 Yuanyuan Lv
Yuanyuan Lv Shan Huang3
Shan Huang3 Tianjiao Zhang
Tianjiao Zhang Bo Gao
Bo Gao
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Genet. , 31 March 2021
Sec. Computational Genomics
Volume 12 - 2021 | https://doi.org/10.3389/fgene.2021.664860
This article is part of the Research Topic Computational Learning Models and Methods Driven by Omics for Biology for “The Fifth China Computer Society Bioinformatics Conference” View all 15 articles
Multilayer networks provide an efficient tool for studying complex systems, and with current, dramatic development of bioinformatics tools and accumulation of data, researchers have applied network concepts to all aspects of research problems in the field of biology. Addressing the combination of multilayer networks and bioinformatics, through summarizing the applications of multilayer network models in bioinformatics, this review classifies applications and presents a summary of the latest results. Among them, we classify the applications of multilayer networks according to the object of study. Furthermore, because of the systemic nature of biology, we classify the subjects into several hierarchical categories, such as cells, tissues, organs, and groups, according to the hierarchical nature of biological composition. On the basis of the complexity of biological systems, we selected brain research for a detailed explanation. We describe the application of multilayer networks and chronological networks in brain research to demonstrate the primary ideas associated with the application of multilayer networks in biological studies. Finally, we mention a quality assessment method focusing on multilayer and single-layer networks as an evaluation method emphasizing network studies.
In recent years, the formulation of multilayer networks has provided new methods for the study of multilevel network systems. Many biological systems comprise interconnected units that can be effectively modeled as networks, which are mathematical structures describing connections between points (Jing et al., 2019; Liu B. et al., 2020; Shao et al., 2020). Complex network systems provide powerful research tools and methods for studying biological fields (Kumari and Verma, 2020; Liu X. et al., 2020; Shao and Liu, 2020), from interactions between genes and proteins (Zhang et al., 2019; Li Z. et al., 2020; Zhai et al., 2020), to the study of tissue and organ functions (Yang et al., 2020), and even human brain study (Zhang J. et al., 2020). The complexity and evolutionary nature of biological systems enable the extensive application of multilayer networks and associated methods. Additionally, ecosystems and evolutionary systems evolve and change over time, and the corresponding network structures for these systems change correspondingly. Furthermore, the reasons for all these changes, particularly topological changes in the course of network structure change, and the importance of network feedback in network structure analyses are all topics worthy of exploration.
A network representation is a simplified description of a more complex, multifaceted system. A social system can include different types of interactions of different biological significance (e.g., cooperation or competition), while standard network approaches usually ignore these interactions or achieve integration through analyzing networks with different edge types separately. In bioinformatics studies using network structures, the progress of each biological system relies on the amount of data and/or new discoveries about unknown biological areas. For example, in the study of transcription-translation relationships between genes and proteins, genes and proteins are represented by nodes, and the correspondence between genes and proteins is represented by links in the network. Therefore, it is necessary to first understand the characteristics of each individual gene and protein, and the methods used to identify these relationships (Lin et al., 2019; Zhang D. et al., 2021; Zhang Z.Y. et al., 2021; Zulfiqar et al., 2021). Only then can the most relevant genes and corresponding proteins for a disease or symptom be identified through clustering or linkage analyses of the network, which further enables the investigation of target therapies for symptoms of disease (Zhu et al., 2018; Iliopoulos et al., 2020). These applications all rely on the data set and on the biological correspondence of genes and proteins.
The definition of a multilayer network varies slightly from one application to another. All edges and nodes in a single network are homogeneous, but in the real world, there is heterogeneity in both the objects represented by the nodes and the connections represented by the edges. Multilayer networks add additional tagging capabilities to traditional networks. That is, tagging terms are added to the traditional network, which can be understood as a composite of simple (single-layer) networks with different tags for complex networks. This is a relatively easy way to understand the definition of complex networks on different systems. Currently, according to different applications and subjects, multilayer networks can be divided into the following types:
(1) Multiplex networks: Networks in which the nodes on different layers are connected by inter-layer edges.
(a) In multi-relational networks, each layer represents a different type of interaction, i.e., different relationships are the distinguishing dimension for building a multilayer network, and the relationships are the tagged labels.
(b) In a temporal network, each layer encodes the same type of interaction at different time points or time windows. That is, time series (time windows) are the tagged labels between layers in a multilayer network.
(2) Interconnected networks: Nodes in different layers do not necessarily represent the same entities and inter-layer edges between different types of nodes may exist.
(a) The network of networks consists of subsystems, which are themselves networks. They are interconnected by interlayer edges between subsystem nodes.
(b) In a contextual network, each layer is interpreted to represent a different type of node. For example, interactions between males in one layer, interactions between females in another layer, and interactions between the sexes in a third layer. These interactions are represented by inter-layer edges.
(c) Spatial networks (also known as geometric networks) can be connected by ecological networks of the subjects moving between various locations.
Multilayer networks are currently used in various fields including physics, chemistry, biology, technology, finance, and social systems because of their inherent structural and functional characteristics. In this review, we briefly introduce the development of multilayer network concepts, techniques, and applications in bioinformatics by reviewing multilayer network applications in bioinformatics, and we summarize the outlook and development of multilayer networks in bioinformatics by analyzing current research.
