 Chandan Roy
Chandan Roy Sudhir Kumar
Sudhir Kumar Rakesh Deo Ranjan2
Rakesh Deo Ranjan2 Velu Govindan
Velu Govindan
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Genet. , 08 November 2022
Sec. Genomics of Plants and the Phytoecosystem
Volume 13 - 2022 | https://doi.org/10.3389/fgene.2022.1045955
This article is part of the Research Topic Functional Genomic Approaches in Molecular Breeding for Crop Improvement View all 7 articles
More than three billion people worldwide suffer from iron deficiency associated anemia and an equal number people suffer from zinc deficiency. These conditions are more prevalent in Sub-Saharan Africa and South Asia. In developing countries, children under the age of five with stunted growth and pregnant or lactating women were found to be at high risk of zinc and iron deficiencies. Biofortification, defined as breeding to develop varieties of staple food crops whose grain contains higher levels of micronutrients such as iron and zinc, are one of the most promising, cost-effective and sustainable ways to improve the health in resource-poor households, particularly in rural areas where families consume some part of what they grow. Biofortification through conventional breeding in wheat, particularly for grain zinc and iron, have made significant contributions, transferring important genes and quantitative trait loci (QTLs) from wild and related species into cultivated wheat. Nonetheless, the quantitative, genetically complex nature of iron and zinc levels in wheat grain limits progress through conventional breeding, making it difficult to attain genetic gain both for yield and grain mineral concentrations. Wheat biofortification can be achieved by enhancing mineral uptake, source-to-sink translocation of minerals and their deposition into grains, and the bioavailability of the minerals. A number of QTLs with major and minor effects for those traits have been detected in wheat; introducing the most effective into breeding lines will increase grain zinc and iron concentrations. New approaches to achieve this include marker assisted selection and genomic selection. Faster breeding approaches need to be combined to simultaneously increase grain mineral content and yield in wheat breeding lines.
Minerals are important components for physical and mental health development in humans. Malnutrition from micronutrient deficiencies, also known as “hidden hunger,” is one of the most challenging health issues globally and particularly in developing countries. Dietary deficiencies of zinc (Zn), iron (Fe), iodine, and vitamin A are most prevalent among children and women. Worldwide, more than three billion people are affected by zinc (Zn), iron (Fe), and vitamin-A deficiencies (Raemaekers, 1988; Beal et al., 2017), with such nutrient deficiencies being particularly high among people in Asia and Sub-Saharan Africa. Fe is an essential part of hemoglobin and myoglobin and being directly involved in oxygen transport, enzymatic functions, energy production and DNA synthesis. Fe deficiency causes anemia (https://www.who.int/health-topics/anaemia); children below 5 years of age (40%) and women at reproductive or lactating stages (30%) are more anemic. Likewise, Zn is a ubiquitous element for every living organism including human beings, acting as a co-factor for more than 300 enzymes and proteins at the cellular and sub-cellular levels during nucleic acid production, metabolism, cell division and differentiation, and the immune system. Zn deficiency impairs physical growth and development and the proper functioning of the immune and reproductive systems and mental acuity, as well as increasing child mortality. On average 20% of women and children are Zn deficient, with a high prevalence in low- and middle-income groups and even among adult men (Gupta et al., 2020b). Another study reported 31.3% Zn deficiency among the children of higher income groups in western Europe and no significant difference was observed between children of the different socio-economic groups (Vreugdenhil et al., 2021). Food supplements and fortification and more diverse diets can help to address micronutrient malnutrition. Iron and zinc supplements, for example, have substantially reduced diarrhea and anaemia in affected segments of the population (Gupta et al., 2020b). Food fortification for vitamin-A and iodine deficiency through pharmaceutical products and iodized salt have reduced related deficiencies, but such products may be unaffordable for low-income families, particularly in developing countries. Reduced consumption of vegetables and fruits is observed due to lower purchasing power in many farm households in Asia and Sub-Saharan Africa; now the COVID-19 outbreak substantially reduced the income of poor households that cut down on their intake of fruits, vegetables, and animal-based foods and increased their dependence on staple grains (Heck et al., 2020). In 2022, restrictions in the movement of farm produce, disrupted supply systems, and continued economic fall-out due to the COVID-19 pandemic and Russia-Ukraine conflict jeopardized food supply chains, including distribution to Central Asia and Africa, and triggered food prices, augmenting global hunger and malnutrition (https://www.worldbank.org/en/topic/agriculture/brief/food-security-update). Increasing nutrient density and bioavailability in staple food crops, as well as their production and distribution at affordable prices, can greatly improve the nutrition of low-income groups.
“Biofortification” refers to increasing essential mineral and vitamin content and bioavailability in edible parts of staple food crops, either through conventional breeding and/or biotechnological interventions, as well as through fertilizer. “Golden rice” is a successful example of improving the beta-carotene content in rice through the transformation of three bio-synthetic pathway genes: phytoene synthase (psy), phytoene desaturase (crtI) and lycopene β-cyclase (lcy) (Ye et al., 2000). Biofortification in staple food crops has progressed through initiatives such as Harvest Plus, the Grand Challenge in Global Health, the India Biofortification Programme, Scaling Up Nutrition (SUN), and Global Alliance for Improved Nutrition (GAIN), among others, and has gained global and local recognition. Numerous countries (Bangladesh, Brazil, China, Colombia, India, Indonesia, Malawi, Nigeria, Pakistan, Panama, Rwanda, Uganda, and Zambia) have included biofortification in their national health and development policies (Virk et al., 2021). Efforts also have been made to speed the seed production and distribution of biofortified crop varieties to reach resource-poor farmers in remote areas. The government of India has proposed the inclusion of fortified wheat, rice, edible oil in mid-day meal programs, the public distribution system and integrated child development program (https://www.livemint.com/Politics/91RAsPJFJLykDywTPHRCeI/Fortified-MidDay-meals-by-December-2019-to-fight-malnutriti.html).
Wheat provides 25% of calories in human diets worldwide and 60% in Central and West Asian countries (Cakmak, 2008). The wheat endosperm is rich in starch but poor in minerals, particularly Zn, Fe, and vitamins (Cakmak and Kutman, 2018). Moreover, minerals and bioactive components of the wheat grain are concentrated in the aleurone layer, which is removed during milling, so the remaining flour generally contains only small amounts of minerals and vitamins. The average Zn concentration in the white flour is around 8 mg per kg, falling short of the daily recommended dietary allowance (RDA) of from 9–19 mg per day for adults (Cakmak, 2007; Gillies et al., 2012; Cakmak and Kutman, 2018). The low mineral contents in rice and wheat is due to the deficiencies of these substances, particularly of Zn, in the soils of most of the areas where the crops are grown. Finally, breeding for high yielding wheat genotypes since the mid-20th century has reduced genetic variation for grain Zn and Fe in modern cultivars from the levels found in landraces and genotypes released during Green Revolution (Debnath et al., 2021).
Through agronomic and genetic interventions, mineral density can be enhanced (Cakmak, 2008). Genetic biofortification is the most sustainable and cost-effective approach to alleviate micronutrient malnutrition among lower-middle income consumers. Based on estimated average micronutrient requirements; average staple food crop intakes; mineral losses incurred during harvesting, processing, and storage; and micronutrient bioavailablility, a minimum target levels have been set for different minerals in staple food crops (Bouis et al., 2011). To meet out 30 and 40% of estimated average requirements for Fe and Zn, respectively for children and adult women, the target levels of 59 ppm for Fe and 38 ppm for Zn are set in wheat grain, considering the baseline of 30 ppm for Fe and 24 ppm for Zn. Large-scale screening of germplasm, identification of genotypes, mapping of underpinning genomic regions, and their use to develop high-yielding, disease resistant, biofortified wheat genotypes are the major focuses of genetic biofortification research. Wild relatives carry higher levels of Zn and Fe in their grain than cultivated wheat genotypes. The locus Gpc-B1 for grain protein, Zn, and Fe was detected and fine mapped in Triticum durum ssp dicoccoides and can serve as a source to enhance these traits in cultivated wheat (Uauy et al., 2006). Genome sequence information of T. aestivum cv. Chinese Spring (Mayer et al., 2014) and a highly annotated chromosome level genome sequence (Ref Seqv1.0; Appels et al., 2018) can be used for gene discovery, cloning, and functional analysis of genome targeting to grain Zn and Fe. In this article we review the status of wheat biofortification and how genomic resources can be used in breeding for biofortified wheat genotypes.
Initial research on agronomic biofortification to increase grain Zn in wheat took place in Turkey (Cakmak et al., 1996; Cakmak et al., 1999). Mineral density can be modulated by soil Zn and Fe status and their availability for the crop. Soil zinc and iron availability depends on edaphic factors such as soil physical and chemical properties, micronutrient availability, pH, the status of soil moisture and organic matter, grain filling duration, and the timing of senescence (Cakmak and Kutman, 2018). Irrespective of this, wheat grain zinc and iron concentrations can be increased by applying these elements directly to wheat (Yilmaz et al., 1997). Zn and Fe can be applied in inorganic or organic forms but the most commonly used are ZnSO4 and FeSO4, due to their ready availability and low cost. Soil and foliar applications of zinc fertilizer increase grain Zn density in wheat and can positively affect agronomic parameters such as the number of tillers, thousand kernel weight, chlorophyll content, and biological yield (Rizwan et al., 2018; Dimkpa et al., 2020). Hussain et al. (2012b) reported that soil applications of Zn increased whole grain zinc concentrations up to 95% and grain yield by 29%. However, foliar application was found to be more effective than soil application to increase zinc and iron concentrations in wheat. Foliar application is also advantageous under stress conditions, particularly drought, to avoid impairment of mineral uptake due to lack of soil moisture. Foliar applications of ZnSO4 @ 0.5% and FeSO4 @ 1% were more effective than applying to soil alone or both soil and foliar applications (Ramzan et al., 2020), and foliar application during grain filling resulted in a more effective translocation of zinc to the grain. Spraying zinc fertilizer at the grain development stage improved grain zinc concentration by 68% (Zhang et al., 2010). Unlike for zinc, iron fertilization of soils is not effective at increasing grain iron concentrations (Gupta, 1991), but in several studies the foliar application of iron increased grain Fe concentration up to 28% (Zhang et al., 2010); 21% (Pahlavan-Rad and Pessarakli, 2009), and 14% (Aciksoz et al., 2011b). Application of N-fertilizers along with FeSO4 increased iron concentrations in shoots as well as in the grain (Aciksoz et al., 2011b; Singh et al., 2018). Previous reports showed that N application increased the secretion of phytosederophore that chelate Fe in the soil and Yellow Stripe 1 protein act as Fe transporter (Aciksoz et al., 2011a). Seed priming is another way to deliver minerals to plants. Zinc also improves coleoptile and radical growth and germination in wheat (Ozturk et al., 2006). The recent practice of applying nano particles (NP) to the soil is becoming popular as a cost effective, environmentally-friendly approach to reduce fertilizer losses and raise productivity and profitability. Applying ZnO-NP and Fe-NP in wheat enhanced grain yield, biomass, chlorophyll content, and drought tolerance (Dimkpa et al., 2020). Nano zinc and iron treatments significantly increased Zn and Fe concentration in the roots, shoots, and grain (Rizwan et al., 2018; Dimkpa et al., 2020). Applying ZnO-NP or Fe-NP alone or coated with NPK can reduce farmers’ costs and, in case of zinc, it is cheaper than ZnSO4.
Whereas, foliar and soil applications of Zn and Fe must be repeated in each crop cycle, genetic biofortification provides a long-term solution against micronutrient malnutrition. Genetic biofortification is complex, involving several physiological pathways and proteins right from soil uptake of minerals to their accumulation in the grain. Targeting the genes or proteins that play key roles in mineral sequestration may increase grain Zn and Fe. The two major approaches that may be used are selective plant breeding and transgenic breeding, to develop biofortified crop genotypes. Transgenic approaches and outputs are subject to stringent policies in many countries and their use is restricted or prohibited, so this paper focuses on selective breeding. Mainstreaming biofortification as a selection priority in breeding programs—along with yield, disease resistance, end-use preferences, and environmental adaption—is paramount, and includes the use of micronutrient dense parental lines and setting grain micronutrient target levels. The review consists of three major sections: the first section deals with physiological and molecular mechanism of uptake, translocation and grain sequestration of Zn and Fe; second section deals with breeding for grain zinc and iron including mapping of gene(s) and QTLs; and the third section deals with new breeding techniques which need to be adopted with conventional breeding.
Complex physiological and metabolic processes are associated with the uptake and accumulation of Zn and Fe into the grain. Broadly, there are three kinds of mineral uptake mechanisms: 1) direct uptake of Zn2+ and Fe2+ molecules via the root system with the help of ZRT-IRT like proteins; 2) where Zn2+ and Fe3+ molecules are chelated first and then taken up through roots; and 3) a combination of 1 and 2. Cereals and millets (rice, maize, wheat, barley, pearl millet, among others) use mechanism 2 to extract Fe and Zn from the rhizosphere into the root system, where secretion of phytosiderophores (PS), proteins of the Mugineic acid (MA) family, help in chelating Fe and Zn and form a complex of Fe3+-MA or Zn2+-MA (Marschner and Romheld, 1994; Nozoye et al., 2015). The role of MA is well understood in Fe uptake and transport in rice and barley, but also showed its affinity towards molecules like Zn and Cu. In zinc-deficient wheat plants, more MA was released (Cakmak et al., 1994). Methionine acts as a precursor of MA synthesis, which is converted to S-adenosyl-L-methionine (SAM) with the help of the enzyme SAM synthetase (Mori and Nishizawa, 1987). In a subsequent reaction, SAM is converted to nicotianamine (NA) by NA synthase (NAS) followed by 3″-keto acid and 2′-deoxymugineic acid (DMA) by NA aminotransferase (NAAT) and DMA synthase (DMAS), respectively (Shojima et al., 1990). In some species, DMA is further converted to MA. In graminaceous crops MA is commonly produced, whereas NA is most common in non-graminaceous crops. In wheat, MA producing genes such as TaNAS, TaNAAT, and TaDMAS were identified (Pearce et al., 2014). MA secreted in the root zone chelates iron along with zinc, manganese (Mn), and copper (Cu). The complexes of Fe3+-MA or Zn2+-MA are taken up by roots. This mechanism is well established for Fe uptake in rice, barley and maize, but little is known about the genetic mechanism of metal uptake and its transport in wheat. Using the knowledge of model crop species, it is possible to identify the candidate genes in wheat of potential use for breeding.
