 Soniza Vieira Alves-Leon1,2*†
Soniza Vieira Alves-Leon1,2*† Cristina dos Santos Ferreira3†
Cristina dos Santos Ferreira3† Alice Laschuk Herlinger4†
Alice Laschuk Herlinger4† Fabricia Lima Fontes-Dantas1†
Fabricia Lima Fontes-Dantas1† Fernanda Cristina Rueda-Lopes5
Fernanda Cristina Rueda-Lopes5 Ronaldo da Silva Francisco Jr.3
Ronaldo da Silva Francisco Jr.3 João Paulo da Costa Gonçalves1,2
João Paulo da Costa Gonçalves1,2 Amanda Dutra de Araújo1,2
Amanda Dutra de Araújo1,2 Cláudia Cecília da Silva Rêgo1,2
Cláudia Cecília da Silva Rêgo1,2 Luiza Mendonça Higa4
Luiza Mendonça Higa4 Alexandra Lehmkuhl Gerber3
Alexandra Lehmkuhl Gerber3 Ana Paula de Campos Guimarães3
Ana Paula de Campos Guimarães3 Mariane Talon de Menezes4
Mariane Talon de Menezes4 Marcelo Calado de Paula Tôrres4Richard Araújo Maia4
Marcelo Calado de Paula Tôrres4Richard Araújo Maia4 Bruno Miceli Gonzalez Nogueira1Laise Carolina França1Marcos Martins da Silva6Christian Naurath7
Bruno Miceli Gonzalez Nogueira1Laise Carolina França1Marcos Martins da Silva6Christian Naurath7 Aline Saraiva da Silva Correia7Claudia Cristina Ferreira Vasconcelos8
Aline Saraiva da Silva Correia7Claudia Cristina Ferreira Vasconcelos8 Amilcar Tanuri4
Amilcar Tanuri4 Orlando Costa Ferreira Jr.4
Orlando Costa Ferreira Jr.4 Cynthia Chester Cardoso4
Cynthia Chester Cardoso4 Renato Santana Aguiar9
Renato Santana Aguiar9 Ana Tereza Ribeiro de Vasconcelos3*
Ana Tereza Ribeiro de Vasconcelos3*- 1Translational Neuroscience Laboratory, Rio de Janeiro State Federal University, Rio de Janeiro, Brazil
- 2Department of Neurology/Reference and Research Center for Multiple Sclerosis and Other Central Nervous System Idiopathic Demyelinating Inflammatory Diseases, Clementino Fraga Filho University Hospital, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil
- 3Bioinformatics Laboratory, National Laboratory for Scientific Computing, Petrópolis, Brazil
- 4Molecular Virology Laboratory, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil
- 5Department of Radiology, Antônio Pedro University Hospital, Fluminense Federal University, Niterói, Brazil
- 6Department of Clinical Medicine, Clementino Fraga Filho University Hospital, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil
- 7Federal Hospital Cardoso Fontes, Ministry of Health, Rio de Janeiro, Brazil
- 8Department of Neurology, University Hospital Gafrée e Guinle, Rio de Janeiro State Federal University, Rio de Janeiro, Brazil
- 9Department of Genetics, Ecology and Evolution, Institute of Biological Sciences, Federal University of Minas Gerais, Belo Horizonte, Brazil
Chikungunya virus (CHIKV) is a re-emergent arbovirus that causes a disease characterized primarily by fever, rash and severe persistent polyarthralgia, although <1% of cases develop severe neurological manifestations such as inflammatory demyelinating diseases (IDD) of the central nervous system (CNS) like acute disseminated encephalomyelitis (ADEM) and extensive transverse myelitis. Genetic factors associated with host response and disease severity are still poorly understood. In this study, we performed whole-exome sequencing (WES) to identify HLA alleles, genes and cellular pathways associated with CNS IDD clinical phenotype outcomes following CHIKV infection. The cohort includes 345 patients of which 160 were confirmed for CHIKV. Six cases presented neurological manifestation mimetizing CNS IDD. WES data analysis was performed for 12 patients, including the CNS IDD cases and 6 CHIKV patients without any neurological manifestation. We identified 29 candidate genes harboring rare, pathogenic, or probably pathogenic variants in all exomes analyzed. HLA alleles were also determined and patients who developed CNS IDD shared a common signature with diseases such as Multiple sclerosis (MS) and Neuromyelitis Optica Spectrum Disorders (NMOSD). When these genes were included in Gene Ontology analyses, pathways associated with CNS IDD syndromes were retrieved, suggesting that CHIKV-induced CNS outcomesmay share a genetic background with other neurological disorders. To our knowledge, this study was the first genome-wide investigation of genetic risk factors for CNS phenotypes in CHIKV infection. Our data suggest that HLA-DRB1 alleles associated with demyelinating diseases may also confer risk of CNS IDD outcomes in patients with CHIKV infection.
Introduction
Chikungunya fever is typically described as a mild disease in which clinical symptoms are mainly related to articular pain and general malaise (Burt et al., 2017). Chikungunya virus (CHIKV) is an arthropod-borne virus from the genus Alphavirus, family Togaviridae, transmitted by the bite of Aedes mosquitoes and endemic in tropical countries (Pialoux et al., 2007). CHIKV infection is considered endemic in the Southeast Region of Brazil, with 1,159.4 cases/100,000 inhabitants having been registered in 2019 (BRASIL, 2020). However, involvement of central and peripheral nervous systems has been reported in 0.3 to 1% (Cerny et al., 2017; Anand et al., 2019), which are considered atypical manifestations of CHIKV.
The central nervous system (CNS) manifestations during CHIKV infections seem not to be different from other viruses like the Zika virus (Alves-Leon et al., 2019). White and gray matter are frequently affected in CNS cases and clinical findings sometimes mimic inflammatory demyelinating diseases (IDD) of the brain and spinal cord, mainly with clinical phenotypes of acute disseminated encephalomyelitis (ADEM), multiple sclerosis (MS), longitudinal extensive myelitis, multifocal myelitis, optic neuritis, and neuromyelitis optic spectrum disorders (NMOSD) (Mehta et al., 2018).
The immune process linked to CNS demyelination is triggered at least in part by post-infectious inflammation. The exact mechanism that leads to the development of these neurological manifestations in a small number of individuals with CHIKV infection is still unknown. In this context, ADEM and Myelitis may show similarities at clinical onset, such as disturbance of immune tolerance to myelin proteins and association with genetic variations mainly in HLA alleles, with presentation of MS, NMOSD and other non-MS autoimmune diseases of the CNS (Alves-Leon et al., 2009; Mowry and Waubant, 2010; Tobin et al., 2014; Lin et al., 2016; Höftberger and Lassmann, 2017).
Several viral and bacterial pathogens and different vaccines have been associated with ADEM and Myelitis (Menge et al., 2005; Yeshokumar and Waubant, 2017). Nevertheless, the discussion on whether CNS CHIKV manifestations are due to direct viral action or an autoimmune response triggered by the infection in a genetically susceptibility individual remains unclear. Variants in TLR7 (rs179010) have already been associated with susceptibility to infection, while TLR3 (rs6552950) was associated with disease severity and specific neutralizing antibody response (Her et al., 2015; Dutta and Tripathi, 2017). Other host genetic variants in CD20913 and HLA alleles were associated with disease susceptibility or different clinical outcomes in CHIKV infection (Chaaithanya et al., 2016). Recently, patients with post-infectious neurologic syndromes (PINS) presenting mainly myelitis (frequency of 44%) and encephalomyelitis (frequency of 44%) were genotyped using the reference panel of the 1000 Genomes Project, but no risk alleles for MS were found (Martinelli-Boneschi et al., 2020).
The neurological manifestations of CHIKV infection make the differential diagnosis of immune-mediated PINS that affect the brain and spinal cord a challenge for neurologists. Since such outcomes are rare, it is not surprising that little is known about their genetic background. In regions such as Rio de Janeiro State, where CHIKV is highly incident, it is not unlikely that some CNS IDDs otherwise considered idiopathic, may, in fact, result from a previous or current CHIKV infection. Therefore, in order to better characterize the CHIKV-related CNS IDD phenotype, we herein provide a detailed clinical and radiological description of 6 patients presenting ADEM or Myelitis following CHIKV infection. Moreover, we performed whole exome sequencing (WES) analyses to investigate the genetic background and biological pathways associated with neurologic phenotypes of CHIKV infection.
Materials and Methods
Study Design and Patients
This is a prospective study, which included a convenience sampling of patients from an ongoing active cohort study in different hospitals in Rio de Janeiro and patients referred by Laboratório Central Noel Nutels (LACEN) in Rio de Janeiro, between July 2016 and June 2019. During this period, the health department from the Rio de Janeiro municipality registered 28,674 Dengue, 3,897 Zika and 51,626 Chikungunya cases with an increasing number of Chikungunya infections overcoming other arboviruses during the last 2 years 2018–2019. The cohort included 345 patients with 160 confirmed positive CHIKV infections investigated by direct detection of virus (RT-qPCR) or antibody detection (ELISA and PRNT evaluation). All participants were evaluated by a team of neurologists. When any neurological manifestation was detected, blood or cerebrospinal fluid (CSF) and imaging tests were requested to perform the differential diagnosis. Patient history, complete physical and neurological examination was performed, including details of serum and CSF testing, panel for autoimmune and other infectious diseases. Neuroimaging, nerve conduction studies, and electromyography were also used for classification of CNS or peripheral nervous system outcomes. After reviewing each case, patients who met the diagnostic criteria by use of standardized case definitions were classified as having CNS IDD phenotype. The other patients were classified without neurological manifestations or with other neurological diseases. Briefly, the patients diagnosed as ADEM fulfill diagnostic criteria (Sejvar et al., 2007; Krupp et al., 2013) of this type of IDD due the presentation of acute encephalopathy manifestations characterized by headache, epileptic seizures and mental confusion associated with brain and/or spinal cord white and gray matter lesions, some of them tumefactive. The diagnosis of Myelitis was based on development of sensory, motor, or autonomic dysfunction attributable to the spinal cord in conjunction with evidence of inflammation and exclusion of extra-axial compressive cause by neuroimaging in accordance with Transverse Myelitis Consortium Working Group (Transverse Myelitis Consortium Working Group, 2002). From our cohort the 6 patients (0.37%) who presented CNS IDD phenotype manifestations following CHIKV infections were selected for exome analysis. Six patients meeting the inclusion criteria who had no IDD were selected for exome analysis, as a baseline for comparisons. Individuals selected for this control group were paired by age and sex with the cases group. The study was approved by the local Ethics Committee (protocol number: 69411317.6.0000.5258) and a written informed consent was signed by all participants by the time of inclusion in the study. The demographic and systemic characteristics of CHIKV infection in these patients are described in Table 1. The CNS IDD manifestations included three cases of ADEM and three cases of Myelitis.
