, 2019)]. For example, the timing and mechanism of the mammalian mtDNA “bottleneck” remains debated, with several mechanisms taking place at various developmental and lifetime stages potentially responsible (Johnston et al., 2015; Zhang et al., 2018; Edwards et al., 2021). Segregation in other taxa is less studied still and may involve a role for gene conversion (Edwards et al., 2021; Broz et al., 2022). Both selection and segregation are also shaped by mitochondrial dynamics and network structure within cells (Gilad et al., 2008; Tam et al., 2013; Aryaman et al., 2019; Lieber et al., 2019; Edwards et al., 2021; Glastad and Johnston, 2022). Our model is deliberately coarse-grained and thus does not depend on the microscopic details of these processes, just their net effect between generations. We intend here to demonstrate the general principles behind this behaviour: detailed quantitative comparison between the predictions of our model and the magnitude of these effects in individual species will constitute followup work.
Regarding the qualitative predictions that our theory makes, technological advances have revealed the ubiquitous presence of mtDNA “microheteroplasmy,” the limited presence of a range of genetically diverse mtDNA types, in mammalian cells (Morris et al., 2017; Aryaman et al., 2019). The prediction that low-level heteroplasmy is present and inherited between generations also has experimental support in humans (Guo et al., 2013). Rapid shifting of mtDNA types, even on the timescale of a single generation, was originally observed decades ago in cattle (Ashley et al., 1989) (giving rise to the bottleneck picture). The phenomenon of “substoichiometric shifting” (SSS) in plant biology provides another example (Ricardo et al., 2003; McCauley, 2002013). In SSS, mitotypes that are initially present at levels much less than one per cell in one generation can rapidly come to dominate organisms in the next generation, providing a dramatic example of fast changing mtDNA populations. Plant mtDNA undergoes frequent recombination, which theory (Edwards et al., 2021) and recent experiments (Broz et al., 2022) suggests can provide a powerful segregation effect that may contribute to this rapid shifting. Our theory would suggest that the unusual strength of this capacity for segregation in plants may have evolved due to their status as sessile organisms, consequently exposed to substantial environmental fluctuations (Johnston, 2018). Conversely, we may expect more limited segregation to be observed in organisms less subject to environmental fluctuations. Comparing the extent of mutational change and the capacity and extent of oDNA segregation across organisms from diverse environments, accounting for phylogenetic correlations, would allow further testing of this hypothesis.
Publicly available datasets were analyzed in this study. This data can be found here: www.github.com/StochasticBiology/organelle-evolvability.
IJ conceived the study, wrote the code and performed the simulations, analysed the data, drafted the manuscript, and critically revised the manuscript; AR conceived the study, analysed the data, and critically revised the manuscript.
This project has received funding from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme [Grant agreement No. 805046 (EvoConBiO) to IJ].
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2022.974472/full#supplementary-material
SUPPLEMENTARY FIGURE S1 | Limited influence of demographic parameters on adaptation behaviour. Mean proportion of adapted oDNAs following an environmental change, with and without segregation for different population sizes N and offspring numbers Noff.
SUPPLEMENTARY FIGURE S2 | Limited influence of oDNA copy number on adaptation behaviour. Mean proportion of adapted oDNAs following an environmental change, with and without segregation for different cellular oDNA copy numbers n.
SUPPLEMENTARY FIGURE S3 | Mutations inducing dysfunctional oDNA limit feasible mutation rates but preserve adaptation behaviour.(A) Mean mutant proportion h and (B) mean dysfunctional proportion hD following an environmental change, with different α, where µD = αµ is the mutation rate leading to dysfunctional oDNA. In black regions, population viability was lost (all oDNAs in a cell became dysfunctional). This viability loss appears patchily in some regions due to sampling (each pixel is the average of ten simulations; if any of these simulations lose all functional oDNA, viability loss is reported). Outside these regions, segregation and mutation facilitated adaptation as in previous cases.