The definition and methodology of multilayer network models in bioinformatics depends on the specific research problem. Organisms can be classified into different systems under different levels, and that system usually changes dynamically with time. Therefore, usually the representation of bioinformatics related networks varies depending on the specific biological system. In this review, we classify research topics into different categories according to the different levels of biological systems. As shown in Figure 1, multilayer networks in bioinformatics can be classified into five major categories.
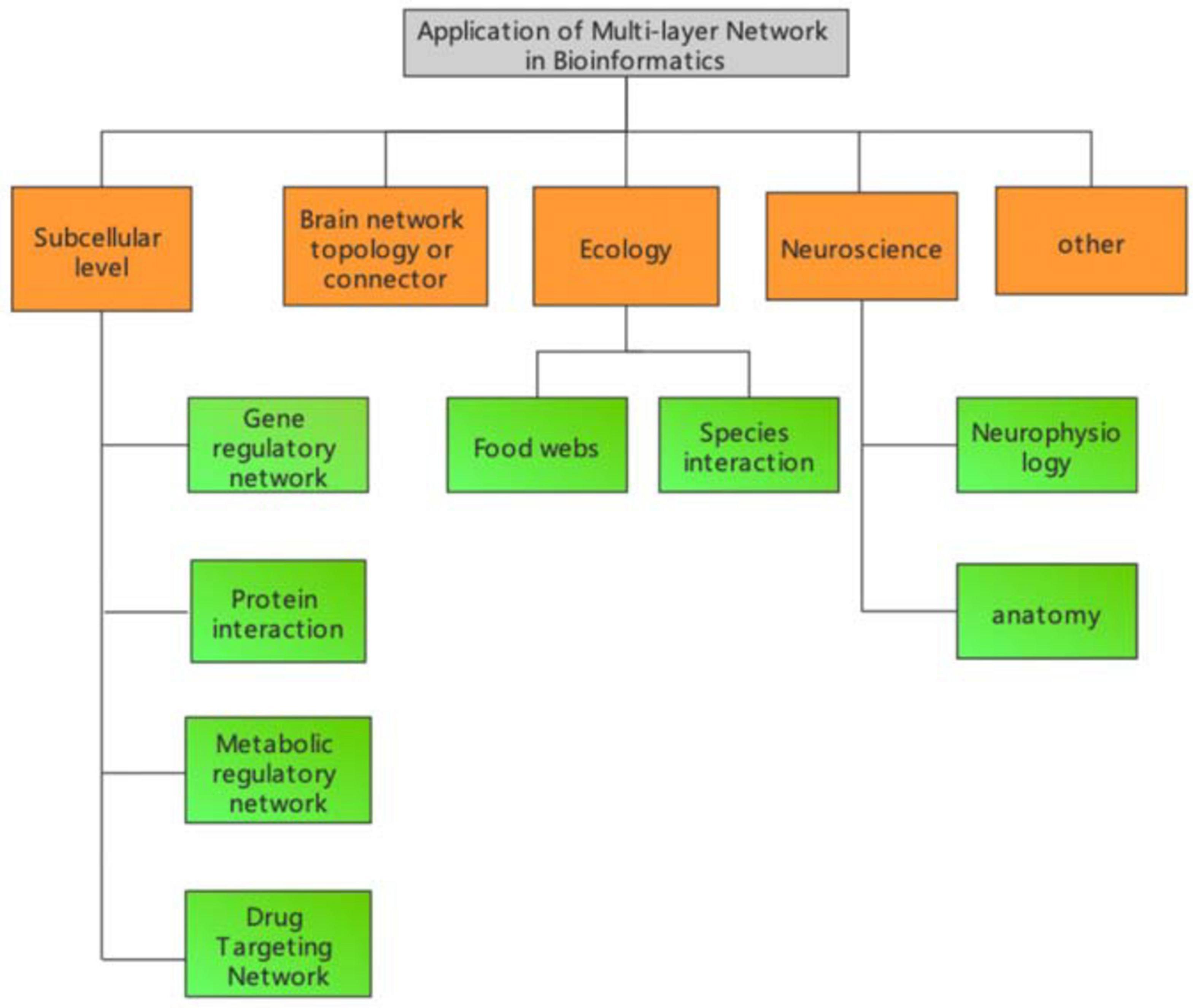
Figure 1. Multilayer network application area framework in bioinformatics.
As the understanding of DNA structure and function has gradually improved (Liu M.L. et al., 2020), understanding the relationships between genes and proteins, genes and disease, and disease and drugs has greatly evolved. For example, the correspondence between genes and disease has been mined through network structures, where the method utilizes a joint learning approach using the functional and connectivity patterns of proteins to predict disease-gene relationships using human interactome networks. In contrast to other data structures, interactomes are characterized by a high degree of incompleteness and lack of explicit negative knowledge, which makes prediction particularly challenging. To maximize potential information in the network, a second-order random walk procedure named random walker (RW^2) is applied in these studies. The random walker is able to learn rich representations of disease gene (or gene product) characteristics. This method has successfully compared with the best-known disease gene prediction systems and other state-of-the-art graph-based methods.