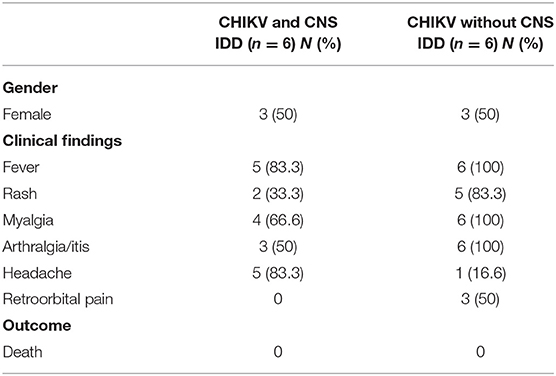
Table 1. Frequency of clinical, neurological manifestations, and in-hospital outcome.
Neuroimaging Diagnosis
Magnetic resonance imaging (MRI) was performed using 1.5 or 3.0 Tesla scanners, depending on the clinical availability. The protocols included brain, cervical and spinal cord MRI according to clinical symptoms. Brain MRI included 3-dimension fluid-attenuated inversion-recovery sequences (3D FLAIR), 3D T1 weighted-images (WI), T2-WI, diffusion WI (DWI), short-tau inversion recovery (STIR) for optic nerve analysis. Spinal cord MRI included sagittal STIR, T2-WI, and T1-WI and axial T2-WI. Brain and spinal cord fat suppressed (FS) T1-WI post-gadolinium scans were also obtained, on axial and sagittal planes.
Chikungunya Diagnosis
Blood and/or urine samples were collected at patient admission. Cerebrospinal fluid was also collected from patients with CNS IDD and stored at −80°C until further analysis. Biological samples were blindly tested for Zika (ZIKV), CHIKV, and Dengue (DENV) virus. Samples selected for exome analysis had their molecular and serological diagnosis confirmed as described below.
Virus RNA was obtained from serum, urine or CSF samples using the QIAmp MinElute Virus Spin Kit (Qiagen), according to manufacturer's instructions. RNA was eluted in 50 and 5 μL were used for virus genome detection using the Go-Taq Probe 1-step RT-qPCR System (Promega) and previously designed primers targeting CHIKV, DENV, and ZIKV (Lanciotti et al., 2007, 2008; Menting et al., 2011). Antibodies against CHIKV infection (IgG and IgM) were investigated by MAC-ELISA (Euroimmun) using serum samples, according to manufacturer's instructions. Plaque-reduction neutralization tests (PRNT) were also performed to detect neutralizing antibodies against CHIKV and to exclude cross-reaction with other arboviruses circulating in the same region. Briefly, plasma samples were incubated at 58°C for 30 min to inactivate CHIKV and limit complement effect. Serial dilutions (from 1:5 to 1:2,560) of heat-inactivated plasma were incubated with an equal volume containing 100 plaque forming units (PFU) of CHIKV isolate BHI3745/H804709 (Accession Number KP164570.1) for 1 h at 37° C. The virus-sera mixture was inoculated into confluent monolayers of VERO cells. After 1 h, inoculum was removed and semisolid medium (1.2% carboxymethylcellulose in alpha-MEM supplemented with 1% fetal bovine serum) was added. Cells were further incubated for 30 h and then fixed with 4% formaldehyde solution. Cells were stained with crystal violet dye solution. PRNT end-point titers are expressed as the reciprocal of the last serum dilution showing reduction in plaque counts ≥90%. PRNT90 titer ≥20 was considered positive for the presence of neutralizing antibodies against CHIKV.
DNA Extraction and Whole Exome Sequencing
Genomic DNA (gDNA) was obtained from blood cells using the MasterPure Complete DNA and RNA Purification Kit (Lucigen). Integrity of gDNA was accessed by agarose gel electrophoresis and quantification was performed using the Qubit dsDNA HS Assay (Thermo Fisher Scientific). Libraries were prepared using Illumina TruSeq Exome Library Prep kit according to the manufacturer's protocol. Sequencing was performed with the Illumina NextSeq® 500/550 High Output Kit v2 (150 cycles), generating 2 × 75 bp paired-end reads. Raw sequencing data files in FASTQ format generated in the Illumina platform were preprocessed in fastqc and trimmomatic for quality assessment and trimming analysis, respectively. Sequenced reads were aligned to the GRCh38 human reference genome using Burrows-Wheeler Aligner (BWA) (Li and Durbin, 2009). Additional BAM file manipulations were performed with Samtools (Li et al., 2009) for sorting and mapping quality filtration (Q30). Duplicate reads were marked using Picard MarkDuplicates tool (http://broadinstitute.github.io/picard). Base quality recalibration and variant calling were performed using BQSR and HaplotypeCaller from Genome Analysis Toolkit (GATK) (McKenna et al., 2010).
Detection of Pathogenic and Potentially Pathogenic Variants
In order to select potential pathogenic variants, we establish two sets of genes. First, a panel of genes previously related to CHIKV infection and neurological impairment was established based on a literature review. We next extended the gene panel by retrieving genes related to enriched Gene Ontology (GO) terms such as pathogen response, immune response, metabolic regulation and expression regulation. To investigate the relationship between the two sets of genes, a connectome analysis was performed (Itan et al., 2013) by estimating the pairwise protein-protein interaction between genes. A p ≤ 0.05 was applied to filter interactions. Next, the Mendelian inheritance model was retrieved for each gene by querying the OMIM database. Finally, we prioritize possibly damaging variants mapped in the final gene list for each patient (Supplementary Figure 1).
Gene-based variant findings were further filtered following functional annotation. Only protein-altering variants were selected, including truncating variants (stop gain/loss, start loss, or frameshift), missense variants, canonical splice-site variants, and inframe indels affecting protein-coding regions. Overall, these functional annotations correspond to high and moderate impact annotation. Variant pathogenicity was predicted using computational tools such as SIFT (Sorting Intolerant from Tolerant) (Ng and Henikoff, 2003), PolyPhen (Polymorphism Phenotyping) (Adzhubei et al., 2013), CADD (Combined Annotation-Dependent Depletion) (Kircher et al., 2014), and LoFtool (Fadista et al., 2017) according to the Ensembl Variant Effect Predictor database (VEP) (McLaren et al., 2016). In addition, American College of Medical Genetics and Genomics (ACMG) (Richards et al., 2015) and ClinVar (Landrum et al., 2018) recommendation guidelines were also assigned. After these filtration steps, we researched the population frequency of the variants in 1000 Genomes Project, Exome Aggregation Consortium (ExAC) and Genome Aggregation Database (gnomAD). The analysis pipeline is shown in Supplementary Figure 2.
HLA Typing From Exome Data
The HLA alleles were predicted from WES reads with the HLAminer tool (Warren et al., 2012). Sequences in FASTQ format were mapped against chromosome 6 (GRCh38) using bowtie2 (Langmead and Salzberg, 2012). To predict HLA alleles, genomic sequences of classes I and II HLA were retrieved from the IMGT/HLA database in FASTA format. WES sequences were assembled into 200 bp contigs using the TASR tool (Warren and Holt, 2011). Contigs were aligned to HLA reference sequences using BLAST (Basic Local Alignment Tool). Aligned sequences were then used to predict HLA alleles.
Pathway Enrichment Analysis
The biological significance of the filtered gene set was explored by enrichment analysis of the GO term for the biological process. Enrichment analysis was conducted using the ClusterProfiler R package v.3.18.0 (Yu et al., 2012) to produce gene clusters (≥2 genes/cluster). We filtered GO terms with a p-value and q-value (Benjamini & Hochberg (BH) method) smaller than 0.05.
Results
This is a series case report describing clinical and neurological outcomes and exome analysis associated with CHIKV infection. In the following sections we describe the neuroimage findings in the CHIKV-confirmed patients followed by the genetic variants and cellular pathways in patients harboring CNS outcomes.
Central Nervous System IDD Phenotypes in Patients With CHIKV Infection
During the period of this study, our multidisciplinary team assisted 345 patients, 160 of which had CHIKV infection, and the remaining were diagnosed with either Dengue or Zika infection. Within the CHIKV positive patients, sixteen presented involvement of the nervous system, and six were classified as having neurological manifestations mimetizing CNS IDD. For all patients that we perform the exome (with and without IDD), the most frequent systemic arbovirus symptoms overall were fever [11 (92%)], myalgia [10 (83%)], and arthralgia [9 (75%)]. Interestingly only one patient with IDD phenotype following CHIKV infection was completely asymptomatic for the classic signs and symptoms of the infection. These patients never had any previous demyelinating manifestations. All patients with CNS IDD phenotype underwent imaging which showed the extent of the inflammatory lesion (Figure 1, Supplementary Figures 3–8). Clinical and radiological characteristics are summarized in Table 2 and detailed in Supplementary Table 1, which shows all clinical, infectious and neurological features of each patient, as well as the exams at the time of the neurological condition.
