
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 28 October 2020
Sec. Evolutionary and Population Genetics
Volume 11 - 2020 | https://doi.org/10.3389/fgene.2020.555843
This article is part of the Research Topic Coping with Climate Change: A Genomic Perspective on Thermal Adaptation View all 11 articles
 Jesper G. Sørensen1*
Jesper G. Sørensen1* Tommaso Manenti1†
Tommaso Manenti1† Jesper S. Bechsgaard1
Jesper S. Bechsgaard1 Mads F. Schou2
Mads F. Schou2 Torsten N. Kristensen3
Torsten N. Kristensen3 Volker Loeschcke1
Volker Loeschcke1Organisms are exposed to temperatures that vary, for example on diurnal and seasonal time scales. Thus, the ability to behaviorally and/or physiologically respond to variation in temperatures is a fundamental requirement for long-term persistence. Studies on thermal biology in ectotherms are typically performed under constant laboratory conditions, which differ markedly from the variation in temperature across time and space in nature. Here, we investigate evolutionary adaptation and environmentally induced plastic responses of Drosophila simulans to no fluctuations (constant), predictable fluctuations or unpredictable fluctuations in temperature. We whole-genome sequenced populations exposed to 20 generations of experimental evolution under the three thermal regimes and examined the proteome after short-term exposure to the same three regimes. We find that unpredictable fluctuations cause the strongest response at both genome and proteome levels. The loci showing evolutionary responses were generally unique to each thermal regime, but a minor overlap suggests either common laboratory adaptation or that some loci were involved in the adaptation to multiple thermal regimes. The evolutionary response, i.e., loci under selection, did not coincide with induced responses of the proteome. Thus, genes under selection in fluctuating thermal environments are distinct from genes important for the adaptive plastic response observed within a generation. This information is key to obtain a better understanding and prediction of the effects of future increases in both mean and variability of temperatures.
It is well known that different adaptive responses for coping with stressful temperature conditions exist. Within generations, organisms can respond plastically to environmental changes (Pigliucci, 1996, 2001; Lande, 2009), while evolutionary responses may occur through both changes in trait means and in the level of plasticity (Hoffmann and Parsons, 1989; Williams et al., 2008; Lande, 2009; Kristensen et al., 2018). It is debated whether plasticity or evolutionary responses constitute the main contributor to temperature adaptation in small ectothermic animals (Gunderson and Stillman, 2015; Sgrò et al., 2016; Sørensen et al., 2016a). Evolutionary change in trait means is better understood and does provide evidence for local adaptation (e.g., Hoffmann et al., 2002; Kellermann et al., 2012). However, upper thermal limits seem to be evolutionary constrained in some small ectothermic insects (particularly studied in Drosophila) (Kellermann et al., 2012; Schou et al., 2014; but see discussion in Logan and Cox, 2020), while not in some species of phytoplankton (Kontopoulos et al., 2020). The constraint among species of Drosophila is not founded in an apparent lack of additive genetic variation, as significant levels of genetic variation for heat tolerance in the same species of Drosophila have been documented (Williams et al., 2012; Castaneda et al., 2019). Theory predicts that evolution of plasticity should be favored in predictably variable environments (Lande, 2009; Ashander et al., 2016). However, for plasticity in upper (and lower) thermal tolerance empirical evidence supporting this hypothesis is scarce (Gunderson and Stillman, 2015). Thus, it is not clear how and how much small ectothermic animals can and will respond to a warming climate.
Under natural conditions, organisms are exposed to temperatures that vary on diurnal (multiple exposures within a generation for most insect species) and seasonal (often across multiple generations in insects) scales. Diurnal fluctuations are often comparable to seasonal variation in magnitude, but characterized by temperature changes which occur much faster than temperature change across seasons. For example, according to the Danish Meteorological Institute, the difference in mean temperature among seasons in Denmark is ∼15°C (based on monthly mean temperatures 2006–20151). Diurnal temperatures can attain a similar range between a cold night and a warm summer day, where 10 and 25°C, respectively, can be found within the same or a few days. The variability in temperature is expected to further increase with increasing heat waves under climate change (Perkins-Kirkpatrick and Lewis, 2020). Under these conditions, it can be questioned whether evolutionary change in trait means is adequate to maintain fitness sufficient high for a species to persist (Araujo et al., 2013; Radchuk et al., 2019). Phenotypic plasticity has been suggested to be more likely to accommodate rapidly changing temperatures and extreme events (Gienapp et al., 2008; Merilä and Hendry, 2014). However, while large differences among Drosophila species in plasticity of different traits exist, within species plasticity in heat tolerance (thermal acclimation capacity) is evolving slowly; plasticity does not to differ among geographically distinct populations of the same species, and is not lost when natural populations are kept at constant temperature in the laboratory (up to 28 generations) (Overgaard et al., 2011; van Heerwaarden et al., 2014; Fragata et al., 2016; Manenti et al., 2017). Furthermore, plasticity does not change under experimental evolution (for 20 generations) in thermal regimes with different variability and predictability (Manenti et al., 2015). An explanation for this could be constraints bounded in the genetic architecture of basal and acclimation trait values (Gerken et al., 2015; see also Lecheta et al., 2020). Temperature fluctuations seem to induce heat tolerance through mechanisms different from mechanisms induced by constant temperatures. For example, thermal fluctuations induce a transcriptomic response that is different from the response induced by differences in mean temperature and from the classic heat stress response induced by increasing thermal stress (Sørensen et al., 2016b; Manenti et al., 2018). However, we have little knowledge on the molecular responses to fluctuating environments, and this limits our understanding of acclimation capacity, evolutionary constraints and trade-offs as well as the costs of induced plastic responses.
A number of evolutionary and ecological studies have recently focused on fitness consequences of short- or long-term variation of temperatures (Ketola et al., 2004; Kingsolver et al., 2009; Hallsson and Bjorklund, 2012; Manenti et al., 2014, 2015; Kellermann et al., 2015; Simões et al., 2020). The use of temperature fluctuations in laboratory experiments has been argued to have a greater ecological relevance compared to constant ones, as they are a better proxy of a natural environment (Boyce et al., 2006; Schreiber, 2010). This study aimed to investigate the molecular responses to environments that differ in amplitude and predictability of daily temperature within the life span of the organism studied. We did this using a well-known insect model system (Drosophila simulans), which can easily be collected and manipulated (e.g., applying laboratory natural selection in replicated lines) in the laboratory. To investigate plastic responses to thermal fluctuations we compared the induced proteomic expression profiles among thermal regimes prior to experimental evolution. To investigate evolutionary responses to the thermal regimes we applied full genome sequencing. Specifically, we aimed to identify candidate mechanisms for plastic responses to temperature fluctuations in the proteome, and the genomic patterns of selection responses, of replicate lines exposed to twenty generations of selection in constant, predictable or unpredictable fluctuating thermal regimes. We expect that evolutionary responses act on cis-regulatory elements and will be detected in regions of the genome that encodes proteins inducible by temperature fluctuations (plasticity genes), if inducible and evolved mechanisms of heat tolerance are shared (as predicted if plasticity evolves by genetic assimilation, Pigliucci et al., 2006). Alternatively, evolutionary responses may occur in trans-regulatory elements and observed at genome level will be independent of the proteins induced by temperature fluctuations. Furthermore, we expect that evolutionary responses will be most pronounced in the predictably fluctuating environment if amplitude drives evolution. Alternatively, evolutionary responses will be most pronounced in the unpredictably fluctuating if predictability drives evolution.
