
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 09 January 2023
Sec. Cancer Genetics and Oncogenomics
Volume 13 - 2022 | https://doi.org/10.3389/fgene.2022.1073232
This article is part of the Research Topic Molecular Mechanisms of Prostate Cancer Initiation and Progression View all 4 articles
 Bao Zhang1*†
Bao Zhang1*† Shenghan Wang1†
Shenghan Wang1† Zhichao Fu2†Qiang Gao1Lin Yang1Zhentao Lei1Yuqiang Shi1Kai Le1Jie Xiong1
Zhichao Fu2†Qiang Gao1Lin Yang1Zhentao Lei1Yuqiang Shi1Kai Le1Jie Xiong1 Siyao Liu2Jiali Zhang2
Siyao Liu2Jiali Zhang2 Junyan Su2Jing Chen2
Junyan Su2Jing Chen2 Mengyuan Liu2,3
Mengyuan Liu2,3 Beifang Niu2,3,4*
Beifang Niu2,3,4*Background: Prostate cancer (PCa) is the second most common cancer among men worldwide. Perineural invasion (PNI) was a prominent characteristic of PCa, which was recognized as a key factor in promoting PCa progression. As a complex and heterogeneous disease, its true condition is difficult to explain thoroughly with conventional bulk RNA sequencing. Thus, an improved understanding of PNI-PCa progression at the single-cell level is needed.
Methods: In this study, we performed scRNAseq on tumor tissues of three PNI-PCa patients. Principal component analysis (PCA) and Uniform manifold approximation and projection (UMAP) were used to reduce dimensionality and visualize the cellular composition of tumor tissues. The differently expressed genes among each cluster were identified by EdgeR. GO enrichment analysis was used to understand the roles of genes within the clusters. Pseudotime cell trajectory was used to reveal the molecular pathways underlying cell fate decisions and identify genes whose expression changed as the cells underwent transition. We applied CellPhoneDB to identify cell-cell interactions among the epithelial and neural cells in PNI-PCa.
Results: Analysis of the ∼17,000 single-cell transcriptomes in three PNI prostate cancer tissues, we identified 12 major cell clusters, including neural cells and two epithelial subtypes with different expression profiles. We found that basal/intermediate epithelial cell subtypes highly expressed PCa progression-related genes, including PIGR, MMP7, and AGR2. Pseudotime trajectory analysis showed that luminal epithelial cells could be the initiating cells and transition to based/intermediate cells. Gene ontology (GO) enrichment analysis showed that pathways related to cancer progressions, such as lipid catabolic and fatty acid metabolic processes, were significantly enriched in basal/intermediate cells. Our analysis also suggested that basal/intermediate cells communicate closely with neural cells played a potential role in PNI-PCa progression.
Conclusion: These results provide our understanding of PNI-PCa cellular heterogeneity and characterize the potential role of basal/intermediate cells in the PNI-PCa progression.
PCa is currently the most commonly diagnosed malignancy in the worldwide male population and one of the frequent leading causes of cancer-related death in men (Siegel et al., 2020). PNI is one of the characteristics of PCa and a prognostic factor in radical prostatectomy specimens. It is defined as cancer cell invasion in, around, and through nerves and is a well-established mechanism of tumor spread. In recent years, it has been demonstrated that PNI was associated with poor survival and increased risk for metastasis in PCa (Wu et al., 2019; Lin et al., 2020).
The majority of prostate cancers are carcinomas, which originate from epithelial cells. There is evidence implicating epithelial in prostate cancer (PCa) progression. Under certain physiological and pathological conditions, epithelial cells differentiate into mesenchyme, accompanied by changes in cell morphology and related genes (Brabletz et al., 2018), which is called epithelial mesenchymal transition (EMT). Emerging evidence demonstrates that the EMT process can promote tumor cell invasion and metastasis by loss of cell-cell connections and cell polarity (Dudas et al., 2020). Furthermore, many studies have reported the effects of PNI on patients’ recurrence, metastasis, and prostate cancer-specific survival. In vitro experiments have illustrated that prostate cancer cells in close contact with nerves exhibit a reduced rate of apoptosis (Deborde and Wong, 2017). A meta-analysis by Zhang et al. (2018) demonstrated that the presence of PNI in PCa was associated with a higher risk of biochemical recurrence, whether undergoing radical prostate prostatectomy or radiotherapy. Importantly, recent studies have identified several mechanisms by which neural infiltration promotes disease development and metastasis. These included PNI, which results in PCa-related axonotmesis, neurotmesis, and cancer cell proliferation and migration (Fromont et al., 2012; Zhang et al., 2016). Moreover, some studies have reported that the progression of cancer results from diverse signaling transduction between cancer cells and the neural microenvironment driven by chemokines, neurotrophic factors, and axon guidance molecules through ligand-receptor interactive mode. Specifically, monoamine oxidase A (MAOA), nerve growth factor (NGF), semaphorin4D (SEMA-4D), and C-X-C motif chemokine ligand 12 (CXCL12) were recently demonstrated to have an active role in promoting PNI in PCa and may be critical mediators in the pathogenesis of PNI (Zhang et al., 2008; Binmadi et al., 2012; Liss et al., 2014; Yin et al., 2021).
Although several molecular mechanisms have been discovered involving prostate tumor development and progression, most of these studies have focused on bulk tissue samples or cell populations. As PCa is a highly heterogeneous tumor, its true condition is difficult to explain thoroughly with conventional bulk RNA sequencing, which leaves a gap in our understanding of the mechanism underlying cancer progression at the single-cell level and hampers the development of novel therapeutic strategies for PCa treatment. scRNA-seq has become the primary tool for characterizing the transcriptomes in thousands of individual cells and provides unprecedented opportunities to understand the cellular heterogeneity and interaction of individual cells (Aibar et al., 2017; Kumar et al., 2018). Although this technology has been applied to several types of cancer in multiple cancers (Chen et al., 2021; He et al., 2021), there are few studies on PNI-PCa. Thus, further analysis of PNI-PCa at the single-cell level is essential for discovering the causes contributing to disease progression.
In this study, we conduct a comprehensive evaluation of prostate tumor tissues by scRNA-seq to analyze their cellular compositions and explore the specific cell subtypes associated with tumor progression of PNI-PCa. Our results indicated that the basal/intermediate epithelial cells are associated with the progression of PNI-PCa. Further, we showed that the interactions between basal/intermediate epithelial cell subtypes and neural cells subtypes through CXCL and CCL signaling network pathways promote tumor progression. These findings have important implications for our understanding of the mechanistic related to PNI-PCa progression and provide some new references for the therapeutic strategy for PNI-PCa.
The tissue samples were obtained from three PNI-PCa patients who underwent surgical resection at the Department of Urology, Aerospace Center Hospital. All patients had lesions visible on preoperative Magnetic Resonance Imaging (MRI) or ultrasound and were later pathologically diagnosed as PCa with perineural invasion (Supplementary Figure S1). All patients provided informed consent and the study protocol permitted by the Ethics Committee of Aerospace Center Hospital.
The tumor tissues were dissociated using Multi-tissue dissociation kit 2 (Miltenyi) after being washed in ice-cold RPMI1640. DNase treatment was performed according to the manufacturer’s protocols. After removing erythrocytes (Miltenyi 130-094-183), cell count and viability were determined by Cell Analyzer (Countstar® Rigel S2) with AO/PI reagent. Then debris and dead cell removal were decided on whether to be performed or not (Miltenyi 130-109-398/130-090-101). Finally, new cells were washed twice in the RPMI1640 and resuspended at 1 × 106 cells per ml in 1 × PBS and .04% bovine serum albumin.