The gene family “transfer of MAs” (TOM) plays important role in the uptake of soil iron and its translocation. TOM1 was first identified in rice and barley as facilitating MA secretions from roots to soil (Nozoye et al., 2011) and their transport. TOM2 and TOM3 are homologous to TOM1 and transport PS into the plant. Over expression of TOM2 using a GUS-promoter in rice allowed scientists to locate it in roots, shoots and seeds, with the highest expression in basal plant tissues (Nozoye et al., 2015). Several zinc-induced facilitator (ZIFL) genes have been identified and their role in zinc homeostasis assessed. ZIFL acts as an efflux transporter of vacuolar NA. In rice, ZIFL transports MA synthesized in the roots. In wheat, 15 TaZIFL proteins distributed on chromosomes 3, 4, and 5 have been characterized; genes like TaZIFL2.3, TaZIFL4.1, TaZIFL4.2, TaZIFL5, TaZIFL6.1, and TaZIFL6.2, were upregulated in roots under zinc and Fe deficient conditions (Sharma et al., 2019).
Other than MA, nicotianamine is produced by roots that also help in chelating soil Fe and Zn. NAS increases the production of NA and DMA, which improves the uptake and translocation of chelated metals. Overexpression of rice nicotianamine synthase genes, the OsNAS2 gene in wheat, increases the uptake and translocation of grain zinc in shoots as well as the grain (Singh et al., 2017).
A complex mechanism of metal transport, chelation and sequestration helps plants to avoid metal toxicity. Plants produce several transporter proteins varying in their substrate, expression, and locations. Colangelo and Guerinot (2006) provide a detailed discussion of the P1B-ATPase family, the CDF family as metal efflux proteins, ZIP, yellow stripe-1 (YSL), the natural resistance associated macrophage protein (NRAMP), and the copper uptake proteins (COPT) as metal uptake proteins. Heavy metal transporting P type ATPase (HMA), a protein of the P1B-ATPase family, act as a Zn, Cd, and Pb transporter. Arabidopsis mutants of hma2 and hma4 showed lower Zn uptake than the wild type (Hussain et al., 2004). HMA helps Zn to move from roots into shoots and plays a role in xylem loading and unloading. In wheat, 32 HMA protein-producing genes were detected on chromosomes 2A, 2B, 2D, 4A, 4D, 5A, 5B, 5D, 6A, 6B, 6D, 7A, 7B, and 7D (Zhou et al., 2019). Overexpression of the TaHMA2 gene in wheat increased the Zn concentration in the shoot as well as in the grain; however, grain Zn concentration was limited to the embryo and aleurone layer (Tan et al., 2013). Similarly, metal tolerance protein (MTP) is another transmembrane metal transporter protein of the CDF family located in the vacuole membrane of roots, shoots, and leaves and helps in Zn transport into the vacuole. Knocking out the mtp1 gene in Arabidopsis reduced Zn accumulation in various plant tissues (Desbrosses-Fonrouge et al., 2005). NRAMP is another protein located in the vacuole membrane and which takes active part in Fe transport. Eight homologues of TaNRAMP genes have been identified in wheat (Borrill et al., 2014). Other genes known as vacuolar iron transporter (VITs) genes help transport of iron into the grain and offer a potential target for iron biofortification. Wheat has two functional VIT genes, TaVIT1 and TaVIT2 on chromosomes 2 and 5, each with three homeologs from the A, B, and D genome. Overexpression of TaVIT2 increased grain iron 2-fold (Connorton et al., 2017).
Several proteins belong to the yellow stripe (YSL) family, a complex of transporter proteins that help in the uptake of chelated metals in the soil into roots and their subsequent transport to stems, leaves, and grain. First detected in maize, YSL-1 is involved in the uptake of metals that form complexes with PS or NA and its role in Fe3+-MA transport has been well established (Curie et al., 2001). Rice genes OsYSL15 express in the seed and are involved in seed germination, while OsYSL2 plays an important role in phloem transport and transport into the seed (Nozoye et al., 2015). Likewise, zinc-induced transporter family (ZIP) proteins are involved in metal uptake and transport. In wheat, 14 and 19 members of YSL and ZIP gene families have been characterized in up and down regulation during senescence and which play key roles in metal transport from the cytoplasm to the phloem and the phloem to the grain (Pearce et al., 2014). Several YSL, ZIP, and NRAMP homologs are involved in the uptake, transport, and remobilization of Fe and Zn in wheat (Table 1). Overexpression of the specific transporter protein using a tissue-specific promoter could be a target area for research to increase the grain mineral concentrations.

TABLE 1. Summary of genes involved in uptake, transport and grain accumulation of Zn and Fe in wheat.
Minerals deposition in grain occurs through the direct uptake of Zn/Fe by roots at grain filling stage, or remobilization of stored Zn/Fe from leaves and stems. Studies in rice to understand the contribution through continued root uptake and remobilization of stored minerals into the grain observed that minerals are accumulated via both processes. Under conditions of adequate minerals in the soil, continued root uptake plays a major role in Zn/Fe deposition into the grain while, under mineral deficient conditions, remobilization of stored Zn/Fe from leaves and stems contributes more to rice grain mineral content (Sperotto, 2013). A similar result was observed for grain Zn accumulation in winter wheat by Liu et al. (2019a), also reported a critical value for soil Zn availability (7.15 mg kg−1 DTPA Zn concentration) for wheat, above which the direct uptake of soil Zn during grain filling is predominant and below which remobilization is the major source of Zn in the grain. Remobilization of Zn/Fe depends on the xylem to phloem loading and mobility. In rice, grain Zn concentrations depend on plants’ ability to redistribute Zn from older leaves and stems, as well as phloem mobilization of Zn (Wu et al., 2010). Significant contributions of the flag and penultimate leaves to wheat grain yield have been observed (Roy et al., 2021a), but there are no detailed studies concerning the proportionate remobilization of Zn and Fe from different wheat plant parts. Uauy et al. (2006) observed an abundance of Zn, Fe and proteins in wheat flag leaves during grain filling, in plants carrying the Gpc-B1 locus.
Micro elemental analysis revealed higher concentrations of Zn, Fe and other bioactive elements in the embryo and grain aleurone layer; achieving localized increases of mineral concentrations in the endosperm is challenging. To better understand mineral translocation and deposition in the grain, experiments in model crop plants identified key genes. Ferritin is a Fe storage protein located in the plastid and readily bio-available. Enhancing FERRITIN gene expression is important for Fe-biofortification. Genetically transforming rice using the soybean FERRITIN gene under the seed-specific promoter gene Glu-B1 (Goto et al., 1999) and the pea FERRITIN gene under the Gt-1 promoter (Lucca et al., 1999) increased endosperm iron levels. The soybean FERRITIN gene under the control of the maize ubiquitin promoter was expressed in wheat, resulting in higher iron concentrations in both plant tissue and the grain, but with much higher levels in the former (Drakakaki et al., 2000). Wheat carries two ferritin genes, TaFer1 and TaFer2, located in chromosomes 4 and 5 and each with three homoalleles in hexaploid wheat. Overexpression of the endogenous TaFer1-A gene targeting the endosperm raised wheat grain Fe levels by 50%–58% (Borg et al., 2012).
The endogenous vacuolar transporter (TaVIT2) gene using a promoter of the GLU-1D-1 gene achieved a more than 2-fold increase in Fe concentrations in white flour (Connorton et al., 2017). In another attempt, the constitutive expression in bread wheat of the rice OsNAS gene that produces chelators such as nicotianamine (NA) and 2′-deoxymugineic acid (DMA) improved grain Fe and Zn concentrations (Beasley et al., 2019).
Effective genetic biofortification approaches for wheat should focus on increasing mineral uptake from the soil and translocation and remobilization into grains, combining genes for higher metal uptake and translocation (NAS, NAAT, DMAS, TOM, YSL, ZIP etc.) and genes that affect targeted transport to the endosperm (VIT, FERITTIN etc.). Singh et al. (2017) developed wheat transgenic lines using the rice OsNAS2 and bean PvFERRITIN genes separately and combining both genes and observed increased grain Zn and Fe concentrations in all three cases. The OsNAS2 gene was most effective; a grain of related transgenic wheat lines contained 93.1 ppm of Fe and 140.6 ppm of Zn (Singh et al., 2017) and there was a more than 2-fold increase in grain Fe, compared to a 1.6-fold increase with the combined expression of both genes. This differs from the case for rice, where the combined expression of OsNAS2 and FERRITIN gave a 6-fold increase in grain Fe content (Trijatmiko et al., 2016), suggesting that the two genes may not be synergistic in wheat and the overexpression of endogenous genes in wheat could be an alternative for increasing endospermic mineral expression. However, the very low efficiency of transformation in wheat, compared with crops like rice and barley, and of transgene expression in hexaploid wheat needs to be considered.
Minerals in cereal and legume grains are less bioavailable. The presence of phytic acid (PA), a phosphorus (P) storage protein in seed representing 65%–85% of seed P, acts as a chelator for cations of Ca2+, Mg2+, Zn2+, Mn2+, and Fe3+ and reduces their absorption in the intestine (Bhati et al., 2016). Thus, the bioavailability of grain Zn and Fe depends on proportionate phytic acid content in the grain or diet. The molar ratio of phytate: Zn/Fe can be used as a determinant of mineral bioavailability; an increase in the molar ratio indicates lower adsorption of Zn and Fe. Morris and Ellis (1989) determined critical ratios of phytate: Fe > 1 (Hallberg et al., 1989) and phytate: Zn > 15. Genotypes with lower phytic acid lines of PA: Zn < 0.4 and PA: Fe < 5 or 5–10 are preferable for biofortification (Gupta et al., 2022). Assessment of phytic acid in wheat genotypes showed higher genetic variability. Average PA in wheat grain ranges from 7.1 to 11.1 mg g−1, giving a molar ratio for PA: Zn from 24 to 41, in a set of 65 wheat genotypes from Pakistan (Hussain et al., 2012a). Another study involving 42 durum wheat genotypes showed PA variation from 0.462 to 0.952% and ranges for molar ratios of PA: Zn and PA: Fe of 16.9–23.6 and 12.1–29.6, respectively (Magallanes-Lopez et al., 2017). Wen et al. (2022) evaluated 330 wheat genotypes from CIMMYT, Mexico and reported 0.9%–1.72% range in PA content. Genome wide association studies revealed six stable genomic regions and the effect of four of the six region could reduce PA content from 1.21% to 1.13% which could potentially increase grain Zn and Fe bioavailability by 7%. In comparison, the PA content in T. monococcum, T. turgidum and T. aestivum was less in diploids than in tetraploids or hexaploids (Bilgrami et al., 2018). Seasonal variation of PA was also observed a significant difference in PA between tetraploids and hexaploids was recorded in the fall but was non-significant in the spring. Likewise, the PA: Zn molar ratio was less in T. monococcum (2.15), than T. turgidum (2.66) and T. aestivum (3.15) (Bilgrami et al., 2018). Understanding the physiological and metabolic pathways their underlying genes related to PA biosynthesis allow researchers to mitigate/reduce/control PA activity in grains. Phytic acid is produced during grain development and abscisic acid (ABA) and gibberellic acid (GA) regulates its accumulation during seed maturation. PA is synthesized from glucose-6-phosphate through a series of phosphorylation reactions by several inositol phosphate kinases (IPK) proteins. Genes such as TaIMP, TaITPK1-4, TaPLC1, TaIPK1, TaIPK2 were identified as being involved in PA biosynthesis in wheat (Aggarwal et al., 2015). A protein, TaABCC13, acts as a PA transporter and showed a pleiotropic effect on seed germination and root development (Bhati et al., 2016). Improving mineral bio-availability is possible by reducing PA expression in grain and also can be achieved through over-expression of the phytase enzyme, which degrades phytic acid and release minerals.
Approaches such as mutagenic treatment and transgenic development were adopted to develop low phytic acid mutant lines of maize, rice, barley and soybean. Maize low phytic acid mutants can be grouped into lpa1, lpa2, and lpa3 mutants; lpa1 affects transporter proteins that packages PA into vacuoles; lpa2 impairs the production of the inositol phosphate kinase enzyme (IPK1) that phosphorylates inositol-5-phosphate to PA and lpa3 impairs myo-inositol kinase that phosphorylates myo-inositol to inositol monophosphate (Singh et al., 2020). Plants with lpa mutations produced seeds with normal levels of phosphorus but greatly reduced PA bound phosphorus. Low phytic acid mutants of wheat showed a two-fold increase in grain Zn and Fe concentrations (Guttieri et al., 2004; Kenzhebayeva et al., 2019). In mutant lines, the distribution of grain P was altered, increasing the P content in the central endosperm and lowering it in the aleurone layer. But negative pleiotropic effects in lpa mutant line Js-12-LPA included reduced yield and height and weak straw, thus limiting the promise of lpa mutation for breeding (Guttieri et al., 2004).
Increasing grain phytase activity can readily increase nutrient bioavailablility. Endogenous phytase is produced during seed germination and releases the P from the PA, but in dry seed and flour, digestive tract phytase activity is very low or absent. Overexpression of the Aspergillus niger phytase gene (phyA) resulted in a 4-fold increase in phytase activity in wheat (Brinch-Pedersen et al., 2000). Detailed studies about increasing phytase activity in wheat are not available. Orthologous genetic information from other related species can be used to identify the gene families involved in PA biosynthesis and degradation and facilitate their use to develop wheat cultivars whose grain features low levels of phytic acid.