Two populations of D. simulans both collected at the same field site close to Bologna, Italy, were used in this study. The first was collected in August 2012 (referred to as the collection of ‘D. simulans 2012’) and the second was collected at the same field site in August 2014 (referred to as the collection of ‘D. simulans 2014’). Flies were throughout maintained in plastic bottles containing 50 mL of standard oatmeal-sugar-yeast-agar Drosophila medium at 23°C and a 16:8 h light:dark cycle. All experimental flies were generated using density control by transferring 40–45 eggs to plastic shell-vials with 7 mL medium. The first population (D. simulans 2012) was used to investigate the evolutionary response to selection in different thermal regimes, while the second population (D. simulans 2014) was used to investigate the plasticity induced by the same regimes, respectively (see Figure 1). The three regimes all had a mean temperature of 23°C, and were either Constant (C), Predictably fluctuating (PF), or Unpredictably fluctuating (UF) in temperature. Custom build programmable thermal cabinets maintained a 16:8 h light:dark cycle, with the C regime maintained at 23°C throughout, while the PF regime followed a 23–28–23°C sine curve during the light phase and a 23–13–23°C sine curve during the dark phase. The UF regime followed the same sine functions, but with a randomly sampled high temperature point between 23 and 28°C during the light phase and a randomly sampled low temperature point between 23 and 13°C during the dark phase (see Figure 2). The thermal regimes are described further in Manenti et al. (2015).
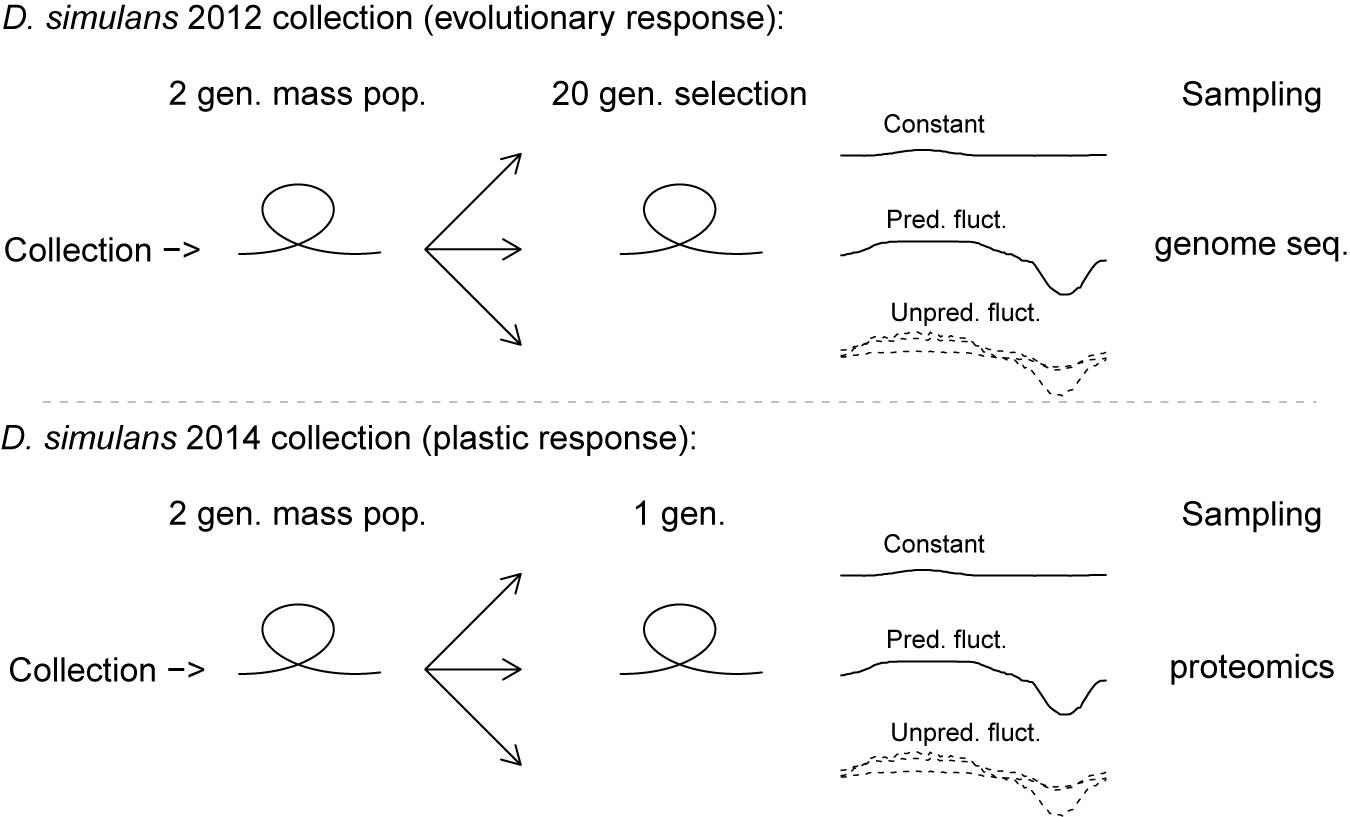
Figure 1. Infographic outlining the design of the study of laboratory responses of D. simulans to constant, predictably and unpredictable fluctuating thermal environments. Top part of the figure shows the design of the selection experiment used to evaluate evolutionary responses by genome sequencing. The lower part shows the design of the phenotypic plasticity experiment used to evaluate inducible responses by proteomics. In both experiments the thermal regimes contained independent biological controls. The plotted temperature profiles of the constant, the predictable fluctuating and the unpredictable fluctuating thermal regimes represent the realized cabinet temperatures (temperature data and the thermal regimes are described in more detail in Figure 2).