Single-cell RNA-Seq libraries were prepared using SeekOne® MM Single Cell 3′ library preparation kit Briefly, the appropriate number of cells were loaded into the flow channel of the SeekOne® MM chip, which had 170,000 microwells and was allowed to settle in microwells by gravity. After removing the unsettled cells, sufficient Cell Barcoded Magnetic Beads (CBBs) were pipetted into the flow channel and allowed to settle in microwells with the help of a magnetic field. Next excess CBBs were rinsed out, and cells in the MM chip were lysed to release RNA, which was captured by the CBB in the same microwell. Then all CBBs were collected, and reverse transcription was performed at 37°C for 30 min to label cDNA with cell barcode on the beads. Further, Exonuclease I treatment was performed to remove unused primer on CBBs. Subsequently, barcoded cDNA on the CBBs was hybridized with a random primer which had reads 2 SeqPrimer sequences on the 5′ end and could extend to form the second strand DNA with cell barcode on the 3’ end. The resulting second strand DNA was denatured off the CBBs, purified, and amplified in polymerase chain reaction (PCR). The amplified cDNA product was then cleaned to remove unwanted fragments and added to a full-length sequencing adapter and sample index by indexed PCR. The indexed sequencing libraries were cleaned up with SPRI beads, quantified using quantitative PCR (KAPA Biosystems KK4824), and then sequenced (PE150) by Illumina NovaSeq 6,000 platform.
The raw sequencing data were processed by fastp (version .20.0) (Chen et al., 2018) firstly to trim primer sequence and low-quality bases. And then, we used SeekOne®Tools to process sequence data and aligned it to human GRCh38 to obtain the gene expression matrix. We used Seurat (version 4.0.3) to filter low-quality cells. The UMI counts per cell should generally be above 300, and belonging to mitochondrial genes was also omitted. The remaining cells were used for the following analysis.
The Seurat package “NormalizeData” function was used to normalize the matrix data of single-cell counts, where gene expression for each cell was normalized by dividing the total expression counts, multiplied by a scale factor of 10,000, and then log-transformed. We conducted normalization and then performed principal component analysis (PCA) on the normalized expression matrix using highly variable genes identified by the Seurat R package (version 4.0.0) (Satija et al., 2015). Following the results of PCA, the top 30 PCs were selected with a resolution parameter equal to .8. The cell clusters were visualized using the uniform manifold approximation and projection (UMAP). The “FindAllMarkers” function in Seurat was used to identify differentially expressed marker genes for each cell type. Marker genes were selected as those with adjusted p values less than .05, average logFC larger than .25, and percentage of cells with expression higher than .25. Cell type identities were determined based on the expression of known markers in the Cell Marker database (Zhang et al., 2019).
The differentially expressed genes DEGs were identified using the edgeR package and filtered by |log2FoldChange| ≥ 1 and FDR <.05. GO enrichment analysis of DEGs involved in the biological process was implemented by the “ClusterProfiler” package in R software.
We used CellPhoneDB to infer cell-cell interactions, processed and normalized logarithmic counts, and cell type annotations were used as an input. To shrink down the most relevant interactions, specific interactions sorted by ligand/receptor expression were investigated in over 10% of the cells in a cluster, where the log2 mean expression of the ligand/receptor pair is greater than 0.
Monocle (version 2.3.6) (Trapnell et al., 2014) was used to estimate lineage differentiation between cell populations using default parameters. First, a CellDataSet object was created with the parameter expressionFamily as negbinomial, then genes used for pseudotime ordering were selected using the ‘dispersionTable’ Function. The ‘DDRTree’ method was utilized for dimension reduction and cell ordering along the pseudo time trajectory. Branch analysis was performed with the branched expression analysis modeling (BEAM) function. The significantly changed genes in the branch point were clustered using “plot_genes_branched_pseudotime” functions according to the distinct patterns of gene expression change.
To understand PNI prostate tumor heterogeneity at single-cell resolution, we obtained tissue samples from patients diagnosed as PCa with perineural invasion and performed single-cell RNA sequencing. The study workflow is shown in Figure 1A. After data processing and normalization, we obtained 17,561 qualified cells for further analysis. Next, we examined the cellular composition with unbiased clustering across all cells by PCA and visualized by UMAP for Dimension Reduction (Figure 1B). To identify the different major cell types, we analyzed the expression level of the cluster-specific gene markers in each cluster identified by examining DEGs, allowing us to categorize these clusters into 12 main clusters: T lymphocyte, natural killer cell, B lymphocyte, macrophage, neutrophil, mast cell, endothelial cell, fibroblast, smooth muscle cell (SMC), neural cell and cell (Figure 1C). Strikingly, we found that the epithelial cells population was divided into two major subsets: luminal and basal/intermediate cells. Differential gene expression analysis results in epithelial cell subtypes showed that basal/intermediate cells exhibited much higher expression levels of basal markers, including KRT5, KRT15, and KRT17, compared with other clusters. The gene of KLK11 was specifically expressed in the luminal cells, which could potentially be used as specific markers for the detection of luminal cells in PNI-PCa. An analysis of prostate cancer-specific gene expression levels in each cell type was performed to evaluate the malignancy of these cell clusters. We found that the prostate cancer-specific genes, including KLK3, KLK2, FOLH1, and ACPP, are expressed mainly in luminal cells (Figure 1D). Furthermore, we identified a subset of neural cells according to the high expression level of marker gene CSF3 (Figure 1C). We further grouped neuronal cells into four clusters based on the scRNA-seq data (Supplementary Figure S2).
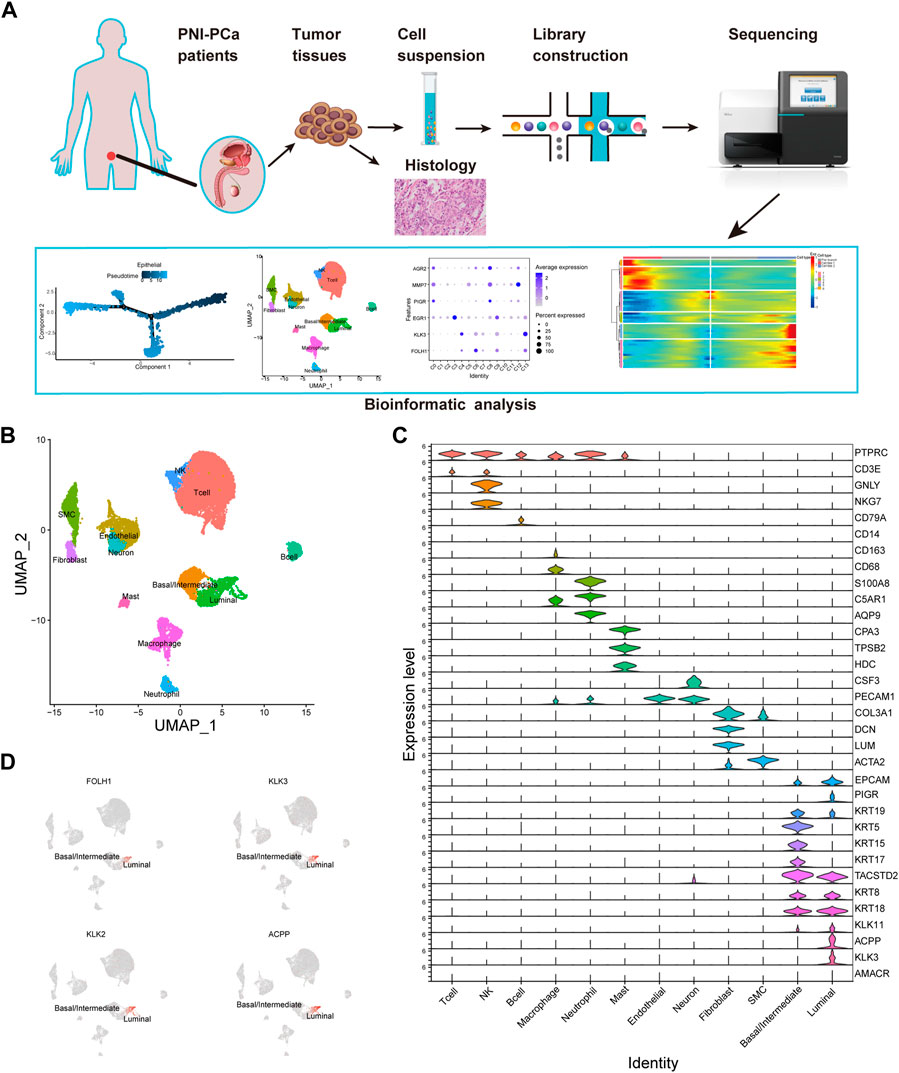
FIGURE 1. Diverse cell types in PNI-PCa were identified by single-cell sequencing. (A) Single-cell sequencing flow chart. (B) UMAP exhibits the cell clusters in PNI-PCa and different colors labeled for different cell types according to their featured gene expression profiles. (C) Violin plots show the expression levels of marker genes for each cell type. (D) UMAP shows the expression of prostate cancer-specific genes localized specifically to the luminal epithelial cells.
