
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet., 30 March 2022
Sec. Behavioral and Psychiatric Genetics
Volume 13 - 2022 | https://doi.org/10.3389/fgene.2022.837123
 Wei Zhang1
Wei Zhang1 Xiaodong Deng1,2Huan Liu3Jianlin Ke2Mingliang Xiang2
Xiaodong Deng1,2Huan Liu3Jianlin Ke2Mingliang Xiang2 Ying Ma4Lixia Zhang2Ming Yang2,5
Ying Ma4Lixia Zhang2Ming Yang2,5 Yun Liu2,6*Feijun Huang1*
Yun Liu2,6*Feijun Huang1*Objective: Amphetamine-type stimulant (ATS) and opioid dependencies are chronic inflammatory diseases with similar symptoms and common genomics. However, their coexpressive genes have not been thoroughly investigated. We aimed to identify and verify the coexpressive hub genes and pathway involved in the pathogenesis of ATS and opioid dependencies.
Methods: The microarray of ATS- and opioid-treatment mouse models was obtained from the Gene Expression Omnibus database. GEO2R and Venn diagram were performed to identify differentially expressed genes (DEGs) and coexpressive DEGs (CDEGs). Functional annotation and protein–protein interaction network detected the potential functions. The hub genes were screened using the CytoHubba and MCODE plugin with different algorithms, and further validated by receiver operating characteristic analysis in the GSE15774 database. We also validated the hub genes mRNA levels in BV2 cells using qPCR.
Result: Forty-four CDEGs were identified between ATS and opioid databases, which were prominently enriched in the PI3K/Akt signaling pathway. The top 10 hub genes were mainly enriched in apoptotic process (CD44, Dusp1, Sgk1, and Hspa1b), neuron differentiation, migration, and proliferation (Nr4a2 and Ddit4), response to external stimulation (Fos and Cdkn1a), and transcriptional regulation (Nr4a2 and Npas4). Receiver operating characteristic (ROC) analysis found that six hub genes (Fos, Dusp1, Sgk1, Ddit4, Cdkn1a, and Nr4a2) have an area under the curve (AUC) of more than 0.70 in GSE15774. The mRNA levels of Fos, Dusp1, Sgk1, Ddit4, Cdkn1a, PI3K, and Akt in BV2 cells and GSE15774 with METH and heroin treatments were higher than those of controls. However, the Nr4a2 mRNA levels increased in BV2 cells and decreased in the bioinformatic analysis.
Conclusions: The identification of hub genes was associated with ATS and opioid dependencies, which were involved in apoptosis, neuron differentiation, migration, and proliferation. The PI3K/Akt signaling pathway might play a critical role in the pathogenesis of substance dependence.
The amphetamine-type stimulants (ATS) and opioids are the major powerful and highly addictive drugs worldwide and nationwide, including 3,4-methylenedioxy-methamphetamine (MDMA), methamphetamine (METH), heroin, morphine, and opium (https://www.unodc.org/unodc/en/data-and analysis/wdr2021.html). Ellis et al. reported that the ATS- and opioid-exposure patients increased from 18.8% in 2011 to 34.2% in 2017 (Ellis et al., 2018). An acute dose of drug treatment led to neuronal death in the frontal cortex, striatum, and substantia nigra in animals, whereas repeated drug administration/chronic exposure led to neuronal loss in the hippocampus, frontal cortex, and striatum (Sabrini et al., 2020). Chronic and repeated administration could also cause tolerance and dependence, recurrent encephalopathy (e.g., neuronal degeneration and/or damage, neuroinflammation), and epigenetic modifications (e.g., histone modifications, DNA methylation, and noncoding RNAs) (Feng and Nestler 2013). Polysubstance dependence, especially opioid and ATS dependence, is evolving into an epidemic drug use pattern worldwide (Jones et al., 2019; Cicero et al., 2020). There are similar target genes (e.g., FAAH, BDNF, DRD4, and OPRM1) and mechanisms (e.g., DNA damage, apoptosis, neurotoxicity, neuroinflammation, and epigenetic modifications) between ATS and opioid dependence (Lopez-Moreno et al., 2012; Feng and Nestler 2013; Górska et al., 2014; Silva et al., 2014; Moratalla et al., 2017; Doris et al., 2019; Zhang et al., 2020; Lopez-Leon et al., 2021). However, the potential coexpressive genes and pathogenesis of ATS and opioid dependence have not been thoroughly investigated. The coexpressive genes were widely applied to discern candidate biomarkers and therapeutic targets for Alzheimer’s disease, schizophrenia, immune-mediated inflammatory diseases, and cancer (Zhu et al., 2016; van Dam et al., 2017; Kakati et al., 2019; Chen Y. et al., 2020; Wang et al., 2021). We hypothesized that coexpressive target genes and signaling pathways may provide further insight into the common pathophysiological process of ATS and opioid dependencies.
Following the development of bioinformatic technology, RNA sequencing (RNAseq) and high-throughput microarray had been widely used to explore and detect the biomarkers, functional annotation, and molecular mechanism of a variety of diseases in vivo and in vitro in the past decades (Kang et al., 2021; Sun et al., 2021; Xu et al., 2021). All the databases could be downloaded and reanalyzed freely. As the most economic and effective technique, bioinformatic analysis was performed to identify candidate hub genes. Previous microarrays mainly focused on a single drug, such as nicotine (Jung et al., 2016), alcohol (Smith et al., 2020), morphine (Skupio et al., 2017), heroin (Kuntz-Melcavage et al., 2009), MDMA (Eun et al., 2010), METH (Palmer et al., 2005), or cocaine (Renthal et al., 2009). They mainly focused on comparatively small samples, a single timepoint, or individual reward brain regions (Piechota et al., 2010; Piechota et al., 2012). Therefore, we conducted the integrated bioinformatic analysis with all conditions and timepoints in all GEO databases. Receiver operating characteristic (ROC) analysis and quantitative real-time PCR (qPCR) were performed to validate the hub genes and key factors of the signaling pathway between ATS and opioid treatments.
The study was approved by the Ethics Committee of Sichuan University. The microarrays were systematically extracted from the Gene Expression Omnibus (GEO) databases (http://www.ncbi.nlm.nih.gov/geo/) with several keywords: “heroin” or “morphine” or “opioids” or “opium,” “Amphetamine” or “methylamphetamine” or “METH” or “MA” or “3,4-methylenedioxymethamphetamine” (Barrett et al., 2010). The inclusion criteria include the following: 1) Databases were restricted in “expression profiling by microarrays.” 2) The organism was limited to “Mus musculus” brain. 3) The original microarrays should contain cases and controls. The exclusion criteria were the following: 1) The mice were intervened by other drugs. 2) Transgenic mice. 3) Other tissues. In addition, for overlapping databases, only the maximum samples were included. The workflow of the study is shown in Figure 1.
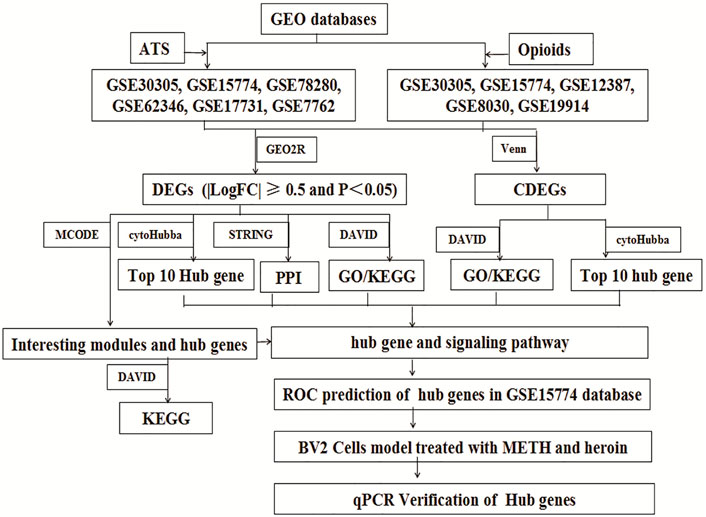
FIGURE 1. Flow diagram of the study design.
GEO2R (https://www.ncbi.nlm.nih.gov/geo/geo2r) was utilized to screen and identify the DEGs from all the eligible databases, respectively (Davis and Meltzer 2007). If a gene had multiple probes on the same chip, the average value of all probes would be taken as the gene expression value. If the genes lacked probes, they were removed. The DEGs were defined with |log2 fold change (FC)| ≥ 0.5 and p < 0.05 in the study. The coexpressive DEGs (CDEGs) were the overlapped DEGs of ATS and opioid treatment, which were screened by a Venn diagram (version 2.1.0, https://bioinfogp.cnb.csic.es/tools/venny/index.html).
Gene Ontology (GO) (Ashburner et al., 2000) and Kyoto Encyclopedia of Genes and Genomes (KEGG) (Kanehisa et al., 2019) pathway enrichment analyses of DEGs and CDEGs were applied to explore the biological function and signaling pathway by the online tool Database for Annotation, Visualization, and Integrated Discovery (DAVID, version 6.8 https://david.ncifcrf.gov/). A term with p < 0.05 was identified as the critical threshold for significant enrichment.
Protein–protein interaction (PPI) network revealed the specific and nonspecific interactions of proteins by the Search Tool for the Retrieval of Interacting Genes database (STRING, https://string-db.org/) (Szklarczyk et al., 2019). The minimum required interaction score of more than 0.4 and p-value less than 0.05 was statistically significant (Fang et al., 2021; Zhou et al., 2021). The Cytoscape software (version 3.6.1) was applied to visualize the PPI network (Shannon et al., 2003). Besides, the Molecular Complex Detection (MCODE) plugin in the Cytoscape software was used to identify the interesting modules (selection criteria: degree cutoff = 2, K-core = 2, and node score cutoff = 0.2) (Bader and Hogue, 2003; Bandettini et al., 2012; Su et al., 2021).
Four most effective algorithms in the CytoHubba plugin in Cytoscape software were performed to identify hub genes, including the maximal clique centrality (MCC), density of maximum neighborhood component (DMNC), maximum neighborhood component (MNC), and degree. A high score indicated that the target was closely related to the disease and was a possible hub gene. The MCODE plugin in the Cytoscape software were also used to identify the hub genes by MCODE score (top three connective genes in interesting modules) (Chin et al., 2014; Nangraj et al., 2020; Qi et al., 2021).
The TF-targeted top 20 hub genes of the PPI network were predicted with the iRegulon plugin in the Cytoscape software, which integrated information from the larger modify and track collections. We obtained the data predicted by the track discovery of existing regulatory databases, which included data validated by ChIP-seq, DHS-seq, or FAIRE-seq. TF-target pairs with normalized enrichment score (NES) >4 were chosen (Janky et al., 2014).