SUPPLEMENTARY FIGURE S4 | Adaptation behaviour under alternative fitness functions. Mean proportion of adapted genomes for (A) quadratic fitness penalty 4ε (h − 1/2)2; (B) large threshold (fixed penalty ε for h > 0.5); (C) small threshold (fixed penalty ε for h > 0.1). As before, adaptation is facilitated by higher segregation strength (right).
SUPPLEMENTARY FIGURE S5 | Evolution of the bottleneck under fixed mutation rates. Segregation strength σ emerging from evolutionary simulations (as in the main text) in fluctuating environments, for different fixed mutation rate µ and fitness penalty ε . At lower µ, stronger segregation evolves to compensate.
Acton, B. M., Lai, I., Shang, X., Jurisicova, A., and Casper, R. F. (2007). Neutral mitochondrial heteroplasmy alters physiological function in mice. Biol. Reprod. 77 (3), 569–576. doi:10.1095/biolreprod.107.060806
Aryaman, J., Johnston, I. G., and Jones, N. S. (2019). Mitochondrial heterogeneity. Front. Genet. 9, 718. doi:10.3389/fgene.2018.00718
Ashley, M. V., Laipis, P. J., and Hauswirth, W. W. (1989). Rapid segregation of heteroplasmic bovine mitochondria. Nucleic Acids Res. 17 (18), 7325–7331. doi:10.1093/nar/17.18.7325
Bergstrom, C. T., and Pritchard, J. (1998). Germline bottlenecks and the evolutionary maintenance of mitochondrial genomes. Genetics 149 (4), 2135–2146. doi:10.1093/genetics/149.4.2135
Birky, C. W. (2001). The inheritance of genes in mitochondria and chloroplasts: Laws, mechanisms, and models. Annu. Rev. Genet. 35 (1), 125–148. doi:10.1146/annurev.genet.35.102401.090231
Blier, P. U., Dufresne, F., and Burton, R. S. (2001). Natural selection and the evolution of mtdna-encoded peptides: Evidence for intergenomic co-adaptation. Trends Genet. 17 (7), 400–406. doi:10.1016/s0168-9525(01)02338-1
Brown, D. T., Samuels, D. C., Michael, E. M., Turnbull, D. M., and Chinnery, P. F. (2001). Random genetic drift determines the level of mutant mtdna in human primary oocytes. Am. J. Hum. Genet. 68 (2), 533–536. doi:10.1086/318190
Broz, A. K., Keene, A., Fernandes Gyorfy, M., Hodous, M., Johnston, I. G., and Sloan, D. B (2022). Sorting of mitochondrial and plastid heteroplasmy in Arabidopsis is extremely rapid and depends on MSH1 activity Proceedings of the National Academy of Sciences, 119 (34), p.e2206973119.
Burgstaller, J. P., Kolbe, T., Havlicek, V., Hembach, S., Poulton, J., Piálek, J., et al. (2018). Large-scale genetic analysis reveals mammalian mtdna heteroplasmy dynamics and variance increase through lifetimes and generations. Nat. Commun. 9 (2488), 1–12. doi:10.1038/s41467-018-04797-2
Christie, J. R., Schaerf, T. M., and Beekman, M. (2015). Selection against heteroplasmy explains the evolution of uniparental inheritance of mitochondria. PLoS Genet. 11 (4), e1005112. doi:10.1371/journal.pgen.1005112
Dowling, D. K., Abiega, K. C., and Arnqvist, G. (2007). Temperature-specific outcomes of cytoplasmic-nuclear interactions on egg-to-adult development time in seed beetles. Evolution 61 (1), 194–201. doi:10.1111/j.1558-5646.2007.00016.x
Drost, J. B., and Lee, W. R. (1995). Biological basis of germline mutation: Comparisons of spontaneous germline mutation rates among drosophila, mouse, and human. Environ. Mol. Mutagen. 25 (2), 48–64. doi:10.1002/em.2850250609
E Bendall, K., Macaulay, V. A., JulietBaker, R., and Sykes, B. C. (1996). Heteroplasmic point mutations in the human mtdna control region. Am. J. Hum. Genet. 59 (6), 1276–1287.