To further explore the potential roles of epithelial cells in PNI-PCa, we performed a graph-based clustering analysis, and 14 clusters were identified (C0-C13, Figure 2A). To characterize these epithelial clusters, we used VlnPlot to examine the expression patterns of the specific marker genes individually. The results showed that the genes including KRT5, KRT15, KRT19, and KRT17, exhibited higher expression levels in C0, C1, C2, C3, C5, C8, C10, and C12 (Figure 2B), which were identified as basal/intermediate cells subtypes. Other epithelial cells, including C4, C6, C7, C9, C11, and C13, were identified as luminal cells based on the expression of specific markers KLK3 and ACPP.
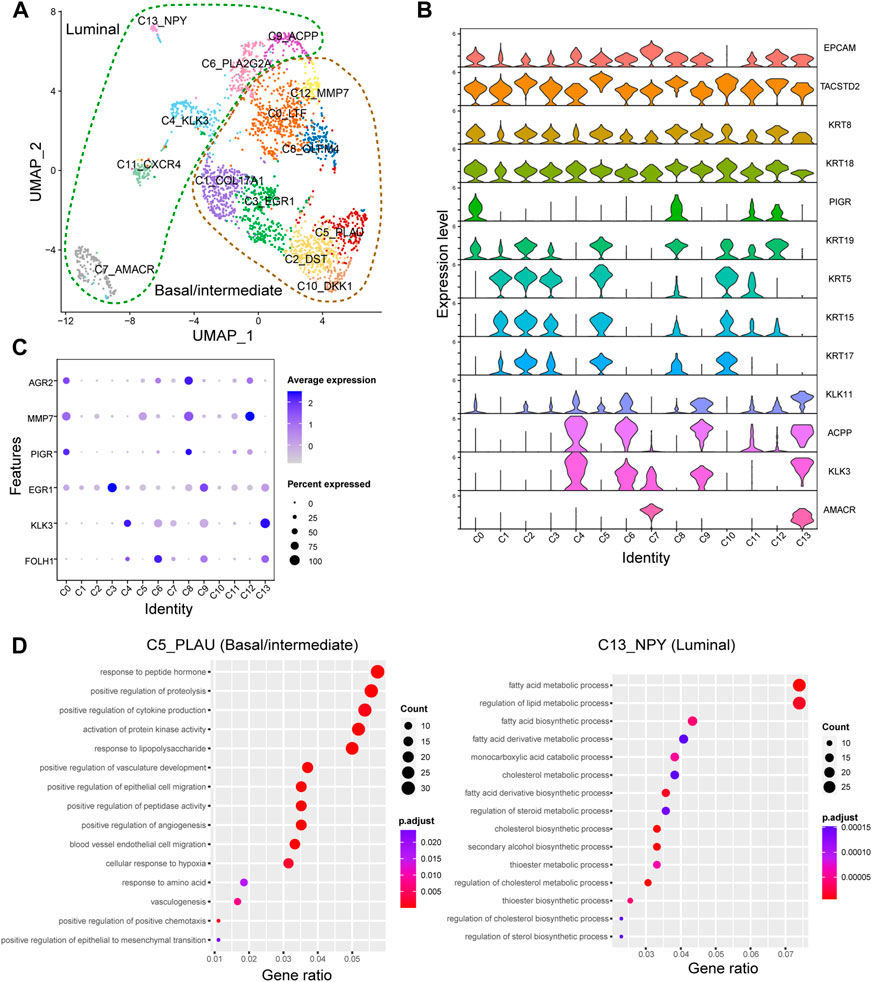
FIGURE 2. The epithelial cell type classifications in PNI-PCa tumor tissue. (A) Epithelial cells are divided into 13 different subgroups, UMAP of epithelial cells annotated by cell types. (B) Violin plots of marker genes across the epithelial cell subtypes. (C) The relative expression of FOLH1, KLK3, EGR1, PIGR, AGR2, and MMP7 genes in different epithelial subgroups. (D) GO enrichment analysis for DEGs in basal/intermediate cell subtype C5 and luminal cell subtype C13, respectively.
To identify the transcriptional states of epithelial cell subtypes, we used the edgeR package to examine gene expression patterns in these cells. The luminal cells, including C4, C6, C7, C9, and C13, showed significantly higher levels of expression of PCa markers genes FOLH1 and KLK3 (Figure 2C). Compared with other epithelial clusters, the luminal cell subtypes of C9 exhibited a higher expression level of EGR1. It has been reported that the expression level of EGR1 was increased in PCa and related to the occurrence, progression, and metastasis of PCa (Li et al., 2019). The human anterior gradient-2 (AGR2), a urinary marker for PCa, was highly expressed in C0, C8, and C12. It is worth noting that PIGR and MMP7 significantly high expressions were detected in the basal/intermittent C0 and C8. In previous studies, PIGR has demonstrated a role in promoting cellular transformation and proliferation. It has also been reported that MMP7 can promote PCa by inducing epithelial-mesenchymal transition (Yue et al., 2017; Zhang et al., 2017). These results suggested that basal/intermediate cell subtypes may be involved in tumor progression. Next, GO enrichment analysis was performed to examine further the potential roles of each epithelial cluster in PNI-PCa progression. We found that the genes upregulated in the luminal cell subtype of C13 were mainly related to metabolic processes involved in cancer progressions, including fatty acid metabolism, lipid metabolism, and steroid biosynthesis (Figure 2D), indicating that luminal cells were indeed malignant. Additionally, the genes upregulated in basal/intermediate cell subtypes C5, were associated with tumor migration processes such as epithelial-mesenchymal transition, vasculogenesis, and endothelium development (Figure 2D), indicating that basal/intermediate cell subtypes were associated with PNI-PCa progression.