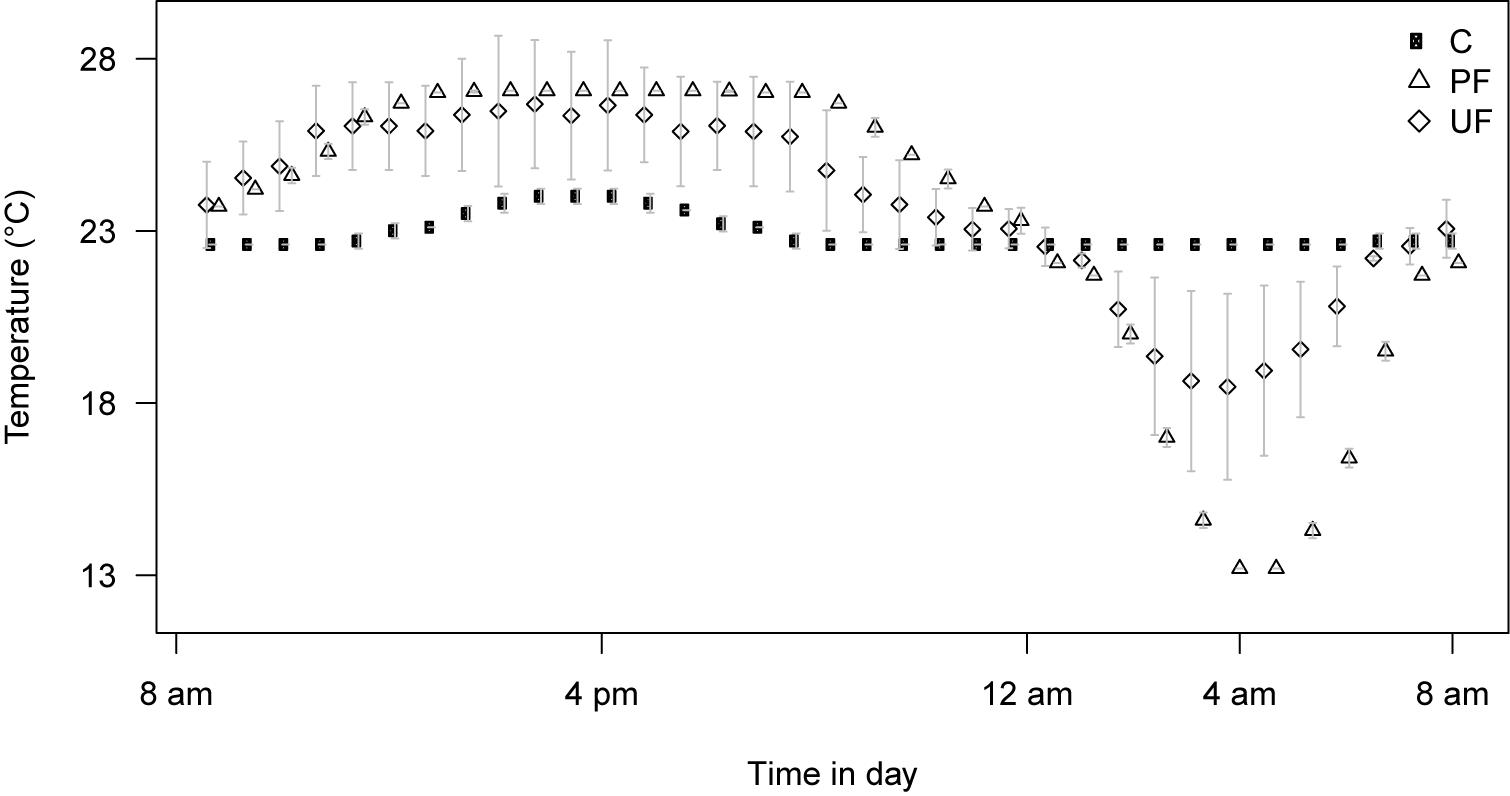
Figure 2. Temperature profile of the three thermal regimes: Constant temperature (C), Predictable fluctuating (PF), and Unpredictable fluctuating (UF). The profiles represent average ± SD for 5 days of recording (points are jittered for better evaluation of error bars). Data and figure modified from Manenti et al. (2014). The average temperature of all regimes is 23°C and flies are exposed to 16:8 h light:dark cycles (note that the light generate a small increase in temperature in the constant regime). The predictable fluctuating regime was programmed to reach 27°C during the light period and 13°C during the dark period, respectively. Low error bars indicate that this was achieved. The unpredictable fluctuating regime was programmed to reach a randomly determined setpoint between 23 and 27°C during the light period and between 23 and 13°C during the dark period, respectively. The average temperatures closer to 23°C and higher SD, respectively, indicate that the fluctuations were on average smaller in amplitude, but unpredictable among days.
The D. simulans 2012 population was used to establish a mass population based on around 350 field caught inseminated females, where after flies were randomly divided into three selection regimes (C, PF, and UF). Each selection regime had three independent biological replicates, each based on three bottles (mixed within replicates each generation) with a combined population size of >500 flies. The selection regimes and maintenance procedures are described further in Manenti et al. (2015). After 20 generations of laboratory natural selection, we froze 250 density controlled females from each biological replicate within each rearing regime, resulting in a total number of 9 samples. These pooled samples were used for full genome sequencing to investigate genomic differences in the three regimes after laboratory experimental evolution.
The D. simulans 2014 collection was used to establish a mass population based on 12 bottles of larvae that were offspring of the field collected flies (>500 adult females). This mass population was maintained for two generations in the laboratory at 23°C, before vials with 40 ± 3 eggs were collected and distributed among the three thermal regimes where they developed. Upon emergence, the flies were transferred to fresh food vials and allowed to mature in their respective thermal regimes. When flies were 3–5 days old, flies from 2–3 vials were combined and flash frozen in liquid nitrogen (without anesthesia) to be assayed for the induced response of the proteome. Sampling was performed at 9 am, when the temperature of all three regimes was around 23°C. We sorted females from these samples using a stereomicroscope placed in a 5°C room, with the flies lying on a thin sheet of plastic on top of dry ice to keep the flies as cold as possible. We collected 3 samples of 50 females from each regime (C, PF, and UF, respectively) of D. simulans 2014, which were used to investigate the inducible proteomic response to the fluctuating regimes. Furthermore, as the original samples of the founder population before selection collected from the 2012 population were lost (in the initial batch sent for sequencing), we included unselected, density controlled flies collected from D. simulans 2014 as an alternative selection control in the full genome sequencing study. For this purpose three replicates of 150 individual females were collected as described above.
We extracted DNA from 150 females pooled from each of the 12 lines (three replicates of the founder D. simulans 2014 collection, and one sample of each of the three replicated selection lines (C, PF, and UF regimes, respectively) from the D. simulans 2012 collection). DNA was Illumina sequenced (100 bp PE) by BGI Hong Kong Co., Limited. The raw reads were quality filtered using TrimGalore2 (parameters; –quality 20, –length 75). The filtered reads were mapped to the D. simulans reference genome (ASM75419v33) using bwa (version 0.7.5a, mem algorithm, default parameters). Samtools (version 1.6.0) (Li et al., 2009) was used to convert sam to bam files, to sort bam files, to remove duplicates, and to make mpileup files (mpileup −6 –q 20 –d 100). In total 93,821,525 sites were analyzed. The following analyses were done using PoPoolation2 (Kofler et al., 2011); (1) indels were identified and removed (identify-indel-region.pl –min-count 10 –indel-window 5, filter-sync-by-gft.pl), (2) resulting mpileup files were converted to synchronized (sync) files (mpileup2sync.pl –min-qual 20), and (3) downsampled to coverage 40 (downsample-synchronized.pl –target-coverage 40 –max-coverage 10000 –method fraction).