Edwards, D. M., Røyrvik, E. C., Chustecki, J. M., Giannakis, K., Glastad, R. C., Radzvilavicius, A. L., et al. (2021). Avoiding organelle mutational meltdown across eukaryotes with or without a germline bottleneck. PLoS Biol. 19 (4), e3001153. doi:10.1371/journal.pbio.3001153
Fraser, D., and Kaern, M. (2009). A chance at survival: Gene expression noise and phenotypic diversification strategies. Mol. Microbiol. 71 (6), 1333–1340. doi:10.1111/j.1365-2958.2009.06605.x
Freyer, C., Lynsey, M., Mourier, A., James, B., Koolmeister, C., Milenkovic, D., et al. (2012). Variation in germline mtdna heteroplasmy is determined prenatally but modified during subsequent transmission. Nat. Genet. 44 (11), 1282–1285. doi:10.1038/ng.2427
Garone, C., Minczuk, M., Zhang, H., Burr, S. P., and Chinnery, P. F. (2018). The mitochondrial dna genetic bottleneck: Inheritance and beyond. Essays Biochem. 62 (3), 225–234. doi:10.1042/EBC20170096
Gilad, T., Hyde, B., and Orian, S. (2008). Mitochondrial fusion, fission and autophagy as a quality control axis: The bioenergetic view. Biochim. Biophys. Acta 1777 (9), 1092–1097. doi:10.1016/j.bbabio.2008.05.001
Glastad, R. C., and Johnston, I. Mitochondrial network structure controls cell-to-cell mtdna variability generated by cell divisions. bioRxiv, 2022.
Guo, Y., Li, C-I., Sheng, Q., Winther, J. F., Cai, Q., Boice, J. D., et al. (2013). Very low-level heteroplasmy mtdna variations are inherited in humans. J. Genet. genomics 40 (12), 607–615. doi:10.1016/j.jgg.2013.10.003
Hoitzing, H., Gammage, P. A., Van Haute, L., Minczuk, M., Johnston, I. G., and Jones, N. S. (2019). Energetic costs of cellular and therapeutic control of stochastic mitochondrial dna populations. PLoS Comput. Biol. 15 (6), e1007023. doi:10.1371/journal.pcbi.1007023
James, B. W. (1992). Intracellular selection, conversion bias, and the expected substitution rate of organelle genes. Genetics 130 (4), 939–946. doi:10.1093/genetics/130.4.939
Jenuth, J. P., Peterson, A. C., Fu, K., and Shoubridge, E. A. (1996). Random genetic drift in the female germline explains the rapid segregation of mammalian mitochondrial dna. Nat. Genet. 14 (2), 146–151. doi:10.1038/ng1096-146
Johnston, I. G., and Burgstaller, J. P. (2019). Evolving mtdna populations within cells. Biochem. Soc. Trans. 31, 1367–1382. doi:10.1042/BST20190238
Johnston, I. G., Burgstaller, J. P., Havlicek, V., Kolbe, T., Rülicke, T., Brem, G., et al. (2015). Stochastic modelling, Bayesian inference, and new in vivo measurements elucidate the debated mtDNA bottleneck mechanism. Elife 4, e07464. doi:10.7554/eLife.07464
Johnston, I. G. (2018). Tension and resolution: Dynamic, evolving populations of organelle genomes within plant cells. Mol. plant 12. doi:10.1016/j.molp.2018.11.002
Johnston, I. G. (2019). Varied mechanisms and models for the varying mitochondrial bottleneck. Front. Cell Dev. Biol. 7, 294. doi:10.3389/fcell.2019.00294
Kirschner, M., and Gerhart, J. (1998). Evolvability. Proc. Natl. Acad. Sci. U. S. A. 95 (15), 8420–8427. doi:10.1073/pnas.95.15.8420
Kivisild, T., Shen, P., Wall, D. P., Bao, D., Sung, R., Davis, K., et al. (2006). The role of selection in the evolution of human mitochondrial genomes. Genetics 172 (1), 373–387. doi:10.1534/genetics.105.043901
Krakauer, D. C., Mira, A., and MirA, A. (1999). Mitochondria and germ-cell death. Nature 400 (6740), 125–126. doi:10.1038/22026
Lane, N. (2012). The problem with mixing mitochondria. Cell 151 (2), 246–248. doi:10.1016/j.cell.2012.09.028
Latorre-Pellicer, A., Lechuga-Vieco, A. V., Johnston, I. G., Hämäläinen, R. H., Pellico, J., Justo-Méndez, R., et al. (2019). Regulation of mother-to-offspring transmission of mtDNA heteroplasmy. Cell metab, 30 (6), 1120–1130.