To explore the relationship among these epithelial cell subtypes and identify the association between the genes associated with epithelial cell reprogramming, we pooled the epithelial cell subtypes and constructed a transcriptional trajectory of these cells on a pseudoprime scale. Analysis of the pseudotemporal trajectory presented two prominent paths for the epithelial cell subtypes (Figure 3A). The luminal cell subtypes, including C4, C6, C7, C9, and C13, were shown at the beginning of the trajectory (Figure 3B). The base/intermediate cells subtypes of C0, C8, and C12 appeared at the end of the trajectory branch 1, and branch 2 is composed of other basal/intermediate cells, including C1, C2, C3, C5, and C10 (Figure 3B). These results indicated that luminal epithelial cells could be the initiating cells and transition to based/intermediate cells.
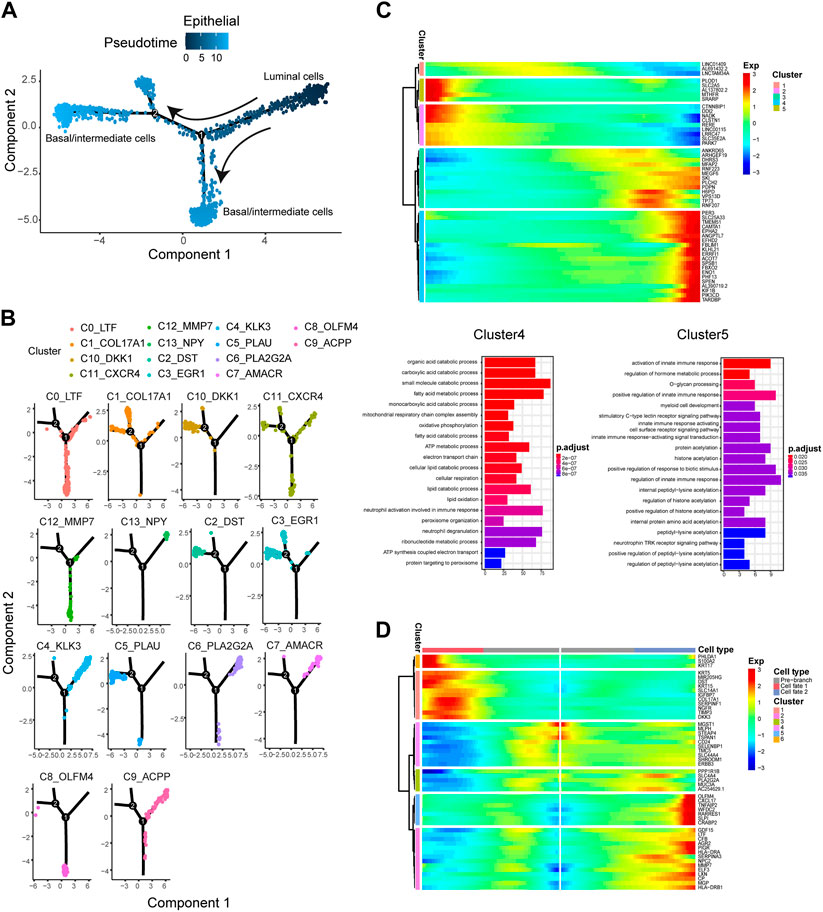
FIGURE 3. The pseudotime trajectory analysis of epithelial cells and identifying genes varied during the trajectory. (A) Differentiation trajectory of epithelial cells constructed by Monocle. (B) The epithelial cell subtypes differentiation trajectory plot. (C) Upper panel: The top 50 DEGs with the most significant expression level change over the pseudotime trajectory were classified into 5 clusters. Lower panel: GO molecular function enrichment heatmap of Cluster 4 and 5. (D) The heatmap shows the expression patterns of the top 50 significant genes in branched expression analysis modeling.
We then analyzed the changes in the dynamic expression of genes along the trajectory (Figure 3C). We identified some of the genes with the most significant changes in expression, including EPHA2, PDPN, SPSB1, and FBXO2, which have been reported to be related to multiple cancer metastasis or cell movement (Feng et al., 2014; Sun et al., 2018; Hu et al., 2020). In addition, from the initial stage of the pseudotime trajectory (luminal cell subtypes) to the terminal stage (basal/intermediate cell subtypes), we found some genes with pseudotemporal expression patterns clustered into five specific clusters. Go enrichment analysis was performed to reveal the different functions of these gene clusters. We identified genes cluster 4, which were highly expressed at the end stage of the pseudotime trajectory (Figure 3C) and mainly involved in PCa progression-related processes, including fatty acid metabolic process, organic acid catabolic process, and lipid catabolic process. Otherwise, the genes cluster 5 were highly expressed at the beginning stage of the pseudotime trajectory and enriched in activation of the innate immune response (Figure 3C).
Furthermore, BEAM analysis was performed to identify the cell cluster marker genes that change as epithelial cell subtypes pass from the luminal to the basal/intermediate stage. We performed a branched heatmap to show the expression dynamic of the top 50 significant genes in different cell fate branches (Figure 3D). From pre-branch (luminal cell subtypes) to cell fate 2 (Basal/intermediate cell subtypes including C1, C2, C3, C5, and C10), we found that two gene clusters (cluster 2 and cluster 5) were highly expressed in fate2. In the two gene clusters, some genes associated with tumorigenesis and cancer progression, such as AGR2, MMP7, LXN, PIGR, and CRABP2, were identified (Figures 3D). Additionally, from pre-branch (luminal cell subtypes) to cell fate 1 (basal/intermediate cell subtypes including C0, C8, and C12), gene cluster 1 and cluster 6 were highly expressed in fate1. We demonstrated that the expression level of KRT17, S100A2, and KRT15 was significantly higher in cluster 1 (Figure 3D). It had been reported that these genes were closely related to tumor progression and metastasis (Wang et al., 2019; Rao et al., 2020; Zhang et al., 2022).
Studies have shown that PNI in the tumor can contribute to the prognosis of PNI-PCa. Emerging evidence also suggested that the interaction between cells played a crucial role in tumor development. We hypothesized that the interactions between epithelial cells, especially basal/intermediate cells and neural cells, might facilitate the malignant growth of PNI-PCa. Based on this assumption, we characterized cell-cell communication mediated by ligand-receptor interactions between neural and epithelial cells in tumors using scRNA-seq data. Our results showed that there were significant interactions between epithelial cell subtypes and neural cells, and these subtypes of the epithelial cell were predominantly basal/intermediate cells (Figure 4A). The attraction strengths of ligand-receptor pairs between the epithelial and neural cells were calculated to infer molecular interactions mediating cell-cell interactions. Among the ligand-receptor pairs pertaining to epithelial and neural cells, CXCL1-ACKR1 and CXCL8-ACKR1 were significantly enriched in basal/intermediate C5, C8, and C12 (Figure 4B). As members of the CXC chemokine family, CXCL1 and CXCL8 have reported an association with the development of multiple cancers (Chen et al., 2020; Unver, 2021). In addition, another ligand-receptor pair involving interactions between epithelial cell subtypes and neural cell subsets were CCL2-CCR2 and CCL2-ACKR1, which mainly enriched in basal/intermediate C2 and C10 (Figure 4B). Compared with other epithelial cells, higher expression of CCL2 was detected in basal/intermediate C2, C3, and C10 (Figure 4C). It has been reported that the chemokine ligand CCL2 and its receptor ACKR1 were implicated in the initiation and metastasis in multiple cancers, which suggested the potential role of the interaction between basal/intermediate epithelial and neural cells in PNI-PCa progression.