Understanding the nature of gene action and inheritance for micronutrient accumulation in the wheat grain is a prerequisite for improving the trait. Genetic analysis revealed the quantitative nature of inheritance for grain Zn and Fe contents, making improvement through conventional breeding is slow. Few studies exist regarding the nature of gene action for grain micronutrient content in wheat. One of the studies reported the predominance of additive gene action for grain Fe and dominance and duplicate epistasis for grain Zn in bread wheat (Amiri et al., 2020). Holasoua et al. (2021) reported additive gene action as being significant for both grain Zn and Fe. Higher heritability is paramount for increased genetic gains through selection. High heritability for grain Fe and moderately low heritability for grain Zn were reported by Amiri et al. (2020). Moderate-to-high heritability was detected in other studies (Velu et al., 2016a; Narendra et al., 2021). Low heritability for grain Zn and Fe content was also reported by Joshi et al. (2010). The existence of moderate-to-low heritability and the predominance of dominance and duplicate epistasis suggests advanced generation selection for the traits. High genotype × environment (G × E) interaction for grain Zn and Fe was reported under multilocation testing (Joshi et al., 2010; Velu et al., 2016a). Numerous factors affect grain Zn and Fe content, including soil nutrient availability (Alloway, 2009), soil moisture and organic matter (Cakmak, 2008; Pal et al., 2009), pH (Kirk and Bajita, 1995; Rupa and Tomar, 1999), tillage (Stipesevic et al., 2009), and soil N availability (Singh et al., 2018). Wheat grown under stress conditions showed higher grain Zn and Fe concentrations than crops grown under optimum conditions. Elevated temperatures and drought stress at post-anthesis increased grain Zn concentrations in wheat (Velu et al., 2016a; Narendra et al., 2021). This may be a result of the production of smaller grains and an increased aleurone: endosperm ratio under stress, particularly given that the overall Zn and Fe yield per unit area was higher in non-stressed environments (Velu et al., 2016a). For iron, there was no significant change in grain concentrations between genotypes grown under stressed or optimum conditions (Narendra et al., 2021).
A significant association between grain Zn and Fe contents has been observed (Velu et al., 2016a; Velu et al., 2019; Narendra et al., 2021), indicating improvement in micronutrient levels of one will lead to improvement in the other. This may be due to commonly associated proteins and enzymes for Zn and Fe uptake and translocation to the grains. However, studies have also found a negative association of grain Zn and Fe with grain yield (Morgounov et al., 2007; Narendra et al., 2021). Velu et al. (2016a) reported a negative association of grain yield with grain Zn but no association with grain Fe content. Simultaneous improvement of both traits is challenging. The significant negative association of grain yield with mineral concentrations could be due to a dilution effect, where high-yielding genotypes are contributing less grain Zn and Fe than photosynthates to the grain. In a study comparing modern wheat and rice cultivars with the cultivars released 50 years earlier, it was found that modern cultivars have less capacity to sequester Zn and Fe in the grain than earlier cultivars (Debnath et al., 2021). A study by Hao et al. (2021) found that wheat landraces carry higher grain Zn contents than cultivars and that landraces accumulated more grain Zn under foliar applications of the mineral.
With intensive breeding efforts, high-yielding, mineral dense wheat genotypes can be developed. An example is BARI Gom 33, developed by CIMMYT and released for commercial cultivation in Bangladesh and which offers a 7–8 ppm Zn advantage (Velu et al., 2019), as well as resistance to wheat blast (Roy et al., 2021b). Identification of high-yielding wheat genotypes with higher root uptakes or with a higher capacity to remobilize stored minerals or both is important. Grain Fe was improved with the supply of N in soil and foliar application (Aciksoz et al., 2011b), indicating the presence of useful genes for a higher uptake of N and Fe. In addition, a significant positive association of grain protein content with grain Zn and Fe has been reported (Morgounov et al., 2007). The locus TaNAM-B1 was detected to correspond higher levels of grain protein, Zn, and Fe (Uauy et al., 2006).
Interestingly, wild species, landraces and synthetic hexaploid wheats (SHW) are probable sources of grain iron and zinc in wheat. Monasterio and Graham (2000) evaluated around 3000 CIMMYT genotypes for grain Zn and Fe concentrations. T. tauschii genotypes had average and maximum iron concentrations of 76 and 99 ppm, with an average and maximum zinc concentrations of 50 and 68.9 ppm, respectively. T. monococcum had maximum iron and zinc levels of 70 and 131 ppm, respectively. T. dicoccoides and T. dicoccon showed zinc levels of 142 and 135 ppm. A core collection of Asian bread and durum wheat in the CIMMYT gene bank showed grain Zn levels ranging from 16.85 to 60.77 ppm and, for grain Fe, from 26.26 to 68.78 ppm (Velu et al., 2011). Ninety diploid and tetraploid wheat progenitors were screened for grain Zn and Fe content, few accessions of D and S genome species including Aegilops tauschii (D), Ae. kotschyi (US), Ae. speltoides (S), Ae. longissima (S) and Ae. bicornis (S) were found promising (Chhuneja et al., 2006). Grain Zn level up to 115.4 ppm (Syn43 [T. durum (Yuk) · Ae. tauschii (864)], 90.4 ppm (Ae. tauschii acc. 14,129) and grain Fe of 109.4 ppm (Ae. tauschii acc. 14,102) and 104.4 ppm (Ae. kotschyi acc. 3,573) were recorded. Another study reported non-progenitor wild species with S, U and M genome carrying 3–4 times higher grain Zn and Fe than the bread and durum wheat (Rawat et al., 2009b). Tetraploids showed higher concentrations of grain Zn and Fe than hexaploids, with ranges for Zn of 45–177 ppm in T. boeoticum, 20–159 in T. dococcoides, 29–89 ppm in T. monococcum and, for grain Fe, 41–92 ppm in T. boeoticum, 28–78 ppm in T. dococcoides, and 34–85 ppm in T. monococcum. This compared to 15–61 ppm for grain Zn and 24–51 ppm for grain Fe in T. aestivum and 18–50 ppm for grain Zn and 10–50 ppm for grain Fe in T. durum (Cakmak et al., 2000). Other studies have also reported the high genetic potential for grain Zn and Fe contents in diploid and tetraploid wild species such as T. monococcum, T. turgidum ssp dicoccoides, T. boeoticum, Aegilops tauschii and landraces (Cakmak, 2007; Velu et al., 2014). Grain Zn in T. turgidum ssp dicoccoides varied from 69 to 139 ppm; Fe concentrations from 44 to 88 ppm and protein from 164 to 382 g/kg (Peleg et al., 2007). A few accessions of T. turgidum ssp dicoccoides showed both high grain Zn and Fe concentrations. In most cases, the cultivated wheat genotypes showed lower levels of grain Zn and Fe. A range of grain Zn and Fe was 21–35 ppm and 22–34 ppm, respectively (Tang et al., 2008), 26–40 ppm and 35–56 ppm, respectively (Peterson et al., 1986), 29–46 ppm and 34–66 ppm, respectively (Ficco et al., 2009), 8–12 ppm and 29–38 ppm, respectively (Cakmak, 2000) were reported in the cultivated wheat genotypes.
Genomic regions associated with higher micronutrient density can be transferred from wild relatives to cultivated varieties. Wheat secondary and tertiary gene pools can be used to broaden the genetic base for grain mineral density through pre-breeding or the production of synthetic hexaploid wheat useful for the breeders. Chromosome addition lines from Ae. peregrina, Ae. longissima and Ae. umbellulata have been found to carry genes for high Zn and Fe in the grain and roots and control the release of high levels of mugineic acid in the root zone (Neelam et al., 2012). Synthetic amphidiploids produced from T. aestivum (AABBDD) and Ae. kotschyi (UUS1S1) carrying grain Zn and Fe more than double of the hexaploid parents (Rawat et al., 2009a).
The addition of rye chromosomes 1R and 7R improved the Zn efficiency of wheat cultivars. Increased zinc concentrations and contents were found in the shoots of chromosome addition lines (Cakmak et al., 1997). One of the most significant achievements in wheat breeding is the addition to hexaploid wheat of the 1BL:1RS rye chromosome segment, which has proved an invaluable source of resistance to stem, leaf, and yellow rust and powdery mildew, as well as providing wide adaptation, enhanced yield, and abiotic stress tolerance (Kumar et al., 2003). Likewise, the 2NS segment from Aegilops ventricosa is associated with cyst nematode resistance (Williamson et al., 2013), lodging resistance (Singh et al., 2019), and resistance to wheat blast, one of the devastating diseases of wheat (Roy et al., 2021b; Singh et al., 2021; Phuke et al., 2022). A study using Ae. longissima × T. turgidum ssp durum crosses showed that amphidiploids received genes from Ae. longissima for higher grain Zn and Fe contents than their durum parents (Tiwari et al., 2008). Considering the importance of wild relatives as a source of important genes for cultivated wheat, CIMMYT started production of SHW during the 1980s and many of these synthetics provided enhanced resilience against biotic and abiotic stresses (Mujeeb-Kazi, 1995). SHW developed from crossed of T. turgidum ssp turgidum x T. tauschii, T. turgidum ssp durum x T. tauschii, and T. turgidum ssp dicoccoides x T. tauschii produced higher grain Zn and Fe than parental accessions (Zhao et al., 2017). SHW developed by CIMMYT using T. spelta, T. turgidum var diccocoides, and Ae. squarosa were found to be promising for grain mineral concentration (Calderini and Ortiz-Monasterio, 2003; Crespo-Herrera et al., 2016). Calderini and Ortiz-Monasterio (2003) reported SHW carrying 25%–30% higher grain Zn, Fe and manganese (Mn). Using SHW, several novel genes responsible for high grain Zn and Fe have been transferred into CIMMYT elite wheat breeding lines; some were tested in Bangladesh, India, and Pakistan and released for commercial cultivation (Khokhar et al., 2018; Velu et al., 2018).
Attempts to detect the genomic regions associated with variation for grain protein, zinc and iron in cultivated and wild relatives of wheat have identified the relevant chromosome locations. For grain protein content, the locus GPC-B1 was mapped on chromosome 6B (DIC-6B) in the durum wheat genotype LND, a D genome disomic substitution line of durum wheat variety LANGDON (LND) from T. turgidum var dicoccoides (accession FA-15-3) (Joppa et al., 1997). The DIC-6B allele encodes a protein specific to the NAC transcription factor family and was found to be similar to the Arabidopsis No Apical Meristem (NAM) protein; therefore, it has been named NAM-B1 (Uauy et al., 2006). The NAC transcription factor is associated with auxin signaling, responses under biotic and abiotic stress, and early leaf senescence. This pleiotropic gene also increases grain protein, Zn, and Fe and triggers early senescence in the plant. Cultivated wheat carries a non-functional allele of the same gene with a 1 bp insertion of thymine base at position 11 that caused a frameshift mutation (Uauy et al., 2006). The presence of the DIC-6B locus increased durum wheat grain Zn, Fe, and Mn levels by 12, 18, and 29%, as well as resulting in a 38% higher protein accumulation than occurs in durum wheat without the locus (Distelfeld et al., 2007). Discovery of high-throughput, tightly-linked markers for GPC-B1 facilitated the marker-assisted transfer into the cultivated wheat. Through marker assisted breeding, the GPC-B1 locus was transferred into the Indian wheat variety HUW 468; some derivative lines showed significantly higher levels of grain Zn, Fe, and protein (Vishwakarma et al., 2014). Use of the DIC allele was a breakthrough for quality improvement in wheat worldwide. Wheat varieties with more than 14% GPC using the DIC locus were developed in Canada (https://www.grainscanada.gc.ca/en/grain-research/scientific-reports/pdf/canadian-wheat.pdf) and Australia (https://seedworld.com/australian-researchers-develop-high-protein-wheat/). Cultivars “Lassik” and “Farnum” in hexaploid wheat and “Westmore” and “Desert King High Protein” in durum wheat in United States; and “Lallian,” “Somerset” and “Burnside” in hexaploid wheat in Canada have been developed (Balyan et al., 2013). Recently, Indian scientists have developed a biofortified wheat variety MACS 4028 that contains 14.7% GPC and above 40 ppm zinc and iron (https://dst.gov.in/scientists-ari-pune-develop-biofortified-high-protein-wheat-variety). Major constraints on the expression of GPC-B1 appear to be a genetic background and environmental conditions, particularly high temperatures (Carter et al., 2012), making breeding to develop high-yielding, biofortified genotypes using the locus is a challenge, particularly for areas where terminal heat and drought are constraints and may lead to early senescence.
Selecting genotypes for highly quantitative traits that also show high G × E interaction is difficult through phenotypic assessment. QTL detection and their use through marker assisted selection can improve selection efficiency. The high environmental influence, narrow range of variation, and tedious phenotyping procedures for estimating grain Zn and Fe have made QTL mapping challenging. Phenotyping through inductively coupled plasma spectrometry (ICP-S) and, recently, use of energy-dispersive X-ray fluorescence spectrometry (ED-XRF) has facilitated the screening of large samples, then the atomic absorption spectrometry (AAS).
