 Sihle E. Mabhida1,2Jyoti R. Sharma1Teke Apalata3Charity Masilela4Sibusiso Nomatshila5Lawrence Mabasa1Hannah Fokkens1,7Mongi Benjeddou2Babu Muhamed6Samukelisiwe Shabalala1Rabia Johnson1,7*
Sihle E. Mabhida1,2Jyoti R. Sharma1Teke Apalata3Charity Masilela4Sibusiso Nomatshila5Lawrence Mabasa1Hannah Fokkens1,7Mongi Benjeddou2Babu Muhamed6Samukelisiwe Shabalala1Rabia Johnson1,7*- 1Biomedical Research and Innovation Platform, SAMRC, Tygerberg, South Africa
- 2Department of Biotechnology, Faculty of Natural Science, University of the Western Cape, Cape Town, South Africa
- 3Division of Medical Microbiology, Department of Laboratory-Medicine and Pathology, Faculty of Health Sciences, Walter Sisulu University and National Health Laboratory Services, Mthatha, South Africa
- 4Department of Biochemistry, North-West University, Mmabatho, South Africa
- 5Division of Preventive Medicine and Health Behavior, Department of Public Health, Faculty of Health Sciences, Walter Sisulu University, Mthatha, South Africa
- 6Division of Infections Disease - University of Tennessee Health Sciences Center (UTHSC), Memphis, TN, United States
- 7Centre for Cardio-Metabolic Research in Africa, Division of Medical Physiology, Faculty of Medicine and Health Sciences, Stellenbosch University, Tygerberg, South Africa
Aims: The current study sought to investigate the association between the methylenetetrahydrofolate reductase (MTHFR) variant (rs1801133) and the risk of developing hypertension (HTN) in an indigenous South African population.
Methods: A total of 442 participants (hypertensive, n = 279 and non-hypertensive, n = 163) from the indigenous tribe residing in Mthatha, Eastern Cape (South Africa) were recruited. HTN was defined as a systolic (SBP) and diastolic blood pressure (DBP) of ≥130/80 mmHg following American Heart Association guidelines. The genotyping of MTHFR (rs1801133) was assessed using MassARRAY® System. Thereafter, the association between rs1801133 in various genetic models and HTN was determined by logistic regression model analysis. Furthermore, the interaction between rs1801133 and selected risk factors on HTN was performed using the open-source multifactor dimensionality reduction (MDR).
Results: The low frequency of the T allele (5%) was also observed when compared with the C allele (95%) in both cases and controls. After adjusting for confounding factors (gender, smoking status, BMI, and blood glucose levels), there were no significant associations were observed between rs1801133 and the risk of HTN in all genetic models: genotypic (OR 0.75, 95% CI 0.29–1.95, p = 0.56), dominant (OR 0.86, 95% CI 0.35–2.16, p = 0.75), co-dominant (OR 1.33, 95% CI 0.51–3.48, p = 0.55) and allelic (OR 0.80, 95% CI 0.49–1.62, p = 0.70) in logistic regression analysis. However, a significant interaction was reported among rs1801133, age, and gender (p < 0.0001) with the risk of HTN.
Conclusion: The present study reports on the lack of association between MTHFR (rs1801133) and the risk of HTN in an indigenous South African tribe. However, an interaction between gender, age, and rs1801133 was observed. Thus, future studies with a large sample size are required to further validate these findings.
Introduction
Hypertension (HTN) is a multifactorial disorder with genetic, environmental, and demographic factors contributing to its prevalence and progression. Over the past years, numerous researchers have uncovered the significant role of inter-individual genetic variations in the development, progression, and control of the disease (Tuder et al., 2013; Fontana et al., 2014; Bayramoglu et al., 2015). Indeed, genome-wide association studies have successfully identified several genetic variants linked to the development and the progression of HTN (Padmanabhan et al., 2008; Li et al., 2012), including methylenetetrahydrofolate reductase (MTHFR) gene variants, with rs1801133 being the most common and widely studied variant located in this gene (Xi et al., 2013; Fan et al., 2016). However, their role in the pathophysiology of HTN particularly among individuals of African descent remains unclear (Amrani-Midoun et al., 2016; Ghogomu et al., 2016).
The MTHFR gene encodes methylenetetrahydrofolate reductase, a key regulatory enzyme in folate and homocysteine (Hcy) metabolism, and in the production of nitric oxide, a potent vasodilator that is important in blood pressure regulation (Goyette et al., 1994; McMahon et al., 2016). The rs1801133 polymorphism (677 C > T) is found on exon 4 of the MTHFR gene. The consequence of this polymorphism is a change of amino acid 222 from alanine to valine, leading to a decrease in MTHFR activity and an increase in plasma Hcy levels (Tawakol et al., 1997; Heux et al., 2004). Literature suggests that elevated plasma concentration of Hcy may injure the vascular endothelium, resulting in HTN (Tawakol et al., 1997; Heux et al., 2004). While other studies have shown that rs1801133 was associated with HTN in populations (Bouzidi et al., 2020; Wu et al., 2022).
Furthermore, these studies established that carriers of rs1801133 present with high levels of plasma Hcy and are more likely to develop HTN (Guenther et al., 1999; Wu et al., 2022). Whereas a reduction in MTHFR activity was linked with the loss of riboflavin, a cofactor associated with a decrease of Hcy among carriers of the MTHFR variant (Guenther et al., 1999; Yamada et al., 2001; McNulty et al., 2006). Most importantly, accumulative studies have suggested that rs1801133 polymorphism could be an independent risk factor for HTN in different ethnic groups (Heux et al., 2004; Ilhan et al., 2008; Nassereddine et al., 2015), and it has been associated with severe diastolic HTN in pregnant women (Kosmas et al., 2004). Even so, conflicting results have been obtained, particularly among African population studies (Al-Kordy et al., 2012; Nassereddine et al., 2015; Amrani-Midoun et al., 2016; Ghogomu et al., 2016). Thus, several authors have emphasized the need to conduct more African studies to further validate the role of MTHFR polymorphisms in the pathogenesis of HTN (Amrani-Midoun et al., 2016; Ghogomu et al., 2016; Olczak et al., 2021).