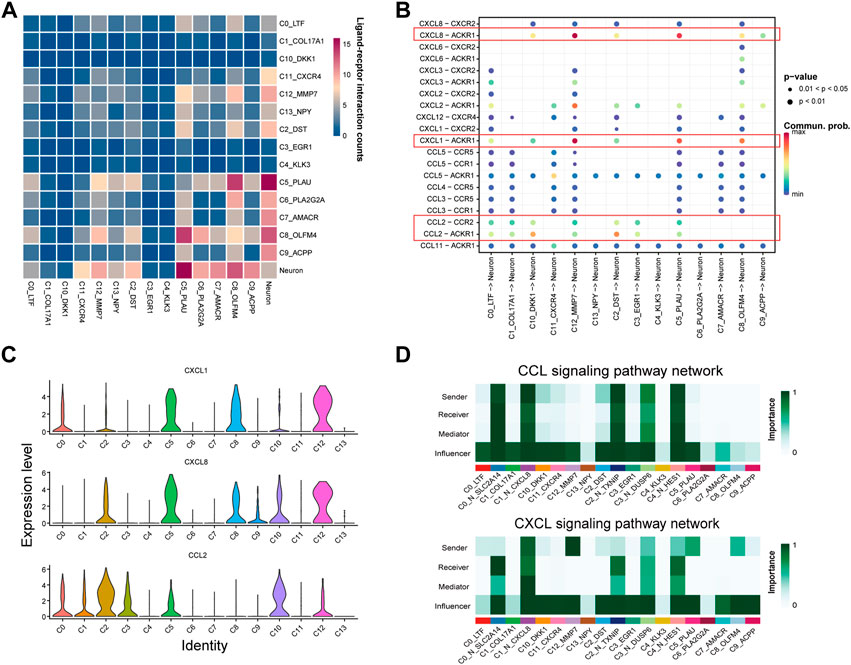
FIGURE 4. Cell interaction networks in PNI-PCa. (A) Heatmap shows the number of potential ligand-receptor pairs between epithelial cell subtypes and neural cell sells predicted by Cellphone DB. The more red the color, the stronger cell-cell communication. (B) Bubble plot of ligand-receptor interactions between epithelial cell subtypes and neural cells, where the bubble size represents the p-value (the p-value of all bubbles is less than .05), the bigger the bubble, the smaller the p-value. The color represents cell interaction strength, the redder the color, the stronger the interaction. (C) Bar chart showing the expression levels of CCL2, CXCL1, and CXCL8 in each epithelial cell subgroup. (D) The communication probability of neural and epithelial cells on the CXCL and CCL signaling pathways.
We further dissected CXCL and CCL signaling of communication probability between neural cell and epithelial cell subtypes. In the CXCL signaling network, basal/intermediate cell subtypes C5, C8, and C12 were the strong senders, all the neural cells were receivers, and the neural cell subtypes, including C0_N_SLC2A14 and C4_N_HES1, were most abundant (Figure 4D). For the CCL signaling network, basal/intermediate cell subtypes C2 and C10 were the major signal senders, and all the neural cell populations were the signal receivers (Figure 4D). Collectively, our results suggested that the cells communicate between basal/intermediate cell subtypes and neural cells through the CXCL and CCL signaling network, which may be one of the critical factors in promoting cancer progression and metastasis in PNI-PCa.
PNI is the process in which cancer cells penetrate along the neural commonly observed in PCa. In recent years, PNI has been confirmed to be associated with the progression and metastatic processes of PCa (Jia et al., 2017; Delahunt et al., 2020). Some studies also have revealed several molecular mechanisms of their influence on PCa progression at bulk tissue samples and cell populations level (Alajati et al., 2020; Pomerantz et al., 2020). However, the underlying molecular mechanisms affecting the progression of PNI-PCa at the single-cell level have been unclear. In our study, we performed scRNA-seq using tumor tissues obtained from PNI-PCa patients, and 12 cell types were identified, including two types of epithelial clusters and neural clusters. We found that basal/intermediate cell subtypes play an important role in PNI-PCa progression. In addition, the intercellular communication between basal/intermediate cells and neural cells via CCL and CXCL signalling pathways may be a possible molecular mechanism for PNI-PCa progression. Our results provide a more holistic interrogation of PNI-PCa progression at an unprecedented single-cell resolution. As far as we know, little has been reported about this aspect of the study. Several lines of evidence support that most prostate cancer originates from luminal cells (Chua et al., 2018; Guo et al., 2020). Consistent with previous studies, our results showed that prostate cancer-specific genes are mainly expressed in luminal cells. A previous study indicated that basal cells from human primary benign prostate tissue could cause PCa in immunodeficient mice (Goldstein et al., 2010), implying the potential roles of basal cells in tumorigenesis. In this study, we found that basal/intermediate cell populations were closely related to the progression of PNI-PCa. Compared to other luminal cell subtypes, basal/intermediate cell subtypes C8 has a higher expression level of AGR2, which was reported to be overexpressed in PCa compared to benign tissue and played an essential role in cancer spread (Takabatake et al., 2021; Maarouf et al., 2022). Furthermore, our results revealed that some genes associated with the migration and development of PCa (Chen et al., 2016; Yue et al., 2017), including MMP7 and PIGR, were highly expressed in basal/intermediate cell subtypes of C0 and C12. GO enrichment analysis also showed that pathways related to cancer progression were significantly enriched in basal/intermediate cells. These results suggested the pivotal roles that basal/intermediate cells play in PNI-PCa progression.
To further investigate this, we next performed pseudotime analysis on epithelial cells. The results demonstrated that luminal cells transform into basal/intermediate cells as cancer progresses. Notably, we identified that ENO1 and EPHA2 were highly expressed at the end of preudotime trajectorypse. EPHA2 is a member of the Eph family of receptor tyrosine kinases (RTK), expressed in most epithelial cells and associated with aggressive behavior in PCa(Kurose et al., 2019). In multiple malignancies, the expression of α-enolase (ENO1) is related to the progression and metastasis of the tumor (Hou et al., 2021; Li et al., 2021). Together, the results above demonstrated the effect of basal/intermediate epithelial cell subtypes on PNI-PCa progression.