A large number of candidate disease-causing genes can be sequenced and checked for variation to help determine relationships between disease and genes (Zhang Z.M. et al., 2020). Many different computational methods have been developed to address this challenge. The observation that genes associated with similar diseases have a higher probability of interaction, many of these methods rely on the analysis of the topological properties of biological networks. However, the incomplete and noisy nature of biological networks is an important challenge. Two-step framework for disease gene ranking: (1) Construct a reliable functional connectivity network using sequence information and machine learning techniques. And (2) rank disease–gene relationships on the basis of that constructed functional connectivity network. Unlike other functional connectivity network-based frameworks that use functional connectivity networks based on the integration of various low-quality biological data, protein sequences can be used as comprehensive data to construct a reliable initial network. Additionally, the physicochemical properties of amino acids can be used to supplement hypotheses of protein function. In conclusion, our assessment of these methods indicates high efficiency and effectiveness for constructing functional linkage networks for disease genes (Wang et al., 2008; Jiang et al., 2010, 2013; Cheng et al., 2018; Zeng et al., 2018).
Gene function can also be determined by collecting biological data. For example, the Drosophila ovary epidermal cells (ECs) externally control the maintenance and progeny differentiation of germ line stem cells (GSC). In this study, the role of 173 EC genes that control GSC maintenance and progeny differentiation were identified using a Drosophila in vivo systemic RNAi approach (Zeng et al., 2016; Zou et al., 2016; Wang et al., 2019). Among the identified genes, 10 and 163 genes were required by ECs for GSC maintenance and progeny differentiation, respectively. The genes required for progeny differentiation were classified into different functional categories, including transcription, mRNA splicing, protein degradation, signal transduction, and cytoskeleton regulation (Cao et al., 2019). In addition, GSC progeny differentiation defects caused by defective ECs were often associated with BMP signaling elevation, indicating that preventing BMP signaling is a general functional feature of the differentiation niche. Finally, EC exon junction complex (EJC) components were identified as required for EC maintenance and the prevention of BMP signaling, and thus the promotion of GSC progeny differentiation. Therefore, this study identifies the major regulators of the differentiation niche and provides important insights into the external control of stem cell progeny differentiation.
Corresponding network structures for different biological data and specific subjects can also be designed to analyse specific biological systems (Zeng et al., 2016; Jiang et al., 2017; Liu et al., 2017). Currently, at the subcellular level, these networks mainly include gene regulatory networks (Wang et al., 2010; Ding et al., 2011; Jiang et al., 2014; Cheng et al., 2019; Konda et al., 2019; Liu L. et al., 2019; Mortezaeefar et al., 2019; Hong et al., 2020), protein functional networks (Guo et al., 2011, 2013, 2014; Sikandar et al., 2019; Tao et al., 2020; Liu et al., 2021), metabolic regulatory networks (Jin et al., 2020), and drug targeting networks (Wei et al., 2014; Ding et al., 2017, 2019, 2020a,b; Jin Q. et al., 2019; Jin S. et al., 2019; Srivastava et al., 2019; Zhao et al., 2019; Zeng et al., 2020).
The study of human brain functional and structural mechanisms using brain networks is also a hot field. Currently, research has mainly studied brain function by acquiring the brain waves of subjects, and the functional partitions of the brain have been predominantly obtained by functional experiments or magneto encephalography. This portion of our review will be introduced in detail in the next section.
Modern network theory is increasingly applied to neuroscience to understand neurophysiology and anatomy at different scales and under experimentally attainable physiological and pathophysiological conditions. The first attempt was made at the micro anatomical level of individual neurons. Watts and Strogatz analyzed the anatomical connections of the nervous system of Caenorhabditis elegans where neurons represent the nodes and synaptic or gap connections of a neural network. Their study revealed a highly clustered and efficient network, thus representing the first evidence of a real neural system with a small-world network. Later graph-theoretical approaches focused on morphological representations or dynamic correlations of the electrical stimulation activity of neuronal networks.
Network-based analyses have also been useful to address several questions in ecology and issues in conservation. The first study was carried out in a contextual network of so-called species interactions. Food webs are one of the fundamental issues in ecological studies, and despite the rather high variability detected in network structure, food webs present a complex topology similar to other types of real networks and host-parasite networks. One of the main advantages of these approaches is the direct assessment of the robustness and sensitivity of a given ecosystem to species loss or other perturbations. Another network type widely used in ecology is connected landscape mapping, where nodes typically represent patches on the landscape. The resulting spatial networks describe the linkages between processes and patterns that characterize the landscape, thus providing an effective way to assess important issues such as the effects of species dispersal or habitat loss.
Understanding the interactions between different species in a community and responses to environmental change is a central goal of ecology. However, defining the network structure of microbial communities is very challenging because of associated extremely diverse and unexplored states. Although recent developments in metagenomic technologies, such as high-throughput sequencing and functional gene arrays, have provided revolutionary tools for analyzing microbial community structure, it is still difficult to study network interactions in microbial communities based on high-throughput meta genomic data. A mathematical and bioinformatics framework for constructing molecular ecological networks (MENs) based on Random Matrix Theory (RMT) has been proposed. The remarkable feature of this approach compared with other network construction methods is that the network is automatically defined and robust to noise, thus providing a good solution to several common problems associated with high throughput.
The brain is the control center of most animal activities, and it has been the goal of many researchers to unravel the mystery of the brain and simulate the human brain with external devices such as computers. Before that, the structure and mechanism of the brain needs to be clarified, and it is costly to study the human brain because of its complexity. The human brain is a complex system organized by the structural and functional relationships among its components (Liu et al., 2018; Song et al., 2018; Liu G. et al., 2019). Recent experimental advances have led to unprecedented amounts of data that describe the structure and function of the brain, and it is now possible to model the brain as a network by measuring pairwise interactions between its various units. This modeling can be performed across multiple scales, where network nodes represent units of the brain, including proteins, neurons, brain regions, or other measurement units. Recording techniques such as functional magnetic resonance imaging (fMRI), magneto encephalography (MEG), and electroencephalography (EEG) are capable of capturing brain dynamics across time and across multiple frequency bands.