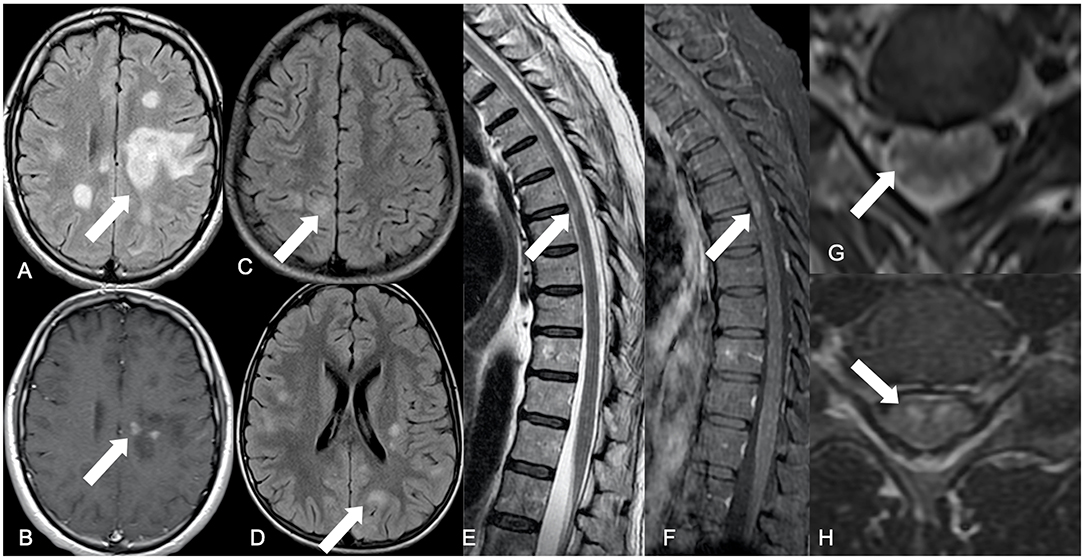
Figure 1. Summary of main neuroimaging findings. Brain axial FLAIR showing a tumefactive hyperintense lesion located in the frontal-parietal transition of the left hemisphere (white arrow) (A), with punctate discontinued contrast enhancement on the corresponding axial post-contrast T1-WI (white arrow) (B), representing an ADEM pattern. An encephalitis pattern can be seen on brain FLAIR axial images (white arrows) with cortical-subcortical hyperintense diffuse lesions (C,D). Dorsal spinal cord sagittal T2-WI with elongated areas of peripherally located hypersignal intensity among the white matter tracts (white arrow) (E), associated with contrast enhancement on sagittal FS T1-WI (white arrow) (F). Axial T2-WI have elongated hypersignal images in a radial distribution, from the central spinal cord gray matter to the peripheral white matter (white arrow) (G). Axial T2-WI with hypersignal affecting the gray matter anterior horn, in a polio-like pattern (white arrow) (H).

Table 2. Neurological manifestations associated with CNS IDD phenotype diagnosis.
Identification of Genetic Variants, Functional Annotation of Genes in Exomes Associated With CNS IDD
On average, 98% of the reads from the WES mapped to the reference genome. Targeted exonic regions achieved 121-fold mean sequence coverage with 91% of bases covered at >20-fold. Genetic variants were filtered based on the impact in protein changes, computational predictions of pathogenicity and population frequency. Overall, we interrogated variants with deleterious profiles according to different algorithms (SIFT, PolyPhen, CADD, and LoFtool), low frequency variants in databases from the 1,000 Genomes Project, ExAC, or gnomAD and variants with pathogenic profiles according to ACMG and ClinVar recommendations. In total, a set of 30 promising variants was defined (Figure 2, Supplementary Table 2). Specifically, in the 6 neurologically affected patients, we found 24 variants in 23 genes, 20 were found exclusively in these patients, while four variants were also found among controls. Most of the identified variants have minor allele frequencies lower than 0.01, and we were unable to identify any variant that explains the CNS IDD manifestations when compared with the unaffected group or a cohort of 609 aged Brazilian individuals (Naslavsky et al., 2017).
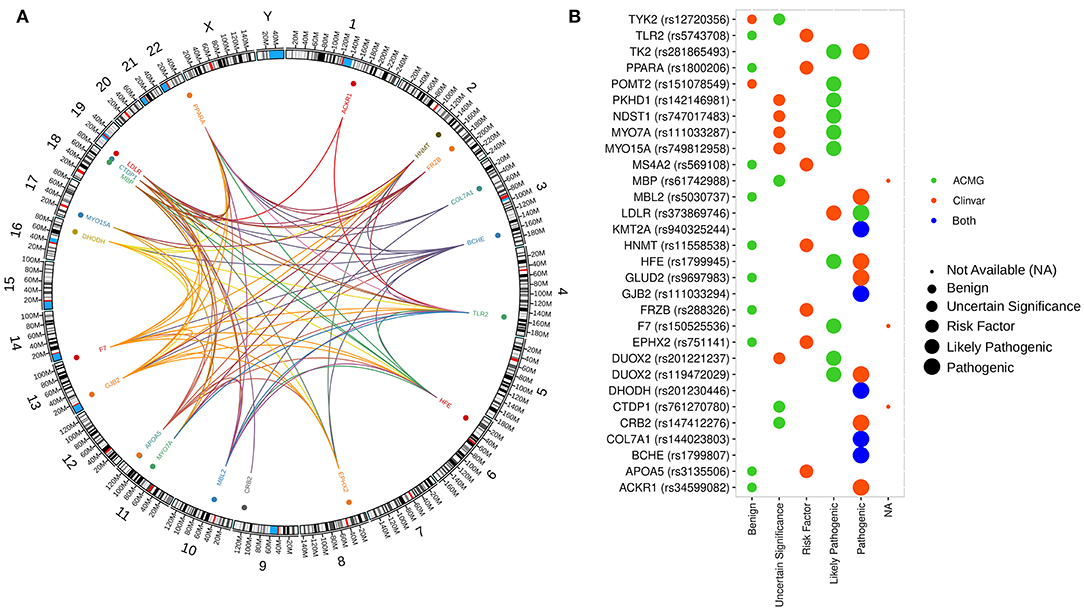
Figure 2. Distribution and interaction between genetic variants and gene ontology across the human genome. (A) Genomic map showing the distribution of genes with predicted pathogenic variants selected by our analysis. Colored dots represented the localization of genes with at least one variant in all patients. The innermost layer represents the connections between the functional annotation of the genes. (B) Pathogenicity classification of genetic variants from our study according to ClinVar and ACMG guideline recommendations.
A phenotype-based analysis allowed the identification of 4 genes (TLR2, MBP, TYK2, and CTDP1) related to CHIKV infection with neurological involvement that harbor variants with pathogenicity prediction. TLR2 is a pattern-recognition receptor associated with cell-mediated immunity, which has been associated mainly with recurrent mycobacterial infection. A role in neurogenesis has also been described (OMIM #607948). The MBP gene encodes a transmembrane IgE receptor associated with susceptibility to multiple sclerosis (OMIM#126200) and the TYK2 gene encodes a tyrosine kinase associated to type I and III interferon signaling, playing a role in antiviral immunity. Deficiency in TYK2 is associated with Immunodeficiency 35 phenotype and increasing susceptibility to viral infection (OMIM#611521). CTDP1 has been associated with congenital cataract, facial dysmorphism, and neuropathy (OMIM#604168) phenotypes. Notably, variant rs5030737 [c.154C>T, p.Arg52Cys] in MBL2, previously associated with mannose-binding lectin deficiency and chronic infections (OMIM#614372), was also detected in a CNS IDD case.
Afterwards, classic HLA alleles [HLA-A, HLA-B, HLA-C, HLA-DP (DPA1, DPB1 and DPB2), HLA-DQ (DQA1, DQA2 and DQB1) and HLA-DR (DRA, DRB1, 3, 4, and 5)] were predicted from WES data for each individual, along with non-classic HLA-G, HLA-E, and HLA-F alleles (Supplementary Table 3, Supplementary Figure 2).
To briefly describe the main variations found in each patient with CNS IDD phenotypes, we highlight that case 1 (18ZC026) harbored four pathogenic, missense variants at PKHD1, TYK2, FRZB, and HNMT genes, in addition to the haplotype DRB3*02:02-DQA1*05:01, associated with susceptibility to Graves' disease (Chen et al., 2000), and DQA1*05:01-DQB1*02:01-DRB1*03:01 haplotype, associated with risk of NMOSD (Zéphir et al., 2009; Brum et al., 2010; Deschamps et al., 2011) (Table 3, Supplementary Table 2). Case 2 (19ZC131) has eight missense variants within several genes, including Gly407Ser at LDLR, associated with familial hypercholesterolemia 1 phenotype (OMIM#143890) and also the rs5030737 polymorphism in the MBL2 gene, previously associated with mannose-binding lectin deficiency and chronic infections (OMIM#614372). This patient also carries DRB1*15 and DPA1*03:01 HLA alleles, which were significantly associated with ADEM (Alves-Leon et al., 2009), and haplotype DRB3*02:02-DQA1*05:01, associated with Graves' disease (Chen et al., 2000; Table 3, Supplementary Table 2).
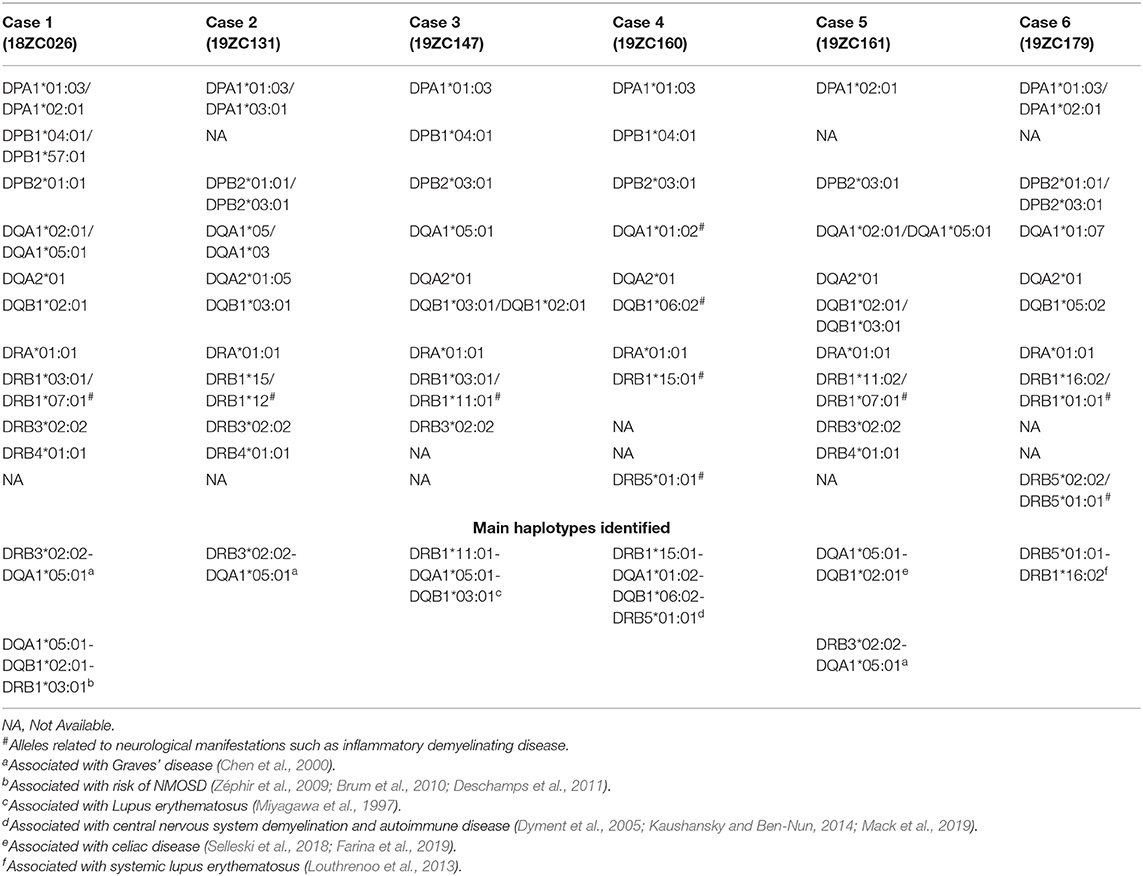
Table 3. Distribution of Class II HLA alleles by CHIKV-related CNS IDD cases.