PNI is a form of cancer progression where cancer cells invade along nerves. Prostate cancer cell interaction with nerves induces a neurogenesis response that promotes the initiation and metastasis of PCa (Magnon et al., 2013). The study of the PNI mechanism shows that ligand-receptor interactions between nerves and cancer cells can promote the process of PNI(Marchesi et al., 2010; Binmadi and Basile, 2011). In our results, we identified that basal/intermediate cell subtypes were the cells that communicate most closely with nerve cells through CCL2-ACKR1, CXCL1-ACKR1, and CXCL8-ACKR1 signaling. CCL2 is overexpressed in various cancers, including glioma, prostate, and breast (Tsaur et al., 2015; Yang et al., 2021; Zhong et al., 2021). Recent studies have implicated the role of chemokine ligand CCL2 and its receptor CCR2 in the progression of multiple cancers (Fang et al., 2021; Xu et al., 2021). Furthermore, some studies have found that CCL2, with its receptor CCR2, played a significant role in PNI in PCa and promoted the metastasis of PCa, using both animal models and a review of human surgical specimens (Ding et al., 2015; He et al., 2015). In our study, we also identified that CXCL1 and CXCL8 were significantly stronger expressed in basal/intermediate cells. It is well known that these chemokines are widely expressed on cancer cells and associated with malignant tumors. Several studies have demonstrated that CXCL1 and CXCL8, as well as their cognate receptors, can mediate tumor growth, proliferation, and metastasis of malignant cells, including PCa (Adekoya and Richardson, 2020; Łukaszewicz-Zając et al., 2020). This evidence suggested that communication between basal/intermediate cells and neural cells plays a critical role in promoting the development and progression of PNI-PCa.Notably, it has been reported that neural cells were related to resistance to antiandrogen in patients with PCa. Li et al. (2016) found that neuronal genes were highly expressed during prostate cancer progression, resulting in the development a neuronal phenotype of PCa. Moreover, their results also revealed that the amount of neuroendocrine cells increases during antiandrogen PCa, leading to therapy resistance. Although many existing studies have shown that the relationship between basal cells and PCa (Xin, 2019), the effects of basal cells on therapy resistance are still not fully understood. In a recent study, the basal cell gene-expression profile was found to be significantly enriched in castration-resistant and advanced prostate cancers (Zhang et al., 2016), which indicated basal cells might play a potential role in the development of resistance to anti-androgens in PCa. In a study published by Marhold et al. showed that basal cells were involved in primary resistance to androgen receptor inhibition in the TRAMP mouse model (Marhold et al., 2022). In addition, increasing evidence suggested that androgen signaling through the androgen receptor (AR) contributes to the promotion of castration-resistant prostate cancer (CRPC) (Zhou et al., 2017). Olar et al. (2014) found that increased perineural invasion diameter was associated with higher levels of AR in PC. These results seem to indicate a potential role of the PNI in the development of CRPC. However, more in-depth studies are required to elucidate these issues.
Although these studies reveal important discoveries, several limitations need to be considered. The small number of patients and samples is a limitation of this study. Another limitation of the current study is lacking of in vitro experiments to provide a deeper mechanistic investigation. Thus, further studies are required.
In conclusions, we systemically characterize the heterogeneity and gene expression landscapes of diverse cell types residing in PNI-PCa by using a single-cell RNA sequencing approach. Importantly, we revealed that the intercellular communication between basal/intermediate epithelial cell subtypes and neural cells through the ligand-receptor interaction pathway could affect the progression of PNI-PCa. These findings have important implications for our understanding of the mechanistic related to PNI-PCa progression and provided some new references for the therapeutic strategy for PNI-PCa.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: The raw sequencing data from this study were uploaded to the Genome Sequence Archive (GSA, https://bigd.big.ac.cn/gsa/); with accession number PRJCA013564.
The studies involving human participants were reviewed and approved by The Ethics Committee of Aerospace Center Hospital. The patients/participants provided their written informed consent to participate in this study.
BZ and BN conceived and designed the study; QG, LY, and ZL collected the patients’ samples and the clinical data; ZF and SW wrote the manuscript; YS, KL, SL, JC, JZ, JS, and JX analyzed the data; BZ, BN, SW, ML, and ZF revised the paper, All authors contributed to the article and approved the submitted version.
Authors ZF, SL, JZ, JS, JC, ML, and BN were employed by the company ChosenMed Technology (Beijing) Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2022.1073232/full#supplementary-material
SUPPLEMENTARY FIGURE S1 | (A–C) Pathological results revealed perineural invasion of the prostate cancer cells (hematoxylin and eosin staining) in three PCa patients.
SUPPLEMENTARY FIGURE S2 | Five subtypes generated from neural cells are demonstrated by UMAP.
Adekoya, T. O., and Richardson, R. M. (2020). Cytokines and chemokines as mediators of prostate cancer metastasis. Int. J. Mol. Sci. 21 (12), 4449. doi:10.3390/ijms21124449
Aibar, S., González-Blas, C. B., Moerman, T., Huynh-Thu, V. A., Imrichova, H., Hulselmans, G., et al. (2017). Scenic: Single-cell regulatory network inference and clustering. Nat. Methods 14 (11), 1083–1086. doi:10.1038/nmeth.4463
Alajati, A., D'Ambrosio, M., Troiani, M., Mosole, S., Pellegrini, L., Chen, J., et al. (2020). CDCP1 overexpression drives prostate cancer progression and can be targeted in vivo. J. Clin. Invest. 130 (5), 2435–2450. doi:10.1172/jci131133
Binmadi, N. O., and Basile, J. R. (2011). Perineural invasion in oral squamous cell carcinoma: A discussion of significance and review of the literature. Oral Oncol. 47 (11), 1005–1010. doi:10.1016/j.oraloncology.2011.08.002
Binmadi, N. O., Yang, Y. H., Zhou, H., Proia, P., Lin, Y. L., Batista De Paula, A. M., et al. (2012). Plexin-B1 and semaphorin 4D cooperate to promote perineural invasion in a RhoA/ROK-dependent manner. Am. J. Pathol. 180 (3), 1232–1242. doi:10.1016/j.ajpath.2011.12.009
Brabletz, T., Kalluri, R., Nieto, M. A., and Weinberg, R. A. (2018). EMT in cancer. Nat. Rev. Cancer 18 (2), 128–134. doi:10.1038/nrc.2017.118
Cancer Genome Atlas Network (2015). Genomic classification of cutaneous melanoma. Cell 161 (7), 1681–1696. doi:10.1016/j.cell.2015.05.044
Chen, H., Chen, P., Huang, J., Selegård, R., Platt, M., Palaniappan, A., et al. (2016). Detection of matrilysin activity using polypeptide functionalized reduced graphene oxide field-effect transistor sensor. Anal. Chem. 88 (6), 2994–2998. doi:10.1021/acs.analchem.5b04663
Chen, S., Zhou, Y., Chen, Y., and Gu, J. (2018). fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 34 (17), i884–i890. doi:10.1093/bioinformatics/bty560
Chen, S., Zhu, G., Yang, Y., Wang, F., Xiao, Y. T., Zhang, N., et al. (2021). Single-cell analysis reveals transcriptomic remodellings in distinct cell types that contribute to human prostate cancer progression. Nat. Cell Biol. 23 (1), 87–98. doi:10.1038/s41556-020-00613-6
Chen, X., Chen, R., Jin, R., and Huang, Z. (2020). The role of CXCL chemokine family in the development and progression of gastric cancer. Int. J. Clin. Exp. Pathol. 13 (3), 484–492.