Recent neuroscience research has also exploited the versatility of multilayer frameworks to model complex relationships in neural data. For example, given fMRI and diffusion tensor imaging (DTI) for a single subject, a multilayer network can be constructed, with one layer representing the fMRI network and another layer representing the DTI network. Using the fMRI data, a functional network can be constructed in which the nodes represent brain regions and the edges represent the coherence between regional activities. On the basis of DTI data, a structural network can be constructed by dividing the brain into regions and then measuring the strength of physical connections between these regions. Finally, considering each network as a layer in a multilayer network, the edges of a brain region in the fMRI layer can be added to the DTI layer to form a multilayer network.
The brain is an inherently dynamic system, and the performance of cognition requires dynamic reconfiguration of a highly evolved network of brain regions, which interact in complex and transient communications. However, an accurate description of these reconfiguration processes during human cognitive function remains elusive (Liu and Jiang, 2016). Therefore, many studies have used temporal networks to investigate the dynamic cluster structure of brain networks and reveal the underlying human brain dynamic changes during learning. Temporal networks that contain temporal information have the advantage of retaining the full information of the data without aggregating connections into individual networks.
When we complete different cognitive vision tasks, we subdivide the regional time series into time windows of variable size, and determine the impact of the time window size on the observed dynamics. Specifically, we applied a multilayer community detection algorithm to identify temporal communities, and we computed network flexibility to quantify the changes in these communities over time. Within our frequency band of interest, large and small time windows were associated with the brain network flexibility within a narrow range, while medium time windows were associated with broad network flexibility. Using medium time windows of 75–100 s, we identified brain regions with low flexibility (considered core regions and observed in visual and attentional areas) and brain regions with high flexibility (considered peripheral regions and observed in subcortical and temporal lobe regions) by comparison with appropriate control dynamic network models. In general, this work demonstrates the effect of time window size on the network dynamics observed during task execution, providing practical considerations when selecting time windows in dynamic network analysis. More generally, this work reveals organizing principles for functional brain connections that are inaccessible to static network approaches.
The hypothesis that human executive functions arise from the dynamic interactions of multiple networks has been tested in previous research (Ding et al., 2019). To corroborate this research, we investigated a key executive function (FCD), namely arbitrary visuomotor mapping. MEG and intracranial EEG were recorded using high gamma activity brain connectivity analysis. We then generated visuomotor mapping using the dynamic interactions of three partially overlapping cortico-cortical and cortico-subcortical functional connectivity (FC) networks. First, visual and parietal regions were coordinated with sensorimotor and premotor regions. Second, dorsal fronto parietal circuits dominated by sensorimotor and associative frontal striatal networks were incorporated. Finally, bilateral sensory-motor areas were coordinated with the cortico-cortical hemisphere between the left fronto parietal network and the visual areas. Our study argued that these networks reflect the processing of visual information, the emergence of visuomotor plans, and the processing of somatosensory responses or action outcomes. Thus, our study demonstrates that visuomotor integration exists in the dynamic reconfiguration of multiple cortico-cortical and cortico-subcortical FC networks. More generally, the approach demonstrates that optokinetic-related FC is unstable and shows task performance-related switching dynamics and regional flexibility on a time scale. In addition, our optokinetic-related FC has sparse connectivity with a density of 10%. Taken together, these findings shed light on the relationship between dynamic network reconfiguration and short-time executive function and provide a candidate start point for the better understanding of cognitive structure.
A vast number of multilayer network applications exists in bioinformatics, but the application of multilayer networks in any subfield of bioinformatics still relies on the acquisition and accumulation of bioinformatics data, and brain research is no exception. Therefore, interdisciplinary collaboration is a very efficient and necessary option. Brain structure and functionality are gradually understood, driven by brain data acquisition. According to these studies, the dynamic modeling of brain function by combining temporal dimensions is an effective means of study. Perhaps as research progresses, new data dimensions will be added (Wang et al., 2018, 2020; Wei et al., 2018a; Ding et al., 2019; Liu B. et al., 2019; Su et al., 2019b; Dao et al., 2020; Li J. et al., 2020; Lv et al., 2020).
Multilayer (complex) networks have been an effective tool for studying complex problems in recent years and are currently being used in a variety of fields. As systems biology develops, multilayer networks are applicable to many aspects and research areas within the field. Because of dataset availability, these networks are currently more often applied to genetics and brain research. However, as research progresses, it should become easier to unravel structural and functional fogs in biology on one hand, and on the other hand, research in this area will prove beneficial to the understanding of biological principles in general to better serve all people. In view of current research status, our review has presented the following ideas and prospects:
(1) The development of biology is promoted by the joint development of various fields, and the application of multilayer networks in bioinformatics depends on the accumulation of biological data and the development of computer-related theories. Therefore, as an interdisciplinary subject, it needs the collaborative work of interdisciplinary experts.
(2) Because of the complexity and dynamic change of biological systems, time-series multilayer networks with the addition of temporal information will have more and more applications in the simulation of dynamic processes in the study of genes, disease, drug discovery, and brain research.
(3) Exploring the communication mode between tissue cells in the form of multi-layer network is to study the interaction (functionality) between structures on the basis of the network represented by the structure.