Using the samples from the unselected ‘D. simulans 2014’ as a base population, we estimated consistent allele frequency changes that have occurred during the 20 generations of experimental evolution in the three replicate populations from each of three different thermal selection regimes; constant (C), predictable fluctuating (PF) and unpredictable fluctuating (UF) temperatures. We expect genetic drift to have an equally strong impact across the three selection regimes given that population sizes were kept constant across selection regimes and replicates. Therefore consistent differences in the amount of genomic change among selection regimes will reflect differences in the strength of selection pressure. We performed the Cochran-Mantel-Haenszel (CMH) test (Agresti, 2002) (popoolation2: cmh-test.pl) to identify consistent changes (relative to the base population) in allele frequencies at all polymorphic sites (n = 2,332,305) across the entire genome (the very small chromosome 4 was not analyzed). We note that these SNPs do not represent independent loci due to linkage. To obtain an overall indication of the strength of selection pressure in the three selection regimes, we counted the number of SNPs that had higher than an arbitrarily chosen threshold p-value [–log10(p) > 7] for each selection regime and chromosome separately. Using R (vs. 3.6.1) (R Core Team, 2019) we made Manhattan plots for each chromosome and selection line by plotting the negative log10-transformed p-values as a function of chromosome position.
Consistent differences in SNP frequencies among replicates of each pair of selection regimes (C-PF, C-UP, and PF-UF) were identified using the CMH test (popoolation2: cmh-test.pl). As such differences can be the product of both random genetic drift and selection, and the identification of loci under selection is challenging. We quantified genetic drift by performing CMH tests between the first two replicates from within each of the three thermal regimes. The resulting distribution of p-values across the genome is a good representation of the pattern that we expect due to drift and other sources of structure in our data. Using this distribution we selected two thresholds, more or less conservative, to detect segments of the genome where frequency changes between two selection regimes are consistent enough that we can interpret it as a product of adaptation (0.001 and 0.0001% percentile). Quantification of drift was also done using the other pairs of replicates within thermal regimes, which produced similar patterns (see Supplementary Table 1). Convergent evolution across selection regimes may occur as a consequence of both laboratory adaptation and other similarities among the three selection regimes. We quantified the pairwise overlap of significant SNPs between two selection regimes by creating 30 bins of SNP significance in one regime, and estimating the proportion of SNPs in each bin which has been under selection (according to the most conservative threshold) in another regime. In the case of independent evolution in the two regimes of a pair, we expected the proportion of SNPs in one regime to be independent of the significance level of the bins in the other regime. Alternatively, an overlap would result in an increased proportion of SNPs under selection in one regime as the significance level of bins increased (p-values decrease) in the other regime. We performed the same analyses with randomized p-values of one of the regimes as a point of reference under the null-expectation. Finally, we used the create-genewise-sync.pl script in PoPoolation2 (Kofler et al., 2011) to only analyze SNPs located in genes. This was also done for each pair of selection lines (C-PF, C-UP, and PF-UF). Similarly, we used the CMH test for each SNP independently, but averaged p-values across each gene. We only considered genes with more than 20 SNPs.
The proteomic investigation was generally performed as described in Sørensen et al. (2017). Briefly, proteins were extracted (lysis buffer: 100 mM Triethylammonium bicarbonate (TEAB) with 0.5% sodium dodecyl sulfate (SDS) and cOmplete ULTRA Tablets (Roche Diagnostics) protease inhibitor), mechanically homogenized (Bio101, Thermo Savant FastPrep FP120 cell disruptor) on ice between cycles to keep samples cold. Protein concentration of the homogenate was determined using a QubitTM Fluorometer (Invitrogen, Life Technologies) and the Qubit Protein assay kit after centrifugation. Each sample (200 μg protein) was precipitated, dried and re-dissolved in 40 μL Dissolution Buffer with 2 μL Denaturant (iTRAQ® Reagents, AB Sciex). Proteins were reduced (Reducing Reagent, iTRAQ® Reagents, AB Sciex) and subsequently alkylated (Cysteine Blocking Reagent, iTRAQ® Reagents, AB Sciex). Proteins were then enzymatically digested over-night to peptides using a 1:50 trypsin:protein ratio (Sequencing Grade Modified Trypsin, Promega Biotech AB). Each sample was labeled with isobaric tags for relative and absolute quantitation (iTRAQ) (Pottiez et al., 2012). Hereafter, all three samples within one replicate and a common reference were pooled for fractionation and purification. Protein samples were fractioned on columns and fractions were subsequently eluted by increasing pH. Mass spectrometry analysis was performed by high-resolution electrospray ionization tandem mass spectrometry (ESI-MS/MS) (Köcher et al., 2009). Reverse phase nanoLC separation (Dionex UltiMate RSLCnano System) was performed online coupled to the mass spectrometer (LTQ Orbitrap Velos). Three technical replicates of each pooled sample replicate were analyzed, each using a slightly different gradient to increase protein coverage. Tandem mass spectrometry parameters were: positive mode, MS scan range 300–1600 with resolution at 30,000, MS/MS fragmentation was performed using HCD (higher-energy collisional dissociation) on the 20 most intense ions with a normalized collision energy of 40, dynamic exclusion of 90 s and a minimum signal threshold of 10,000.
All raw data from triplicate injections of each fraction were searched and identified (Proteome Discoverer 1.4, Reporter Ions Quantifier, Percolator validator, Mascot 2.4 Search Engine and Swiss-Prot database restricted to taxonomy ‘Drosophila’), with the Percolator algorithm using semi-supervised machine learning and a target-decoy search strategy with reversed sequences to identify correct peptide sequence matches (Käll et al., 2007; Spivak et al., 2009). Search parameters were precursor mass tolerance 10 ppm and fragment mass tolerance 0.1 Da, maximum two missed cleavages, quantification method iTRAQ 4-plex. The strict target FDR was 0.01 for high and the relaxed target FDR was 0.05 for medium confident peptide matches. Protein quantification was based exclusively on unique peptides and among replicate experimental bias correction (Latosinska et al., 2015). The proteomics analysis identified 1319 unique protein IDs in the total data (dataset available as Supplementary Data: ‘Table 1.xlsx’). Of these, 1001 unique protein IDs were detected in at least two out of three replicate samples for all selection regimes and were retained for analysis. We only accepted proteins that were detected in two of the three samples for all regimes, to avoid a strong bias from non-detected proteins. We performed ANOVA using regimes as a categorical variable. Pairwise (post hoc ANOVA) comparisons between selection lines maintained in the constant, the predictably and the unpredictably fluctuating regimes were performed on the resulting dataset. All analyses were performed using the statistical software R (vs. 3.6.1) (R Core Team, 2019) (R code available upon request to the authors).