Chua, C. W., Epsi, N. J., Leung, E. Y., Xuan, S., Lei, M., Li, B. I., et al. (2018). Differential requirements of androgen receptor in luminal progenitors during prostate regeneration and tumor initiation. Elife 7, e28768. doi:10.7554/eLife.28768
Deborde, S., and Wong, R. J. (2017). How Schwann cells facilitate cancer progression in nerves. Cell Mol. Life Sci. 74 (24), 4405–4420. doi:10.1007/s00018-017-2578-x
Delahunt, B., Murray, J. D., Steigler, A., Atkinson, C., Christie, D., Duchesne, G., et al. (2020). Perineural invasion by prostate adenocarcinoma in needle biopsies predicts bone metastasis: Ten year data from the TROG 03.04 RADAR Trial. Histopathology 77 (2), 284–292. doi:10.1111/his.14107
Ding, X., Yang, D. R., Lee, S. O., Chen, Y. L., Xia, L., Lin, S. J., et al. (2015). TR4 nuclear receptor promotes prostate cancer metastasis via upregulation of CCL2/CCR2 signaling. Int. J. Cancer 136 (4), 955–964. doi:10.1002/ijc.29049
Dudas, J., Ladanyi, A., Ingruber, J., Steinbichler, T. B., and Riechelmann, H. (2020). Epithelial to mesenchymal transition: A mechanism that fuels cancer radio/chemoresistance. Cells 9 (2), 428. doi:10.3390/cells9020428
Fang, W. B., Sofia Acevedo, D., Smart, C., Zinda, B., Alissa, N., Warren, K., et al. (2021). Expression of CCL2/CCR2 signaling proteins in breast carcinoma cells is associated with invasive progression. Sci. Rep. 11 (1), 8708. doi:10.1038/s41598-021-88229-0
Feng, Y., Pan, T. C., Pant, D. K., Chakrabarti, K. R., Alvarez, J. V., Ruth, J. R., et al. (2014). SPSB1 promotes breast cancer recurrence by potentiating c-MET signaling. Cancer Discov. 4 (7), 790–803. doi:10.1158/2159-8290.Cd-13-0548
Fromont, G., Godet, J., Pires, C., Yacoub, M., Dore, B., and Irani, J. (2012). Biological significance of perineural invasion (PNI) in prostate cancer. Prostate 72 (5), 542–548. doi:10.1002/pros.21456
Goldstein, A. S., Huang, J., Guo, C., Garraway, I. P., and Witte, O. N. (2010). Identification of a cell of origin for human prostate cancer. Science 329 (5991), 568–571. doi:10.1126/science.1189992
Guo, W., Li, L., He, J., Liu, Z., Han, M., Li, F., et al. (2020). Single-cell transcriptomics identifies a distinct luminal progenitor cell type in distal prostate invagination tips. Nat. Genet. 52 (9), 908–918. doi:10.1038/s41588-020-0642-1
He, M. X., Cuoco, M. S., Crowdis, J., Bosma-Moody, A., Zhang, Z., Bi, K., et al. (2021). Transcriptional mediators of treatment resistance in lethal prostate cancer. Nat. Med. 27 (3), 426–433. doi:10.1038/s41591-021-01244-6
He, S., He, S., Chen, C. H., Deborde, S., Bakst, R. L., Chernichenko, N., et al. (2015). The chemokine (CCL2-CCR2) signaling axis mediates perineural invasion. Mol. Cancer Res. 13 (2), 380–390. doi:10.1158/1541-7786.Mcr-14-0303
Hou, J. Y., Cao, J., Gao, L. J., Zhang, F. P., Shen, J., Zhou, L., et al. (2021). Upregulation of α enolase (ENO1) crotonylation in colorectal cancer and its promoting effect on cancer cell metastasis. Biochem. Biophys. Res. Commun. 578, 77–83. doi:10.1016/j.bbrc.2021.09.027
Hu, L., Zhang, P., Sun, W., Zhou, L., Chu, Q., and Chen, Y. (2020). PDPN is a prognostic biomarker and correlated with immune infiltrating in gastric cancer. Med. Baltim. 99 (19), e19957. doi:10.1097/md.0000000000019957
Jia, Z. W., Chang, K., Dai, B., Kong, Y. Y., Wang, Y., Qu, Y. Y., et al. (2017). Factors influencing biochemical recurrence in patients who have received salvage radiotherapy after radical prostatectomy: A systematic review and meta-analysis. Asian J. Androl. 19 (4), 493–499. doi:10.4103/1008-682x.179531
Kumar, M. P., Du, J., Lagoudas, G., Jiao, Y., Sawyer, A., Drummond, D. C., et al. (2018). Analysis of single-cell RNA-seq identifies cell-cell communication associated with tumor characteristics. Cell Rep. 25 (6), 1458–1468. e1454. doi:10.1016/j.celrep.2018.10.047
Kurose, H., Ueda, K., Kondo, R., Ogasawara, S., Kusano, H., Sanada, S., et al. (2019). Elevated expression of EPHA2 is associated with poor prognosis after radical prostatectomy in prostate cancer. Anticancer Res. 39 (11), 6249–6257. doi:10.21873/anticanres.13834
Li, H. J., Ke, F. Y., Lin, C. C., Lu, M. Y., Kuo, Y. H., Wang, Y. P., et al. (2021). ENO1 promotes lung cancer metastasis via HGFR and WNT signaling-driven epithelial-to-mesenchymal transition. Cancer Res. 81 (15), 4094–4109. doi:10.1158/0008-5472.Can-20-3543
Li, L., Ameri, A. H., Wang, S., Jansson, K. H., Casey, O. M., Yang, Q., et al. (2019). EGR1 regulates angiogenic and osteoclastogenic factors in prostate cancer and promotes metastasis. Oncogene 38 (35), 6241–6255. doi:10.1038/s41388-019-0873-8
Li, Q., Zhang, C. S., and Zhang, Y. (2016). Molecular aspects of prostate cancer with neuroendocrine differentiation. Chin. J. Cancer Res. 28 (1), 122–129. doi:10.3978/j.issn.1000-9604.2016.01.02
Lin, S. X., Zheng, Y., Wu, S., Blute, M. L., Dahl, D. M., and Wu, C. L. (2020). Impact of biopsy perineural invasion on younger prostate cancer patients after radical prostatectomy. Scand. J. Urol. 54 (6), 475–480. doi:10.1080/21681805.2020.1817143
Liss, M. A., Gordon, A., Morales, B., Osann, K., Skarecky, D., Lusch, A., et al. (2014). Urinary nerve growth factor as an oncologic biomarker for prostate cancer aggressiveness. Urol. Oncol. 32 (5), 714–719. doi:10.1016/j.urolonc.2014.01.018
Łukaszewicz-Zając, M., Pączek, S., Mroczko, P., and Kulczyńska-Przybik, A. (2020). The significance of CXCL1 and CXCL8 as well as their specific receptors in colorectal cancer. Cancer Manag. Res. 12, 8435–8443. doi:10.2147/cmar.S267176
Maarouf, A., Boissard, A., Henry, C., Leman, G., Coqueret, O., Guette, C., et al. (2022). Anterior gradient protein 2 is a marker of tumor aggressiveness in breast cancer and favors chemotherapy-induced senescence escape. Int. J. Oncol. 60 (1), 5. doi:10.3892/ijo.2021.5295
Magnon, C., Hall, S. J., Lin, J., Xue, X., Gerber, L., Freedland, S. J., et al. (2013). Autonomic nerve development contributes to prostate cancer progression. Science 341 (6142), 1236361. doi:10.1126/science.1236361
Marchesi, F., Piemonti, L., Mantovani, A., and Allavena, P. (2010). Molecular mechanisms of perineural invasion, a forgotten pathway of dissemination and metastasis. Cytokine Growth Factor Rev. 21 (1), 77–82. doi:10.1016/j.cytogfr.2009.11.001
Marhold, M., Udovica, S., Topakian, T., Horak, P., Horvat, R., Tomasich, E., et al. (2022). MALAT1 fusions and basal cells contribute to primary resistance against androgen receptor inhibition in TRAMP mice. Cancers (Basel) 14 (3), 749. doi:10.3390/cancers14030749