In addition to the structural and functional aspects of multilayer network research, methods to efficiently evaluate and assess the results of multilayer networks remains an importan tissue. The evaluation of the algorithmic complexity of multilayer networks has been proposed to assess if and when the multilayer representation of a system is qualitatively superior to classical single-layer aggregation network approaches (Wei et al., 2017a,b,c, 2018b, 2019; Su et al., 2019a, 2020a,b; Wang D. et al., 2021; Wang H. et al., 2021; Zhao et al., 2017).
YL contributed to conception and design of the study and wrote the first draft of the manuscript. SH organized the database. BG performed the statistical analysis. TZ, YL, and BG wrote sections of the manuscript. All authors contributed to manuscript revision, read, and approved the submitted version.
This work was supported by the National Natural Science Foundation of China (No. 62002087).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Steven M. Thompson, from LiwenBianji, Edanz Editing China (www.liwenbianji.cn/ac), for editing the English text of a draft of this manuscript.
Cao, S., Wang, Y., and Tang, Z. (2019). Adaptive elman model of gene regulation network based on time series data. Curr. Bioinform. 14, 551–561. doi: 10.2174/1574893614666190126145431
Cheng, L., Hu, Y., Sun, J., Zhou, M., and Jiang, Q. (2018). DincRNA: a comprehensive web-based bioinformatics toolkit for exploring disease associations and ncRNA function. Bioinformatics 34, 1953–1956. doi: 10.1093/bioinformatics/bty002
Cheng, L., Wang, P., Tian, R., Wang, S., Guo, Q., Luo, M., et al. (2019). LncRNA2Target v2.0: a comprehensive database for target genes of lncRNAs in human and mouse. Nucleic Acids Res. 47, D140–D144.
Dao, F. Y., Lv, H., Zhang, D., Zhang, Z. M., Liu, L., and Lin, H. (2020). DeepYY1: a deep learning approach to identify YY1-mediated chromatin loops. Brief. Bioinform. doi: 10.1093/bib/bbaa356 Online ahead of print.
Ding, H., Luo, L. F., and Lin, H. (2011). Entropy production rate changes in lysogeny/lysis switch regulation of Bacteriophage Lambda. Commun. Theor. Phys. 55, 371–375. doi: 10.1088/0253-6102/55/2/31
Ding, Y., Tang, J., and Guo, F. (2017). Identification of drug-target interactions via multiple information integration. Inform. Sci. 418, 546–560. doi: 10.1016/j.ins.2017.08.045
Ding, Y., Tang, J., and Guo, F. (2019). Identification of drug-side effect association via multiple information integration with centered kernel alignment. Neurocomputing 325, 211–224. doi: 10.1016/j.neucom.2018.10.028
Ding, Y., Tang, J., and Guo, F. (2020a). Identification of drug-target interactions via dual laplacian regularized least squares with multiple kernel fusion. Knowl. Based Syst. 204:106254. doi: 10.1016/j.knosys.2020.106254
Ding, Y., Tang, J., and Guo, F. (2020b). Identification of drug-target interactions via fuzzy bipartite local model. Neural Comput. Appl. 23, 10303–10319. doi: 10.1007/s00521-019-04569-z
Guo, F., Li, S. C., and Wang, L. (2011). Protein-protein binding sites prediction by 3D structural similarities. J. Chem. Inform. Modeling 51, 3287–3294. doi: 10.1021/ci200206n
Guo, F., Li, S. C., Du, P., and Wang, L. (2014). Probabilistic models for capturing more physicochemical properties on protein-protein interface. J. Chem. Inform. Modeling 54, 1798–1809. doi: 10.1021/ci5002372
Guo, F., Li, S. C., Ma, W., and Wang, L. (2013). Detecting protein conformational changes in interactions via scaling known structures. J. Comput. Biol. 20, 765–779. doi: 10.1089/cmb.2013.0069
Hong, Z., Zeng, X., Wei, L., and Liu, X. (2020). Identifying enhancer–promoter interactions with neural network based on pre-trained DNA vectors and attention mechanism. Bioinformatics 36, 1037–1043.
Iliopoulos, A. C., Beis, G., Apostolou, P., and Papasotiriou, I. (2020). Complex networks, gene expression and cancer complexity: a brief review of methodology and applications. Curr. Bioinform. 15, 629–655. doi: 10.2174/1574893614666191017093504
Jiang, Q., Hao, Y., Wang, G., Juan, L., Zhang, T., Teng, M., et al. (2010). Prioritization of disease microRNAs through a human phenome-microRNAome network. BMC Syst. Biol. 4, (Suppl. 1):S2.
Jiang, Q., Jin, S., Jiang, Y., Liao, M., Feng, R., Zhang, L., et al. (2017). Alzheimer’s disease variants with the genome-wide significance are significantly enriched in immune pathways and active in immune cells. Mol. Neurobiol. 54, 594–600. doi: 10.1007/s12035-015-9670-8
Jiang, Q., Wang, G., Jin, S., Li, Y., and Wang, Y. (2013). Predicting human microRNA-disease associations based on support vector machine. Int. J. Data Min. Bioinform. 8, 282–293. doi: 10.1504/ijdmb.2013.056078
Jiang, Q., Wang, J., Wang, Y., Ma, R., Wu, X., and Li, Y. (2014). TF2LncRNA: identifying common transcription factors for a list of lncRNA genes from ChIP-Seq data. Biomed. Res. Int. 2014:317642.