We investigated whether SNPs under selection were physically linked to genes encoding proteins and transcripts responding to the thermal regimes. To visualize the locations, the genomic location of 204 transcripts that were previously found to show an evolutionary response to the fluctuating temperatures in their expression level (Manenti et al., 2018) were indicated on the Manhattan plots showing the divergence among selection lines. The genomic location of 34 loci that in this study were found to show a plastic response in protein expression after exposure to the three thermal regimes (see below) were also indicated in the Manhattan plots. To test if the overlap visualized on Manhattan plots was different from the null expectation, we estimated the observed distance from SNPs under selection to (1) genes with protein expression responding to the thermal regimes (plastic responses), and (2) genes with RNA expression responding to selection in the thermal regimes. These distances were compared to distances to a null expectation (distances to random genes). We estimated confidence intervals of the deviation from the null expectation by producing 10,000 random gene sets, each corresponding to the number of significant genes, and for each gene set estimating the average distance to nearest SNP under selection. We then subtracted these 10,000 estimates of distance to random gene sets, from the observed distance to the significant gene set. Distances between SNPs under selection and significant genes being smaller than distances between SNPs under selection and random genes would indicate linkage.
Populations exposed to unpredictable fluctuations showed the largest number of consistently differentiated SNPs as compared to the base population (Table 1). This was consistent across chromosome arms, with the exception of chromosome 2L. The unpredictable fluctuations therefore likely expose the flies to a stronger selection pressure than the constant and predictably fluctuating temperatures, which both showed a lower number of consistently differentiated SNPs (Table 1). However, consistent differentiation to the base population was found among lines from all three thermal regimes suggesting that all thermal regimes imposed selection. The results of the CMH analyses were visualized as Manhattan plots for each chromosome and thermal regime (Supplementary Figure 1).
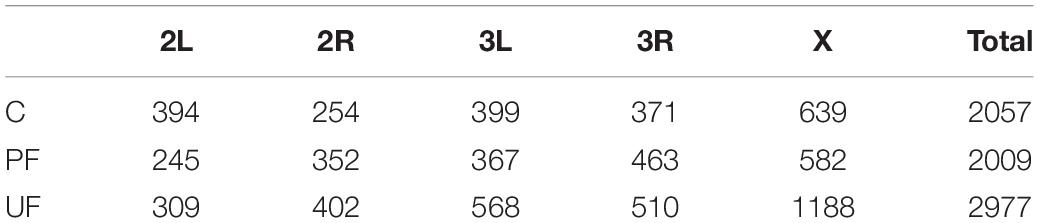
Table 1. Number of SNPs that show consistent allele frequency changes in each selection regime compared to the base population as estimated by CMH test [–log10(p) > 7] at the different chromosomes.
The number of SNPs under selection according to the drift thresholds (see Supplementary Figure 2) was highest in populations exposed to the unpredictable fluctuating temperatures (Table 2). This result was consistent when selection thresholds were estimated using the other possible within-regime population pairs to quantify genetic drift, although as expected the exact threshold and therefore absolute number of SNPs inferred to be under selection varied (Supplementary Table 1). There was only a small overlap among the SNPs under selection between regimes. The magnitude of this overlap was similar in all pairwise comparisons and across all chromosomes (Figure 3: chromosome 2L, Supplementary Figure 3: all chromosomal segments). From visual inspection of the Manhattan plots comparing pairs of selection regimes (Figure 4 and Supplementary Figure 4), it is evident that the selection responses, resulting in divergence between the three selection regimes, were not limited to a single or few loci with large consistent allele frequency changes. Rather it seems that several loci, showing smaller but consistent allele frequency changes, are involved.
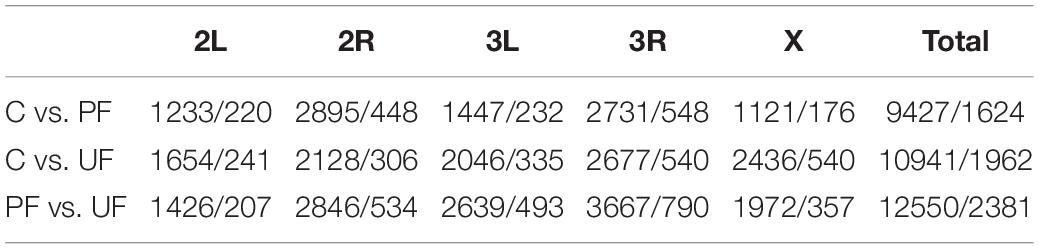
Table 2. Number of SNPs that show consistent allele frequency changes among thermal regimes and have a p-value lower than the 0.001 and 0.0001% (0.001%/0.0001%, respectively) percentile of the drift analysis at the different chromosomes and selection regimes as estimated by CMH test.
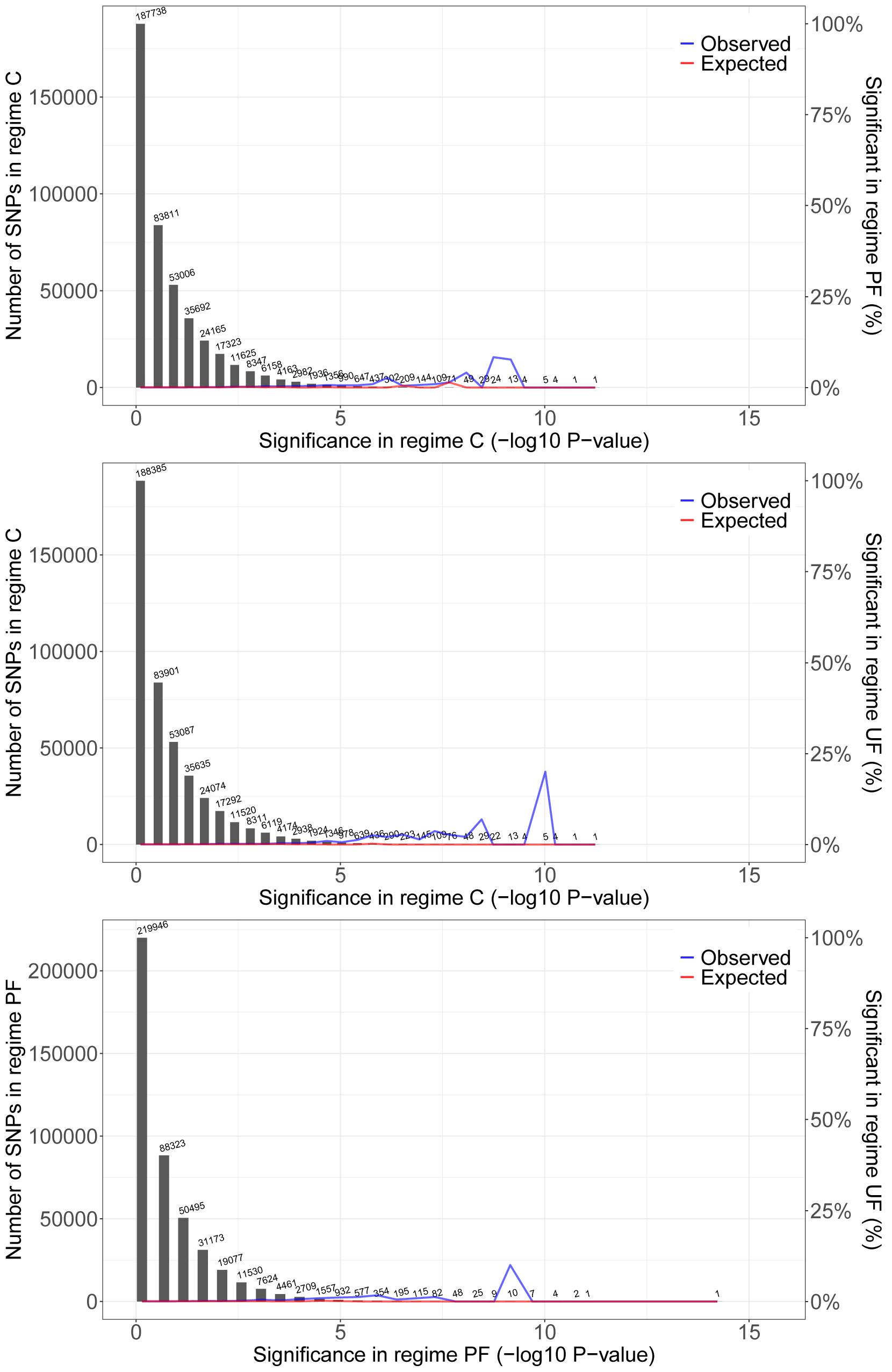
Figure 3. Pairwise overlap of significant SNPs among thermal regimes. Plot shows the distribution of SNP significance [–log10(p-values)] in one regime (gray bars), the observed proportion of SNPs that overlap with significant SNPs in the second regime, and the proportion of SNPs with randomized significance in the second regime that overlap (null-expectation). There is a consistent signal of overlap of SNPs with low p-values (in a small number of SNPs) in all pairwise comparisons. Thus, of the few SNPs with low p-values [high –log10(p)] in one selection regime a larger proportion than expected by chance also have low p-values in the other selection regimes. Only SNPs from chromosome arm 2L are presented here. Results are consistent across chromosomes (see Supplementary Figure 3).