Undoubtedly, HTN as a chronic medical condition, has contributed significantly to the rapid rise in global premature deaths, especially among Black African adults (Adeniyi et al., 2016; Sharma et al., 2021). Moreover, the devastating outcome has been the limited data currently informing on the incidence and pathological mechanisms driving the onset and progression of this medical condition among black African adults (Mabhida et al., 2021). Studying the genetics of HTN can revolutionize our understanding the pathophysiology of the disease and the discovery of effective treatment strategies. Importantly, our research group has actively explored this research niche within an indigenous South African population. Our group has demonstrated a high prevalence (75%) of HTN in the village of Mthatha, Eastern Cape (Adeniyi et al., 2016; Sharma et al., 2021). We further conducted a narrative synthesis of the literature that predicted a possible association between MTHFR rs1801133 and the development of HTN among individuals of African origin (Mabhida et al., 2022). Our findings highlighted the necessity of performing genomic studies on indigenous African individuals, as very few studies have addressed this research niche in a purely black African population. Thus, this study aimed to investigate the association between MTHFR (rs1801133) and HTN in the indigenous South African population of Mthatha in the Eastern Cape province of South Africa. Additionally, the study aimed to explore the interaction between single nucleotide polymorphism (SNP) and sociodemographic as well as their interaction with the risk of HTN.
Material and methods
Ethical clearance
The ethical approval for this study was provided by the Senate Research Committee of the University of the Western Cape (Ethics clearance number BM19/8/19), Water Sisulu University (073/15), and the South African Medical Research Council (EC028-8/2020). The study conforms to the ethical guidelines of the Declaration of Helsinki (Puri et al., 2009). Written informed consent in both English and IsiXhosa was obtained from all the study participants.
Power and sample size estimation
The appropriate sample and power size were estimated using the Power and Sample Size Program (PS version 3.1.2). The study initially planned to recruit approximately 430 participants (1:1 ratio). “Previous data indicate that the probability of exposure among controls is 0.75. If the true odds ratio for disease in exposed subjects relative to unexposed subjects is 2, we will need to study 215 case patients and 215 control patients to be able to reject the null hypothesis that this odds ratio equals 1 with a probability (power) 0.8. The Type I error probability associated with this test of this null hypothesis is 0.05 (Dupont and Plummer, 1990; Dupont and Plummer, 1998)”.
Study design and patient selection
A total of 442 participants belonging to the indigenous Xhosa tribe in the Eastern Cape province of South Africa, aged 18 years and above were recruited from four districts (OR Tambo, Alfred Nzo, Chris Hani, and Joe Gqabi). Participants were divided into two groups 1) the non-hypertensive group (n = 163), and 2) hypertensive group (n = 279). HTN was defined as average SBP or DBP of ≥130/80 mmHg or using at least one class of antihypertensive medication following the American Heart Association guidelines (AHA) (Whelton et al., 20172018). Blood pressure values were measured 3 consecutive times, after 10 min of seated rest before the first measurement and 5 min intervals between each measurement with a manual aneroid sphygmomanometer. SBP and DBP were determined by the first and the fifth Korotkoff sounds. The average of three consecutive measurements to the nearest 2 mmHg was recorded, with a time interval of at least 2 min. Participants were selected in general consultation in the same centres.
Data collection and sampling procedure
The data records were collected using the World Health Organization (WHO) STEPwise questionnaire which was uploaded onto the Research Electronic Data Capture (REDCap), a web-based application for building and managing online surveys and databases. Data collected included information on environmental or sociodemographic factors (smoking, age, gender), anthropometric measurements (body mass index, BMI), and blood glucose levels. All selected variables were defined as previously described by Sharma et al. (2021) (Sharma et al., 2021) Venous blood (5 ml) was collected in EDTA tubes. The samples were stored at −80°C until used. A blood sample was also taken after an overnight fast to measure the glucose levels.
Genomic DNA isolation and genotyping
Peripheral blood samples were collected from all subjects and the DNA was extracted using a QIAamp DNA Blood Midi kit (Qiagen, Valencia, CA, United States) as per the manufacturers’ instructions. DNA quantity and quality were determined using a Nano-Drop™ 2000/2000c UV/VIS Spectrophotometer (ThermoScientific™). Thereafter, the presence of MTHFR (rs1801133) was analyzed using MassARRAY® System (Agena Bioscience™).
Statistical analysis
Statistical analyses were performed using Stata/IC version 17.0 (Stata Corp, United States). The general characteristics of the participants were expressed as frequency (percentages). A chi-square test (non-parametric) was used to evaluate the genotypic distribution of the MTHFR (rs1801133) variant. Logistic regression analysis was used to examine the potential effects of the selected SNP (rs1801133) on the risk of developing HTN under various genetic models (genotypic, dominant, recessive, co-dominant, and allelic models) (Fan et al., 2016). A method suggested by Thakkinstian et al. (2005) was used to select the most appropriate genetic model. Multivariate regression analysis was carried out between rs1801133 variant and independent variables such as age, gender, smoking status, BMI and blood glucose levels. The probability of exposure given the outcomes (OR) and their 95% confidence intervals (CI) was used to present the association between the SNP and the risk of HTN. A two-tailed statistical significance was evaluated by using a p-value of <0.05. Interactions between rs1801133 and selected environmental factors on HTN were detected using the open-source multifactor dimensionality reduction (MDR) software package version 3.0.2. Minor allele frequency (MAF) and Hardy–Weinberg equilibrium (HWE) tests were calculated using Genetic Analysis in Excel (GenAIEx) Version 6.5, where a p-value < 0.05 indicated a deviation from the Hardy Weinberg principle.
Results
General characteristics of the study cohort
The present study sampled 442 individuals over the age of 18 years, of whom 19% (n = 82) were males and 81% (n = 360) were females. The mean for age was 46.96 ± 14.55 years. The distribution of baseline characteristics of categorical variables (age, gender, smoking status, BMI, and blood glucose levels) is summarized in Table 1.