Efforts have been made in the last two decades to dissect the genetic components governing grain Zn and Fe content in wheat. A number of QTLs associated with the genetic variation of grain mineral content have been detected. QTL mapping was performed on the diverse genetic backgrounds using bi-parental and association mapping population and several QTLs of major and minor allelic variations were detected (Table 2). Most of the QTL mapping studies have focused on the background of SHW or wild relatives of tetraploid or diploid species, due to the presence of higher levels of genetic variation. One early study has reported the GPC-B1 locus, which has been cloned and sequenced (Uauy et al., 2006). Peleg et al. (2009) used recombinant inbred lines (RILs) developed from T. durum (cv. Langdon) × wild emmer wheat (accession #G18-16) and identified 82 QTLs for different minerals, six of which were associated with grain Zn and located on chromosomes 2A, 5A, 6B, 7A, 7B, and 11 of which linked to grain Fe were found on chromosomes 2A, 2B, 3A, 3B, 4B, 5A, 6A, 6B, 7A, and 7B. Using interspecific crosses between Triticum boeoticum (pau5088) × Triticum monococcum (pau14087), 2 QTLs for grain Fe were detected on chromosomes 2A and 7A and 1 QTL for grain Zn on 7A (Tiwari et al., 2009). Genes for enhanced grain Zn and Fe content were observed on chromosome 2S and 7U of Ae. kotschyi and substitution of 2S and 7U for the homoeologous A genome increased up to 117.4% grain Fe and 136% grain Zn over T. aestivum cv. WH 711 (Tiwari et al., 2010). A number of SHW were identified as potential donors and their underlying QTLs were detected. Using a SHW × T. spelta recombinant inbred line population, 12 QTLs for grain Zn and 7 QTLs for grain Fe were detected (Crespo-Herrera et al., 2017). The QTL Qgrain Zn.cimmyt-7B_1P2 for grain Zn on chromosome 7B, with a maximum phenotypic variation explained (PVE) of 32.7% and QTL Qgrain Fe.cimmyt-4A_P2 on chromosome 4A with 21.14% for grain Fe can be used in marker assisted breeding. Pleiotropic QTLs can be targeted through marker assisted selection for simultaneous improvement of multiple traits including yield. QTLs detected on chromosomes 2A, 5A, 6B for grain Zn were co-localized for high GPC and 5 loci on chromosomes 2A, 2B, 5A, 6A, and 7B for high grain Fe and GPC, indicating the potential correlated improvement for grain content of both minerals and proteins (Peleg et al., 2009). Other studies have reported common genomic regions. QTL for grain Zn content on 7B was co-localized with a QTL for grain Fe (Crespo-Herrera et al., 2017). Common genetic regions influencing high GPC, Zn and Fe contents were also found on chromosomes 2A and 5A (Peleg et al., 2009) and on 2A (Krishnappa et al., 2017). Although few, QTLs were detected in the hexaploid wheat background is advantageous, as they can be transferred through breeding without linkage drag, cross compatibility issues, or much time. Using RILs from a cross between a Chinese wheat line (with the lineage Hong Hua Mai/. ../Blouk #1) and the commercial bread wheat cultivar Roelfs F2007, 10 QTLs for grain Zn, 9 for grain Fe, 5 for GPC, and 36 for agronomic traits were detected (Liu et al., 2019b). Pleiotropic QTLs for grain Zn and Fe content on chromosome 3D and for grain Zn, GPC and thousand kernel weight were detected on chromosome 2B. Shi et al. (2008) used a double haploid population derived from two winter wheat cultivars and mapped 4 and 7 genomic locations for grain Zn concentration and grain Zn content, respectively. Four QTLs on chromosomes 4A, 4D, 5A and 7A were identified for grain Zn concentration and content and may facilitate simultaneous improvement of grain Zn content and concentration. Surprisingly, collocated QTLs for grain Zn and phosphorus (P) were found on chromosomes 4A and 4D, suggesting the potential for their correlated breeding improvement, even though the elements share an antagonistic relationship for plant uptake in soils.
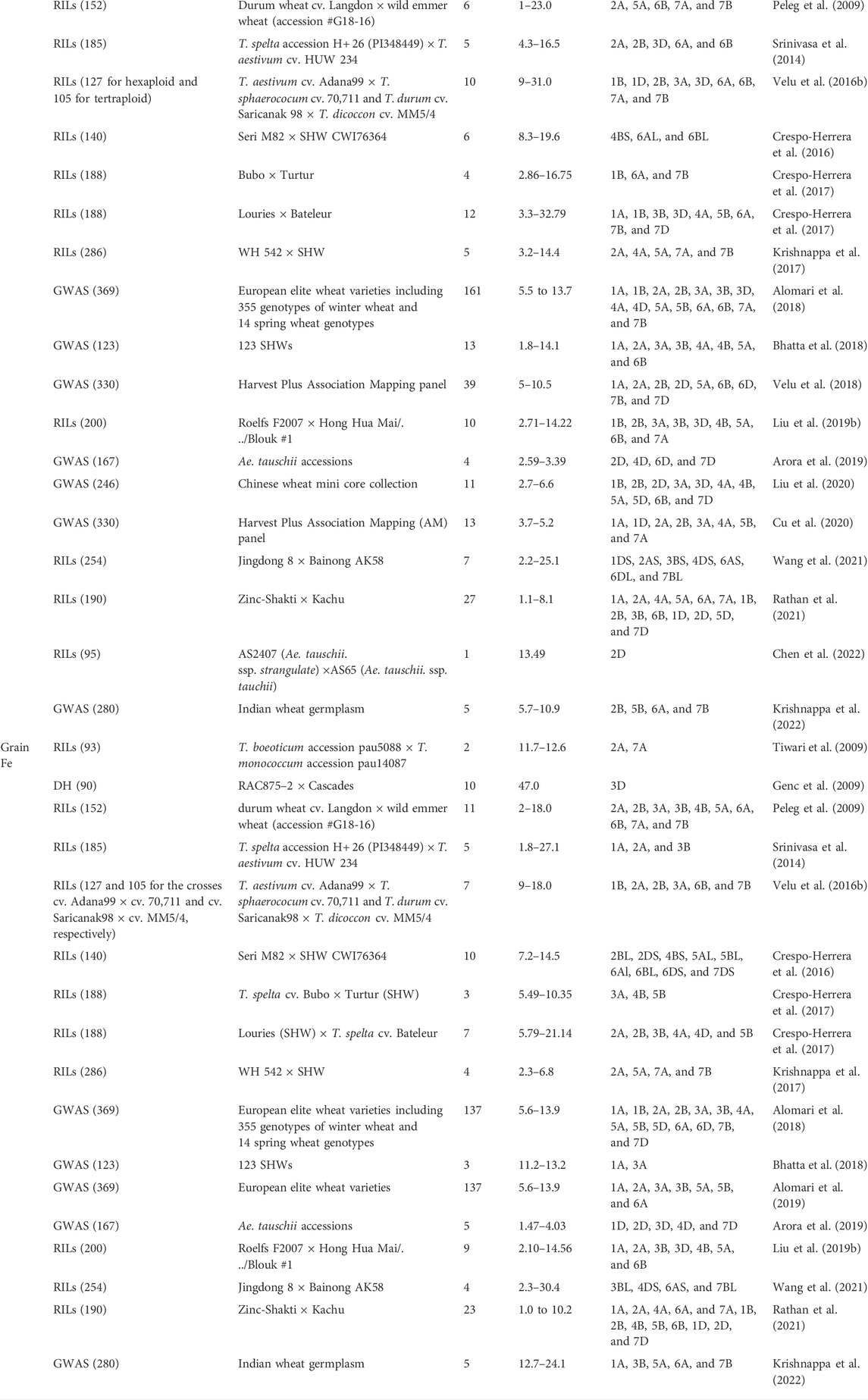
TABLE 2. Mapping population, number of QTLs detected along with their percent of phenotypic variation explained (PVE%) and chromosome carrying the QTLs for grain zinc and iron in wheat.
Genome wide association studies (GWAS) use potential natural variations to map significant marker trait associations (MTA), and provide the advantages of high resolution, high allele coverage over the bi-parental mapping population, and elimination of the disadvantages associated with RILs, where variation is confined within the two parental lines. Using GWAS in winter and red wheat cultivars, 161 MTAs were detected on 15 wheat chromosomes. The most significant regions were on chromosomes 3A and 5B and are related to transporter proteins such as those of the ZIP family and signal proteins of the MAPK family (Alomari et al., 2018). GWAS performed using a diverse panel of SHW develop from of T. durum, T. dicoccon, T. spelta, pre-breeding derivatives from T. polonicum, and landraces detected 39 QTLs for grain Zn on chromosomes 1A, 2A, 2B, 2D, 5A, 6B, 6D, 7B, and 7D (Velu et al., 2018). QTLs with large effects were detected on chromosomes 2B and 7B and are associated with transcription factors such as zinc finger motifs and metal ion binding protein (phosphatase), all of which has a role in the additional loading of Zn in wheat grains. Multiple studies have reported QTLs for high Zn and Fe concentrations on chromosomes 2A, 2B, 5A, and 7B (Srinivasa et al., 2014; Alomari et al., 2018; Alomari et al., 2019; Krishnappa et al., 2022). Transfer of these QTLs through marker assisted breeding will certainly improve mineral concentrations in wheat grains. However, detection of QTLs with major effects and stable expression across environments remains a constraint for marker-assisted breeding. Stable QTLs reported in some studies will be initial targets for breeding, but few QTLs were reported to be stable across years or locations. Based on across-location evaluations in India and Mexico, Velu et al. (2018) reported 39 stable MTAs (significant across at least three environments) for grain Zn. Crespo-Herrera et al. (2016) reported a QTL on 4BS that appeared across years governing 19.6% of the PVE for grain Zn and with pleiotropic effects on grain Zn and grain Fe. Alomari et al. (2018) reported a single MTA with minor effects on chromosome 3B and expressed in all 3 years of the study. Cu et al. (2020) reported five MTAs on chromosome 5B expressed over the season and having pleiotropic effects on grain Fe, Mn, Cu, and P contents. Recently, a meta-analysis using QTLs from seven different studies of diverse parental combinations (Shariatipour et al., 2021) detected meta-QTLs (MQTL) on chromosomes 1B, 2B, 4A, 5A, 7A, and 7B with MQTL-1, MQTL-5 and MQTL-7 on chromosomes 1B, 7A, and 7B, respectively, were detected with the highest number of initial QTLs. Functional analysis of candidate genes confined in the MQTL revealed that most of the genes were associated with Zn and Fe homeostasis. The meta-QTL analysis is a powerful tool to reduce the confidence interval for the QTLs by integrating information from independent QTL analyses performed by different authors and helps to identify more reliable QTLs.
Wheat biofortification is challenging, an integrated approach needs to be constructed integrating conventional breeding, biotechnological tools, new breeding techniques, and agronomic measures for immediate soil remediation (Figure 1). In populations with diets heavy in staple crop foods and highly susceptible to micronutrient deficiencies, supplementation through pharmaceutical products and food additives may be required and need support through governmental policies.

FIGURE 1. An integrated approach for eradication of malnutrition including new breeding techniques, supplementation and diversification.
A number of wheat genotypes with higher grain Zn and Fe content have been developed and released for cultivation in Australia, Bangladesh, Bolivia, India, Nepal, Mexico, and Pakistan (Table 3). Constraints associated with selective breeding include limited genetic variation, low heritability, crossability barriers, and linkage drag, to name several. Yield gains through pure line varieties have plateaued and the development of pure line varieties is slow. Working in the joint Rockefeller Foundation-Mexican government Office of Special Studies in the mid-20th century, Nobel Peace laureate Dr. Norman E. Borlaug launched a “shuttle breeding” approach to speed wheat varietal development, allowing two breeding cycles per year and still used by CIMMYT. Double haploids represents another method to rapidly fix genotypes from gametospores of an F1 plant, but are also associated with the flaws of a single generation of recombination and not exposed to diverse environments for hardening. Recently, speed breeding has emerged as an approach to the rapid advancement of generation. In this approach, plants are raised in growth chambers under controlled conditions with extended temperatures and photoperiods (Watson et al., 2018). Among other things, this modulates flowering genes to stimulate the early onset of flowering and, as a result, allows 5–6 generations to be grown in a year and the rate of genetic gains is greatly increased. For instance, genetic gain is ∆G = ihσA/L, where i = selection intensity, h = heritability, σA = standard deviation of additive genetic variance and L = length of breeding cycle interval or generation. Through modification in the selection intensity and accurate phenotyping, genetic advance can be enhanced to some extent. Rapid breeding cycles also lead to higher genetic gain and the early development of homozygous lines. For quantitative traits with low heritability where the selection in advanced generations is practiced, speed breeding can be of great importance to achieve high homozygosity quickly. Speed breeding has been used in breeding for spring wheat, durum wheat, barley, pea, chickpea and canola (Watson et al., 2018). In wheat, speed breeding has been used to generate genotypes resistant to stem rust, yellow rust, and fusarium head blight (Voss-Fels et al., 2019).
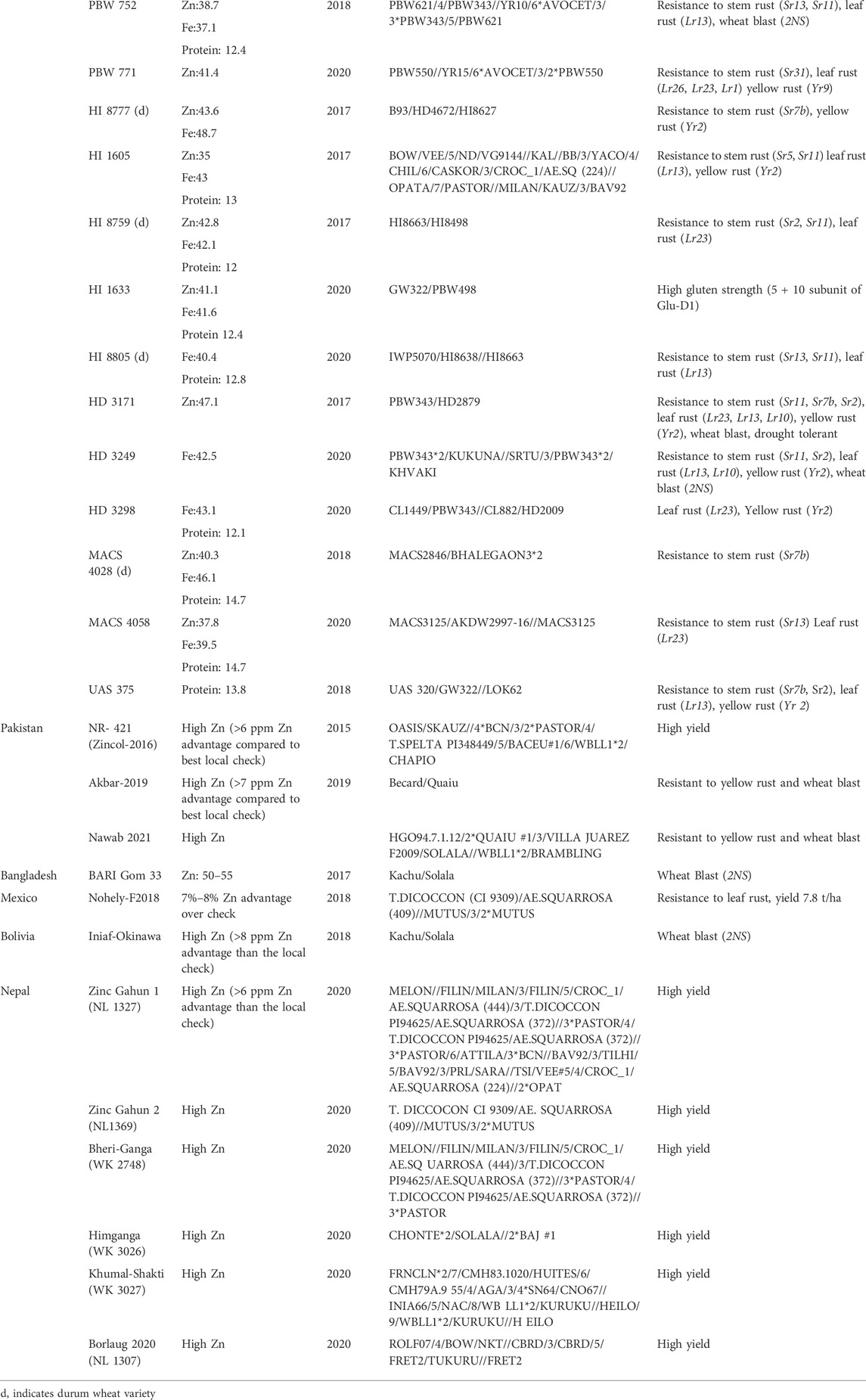
TABLE 3. List of biofortified wheat varieties released for commercial cultivation with their grain micronutrient level (ppm), protein content (%) and additional traits (modified from Gupta et al., 2022).