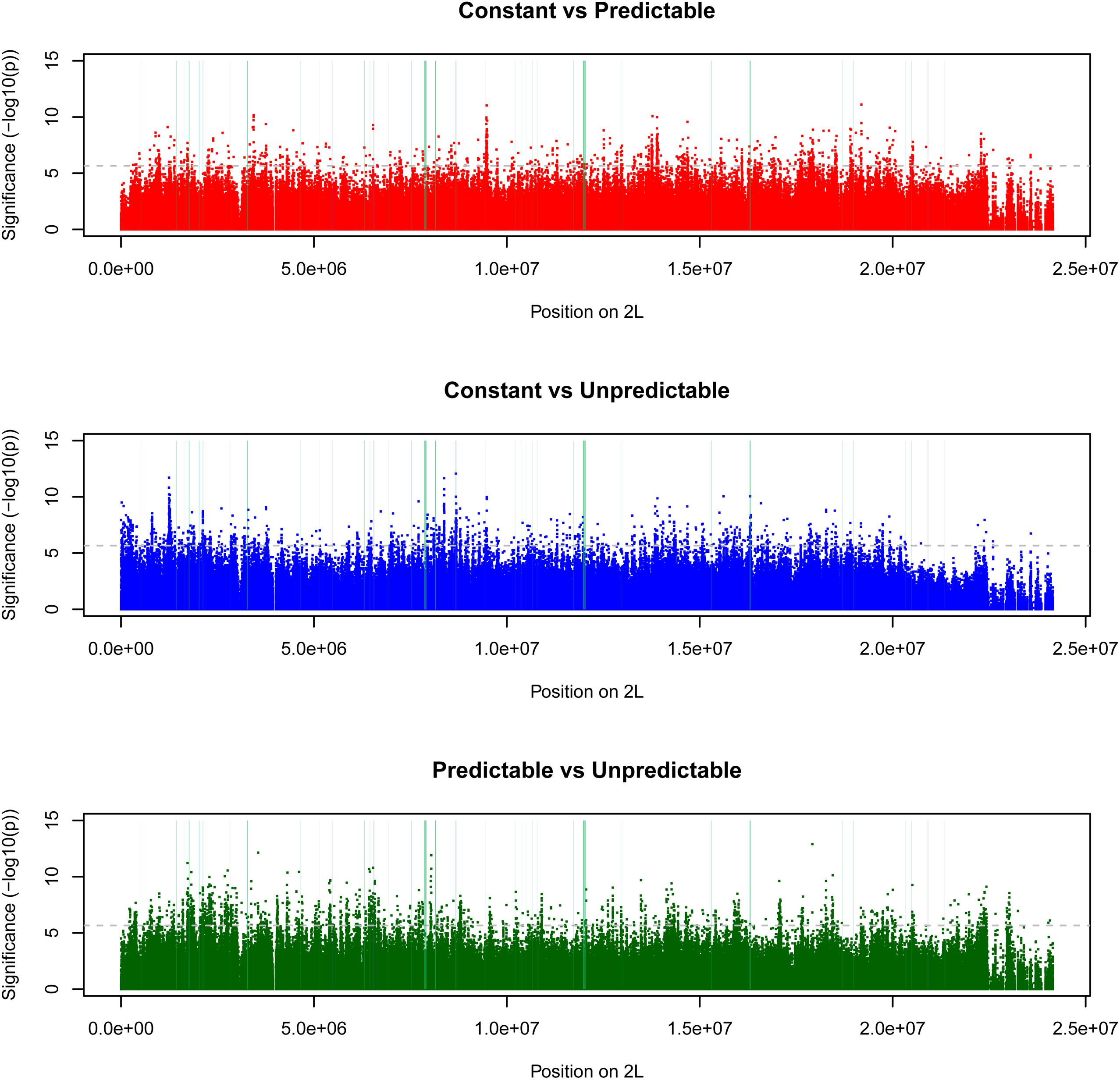
Figure 4. Manhattan plots comparing pairs of selection regimes. Horizontal lines indicate two thresholds used to infer patterns of differentiation due to evolutionary changes across selection regimes; 0.001 and 0.0001% percentiles. SNPs with [–log10(p-values)] above these thresholds can be considered evidence of divergence between selection regimes due to adaptation. Vertical lines indicate positions of loci with diverging gene expression levels (Manenti et al., 2018) (blue) or diverging protein expression levels (green), with width of line representing length of gene. Only SNPs from chromosome arm 2L are presented here. Results are consistent across chromosomes (Supplementary Figure 4).
Based on CMH analyses on SNPs located in genes, we estimated an average p-value per gene by averaging over all SNPs in each gene. We only analyzed genes having 20 SNPs or more. We used a similar approach to quantify drift as outlined above, but only using the 0.001% percentile. From each comparison among selection regimes, we only found a few genes having lower p-values than expected due to drift (Table 3). The comparisons including unpredictable fluctuations showed the highest number of genes suggested to be under selection (C-PF: 17, C-UF: 30, and PF-UF: 40). Note that 12 genes in the drift analysis have p-values lower than the threshold.