Immortalized mouse microglia cells (BV2) were purchased from Procell (Wuhan, China) and incubated with Dulbecco’s modified Eagle’s medium/nutrient mixture F-12 (DMEM/F12) (Gibco, USA, Catalog: 11320033) supplemented with 10% fetal bovine serum (FBS) (BI, USA, Catalog: 04-002-1A), and 1% penicillin and streptomycin (Solarbio, Beijing, China, catalog: P1400). The BV2 cells were seeded in six-well plates at a density of 2.5 × 105 cells/well with 2 ml of complete medium cultured at 37°C in a humidified 5% CO2 atmosphere until the cell confluence reached 60%–70%. Then BV2 cells were treated with different concentrations of METH or heroin for different periods to mimic the drug injury in vitro, respectively. In the study, the relative mRNA levels of CD11b (a marker of activated microglia) substantially increased with METH above 500 μM and heroin above 100 μM, and with a peak response at 2,000 μM of METH and 400 μM of heroin, respectively (Figure 2A). Considering the cell activity and damage (Figure 2B), the BV2 cells were treated with 1,000 μM of METH and 200 μM of heroin after 24 h, respectively. The concentration of drugs did not induce significant cell damage, even in a time course experiment (Figures 2C, D). The concentrations were consistent with previous studies (Lai et al., 2011; Park et al., 2017; Chen X. et al., 2020). Under this condition, the relative mRNA levels of CD11b, TNF-a, and IL-6 were considerably increased (Figures 2E–G). It indicated that the METH and heroin effectively established inflammatory models in vitro.
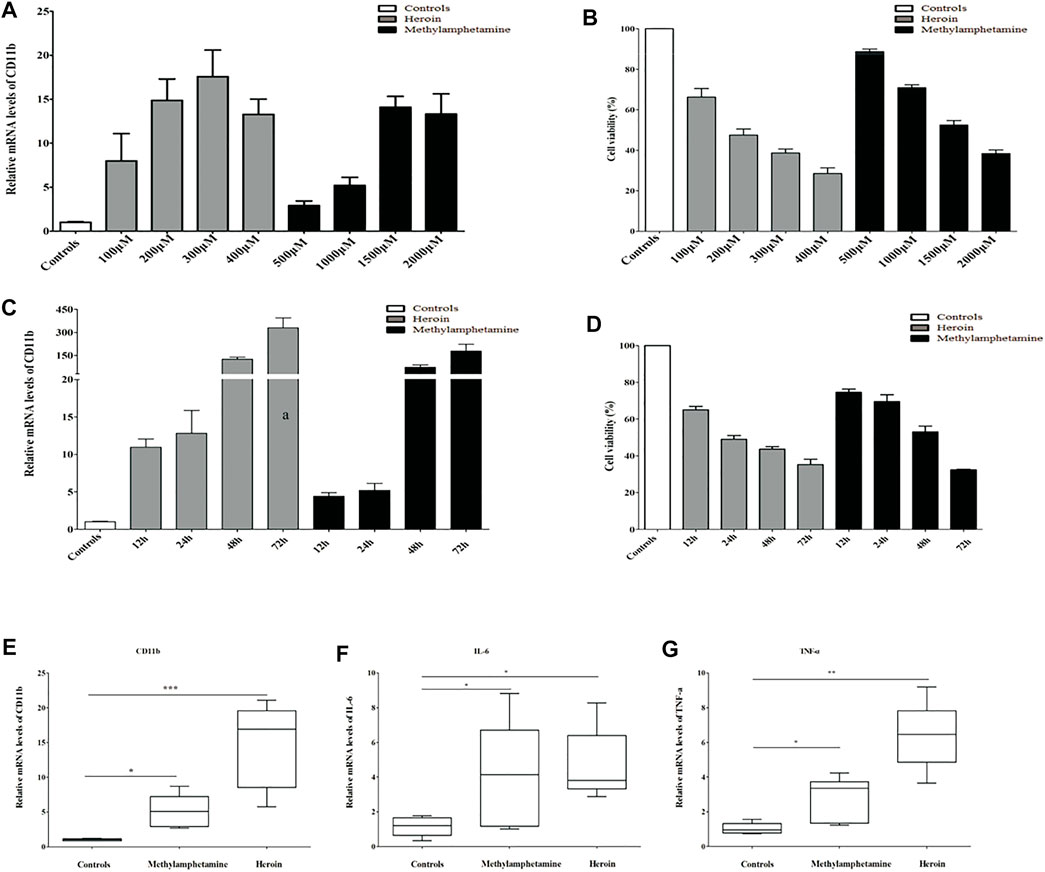
FIGURE 2. METH) and heroin increased the mRNA levels of CD11b and inflammatory cytokine in immortalized mouse microglia cells (BV2) cells. The relative mRNA levels of CD11b by qPCR and cell viability by CCK-8 assay at varied concentrations (A, B) and various timepoints (C, D) between METH and heroin, respectively. The relative mRNA levels of CD11b, IL-6, and TNF-α treated with 1,000 μM METH and 200 μM heroin after 24 h (E–G), respectively. (*p < 0.05, **p < 0.01, ***p < 0.001 compared with the controls).
The qPCR was performed to examine the difference in transcriptional levels of the hub genes and key factors of signaling pathway in BV2 cells after heroin and METH treatment. The primers and amplicon sizes of hub genes are shown in Supplementary Table S1. The qPCR was performed according to the instructions of TaKara TB Green™ Premix Ex Taq™ II by Bio-Rad CFX96TM real-time system (Foster City, CA, USA). All samples were performed in triplicate. The relative mRNA levels of hub genes were calculated by comparing the average of each target gene with the reference glyceraldehyde-3-phosphate dehydrogenase (GAPDH) in the same sample with the 2−ΔΔCt method.
The relative mRNA levels were analyzed with SPSS for Windows software package version 13.0 (SPSS, Inc., Chicago, IL, USA). All data were presented as the mean ± standard deviation (SD) for continuous variables. The Student’s t-test was used if the data have a normal distribution. Otherwise, the Kruskal–Wallis test was used. A two-sided p-value < 0.05 was taken as the level for statistical significance. ROC analysis was performed to explore the predictive accuracy of hub genes in the GSE15774 database. AUC was used to evaluate the sensitivity and specificity of each gene. The genes with an AUC of more than 0.7 and p-value of less than 0.05 were used to evaluate the predictive accuracy of hub genes (Fang et al., 2021; Kang et al., 2021).
According to the inclusion criteria, GSE30305 (Piechota et al., 2012), GSE15774 (Piechota et al., 2010), GSE78280 (Skupio et al., 2017), GSE62346 (Mcadams et al., 2015), GSE17731 (Sanchis-Segura et al., 2009), GSE7762 (Korostynski et al., 2007), GSE12387 (Palmer et al., 2005), GSE8030 (Chin et al., 2008), and GSE19914 (Eun et al., 2010) were included in the study (Table 1). For opioid treatment, 100 opioid samples and 107 controls from the six opioid treatment databases were further analyzed, including GSE30305, GSE15774, GSE78280, GSE62346, GSE17731, and GSE7762. There were 68 ATS samples and 79 controls in five ATS-treatment databases, which included GSE30305, GSE15774, GSE12387, GSE8030, and GSE19914. Additionally, GSE30305 and GSE15774 databases simultaneously contained opioid and ATS treatments. The main administration methods were intraperitoneal (ip) injection (five studies of opioids and four studies of ATS). Single-dose treatment was used in six databases, and multidose treatment was used in three databases (dose range was 10–40 mg/kg for opioids and 2–20 mg/kg for ATS). The duration of administration was 3 to 14 days for all opioid models and three ATS models. All the tissues were involved in brain reward regions, including the striatum, hippocampal, cerebral cortex, and nucleus acumbens (NAc). As shown in Supplementary Figures S1 and S2, the midline of ATS and opioid treatments were matched by boxplot analysis, and the gene expression profiles were comfortable for further study.
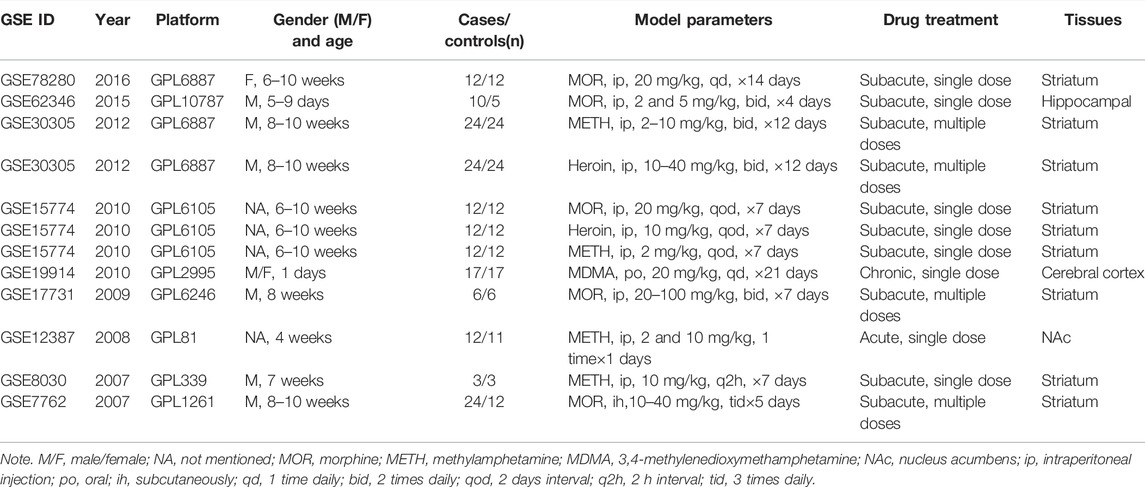
TABLE 1. Basic information of the selected databases in the study.
The number of DEGs in each database by GEO2R is shown in Table 2. The number of DEGs obtained in each microarray varied widely, ranging from 0 to more than 700. There were only two DEGs for opioid treatment, but no DEGs for ATS treatment in the GSE30305 database. A total of 366 DEGs were identified from opioid treatment databases, of which 157 genes were upregulated and 209 genes were downregulated. 1,183 DEGs including 873 upregulated and 310 downregulated genes were identified from ATS treatment databases. The CDEGs between opioid and ATS treatments were identified from the all databases by Venn diagram (Figure 3). There were 44 CDEGs (17 upregulated, 8 downregulated, and 19 discordant) between opioid and ATS treatments including Fos, Ddit4, Sgk1, Dusp1, Nr4a2, Cdkn1a, Hspa1b, CD44, Gngt1, Syk, Tsc22d3, Plin4, Pnlip, Rhpn2, Cldn1, Ace2, Fzd1, Ssbp1, Ppih, Prlr, Rasd1, Txnip, Npas4, Cfap97, Ttll1, Nkx2-4, Tmem252, Foxh1, Cpsf3, Zfp706, Ap1s2, Kcnj2, Gbp3, Nhej1, Ascl1, Taf1d, Gsap, Ern1, Spam1, Nkx1-2, Ttr, Zfp189, Arc, and S100a5.