Genomic selection (GS) is a powerful tool to increase genetic gains, shorten breeding cycles, prediction the performance of an individual in an untested environment, and increase selection accuracy and genetic gain. GS is most effective for the traits where phenotyping is costly—particularly, yield and quality traits. GS can be used for the selection of quantitative traits involving complex physiological mechanisms, each underpinned by multiple genetic variations that require genome-wide selection approaches, rather than targeting single QTLs. GS calculates genomic assisted breeding values (GEBVs) by estimating all available genetic variations in an individual. It uses the genomic best linear unbiased prediction (GBLUP) model and a multi-environment, linear mixed model to estimate correlated environmental structure, to predict the performance of individuals before phenotyping, based on genotyping and pedigree information. In GS, a set of individuals are genotyped and phenotyped as a training population for use to develop the prediction model (Juliana et al., 2018). The efficacy of GS has been tested for multiple traits in several crops, including wheat, for yield (Juliana et al., 2018), leaf rust and yellow rust (Juliana et al., 2017), spot blotch (Juliana et al., 2022b), and wheat blast (Juliana et al., 2022a). GS to improve grain mineral concentrations has been employed in maize (Mageto et al., 2020), rice (Rakotondramanana et al., 2022), and wheat (Velu et al., 2018), with reasonable prediction accuracies. Velu et al. (2018) reported prediction accuracies ranging from 0.331 to 0.694, with an average of 0.542, for grain Zn content and 0.324 to 0.734, with an average of 0.529, for grain Fe content, in the Harvest Plus Association Mapping (HPAM) panel. Prediction accuracy was higher in the environments with high heritability and soil available Zn. Genetic relatedness increases prediction ability and prediction ability sharply declines as relatedness between the training and testing population decreases (Crossa et al., 2010) and when evaluations are conducted in poor environments and with poor phenotyping (Velu et al., 2018). GS becomes more effective with higher prediction accuracies and applied in early generations of selection to retain lines with high breeding value and discarding those not expected to give higher genetic gains in advanced generations. GS can easily be incorporated with speed breeding to increase genetic gain. GS has not been studied in detail for increasing mineral density in wheat. A model has been proposed where GS was accumulated through speed breeding and predicted higher genetic gains over conventional breeding (Voss-Fels et al., 2019). It has been shown that the incorporation of GS with speed breeding can significantly increase grain yield, over conventional phenotypic selection.
Genome editing is a powerful tool in the field of medicine, agriculture and the life sciences, allowing targeted changes in a genotype and avoided the random changes that occur in induced mutations. Genome editing can produce targeted genetic variation where none previously existed. Unlike the case of genetically modified crops, gene editing does not require the insertion of foreign DNA. Deletions, additions, single nucleotides or DNA segment substitution are used to change a target gene. Researchers have developed artificial site-specific nucleases, broadly classified as zinc-finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and clustered regularly interspaced short palindromic repeats (CRISPR)-CRISPR-associated (Cas) nucleases that are designed to effect a targeted, double-stranded break in the DNA (Karmakar et al., 2022). ZFNs consist of a zinc finger DNA binding domain attached with nuclease FokI (Carroll, 2011). TALENs are engineered with a TAL effector binding DNA domain attached with FokI nuclease (Joung and Sander, 2013). CRISPR-Cas nuclease consists of a guide RNA and a Cas nuclease. Using the Cas9 endonuclease, CRISPR-Cas9 is the most extensively used system for genome editing, thanks to its simple design, cost effectiveness, high efficacy, reproducibility and engineering feasibility (Karmakar et al., 2022). A single guide RNA (sgRNA) hybrid consisting of a CRISPR-RNA and a transactivating RNA locate the binding and cleavage site for the Cas9 nuclease. CRISPR-Cas9 genome editing is increasingly used to enhance disease resistance, tolerance to abiotic stresses (drought, heat, salinity) and end-use quality in food crops. The role of genome editing in crop biofortification is being exploited for several traits including vitamin-A enrichment, targeted increases in grain zinc and iron, and reducing anti-nutritional factors in the grain (Kumar et al., 2022). Targeted knockout of the OsAAP6 and OsAAP10 genes using gene editing has reduced protein content in rice (Wang et al., 2020). The CRISPR-Cas9 system was used to create novel OsBADH2 genes in non-aromatic rice lines and created aroma (Kumar et al., 2020). Targeted change in OsNRAMP2 gene enhanced iron remobilization and distribution in rice (Chang et al., 2022). In wheat, CRISPR-Cas9 mediated genome change in the α-gliadin gene resulted in low gluten wheat (Sánchez-León et al., 2018). Reduction in phytic acid levels through the disruption of inositol pentakisphosphate 2-kinase 1 (TaIPK1) improves grain Zn and Fe bioavailability in wheat (Ibrahim et al., 2021). In the future, numerous genes can be targeted through CRISPR-Cas to develop biofortified crops. To promote genome edited technology in crop improvement, Argentina, Brazil, Colombia, Chile, India, and the United States have established regulatory measures separate from those applied for transgenic crops.
The suppression of gene expression through antisense or RNAi technology is a powerful way to modulate biosynthetic pathways. RNAi is preferred over antisense technology, as it is more stable, efficient, and precise. Using a double stranded RNA (dsRNA) molecule, RNAi inhibits the expression of a gene at the transcription and translational levels. It has been used for biotic and abiotic stress tolerance and nutritional quality improvement. RNAi mediated suppression of the inositol pentakisphosphate kinase (IPK1) gene involved in the phytic acid biosynthesis pathway reduced phytic acid level of 28%–56% in wheat (Aggarwal et al., 2018). Similarly, downregulation of the rice inositol triphosphate kinase (OsITP5/6K-1) gene through RNAi resulted in a 42% reduction in phytic acid in rice grains (Karmakar et al., 2020). This technology can be used to elucidate biosynthetic pathways and understand the roles of different genes in Zn and Fe sequestration.
Advancement in next-generation sequencing platforms has enabled the development of a whole genome sequence for Triticum aestivum cv. Chinese Spring (https://www.wheatgenome.org/News/Latest-news/RefSeq-v1.0-URGI), which is the largest cereal genome. Genome sequence information of diploid (AA, DD genome) and tetraploid (AABB) progenitors are also available (Zhou et al., 2020). Sequence information for wheat wild relatives will elucidate the genetic mechanisms and variation for micronutrient levels in wheat grain, paving the way for structural genomics, functional genomics and metabolomics. Integration of multiple omics data with high-throughput techniques has elucidated the complex pathways of plant growth and their responses under biotic and abiotic stress. The availability of other omics technologies (mutagenomics, pangenomics, transcriptomics, proteomics, ionomics, and phenomics) would provide new knowledge on complex biological systems. Huge amounts of data are generated using these technologies; advances in computing and integrated system-based analysis to use and manage data will expand our understanding of genotype and phenotype relationships, allowing wheat breeders to make targeted changes in the pathways of metal homeostasis and develop bio-fortified genotypes.
The review updates about wheat biofortification research include conventional and molecular breeding. Mainstreaming biofortification is essential along with yield, disease resistance and other parameters. Being quantitative, limited genetic variations in the cultivated wheat varieties, improvement through conventional breeding is hard. Available genomic resources are needed to be utilized while breeding for high grain zinc and iron content wheat varieties. An integrated breeding programme including marker assisted selection, genomic approaches and gene editing tools will gear up development of biofortified wheat varieties.
CR conceptualized and drafted the first version; SK collected the information; SK, RR, SRK, and VG edited the article.
Part of the research work was supported by a grant from the Bill and Melinda Gates Foundation (INV003012) and co-funded by the Foreign, Commonwealth and Development Office (FCDO) of the United Kingdom Government to CIMMYT.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer ZW declared a past co-authorship with the author VG to the handling editor.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aciksoz, S. B., Ozturk, L., Gokmen, O. O., Roemheld, V., and Cakmak, I. (2011a). Effect of nitrogen on root release of phytosiderophores and root uptake of Fe(III)-phytosiderophore in Fedeficient wheat plants. Physiol. Plant. 142, 287–296. doi:10.1111/j.1399-3054.2011.01460.x
Aciksoz, S. B., Yazici, A., Ozturk, L., and Cakmak, I. (2011b). Biofortification of wheat with iron through soil and foliar application of nitrogen and iron fertilizers. Plant Soil 349, 215–225. doi:10.1007/s11104-011-0863-2
Aggarwal, S., Kumar, A., Bhati, K. K., Kaur, G., Shukla, V., Tiwari, S., et al. (2018). RNAi-mediated downregulation of inositol pentakisphosphate kinase (IPK1) in wheat grains decreases phytic acid levels and increases Fe and Zn accumulation. Front. Plant Sci. 9, 259. doi:10.3389/fpls.2018.00259
Aggarwal, S., Shukla, V., Bhati, K. K., Kaur, M., Sharma, S., Singh, A., et al. (2015). Hormonal regulation and expression profiles of wheat genes involved during phytic acid biosynthesis pathway. Plants 4, 298–319. doi:10.3390/plants4020298
Ajeesh-Krishna, T. P., Maharajan, T., Victor Roch, G., Ignacimuthu, S., and Antony Ceasar, S. (2020). Structure, function, regulation and phylogenetic relationship of ZIP family transporters of plants. Front. Plant Sci. 11, 662. doi:10.3389/fpls.2020.00662
Alloway, B. J. (2009). Soil factors associated with zinc deficiency in crops and humans. Environ. Geochem. Health 31, 537–548. doi:10.1007/s10653-009-9255-4
Alomari, D. Z., Eggert, K., von Wiren, N., Alqudah, A. M., Polley, A., Plieske, J., et al. (2018). Identifying candidate genes for enhancing grain Zn concentration in wheat. Front. Plant Sci. 9, 1313. doi:10.3389/fpls.2018.01313
Alomari, D. Z., Eggert, K., von Wiren, N., Polley, A., Plieske, J., Ganal, M. W., et al. (2019). Whole-genome association mapping and genomic prediction for iron concentration in wheat grains. Int. J. Mol. Sci. 20, 76. doi:10.3390/ijms20010076
Amiri, R., Bahraminejad, S., Cheghamirza, K., and Arzani, A. (2020). Genetic analysis of iron and zinc concentrations in bread wheat grains. J. Cereal Sci. 95, 103077. doi:10.1016/j.jcs.2020.103077
Appels, R., Eversole, K., Feuillet, C., Keller, B., Rogers, J., Stein, N., et al. (2018). Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 361, eaar7191. doi:10.1126/science.aar7191
Arora, S., Cheema, J., Poland, J., Uauy, C., and Chhuneja, P. (2019). Genome-wide association mapping of grain micronutrients concentration in Aegilops tauschii. Front. Plant Sci. 10, 54. doi:10.3389/fpls.2019.00054
Balyan, H. S., Gupta, P. K., Kumar, S., Dhariwal, R., Jaiswal, V., Tyagi, S., et al. (2013). Genetic improvement of grain protein content and other health-related constituents of wheat grain. Plant Breed. 132, 446–457. doi:10.1111/pbr.12047
Beal, T., Massiot, E., Arsenault, J. E., Smith, M. R., and Hijmans, R. J. (2017). Global trends in dietary micronutrient supplies and estimated prevalence of inadequate intakes. PLoS One 12, e0175554. doi:10.1371/journal.pone.0175554
Beasley, J. T., Bonneau, J. P., Sanchez-Palacios, J. T., Moreno-Moyano, L. T., Callahan, D. L., Tako, E., et al. (2019). Metabolic engineering of bread wheat improves grain iron concentration and bioavailability. Plant Biotechnol. J. 17, 1514–1526. doi:10.1111/pbi.13074
Bhati, K. K., Alok, A., Kumar, A., Kaur, J., Tiwari, S., and Pandey, A. K. (2016). Silencing of ABCC13 transporter in wheat reveals its involvement in grain development, phytic acid accumulation and lateral root formation. J. Exp. Bot. 67, 4379–4389. doi:10.1093/jxb/erw224
Bhatta, M., Baenziger, P. S., Waters, B. M., Poudel, R., Belamkar, V., Poland, J., et al. (2018). Genome-wide association study reveals novel genomic regions associated with 10 grain minerals in synthetic hexaploid wheat. Int. J. Mol. Sci. 19, 10 3237. doi:10.3390/ijms19103237
Bilgrami, S. S., Houshmand, S., Kadivar, M., Fakheri, B., Zandi, P., Shariati, V., et al. (2018). Phytic acid, iron and zinc content in wheat ploidy levels and amphiploids: The impact of genotype and planting seasons. Arch. Agron. Soil Sci. 64, 331–346. doi:10.1080/03650340.2017.1352085
Borg, S., Brinch-Pedersen, H., Tauris, B., Madsen, L. H., Darbani, B., Noeparvar, S., et al. (2012). Wheat ferritins: Improving the iron content of the wheat grain. J. Cereal Sci. 56, 204–213. doi:10.1016/j.jcs.2012.03.005
Borrill, P., Connorton, J. M., Balk, J., Miller, A. J., Sanders, D., and Uauy, C. (2014). Biofortification of wheat grain with iron and zinc: Integrating novel genomic resources and knowledge from model crops. Front. Plant Sci. 5, 53. doi:10.3389/fpls.2014.00053
Bouis, H. E., Hotz, C., McClafferty, B., Meenakshi, J. V., and Pfeiffer, W. H. (2011). Biofortification: A new tool to reduce micronutrient malnutrition. Food Nutr. Bull. 32, S31–S40. doi:10.1177/15648265110321S105
Brinch-Pedersen, H., Olesen, A., Rasmussen, S. K., and Holm, P. B. (2000). Generation of transgenic wheat (Triticum aestivum L.) for constitutive accumulation of an Aspergillus phytase. Mol. Breed. 6, 195–206.