The missense variant (Arg376Trp), located in the DUOX2 gene, was found in cases 2 and 3 (19ZC147). The DUOX2 gene is associated with thyroid dyshormonogenesis 6 phenotype (OMIM#607200). Case 3 also carries a variant in the CRB2 gene, the HLA-DRB1*03 allele related to NMOSD (Zéphir et al., 2009; Brum et al., 2010; Deschamps et al., 2011), and haplotype DRB1*11:01-DQA1*05:01-DQB1*03:01, associated with lupus erythematosus (Miyagawa et al., 1997; Table 3, Supplementary Table 2). Case 4 (19ZC160) has five missense variants, including His63Asp, within the HFE gene, involved in iron absorption and associated with hemochromatosis (OMIM#235200) and microvascular complications of diabetes 7 (OMIM#612635). Variants in the TK2 gene, associated with mitochondrial DNA depletion syndrome 2 (OMIM#609560), and in the MBP gene, associated with susceptibility to multiple sclerosis (OMIM#126200), were also found. Case 4 was also homozygous for variant rs9697983 [c.1492T>G, p.Ser498Ala] in the GLUD2 gene, associated with Parkinson's disease (OMIM#168600). Patient 19ZC160 also carries the HLA haplotype DRB1*15:01-DQA1*01:02-DQB1*06:02-DRB5*01:01, associated with CNS demyelination and autoimmune disease (Dyment et al., 2005; Kaushansky and Ben-Nun, 2014; Mack et al., 2019), and the DPA1*01:03 and DPB1*04:01 alleles, associated with narcolepsy (Cortes et al., 2004; Louthrenoo et al., 2013; Table 3, Supplementary Table 2).
Case 5 (19ZC161) has four missense variants, including rs751141, in the hydrolase EPHX2 gene, associated with hypercholesterolemia (OMIM#143890), and rs761270780, in the CTDP1 gene. Moreover, HLA haplotype DQA1*05:01-DQB1*02:01 and DRB3*02:02-DQA1*05:01, associated with celiac disease (Selleski et al., 2018; Farina et al., 2019) and Graves' disease (Chen et al., 2000), were also found (Table 3, Supplementary Table 2). Case 6 (19ZC179) had two variants, including rs5743708, in TLR2. This patient also carries the HLA-A*03 allele, associated to MS (Naito et al., 1972), B*51:01/B*27:05 alleles, associated with inflammatory disease (Mizuki et al., 1997), DQB1*05:02, associated with Myasthenia gravis (Testi et al., 2012), and the DRB5*01:01-DRB1*16:02, associated with susceptibility to systemic lupus erythematosus (Louthrenoo et al., 2013; Table 3, Supplementary Table 2).
Despite the lack of significantly overrepresented pathways in patients with CHIKV-associated neurologic outcomes, we found that the associated set contains 29 genes annotated by at least one of the 24 GO terms enriched at q < 0.05 (Figure 2, Supplementary Table 4). The enriched GO includes pathways already related with IDD syndrome, such as leukocyte activation involved in the inflammatory response (GO:0002269; p = 2.51E-3) and microglial cell activation (GO:0001774; p = 2.51E-3).
Discussion
In the last decades, the idea of a virus playing a role in the development of IDD has been reinforced in several studies (Lucas et al., 2011). It is believed that infections can trigger such demyelinating lesions by the activation of autoreactive T cells, with primary events taking place outside the CNS further initiating pathogenic T cell reactivation within the CNS, or by molecular mimicry and developing an autoantibody response (Salvetti et al., 2009; Delogu et al., 2011; Anaya et al., 2016; Burnard et al., 2017). Here, we describe ourselves for the first time that CHIKV patients who developed CNS IDD phenotype shared a common genetic signature with diseases such as MS and NMOSD.
During CHIKV infection progression viremia is observed up to the 5–7 days post-illness onset (pio), whereas IgM can be detected from 2 days pio up to 4 months, and IgG from 4 days pio to over one year (Tanabe et al., 2018). By the time of neurological evaluation, all patients described herein were both CHIKV IgM and IgG reactive, whereas presenting a prodrome period varying from 7 to 30 days (from CHIKV-like symptoms to neurological ones), suggesting they were either at the acute phase of the infection, or at the early-chronic phase of the infection. Two important characteristics of nervous system involvement in CHIKV infection are the fact that it is rare and that it can mimic autoimmune inflammatory diseases in adults. Among CHIKV positive patients, sixteen presented nervous system involvement and six were classified as having neurological manifestations mimetizing CNS IDD as ADEM and extensive transverse myelitis. Despite the relatively small sample size, literature shows that 0.3–1% of cases will develop severe neurological manifestations, corroborating our findings (Cerny et al., 2017).
The genome-wide search for genetic variants associated to diseases such as MS, NMOSD, ADEM, and myelitis has suggested that the sharing of molecular pathways between these immune-mediated diseases may be greater than we imagined (Oldstone, 1998; Kakalacheva et al., 2011; Kattimani and Veerappa, 2018; Zamvil et al., 2018). This justifies similar heterogeneous clinical phenotypes associated with various environmental factors, such as the seasonal arbovirus infections that Brazil and other countries have been exposed to. In this context, the viral trigger is an opportunity to investigate the encounter between individual genetic variants that may be associated with the immune response and CNS IDD. Possible explanations to these questions include the phenomenon of molecular mimicry and shared polymorphisms between these immune mediated diseases, both hypotheses to be further explored.
Although we have not identified any variant that could explain susceptibility to IDD syndromes following CHIKV infection in our cohort, these results do not exclude a genetic component for this neurological manifestation. Indeed, the lack of a clear genetic cause reinforces the hypothesis of a multifactorial inheritance. The most significant GO terms were related to regulation of chemokine production (GO:0032642; p = 2.75E-4), endocytosis (GO:0030100; p = 8.87E-4), acute inflammatory response (GO:0006953; p = 2.41E-3) and leukocyte activation involved in inflammatory response (GO:0002269; p = 2.51E-3). They all reflect mechanisms of viral infection, endocytosis and immune mediated inflammatory response (Lindenbach and Rice, 2003; Gao et al., 2020). Interestingly, we also found evidence for microglial cell activation (GO:0001774; p = 2.51E-3) only in patients with ADEM after CHIKV, which is involved in immune regulation and CNS autoimmune disease (Dheen et al., 2007; O'Loughlin et al., 2018). Neuroinflammation is a complex integration of the responses of all cells present within the CNS and consequent microglial activation and leukocyte infiltration (Carson et al., 2006).
The demyelinating lesions found in most MS and NMOSD patients are mainly associated with T cell inflammation, with the involvement of both microglia and infiltrating peripheral macrophages - as revealed by TMEM119 immunohistochemical profiling - in the pathology of the lesions (Popescu and Lucchinetti, 2012; Zrzavy et al., 2017; Hayashida et al., 2020). Interestingly, infection of the CNS by the CHIKV has been linked to microglial activation (Ganesan et al., 2008; Abere et al., 2012). The overproduction of inflammatory modulators and oxidative stress may induce mitochondrial dysfunction of oligodendrocytes in demyelinating diseases, such as MS, that may cause irreversible cell damage and consequently play a key role in CNS demyelination (Liñares et al., 2006; Venugopalan et al., 2014; Banerjee and Mukhopadhyay, 2018; Colavita et al., 2018; Lan et al., 2018). Recently, the International Multiple Sclerosis Genetics Consortium performed a large genetic association study, which identified an enrichment of genetic variants in human microglia but not in astrocytes or neurons (International Multiple Sclerosis Genetics Consortium, 2019).
Large GWAS studies have identified the tyrosine kinase 2 (TYK2) gene as associated with susceptibility to MS (Ban et al., 2009; Mero et al., 2010). In patient 18ZC026 (myelitis), we found the rs12720356 variant in the TYK2 gene. Of note, this SNV introduces a missense mutation (I684S) in the catalytic domain, which maintains residual function in response to IFN-α/β and other pro-inflammatory cytokines. It is known that TYK2 kinase impaired activity impacts type I IFN signaling and alters defense against the virus (Prchal-Murphy et al., 2012). This non-synonymous variant might decrease IFN-I signaling, hindering anti-viral immunity, and if associated with HLA alleles may be related to the development of IDD.
Noteworthy, HLA class II alleles have been linked to a wide range of autoimmune diseases (Svejgaard, 2008; Asgari et al., 2012; Bodis et al., 2018). Here, we found the DR15 haplotype (DRB1*15:01-DQA1*01:02-DQB1*06:02) and the HLA-DRB5*01:01 allele (DR2) in patient 19ZC160, while at least one of the alleles related to these haplotypes was found in the other ADEM patients. It is important to highlight that we found the HLA-DRB1*15:01 allele only in patients with ADEM, which is clinically cataloged as a risk factor for MS (VCV000029757). The literature reports that ADEM can progress similar to MS and these HLA haplotypes are the main genetic risk factor that contribute to the etiology of MS (Schmidt et al., 2007; Etzensperger et al., 2008).
Some authors have already described that the presence of certain HLA alleles may be related to the development of ADEM, especially in childhood (Ramachandran Nair, 2004; Waubant et al., 2016). Patient 19ZC179 carries HLA-DRB1*01:01 and HLA-DRB5*01:01 alleles, which were associated with ADEM in Russian and Korean populations (Idrissova et al., 2003; Oh et al., 2004). We also identified the HLA-DRB1*15:03 allele in patient 19ZC129 without neurological manifestation. This allele has been associated with ADEM in a small case-control study in Brazil, but results still lack replication in larger cohorts (Alves-Leon et al., 2009).