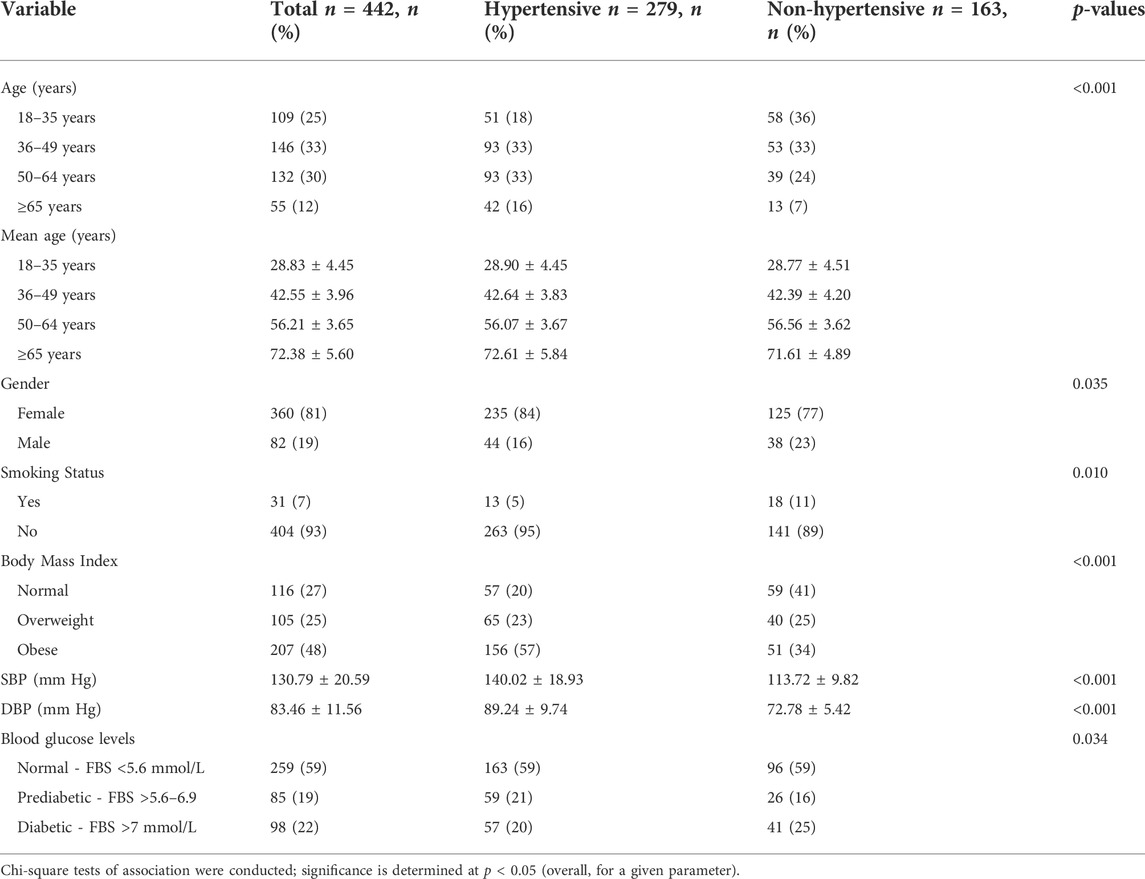
TABLE 1. General characteristics of the study cohort.
Correlation between covariates and hypertension susceptibility
In Table 2, summarize the ORs with corresponding 95% CIs for the association of environmental factors (age, gender, smoking status, BMI, and blood glucose levels) with HTN using univariate analyses. In the unadjusted model, the logistic regression analysis showed a significant association with older age (36 to ≥65) (p = 0.007 and p < 0.001 respectively), being male (p = 0.034), smoking habits (p = 0.012), obese (p < 0.001), and prediabetic/diabetic (p = 0.031) with the increasing risk of developing HTN.
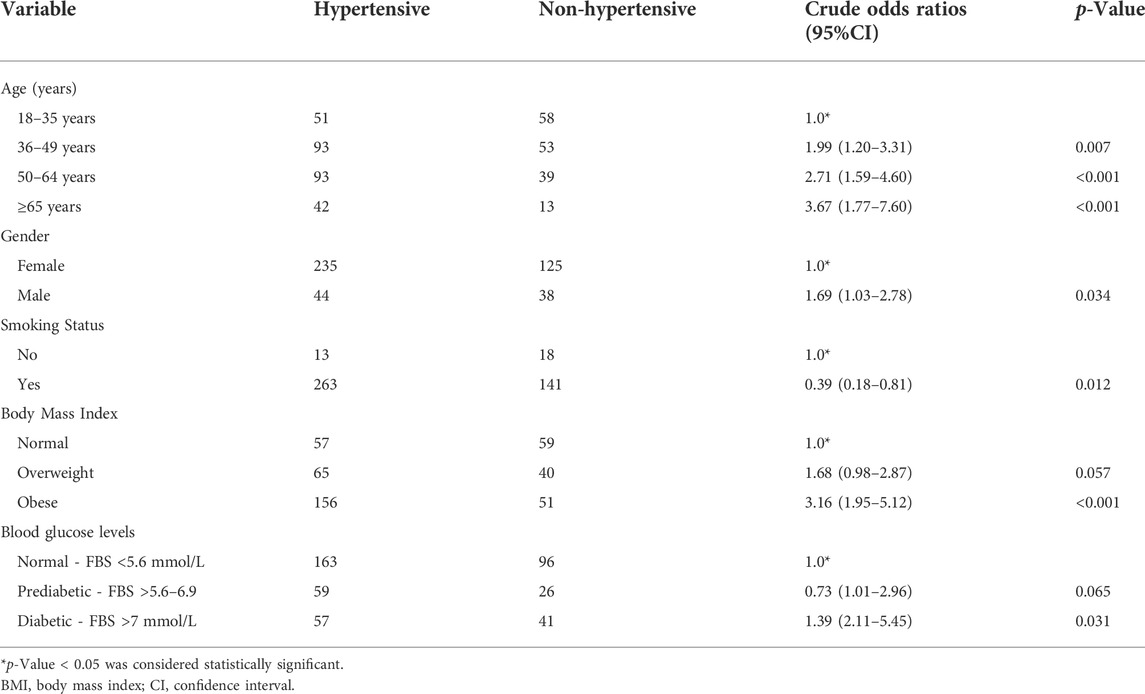
TABLE 2. Associations of age, gender, BMI, smoking status, and blood glucose levels with the risk of developing HTN.
Association between MTHFR (rs1801133) polymorphism and hypertension susceptibility
Table 3 shows the genotypic and allelic frequencies for the MTHFR (rs1801133) variant and the risk of developing HTN under genotypic, dominant, recessive, co-dominant, and allelic. The most frequently observed genotype was the homozygous CC (hypertensive, 91% and non-hypertensive, 89%) followed by heterozygous CT (hypertensive, 8% and non-hypertensive, 11%). The homozygous genotype TT had the lowest frequency in the studied population (hypertensive, 1% and non-hypertensive, 0%). Furthermore, the low frequency of the T allele of rs1801133 (5%) was also observed in both hypertensive and non-hypertensive individuals when compared with the C allele (95%). After testing for Hardy-Weinberg principle using X2 and p < 0.05, all alleles and genotypes were in agreement with the hypothesis (p = 0.069).