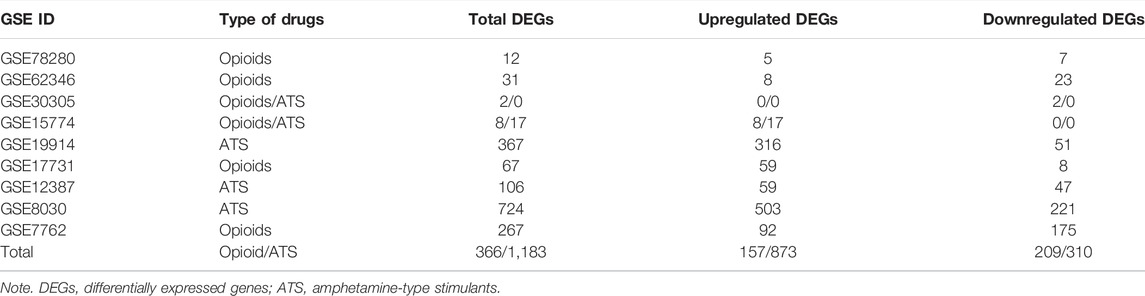
TABLE 2. The number of differentially expressed genes (DEGs) in each database by GEO2R.
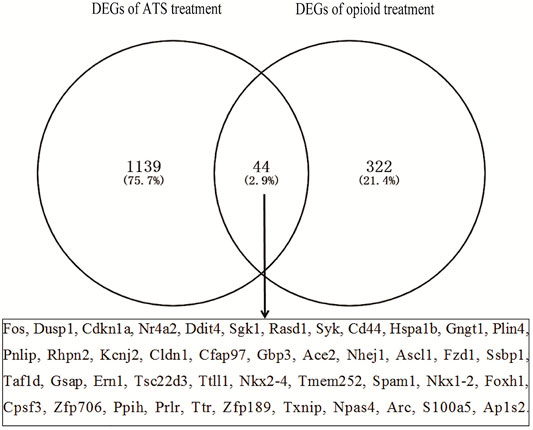
FIGURE 3. The coexpressive differentially expressed genes (CDEGs) in ATS- and opioid-treatment databases by Venn diagram.
GO biological analysis is demonstrated in Figures 4A, B. For DEGs of opioid treatment, there were about 57 terms in the biological process (BP) category, 26 terms in the molecular function (MF) category, and 11 terms in the cellular component (CC) category. Similarly, DEGs of ATS treatment were associated with 280 terms in the BP category, 96 terms in the MF category, and 92 terms in the CC category. KEGG pathway analysis indicated that the DEGs of opioid and ATS treatments were enriched in 18 and 70 signaling pathways, respectively (Figures 4C, D). Functional annotation of CDEGs indicated that 20 enriched GO terms and three KEGG pathways were detected between opioid and ATS treatments (Figure 4E). GO terms were mainly enriched in the BP category including cellular apoptotic process (GO:0043066), regulation of transcription, DNA templated (GO:0006355), neuron migration (GO:0001764) and differentiation (GO:0030182), mitotic cell cycle arrest (GO:0071850), transcriptional regulation (GO:0000122), DNA templated (GO:0045893), response to extracellular stimulus (GO:0031668), drug (GO:0042493), and corticosterone (GO:0051412) (Supplementary Table S3). KEGG pathways were mainly enriched in the PI3K/Akt signaling pathway (mmu04151), circadian entrainment (mmu04713), and cholinergic synapse (mmu04725) (Figure 4E and Supplementary Table S2).
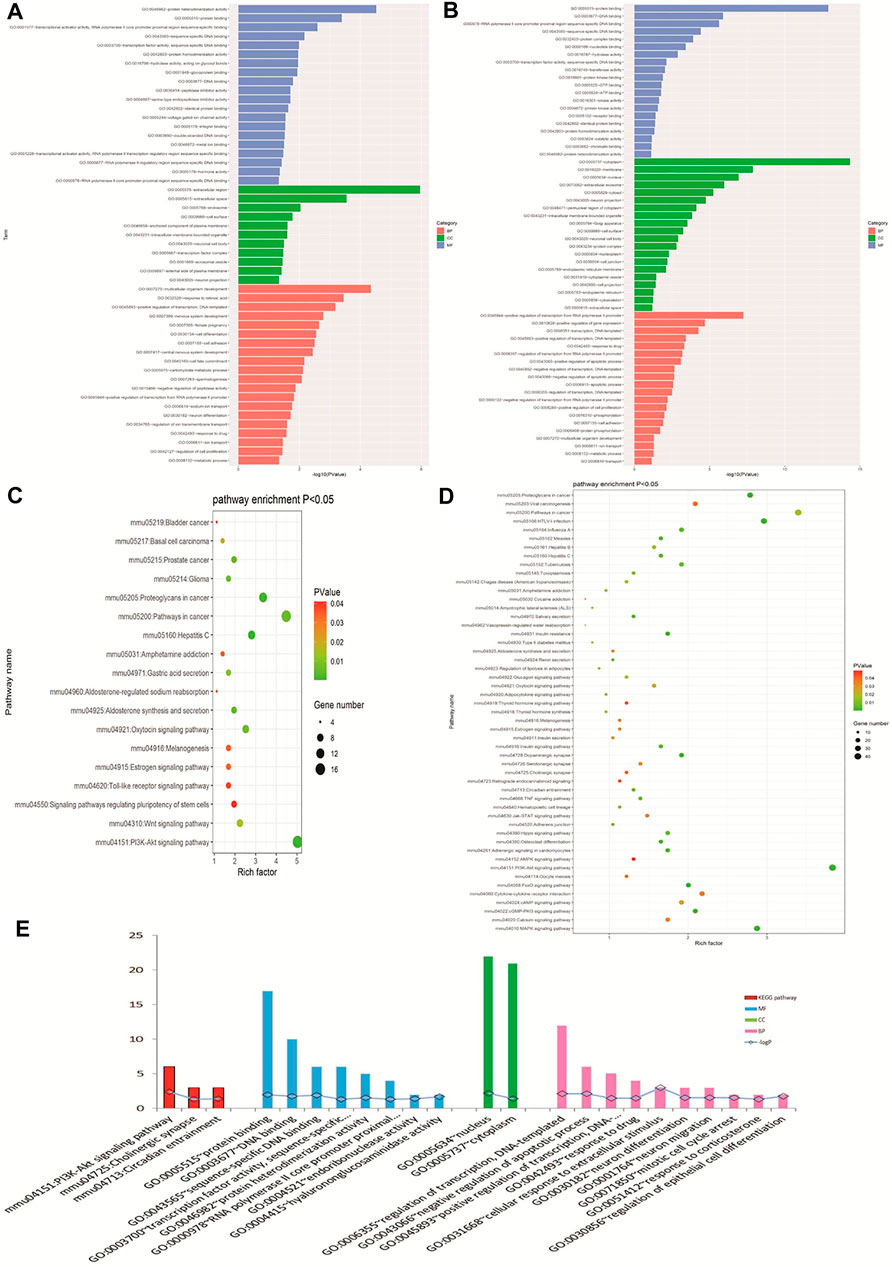
FIGURE 4. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses. GO analysis of DEGs with opioid (A) and ATS treatments (B). KEGG pathway enrichment analysis for DEGs in opioid (C) and ATS treatments (D). GO and KEGG pathway analysis for CDEGs (E).
According to the STRING online database, 342 nodes and 568 edges were detected in the 366 DEGs of opioid treatment, and the average node degree was 3.32. Similarly, 1,128 nodes and 5,888 edges were screened in the 1,183 DEGs of ATS treatment, and the average node degree was 10.4. A total of 44 nodes and 29 edges were screened in the 44 CDEGs of opioid and ATS treatments, and the average node degree was 1.32 (Figure 5A).
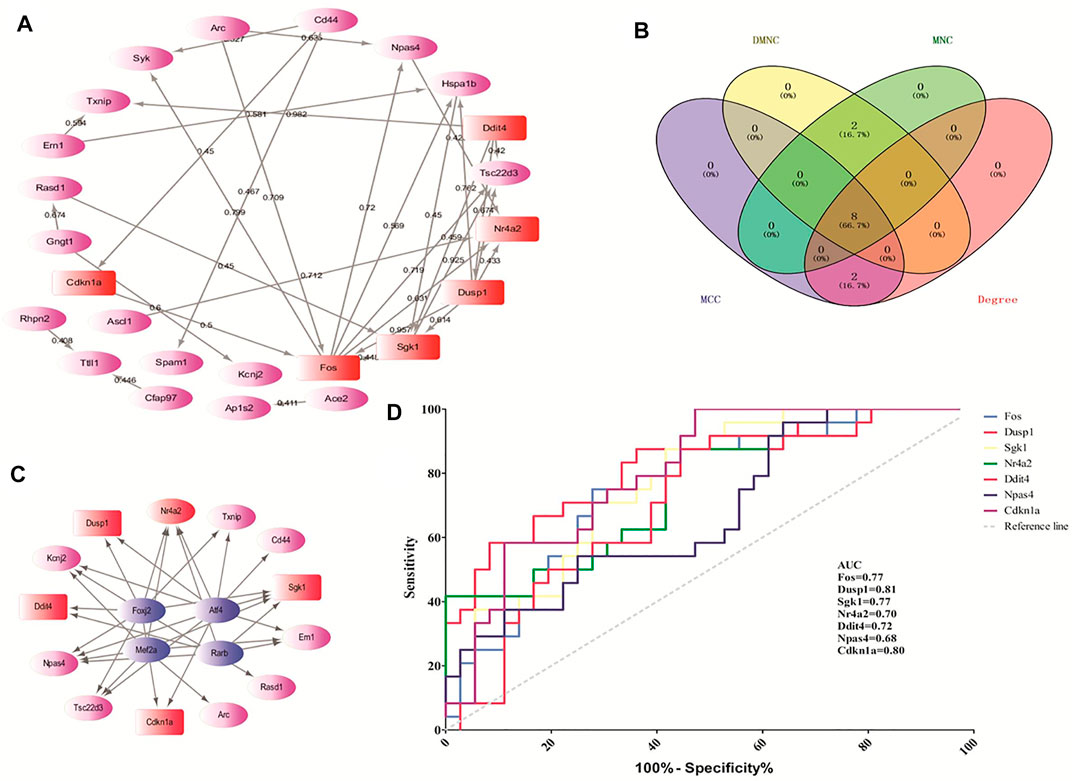
FIGURE 5. The hub genes screened by the CytoHubba plugin and predicted by receiver operating characteristic (ROC) curve. The protein–protein interaction (PPI) network of CDEGs (A). Venn diagram of the top 10 hub genes by MCC, DMNC, MNC, and degree algorithms (B). The PPI network of TF-target genes by the iRegulon plugin (C). The ROC curve-predicted hub genes in GSE15774 (D). Red indicates the hub genes. Pink indicates the other DEGs. Blue indicates TF-target genes.
We screened 29 modules in ATS treatment, and 7 modules in opioid treatment with the MCODE plugin, respectively. In the present study, a MCODE score greater than 4.0 was considered as an interesting module (Qi et al., 2021). Finally, eight interesting modules for ATS treatment and two interesting modules for opioid treatment were selected, respectively (Table 3 and Supplementary Figure S3). Pathway enrichment analysis of the interesting module demonstrated that each module was functionally correlated (Supplementary Table S3).