Olar, A., He, D., Florentin, D., Ding, Y., Wheeler, T., and Ayala, G. (2014). Biological correlates of prostate cancer perineural invasion diameter. Hum. Pathol. 45 (7), 1365–1369. doi:10.1016/j.humpath.2014.02.011
Pomerantz, M. M., Qiu, X., Zhu, Y., Takeda, D. Y., Pan, W., Baca, S. C., et al. (2020). Prostate cancer reactivates developmental epigenomic programs during metastatic progression. Nat. Genet. 52 (8), 790–799. doi:10.1038/s41588-020-0664-8
Rao, X., Wang, J., Song, H. M., Deng, B., and Li, J. G. (2020). KRT15 overexpression predicts poor prognosis in colorectal cancer. Neoplasma 67 (2), 410–414. doi:10.4149/neo_2019_190531N475
Satija, R., Farrell, J. A., Gennert, D., Schier, A. F., and Regev, A. (2015). Spatial reconstruction of single-cell gene expression data. Nat. Biotechnol. 33 (5), 495–502. doi:10.1038/nbt.3192
Siegel, R. L., Miller, K. D., and Jemal, A. (2020). Cancer statistics, 2020. CA Cancer J. Clin. 70 (1), 7–30. doi:10.3322/caac.21590
Sun, X., Wang, T., Guan, Z. R., Zhang, C., Chen, Y., Jin, J., et al. (2018). FBXO2, a novel marker for metastasis in human gastric cancer. Biochem. Biophys. Res. Commun. 495 (3), 2158–2164. doi:10.1016/j.bbrc.2017.12.097
Takabatake, K., Konishi, H., Arita, T., Kataoka, S., Shibamoto, J., Furuke, H., et al. (2021). Anterior gradient 2 regulates cancer progression in TP53-wild-type esophageal squamous cell carcinoma. Oncol. Rep. 46 (6), 260. doi:10.3892/or.2021.8211
Trapnell, C., Cacchiarelli, D., Grimsby, J., Pokharel, P., Li, S., Morse, M., et al. (2014). The dynamics and regulators of cell fate decisions are revealed by pseudotemporal ordering of single cells. Nat. Biotechnol. 32 (4), 381–386. doi:10.1038/nbt.2859
Tsaur, I., Noack, A., Makarevic, J., Oppermann, E., Waaga-Gasser, A. M., Gasser, M., et al. (2015). CCL2 chemokine as a potential biomarker for prostate cancer: A pilot study. Cancer Res. Treat. 47 (2), 306–312. doi:10.4143/crt.2014.015
Unver, N. (2021). Identification of the dominant angiogenic CXCL class chemokines associated with non-small cell lung cancer via bioinformatics tools. Med. Oncol. 38 (6), 68. doi:10.1007/s12032-021-01517-7
Wang, Z., Yang, M. Q., Lei, L., Fei, L. R., Zheng, Y. W., Huang, W. J., et al. (2019). Overexpression of KRT17 promotes proliferation and invasion of non-small cell lung cancer and indicates poor prognosis. Cancer Manag. Res. 11, 7485–7497. doi:10.2147/CMAR.S218926
Wu, S., Lin, X., Lin, S. X., Lu, M., Deng, T., Wang, Z., et al. (2019). Impact of biopsy perineural invasion on the outcomes of patients who underwent radical prostatectomy: A systematic review and meta-analysis. Scand. J. Urol. 53 (5), 287–294. doi:10.1080/21681805.2019.1643913
Xin, L. (2019). Cells of origin for prostate cancer. Adv. Exp. Med. Biol. 1210, 67–86. doi:10.1007/978-3-030-32656-2_4
Xu, M., Wang, Y., Xia, R., Wei, Y., and Wei, X. (2021). Role of the CCL2-CCR2 signalling axis in cancer: Mechanisms and therapeutic targeting. Cell Prolif. 54 (10), e13115. doi:10.1111/cpr.13115
Yang, Q., Zhang, J., Zhang, X., Miao, L., Zhang, W., Jiang, Z., et al. (2021). C-C motif chemokine ligand 2/C-C receptor 2 is associated with glioma recurrence and poor survival. Exp. Ther. Med. 21 (6), 564. doi:10.3892/etm.2021.9996
Yin, L., Li, J., Wang, J., Pu, T., Wei, J., Li, Q., et al. (2021). MAOA promotes prostate cancer cell perineural invasion through SEMA3C/PlexinA2/NRP1-cMET signaling. Oncogene 40 (7), 1362–1374. doi:10.1038/s41388-020-01615-2
Yue, X., Ai, J., Xu, Y., Chen, Y., Huang, M., Yang, X., et al. (2017). Polymeric immunoglobulin receptor promotes tumor growth in hepatocellular carcinoma. Hepatology 65 (6), 1948–1962. doi:10.1002/hep.29036
Zhang, D., Park, D., Zhong, Y., Lu, Y., Rycaj, K., Gong, S., et al. (2016). Stem cell and neurogenic gene-expression profiles link prostate basal cells to aggressive prostate cancer. Nat. Commun. 7, 10798. doi:10.1038/ncomms10798
Zhang, L. J., Wu, B., Zha, Z. L., Qu, W., Zhao, H., Yuan, J., et al. (2018). Perineural invasion as an independent predictor of biochemical recurrence in prostate cancer following radical prostatectomy or radiotherapy: A systematic review and meta-analysis. BMC Urol. 18 (1), 5. doi:10.1186/s12894-018-0319-6
Zhang, Q., Liu, S., Parajuli, K. R., Zhang, W., Zhang, K., Mo, Z., et al. (2017). Interleukin-17 promotes prostate cancer via MMP7-induced epithelial-to-mesenchymal transition. Oncogene 36 (5), 687–699. doi:10.1038/onc.2016.240
Zhang, Q., Xia, T., Qi, C., Du, J., and Ye, C. (2022). High expression of S100A2 predicts poor prognosis in patients with endometrial carcinoma. BMC Cancer 22 (1), 77. doi:10.1186/s12885-022-09180-5
Zhang, S., Qi, L., Li, M., Zhang, D., Xu, S., Wang, N., et al. (2008). Chemokine CXCL12 and its receptor CXCR4 expression are associated with perineural invasion of prostate cancer. J. Exp. Clin. Cancer Res. 27 (1), 62. doi:10.1186/1756-9966-27-62
Zhang, X., Lan, Y., Xu, J., Quan, F., Zhao, E., Deng, C., et al. (2019). CellMarker: A manually curated resource of cell markers in human and mouse. Nucleic Acids Res. 47 (D1), D721–D728. doi:10.1093/nar/gky900
Zhong, X., Wang, X., and Sun, Q. (2021). CCL2/ACKR2 interaction participate in breast cancer metastasis especially in patients with altered lipid metabolism. Med. Hypotheses 158, 110734. doi:10.1016/j.mehy.2021.110734
Keywords: prostate cancer with perineural invasion, intratumor heterogeneity, single-cell RNA sequencing, epithelial basal/intermediate cells, neural cells, progression, intercellular communication
Citation: Zhang B, Wang S, Fu Z, Gao Q, Yang L, Lei Z, Shi Y, Le K, Xiong J, Liu S, Zhang J, Su J, Chen J, Liu M and Niu B (2023) Single-cell RNA sequencing reveals intratumoral heterogeneity and potential mechanisms of malignant progression in prostate cancer with perineural invasion. Front. Genet. 13:1073232. doi: 10.3389/fgene.2022.1073232
Received: 18 October 2022; Accepted: 23 December 2022;
Published: 09 January 2023.
Edited by:
Ata Abbas, Case Western Reserve University, United StatesReviewed by:
Babu Roshan Padmanabhan, University Hospitals Cleveland Medical Center, United StatesCopyright © 2023 Zhang, Wang, Fu, Gao, Yang, Lei, Shi, Le, Xiong, Liu, Zhang, Su, Chen, Liu and Niu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bao Zhang, YmFvenRqQHNpbmEuY29t; Beifang Niu, bml1YmZAY25pYy5jbg==
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.