Lechuga-Vieco, A. V, Latorre-Pellicer, A., Johnston, I. G., Prota, G., Gileadi, U., Justo-Méndez, R., et al. (2020). Cell identity and nucleo-mitochondrial genetic context modulate oxphos performance and determine somatic heteroplasmy dynamics. Sci. Adv. 6 (31), eaba5345. doi:10.1126/sciadv.aba5345
Lestas, I., Vinnicombe, G., and Paulsson, J. (2010). Fundamental limits on the suppression of molecular fluctuations. Nature 467 (7312), 174–178. doi:10.1038/nature09333
Li, M., Rothwell, R., Vermaat, M., Wachsmuth, M., Roland, S., Laros, J. F. J., et al. (2016). Transmission of human mtdna heteroplasmy in the genome of The Netherlands families: Support for a variable-size bottleneck. Genome Res. 26 (4), 417–426. doi:10.1101/gr.203216.115
Lieber, T., Jeedigunta, S. P., Palozzi, J. M., Lehmann, R., and Hurd, T. R. (2019). Mitochondrial fragmentation drives selective removal of deleterious mtdna in the germline. Nature 570 (7761), 380–384. doi:10.1038/s41586-019-1213-4
Lynch, M., and Blanchard, J. L. (1998). Deleterious mutation accumulation in organelle genomes. Mutat. Evol., 29–39.
Lynch, M., Koskella, B., and Schaack, S. (2006). Mutation pressure and the evolution of organelle genomic architecture. Science 311 (5768), 1727–1730. doi:10.1126/science.1118884
Marchington, D. R., Hartshorne, G. M., Barlow, D., and Poulton, J. (1997). Homopolymeric tract heteroplasmy in mtdna from tissues and single oocytes: Support for a genetic bottleneck. Am. J. Hum. Genet. 60 (2), 408–416.