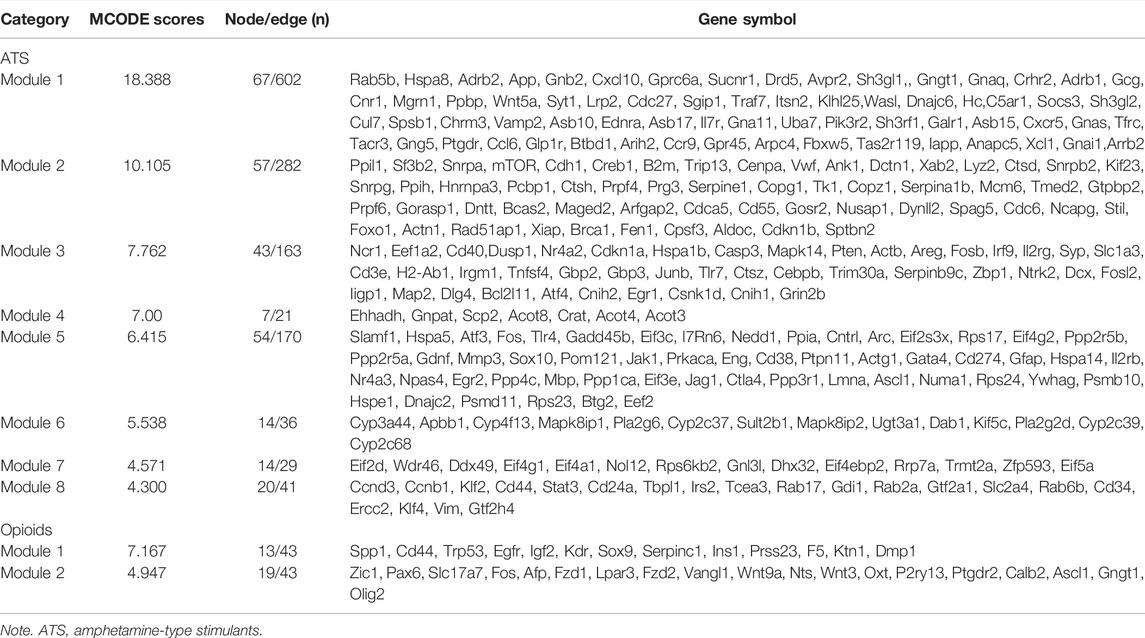
TABLE 3. The information of interesting modules in ATS- and opioid-treatment databases, ranked in descending order of MCODE score.
According to the CytoHubba plugin with MCC algorithm in Cytoscape, the hub genes were significantly different between opioid- and ATS-treatment DEGs (Supplementary Table S4). The Fos, Dusp1, Sgk1, Nr4a2, Ddit4, Hspa1b, Npas4, Cdkn1a, Cd44, and Rasd1 were identified as hub genes (Table 4). Similar results were obtained by DMNC, MNC, and degree algorithms (Table 4 and Figure 5B).
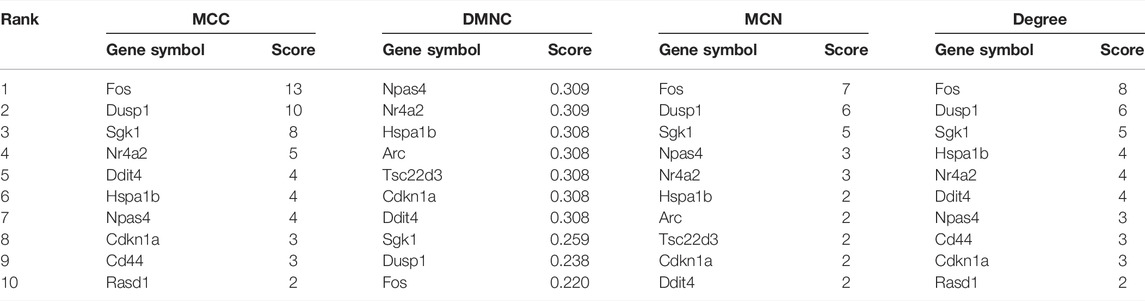
TABLE 4. Maximal clique centrality (MCC), density of maximum neighborhood component (DMNC), maximum neighborhood component (MNC), and degree score of the top 10 hub genes by the CytoHubba plugin in Cytoscape Software.
Based on the MCODE score, we selected the top three targets in each interesting module as the hub genes. In opioid treatment, SPP1, CD44, Trp53, (module 1), Zic1, Pax6, and Slc17a7 (module 2) were screened as hub genes. Similarly, Rab5b, Hspa8, Adrb2 (module 1), Ppil1, Sf3b2, Snrpa (module 2), Ncr1, Eef1a2, CD40 (module 3), Ehhadh, Gnpat, Scp2 (modul4), Slafm1, Hspa5, Atf3 (module 5), Cyp3a44, Apbb1, Cyp4f13 (module 6), Eif2d, Wdr46, Ddx49 (module 7), Ccnd3, Ccnb1, and Klf2 (module 8) were hub genes for ATS treatment (Supplementary Figure S3). Furthermore, most hub genes screened by the CytoHubba plugin were also enriched in the interesting modules.
We found four TFs (Foxj2, Alf4, Rarb, and Mef2a) with NES > 4.5, which may regulate the expression of the top 20 hub genes. These TF-target genes could regulate the expression of Dusp1, Sgk1, Ddit4, Nr4a2, and Cdkn1a (Figure 5C).
In order to verify the predictive accuracy of hub genes, ROC curve analysis was performed in GSE15774. The relative mRNA levels of hub genes in GSE15774 are shown in Table 5. Among the hub genes of ATS- and opioid-treatment databases, the Fos, Dusp1, Sgk1, Ddit4, Npas4, and Cdkn1a mRNA levels were increased, while the Nr4a2 mRNA levels were decreased. Rasd1 mRNA levels in opioid treatment were different from controls, but not in ATS treatment. There were no significant difference in CD44 and Hspa1b mRNA levels in neither ATS nor opioid treatment than those of controls (Table 5). ROC analysis indicated that AUC of Fos, Dusp1, Sgk1, Nr4a2, Ddit4, and Cdkn1a was more than 0.7 and p < 0.05 in the GSE15774 database (Figure 5D). It might be potential hub genes for ATS and opioid dependence and needed to be verified by qPCR in BV2 cells.
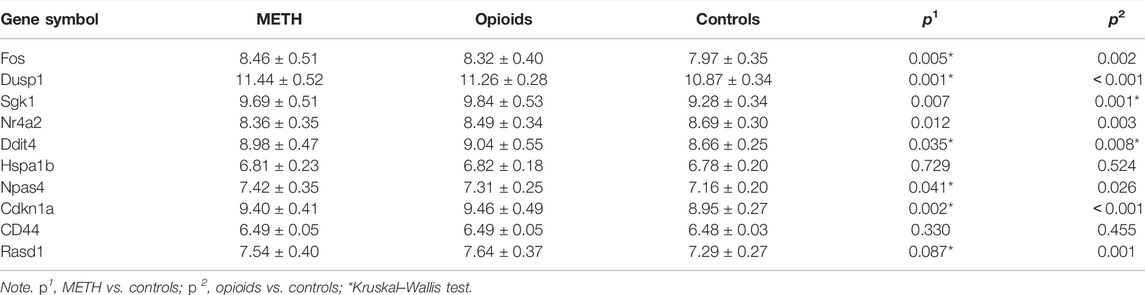
TABLE 5. The relative mRNA levels of hub genes in GSE15774 database (mean ± SD).
The relative mRNA levels of Fos, Dusp1, Sgk1, Ddit4, Cdkn1a, PI3K, and Akt in BV2 cells in both METH and heroin treatments significantly increased compared with those of controls (p < 0.05), respectively (Figure 6 and Supplementary Table S5). The results were consistent with those of the bioinformatic analysis and GSE15774 database. However, the Nr4a2 mRNA level increased in BV2 cells, while it decreased in the bioinformatic analysis and GSE15774 database.
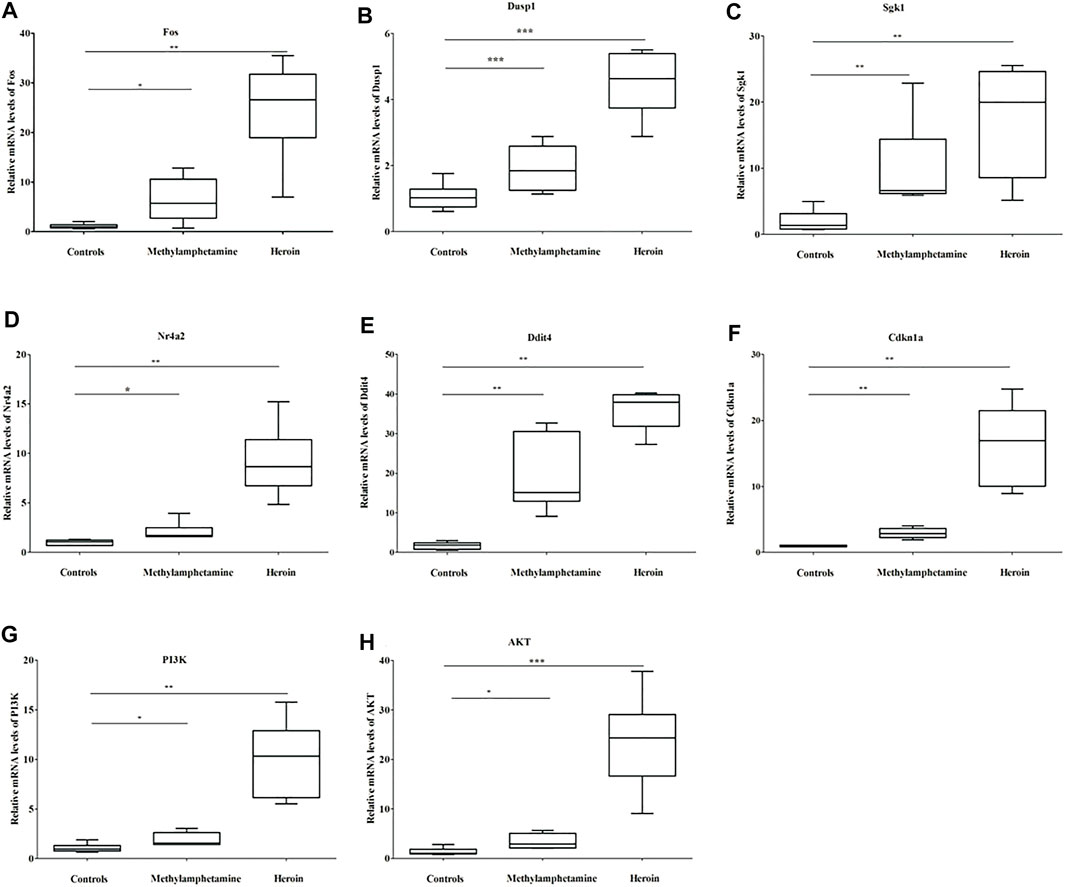
FIGURE 6. The relative mRNA levels of Fos (A), Dusp1 (B), Sgk1 (C), Nr4a2 (D), Ddit4 (E), Cdkn1a (F), PI3K (G), and Akt (H) in BV2 cells treated with 1,000 μM METH and 200 μM heroin after 24 h, respectively (*p < 0.05, **p < 0.01, ***p < 0.001 compared with the controls).