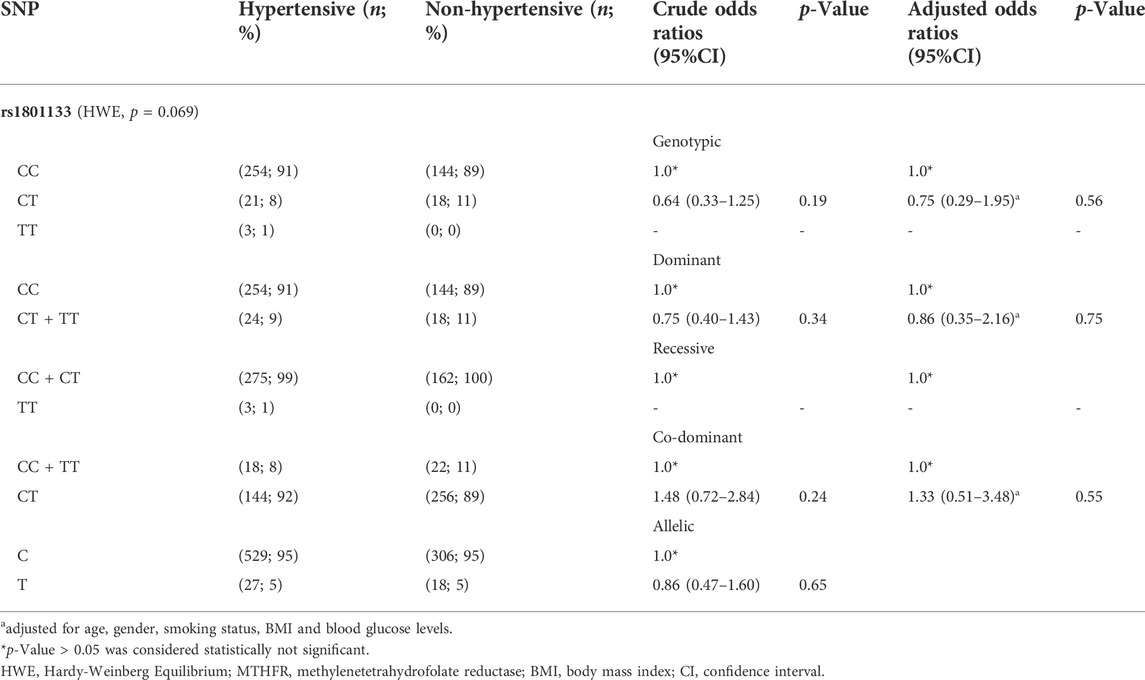
TABLE 3. Association of the MTHFR (rs1801133) polymorphism with risk of developing HTN.
In the univariate analysis, the MTHFR rs1801133 variant was not significantly associated with HTN under various inheritance models [genotypic (OR = 0.75, 95% CI = 0.29–1.95, p = 0.56), dominant (OR = 0.86, 95% CI = 0.35–2.16, p = 0.75), recessive (N/A), co-dominant (OR = 1.33, 95% CI = 1.51–3.48, p = 0.55) and allelic models (OR = 0.80, 95% CI = 0.49–1.62, p = 0.70)]. After adjusting for confounding factors such as age, gender, smoking status, BMI, and blood glucose levels, the direction of association did not change (p > 0.05).
Epistatic interactions between the genotypes of MTHFR (rs1801133) and selected variables (age, gender, smoking status, BMI, and blood glucose levels) were assessed using multifactor dimensionality reduction (MDR) as presented in Table 4. The combination of age and rs1801133 had a high CVC score (9/10), however; the model was not significantly associated with HTN (p = 0.054). On the other hand, the combination of age, rs1801133, and gender (CVC = 7/10) showed a significant association with the risk of HTN (p < 0.0001). Other possible interactions are shown in Figure 1.
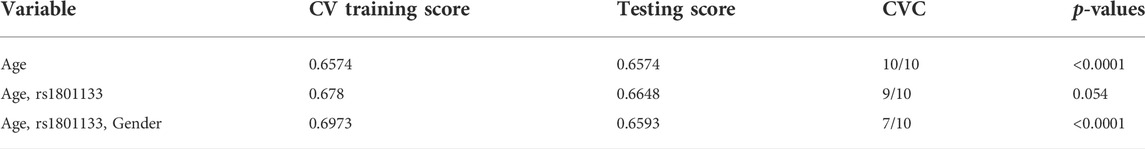
TABLE 4. Interaction models between genetic and sociodemographic factors.
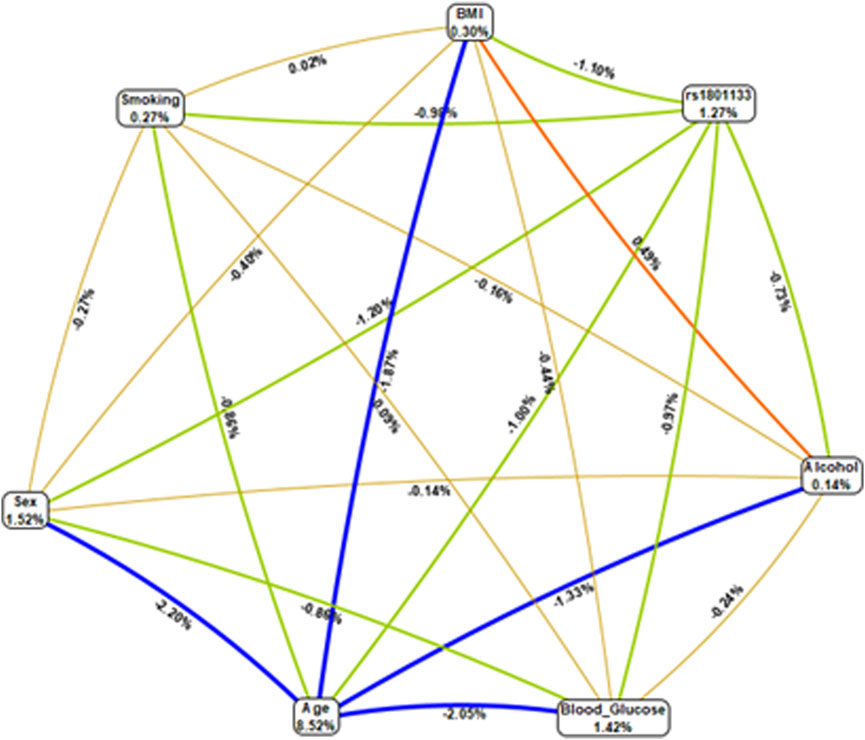
FIGURE 1. Multifactor dimensionality reduction combined attribute network demonstrating all possible interactions between rs1801133 and various environmental factors. Four different colours and various interactions between the genotypes of MTHFR (rs1801133) and selected variables (age, gender, smoking status, BMI, and blood glucose levels) were observed, and each colour represents a possible interaction. The width of the line indicates the strength of the interaction, whereas the thin lines represent weak interactions. Figures <1 and thin lines represent weak interactions. The strongest interactions are represented by figures ≥1 and thick lines. The image was generated using the open-source MDR software package version 3.0.2.