Jin, Q., Meng, Z., Tuan, D. P., Chen, Q., Wei, L., and Su, R. (2019). DUNet: a deformable network for retinal vessel segmentation. Knowl. Based Syst. 178, 149–162. doi: 10.1016/j.knosys.2019.04.025
Jin, S., Zeng, X., Fang, J., Lin, J., Chan, S. Y., and Erzurum, S. C. (2019). applications, a network-based approach to uncover microRNA-mediated disease comorbidities and potential pathobiological implications. NPJ Syst. Biol. Appl. 5, 1–11.
Jin, S., Zeng, X., Xia, F., Huang, W., and Liu, X. (2020). Application of deep learning methods in biological networks. Brief. Bioinform. doi: 10.1093/bib/bbaa043 Online ahead of print.
Jing, F., Zhang, S.-W., and Zhang, S. (2019). Brief survey of biological network alignment and a variant with incorporation of functional annotations. Curr. Bioinform. 14, 4–10. doi: 10.2174/1574893612666171020103747
Konda, A. K., Sabale, P. R., Soren, K. R., Subramaniam, S. P., Singh, P., Rathod, S., et al. (2019). Systems biology approaches reveal a multi-stress responsive WRKY transcription factor and stress associated gene co-expression networks in chickpea. Curr. Bioinform. 14, 591–601. doi: 10.2174/1574893614666190204152500
Kumari, N., and Verma, A. (2020). Analysis of oncogene protein structure using small world network concept. Curr. Bioinform. 15, 732–740. doi: 10.2174/1574893614666191113143840
Li, J., Pu, Y., Tang, J., Zou, Q., and Guo, F. (2020). DeepAVP: a dual-channel deep neural network for identifying variable-length antiviral peptides. IEEE J. Biomed. Health Inform. 24, 3012–3019. doi: 10.1109/jbhi.2020.2977091
Li, Z., Zhang, T., Lei, H., Wei, L., Liu, Y., and Shi, Y. (2020). Research on gastric cancer’s drug-resistant gene regulatory network model. Curr. Bioinform. 15, 225–234. doi: 10.2174/1574893614666190722102557
Lin, H., Liang, Z. Y., Tang, H., and Chen, W. (2019). Identifying Sigma70 promoters with novel pseudo nucleotide composition. IEEE/ACM Trans. Comput. Biol. Bioinform. 16, 1316–1321. doi: 10.1109/tcbb.2017.2666141
Liu, B., Gao, X., and Zhang, H. (2019). BioSeq-Analysis2.0: an updated platform for analyzing DNA, RNA, and protein sequences at sequence level and residue level based on machine learning approaches. Nucleic Acids Res. 47:e127. doi: 10.1093/nar/gkz740
Liu, B., Zhu, Y., and Yan, K. (2020). Fold-LTR-TCP: protein fold recognition based on triadic closure principle. Brief. Bioinform. 21, 2185–2193. doi: 10.1093/bib/bbz139
Liu, G., and Jiang, Q. (2016). Alzheimer’s disease CD33 rs3865444 variant does not contribute to cognitive performance. Proc. Natl. Acad. Sci. U.S.A. 113, E1589–E1590.
Liu, G., Hu, Y., Han, Z., Jin, S., and Jiang, Q. (2019). Genetic variant rs17185536 regulates SIM1 gene expression in human brain hypothalamus. Proc. Natl. Acad. Sci. U.S.A. 116, 3347–3348. doi: 10.1073/pnas.1821550116
Liu, G., Jin, S., Hu, Y., and Jiang, Q. (2018). Disease status affects the association between rs4813620 and the expression of Alzheimer’s disease susceptibility gene TRIB3. Proc. Natl. Acad. Sci. U.S.A. 115, E10519–E10520.
Liu, G., Zhang, F., Jiang, Y., Hu, Y., Gong, Z., Liu, S., et al. (2017). Integrating genome-wide association studies and gene expression data highlights dysregulated multiple sclerosis risk pathways. Mult. Scler. 23, 205–212. doi: 10.1177/1352458516649038
Liu, L., Li, Q. Z., Jin, W., Lv, H., and Lin, H. (2019). Revealing gene function and transcription relationship by reconstructing gene-level chromatin interaction. Comput. Struct. Biotechnol. J. 17, 195–205. doi: 10.1016/j.csbj.2019.01.011
Liu, L., Zhang, L. R., Dao, F. Y., Yang, Y. C., and Lin, H. (2021). A computational framework for identifying the transcription factors involved in enhancer-promoter loop formation. Mol. Ther. Nucleic Acids 23, 347–354. doi: 10.1016/j.omtn.2020.11.011
Liu, M. L., Su, W., Wang, J. S., Yang, Y. H., Yang, H., and Lin, H. (2020). Predicting preference of transcription factors for Methylated DNA using sequence information. Mol. Ther. Nucleic Acids 22, 1043–1050. doi: 10.1016/j.omtn.2020.07.035
Liu, X., Hong, Z., Liu, J., Lin, Y., Alfonso, R.-P., Zou, Q., et al. (2020). Computational methods for identifying the critical nodes in biological networks. Brief. Bioinform. 21, 486–497. doi: 10.1093/bib/bbz011
Lv, H., Dao, F.-Y., Guan, Z.-X., Yang, H., Li, Y.-W., and Lin, H. (2020). Deep-Kcr: accurate detection of lysine crotonylation sites using deep learning method. Brief. Bioinform. bbaa255. doi: 10.1093/bib/bbaa255
Mortezaeefar, M., Fotovat, R., Shekari, F., and Sasani, S. (2019). Comprehensive understanding of the interaction among stress hormones signalling pathways by gene co-expression network. Curr. Bioinform. 14, 602–613. doi: 10.2174/1574893614666190226160742
Shao, J., and Liu, B. (2020). ProtFold-DFG: protein fold recognition by combining Directed Fusion Graph and PageRank algorithm. Brief. Bioinform. doi: 10.1093/bib/bbaa192 Online ahead of print.