Two of the 3 patients who developed myelitis associated to CHIKV infection carry the HLA-DRB1*03:01 allele, linked to NMOSD in Brazilian patients from Rio de Janeiro (Alvarenga et al., 2017), whereas that same allele was identified in only one individual without neurological manifestations. It is important to note that HLA-DRB1*03:01 has also been associated to susceptibility to rheumatoid arthritis, mainly when inherited together with HLA-DRB1*09:01, which is the case of this patient (van Drongelen and Holoshitz, 2017). In addition, the HLA-DRB1*09:01 allele has already been reported as a protective factor for NMOSD in a Japanese population (Matsushita et al., 2020).
All patients who developed myelitis following CHIKV infection carried the HLA-A*23:01 allele, whereas it was identified in only one individual without neurological manifestation. The distribution of HLA-A*23:01 has been analyzed in different Brazilian regions and throughout the world, specifically in different ethnic groups from Rio de Janeiro, and an allele frequency lower than <0.1 has been reported (http://allelefrequencies.net). On the other hand, alleles already reported in the literature as protectors against autoimmune diseases were found only in patients without IDD. For example, HLA-B*44:02 is a protective allele for MS susceptibility, associated with better MRI outcomes in terms of brain parenchymal fraction and T2 hyperintense lesion volume (Healy et al., 2010). Of particular interest, HLA-DRB1*13 alleles are the protective alleles shared by multiple autoimmune diseases and it has already been observed that HLA-DRB1*13:02 confers protection against organ-specific autoimmune diseases and some infectious diseases (Furukawa et al., 2017).
The variant that most caught our attention was rs61742988 (MBP) in patient 19ZC160, who also carries HLA DR15 and DR2 haplotype. To our knowledge, the rs61742988 variant has been described in few publications and classified as having uncertain significance, according to the ACMG guideline (Richards et al., 2015). Independent studies have also shown this mutation in the MBP gene to be associated to clinical course in MS (Zhou et al., 2017) and, in association with HLA-DRB1*15:01, the allele may be considered a predisposing factor for this disorder (Nejati et al., 2017).
Candidate gene approaches or GWAS studies have not been able to clearly establish any markers that predict susceptibility or clinical course after CHIKV infection. Indeed, different disease outcomes are like multifactorial traits, in which phenotypes is determined by the interaction between several genes, environment factors and epigenetic mechanisms (Hill, 2006; Oksenberg and Baranzini, 2010; Paschos and Allday, 2010; Ho et al., 2012; Ladd-Acosta and Fallin, 2016). The exact mechanisms whereby CHIKV infection influences CNS IDDs are not yet well-established and we suggest that in genetically susceptible individuals, molecular mimicry could induce myelin-reactive T cells or autoantibody production, resulting in cross-reaction to both viral and myelin proteins (Oldstone, 1998).
We know there are more than 200 genetic variants in coding and non-coding regions of DNA that are related to susceptibility or progression of MS (Cotsapas and Mitrovic, 2018) and MS genetic burden (MSGB) score, one of the most used concepts today, is based on a weighted scoring algorithm using SNPs associated with MS (Isobe et al., 2016). Therefore, one of the important limitations of our study is related to the non-identification of genetic variants in non-coding regions.
Regarding genotype-phenotype correlations, the most interesting finding in the present study was the specific HLA-DRB1 allele conferring a similar phenotype to MS and NMOSD. Taken together, our data suggest a role for HLA alleles and candidate genes in the development of demyelinating events following CHIKV infection. Further analyses in larger cohorts are required to replicate and functionally validate these results. Questions that still need to be answered include whether exposure to new viral antigens in populations around the world, through deforestation, species migrations and environmental changes, may impact the incidence of autoimmune diseases in the future.
Data Availability Statement
The WES dataset used in our study is publicly available in SRA-NCBI (www.ncbi.nlm.nih.gov/sra), SRA accession PRJNA680041.
Ethics Statement
The studies involving human participants were reviewed and approved by Research Ethics Committee of HUGG/UNIRIO (protocol number: 69411317.6.0000.5258) and a written informed consent was signed by all participants by the time of inclusion in the study. Written informed consent to participate in this study was provided by the participants' legal guardian/next of kin.
Author Contributions
SVA-L: designed the research clinical and data information, neurological evaluation, data analysis, and drafted the manuscript. CdSF and RdSFJ: exome data analysis and drafted the manuscript. ALH: blood samples analysis and initial draft of the manuscript. FLF-D: recruited patients and sorted out clinical information, collected the blood samples, data analysis, and drafted the manuscript. FCR-L: acquisition, analysis of radiological image data, and drafted the manuscript. JPdCG and ADdA: recruited patients and sorted out clinical information, neurological evaluation, and collected the blood samples. CCdSR: recruited patients and sorted out clinical information and neurological evaluation. LMH: blood samples analysis. ALG and APdCG: exome sequencing. MTdM, MCdPT, and RAM: blood samples analysis. BMGN and LCF: collected the blood samples. MMdS, CN, ASdSC, and CCFV: recruited patients and sorted out clinical information, and neurological evaluation. AT: review of the manuscript. OCFJ and RSA: blood samples analyses and review of the manuscript. CCC: data analysis and drafted the manuscript. ATRdV: coordinated the exome experiments, data analysis, drafted the manuscript, and critical review of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by FINEP (Grant No. 01.16.0078.00) and Rede Nacional de Especialistas em ZIKA e Doenças Correlatas (RENEZIKA) (CNPq Number 440779/2016-2), Coordination for the Improvement of Higher Education Personnel (CAPES Number 88887.130752/2016-00), Department of Science and Technology (DECIT No. 14/2016). Brazilian National Council for Scientific and Technological Developmen,and Foundation for Rio de Janeiro State Research (FAPERJ). ATRdV is supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico–CNPq (303170/2017-4) and FAPERJ (E-26/202.903/20). RSA is supported by CNPq 312688/2017-2, 439119/2018-9; FAPERJ 202.922/2018. CCC is supported by FAPERJ (E-26/202.791/2019). RdSFJ is a recipient of a graduate fellowship from CNPq.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We appreciate the support of Secretaria de Saúde do Estado do Rio de Janeiro and Laboratório Central Noel Nutels (LACEN).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2021.639364/full#supplementary-material
Supplementary Figure 1. Flowchart of gene panel prioritization. Strategies of variant filtration based on gene panel selection for CHIKV infection and neurological impairment. The main genes related to CHIKV infection, Myelitis and ADEM were firstly retrieved from the literature. Next, we expanded the gene set by retrieving genes related to enriched GO terms. To further investigate the relationship between the two sets (genes from the literature and recovered from GO) a pairwise protein-protein interaction (connectome) was performed. We retrieved the mendelian inheritance model for each gene by querying the OMIM database. Finally, we prioritize possibly damaging variants mapped in the final genes list for each patient.
Supplementary Figure 2. Flowchart of the pipeline used to filter genetic variants and predict HLA alleles. From fastq files originated by WES, the pipeline was subdivided into two main approaches: (i) variant prioritization and (ii) HLA allele prediction. The set of variants selected was analyzed sequentially using computational tools to predict pathogenicity. They were then filtered by low frequency using population databases and further classified according to clinical guidelines from pathogenicity databases. Prediction of HLA alleles (class I and II) was performed with the HLAminer tool.
Supplementary Figure 3. Central nervous system MRI of case 1. Sagittal STIR (A) and T1 FS post-contrast (B) and sagittal T2 (C) and T1 FS post-contrast (D) of the cervical and dorsal spinal cord respectively show sparse hyperintense focci (white arrows), some with gadolinium enhancement, notably in the cervical-dorsal transition, extending <1 vertebral body, mostly affecting the posterior columns on axial T2 WI (white arrows) (E,F), resembling MS like pattern dissemination.
Supplementary Figure 4. Central nervous system MRI of case 2. Brain axial FLAIR images (A–C) show hyperintense cortical-justacortical lesions identified in the parietal and occipital lobes (white arrows), in the periventricular white matter and adjacent to the occipital horns of the lateral ventricles, characterizing an encephalitis pattern.
Supplementary Figure 5. Central nervous system MRI of case 3. Dorsal spinal cord sagittal T2 WI (A) have multiple peripheral hypersignal intensity focci (white arrows), with contrast enhancement on FS post-contrast T1 WI (B). Tiny focci of hypersignal intensity peripherally located on cervical axial T2 WI (C,D), which are elongated on sagittal T2 WI (white arrow) (E).
Supplementary Figure 6. Central nervous system MRI of case 4. Brain axial FLAIR (A,B) and the corresponding post-contrast T1 WI images (C,D) show tumefactive hipersignal intensity lesions with irregular and discontinued peripheral contrast enhancement (white arrow), determining mass effect predominantly in the frontal-parietal left lobe, such as in an ADEM pattern. Sagittal STIR and post-contrast T1 WI of the cervical (E,F) and dorsal (G,H) spinal cord have confluent and extensive hypersignal lesions (white arrows), with transversal damage in the dorsal area, resembling LETM pattern.
Supplementary Figure 7. Central nervous system MRI of case 5. Sagittal STIR of cervical (A) and dorsal (B) spinal cord with multiple elongated hypersignal images peripherally located along the white matter tracts (white arrows), associated with elongated hypersignal images in a radial distribution, from the central spinal cord gray matter to the peripheral white matter, best identified on axial T2 images (C).
Supplementary Figure 8. Central nervous system MRI of case 6. Hyperintense lesion in the central pons identified on axial FLAIR (A) and T2-WI (white arrows) (B), in the medium cerebellar peduncles. There is diffuse leptomeningeal gadolinium enhancement noticed on post-contrast axial FLAIR images (white arrow) (D) but non-observed on pre-contrast FLAIR (C). Anterior horn diffuse damage is noticed on sagittal T2-WI of the cervical (E) and dorsal (F) spinal cord, which is confirmed on axial images (white arrow) (G,H).
Supplementary Table 1. Clinical and neurological manifestations associated with virus diagnosis and systemic prodrome interval, cerebrospinal fluid results, comorbidities, and outcomes.
Supplementary Table 2. Pathogenic profile variants identified by whole-exome sequencing analysis.
Supplementary Table 3. Distribution of HLA alleles predicted from exomes of 6 patients with CHIKV-related CNS IDD and 6 patients with CHIKV without IDD.
Supplementary Table 4. Gene Ontology categories ranked for the 29 genes filtered in our analysis.