Discussion
Previous data from our group demonstrated a high prevalence (75%) of HTN in the villages of Mthatha, Eastern Cape (Adeniyi et al., 2016; Sharma et al., 2021). Through a narrative synthesis of the literature, we further reported that there is a lack of African-specific data with regards to the association between MTHFR rs1801133 and HTN (Mabhida et al., 2021; Mabhida et al., 2022); thus, highlighting the significance of the current report. The present study aimed to investigate the association between rs1801133 and HTN in an indigenous South African population from the Eastern Cape province.
The current study reports on the lack of association between MTHFR (rs1801133) and the risk of HTN among an indigenous South African population (n = 442). Our findings are consistent with results reported by Amin et al. (Al-Kordy et al., 2012), and Amrani-Midoun et al. (Amrani-Midoun et al., 2016), where they also observed no association between rs1801133 and the risk of developing HTN in a cohort of Egyptian (n = 181) and Algerian (n = 154) adults, respectively. In contrast, a study done by Ghogomu et al. (Ghogomu et al., 2016) showed a significant association between rs1801133 and HTN among a native Bantu ethnic group of the South-West region of Cameroon (n = 91). Furthermore, Nasseredine et al. (Nassereddine et al., 2015) demonstrated that individuals of Moroccan (n = 203) origin who were homozygous carriers of rs1801133 were more likely to develop HTN.
Several studies performed in non-African countries also demonstrated that genotype TT or T allele of rs1801133 may increase the risk of HTN (Fan et al., 2016; Bouzidi et al., 2020; Wu et al., 2022). The discrepancies observed between these studies may be due to geographical location, ethnicity, and epigenetic mechanisms that are involved in the regulation of MTHFR gene expression (Fu et al., 2019; Meng et al., 2021). Suggesting that data linking HTN to rs1801133 remains controversial. Li et al. (Li et al., 2005) and Rosenberg et al. (Rosenberg et al., 2002) showed that the frequency of the T allele varies across different populations. The authors further demonstrated that individuals of African origin presented a low frequency of the T allele (<10%) in comparison to other ethnic groups (Shi et al., 2003; Guéant-Rodriguez et al., 2006; Romero-Sánchez et al., 2015). This was further confirmed by a study conducted by Atadzhanov et al. (Atadzhanov et al., 2014), who also observed a low frequency of the T allele in a Zambian population. In agreement with the later studies (Atadzhanov et al., 2014), our study demonstrated that the T allele and the TT genotype frequencies of MTHFR (rs1801133) were lower (5%) than the C allele (95%) in both hypertensive and non-hypertensive individuals. We, therefore, tested for Hardy-Weinberg equilibrium, and none of the genetic models deviated from the principle. This is an indication that the genotype and allele frequencies in our study population remained constant between generations, and that there was no random mating that occurred. Our findings further suggest that a positive association between MTHFR (rs1801133) polymorphism and HTN among African individuals may not be strong enough to withstand statistical interrogation by false-positive report probability (Meng et al., 2021). Of note, the current study as well as the reference studies (Al-Kordy et al., 2012; Nassereddine et al., 2015; Amrani-Midoun et al., 2016; Ghogomu et al., 2016), were composed of relatively small sample sizes; thus, the observations made remain to be explored in future studies with larger sample sizes.
Some observational studies have shown that genetic changes are inseparable from the impact of environmental factors such as diet, exercise, smoking, and drinking (Anand, 2005; Zhong et al., 2017). As a result, SNP-environment interaction could not be ignored as our existing understanding of the pathogenesis of HTN indicates an interplay between genetic and environmental factors (Fan et al., 2016). Although no association was established between rs1801133 and HTN in the present study, an interaction between rs1801133, age, and gender was observed, suggesting a possible synergistic effect. This was in agreement with the previous studies which also reported that rs1801133, aging, and gender are potential risk factors for elevated Hcy levels (Ma et al., 2020), decreased uric acid (Massie et al., 1991), and folate deficiency (James et al., 2002), which increases the risk of HTN and hyperhomocysteinemia (Nygård et al., 1995; Refsum et al., 2006). Furthermore, Boers et al. (Boers et al., 1985), and Ueland et al. (Ueland et al., 1993), have demonstrated that premenopausal women have lower Hcy levels than men and postmenopausal women (Nygård et al., 1995; Refsum et al., 2006). The reasons for the higher Hcy concentrations at older ages are not well understood, although changes in renal function are certainly involved. Higher Hcy concentrations in men in comparison to women may be explained by differences in muscle mass, hormone, and vitamin status (Refsum et al., 2004). Thus, this suggests that the interaction between age, gender, and the MTHFR (rs1801133) polymorphism in HTN may be partially due to their interactive effects on Hcy concentrations and DNA methylation status (Selhub et al., 1993; Kotchen, 2010; Kato et al., 2015; Liu et al., 2015; Fan et al., 2016). But this hypothesis needs to be further explored in a bigger cohort.
Limitations and strengths of this study
It is noteworthy to mention that several limitations need to be considered in interpreting the findings of our study. Firstly, the sample size of our study was relatively small, of which could very likely introduce random errors and bias, resulting in unreliable results. However, this limitation projects another strength as it is often better to test a new research hypothesis on a small number of participants before progressing to a large number. This is especially important not to conserve resources. Importantly, our findings will be a stepping-stone to larger ongoing clinical and genetic studies of HTN in the South African population. Notably, also highlights another strength of the current report, as very few studies have addressed this research niche in a purely black African population. Importantly, unlike other reports (Nassereddine et al., 2015; Ghogomu et al., 2016), our study did perform an adjustment for confounding factors such as gender, age, and smoking status. This indicates an urgent need to carefully plan African-specific studies with large sample sizes to be able to draw conclusions on the association between rs1801133 and HTN. Secondly, this study used a cross-sectional design, which precludes causal inference, consistent with recruiting participants from only one ethnic group (Xhosa population of Mthatha, Eastern Cape). Further highlights that our findings are generalised to the included study population and individuals residing in similar settings in the province. Lastly, the current project was planned to recruit approximately 430 participants, with a 1:1 ratio of cases to controls. But, due to the high prevalence of the HTN (75%) in our study population, a high number of hypertensive patients were included in the study (n = 279). However, the T allele frequency of study population was not influenced by this limitation as it remained low (5%). Despite these limitations, to the best of our knowledge, this is the first study to investigating the association between the MTHFR (rs1801133) variant and HTN in South Africa, and further explored a unique and important feature of SNP-environment interaction. Overall, these findings remain important in addressing the limited data on the MTHFR variant (rs1801133) and the risk of developing HTN in indigenous populations of South Africa.