Shao, J., Yan, K., and Liu, B. (2020). FoldRec-C2C: protein fold recognition by combining cluster-to-cluster model and protein similarity network. Brief. Bioinform. doi: 10.1093/bib/bbaa144 Online ahead of print.
Sikandar, A., Anwar, W., and Sikandar, M. (2019). Combining sequence entropy and subgraph topology for complex prediction in protein protein interaction (PPI) network. Curr. Bioinform. 14, 516–523. doi: 10.2174/1574893614666190103100026
Song, T., Rodríguez-Patón, A., Zheng, P., and Zeng, X. (2018). Spiking Neural P systems with colored spikes. IEEE Trans. Cogn. Dev. Syst. 10, 1106–1115. doi: 10.1109/tcds.2017.2785332
Srivastava, N., Mishra, B. N., and Srivastava, P. (2019). In-silico identification of drug lead molecule against pesticide exposed-neurodevelopmental disorders through network-based computational model approach. Curr. Bioinform. 14, 460–467. doi: 10.2174/1574893613666181112130346
Su, R., Liu, X., and Wei, L. (2020a). MinE-RFE: determine the optimal subset from RFE by minimizing the subset-accuracy-defined energy. Brief. Bioinform. 21, 687–698. doi: 10.1093/bib/bbz021
Su, R., Liu, X., Wei, L., and Zou, Q. (2019a). Deep-resp-forest: a deep forest model to predict anti-cancer drug response. Methods 166, 91–102. doi: 10.1016/j.ymeth.2019.02.009
Su, R., Liu, X., Xiao, G., and Wei, L. (2020b). Meta-GDBP: a high-level stacked regression model to improve anticancer drug response prediction. Brief. Bioinform. 21, 996–1005. doi: 10.1093/bib/bbz022
Su, R., Wu, H., Xu, B., Liu, X., and Wei, L. (2019b). Developing a multi-dose computational model for drug-induced hepatotoxicity prediction based on toxicogenomics data. IEEE ACM Trans. Comput. Biol. Bioinform. 16, 1231–1239. doi: 10.1109/tcbb.2018.2858756
Tao, Z., Li, Y., Teng, Z., and Zhao, Y. (2020). A method for identifying vesicle transport proteins based on LibSVM and MRMD. Comput. Math. Methods Med. 2020:8926750.
Wang, D., Zhang, Z., Jiang, Y., Mao, Z., Wang, D., Lin, H., et al. (2021). DM3Loc: multi-label mRNA subcellular localization prediction and analysis based on multi-head self-attention mechanism. Nucleic Acids Res. doi: 10.1093/nar/gkab016 Online ahead of print.
Wang, G., Wang, Y., Feng, W., Wang, X., Yang, X. Y., Zhao, Y., Wang, Y., Liu, Y., et al. (2008). Transcription factor and microRNA regulation in androgen-dependent and -independent prostate cancer cells. BMC Genomics 9, (Suppl. 2):S22.
Wang, G., Luo, X., Wang, J., Wan, J., Xia, S., Zhu, H., et al. (2018). MeDReaders: a database for transcription factors that bind to methylated DNA. Nucleic Acids Res. 46, D146–D151.
Wang, G., Wang, Y., Teng, M., Zhang, D., Li, L., and Liu, Y. (2010). Signal transducers and activators of transcription-1 (STAT1) regulates microRNA transcription in interferon gamma-stimulated HeLa cells. PLoS One 5:e11794. doi: 10.1371/journal.pone.0011794
Wang, H., Ding, Y., Tang, J., and Guo, F. (2020). Identification of membrane protein types via multivariate information fusion with Hilbert-Schmidt Independence criterion. Neurocomputing 383, 257–269. doi: 10.1016/j.neucom.2019.11.103
Wang, H., Tang, J., Ding, Y., and Guo, F. (2021). Exploring associations of non-coding RNAs in human diseases via three-matrix factorization with hypergraph-regular terms on center kernel alignment. Brief. Bioinform. doi: 10.1093/bib/bbaa409 Online ahead of print.
Wang, L., Xuan, Z., Zhou, S., Kuang, L., and Pei, T. (2019). A Novel Model for predicting LncRNA-disease associations based on the LncRNA-MiRNA-disease interactive network. Curr. Bioinform. 14, 269–278. doi: 10.2174/1574893613666180703105258
Wei, L., Ding, Y., Su, R., Tang, J., and Zou, Q. (2018a). Prediction of human protein subcellular localization using deep learning. J. Parallel Distrib. Comput. 117, 212–217.