The bioinformatic analysis was conducted to better understand the hub genes and molecular mechanisms of substance dependence. In the study, 44 CDEGs were identified between ATS and opioid databases. The top 10 hub genes were mainly enriched in apoptotic process (CD44, Dusp1, Sgk1, and Hspa1b), neuron differentiation, migration, and proliferation (Nr4a2 and Ddit4), response to external stimulation (Fos and Cdkn1a), and transcriptional regulation (Nr4a2 and Npas4). They were also prominently enriched in the PI3K/Akt signaling pathway. The relative mRNA levels of the aforementioned hub genes were significantly different between cases and controls in the GES15774 database. ROC analysis found that the AUC scores of hub genes (Fos, Dusp1, Sgk1, Nr4a2, Ddit4, and Cdkn1a) were more than 0.70 in GSE15774. It indicated that the hub genes could be accurately predicted. The mRNA levels of Fos, Dusp1, Sgk1, Ddit4, Cdkn1a, PI3K, and AKT were significantly increased in BV2 cells with METH and heroin treatments, respectively. However, the Nr4a2 mRNA levels increased in BV2 cells, and decreased in bioinformatic analysis. These results were consistent with those of the bioinformatic analysis and GSE15774 database.
In the bioinformatic analysis, nine databases (six studies of opioid treatment, and four studies of ATS treatment) were downloaded from the GEO database, and GSE30305 and GSE15774 databases simultaneously contained opioid and ATS treatments. The number of DEGs obtained in each microarray ranged from 0 to more than 700. It might be caused by the differences in detection platform and model parameters including drug types, doses, frequency, and duration of drug administration. GEO2R analysis indicated that a total of 366 DEGs including 157 upregulated and 209 downregulated DEGs in opioid treatment, and 1,183 DEGs including 873 upregulated and 310 downregulated DEGs in ATS treatment were identified, respectively. BP was the main enriched term of those DEGs, including transcriptional regulation, apoptotic process, phosphorylation, cell proliferation and adhesion, and nervous system development in ATS treatment, and nervous system development, transcriptional regulation, cell differentiation and adhesion, and ion transport in opioid treatment. It indicated that the common biological processes involved in ATS and opioid dependencies were apoptosis, nervous system development, cell differentiation, and proliferation (Piechota et al., 2010; Piechota et al., 2012). In the KEGG pathways, the PI3K/Akt signaling pathway was the prominently enriched pathway in either opioid or ATS treatment. Furthermore, there were 44 CDEGs between ATS- and opioid-treatment databases. The results illustrated that ATS and opioid dependencies shared common target genes and molecular mechanism, which were consistent with the previous studies and whole-genome microarray profiling (Piechota et al., 2012; Korostynski et al., 2013; Lopez-Leon et al., 2021).
The PI3K/Akt signaling pathway was the main enriched signaling pathway between ATS and opioid treatments, and was involved in regulating the process of cell survival, growth, proliferation, angiogenesis, transcription, translation, metabolism, and apoptosis (Gaesser and Fyffe-Maricich 2016; Sinha et al., 2018; Ediriweera et al., 2019). Some studies indicated that the PI3K/Akt signaling pathway played an essential role in neuronal survival (Dudek et al., 1997) and neurodegeneration (Wu et al., 2010; Chen et al., 2012). Qiao et al. reported that drugs (e.g., alcohol, heroin, morphine, and METH) could activate the PI3K/Akt signaling pathway in the cortex, and contribute to addiction (Qiao et al., 2018; Li et al., 2020; Meng et al., 2020; Zhu et al., 2021). The Akt phosphorylation in the NAc was related to heroin-seeking behavior (Zhu et al., 2021). Furthermore, we detected that the PI3K and Akt mRNA levels in BV2 cells treated with METH and heroin significantly increased than those of controls. It showed that the PI3K/Akt signaling pathway was the common signaling pathway and played vital roles in ATS and opioid treatments. Interestingly, Sgk1, Ddit4, and Cdkn1a (hub genes) were also enriched in the PI3K/Akt signaling pathway. Sgk1, Ddit4, and Cdkn1a were located in 6q23.2, 10q24.33, and 6p21.2 and encoded at 49 kD with 431 amino acids, 25 kD with 232 amino acids, 18.1 kD with 164 amino acids, respectively (Demetrick et al., 1995; Kobayashi et al., 1999; Ellisen et al., 2002). The hub genes were widely distributed in different tissues, and were involved in multiple physiological functions and pathophysiological conditions, such as hypoxia, ionizing radiation, heat shock, oxidative stress, hormone release, cell proliferation and apoptosis, autophagy, fibrosis disease, ischemia sequelae, neuronal survival, neuroexcitability, and neurodegeneration (Ellisen et al., 2002; Shoshani et al., 2002; Malagelada et al., 2006; Anacker et al., 2013; Canal et al., 2014; Wolff et al., 2014; Jin et al., 2019; Fattahi et al., 2021). The Sgk1, Ddit4, and Cdkn1a mRNA levels in the databases significantly increased, except Sgk1 in GSE78280. The GSE78280 database was the only female model, the difference might be associated with gender (Dumeige et al., 2017). Further study is needed to reveal the exact reason. The relative mRNA levels of Sgk1, Ddit4, and Cdkn1a markedly increased in BV2 cells with METH and heroin treatments than those of controls. The results were consistent with the bioinformatic analysis and other studies on liver injury, traumatic brain injury, anterior cruciate ligament transection, and lung injury (Lang et al., 2008; Chen et al., 2016; Steiner et al., 2016; Wong et al., 2016; Li et al., 2017; Faust et al., 2020; Blazquez-Prieto et al., 2021; Guillot et al., 2021). The data demonstrated that Sgk1, Ddit4, and Cdkn1a might play vital roles in ATS and opioid dependencies (Piechota et al., 2010). Furthermore, function annotation demonstrated that Sgk1, Ddit4, and Cdkn1a were mainly involved in apoptotic process and cell proliferation. It suggested that Sgk1, Ddit4, and Cdkn1a played key roles in ATS and opioid dependencies through neuronal autophagy and apoptosis, and may be potential target genes of drug dependence. Combined with the aforementioned results, we speculated that the PI3K/Akt signaling pathway and related genes could regulate the pathogenesis of drug dependence though autophagy and apoptosis.
With Fos and Dusp1, as the immediate early genes (Wojcieszak et al., 2019), the relative mRNA levels increased in opioid- and most ATS-treatment databases. The results were in line with other studies [e.g., heroin, cocaine, △9-tetrahydrocannabinol (THC), and amphetamine] in different brain reward regions (e.g., NAc, neocortex, posterior caudate, and striatum) (Singh et al., 2005; Mattson et al., 2007; Paolone et al., 2007; Celentano et al., 2009; Fanous et al., 2012; Rubio et al., 2015; Wojcieszak et al., 2019). Fos and Dusp1 locus resided on 14q21–q31 and 5q35.1, encoded approximately 380 and 367 amino acids, respectively (Visvader et al., 1988). Fos and Dusp1 played vital roles in regulating signaling transduction, cell proliferation, and differentiation (Duric et al., 2010). Some studies demonstrated that Fos and Dusp1 were not only deemed as biomarkers of neuronal activity (Hughes and Dragunow 1995) but also a vital initial step in regulating neuroplasticity caused by drugs (Harlan and Garcia 1998). Kuroda et al. even demonstrated that Fos played a neuroprotective role in the progress of neurotoxicity caused by methamphetamine (Kuroda et al., 2010). In the present study, the relative mRNA levels of Fos and Dusp1 also increased in BV2 cells treated with METH and heroin, which was consistent with the study of Takaki et al. in acute METH administration (Takaki et al., 2001; Wojcieszak et al., 2019). However, Beauvais et al. reported that repeated injection of METH did not alter the Fos mRNA levels (Beauvais et al., 2010). It may be that Fos and Dusp1 were the immediate early genes. Interestingly, the mRNA levels of Fos and Dusp1 decreased in the GSE19914. It might be associated with drug species. GSE19914 mainly focused on MDMA and newborns (one pup), while other databases were prominent METH and young/adult mice.
Nr4a2 was predominantly expressed in the midbrain, substantia nigra, and ventral tegmental (Zetterstrom et al., 1997; Torii et al., 1999), which played an essential role in protecting dopaminergic neurons and limiting proinflammatory neurotoxin. Nr4a2 (−/−) mice failed to synthesize brain dopaminergic neurons, which resulted in mice hypoactivity, and died rapidly after birth (Saijo et al., 2009). In the study, Nr4a2 was mainly involved in neuron differentiation and migration, response to external stimulation, and transcriptional regulation (Horvath et al., 2007). Combined with previous studies, we speculated that the Nr4a2 was a potential target gene for substance dependence. However, the Nr4a2 mRNA levels were increased in BV2 cells with METH and heroin treatments, while they decreased in the bioinformatic analysis and GSE15774 database. The qPCR was performed only on BV2 cells, and microarrays were from the C57BL/6 brain musculus, which contained neurons, astrocytes, microglia, and neural intermediants.
There were some limitations in the present study. First, due to database limitations, the microarrays with different model parameters were all included. It might result in biases of selected hub genes. So, the hub genes might not really be involved in the pathogenesis of substance dependence. Further studies are needed to verify the pathophysiological mechanism of these hub genes and pathways that participated in the substance-dependent animal model and humans. Second, the study was based on animal databases and BV2 cells. The results might not be suitable to extrapolate to substance dependence of other animal models and humans. Despite the interspecies differences, animal studies contributed significantly to addiction research and are still of great assistance for future research with a more relevant model of compulsive drug use in humans. Third, the mechanisms of ATS and opioid dependencies might involve both genetic and epigenetic aspects. Epigenetic regulation consequences were beyond alterations in steady-state levels of expressed RNAs, which were far beyond the coverage of microarrays. The next-generation sequencing technologies (e.g., ChIP-seq and RNA-seq) could provide additional information. That work would be the focus of our future studies.
In conclusion, Fos, Dusp1, Sgk1, Nr4a2, Ddit4, and Cdkn1a hub genes were associated with ATS and opioid dependencies. Functional annotation and KEGG pathway analysis suggested that apoptosis, neuron differentiation, migration and proliferation, and the PI3K/Akt signaling pathway might play a critical role in pathogenesis of drug dependence. The findings may be helpful for better understanding of the shared pathogenesis and molecular mechanisms of ATS and opioid dependencies, and consequently detecting the new detection and potential therapeutic targets for drug dependence.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
WZ contributed to conceiving and designing the experiments, analyzing the data, and drafting the manuscript. JK, MX, and XD took responsibility for the data collected and qPCR. HL and MY were responsible for the statistical analyses and cell culture. YM and LZ contributed to interpreting the data and revising the manuscript. YL and FH contributed to conceiving and designing the experiments, evaluating and guiding the full text of the manuscript, and providing economic support. All authors read and approved the final manuscript.