References
Abere, B., Wikan, N., Ubol, S., Auewarakul, P., Paemanee, A., Kittisenachai, S., et al. (2012). Proteomic analysis of chikungunya virus infected microgial cells. PLoS ONE 7:e34800. doi: 10.1371/journal.pone.0034800
Adzhubei, I., Jordan, D. M., and Sunyaev, S. R. (2013). Predicting functional effect of human missense mutations using PolyPhen-2. Curr. Protoc. Hum. Genet. Chapter 7, Unit 7.20. doi: 10.1002/0471142905.hg0720s76
Alvarenga, M. P., Fernandez, O., Leyva, L., Campanella, L., Vasconcelos, C. F., Alvarenga, M., et al. (2017). The HLA DRB1*03:01 allele is associated with NMO regardless of the NMO-IgG status in Brazilian patients from Rio de Janeiro. J. Neuroimmunol. 310, 1–7. doi: 10.1016/j.jneuroim.2017.05.018
Alves-Leon, S. V., Lima, M., da, R., Nunes, P. C. G., Chimelli, L. M. C., Rabelo, K., et al. (2019). Zika virus found in brain tissue of a multiple sclerosis patient undergoing an acute disseminated encephalomyelitis-like episode. Mult. Scler. 25, 427–430. doi: 10.1177/1352458518781992
Alves-Leon, S. V., Veluttini-Pimentel, M. L., Gouveia, M. E., Malfetano, F. R., Gaspareto, E. L., Alvarenga, M. P., et al. (2009). Acute disseminated encephalomyelitis: clinical features, HLA DRB1*1501, HLA DRB1*1503, HLA DQA1*0102, HLA DQB1*0602, and HLA DPA1*0301 allelic association study. Arq. Neuropsiquiatr. 67, 643–651. doi: 10.1590/S0004-282X2009000400013
Anand, K. S., Agrawal, A. K., Garg, J., Dhamija, R. K., and Mahajan, R. K. (2019). Spectrum of neurological complications in chikungunya fever: experience at a tertiary care centre and review of literature. Trop. Doct. 49, 79–84. doi: 10.1177/0049475518825219
Anaya, J.-M., Ramirez-Santana, C., Salgado-Castaneda, I., Chang, C., Ansari, A., and Gershwin, M. E. (2016). Zika virus and neurologic autoimmunity: the putative role of gangliosides. BMC Med. 14:49. doi: 10.1186/s12916-016-0601-y
Asgari, N., Nielsen, C., Stenager, E., Kyvik, K. O., and Lillevang, S. T. (2012). HLA, PTPN22 and PD-1 associations as markers of autoimmunity in neuromyelitis optica. Mult. Scler. 18, 23–30. doi: 10.1177/1352458511417480
Ban, M., Goris, A., Lorentzen, A. R., Baker, A., Mihalova, T., Ingram, G., et al. (2009). Replication analysis identifies TYK2 as a multiple sclerosis susceptibility factor. Eur. J. Hum. Genet. 17, 1309–1313. doi: 10.1038/ejhg.2009.41
Banerjee, N., and Mukhopadhyay, S. (2018). Oxidative damage markers and inflammatory cytokines are altered in patients suffering with post-chikungunya persisting polyarthralgia. Free Radic. Res. 52, 887–895. doi: 10.1080/10715762.2018.1489131
Bodis, G., Toth, V., and Schwarting, A. (2018). Role of human leukocyte antigens (HLA) in autoimmune diseases. Rheumatol. Ther. 5–20. doi: 10.1007/s40744-018-0100-z
BRASIL (2020). Monitoramento Dos Casos de Arboviroses Urbanas Transmitidas Pelo Aedes Aegypti (Dengue, Chikungunya e Zika), Semanas Epidemiológicas 1 a 46. Available online at: https://www.gov.br/saude/pt-br/media/pdf/2020/dezembro/11/boletim_epidemiologico_svs_48.pdf (accessed February 18, 2021).
Brum, D. G., Barreira, A. A., dos Santos, A. C., Kaimen-Maciel, D. R., Matiello, M., Costa, R. M., et al. (2010). HLA-DRB association in neuromyelitis optica is different from that observed in multiple sclerosis. Mult. Scler. 16, 21–29. doi: 10.1177/1352458509350741
Burnard, S., Lechner-Scott, J., and Scott, R. J. (2017). EBV and MS: major cause, minor contribution or red-herring? Mult. Scler. Relat. Disord. 16, 24–30. doi: 10.1016/j.msard.2017.06.002
Burt, F. J., Chen, W., Miner, J. J., Lenschow, D. J., Merits, A., Schnettler, E., et al. (2017). Chikungunya virus: an update on the biology and pathogenesis of this emerging pathogen. Lancet Infect. Dis. 17, e107–e117. doi: 10.1016/S1473-3099(16)30385-1
Carson, M. J., Thrash, J. C., and Walter, B. (2006). The cellular response in neuroinflammation: the role of leukocytes, microglia and astrocytes in neuronal death and survival. Clin. Neurosci. Res. 6, 237–245. doi: 10.1016/j.cnr.2006.09.004
Cerny, T., Schwarz, M., Schwarz, U., Lemant, J., Gérardin, P., and Keller, E. (2017). The range of neurological complications in chikungunya fever. Neurocrit. Care 27, 447–457. doi: 10.1007/s12028-017-0413-8
Chaaithanya, I. K., Muruganandam, N., Surya, P., Anwesh, M., Alagarasu, K., and Vijayachari, P. (2016). Association of oligoadenylate synthetase gene cluster and DC-SIGN (CD209) gene polymorphisms with clinical symptoms in chikungunya virus infection. DNA Cell Biol. 35, 44–50. doi: 10.1089/dna.2015.2819
Chen, Q. Y., Nadell, D., Zhang, X. Y., Kukreja, A., Huang, Y. J., Wise, J., et al. (2000). The human leukocyte antigen HLA DRB3*020/DQA1*0501 haplotype is associated with Graves' disease in African Americans. J. Clin. Endocrinol. Metab. 85, 1545–1549. doi: 10.1210/jc.85.4.1545
Colavita, F., Vita, S., Lalle, E., Carletti, F., Bordi, L., Vincenti, D., et al. (2018). Overproduction of IL-6 and Type-I IFN in a lethal case of chikungunya virus infection in an elderly man during the 2017 Italian outbreak. Open Forum Infect. Dis. 5:ofy276. doi: 10.1093/ofid/ofy276
Cortes, L. M., Baltazar, L. M., Lopez-Cardona, M. G., Olivares, N., Ramos, C., Salazar, M., et al. (2004). HLA class II haplotypes in Mexican systemic lupus erythematosus patients. Hum. Immunol. 65, 1469–1476. doi: 10.1016/j.humimm.2004.09.008
Cotsapas, C., and Mitrovic, M. (2018). Genome-wide association studies of multiple sclerosis. Clin. Transl. Immunology 7:e1018. doi: 10.1002/cti2.1038
Delogu, L. G., Deidda, S., Delitala, G., and Manetti, R. (2011). Infectious diseases and autoimmunity. J. Infect. Dev. Ctries. 5, 679–687. doi: 10.3855/jidc.2061
Deschamps, R., Paturel, L., Jeannin, S., Chausson, N., Olindo, S., Béra, O., et al. (2011). Different HLA class II (DRB1 and DQB1) alleles determine either susceptibility or resistance to NMO and multiple sclerosis among the French Afro-Caribbean population. Mult. Scler. 17, 24–31. doi: 10.1177/1352458510382810
Dheen, S. T., Kaur, C., and Ling, E.-A. (2007). Microglial activation and its implications in the brain diseases. Curr. Med. Chem. 14, 1189–1197. doi: 10.2174/092986707780597961
Dutta, S. K., and Tripathi, A. (2017). Association of toll-like receptor polymorphisms with susceptibility to chikungunya virus infection. Virology 511, 207–213. doi: 10.1016/j.virol.2017.08.009
Dyment, D. A., Herrera, B. M., Cader, M. Z., Willer, C. J., Lincoln, M. R., Sadovnick, A. D., et al. (2005). Complex interactions among MHC haplotypes in multiple sclerosis: susceptibility and resistance. Hum. Mol. Genet. 14, 2019–2026. doi: 10.1093/hmg/ddi206
Etzensperger, R., McMahon, R. M., Jones, E. Y., and Fugger, L. (2008). Dissection of the multiple sclerosis associated DR2 haplotype. J. Autoimmun. 31, 201–207. doi: 10.1016/j.jaut.2008.04.016
Fadista, J., Oskolkov, N., Hansson, O., and Groop, L. (2017). LoFtool: a gene intolerance score based on loss-of-function variants in 60 706 individuals. Bioinformatics 33, 471–474. doi: 10.1093/bioinformatics/btv602
Farina, F., Picascia, S., Pisapia, L., Barba, P., Vitale, S., Franzese, A., et al. (2019). HLA-DQA1 and HLA-DQB1 alleles, conferring susceptibility to celiac disease and type 1 diabetes, are more expressed than non-predisposing alleles and are coordinately regulated. Cells 8:751. doi: 10.3390/cells8070751
Furukawa, H., Oka, S., Tsuchiya, N., Shimada, K., Hashimoto, A., Tohma, S., et al. (2017). The role of common protective alleles HLA-DRB1*13 among systemic autoimmune diseases. Genes Immun. 18, 1–7. doi: 10.1038/gene.2016.40
Ganesan, K., Diwan, A., Shankar, S. K., Desai, S. B., Sainani, G. S., and Katrak, S. M. (2008). Chikungunya encephalomyeloradiculitis: report of 2 cases with neuroimaging and 1 case with autopsy findings. AJNR Am. J. Neuroradiol. 29, 1636–1637. doi: 10.3174/ajnr.A1133
Gao, Y., Wang, K.-X., Wang, P., Li, X., Chen, J.-J., Zhou, B.-Y., et al. (2020). A novel network pharmacology strategy to decode mechanism of lang chuang wan in treating systemic lupus erythematosus. Front. Pharmacol. 11:512877. doi: 10.3389/fphar.2020.512877
Hayashida, S., Masaki, K., Suzuki, S. O., Yamasaki, R., Watanabe, M., Koyama, S., et al. (2020). Distinct microglial and macrophage distribution patterns in the concentric and lamellar lesions in Baló's disease and neuromyelitis optica spectrum disorders. Brain Pathol. 30, 1144–1157. doi: 10.1111/bpa.12898
Healy, B. C., Liguori, M., Tran, D., Chitnis, T., Glanz, B., Wolfish, C., et al. (2010). HLA B*44: protective effects in MS susceptibility and MRI outcome measures. Neurology 75, 634–640. doi: 10.1212/WNL.0b013e3181ed9c9c