Wei, L., Liao, M., Gao, Y., Ji, R., He, Z., and Zou, Q. (2014). Improved and promising identification of human micrornas by incorporating a high-quality negative set. IEEE ACM Trans. Comput. Biol. Bioinform. 1, 192–201. doi: 10.1109/tcbb.2013.146
Wei, L., Tang, J., and Zou, Q. (2017a). Local-DPP: an improved DNA-binding protein prediction method by exploring local evolutionary information. Inform. Sci. 384, 135–144. doi: 10.1016/j.ins.2016.06.026
Wei, L., Wan, S., Guo, J., and Wong, K. K. L. (2017b). A novel hierarchical selective ensemble classifier with bioinformatics application. Artif. Intell. Med. 83, 82–90. doi: 10.1016/j.artmed.2017.02.005
Wei, L., Xing, P., Shi, G., Ji, Z., and Zou, Q. (2019). Fast prediction of protein methylation sites using a sequence-based feature selection technique. IEEE ACM Trans Comput. Biol. Bioinform. 16, 1264–1273. doi: 10.1109/tcbb.2017.2670558
Wei, L., Xing, P., Zeng, J., Chen, J., Su, R., and Guo, F. (2017c). Improved prediction of protein-protein interactions using novel negative samples, features, and an ensemble classifier. Artif. Intell. Med. 83, 67–74. doi: 10.1016/j.artmed.2017.03.001
Wei, L., Zhou, C., Chen, H., Song, J., and Su, R. (2018b). ACPred-FL: a sequence-based predictor using effective feature representation to improve the prediction of anti-cancer peptides. Bioinformatics 34, 4007–4016.
Yang, Q., Wu, J., Zhao, J., Xu, T., Han, P., and Song, X. (2020). The expression profiles of lncRNAs and their regulatory network during Smek1/2 knockout mouse neural stem cells differentiation. Curr. Bioinform. 15, 77–88. doi: 10.2174/1574893614666190308160507
Zeng, X. X., Liu, L., Lu, L. Y., and Zou, Q. (2018). Prediction of potential disease-associated microRNAs using structural perturbation method. Bioinformatics 34, 2425–2432. doi: 10.1093/bioinformatics/bty112
Zeng, X., Song, X., Ma, T., Pan, X., Zhou, Y., and Hou, Y. (2020). Repurpose open data to discover therapeutics for COVID-19 using deep learning. J. Proteome Res. 19, 4624–4636. doi: 10.1021/acs.jproteome.0c00316
Zeng, X., Zhang, X., and Zou, Q. (2016). Integrative approaches for predicting microRNA function and prioritizing disease-related microRNA using biological interaction networks. Brief. Bioinform. 17, 193–203. doi: 10.1093/bib/bbv033
Zhai, Y., Chen, Y., Teng, Z., and Zhao, Y. (2020). Identifying antioxidant proteins by using amino acid composition and protein-protein interactions. Front. Cell Dev. Biol. 8:591487.
Zhang, D., Chen, H.-D., Zulfiqar, H., Yuan, S.-S., Huang, Q.-L., Zhang, Z.-Y., et al. (2021). iBLP: an XGBoost-based predictor for identifying bioluminescent proteins. Comput. Math. Methods Med. 2021:6664362.
Zhang, J., Feng, J., and Wu, F.-X. (2020). Finding community of brain networks based on neighbor index and DPSO with dynamic crossover. Curr. Bioinform. 15, 287–299. doi: 10.2174/1574893614666191017100657
Zhang, W., Li, W., Zhang, J., and Wang, N. (2019). Data integration of hybrid microarray and single cell expression data to enhance gene network inference. Curr. Bioinform. 14, 255–268. doi: 10.2174/1574893614666190104142228
Zhang, Z. M., Tan, J. X., Wang, F., Dao, F. Y., Zhang, Z. Y., and Lin, H. (2020). Early diagnosis of hepatocellular carcinoma using machine learning method. Front. Bioeng. Biotechnol. 8:254.
Zhang, Z. Y., Yang, Y. H., Ding, H., Wang, D., Chen, W., and Lin, H. (2021). Design powerful predictor for mRNA subcellular location prediction in Homo sapiens. Brief. Bioinform. 22, 526–535. doi: 10.1093/bib/bbz177
Zhao, X., Chen, L., Guo, Z.-H., and Liu, T. (2019). Predicting drug side effects with compact integration of heterogeneous networks. Curr. Bioinform. 14, 709–720. doi: 10.2174/1574893614666190220114644
Zhao, Y., Wang, F., Chen, S., Wan, J., and Wang, G. (2017). Methods of MicroRNA promoter prediction and transcription factor mediated regulatory network. Biomed Res. Int. 2017:7049406.
Zhu, L., Su, F., Xu, Y., and Zou, Q. (2018). Network-based method for mining novel HPV infection related genes using random walk with restart algorithm. BBA Mol. Basis Dis. 1864, 2376–2383. doi: 10.1016/j.bbadis.2017.11.021
Zou, Q., Li, J., Song, L., Zeng, X., and Wang, G. (2016). Similarity computation strategies in the microRNA-disease network: a survey. Brief. Funct. Genomics 15, 55–64.
Keywords: multilayer networks, bioinformatics, brain network structure, biological systems, chronological networks
Citation: Lv Y, Huang S, Zhang T and Gao B (2021) Application of Multilayer Network Models in Bioinformatics. Front. Genet. 12:664860. doi: 10.3389/fgene.2021.664860
Received: 06 February 2021; Accepted: 26 February 2021;
Published: 31 March 2021.
Edited by:
Chunhou Zheng, Anhui University, ChinaReviewed by:
Jin-Xing Liu, Qufu Normal University, ChinaCopyright © 2021 Lv, Huang, Zhang and Gao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tianjiao Zhang, enRqLmhpdEBnbWFpbC5jb20=; Bo Gao, MTY3ODcyOTU4OEBxcS5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.