Conclusion
In the present study, we demonstrated the lack of association between MTHFR (rs1801133) and HTN among individuals of Xhosa origin. However, a potential synergistic effect of the rs1801133 with age and gender on HTN susceptibility was observed. The current study has laid a foundation for future studies by highlighting the significance of considering the impact of environmental factors in genetic association studies. Furthermore, future studies with a large sample size are required to further validate our findings.
Data availability statement
The original contributions presented in the study are included in the article/supplementary materials, further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving human participants were reviewed and approved by University of the Western Cape (Ethics clearance number BM19/8/19), Water Sisulu University (073/15), and the South African Medical Research Council (EC028-8/2020). The patients/participants provided their written informed consent to participate in this study.
Author contributions
SM and RJ conceptualized and designed the study. SM acquired the data. SM, CM, JS, and SS, analysed and interpreted the data and drafted the manuscript. BM, SN, SM, SS, TA, LM, JS, MB, HF, and RJ were involved in the critical revision of the manuscript for important intellectual content. All authors have read and approved the final version of the manuscript.
Funding
This research was funded by the South Africa Medical Research Council’s SHIP-RFA-01-2019 Strategic Health Innovation Partnership (Pharmacogenomics in Precision Medicine), Bio-medical Research and Innovation Platform baseline funding as well as Division of Research Ca-pacity Development Internship Scholarship Program (RCD), and the National Research Founda-tion (NRF) Competitive Program for Rated Researchers (UID120812) NRF bursary (No: MND190721457781).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adeniyi, O. V., Yogeswaran, P., Longo-Mbenza, B., and Ter Goon, D. (2016). Uncontrolled hypertension and its determinants in patients with concomitant type 2 diabetes mellitus (T2DM) in rural South Africa. PloS one 11 (3), e0150033. doi:10.1371/journal.pone.0150033
Al-Kordy, H., Amin, H. A. K., Fahmy, H., Aziz, A., Leheta, O. F., Kamal, H., et al. (2012). Methylenetetrahydrofolate reductase (677C/T) polymorphism in myocardial infarction and hypertension. Am. J. Biochem. Biotechnol. 8 (3), 150–156. doi:10.3844/ajbbsp.2012.150.156
Amrani-Midoun, A., Kiando, S. R., Treard, C., Jeunemaitre, X., and Bouatia-Naji, N. (2016). The relationship between MTHFR C677T gene polymorphism and essential hypertension in a sample of an Algerian population of Oran city. Int. J. Cardiol. 225, 408–411. doi:10.1016/j.ijcard.2016.10.027
Anand, S. S. (2005). The value of studying gene-environment interactions in culturally diverse populations. Can. J. Physiol. Pharmacol. 83 (1), 42–46. doi:10.1139/y05-004
Atadzhanov, M., Mwaba, M. H., Mukomena, P. N., Lakhi, S., Mwaba, P., Rayaprolu, S., et al. (2014). Frequency of APOE, MTHFR and ACE polymorphisms in the Zambian population. BMC Res. Notes 7 (1), 194. doi:10.1186/1756-0500-7-194
Bayramoglu, A., Urhan Kucuk, M., Guler, H. I., Abaci, O., Kucukkaya, Y., and Colak, E. (2015). Is there any genetic predisposition of MMP-9 gene C1562T and MTHFR gene C677T polymorphisms with essential hypertension? Cytotechnology 67 (1), 115–122. doi:10.1007/s10616-013-9665-0
Boers, G. H., Smals, A. G., Trijbels, F. J., Fowler, B., Bakkeren, J. A., Schoonderwaldt, H. C., et al. (1985). Heterozygosity for homocystinuria in premature peripheral and cerebral occlusive arterial disease. N. Engl. J. Med. 313 (12), 709–715. doi:10.1056/NEJM198509193131201
Bouzidi, N., Hassine, M., Fodha, H., Ben Messaoud, M., Maatouk, F., Gamra, H., et al. (2020). Association of the methylene-tetrahydrofolate reductase gene rs1801133 C677T variant with serum homocysteine levels, and the severity of coronary artery disease. Sci. Rep. 10 (1), 10064. doi:10.1038/s41598-020-66937-3
Dupont, W. D., and Plummer, W. D. (1998). Power and sample size calculations for studies involving linear regression. Control. Clin. Trials 19 (6), 589–601. doi:10.1016/s0197-2456(98)00037-3
Dupont, W. D., and Plummer, W. D. (1990). Power and sample size calculations: A review and computer program. Control. Clin. Trials 11 (2), 116–128. doi:10.1016/0197-2456(90)90005-m
Fan, S., Yang, B., Zhi, X., Wang, Y., Wei, J., Zheng, Q., et al. (2016). Interactions of methylenetetrahydrofolate reductase C677T polymorphism with environmental factors on hypertension susceptibility. Int. J. Environ. Res. Public Health 13 (6), 601. doi:10.3390/ijerph13060601
Fontana, V., McDonough, C. W., Gong, Y., El Rouby, N. M., Sa, A. C. C., Taylor, K. D., et al. (2014). Large-scale gene-centric analysis identifies polymorphisms for resistant hypertension. J. Am. Heart Assoc. 3 (6), e001398. doi:10.1161/JAHA.114.001398
Fu, L., Li, Y. N., Luo, D., Deng, S., Wu, B., and Hu, Y. Q. (2019). Evidence on the causal link between homocysteine and hypertension from a meta‐analysis of 40 173 individuals implementing Mendelian randomization. J. Clin. Hypertens. 21 (12), 1879–1894. doi:10.1111/jch.13737
Ghogomu, S., Ngolle, N. E., Mouliom, R. N., and Asa, B. F. (2016). Association between the MTHFR C677T gene polymorphism and essential hypertension in South West Cameroon. Genet. Mol. Res. 15 (1), 28. doi:10.4238/gmr.15017462
Goyette, P., Sumner, J. S., Milos, R., Duncan, A. M., Rosenblatt, D. S., Matthews, R. G., et al. (1994). Human methylenetetrahydrofolate reductase: Isolation of cDNA, mapping and mutation identification. Nat. Genet. 7 (2), 195–200. doi:10.1038/ng0694-195
Guéant-Rodriguez, R.-M., Gueant, J. L., Debard, R., Thirion, S., Hong, L. X., Bronowicki, J. P., et al. (2006). Prevalence of methylenetetrahydrofolate reductase 677T and 1298C alleles and folate status: A comparative study in Mexican, west african, and European populations. Am. J. Clin. Nutr. 83 (3), 701–707. doi:10.1093/ajcn.83.3.701