Her, Z., Teng, T.-S., Tan, J. J. L., Teo, T.-H., Kam, Y.-W., Lum, F.-M., et al. (2015). Loss of TLR3 aggravates CHIKV replication and pathology due to an altered virus-specific neutralizing antibody response. EMBO Mol. Med. 7, 24–41. doi: 10.15252/emmm.201404459
Hill, A. V. S. (2006). Aspects of genetic susceptibility to human infectious diseases. Annu. Rev. Genet. 40, 469–486. doi: 10.1146/annurev.genet.40.110405.090546
Ho, S.-M., Johnson, A., Tarapore, P., Janakiram, V., Zhang, X., and Leung, Y.-K. (2012). Environmental epigenetics and its implication on disease risk and health outcomes. ILAR J. 53, 289–305. doi: 10.1093/ilar.53.3-4.289
Höftberger, R., and Lassmann, H. (2017). “Chapter 19 - inflammatory demyelinating diseases of the central nervous system,” in Handbook of Clinical Neurology, eds. G. G. Kovacs, and I. Alafuzoff (Vienna: Elsevier), 263–283. doi: 10.1016/B978-0-12-802395-2.00019-5
Idrissova, Z. R., Boldyreva, M. N., Dekonenko, E. P., Malishev, N. A., Leontyeva, I. Y., Martinenko, I. N., et al. (2003). Acute disseminated encephalomyelitis in children: clinical features and HLA-DR linkage. Eur. J. Neurol. 10, 537–546. doi: 10.1046/j.1468-1331.2003.00639.x
International Multiple Sclerosis Genetics Consortium (2019). Multiple sclerosis genomic map implicates peripheral immune cells and microglia in susceptibility. Science 365:eaav7188. doi: 10.1126/science.aav7188
Isobe, N., Keshavan, A., Gourraud, P.-A., Zhu, A. H., Datta, E., Schlaeger, R., et al. (2016). Association of HLA genetic risk burden with disease phenotypes in multiple sclerosis. JAMA Neurol. 73, 795–802. doi: 10.1001/jamaneurol.2016.0980
Itan, Y., Zhang, S.-Y., Vogt, G., Abhyankar, A., Herman, M., Nitschke, P., et al. (2013). The human gene connectome as a map of short cuts for morbid allele discovery. Proc. Natl. Acad. Sci. U.S.A. 110, 5558–5563. doi: 10.1073/pnas.1218167110
Kakalacheva, K., Münz, C., and Lünemann, J. D. (2011). Viral triggers of multiple sclerosis. Biochim. Biophys. Acta 1812, 132–140. doi: 10.1016/j.bbadis.2010.06.012
Kattimani, Y., and Veerappa, A. M. (2018). Complex interaction between mutant HNRNPA1 and gE of varicella zoster virus in pathogenesis of multiple sclerosis. Autoimmunity 51, 147–151. doi: 10.1080/08916934.2018.1482883
Kaushansky, N., and Ben-Nun, A. (2014). DQB1*06:02-associated pathogenic anti-myelin autoimmunity in multiple sclerosis-like disease: potential function of DQB1*06:02 as a disease-predisposing allele. Front. Oncol. 4:280. doi: 10.3389/fonc.2014.00280
Kircher, M., Witten, D. M., Jain, P., O'Roak, B. J., Cooper, G. M., and Shendure, J. (2014). A general framework for estimating the relative pathogenicity of human genetic variants. Nat. Genet. 46, 310–315. doi: 10.1038/ng.2892
Krupp, L. B., Tardieu, M., Amato, M. P., Banwell, B., Chitnis, T., Dale, R. C., et al. (2013). International Pediatric multiple sclerosis study group criteria for pediatric multiple sclerosis and immune-mediated central nervous system demyelinating disorders: revisions to the 2007 definitions. Mult. Scler. 19, 1261–1267. doi: 10.1177/1352458513484547
Ladd-Acosta, C., and Fallin, M. D. (2016). The role of epigenetics in genetic and environmental epidemiology. Epigenomics 8, 271–283. doi: 10.2217/epi.15.102
Lan, M., Tang, X., Zhang, J., and Yao, Z. (2018). Insights in pathogenesis of multiple sclerosis: nitric oxide may induce mitochondrial dysfunction of oligodendrocytes. Rev. Neurosci. 29, 39–53. doi: 10.1515/revneuro-2017-0033
Lanciotti, R. S., Kosoy, O. L., Laven, J. J., Panella, A. J., Velez, J. O., Lambert, A. J., et al. (2007). Chikungunya virus in US travelers returning from India, 2006. Emerging Infect. Dis. 13, 764–767. doi: 10.3201/eid1305.070015
Lanciotti, R. S., Kosoy, O. L., Laven, J. J., Velez, J. O., Lambert, A. J., Johnson, A. J., et al. (2008). Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerging Infect. Dis. 14, 1232–1239. doi: 10.3201/eid1408.080287
Landrum, M. J., Lee, J. M., Benson, M., Brown, G. R., Chao, C., Chitipiralla, S., et al. (2018). ClinVar: improving access to variant interpretations and supporting evidence. Nucleic Acids Res. 46, D1062–D1067. doi: 10.1093/nar/gkx1153
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Li, H., and Durbin, R. (2009). Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754–1760. doi: 10.1093/bioinformatics/btp324
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., et al. (2009). The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079. doi: 10.1093/bioinformatics/btp352
Lin, J., Li, X., and Xia, J. (2016). Th17 cells in neuromyelitis optica spectrum disorder: a review. Int. J. Neurosci. 126, 1051–1060. doi: 10.3109/00207454.2016.1163550
Liñares, D., Taconis, M., Maña, P., Correcha, M., Fordham, S., Staykova, M., et al. (2006). Neuronal nitric oxide synthase plays a key role in CNS demyelination. J. Neurosci. 26, 12672–12681. doi: 10.1523/JNEUROSCI.0294-06.2006
Lindenbach, B. D., and Rice, C. M. (2003). “Molecular biology of flaviviruses,” in Advances in Virus Research (Academic Press), 23–61. doi: 10.1016/S0065-3527(03)59002-9
Louthrenoo, W., Kasitanon, N., Wichainun, R., Wangkaew, S., Sukitawut, W., Ohnogi, Y., et al. (2013). The genetic contribution of HLA-DRB5*01:01 to systemic lupus erythematosus in Thailand. Int. J. Immunogenet. 40, 126–130. doi: 10.1111/j.1744-313X.2012.01145.x
Lucas, R. M., Hughes, A. M., Lay, M.-L. J., Ponsonby, A.-L., Dwyer, D. E., Taylor, B. V., et al. (2011). Epstein-barr virus and multiple sclerosis. J. Neurol. Neurosurg. Psychiatr. 82, 1142–1148. doi: 10.1136/jnnp-2011-300174
Mack, S. J., Udell, J., Cohen, F., Osoegawa, K., Hawbecker, S. K., Noonan, D. A., et al. (2019). High resolution HLA analysis reveals independent class I haplotypes and amino-acid motifs protective for multiple sclerosis. Genes Immun. 20, 308–326. doi: 10.1038/s41435-017-0006-8
Martinelli-Boneschi, F., Currò, R., Santoro, S., Berzero, G., Sorosina, M., Ferrè, L., et al. (2020). Impact of multiple sclerosis risk loci in postinfectious neurological syndromes. Mult. Scler. Relat. Disord. 44:102326. doi: 10.1016/j.msard.2020.102326
Matsushita, T., Masaki, K., Isobe, N., Sato, S., Yamamoto, K., Nakamura, Y., et al. (2020). Genetic factors for susceptibility to and manifestations of neuromyelitis optica. Ann. Clin. Transl. Neurol. 7, 2082–2093. doi: 10.1002/acn3.51147
McKenna, A., Hanna, M., Banks, E., Sivachenko, A., Cibulskis, K., Kernytsky, A., et al. (2010). The genome analysis toolkit: a mapreduce framework for analyzing next-generation DNA sequencing data. Genome Res. 20, 1297–1303. doi: 10.1101/gr.107524.110
McLaren, W., Gil, L., Hunt, S. E., Riat, H. S., Ritchie, G. R. S., Thormann, A., et al. (2016). The ensembl variant effect predictor. Genome Biol. 17:122. doi: 10.1186/s13059-016-0974-4
Mehta, R., Gerardin, P., de Brito, C. A. A., Soares, C. N., Ferreira, M. L. B., and Solomon, T. (2018). The neurological complications of chikungunya virus: a systematic review. Rev. Med. Virol. 28:e1978. doi: 10.1002/rmv.1978
Menge, T., Hemmer, B., Nessler, S., Wiendl, H., Neuhaus, O., Hartung, H.-P., et al. (2005). Acute disseminated encephalomyelitis: an update. Arch. Neurol. 62, 1673–1680. doi: 10.1001/archneur.62.11.1673
Menting, S., Thai, K. T. D., Nga, T. T. T., Phuong, H. L., Klatser, P., Wolthers, K. C., et al. (2011). Internally controlled, generic real-time PCR for quantification and multiplex real-time PCR with serotype-specific probes for serotyping of dengue virus infections. Adv. Virol. 2011:514681. doi: 10.1155/2011/514681
Mero, I.-L., Lorentzen, A. R., Ban, M., Smestad, C., Celius, E. G., Aarseth, J. H., et al. (2010). A rare variant of the TYK2 gene is confirmed to be associated with multiple sclerosis. Eur. J. Hum. Genet. 18, 502–504. doi: 10.1038/ejhg.2009.195
Miyagawa, S., Shinohara, K., Kidoguchi, K., Fujita, T., Fukumoto, T., Hashimoto, K., et al. (1997). Neonatal lupus erythematosus: studies on HLA class II genes and autoantibody profiles in Japanese mothers. Autoimmunity 26, 95–101. doi: 10.3109/08916939709003852
Mizuki, N., Ohno, S., Ando, H., Chen, L., Palimeris, G. D., Stavropoulos-Ghiokas, E., et al. (1997). A strong association between HLA-B*5101 and Behçet's disease in Greek patients. Tissue Antigens 50, 57–60. doi: 10.1111/j.1399-0039.1997.tb02835.x
Mowry, E. M., and Waubant, E. (2010). Pediatric multiple sclerosis. Continuum 16, 181–192. doi: 10.1212/01.CON.0000389941.99907.c8