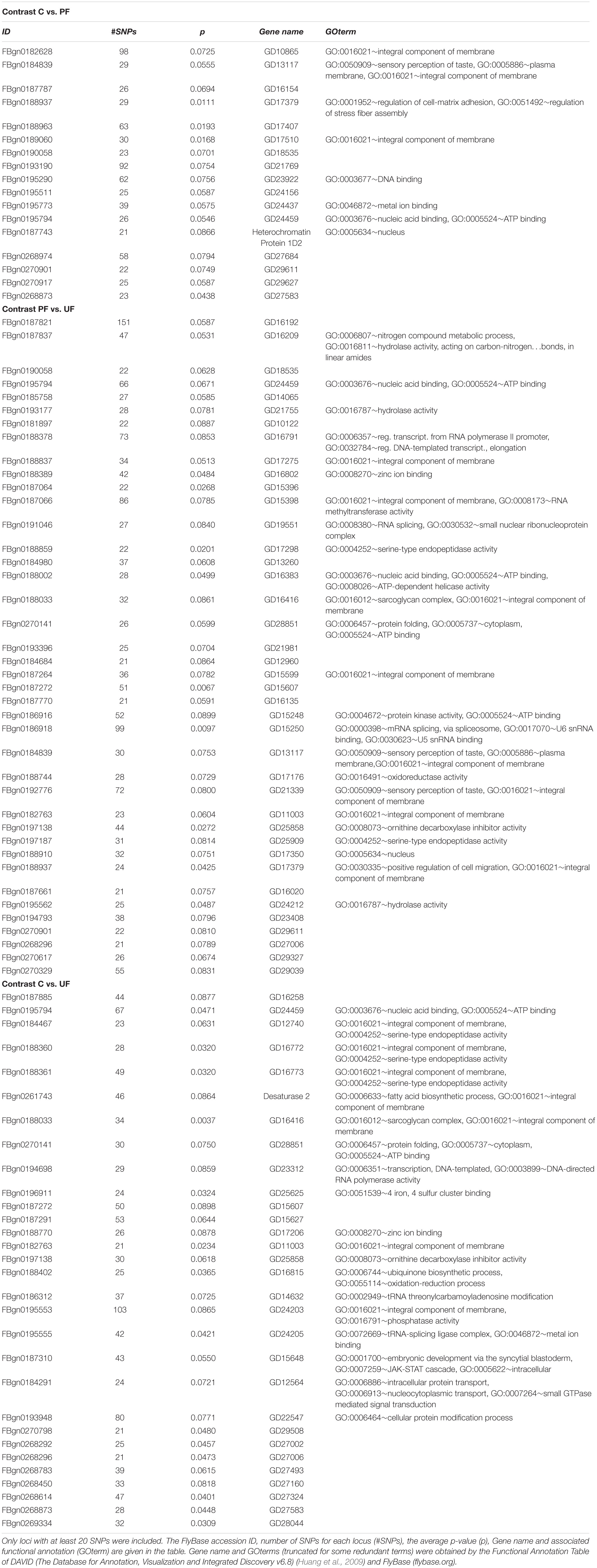
Table 3. Loci showing average p-values below what was estimated for the 99.99% drift values in the contrasts of Constant vs. Predictable fluctuating temperature (C vs. PF), Predictable fluctuating vs. Unpredictable fluctuating temperature (PF vs. UF) or Constant vs. Unpredictable fluctuating temperature (C vs. UF), respectively.
The functional annotation of the identified genes was only available for about half the genes (Table 3). Attempting to identify likely D. melanogaster orthologs resulted in ‘uncharacterized proteins.’ Among the characterized genes, many were involved in regulation of expression of the genome, i.e., processes related to transcription, translation and post-transcriptional regulation or regulation (e.g., mRNA, splicing, tRNAs, chaperonins involved in protein folding). Additionally, the desaturase gene Desat 2, involved in the modifications of fatty acids, was identified.
Of the 1001 analyzed proteins, 34 proteins showed differences in their expression levels among the three thermal regimes. Post hoc pairwise comparisons showed that none of the proteins were differentially expressed among all three regimes. Most of protein expression differences were found between the unpredictably fluctuating as compared to either the constant (24 proteins) or the predictably fluctuating (14 proteins) regime, respectively (Table 4). Eight of these proteins were shared between the two contrasts and thus showed unique expression in the unpredictable regime. The proteins showing differential expression among flies exposed to the different thermal regimes covered a range of biological functions. Most notable are proteins known to respond to environmental cues, including several heat shock proteins (Hsp60, Hsc70-5, and Hsp83), turandot proteins (Turandot A, Turandot X), a metallothionein and two odorant-binding proteins. We also found several signaling proteins (Calmodulin, Pellino) as well as proteins involved in the transcriptional and translational machinery (ribosomal proteins, elongation and translation initiation factors) and turnover (Ubiquitin-conjugating enzymes).
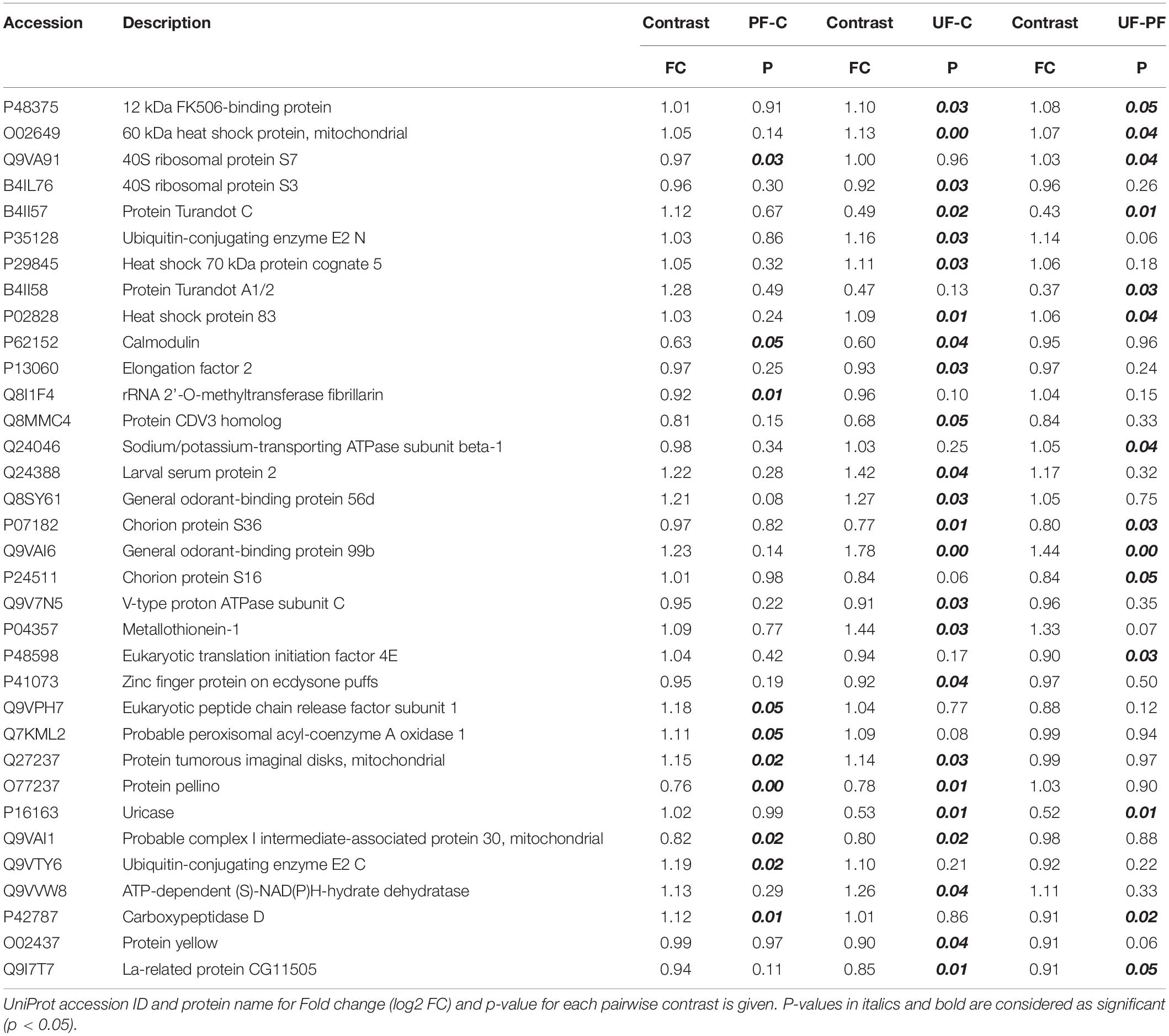
Table 4. Proteomics results.