Guenther, B. D., Sheppard, C. A., Tran, P., Rozen, R., Matthews, R. G., and Ludwig, M. L. (1999). The structure and properties of methylenetetrahydrofolate reductase from Escherichia coli suggest how folate ameliorates human hyperhomocysteinemia. Nat. Struct. Biol. 6 (4), 359–365. doi:10.1038/7594
Heux, S., Morin, F., Lea, R. A., Ovcaric, M., Tajouri, L., and Griffiths, L. R. (2004). The methylentetrahydrofolate reductase gene variant (C677T) as a risk factor for essential hypertension in Caucasians. Hypertens. Res. 27 (9), 663–667. doi:10.1291/hypres.27.663
Ilhan, N., Kucuksu, M., Kaman, D., and Ozbay, Y. (2008). The 677 C/T MTHFR polymorphism is associated with essential hypertension, coronary artery disease, and higher homocysteine levels. Arch. Med. Res. 39 (1), 125–130. doi:10.1016/j.arcmed.2007.07.009
James, S. J., Melnyk, S., Pogribna, M., Pogribny, I. P., and Caudill, M. A. (2002). Elevation in S-adenosylhomocysteine and DNA hypomethylation: Potential epigenetic mechanism for homocysteine-related pathology. J. Nutr. 132 (8), 2361S–2366S. doi:10.1093/jn/132.8.2361S
Kato, N., Loh, M., Takeuchi, F., Verweij, N., Wang, X., Zhang, W., et al. (2015). Trans-ancestry genome-wide association study identifies 12 genetic loci influencing blood pressure and implicates a role for DNA methylation. Nat. Genet. 47 (11), 1282–1293. doi:10.1038/ng.3405
Kosmas, I. P., Tatsioni, A., and Ioannidis, J. P. (2004). Association of C677T polymorphism in the methylenetetrahydrofolate reductase gene with hypertension in pregnancy and pre-eclampsia: A meta-analysis. J. Hypertens. 22 (9), 1655–1662. doi:10.1097/00004872-200409000-00004
Kotchen, T. A. (2010). Obesity-related hypertension: Epidemiology, pathophysiology, and clinical management. Am. J. Hypertens. 23 (11), 1170–1178. doi:10.1038/ajh.2010.172
Li, D., Ahmed, M., Li, Y., Jiao, L., Chou, T. H., Wolff, R. A., et al. (2005). 5, 10-Methylenetetrahydrofolate reductase polymorphisms and the risk of pancreatic cancer. Cancer Epidemiol. Biomarkers Prev. 14 (6), 1470–1476. doi:10.1158/1055-9965.EPI-04-0894
Li, M. J., Wang, P., Liu, X., Lim, E. L., Wang, Z., Yeager, M., et al. (2012). GWASdb: A database for human genetic variants identified by genome-wide association studies. Nucleic Acids Res. 40 (D1), D1047–D1054. doi:10.1093/nar/gkr1182
Liu, X.-D., Gao, B., Sun, D., Shi, M., Ma, Y. Y., Liu, Z. R., et al. (2015). Prevalence of hyperhomocysteinaemia and some of its major determinants in shaanxi province, China: A cross-sectional study. Br. J. Nutr. 113 (4), 691–698. doi:10.1017/S0007114514004218
Ma, C., Zhao, Y., and Liu, Z. (2020). Vitamin D provides benefit based on the proinflammatory effects of homocysteine in elderly patients with type 2 diabetes mellitus. Clin. Ther. 42 (10), 2010–2020. doi:10.1016/j.clinthera.2020.08.009
Mabhida, S. E., Mashatola, L., Kaur, M., Sharma, J. R., Apalata, T., Muhamed, B., et al. (2021). Hypertension in african populations: Review and computational insights. Genes 12 (4), 532. doi:10.3390/genes12040532
Mabhida, S. E., Muhamed, B., Sharma, J. R., Apalata, T., Nomatshila, S., Mabasa, L., et al. (2022). Methylenetetrahydrofolate reductase polymorphism (rs1801133) and the risk of hypertension among african populations: A narrative synthesis of literature. Genes 13 (4), 631. doi:10.3390/genes13040631
Massie, H. R., Shumway, M. E., and Whitney, S. J. (1991). Uric acid content of Drosophila decreases with aging. Exp. Gerontol. 26 (6), 609–614. doi:10.1016/0531-5565(91)90078-z
McMahon, A., McNulty, H., Hughes, C. F., Strain, J. J., and Ward, M. (2016). Novel approaches to investigate one-carbon metabolism and related B-vitamins in blood pressure. nutrients 8 (11), 720. doi:10.3390/nu8110720
McNulty, H., Dowey, L. R. C., Strain, J. J., Dunne, A., Ward, M., Molloy, A. M., et al. (2006). Riboflavin lowers homocysteine in individuals homozygous for the MTHFR 677C→ T polymorphism. Circulation 113 (1), 74–80. doi:10.1161/CIRCULATIONAHA.105.580332
Meng, H., Huang, S., Yang, Y., He, X., Fei, L., and Xing, Y. (2021). Association between MTHFR polymorphisms and the risk of essential hypertension: An updated meta-analysis. Front. Genet. 12, 698590. doi:10.3389/fgene.2021.698590
Nassereddine, S., Kassogue, Y., Korchi, F., Habbal, R., and Nadifi, S. (2015). Association of methylenetetrahydrofolate reductase gene (C677T) with the risk of hypertension in Morocco. BMC Res. Notes 8 (1), 775. doi:10.1186/s13104-015-1772-x
Nygård, O., Vollset, S. E., Refsum, H., Stensvold, I., TverdAl, A., Nordrehaug, J. E., et al. (1995). Total plasma homocysteine and cardiovascular risk profile: The hordaland homocysteine study. Jama 274 (19), 1526–1533. doi:10.1001/jama.1995.03530190040032
Olczak, K. J., Taylor-Bateman, V., Nicholls, H. L., Traylor, M., Cabrera, C. P., and Munroe, P. B. (2021). Hypertension genetics past, present and future applications. J. Intern. Med. 290, 1130–1152. doi:10.1111/joim.13352
Padmanabhan, S., Melander, O., Hastie, C., Menni, C., Delles, C., Connell, J. M., et al. (2008). Hypertension and genome-wide association studies: Combining high fidelity phenotyping and hypercontrols. J. Hypertens. 26 (7), 1275–1281. doi:10.1097/HJH.0b013e3282ff634f
Puri, K., Suresh, K. R., Gogtay, N. J., and Thatte, U. M. (2009). Declaration of Helsinki, 2008: Implications for stakeholders in research. J. Postgrad. Med. 55 (2), 131–134. doi:10.4103/0022-3859.52846
Refsum, H., Nurk, E., Smith, A. D., Ueland, P. M., Gjesdal, C. G., Bjelland, I., et al. (2006). The hordaland homocysteine study: A community-based study of homocysteine, its determinants, and associations with disease. J. Nutr. 136 (6), 1731S–1740S. doi:10.1093/jn/136.6.1731S
Refsum, H., Smith, A. D., Ueland, P. M., Nexo, E., Clarke, R., McPartlin, J., et al. (2004). Facts and recommendations about total homocysteine determinations: An expert opinion. Clin. Chem. 50 (1), 3–32. doi:10.1373/clinchem.2003.021634
Romero-Sánchez, C., Gomez-Gutierrez, A., Gomez, P. E., Casas-Gomez, M. C., and Briceno, I. (2015). C677T (RS1801133 ) MTHFR gene polymorphism frequency in a colombian population. Colomb. Med. 46 (2), 75–79.