Naito, S., Namerow, N., Mickey, M. R., and Terasaki, P. I. (1972). Multiple sclerosis: association with HL-A3. Tissue Antigens 2, 1–4. doi: 10.1111/j.1399-0039.1972.tb00111.x
Naslavsky, M. S., Yamamoto, G. L., de Almeida, T. F., Ezquina, S. A. M., Sunaga, D. Y., Pho, N., et al. (2017). Exomic variants of an elderly cohort of Brazilians in the ABraOM database. Hum. Mutat. 38, 751–763. doi: 10.1002/humu.23220
Nejati, P., Attar, M., Rahimian, M., Fathi, D., and Shahbazi, M. (2017). Combination of myelin basic protein gene polymorphisms with HLA-DRB1*1501 in Iranian patients with multiple sclerosis. Iran. J. Immunol. 14, 231–239. Available online at: https://iji.sums.ac.ir/article_39313.html
Ng, P. C., and Henikoff, S. (2003). SIFT: predicting amino acid changes that affect protein function. Nucleic Acids Res. 31, 3812–3814. doi: 10.1093/nar/gkg509
Oh, H. H., Kwon, S. H., Kim, C. W., Choe, B. H., Ko, C. W., Jung, H. D., et al. (2004). Molecular analysis of HLA class II-associated susceptibility to neuroinflammatory diseases in Korean children. J. Korean Med. Sci. 19, 426–430. doi: 10.3346/jkms.2004.19.3.426
Oksenberg, J. R., and Baranzini, S. E. (2010). Multiple sclerosis genetics–is the glass half full, or half empty? Nat. Rev. Neurol. 6, 429–437. doi: 10.1038/nrneurol.2010.91
Oldstone, M. B. (1998). Molecular mimicry and immune-mediated diseases. FASEB J. 12, 1255–1265. doi: 10.1096/fasebj.12.13.1255
O'Loughlin, E., Madore, C., Lassmann, H., and Butovsky, O. (2018). Microglial phenotypes and functions in multiple sclerosis. Cold Spring Harb. Perspect. Med. 8:a028993. doi: 10.1101/cshperspect.a028993
Paschos, K., and Allday, M. J. (2010). Epigenetic reprogramming of host genes in viral and microbial pathogenesis. Trends Microbiol. 18, 439–447. doi: 10.1016/j.tim.2010.07.003
Pialoux, G., Gaüzère, B.-A., Jauréguiberry, S., and Strobel, M. (2007). Chikungunya, an epidemic arbovirosis. Lancet Infect. Dis. 7, 319–327. doi: 10.1016/S1473-3099(07)70107-X
Popescu, B. F. G., and Lucchinetti, C. F. (2012). Pathology of demyelinating diseases. Annu. Rev. Pathol. 7, 185–217. doi: 10.1146/annurev-pathol-011811-132443
Prchal-Murphy, M., Semper, C., Lassnig, C., Wallner, B., Gausterer, C., Teppner-Klymiuk, I., et al. (2012). TYK2 kinase activity is required for functional type I interferon responses in vivo. PLoS ONE 7:e39141. doi: 10.1371/journal.pone.0039141
Ramachandran Nair, R. (2004). Acute disseminated encephalomyelitis: clinical features and HLA-DR linkage. Eur. J. Neurol. 11:354; author reply 354–355. doi: 10.1111/j.1468-1331.2004.00806.x
Richards, S., Aziz, N., Bale, S., Bick, D., Das, S., Gastier-Foster, J., et al. (2015). Standards and guidelines for the interpretation of sequence variants: a joint consensus recommendation of the American college of medical genetics and genomics and the association for molecular pathology. Genet. Med. 17, 405–424. doi: 10.1038/gim.2015.30
Salvetti, M., Giovannoni, G., and Aloisi, F. (2009). Epstein-barr virus and multiple sclerosis. Curr. Opin. Neurol. 22, 201–206. doi: 10.1097/WCO.0b013e32832b4c8d
Schmidt, H., Williamson, D., and Ashley-Koch, A. (2007). HLA-DR15 haplotype and multiple sclerosis: a HuGE review. Am. J. Epidemiol. 165, 1097–1109. doi: 10.1093/aje/kwk118
Sejvar, J. J., Kohl, K. S., Bilynsky, R., Blumberg, D., Cvetkovich, T., Galama, J., et al. (2007). Encephalitis, myelitis, and acute disseminated encephalomyelitis (ADEM): case definitions and guidelines for collection, analysis, and presentation of immunization safety data. Vaccine 25, 5771–5792. doi: 10.1016/j.vaccine.2007.04.060
Selleski, N., Almeida, L. M., de Almeida, F. C., Pratesi, C. B., Medeiros Nóbrega, Y. K., Gandolfi, L., et al. (2018). Prevalence Of celiac disease predisposing genotypes, including HLA-DQ2.2 variant, in Brazilian children. Arq. Gastroenterol. 55, 82–85. doi: 10.1590/s0004-2803.201800000-16
Svejgaard, A. (2008). The immunogenetics of multiple sclerosis. Immunogenetics 60, 275–286. doi: 10.1007/s00251-008-0295-1
Tanabe, I. S. B., Tanabe, E. L. L., Santos, E. C., Martins, W. V., Araújo, I. M. T. C., Cavalcante, M. C. A., et al. (2018). Cellular and molecular immune response to chikungunya virus infection. Front. Cell. Infect. Microbiol. 8:345. doi: 10.3389/fcimb.2018.00345
Testi, M., Terracciano, C., Guagnano, A., Testa, G., Marfia, G. A., Pompeo, E., et al. (2012). Association of HLA-DQB1*05:02 and DRB1*16 alleles with late-onset, nonthymomatous, AChR-Ab-positive myasthenia gravis. Autoimmune Dis. 2012:541760. doi: 10.1155/2012/541760
Tobin, W. O., Weinshenker, B. G., and Lucchinetti, C. F. (2014). Longitudinally extensive transverse myelitis. Curr. Opin. Neurol. 27, 279–289. doi: 10.1097/WCO.0000000000000093
Transverse Myelitis Consortium Working Group (2002). Proposed diagnostic criteria and nosology of acute transverse myelitis. Neurology 59, 499–505. doi: 10.1212/WNL.59.4.499
van Drongelen, V., and Holoshitz, J. (2017). Human leukocyte antigen-disease associations in rheumatoid arthritis. Rheum. Dis. Clin. North Am. 43, 363–376. doi: 10.1016/j.rdc.2017.04.003
Venugopalan, A., Ghorpade, R. P., and Chopra, A. (2014). Cytokines in acute chikungunya. PLoS ONE 9:e111305. doi: 10.1371/journal.pone.0111305
Warren, R. L., Choe, G., Freeman, D. J., Castellarin, M., Munro, S., Moore, R., et al. (2012). Derivation of HLA types from shotgun sequence datasets. Genome Med. 4:95. doi: 10.1186/gm396
Warren, R. L., and Holt, R. A. (2011). Targeted assembly of short sequence reads. PLoS ONE 6:e19816. doi: 10.1371/journal.pone.0019816
Waubant, E., Ponsonby, A.-L., Pugliatti, M., Hanwell, H., Mowry, E. M., and Hintzen, R. Q. (2016). Environmental and genetic factors in pediatric inflammatory demyelinating diseases. Neurology 87, S20–S27. doi: 10.1212/WNL.0000000000003029
Yeshokumar, A. K., and Waubant, E. (2017). “Post-infectious acute transverse myelitis,” in Pediatric Demyelinating Diseases of the Central Nervous System and Their Mimics: A Case-Based Clinical Guide, eds. E. Waubant, and T. E. Lotze (Cham: Springer International Publishing), 215–224. doi: 10.1007/978-3-319-61407-6_28
Yu, G., Wang, L.-G., Han, Y., and He, Q.-Y. (2012). clusterProfiler: an R package for comparing biological themes among gene clusters. OMICS 16, 284–287. doi: 10.1089/omi.2011.0118
Zamvil, S. S., Spencer, C. M., Baranzini, S. E., and Cree, B. A. C. (2018). The gut microbiome in neuromyelitis optica. Neurotherapeutics 15, 92–101. doi: 10.1007/s13311-017-0594-z
Zéphir, H., Fajardy, I., Outteryck, O., Blanc, F., Roger, N., Fleury, M., et al. (2009). Is neuromyelitis optica associated with human leukocyte antigen? Mult. Scler. 15, 571–579. doi: 10.1177/1352458508102085
Zhou, Y., Simpson, S. Jr, Charlesworth, J. C., van der Mei, I., Lucas, R. M., Ponsonby, A.-L., et al. (2017). Variation within MBP gene predicts disease course in multiple sclerosis. Brain Behav. 7:e00670. doi: 10.1002/brb3.670
Keywords: whole exome sequence, chikungunya, pathogenic variants, HLA alleles, inflammatory demyelinating diseases, neurological manifestations in viral infection, acute disseminated encephalomyelitis, myelitis
Citation: Alves-Leon SV, Ferreira CdS, Herlinger AL, Fontes-Dantas FL, Rueda-Lopes FC, Francisco RdS Jr, Gonçalves JPdC, de Araújo AD, Rêgo CCdS, Higa LM, Gerber AL, Guimarães APdC, de Menezes MT, Tôrres MCdP, Maia RA, Nogueira BMG, França LC, da Silva MM, Naurath C, Correia ASdS, Vasconcelos CCF, Tanuri A, Ferreira OC Jr, Cardoso CC, Aguiar RS and de Vasconcelos ATR (2021) Exome-Wide Search for Genes Associated With Central Nervous System Inflammatory Demyelinating Diseases Following CHIKV Infection: The Tip of the Iceberg. Front. Genet. 12:639364. doi: 10.3389/fgene.2021.639364
Received: 08 December 2020; Accepted: 08 February 2021;
Published: 17 March 2021.
Edited by:
Tanima Bose, Ludwig Maximilian University of Munich, GermanyReviewed by:
Abdorreza Moghadasi, Tehran University of Medical Sciences, IranKatsuhisa Masaki, University of Chicago Medical Center, United States
Copyright © 2021 Alves-Leon, Ferreira, Herlinger, Fontes-Dantas, Rueda-Lopes, Francisco, Gonçalves, de Araújo, Rêgo, Higa, Gerber, Guimarães, de Menezes, de Paula Tôrres, Maia, Nogueira, França, da Silva, Naurath, Correia, Vasconcelos, Tanuri, Ferreira, Cardoso, Aguiar and de Vasconcelos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Soniza Vieira Alves-Leon, c29uaXphdmllaXJhYWx2ZXNsZW9uQGdtYWlsLmNvbQ==; Ana Tereza Ribeiro de Vasconcelos, YXRydkBsbmNjLmJy
†These authors have contributed equally to this work