Cakmak, I., Derici, R., Torun, I., Braun, H. J., and Schlegel, R. (1997). Role of rey chromosome in improvemengt of zinc efficiency in whet and triticale. Plant Soil 196, 249–253. doi:10.1023/a:1004210309876
Cakmak, I. (2007). Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil 302, 1–17. doi:10.1007/s11104-007-9466-3
Cakmak, I. (2008). Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil 302, 1–17. doi:10.1007/s11104-007-9466-3
Cakmak, I., Gulut, K. Y., Marschner, H., and Graham, R. D. (1994). Efect of zinc and iron deficiency on phytos1derophore release in wheat genotypes differing in zinc efficiency. J. Plant Nutr. 17, 1–17. doi:10.1080/01904169409364706
Cakmak, I., Kalayci, M., Ekiz, H., Braun, H. J., and Yilmaz, A. (1999). Zinc deficiency as a practical problem in plant and human nutrition in Turkey: A NATO-science for stability project. Field Crops Res. 60, 175–188. doi:10.1016/s0378-4290(98)00139-7
Cakmak, I., and Kutman, U. B. (2018). Agronomic biofortification of cereals with zinc: A review. Eur. J. Soil Sci. 69, 172–180. doi:10.1111/ejss.12437
Cakmak, I., Ozkan, H., Braun, H. J., Welch, R. M., and Romheld, V. (2000). Zinc and iron concentrations in seeds of wild, primitive, and modern wheats. Food Nutr. Bull. 21, 401–403. doi:10.1177/156482650002100411
Cakmak, I. (2000). Plant nutrition research: Priorities to meet human needs for food in sustainable ways. Plant Soil 247, 3–24. doi:10.1023/a:1021194511492
Cakmak, I., Yilmaz, A., Ekiz, H., Torun, B., Erenoglu, B., Braun, H. J., et al. (1996). Zinc deficiency as a critical problem in wheat production in Central Anatolia. Plant Soil 180, 165–172. doi:10.1007/bf00015299
Calderini, D. F., and Ortiz-Monasterio, I. (2003). Are synthetic hexaploids a means of increasing grain element concentrations in wheat? Euphytica 134, 169–178. doi:10.1023/b:euph.0000003849.10595.ac
Carroll, D. (2011). Genome engineering with zinc-finger nucleases. Genetics 188, 773–782. doi:10.1534/genetics.111.131433
Carter, A. H., Santra, D. K., and Kidwell, K. K. (2012). Assessment of the effects of the Gpc-B1 allele on senescence rate, grain protein concentration and mineral content in hard red spring wheat (Triticum aestivum L.) from the Pacific Northwest Region of the USA. Plant Breed. 131, 62–68. doi:10.1111/j.1439-0523.2011.01900.x
Chang, J.-D., Xie, Y., Zhang, H., Zhang, S., and Zhao, F.-J. (2022). The vacuolar transporter OsNRAMP2 mediates Fe remobilization during germination and affects Cd distribution to rice grain. Plant Soil, 1–17. doi:10.21203/rs.3.rs-1206604/v1
Chen, H., Liu, T., Zhou, R., Ning, S., Zhang, L., and Zhang, B. (2022). Identification of novel major quantitative trait loci associated with selenium and zinc accumulation in Aegilops tauschii. Res. Sq., 1–18.
Chhuneja, P., Dhaliwal, H. S., Bains, N. S., and Singh, K. (2006). Aegilops kotschyi and Ae. tauschii as a source for higher levels of grain iron and zinc. Plant Br. 125, 531–539.
Colangelo, E. P., and Guerinot, M. L. (2006). Put the metal to the petal: Metal uptake and transport throughout plants. Curr. Opin. Plant Biol. 9, 322–330. doi:10.1016/j.pbi.2006.03.015
Connorton, J. M., Jones, E. R., Rodríguez-Ramiro, I., Fairweather-Tait, S., Uauy, C., and Balk, J. (2017). Wheat vacuolar iron transporter TaVIT2 transports Fe and Mn and is effective for biofortification. Plant Physiol. 174, 2434–2444. doi:10.1104/pp.17.00672
Crespo-Herrera, L. A., Govindan, V., Stangoulis, J., Hao, Y., and Singh, R. P. (2017). QTL mapping of grain Zn and Fe concentrations in two hexaploid wheat RIL populations with ample transgressive segregation. Front. Plant Sci. 8, 1800. doi:10.3389/fpls.2017.01800
Crespo-Herrera, L. A., Velu, G., and Singh, R. P. (2016). Quantitative trait loci mapping reveals pleiotropic effect for grain iron and zinc concentrations in wheat. Ann. Appl. Biol. 169, 27–35. doi:10.1111/aab.12276
Crossa, J., de los Campos, G., Perez, P., Gianola, D., Burgueno, J., Aruas, J. L., et al. (2010). Prediction of genetic values of quantitative traits in plant breeding using pedigree and molecular markers. Genetics 186, 713–724. doi:10.1534/genetics.110.118521
Cu, S. T., Guild, G., Nicolson, A., Velu, G., Singh, R., and Stangoulis, J. (2020). Genetic dissection of zinc, iron, copper, manganese and phosphorus in wheat (Triticum aestivum L.) grain and rachis at two developmental stages. Plant Sci. 291, 110338. doi:10.1016/j.plantsci.2019.110338
Curie, C., Panavience, Z., Loulergue, C., Dellaporta, S. L., Briat, J. F., and Walker, E. L. (2001). Maize yellow stripe1 encodes a membrane protein directly involved in Fe(III) uptake. Nature 409, 346–349. doi:10.1038/35053080
Debnath, S., Mandal, B., Saha, S., Sarkar, D., Batabyal, K., Murmu, S., et al. (2021). Are the modern-bred rice and wheat cultivars in India inefficient in zinc and iron sequestration? Environ. Exp. Bot. 189, 104535. doi:10.1016/j.envexpbot.2021.104535
Desbrosses-Fonrouge, A. G., Voigt, K., Schroder, A., Arrivault, S., Thomine, S., and Kramer, U. (2005). Arabidopsis thaliana MTP1 is a Zn transporter in the vacuolar membrane which mediates Zn detoxification and drives leaf Zn accumulation. FEBS Lett. 579, 4165–4174. doi:10.1016/j.febslet.2005.06.046
Dimkpa, C. O., Andrewsa, J., Sanabria, J., Bindraban, P. S., Singha, U., Elmer, W. H., et al. (2020). Interactive effects of drought, organic fertilizer, and zinc oxide nanoscale and bulk particles on wheat performance and grain nutrient accumulation. Sci. Total Environ. 722, 137808. doi:10.1016/j.scitotenv.2020.137808
Distelfeld, A., Cakmak, I., Peleg, Z., Ozturk, L., Yazici, A. M., Budak, H., et al. (2007). Multiple QTL-effects of wheat Gpc-B1 locus on grain protein and micronutrient concentrations. Physiol. Plant. 129, 635–643. doi:10.1111/j.1399-3054.2006.00841.x
Drakakaki, G., Christou, P., and Stoger, E. (2000). Constitutive expression of soybean ferritin cDNA in transgenic wheat and rice results in increased iron levels in vegetative tissues but not in seeds. Transgenic Res. 9, 445–452. doi:10.1023/a:1026534009483
Ficco, D. B. M., Riefolo, C., Nicastro, G., De Simone, V., Di Gesù, A. M., Beleggia, R., et al. (2009). Phytate and mineral elements concentration in a collection of Italian durum wheat cultivars. Field Crops Res. 111, 235–242. doi:10.1016/j.fcr.2008.12.010
Genc, Y., Verbyla, A. P., Torun, A. A., Cakmak, I., Willsmore, K., Wallwork, H., et al. (2009). Quantitative trait loci analysis of zinc efficiency and grain zinc concentration in wheat using whole genome average interval mapping. Plant Soil 314, 49–66. doi:10.1007/s11104-008-9704-3
Gillies, S. A., Futardo, A., and Henry, R. J. (2012). Gene expression in the developing aleurone and starchy endosperm of wheat. Plant Biotechnol. J. 10, 668–679. doi:10.1111/j.1467-7652.2012.00705.x
Gupta, O. P., Pandey, V., Saini, R., Sneh Narwal, S., Malik, V. K., Khandale, T., et al. (2020a). Identifying transcripts associated with efficient transport and accumulation of Fe and Zn in hexaploid wheat (Triticum aestivum L.). J. Biotechnol. 316, 10 46–55. doi:10.1016/j.jbiotec.2020.03.015
Gupta, O. P., Singh, A. K., Singh, A., Singh, G. P., Bansal, K. C., and Datta, S. K. (2022). Wheat biofortification: Utilizing natural genetic diversity, genome-wide association mapping, genomic selection, and genome editing technologies. Front. Nutr. 9, 826131. doi:10.3389/fnut.2022.826131
Gupta, S., Brazier, A. K. M., and Lowe, N. M. (2020b). Zinc deficiency in low- and middle-income countries: Prevalence and approaches for mitigation. J. Hum. Nutr. Diet. 33, 624–643. doi:10.1111/jhn.12791
Gupta, U. C. (1991). Iron status of crops in Prince Edward Island and effect of soil pH on plant iron concentration. Can. J. Soil Sci. 71, 197–202. doi:10.4141/cjss91-019
Guttieri, M., Bowen, D., Dorsch, J. A., Raboy, V., and Souza, E. (2004). Identification and characterization of a low phytic acid wheat. Crop Sci. 44, 418–1505. doi:10.2135/cropsci2004.0418
Hallberg, L., Brune, M., and Rossander, L. (1989). Iron absorption in man: Ascorbic acid and dose-dependent inhibition by phytate. Am. J. Clin. Nutr. 49, 140–144. doi:10.1093/ajcn/49.1.140
Hao, B., Ma, J., Jiang, L., Wang, X., Bai, Y., Zhou, C., et al. (2021). Effects of foliar application of micronutrients on concentration and bioavailability of zinc and iron in wheat landraces and cultivars. Sci. Rep. 11, 22782. doi:10.1038/s41598-021-02088-3
Heck, S., Campos, H., Barker, I., Okello, J. J., Baral, A., Boy, E., et al. (2020). Resilient agri-food systems for nutrition amidst COVID-19: Evidence and lessons from food-based approaches to overcome micronutrient deficiency and rebuild livelihoods after crises. Food Secur. 12, 823–830. doi:10.1007/s12571-020-01067-2
Holasoua, H. A., Kiaa, S. S. A., Mohammadia, S. A., and Vaheda, M. M. (2021). Pattern of genetic inheritance of grain zinc and iron content, agronomic and biochemical traits in bread wheat using mixed linear models under salinity stress. Res. Sq., 1–25. doi:10.21203/rs.3.rs-367613/v1
Hussain, D., Haydon, M. J., Wang, Y., Wong, E., Sherson, S. M., Young, J., et al. (2004). P-type ATPase heavy metal transporters with roles in essential zinc homeostasis in Arabidopsis. Plant Cell. 16, 1327–1339. doi:10.1105/tpc.020487
Hussain, S., Maqsood, M. A., and Miller, L. V. (2012a). Bioavailable zinc in grains of bread wheat varieties of Pakistan. Cereal Res. Commun. 40, 62–73. doi:10.1556/crc.2011.003
Hussain, S., Maqsood, M. A., Rengel, Z., and Aziz, T. (2012b). Biofortification and estimated human bioavailability of zinc in wheat grains as influenced by methods of zinc application. Plant Soil 361, 279–290. doi:10.1007/s11104-012-1217-4
Ibrahim, S., Saleem, B., Rehman, N., Zafar, S. A., Naeem, M. K., and Khan, M. R. (2021). CRISPR/Cas9 mediated disruption of inositol pentakisphosphate 2- kinase 1 (TaIPK1) reduces phytic acid and improves iron and zinc accumulation in wheat grains. J. Adv. Res. 37, 33–41. doi:10.1016/j.jare.2021.07.006
Joppa, L. R., Du, C., Hart, G. E., and Hareland, G. A. (1997). Mapping a QTL for grain protein in tetraploid wheat (Triticum turgidum L.) using a population of recombinant inbred chromosome lines. Crop Sci. 37, 1586–1589. doi:10.2135/cropsci1997.0011183x003700050030x
Joshi, A. K., Crossa, J., Arun, B., Chand, R., Trethowan, R., Vargas, M., et al. (2010). Genotype×environment interaction for zinc and iron concentration of wheat grain in eastern Gangetic plains of India. Field Crops Res. 116, 268–277. doi:10.1016/j.fcr.2010.01.004