Rosenberg, N., Murata, M., Ikeda, Y., Opare-Sem, O., Zivelin, A., Geffen, E., et al. (2002). The frequent 5, 10-methylenetetrahydrofolate reductase C677T polymorphism is associated with a common haplotype in whites, Japanese, and Africans. Am. J. Hum. Genet. 70 (3), 758–762. doi:10.1086/338932
Selhub, J., Jacques, P. F., Wilson, P. W., Rush, D., and Rosenberg, I. H. (1993). Vitamin status and intake as primary determinants of homocysteinemia in an elderly population. Jama 270 (22), 2693–2698. doi:10.1001/jama.1993.03510220049033
Sharma, J. R., Mabhida, S. E., Myers, B., Apalata, T., Nicol, E., Benjeddou, M., et al. (2021). Prevalence of hypertension and its associated risk factors in a rural black population of Mthatha town, South Africa. Int. J. Environ. Res. Public Health 18 (3), 1215. doi:10.3390/ijerph18031215
Shi, M., Caprau, D., Romitti, P., Christensen, K., and Murray, J. C. (2003). Genotype frequencies and linkage disequilibrium in the CEPH human diversity panel for variants in folate pathway genes MTHFR, MTHFD, MTRR, RFC1, and GCP2. Birth Defects Res. A Clin. Mol. Teratol. 67 (8), 545–549. doi:10.1002/bdra.10076
Tawakol, A., GerhardM., , Wu, J. T., and Creager, M. A. (1997). Hyperhomocyst (e) inemia is associated with impaired endothelium-dependent vasodilation in humans. Circulation 95 (5), 1119–1121. doi:10.1161/01.cir.95.5.1119
Thakkinstian, A., McEvoy, M., Minelli, C., Gibson, P., Hancox, B., Duffy, D., et al. (2005). Systematic review and meta-analysis of the association between {beta}2-adrenoceptor polymorphisms and asthma: A HuGE review. Am. J. Epidemiol. 162 (3), 201–211. doi:10.1093/aje/kwi184
Tuder, R. M., Archer, S. L., Dorfmuller, P., Erzurum, S. C., Guignabert, C., Michelakis, E., et al. (2013). Relevant issues in the pathology and pathobiology of pulmonary hypertension. J. Am. Coll. Cardiol. 62 (25S), D4–D12. doi:10.1016/j.jacc.2013.10.025
Ueland, P. M., Refsum, H., Stabler, S. P., Malinow, M. R., Andersson, A., and Allen, R. H. (1993). Total homocysteine in plasma or serum: Methods and clinical applications. Clin. Chem. 39 (9), 1764–1779. doi:10.1093/clinchem/39.9.1764
Whelton, P. K., Carey, R. M., Aronow, W. S., Casey, D. E., Collins, K. J., Dennison Himmelfarb, C., et al. (20172018). 2017 ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA guideline for the prevention, detection, evaluation, and management of high blood pressure in adults: A report of the American college of cardiology/American Heart association task force on clinical practice guidelines./ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA guideline for the prevention, detection, evaluation, and management of high blood pressure in adults: A report of the American college of cardiology/American Heart association task force on clinical practice guidelines. J. Am. Coll. Cardiol. 71 (19), e127–e248. doi:10.1016/j.jacc.2017.11.006
Wu, H., Huang, Q., Yu, Z., and Zhong, Z. (2022). Association of ALDH2 rs671 and MTHFR rs1801133 polymorphisms with hypertension among Hakka people in Southern China. BMC Cardiovasc. Disord. 22 (1), 128. doi:10.1186/s12872-022-02577-x
Xi, B., Zhao, X., Chandak, G. R., Shen, Y., Cheng, H., Hou, D., et al. (2013). Influence of obesity on association between genetic variants identified by genome-wide association studies and hypertension risk in Chinese children. Am. J. Hypertens. 26 (8), 990–996. doi:10.1093/ajh/hpt046
Yamada, K., Chen, Z., Rozen, R., and Matthews, R. G. (2001). Effects of common polymorphisms on the properties of recombinant human methylenetetrahydrofolate reductase. Proc. Natl. Acad. Sci. U. S. A. 98 (26), 14853–14858. doi:10.1073/pnas.261469998
Keywords: hypertension, methylenetetrahydrofolate reductase gene, MTHFR, single-nucleotide polymorphism, Africa, genetic variation
Citation: Mabhida SE, Sharma JR, Apalata T, Masilela C, Nomatshila S, Mabasa L, Fokkens H, Benjeddou M, Muhamed B, Shabalala S and Johnson R (2022) The association of MTHFR (rs1801133) with hypertension in an indigenous south African population. Front. Genet. 13:937639. doi: 10.3389/fgene.2022.937639
Received: 06 May 2022; Accepted: 30 June 2022;
Published: 22 July 2022.
Edited by:
Ramcés Falfán-Valencia, Instituto Nacional de Enfermedades Respiratorias-México (INER), MexicoReviewed by:
Ahmet Muderrisoglu, Kırıkkale University, TurkeyAsma Amrani Midoun, Oran 1 University, Algeria
Copyright © 2022 Mabhida, Sharma, Apalata, Masilela, Nomatshila, Mabasa, Fokkens, Benjeddou, Muhamed, Shabalala and Johnson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rabia Johnson, cmFiaWEuam9obnNvbkBtcmMuYWMuemE=