This project was supported by the Sichuan Primary Health Development Research Center of NSMC (grant number: SWFZ19-Q-05) and the Science and Technology Strategic Cooperation Project of Nanchong and North Sichuan Medical College (grant number: 20SXQT0212 and 20SXQT0327).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The authors would like to thank the Narcotics Department, Nanchong Public Security Bureau of Sichuan, for kindly gifting the drugs.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2022.837123/full#supplementary-material
Anacker, C., Cattaneo, A., Musaelyan, K., Zunszain, P. A., Horowitz, M., Molteni, R., et al. (2013). Role for the Kinase SGK1 in Stress, Depression, and Glucocorticoid Effects on Hippocampal Neurogenesis. Proc. Natl. Acad. Sci. 110 (21), 8708–8713. doi:10.1073/pnas.1300886110
Ashburner, M., Ball, C. A., Blake, J. A., Botstein, D., Butler, H., Cherry, J. M., et al. (2000). Gene Ontology: Tool for the Unification of Biology. Nat. Genet. 25 (1), 25–29. doi:10.1038/75556
Bader, G. D., and Hogue, C. W. (2003). An Automated Method for Finding Molecular Complexes in Large Protein Interaction Networks. BMC Bioinformatics 4, 2. doi:10.1186/1471-2105-4-2
Bandettini, W. P., Kellman, P., Mancini, C., Booker, O. J., Vasu, S., Leung, S. W., et al. (2012). MultiContrast Delayed Enhancement (MCODE) Improves Detection of Subendocardial Myocardial Infarction by Late Gadolinium Enhancement Cardiovascular Magnetic Resonance: a Clinical Validation Study. J. Cardiovasc. Magn. Reson. 14, 83. doi:10.1186/1532-429X-14-83
Barrett, T., Troup, D. B., Wilhite, S. E., Ledoux, P., Evangelista, C., Kim, I. F., et al. (2010). NCBI GEO: Archive for Functional Genomics Data Sets--10 Years on. Nucleic Acids Res. 39, D1005–D1010. doi:10.1093/nar/gkq1184
Beauvais, G., Jayanthi, S., Mccoy, M. T., Ladenheim, B., and Cadet, J. L. (2010). Differential Effects of Methamphetamine and SCH23390 on the Expression of Members of IEG Families of Transcription Factors in the Rat Striatum. Brain Res. 1318, 1–10. doi:10.1016/j.brainres.2009.12.083
Blázquez-Prieto, J., Huidobro, C., López-Alonso, I., Amado-Rodriguez, L., Martín-Vicente, P., López-Martínez, C., et al. (2021). Activation of P21 Limits Acute Lung Injury and Induces Early Senescence after Acid Aspiration and Mechanical Ventilation. Translational Res. 233, 104–116. doi:10.1016/j.trsl.2021.01.008
Canal, M., Romaní-Aumedes, J., Martín-Flores, N., Pérez-Fernández, V. c., and Malagelada, C. (2014). RTP801/REDD1: a Stress Coping Regulator that Turns into a Troublemaker in Neurodegenerative Disorders. Front. Cel. Neurosci. 8, 313. doi:10.3389/fncel.2014.00313
Celentano, M., Caprioli, D., Di Pasquale, P., Cardillo, V., Nencini, P., Gaetani, S., et al. (2009). Drug Context Differently Regulates Cocaine versus Heroin Self-Administration and Cocaine- versus Heroin-Induced Fos mRNA Expression in the Rat. Psychopharmacology 204 (2), 349–360. doi:10.1007/s00213-009-1467-x
Chen, L.-M., Xiong, Y.-S., Kong, F.-L., Qu, M., Wang, Q., Chen, X.-Q., et al. (2012). Neuroglobin Attenuates Alzheimer-like Tau Hyperphosphorylation by Activating Akt Signaling. J. Neurochem. 120 (1), 157–164. doi:10.1111/j.1471-4159.2011.07275.x
Chen, R., Wang, B., Chen, L., Cai, D., Li, B., Chen, C., et al. (2016). DNA Damage-Inducible Transcript 4 (DDIT4) Mediates Methamphetamine-Induced Autophagy and Apoptosis through mTOR Signaling Pathway in Cardiomyocytes. Toxicol. Appl. Pharmacol. 295, 1–11. doi:10.1016/j.taap.2016.01.017
Chin, M. H., Qian, W.-J., Wang, H., Petyuk, V. A., Bloom, J. S., Sforza, D. M., et al. (2008). Mitochondrial Dysfunction, Oxidative Stress, and Apoptosis Revealed by Proteomic and Transcriptomic Analyses of the Striata in Two Mouse Models of Parkinson's Disease. J. Proteome Res. 7 (2), 666–677. doi:10.1021/pr070546l
Chin, C.-H., Chen, S.-H., Wu, H.-H., Ho, C.-W., Ko, M.-T., and Lin, C.-Y. (2014). cytoHubba: Identifying Hub Objects and Sub-networks from Complex Interactome. BMC Syst. Biol. 8 (Suppl. 4), S11. doi:10.1186/1752-0509-8-S4-S11
Cicero, T. J., Ellis, M. S., and Kasper, Z. A. (2020). Polysubstance Use: A Broader Understanding of Substance Use during the Opioid Crisis. Am. J. Public Health 110 (2), 244–250. doi:10.2105/AJPH.2019.305412
Davis, S., and Meltzer, P. S. (2007). GEOquery: a Bridge between the Gene Expression Omnibus (GEO) and BioConductor. Bioinformatics 23 (14), 1846–1847. doi:10.1093/bioinformatics/btm254
Demetrick, D. J., Matsumoto, S., Harmon, G. J., Okamoto, K., Xiong, Y., Zhang, H., et al. (1995). Chromosomal Mapping of the Genes for the Human Cell Cycle Proteins Cyclin C (CCNC), Cyclin E (CCNE), P21 (CDKN1) and KAP (CDKN3). Cytogenet. Cel Genet 69 (3-4), 190–192. doi:10.1159/000133960
Doris, J. M., Millar, S. A., Idris, I., and O'Sullivan, S. E. (2019). Genetic Polymorphisms of the Endocannabinoid System in Obesity and Diabetes. Diabetes Obes. Metab. 21 (2), 382–387. doi:10.1111/dom.13504
Dudek, H., Datta, S. R., Franke, T. F., Birnbaum, M. J., Yao, R., Cooper, G. M., et al. (1997). Regulation of Neuronal Survival by the Serine-Threonine Protein Kinase Akt. Science 275 (5300), 661–665. doi:10.1126/science.275.5300.661
Dumeige, L., Storey, C., Decourtye, L., Nehlich, M., Lhadj, C., Viengchareun, S., et al. (2017). Sex-Specificity of Mineralocorticoid Target Gene Expression during Renal Development, and Long-Term Consequences. Ijms 18 (2), 457. doi:10.3390/ijms18020457
Duric, V., Banasr, M., Licznerski, P., Schmidt, H. D., Stockmeier, C. A., Simen, A. A., et al. (2010). A Negative Regulator of MAP Kinase Causes Depressive Behavior. Nat. Med. 16 (11), 1328–1332. doi:10.1038/nm.2219
Ediriweera, M. K., Tennekoon, K. H., and Samarakoon, S. R. (2019). Role of the PI3K/AKT/mTOR Signaling Pathway in Ovarian Cancer: Biological and Therapeutic Significance. Semin. Cancer Biol. 59, 147–160. doi:10.1016/j.semcancer.2019.05.012
Ellis, M. S., Kasper, Z. A., and Cicero, T. J. (2018). Twin Epidemics: The Surging Rise of Methamphetamine Use in Chronic Opioid Users. Drug Alcohol Depend. 193, 14–20. doi:10.1016/j.drugalcdep.2018.08.029
Ellisen, L. W., Ramsayer, K. D., Johannessen, C. M., Yang, A., Beppu, H., Minda, K., et al. (2002). REDD1, a Developmentally Regulated Transcriptional Target of P63 and P53, Links P63 to Regulation of Reactive Oxygen Species. Mol. Cel. 10 (5), 995–1005. doi:10.1016/s1097-2765(02)00706-2
Eun, J. W., Kwack, S. J., Noh, J. H., Jung, K. H., Kim, J. K., Bae, H. J., et al. (2010). Identification of post-generation Effect of 3,4-methylenedioxymethamphetamine on the Mouse Brain by Large-Scale Gene Expression Analysis. Toxicol. Lett. 195 (1), 60–67. doi:10.1016/j.toxlet.2010.02.013
Fang, Q., Li, T., Chen, P., Wu, Y., Wang, T., Mo, L., et al. (2021). Comparative Analysis on Abnormal Methylome of Differentially Expressed Genes and Disease Pathways in the Immune Cells of RA and SLE. Front. Immunol. 12, 668007. doi:10.3389/fimmu.2021.668007
Fanous, S., Goldart, E. M., Theberge, F. R. M., Bossert, J. M., Shaham, Y., and Hope, B. T. (2012). Role of Orbitofrontal Cortex Neuronal Ensembles in the Expression of Incubation of Heroin Craving. J. Neurosci. 32 (34), 11600–11609. doi:10.1523/JNEUROSCI.1914-12.2012
Fattahi, F., Saeednejad Zanjani, L., Habibi Shams, Z., Kiani, J., Mehrazma, M., Najafi, M., et al. (2021). High Expression of DNA Damage-Inducible Transcript 4 (DDIT4) Is Associated with Advanced Pathological Features in the Patients with Colorectal Cancer. Sci. Rep. 11 (1), 13626. doi:10.1038/s41598-021-92720-z
Faust, H. J., Zhang, H., Han, J., Wolf, M. T., Jeon, O. H., Sadtler, K., et al. (2020). IL-17 and Immunologically Induced Senescence Regulate Response to Injury in Osteoarthritis. J. Clin. Invest. 130 (10), 5493–5507. doi:10.1172/JCI134091
Feng, J., and Nestler, E. J. (2013). Epigenetic Mechanisms of Drug Addiction. Curr. Opin. Neurobiol. 23 (4), 521–528. doi:10.1016/j.conb.2013.01.001
Gaesser, J. M., and Fyffe-Maricich, S. L. (2016). Intracellular Signaling Pathway Regulation of Myelination and Remyelination in the CNS. Exp. Neurol. 283, 501–511. doi:10.1016/j.expneurol.2016.03.008
Górska, A. M., Noworyta-Sokołowska, K., and Gołembiowska, K. (2014). The Effect of Caffeine on MDMA-Induced Hydroxyl Radical Production in the Mouse Striatum. Pharmacol. Rep. 66 (4), 718–721. doi:10.1016/j.pharep.2014.03.003
Guillot, A., Guerri, L., Feng, D., Kim, S.-J., Ahmed, Y. A., Paloczi, J., et al. (2021). Bile Acid-Activated Macrophages Promote Biliary Epithelial Cell Proliferation through Integrin αvβ6 Upregulation Following Liver Injury. J. Clin. Invest. 131 (9), e132305. doi:10.1172/JCI132305
Harlan, R. E., and Garcia, M. M. (1998). Drugs of Abuse and Immediate-Early Genes in the Forebrain. Mol. Neurobiol. 16 (3), 221–267. doi:10.1007/BF02741385
Horvath, M. C., Kovacs, G. G., Kovari, V., Majtenyi, K., Hurd, Y. L., and Keller, E. (2007). Heroin Abuse Is Characterized by Discrete Mesolimbic Dopamine and Opioid Abnormalities and Exaggerated Nuclear Receptor-Related 1 Transcriptional Decline with Age. J. Neurosci. 27 (49), 13371–13375. doi:10.1523/JNEUROSCI.2398-07.2007
Hughes, P., and Dragunow, M. (1995). Induction of Immediate-Early Genes and the Control of Neurotransmitter-Regulated Gene Expression within the Nervous System. Pharmacol. Rev. 47 (1), 133–178.