Masel, J., and Meredith, V. T. (2010). Robustness and evolvability. Trends Genet. 26 (9), 406–414. doi:10.1016/j.tig.2010.06.002
McCauley, D. E. (2002). Paternal leakage, heteroplasmy, and the evolution of plant mitochondrial genomes. New Phytol. 200 (4), 966–977. doi:10.1111/nph.12431
Mishmar, D., Ruiz-Pesini, E., Golik, P., Macaulay, V., Clark, A. G., Hosseini, S., et al. (2003). Natural selection shaped regional mtdna variation in humans. Proc. Natl. Acad. Sci. U. S. A. 100 (1), 171–176. doi:10.1073/pnas.0136972100
Morris, J., Young-Ji, N., Zhu, H., Lee, J-H., Giang, H., Alexandra, V., et al. (2017). Pervasive within-mitochondrion single-nucleotide variant heteroplasmy as revealed by single-mitochondrion sequencing. Cell Rep. 21 (10), 2706–2713. doi:10.1016/j.celrep.2017.11.031
Muller, H. J. (1964). The relation of recombination to mutational advance. Mutat. Res. 1 (1), 2–9. doi:10.1016/0027-5107(64)90047-8
Neil Howell, S., Kubacka, I., McCullough, D. A., Bindoff, L. A., and Turnbull, D. M. (1992). Mitochondrial gene segregation in mammals: Is the bottleneck always narrow? Hum. Genet. 90 (1-2), 117–120. doi:10.1007/BF00210753
Otten, A. B., Tom, E. J., Derhaag, J. G., Ellen, H., Boesten, I. B. W., Winandy, M., et al. (2016). Differences in strength and timing of the mtdna bottleneck between zebrafish germline and non-germline cells. Cell Rep. 16 (3), 622–630. doi:10.1016/j.celrep.2016.06.023
Pallotti, F., Binelli, G., Fabbri, R., Valentino, M. L., Vicenti, R., Macciocca, M., et al. (2014). A wide range of 3243A>G/tRNALeu(UUR) (MELAS) mutation loads may segregate in offspring through the female germline bottleneck. PLoS One 9 (5), e96663. doi:10.1371/journal.pone.0096663
Pereira, L., Soares, P., Triska, P., van der Waerden, A., Li, B., Radivojac, P., et al. (2014). Global human frequencies of predicted nuclear pathogenic variants and the role played by protein hydrophobicity in pathogenicity potential. Sci. Rep. 4 (1), 7155–7158. doi:10.1038/srep07155
Peter, R., Zaidi, A., Makova, K., and Nielsen, R. (2018). A population phylogenetic view of mitochondrial heteroplasmy. Genetics 208 (3), 1261–1274. doi:10.1534/genetics.118.300711
Picard, M., Zhang, J., Hancock, S., Derbeneva, O., Ryan, G., Golik, P., et al. (2014). Progressive increase in mtDNA 3243A>G heteroplasmy causes abrupt transcriptional reprogramming. Proc. Natl. Acad. Sci. U. S. A. 111 (38), E4033–E4042. doi:10.1073/pnas.1414028111
Radzvilavicius, A., and Johnston, I. G. (2020). Paternal leakage of organelles can improve adaptation to changing environments. bioRxiv. doi:10.1101/2020.12.18.423500
Rand, D. M., and Harrison, R. G. (1986). Mitochondrial dna transmission genetics in crickets. Genetics 114 (3), 955–970. doi:10.1093/genetics/114.3.955
Rebolledo-Jaramillo, B., Su, M. S-W., Stoler, N., McElhoe, J. A., Dickins, B., Blankenberg, D., et al. (2014). Maternal age effect and severe germ-line bottleneck in the inheritance of human mitochondrial dna. Proc. Natl. Acad. Sci. U. S. A. 111 (43), 15474–15479. doi:10.1073/pnas.1409328111
Ricardo, V., Ryan, Y., Elo, A., Christensen, A. C., Meyer-Gauen, G., and Mackenzie, S. A. (2003). Substoichiometric shifting in the plant mitochondrial genome is influenced by a gene homologous to muts. Proc. Natl. Acad. Sci. U. S. A. 100 (10), 5968–5973. doi:10.1073/pnas.1037651100
Rossignol, R., Faustin, B., Rocher, C., Malgat, M., Mazat, J-P., and Letellier, T. (2003). Mitochondrial threshold effects. Biochem. J. 370 (3), 751–762. doi:10.1042/BJ20021594