Joung, J. K., and Sander, J. D. (2013). TALENs: aWidely applicable technology for targeted genome editing. Nat. Rev. Mol. Cell. Biol. 14, 49–55. doi:10.1038/nrm3486
Juliana, P., He, X., Marza, F., Islam, R., Anwar, B., Poland, J., et al. (2022a). Genomic selection for wheat blast in a diversity panel, breeding panel and full-sibs panel. Front. Plant Sci. 7, 745379. doi:10.3389/fpls.2021.745379
Juliana, P., He, X., Poland, J., Roy, K. K., Malaker, P. K., Mishra, V. K., et al. (2022b). Genomic selection for spot blotch in bread wheat breeding panels, full-sibs and half-sibs and index-based selection for spot blotch, heading and plant height. Theor. Appl. Genet. 135, 1965–1983. doi:10.1007/s00122-022-04087-y
Juliana, P., Singh, R. P., Poland, J., Mondal, S., Crossa, J., Montesinos- López, O. A., et al. (2018). Prospects and challenges of applied genomic selection—a new paradigm in breeding for grain yield in bread wheat. Plant Genome 11, 180017. doi:10.3835/plantgenome2018.03.0017
Juliana, P., Singh, R. P., Singh, P. K., Crossa, J., Huerta-Espino, J., Lan, C., et al. (2017). Genomic and pedigree-based prediction for leaf, stem, and stripe rust resistance in wheat. Theor. Appl. Genet. 130, 1415–1430. doi:10.1007/s00122-017-2897-1
Karmakar, A., Bhattacharya, S., Sengupta, S., Ali, N., Sarkar, S. N., Datta, K., et al. (2020). RNAi-mediated silencing of ITPK gene reduces phytic acid content, alters transcripts of phytic acid biosynthetic genes, and modulates mineral distribution in rice seeds. Rice Sci. 27, 315–328. doi:10.1016/j.rsci.2020.05.007
Karmakar, S., Das, P., Panda, D., Xie, K., Baig, M. J., and Molla, K. A. (2022). A detailed landscape of CRISPR-Cas-mediated plant disease and pest management. Plant Sci. 323, 111376. doi:10.1016/j.plantsci.2022.111376
Kenzhebayeva, S., Abekova, A., Atabayeva, S., Yernazarova, G., Omirbekova, N., Zhang, G., et al. (2019). Mutant lines of spring wheat with increased iron, zinc, and micronutrients in grains and enhanced bioavailability for human health. Biomed. Res. Int. 9692053, 9692053. doi:10.1155/2019/9692053
Khokhar, J. S., Sareen, S., Tyagi, B. S., Singh, G., Wilson, L., King, I. P., et al. (2018). Variation in grain Zn concentration, and the grain ionome, in field-grown Indian wheat. PLoS One 13, e0192026. doi:10.1371/journal.pone.0192026
Kirk, G. J. D., and Bajita, J. B. (1995). Root‐induced iron oxidation, pH changes and zinc solubilization in the rhizosphere of lowland rice. New Phytol. 131, 129–137. doi:10.1111/j.1469-8137.1995.tb03062.x
Krishnappa, G., Khan, H., Krishna, H., Kumar, S., Mishra, C. N., Parkash, O., et al. (2022). Genetic dissection of grain iron and zinc, and thousand kernel weight in wheat (Triticum aestivum L.) using genome-wide association study. Sci. Rep. 12, 12444. doi:10.1038/s41598-022-15992-z
Krishnappa, G., Singh, A. M., Chaudhary, S., Ahlawat, A. K., Singh, S. K., Shukla, R. B., et al. (2017). Molecular mapping of the grain iron and zinc concentration, protein content and thousand kernel weight in wheat (Triticum aestivum L.). PLoS One 12, e0174972. doi:10.1371/journal.pone.0174972
Kumar, A., Kaur, G., Goel, P., Bhati, K. K., Kaur, M., Shukla, V., et al. (2018). Genome-wide analysis of oligopeptide transporters and detailed characterization of yellow stripe transporter genes in hexaploid wheat. Funct. Integr. Genomics 19, 1 75–90. doi:10.1007/s10142-018-0629-5
Kumar, A. S., Jaganathan, D., Ramanathan, V., Rahman, H., Palaniswamy, R., Kambale, R., et al. (2020). Creation of novel alleles of fragrance gene OsBADH2 in rice through CRISPR/Cas9 mediated gene editing. PloS One 15, e0237018. doi:10.1371/journal.pone.0237018
Kumar, D., Yadav, A., Ahmad, R., Dwivedi, U. N., and Yadav, K. (2022). CRISPR-based genome editing for nutrient enrichment in crops: A promising approach toward global food security. Front. Genet. 3, 932859. doi:10.3389/fgene.2022.932859
Kumar, S., Kumar, N., Balyan, H. S., and Gupta, P. K. (2003). 1BL.1RS translocation in some Indian bread wheat genotypes and strategies for its use in future wheat breeding. Caryologia 56, 23–30. doi:10.1080/00087114.2003.10589303
Liu, D. Y., Liu, Y. M., Zhang, W., Chen, X. P., and Zou, C. Q. (2019a). Zinc uptake, translocation, and remobilization in winter wheat as affected by soil application of Zn fertilizer. Front. Plant Sci. 10, 426. doi:10.3389/fpls.2019.00426
Liu, J., Wua, B., Singh, R. P., and Velu, G. (2019b). QTL mapping for micronutrients concentration and yield component traits in a hexaploid wheat mapping population. J. Cereal Sci. 88, 57–64. doi:10.1016/j.jcs.2019.05.008
Liu, Y., Chen, Y., Yang, Y., Zhang, Q., Fu, B., Cai, J., et al. (2020). A thorough screening based on QTLs controlling zinc and copper accumulation in the grain of different wheat genotypes. Environ. Sci. Pollut. Res. Int. 28, 12 15043–15054. doi:10.1007/s11356-020-11690-3
Magallanes-Lopez, A. M., Hernandez-Espinosa, N., Velu, G., Posadas-Romano, G., Ordonez-Villegas, V. M. G., Crossa, J., et al. (2017). Variability in iron, zinc and phytic acid content in a worldwide collection of commercial durum wheat cultivars and the effect of reduced irrigation on these traits. Food Chem. 237, 499–505. doi:10.1016/j.foodchem.2017.05.110
Mageto, E. K., Crossa, J., Rodriguez, P. P., Dhliwayo, T., Palacios-Rojas, N., Lee, M., et al. (2020). Genomic prediction with genotype by environment interaction analysis for kernel zinc concentration in tropical maize germplasm. G3 10, 2629–2639. doi:10.1534/g3.120.401172
Marschner, H., and Romheld, V. (1994). Strategies of plants for acquisition of iron. Plant Soil 165, 261–274. doi:10.1007/bf00008069
Mayer, K. F. X., Rogers, J., Dolezel, J., Pozniak, C., Eversole, K., Feuillet, C., et al. (2014). A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 345, 1251788. doi:10.1126/science.1251788
Monasterio, J. I., and Graham, R. D. (2000). Breeding for trace minerals in wheat. Food Nutr. Bull. 21, 392–396. doi:10.1177/156482650002100409
Morgounov, A., Gomez-Becerra, H. F., Abugalieva, A., Dzhunusova, M., Yessimbekova, M., Muminjanov, H., et al. (2007). Iron and zinc grain density in common wheat grown in Central Asia. Euphytica 155, 193–203. doi:10.1007/s10681-006-9321-2
Mori, S., and Nishizawa, N. (1987). Methionine as a dominant precursor of phytosiderophores in Graminaceae plants. Plant Cell. Physiol. 28, 1081–1092.
Morris, E. R., and Ellis, R. (1989). Usefulness of the dietary phytic acid/zinc molar ratio as an index of zinc bioavailability to rats and humans. Biol. Trace Elem. Res. 19, 107–117. doi:10.1007/BF02925452
Mujeeb-Kazi, A. (1995). Interespecific crosses: Hybrid production and utilization. In Utilizing wild grass biodiversity in wheat improvement. 15 Years of wilde cross research at CIMMYT. Editors A. Mujeeb-Kazi, and G. P. Hettel (Mexico: CIMMYT Research Report Mexico, DF), 14–21.
Narendra, M. C., Roy, C., Kumar, S., Virk, P., and De, N. (2021). Effect of terminal heat stress on physiological traits, grain zinc and iron content in wheat (Triticum aestivum L.). Czech J. Genet. Plant Breed. 57, 43–50. doi:10.17221/63/2020-CJGPB
Neelam, K., Rawat, N., Tiwari, V., Prasad, R., Tripathi, S., Randhawa, G., et al. (2012). Evaluation and identification of wheat-Aegilopsaddition lines controlling high grain iron and zinc concentration and mugineic acid production. Cereal Res. Commun. 40, 53–61. doi:10.1556/crc.40.2012.1.7
Nozoye, T., Nagasaka, S., Kobayashi, T., Sato, Y., Uozumi, N., Nakanishi, H., et al. (2015). The phytosiderophore efflux transporter TOM2 is involved in metal transport in rice. J. Biol. Chem. 290, 27688–27699. doi:10.1074/jbc.M114.635193
Nozoye, T., Nagasaka, S., Kobayashi, T., Takahashi, M., Sato, Y., Sato, Y., et al. (2011). Phytosiderophore efflux transporters are crucial for iron acquisition in graminaceous plants. J. Biol. Chem. 286, 5446–5454. doi:10.1074/jbc.M110.180026
Ozturk, L., Yazici, M. A., Yucel, C., Torun, A., Cekic, C., Bagci, A., et al. (2006). Concentration and localization of zinc during seed development and germination in wheat. Physiol. Plant. 128, 144–152. doi:10.1111/j.1399-3054.2006.00737.x
Pahlavan-Rad, M., and Pessarakli, M. (2009). Response of wheat plants to zinc, iron and manganese applications and uptake and concentration of zinc, iron and manganese in wheat grains. Commun. Soil Sci. Plant Anal. 40, 1322–1332.
Pal, S., Datta, S. P., Rattan, R. K., and Singh, A. K. (2009). Diagnosis and amelioration of iron deficiency under aerobic rice. J. Plant Nutr. 31, 919–940. doi:10.1080/01904160802043262
Pearce, S., Tabbita, F., Cantu, D., Buffalo, V., Avni, R., Vazquez-Gross, H., et al. (2014). Regulation of Zn and Fe transporters by the GPC1 gene during early wheat monocarpic senescence. BMC Plant Biol. 14, 368. doi:10.1186/s12870-014-0368-2
Peleg, Z., Cakmak, I., Ozturk, L., Yazici, A., Jun, Y., Budak, H., et al. (2009). Quantitative trait loci conferring grain mineral nutrient concentrations in durum wheat x wild emmer wheat RIL population. Theor. Appl. Genet. 119, 353–369. doi:10.1007/s00122-009-1044-z
Peleg, Z., Saranga, Y., Yazici, A., Fahima, T., Ozturk, L., and Cakmak, I. (2007). Grain zinc, iron and protein concentrations and zinc-efficiency in wild emmer wheat under contrasting irrigation regimes. Plant Soil 306, 57–67. doi:10.1007/s11104-007-9417-z
Peng, F., Wang, C., Cheng, Y., Kang, H., Fan, X., Sha, L., et al. (2018). Cloning and characterization of TpNRAMP3, a metal transporter from polish wheat (Triticum polonicum L.). Front. Plant Sci. 9, 1354. doi:10.3389/fpls.2018.01354
Peterson, C. J., Johnson, V. A., and Mattern, P. J. (1986). Influence of cultivar and environment on mineral and protein concentrations of wheat flour, bran, and grain. Cereal Chem. 63, 118–186.
Phuke, R. M., He, X., Juliana, P., Kabir, M. R., Roy, K. K., Marza, F., et al. (2022). Identification of genomic regions and sources for wheat blast resistance through GWAS in Indian wheat genotypes. Genes. 13 (4), 596. doi:10.3390/genes13040596
Qiao, K., Wang, F., Liang, S., Wang, H., Hu, Z., and Chai, T. (2019). Improved Cd, Zn and Mn tolerance and reduced Cd accumulation in grains with wheat-based cell number regulator TaCNR2. Sci. Rep. 9, 870. doi:10.1038/s41598-018-37352-6
Raemaekers, R. H. (1988). in Helminthosporium sativum: Disease complex on wheat and sources of resistance in Zambia” in wheat production constraints in tropical environments. Editor A. R. Klatt (Mexico DF): CIMMYT), 175–186.