Janky, R. s., Verfaillie, A., Imrichová, H., Van de Sande, B., Standaert, L., Christiaens, V., et al. (2014). iRegulon: from a Gene List to a Gene Regulatory Network Using Large Motif and Track Collections. Plos Comput. Biol. 10 (7), e1003731. doi:10.1371/journal.pcbi.1003731
Jin, H. O., Hong, S. E., Kim, J. Y., Kim, M. R., Chang, Y., Hong, Y., et al. (2019). Induction of HSP27 and HSP70 by Constitutive Overexpression of Redd1 Confers Resistance of Lung Cancer Cells to Ionizing Radiation. Oncol. Rep. 41 (5), 3119–3126. doi:10.3892/or.2019.7036
Jones, C. M., Underwood, N., and Compton, W. M. (2019). Increases in Methamphetamine Use Among Heroin Treatment Admissions in the United States, 2008-17. Addiction 115 (2), 347–353. doi:10.1111/add.14812
Jung, Y., Hsieh, L. S., Lee, A. M., Zhou, Z., Coman, D., Heath, C. J., et al. (2016). An Epigenetic Mechanism Mediates Developmental Nicotine Effects on Neuronal Structure and Behavior. Nat. Neurosci. 19 (7), 905–914. doi:10.1038/nn.4315
Kakati, T., Bhattacharyya, D. K., Barah, P., and Kalita, J. K. (2019). Comparison of Methods for Differential Co-expression Analysis for Disease Biomarker Prediction. Comput. Biol. Med. 113, 103380. doi:10.1016/j.compbiomed.2019.103380
Kanehisa, M., Sato, Y., Furumichi, M., Morishima, K., and Tanabe, M. (2019). New Approach for Understanding Genome Variations in KEGG. Nucleic Acids Res. 47 (D1), D590–D595. doi:10.1093/nar/gky962
Kang, Q., Li, W., Xiao, J., Yu, N., Fan, L., Sha, M., et al. (2021). Integrated Analysis of Multiple Microarray Studies to Identify Novel Gene Signatures in Preeclampsia. Placenta 105, 104–118. doi:10.1016/j.placenta.2021.01.023
Kobayashi, T., Deak, M., Morrice, N., and Cohen, P. (1999). Characterization of the Structure and Regulation of Two Novel Isoforms of Serum- and Glucocorticoid-Induced Protein Kinase. Biochem. J. 344, 189–197. doi:10.1042/bj3440189
Korostynski, M., Piechota, M., Kaminska, D., Solecki, W., and Przewlocki, R. (2007). Morphine Effects on Striatal Transcriptome in Mice. Genome Biol. 8 (6), R128. doi:10.1186/gb-2007-8-6-r128
Korostynski, M., Piechota, M., Dzbek, J., Mlynarski, W., Szklarczyk, K., Ziolkowska, B., et al. (2013). Novel Drug-Regulated Transcriptional Networks in Brain Reveal Pharmacological Properties of Psychotropic Drugs. BMC Genomics 14 (1), 606. doi:10.1186/1471-2164-14-606
Kuntz-Melcavage, K. L., Brucklacher, R. M., Grigson, P. S., Freeman, W. M., and Vrana, K. E. (2009). Gene Expression Changes Following Extinction Testing in a Heroin Behavioral Incubation Model. BMC Neurosci. 10 (1), 95. doi:10.1186/1471-2202-10-95
Kuroda, K. O., Ornthanalai, V. G., Kato, T., and Murphy, N. P. (2010). FosB Null Mutant Mice Show Enhanced Methamphetamine Neurotoxicity: Potential Involvement of FosB in Intracellular Feedback Signaling and Astroglial Function. Neuropsychopharmacol 35 (3), 641–655. doi:10.1038/npp.2009.169
Lai, B., Pu, H., Cao, Q., Jing, H., and Liu, X. (2011). Activation of Caspase-3 and C-Jun NH2-terminal Kinase Signaling Pathways Involving Heroin-Induced Neuronal Apoptosis. Neurosci. Lett. 502 (3), 209–213. doi:10.1016/j.neulet.2011.07.046
Lang, C. H., Frost, R. A., and Vary, T. C. (2008). Acute Alcohol Intoxication Increases REDD1 in Skeletal Muscle. Alcohol. Clin. Exp. Res. 32 (5), 796–805. doi:10.1111/j.1530-0277.2008.00637.x
Li, B., Chen, R., Chen, L., Qiu, P., Ai, X., Huang, E., et al. (2017). Effects of DDIT4 in Methamphetamine-Induced Autophagy and Apoptosis in Dopaminergic Neurons. Mol. Neurobiol. 54 (3), 1642–1660. doi:10.1007/s12035-015-9637-9
Li, Z., Peng, X., Jia, X., Su, P., Liu, D., Tu, Y., et al. (2020). Spinal Heat Shock Protein 27 Participates in PDGFRβ‐mediated Morphine Tolerance through PI3K/Akt and P38 MAPK Signalling Pathways. Br. J. Pharmacol. 177 (22), 5046–5062. doi:10.1111/bph.15169
Lopez-Leon, S., González-Giraldo, Y., Wegman-Ostrosky, T., and Forero, D. A. (2021). Molecular Genetics of Substance Use Disorders: An Umbrella Review. Neurosci. Biobehav. Rev. 124, 358–369. doi:10.1016/j.neubiorev.2021.01.019
López-Moreno, J. A., Echeverry-Alzate, V., and Bühler, K.-M. (2012). The Genetic Basis of the Endocannabinoid System and Drug Addiction in Humans. J. Psychopharmacol. 26 (1), 133–143. doi:10.1177/0269881111416689
Malagelada, C., Ryu, E. J., Biswas, S. C., Jackson-Lewis, V., and Greene, L. A. (2006). RTP801 Is Elevated in Parkinson Brain Substantia Nigral Neurons and Mediates Death in Cellular Models of Parkinson's Disease by a Mechanism Involving Mammalian Target of Rapamycin Inactivation. J. Neurosci. 26 (39), 9996–10005. doi:10.1523/JNEUROSCI.3292-06.2006
Mattson, B., Crombag, H., Mitchell, T., Simmons, D., Kreuter, J., Morales, M., et al. (2007). Repeated Amphetamine Administration outside the home Cage Enhances Drug-Induced Fos Expression in Rat Nucleus Accumbens. Behav. Brain Res. 185 (2), 88–98. doi:10.1016/j.bbr.2007.07.024
Mcadams, R. M., Mcpherson, R. J., Beyer, R. P., Bammler, T. K., Farin, F. M., and Juul, S. E. (2015). Dose-dependent Effects of Morphine Exposure on mRNA and microRNA (miR) Expression in hippocampus of Stressed Neonatal Mice. PLoS One 10 (4), e0123047. doi:10.1371/journal.pone.0123047
Meng, X., Zhang, C., Guo, Y., Han, Y., Wang, C., Chu, H., et al. (2020). TBHQ Attenuates Neurotoxicity Induced by Methamphetamine in the VTA through the Nrf2/HO-1 and PI3K/AKT Signaling Pathways. Oxid. Med. Cell Longev. 2020, 1–13. doi:10.1155/2020/8787156
Moratalla, R., Khairnar, A., Simola, N., Granado, N., García-Montes, J. R., Porceddu, P. F., et al. (2017). Amphetamine-related Drugs Neurotoxicity in Humans and in Experimental Animals: Main Mechanisms. Prog. Neurobiol. 155, 149–170. doi:10.1016/j.pneurobio.2015.09.011
Nangraj, A. S., Selvaraj, G., Kaliamurthi, S., Kaushik, A. C., Cho, W. C., and Wei, D. Q. (2020). Integrated PPI- and WGCNA-Retrieval of Hub Gene Signatures Shared between Barrett's Esophagus and Esophageal Adenocarcinoma. Front. Pharmacol. 11, 881. doi:10.3389/fphar.2020.00881
Palmer, A. A., Verbitsky, M., Suresh, R., Kamens, H. M., Reed, C. L., Li, N., et al. (2005). Gene Expression Differences in Mice Divergently Selected for Methamphetamine Sensitivity. Mamm. Genome 16 (5), 291–305. doi:10.1007/s00335-004-2451-8
Paolone, G., Conversi, D., Caprioli, D., Bianco, P. D., Nencini, P., Cabib, S., et al. (2007). Modulatory Effect of Environmental Context and Drug History on Heroin-Induced Psychomotor Activity and Fos Protein Expression in the Rat Brain. Neuropsychopharmacol 32 (12), 2611–2623. doi:10.1038/sj.npp.1301388
Park, J.-H., Seo, Y. H., Jang, J.-H., Jeong, C.-H., Lee, S., and Park, B. (2017). Asiatic Acid Attenuates Methamphetamine-Induced Neuroinflammation and Neurotoxicity through Blocking of NF-kB/STAT3/ERK and Mitochondria-Mediated Apoptosis Pathway. J. Neuroinflamm. 14 (1), 240. doi:10.1186/s12974-017-1009-0
Piechota, M., Korostynski, M., Solecki, W., Gieryk, A., Slezak, M., Bilecki, W., et al. (2010). The Dissection of Transcriptional Modules Regulated by Various Drugs of Abuse in the Mouse Striatum. Genome Biol. 11 (5), R48. doi:10.1186/gb-2010-11-5-r48
Piechota, M., Korostynski, M., Sikora, M., Golda, S., Dzbek, J., and Przewlocki, R. (2012). Common Transcriptional Effects in the Mouse Striatum Following Chronic Treatment with Heroin and Methamphetamine. Genes, Brain Behav. 11 (4), 404–414. doi:10.1111/j.1601-183X.2012.00777.x
Qi, B., Chen, J.-H., Tao, L., Zhu, C.-M., Wang, Y., Deng, G.-X., et al. (2021). Integrated Weighted Gene Co-expression Network Analysis Identified that TLR2 and CD40 Are Related to Coronary Artery Disease. Front. Genet. 11, 613744. doi:10.3389/fgene.2020.613744
Qiao, X., Gai, H., Su, R., Deji, C., Cui, J., Lai, J., et al. (2018). PI3K-AKT-GSK3β-CREB Signaling Pathway Regulates Anxiety-like Behavior in Rats Following Alcohol Withdrawal. J. Affective Disord. 235, 96–104. doi:10.1016/j.jad.2018.04.039
Renthal, W., Kumar, A., Xiao, G., Wilkinson, M., Covington, H. E., Maze, I., et al. (2009). Genome-wide Analysis of Chromatin Regulation by Cocaine Reveals a Role for Sirtuins. Neuron 62 (3), 335–348. doi:10.1016/j.neuron.2009.03.026
Rubio, F. J., Liu, Q.-R., Li, X., Cruz, F. C., Leao, R. M., Warren, B. L., et al. (2015). Context-Induced Reinstatement of Methamphetamine Seeking Is Associated with Unique Molecular Alterations in Fos-Expressing Dorsolateral Striatum Neurons. J. Neurosci. 35 (14), 5625–5639. doi:10.1523/JNEUROSCI.4997-14.2015