Roze, D., Rousset, F., and Michalakis, Y. (2005). Germline bottlenecks, biparental inheritance and selection on mitochondrial variants: A two-level selection model. Genetics 170 (3), 1385–1399. doi:10.1534/genetics.104.039495
Ruiz-Pesini, E., Marie, T., Vincent, P., Poole, J. C., Brandon, M. C., Mishmar, D., et al. (2006). An enhanced mitomap with a global mtdna mutational phylogeny. Nucleic Acids Res. 35 (1), D823–D828. doi:10.1093/nar/gkl927
Ruiz-Pesini, E., Mishmar, D., Martin, B., Vincent, P., and Wallace, D. C. (2004). Effects of purifying and adaptive selection on regional variation in human mtdna. Science 303 (5655), 223–226. doi:10.1126/science.1088434
Sharpley, M. S., Marciniak, C., Eckel-Mahan, K., McManus, M., Crimi, M., Waymire, K., et al. (2012). Heteroplasmy of mouse mtdna is genetically unstable and results in altered behavior and cognition. Cell 151 (2), 333–343. doi:10.1016/j.cell.2012.09.004
Stewart, J. B., and Chinnery, P. F. (2015). The dynamics of mitochondrial dna heteroplasmy: Implications for human health and disease. Nat. Rev. Genet. 16 (9), 530–542. doi:10.1038/nrg3966
Sun, C., Kong, Q-P., and Zhang, Y-P. (2007). The role of climate in human mitochondrial dna evolution: A reappraisal. Genomics 89 (3), 338–342. doi:10.1016/j.ygeno.2006.11.005
Tam, Z. Y., Gruber, J., Halliwell, B., and Gunawan, R. (2013). Mathematical modeling of the role of mitochondrial fusion and fission in mitochondrial dna maintenance. PloS one 8 (10), e76230. doi:10.1371/journal.pone.0076230
Tufto, J. (2015). Genetic evolution, plasticity, and bet-hedging as adaptive responses to temporally autocorrelated fluctuating selection: A quantitative genetic model. Evolution 69 (8), 2034–2049. doi:10.1111/evo.12716
Wagner, A. (2008). Robustness and evolvability: A paradox resolved. Proc. Biol. Sci. 275, 91–100. doi:10.1098/rspb.2007.1137
Wallace, D. C. (2010). Bioenergetics, the origins of complexity, and the ascent of man. Proc. Natl. Acad. Sci. U. S. A. 107 (2), 8947–8953. doi:10.1073/pnas.0914635107
Wallace, D. C., and Chalkia, D. (2013). Mitochondrial dna genetics and the heteroplasmy conundrum in evolution and disease. Cold Spring Harb. Perspect. Biol. 5 (11), a021220. doi:10.1101/cshperspect.a021220
Wei, W., Tuna, S., Keogh, M. J., Beales, P. L., Bennett, D. L., Black, G. C., et al. (2019). Germline selection shapes human mitochondrial dna diversity. Science 364 (6442), eaau6520. doi:10.1126/science.aau6520
Wilson, I. J., J Carling, P., Alston, C. L., Vasileios, I., Pyle, A., Hudson, G., et al. (2016). Mitochondrial dna sequence characteristics modulate the size of the genetic bottleneck. Hum. Mol. Genet. 25 (5), 1031–1041. doi:10.1093/hmg/ddv626
Wright, S. (1942). Statistical genetics and evolution. Bull. Amer. Math. Soc. 48 (4), 223–246. doi:10.1090/s0002-9904-1942-07641-5
Xu, J. (2005). The inheritance of organelle genes and genomes: Patterns and mechanisms. Genome 48 (6), 951–958. doi:10.1139/g05-082
Keywords: mtDNA, ptDNA, evolution, evolvability, heteroplasmy, changing environments, bottleneck
Citation: Radzvilavicius AL and Johnston IG (2022) Organelle bottlenecks facilitate evolvability by traversing heteroplasmic fitness valleys. Front. Genet. 13:974472. doi: 10.3389/fgene.2022.974472
Received: 21 June 2022; Accepted: 11 October 2022;
Published: 28 October 2022.
Edited by:
Henry H. Heng, Wayne State University, United StatesReviewed by:
Arslan A. Zaidi, University of Pennsylvania, United StatesCopyright © 2022 Radzvilavicius and Johnston. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Iain G. Johnston, aWFpbi5qb2huc3RvbkB1aWIubm8=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.