Rakotondramanana, M., Tanaka, R., Pariasca-Tanaka, J., Stangoulis, J., Grenier, C., and Wissuwa, M. (2022). Genomic prediction of zinc-biofortification potential in rice gene bank accessions. Theor. Appl. Genet. 135, 2265–2278. doi:10.1007/s00122-022-04110-2
Ramzan, Y., Hafeez, M. B., Khan, S., Nadeem, M., Rahman, S., Batool, S., et al. (2020). Biofortification with zinc and iron improves the grain quality and yield of wheat crop. Int. J. Plant Prod. 14, 501–510. doi:10.1007/s42106-020-00100-w
Rathan, N. D., Sehgal, D., Thiyagarajan, K., Singh, R., Singh, A. M., and Velu, G. (2021). Identification of genetic loci and candidate genes related to grain zinc and iron concentration using a zinc-enriched wheat ‘zinc-shakti. Front. Genet. 12, 652653. doi:10.3389/fgene.2021.652653
Rawat, N., Tiwari, V. K., Neelam, K., Randhawa, G. S., Singh, K., Chhuneja, P., et al. (2009a). Development and characterization of Triticum aestivum–Aegilops kotschyi amphiploids with high grain iron and zinc contents. Plant Genet. Resour. 7 (3), 271–280. doi:10.1017/s1479262109356592
Rawat, N., Tiwari, V. K., Singh, N., Randhawa, G. S., Singh, K., Chhuneja, P., et al. (2009b). Evaluation and utilization of Aegilops and wild Triticum species for enhancing iron and zinc content in wheat. Genet. Resour. Crop Evol. 56, 53–64. doi:10.1007/s10722-008-9344-8
Rizwan, M., Ali, S., Ali, B., Adrees, M., Arshad, M., Hussain, A., et al. (2018). Zinc and iron oxide nanoparticles improved the plant growth and reduced the oxidative stress and cadmium concentration in wheat. Chemosphere 214, 269–277. doi:10.1016/j.chemosphere.2018.09.120
Roy, C., Chattopadhyay, T., Ranjan, R. D., Hasan, W. U., Kumar, A., and De, N. (2021a). Association of leaf chlorophyll content with the stay-green trait and grain yield in wheat grown under heat stress conditions. Czech J. Genet. Plant Breed. 57, 140–148. doi:10.17221/45/2021-CJGPB
Roy, C., Juliana, P., Kabir, M. R., Roy, K. K., Gahtyari, N. C., Marza, F., et al. (2021b). New genotypes and genomic regions for resistance to wheat blast in South asian germplasm. Plants 10, 2693. doi:10.3390/plants10122693
Rupa, T. R., and Tomar, K. P. (1999). Zinc desorption kinetics as influenced by pH and phosphorus in soils. Commun. Soil Sci. Plant Anal. 30, 1951–1962. doi:10.1080/00103629909370345
Sánchez-León, S., Gil-Humanes, J., Ozuna, C. V., Giménez, M. J., Sousa, C., Voytas, D. F., et al. (2018). Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol. J. 16, 902–910. doi:10.1111/pbi.12837
Shariatipour, N., Heidari, B., and Richards, C. M. (2021). Meta-analysis of QTLome for grain zinc and iron contents in wheat (Triticum aestivum L.). Euphytica 217, 86. doi:10.1007/s10681-021-02818-8
Sharma, S., Kaur, G., Kumar, A., Meena, V., Kaur, J., and Pandey, A. K. (2019). Overlapping transcriptional expression response of wheat zinc-induced facilitator-like transporters emphasize important role during Fe and Zn stress. BMC Mol. Biol. 20, 22. doi:10.1186/s12867-019-0139-6
Sharma, S., Kaur, G., Kumar, A., Meena, V., Ram, H., Kaur, J., et al. (2020). Gene expression pattern of vacuolar-iron transporter-like (VTL) genes in hexaploid wheat during metal stress. Plants 9, 229. doi:10.3390/plants9020229
Shi, R., Li, H., Tong, Y., Jing, R., Zhang, F., and Zou, C. (2008). Identification of quantitative trait locus of zinc and phosphorus density in wheat (Triticum aestivum L.) grain. Plant Soil 306, 95–104. doi:10.1007/s11104-007-9483-2
Shojima, S., Nishizawa, N. K., Fushiya, S., Nozoe, S., Irifune, T., and Mori, S. (1990). Biosynthesis of phytosiderophores: In vitro biosynthesis of 2-deoxymugineic acid from l-methionine and nicotianamine. Plant Physiol. 93, 1497–1503. doi:10.1104/pp.93.4.1497
Singh, A., Karjagi, C., and Rakshit, S. (2020). Minimally altering a critical kinase for low-phytate maize. Sci. Rep. 10, 6324. doi:10.1038/s41598-020-63016-5
Singh, B. R., Timsina, Y. N., Lind, O. C., Cagno, S., and Janssens, K. (2018). Zinc and iron concentration as affected by nitrogen fertilization and their localization in wheat grain. Front. Plant Sci. 9, 307. doi:10.3389/fpls.2018.00307
Singh, D., Wang, X., Kumar, U., Gao, L., Noor, M., Imtiaz, M., et al. (2019). High-throughput phenotyping enabled genetic dissection of crop lodging in wheat. Front. Plant Sci. 10, 394. doi:10.3389/fpls.2019.00394
Singh, P. K., Gahtyari, N. C., Roy, C., Roy, K. K., He, X., Tembo, B., et al. (2021). Wheat blast: A disease spreading by intercontinental jumps and its management strategies. Front. Plant Sci. 12, 710707. doi:10.3389/fpls.2021.710707
Singh, S. P., Keller, B., Gruissem, W., and Bhullar, N. K. (2017). Rice NICOTIANAMINE SYNTHASE 2 expression improves dietary iron and zinc levels in wheat. Theor. Appl. Genet. 130, 283–292. doi:10.1007/s00122-016-2808-x
Sperotto, R. A. (2013). Zn/Fe remobilization from vegetative tissues to rice seeds: should I stay or should I go? Ask Zn/Fe supply. Front. Plant Sci. 4, 464. doi:10.3389/fpls.2013.00464
Srinivasa, J., Arun, B., Mishra, V. K., Singh, G. P., Velu, G., Babu, R., et al. (2014). Zinc and iron concentration QTL mapped in a Triticum spelta × T. aestivum cross. Theor. Appl. Genet. 127, 1643–1651. doi:10.1007/s00122-014-2327-6
Stipesevic, B., Jug, D., Jug, I., Tolimir, M., and Cvijovic, M. (2009). Winter wheat and soybean zinc uptake in different soil tillage systems. Cereal Res. Commun. 37, 305–310. doi:10.1556/crc.37.2009.2.19
Tan, J., Wang, J., Chai, T., Zhang, Y., Feng, S., Li, Y., et al. (2013). Functional analyses of TaHMA2, a P1B-type ATPase in wheat. Plant Biotechnol. J. 11, 420–431. doi:10.1111/pbi.12027
Tang, J. W., Zou, C. Q., He, Z. H., Shi, R., Ortiz-Monasterio, I., Qu, Y., et al. (2008). Mineral element distributions in milling fractions of Chinese wheats. J. Cereal Sci. 48, 821–828. doi:10.1016/j.jcs.2008.06.008
Tiwari, V. K., Rawat, N., Chhuneja, P., Neelam, K., Aggarwal, R., Randhawa, G. S., et al. (2009). Mapping of quantitative trait Loci for grain iron and zinc concentration in diploid A genome wheat. J. Hered. 100, 771–776. doi:10.1093/jhered/esp030
Tiwari, V. K., Rawat, N., Neelam, K., Kumar, S., Randhawa, G. S., and Dhaliwal, H. S. (2010). Substitutions of 2S and 7U chromosomes of Aegilops kotschyi in wheat enhance grain iron and zinc concentration. Theor. Appl. Genet. 121, 259–269. doi:10.1007/s00122-010-1307-8
Tiwari, V. K., Rawat, N., Neelam, K., Randhawa, G. S., Singh, K., Chhuneja, P., et al. (2008). Development of Triticum turgidum ssp. durum–Aegilops longissima amphiploids with high iron and zinc content through unreduced gamete formation in F1 hybrids. Genome 51, 757–766. doi:10.1139/G08-057
Trijatmiko, K. R., Dueñas, C., Tsakirpaloglou, N., Torrizo, L., Arines, F. M., Adeva, C., et al. (2016). Biofortified indica rice attains iron and zinc nutrition dietary targets in the field. Sci. Rep. 6, 19792. doi:10.1038/srep19792
Uauy, C., Distelfeld, A., Fahima, T., Blechl, A., and Dubcovsky, J. (2006). A NAC gene regulating senescence improves grain protein, zinc, and iron content in wheat. Science 314, 1298–1301. doi:10.1126/science.1133649
Vatansever, R., Filiz, E., and Eroglu, S. (2017). Genome-wide exploration of metal tolerance protein (MTP) genes in common wheat (Triticum aestivum): Insights into metal homeostasis and biofortification. Biometals 30, 217–235. doi:10.1007/s10534-017-9997-x
Velu, G., Crespo-Herrera, L., Guzman, C., Huerta, J., Payne, T., and Singh, R. P. (2019). Assessing genetic diversity to breed competitive biofortified wheat with enhanced grain Zn and Fe concentrations. Front. Plant Sci. 9, 1971. doi:10.3389/fpls.2018.01971
Velu, G., Guzman, C., Mondal, S., Autrique, J. E., Huerta, J., and Singh, R. P. (2016a). Effect of drought and elevated temperature on grain zinc and iron concentrations in CIMMYT spring wheat. J. Cereal Sci. 69, 182–186. doi:10.1016/j.jcs.2016.03.006
Velu, G., Ortiz-Mona, I., Singh, R. P., and Payne, T. (2011). Variation for grain micronutrients concentration in wheat core-collection accessions of diverse origin. Asian J. Crop Sci. 3, 43–48. doi:10.3923/ajcs.2011.43.48
Velu, G., Ortiz-Monasterio, I., Cakmak, I., Hao, Y., and Singh, R. P. (2014). Biofortification strategies to increase grain zinc and iron concentrations in wheat. J. Cereal Sci. 59, 365–372. doi:10.1016/j.jcs.2013.09.001
Velu, G., Singh, R. P., Crespo-Herrera, L., Juliana, P., Dreisigacker, S., Valluru, R., et al. (2018). Genetic dissection of grain zinc concentration in spring wheat for mainstreaming biofortification in CIMMYT wheat breeding. Sci. Rep. 8, 13526. doi:10.1038/s41598-018-31951-z
Velu, G., Tutus, Y., Gomez-Becerra, H. F., Hao, Y., Demir, L., Kara, R., et al. (2016b). QTL mapping for grain zinc and iron concentrations and zinc efficiency in a tetraploid and hexaploid wheat mapping populations. Plant Soil 411, 81–99. doi:10.1007/s11104-016-3025-8
Virk, P. S., Andersson, M. S., Arcos, J., Govindaraj, M., and Pfeiffer, W. H. (2021). Transition from targeted breeding to mainstreaming of biofortification traits in crop improvement programs. Front. Plant Sci. 12, 703990. doi:10.3389/fpls.2021.703990
Vishwakarma, M. K., Mishra, V. K., Gupta, P. K., Yadav, P. S., Kumar, H., and Joshi, A. K. (2014). Introgression of the high grain protein gene Gpc-B1 in an elite wheat variety of Indo-Gangetic Plains through marker assisted backcross breeding. Curr. Plant Biol. 1, 60–67. doi:10.1016/j.cpb.2014.09.003
Voss-Fels, K. P., Herzog, E., Dreisigacker, S., Sukumaran, S., Watson, A., Frisch, M., et al. (2019). in SpeedGS to accelerate genetic gain in spring wheat” in applications of genetic and genomic research in cereals. Editors T. Miedaner, and V. Korzun (Woodhead Publishing-Elsevier), 303–327. doi:10.1007/978-3-030-90673-3_30
Vreugdenhil, M., Akkermans, M. D., van der Merwe, L. F., van Elburg, R. M., van Goudoever, J. B., and Brus, F. (2021). Prevalence of zinc deficiency in healthy 1–3-year-old children from three western European countries. Nutrients 13, 3713. doi:10.3390/nu13113713
Wang, S., Yang, Y., Guo, M., Zhong, C., Yan, C., and Sun, S. (2020). Targeted mutagenesis of amino acid transporter genes for rice quality improvement using the CRISPR/Cas9 system. Crop J. 8, 457–464. doi:10.1016/j.cj.2020.02.005
Wang, Y., Xu, X., Hao, Y., Zhang, Y., Liu, Y., Pu, Z., et al. (2021). QTL mapping for grain zinc and iron concentrations in bread wheat. Front. Nutr. 8, 680391. doi:10.3389/fnut.2021.680391
Watson, A., Ghosh, S., Williams, M. J., Cuddy, W. S., Simmonds, J., Rey, M.-D., et al. (2018). Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 4, 23–29. doi:10.1038/s41477-017-0083-8
Wen, Z., Juliana, P., Dhugga, H. S., Pacheco, M., Martínez, U. I., Aguilar, A., et al. (2022). Genome-wide association study of phytic acid in wheat grain unravels markers for improving biofortification. Front. Plant Sci. 13, 830147. doi:10.3389/fpls.2022.830147
Williamson, V. M., Thomas, V., Ferris, H., and Dubcovsky, J. (2013). An Aegilops ventricosa translocation confers resistance against root-knot nematodes to common wheat. Crop Sci. 53, 1412–1418. doi:10.2135/cropsci2012.12.0681
Wu, C. Y., Lu, L. L., Yang, X. E., Feng, Y., Wei, Y. Y., Hao, H. L., et al. (2010). Uptake, translocation, and remobilization of zinc absorbed at different growth stages by rice genotypes of different Zn densities. J. Agric. Food Chem. 58, 6767–6773. doi:10.1021/jf100017e
Ye, X., Al-babili, S., Kloti, A., Zhang, J., Lucca, P., Beyer, P., et al. (2000). Engineering the provitamin A (β-Carotene) biosynthetic pathway into (Carotenoid-Free) rice endosperm. Science 287, 303–305. doi:10.1126/science.287.5451.303
Yilmaz, A., Ekiz, H., Torun, B., Gultekin, I., Karanlik, S., Bagci, S. A., et al. (1997). Effect of different zinc application methods on grain yield and zinc concentration in wheat cultivars grown on zinc-deficient calcareous soils. J. Plant Nutr. 20, 461–465 461471. doi:10.1080/01904169709365267
Zhang, Y., Shi, R., Rezaul, K. M. D., Zhang, F., and Zou, C. (2010). Iron and zinc concentrations in grain and flour of winter wheat as affected by foliar application. J. Agric. Food Chem. 58, 12268–12274. doi:10.1021/jf103039k
Zhao, D., Zhang, B., Chen, W., Liu, B., Zhang, L., Zhang, H., et al. (2017). Comparison of zinc, iron, and selenium accumulation between synthetic hexaploid wheat and its tetraploid and diploid parents. Can. J. Plant Sci. 97, 692–701. doi:10.1139/cjps-2016-0320
Zhou, M., Zheng, S., Liu, R., Lu, L., Zhang, C., Zhang, L., et al. (2019). The genome-wide impact of cadmium on microRNA and mRNA expression in contrasting Cd responsive wheat genotypes. BMC Genomics 20, 615. doi:10.1186/s12864-019-5939-z
Keywords: malnutrition, QTL mapping, GWAS-genome-wide association study, speed breeding, new breeding techniques (NBTs), biofortification, genomic selection
Citation: Roy C, Kumar S, Ranjan RD, Kumhar SR and Govindan V (2022) Genomic approaches for improving grain zinc and iron content in wheat. Front. Genet. 13:1045955. doi: 10.3389/fgene.2022.1045955
Received: 16 September 2022; Accepted: 24 October 2022;
Published: 08 November 2022.
Edited by:
Zeba I. Seraj, University of Dhaka, BangladeshCopyright © 2022 Roy, Kumar, Ranjan, Kumhar and Govindan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chandan Roy, Y2hhbmRhbi5yb3k0M0BnbWFpbC5jb20=; Velu Govindan, dmVsdUBjZ2lhci5vcmc=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.