Sabrini, S., Russell, B., Wang, G., Lin, J., Kirk, I., and Curley, L. (2020). Methamphetamine Induces Neuronal Death: Evidence from Rodent Studies. Neurotoxicology 77, 20–28. doi:10.1016/j.neuro.2019.12.006
Saijo, K., Winner, B., Carson, C. T., Collier, J. G., Boyer, L., Rosenfeld, M. G., et al. (2009). A Nurr1/CoREST Pathway in Microglia and Astrocytes Protects Dopaminergic Neurons from Inflammation-Induced Death. Cell 137 (1), 47–59. doi:10.1016/j.cell.2009.01.038
Sanchis-Segura, C., Lopez-Atalaya, J. P., and Barco, A. (2009). Selective Boosting of Transcriptional and Behavioral Responses to Drugs of Abuse by Histone Deacetylase Inhibition. Neuropsychopharmacol 34 (13), 2642–2654. doi:10.1038/npp.2009.125
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 13 (11), 2498–2504. doi:10.1101/gr.1239303
Shoshani, T., Faerman, A., Mett, I., Zelin, E., Tenne, T., Gorodin, S., et al. (2002). Identification of a Novel Hypoxia-Inducible Factor 1-Responsive Gene, RTP801 , Involved in Apoptosis. Mol. Cel. Biol. 22 (7), 2283–2293. doi:10.1128/MCB.22.7.2283-2293.2002
Silva, C. D., Neves, A. F., Dias, A. I., Freitas, H. J., Mendes, S. M., Pita, I., et al. (2014). A Single Neurotoxic Dose of Methamphetamine Induces a Long-Lasting Depressive-like Behaviour in Mice. Neurotox. Res. 25 (3), 295–304. doi:10.1007/s12640-013-9423-2
Singh, M. E., Mcgregor, I. S., and Mallet, P. E. (2005). Repeated Exposure to Δ9-tetrahydrocannabinol Alters Heroin-Induced Locomotor Sensitisation and Fos-Immunoreactivity. Neuropharmacology 49 (8), 1189–1200. doi:10.1016/j.neuropharm.2005.07.008
Sinha, D., Kalimutho, M., Bowles, J., Chan, A.-L., Merriner, D. J., Bain, A. L., et al. (2018). Cep55 Overexpression Causes Male‐specific Sterility in Mice by Suppressing Foxo1 Nuclear Retention through Sustained Activation of PI3K/Akt Signaling. FASEB j. 32 (9), 4984–4999. doi:10.1096/fj.201701096RR
Skupio, U., Sikora, M., Korostynski, M., Wawrzczak-Bargiela, A., Piechota, M., Ficek, J., et al. (2017). Behavioral and Transcriptional Patterns of Protracted Opioid Self-Administration in Mice. Addict. Biol. 22 (6), 1802–1816. doi:10.1111/adb.12449
Smith, M. L., Lopez, M. F., Wolen, A. R., Becker, H. C., and Miles, M. F. (2020). Brain Regional Gene Expression Network Analysis Identifies Unique Interactions between Chronic Ethanol Exposure and Consumption. PLoS One 15 (5), e0233319. doi:10.1371/journal.pone.0233319
Steiner, J. L., Kimball, S. R., and Lang, C. H. (2016). Acute Alcohol-Induced Decrease in Muscle Protein Synthesis in Female Mice Is REDD-1 and mTOR-Independent. Alcohol Alcohol. 51 (3), 242–250. doi:10.1093/alcalc/agv105
Su, W., Zhao, Y., Wei, Y., Zhang, X., Ji, J., and Yang, S. (2021). Exploring the Pathogenesis of Psoriasis Complicated with Atherosclerosis via Microarray Data Analysis. Front. Immunol. 12, 667690. doi:10.3389/fimmu.2021.667690
Sun, F., Zhou, J. L., Peng, P. J., Qiu, C., Cao, J. R., and Peng, H. (2021). Identification of Disease-Specific Hub Biomarkers and Immune Infiltration in Osteoarthritis and Rheumatoid Arthritis Synovial Tissues by Bioinformatics Analysis. Dis. Markers 2021, 1–17. doi:10.1155/2021/9911184
Szklarczyk, D., Gable, A. L., Lyon, D., Junge, A., Wyder, S., Huerta-Cepas, J., et al. (2019). STRING V11: Protein-Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-wide Experimental Datasets. Nucleic Acids Res. 47 (D1), D607–D613. doi:10.1093/nar/gky1131
Takaki, M., Ujike, H., Kodama, M., Takehisa, Y., Nakata, K., and Kuroda, S. (2001). Two Kinds of Mitogen-Activated Protein Kinase Phosphatases, MKP-1 and MKP-3, Are Differentially Activated by Acute and Chronic Methamphetamine Treatment in the Rat Brain. J. Neurochem. 79 (3), 679–688. doi:10.1046/j.1471-4159.2001.00615.x
Torii, T., Kawarai, T., Nakamura, S., and Kawakami, H. (1999). Organization of the Human Orphan Nuclear Receptor Nurr1 Gene. Gene 230 (2), 225–232. doi:10.1016/s0378-1119(99)00064-5
van Dam, S., Võsa, U., van der Graaf, A., Franke, L., and de Magalhães, J. P. (2017). Gene Co-expression Analysis for Functional Classification and Gene-Disease Predictions. Brief. Bioinform. 19 (4), 575–592. doi:10.1093/bib/bbw139
Visvader, J., Sassone-Corsi, P., and Verma, I. M. (1988). Two Adjacent Promoter Elements Mediate Nerve Growth Factor Activation of the C-Fos Gene and Bind Distinct Nuclear Complexes. Proc. Natl. Acad. Sci. 85 (24), 9474–9478. doi:10.1073/pnas.85.24.9474
Wang, Z., Guo, M., Ai, X., Cheng, J., Huang, Z., Li, X., et al. (2021). Identification of Potential Diagnostic and Prognostic Biomarkers for Colorectal Cancer Based on GEO and TCGA Databases. Front. Genet. 11, 602922. doi:10.3389/fgene.2020.602922
Wojcieszak, J., Andrzejczak, D., Szymańska, B., and Zawilska, J. B. (2019). Induction of Immediate Early Genes Expression in the Mouse Striatum Following Acute Administration of Synthetic Cathinones. Pharmacol. Rep. 71 (6), 977–982. doi:10.1016/j.pharep.2019.05.011
Wolff, N. C., Mckay, R. M., and Brugarolas, J. (2014). REDD1/DDIT4-independent mTORC1 Inhibition and Apoptosis by Glucocorticoids in Thymocytes. Mol. Cancer Res. 12 (6), 867–877. doi:10.1158/1541-7786.MCR-13-0625
Wong, Y.-H., Wu, C.-C., Wu, J., Lai, H.-Y., Chen, K.-Y., Jheng, B.-R., et al. (2016). Temporal Genetic Modifications after Controlled Cortical Impact-Understanding Traumatic Brain Injury through a Systematic Network Approach. Ijms 17 (2), 216. doi:10.3390/ijms17020216
Wu, M., Wang, B., Fei, J., Santanam, N., and Blough, E. R. (2010). Important Roles of Akt PKB Signaling in the Aging Process. Front. Biosci. S2, 1169–1188. doi:10.2741/s125
Xu, H., Xie, Y., Sun, Y., Guo, R., Lv, D., Li, X., et al. (2021). Integrated Analysis of Multiple Microarray Studies to Identify Potential Pathogenic Gene Modules in Preeclampsia. Exp. Mol. Pathol. 120, 104631. doi:10.1016/j.yexmp.2021.104631
Chen, X., Qiu, F., Zhao, X., Lu, J., Tan, X., Xu, J., et al. (2020). Astrocyte-Derived Lipocalin-2 Is Involved in Mitochondrion-Related Neuronal Apoptosis Induced by Methamphetamine. ACS Chem. Neurosci. 11 (8), 1102–1116. doi:10.1021/acschemneuro.9b00559
Chen, Y., Li, H., Lai, L., Feng, Q., and Shen, J. (2020). Identification of Common Differentially Expressed Genes and Potential Therapeutic Targets in Ulcerative Colitis and Rheumatoid Arthritis. Front. Genet. 11, 572194. doi:10.3389/fgene.2020.572194
Zetterström, R. H., Solomin, L., Jansson, L., Hoffer, B. J., Olson, L., and Perlmann, T. (1997). Dopamine Neuron Agenesis in Nurr1-Deficient Mice. Science 276 (5310), 248–250. doi:10.1126/science.276.5310.248
Zhang, W., Liu, H., Deng, X. D., Ma, Y., and Liu, Y. (2020). FAAH Levels and its Genetic Polymorphism Association with Susceptibility to Methamphetamine Dependence. Ann. Hum. Genet. 84 (3), 259–270. doi:10.1111/ahg.12368
Zhou, S., Liu, S., Liu, X., and Zhuang, W. (2021). Bioinformatics Gene Analysis of Potential Biomarkers and Therapeutic Targets for Unstable Atherosclerotic Plaque-Related Stroke. J. Mol. Neurosci. 71 (5), 1031–1045. doi:10.1007/s12031-020-01725-2
Zhu, L., Ding, Y., Chen, C.-Y., Wang, L., Huo, Z., Kim, S., et al. (2016). MetaDCN: Meta-Analysis Framework for Differential Co-expression Network Detection with an Application in Breast Cancer. Bioinformatics 33 (8), 1121–1129. doi:10.1093/bioinformatics/btw788
Keywords: amphetamine-type stimulants (ATS), opioids, differentially expressed genes (DEGs), PI3K/Akt pathway, apoptosis, hub gene
Citation: Zhang W, Deng X, Liu H, Ke J, Xiang M, Ma Y, Zhang L, Yang M, Liu Y and Huang F (2022) Identification and Verification of Potential Hub Genes in Amphetamine-Type Stimulant (ATS) and Opioid Dependence by Bioinformatic Analysis. Front. Genet. 13:837123. doi: 10.3389/fgene.2022.837123
Received: 16 December 2021; Accepted: 21 February 2022;
Published: 30 March 2022.
Edited by:
Błażej Misiak, Wroclaw Medical University, PolandReviewed by:
Laura Beth Kozell, Oregon Health and Science University, United StatesCopyright © 2022 Zhang, Deng, Liu, Ke, Xiang, Ma, Zhang, Yang, Liu and Huang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yun Liu, eHl1bjIwMDVAMTYzLmNvbQ==; Feijun Huang, aGZqNjAxMjNAaG90bWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.