The loci that have shown evolutionary responses in their level of transcription (Manenti et al., 2018) or proteome expression (this study) did not show obvious signs of physical linkage to SNPs under selection in the three thermal regimes. The average distance from these SNPs to nearest genes showing expression responses were in no comparison shorter than their average distance to random genes (Figure 5) suggesting that (1) evolutionary responses in gene expression to the thermal regimes are not caused by selection on cis-elements, and (2) genes that respond plastically to the thermal regimes were not under strong selection.
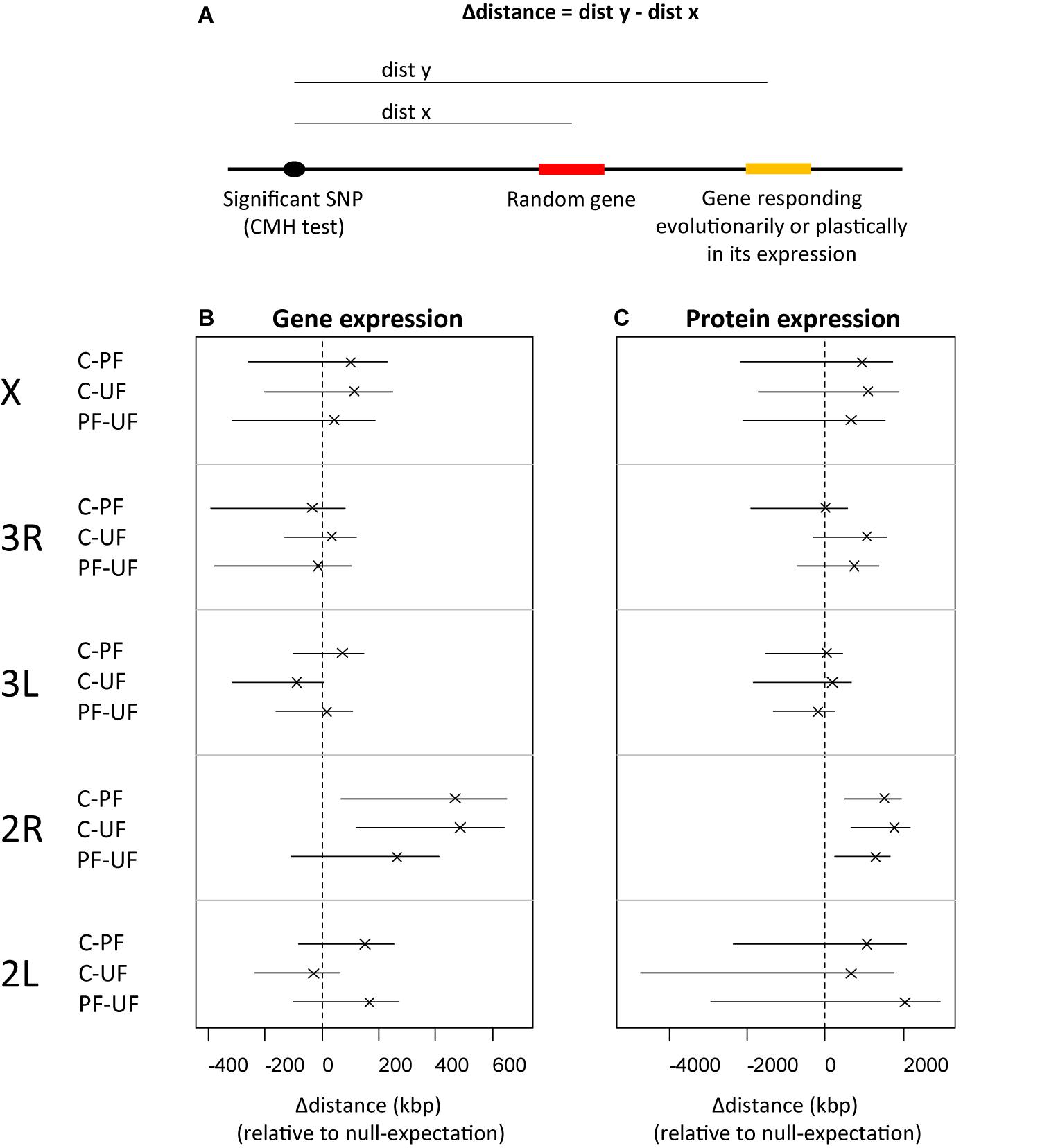
Figure 5. Represents an analysis of distance between SNPs detected to be under selection in the thermal regimes detected by the CMH test and genes that showed an evolutionary response in their gene expression and genes that show a plastic response in their proteome expression. (A) illustration of the metric used: Δdistance (the average distance between SNPs under selection and nearest gene showing expression response – the average distance between SNPs under selection and nearest gene out of a random set of 204 genes in gene expression and 34 genes in protein expression). (B,C) Δdistance calculated for each chromosome and thermal regime comparison. Crosses (x) are medians and horizontal lines (–) are 95% confidence intervals both based on 10000 random sets of genes. (B) Shows results of genes that show an evolutionary response in gene expression and (C) the results of genes that show a plastic response in proteome expression.
In this study, we applied thermal regimes with temperature variation on replicated populations of D. simulans to mimic naturally occurring daily variation (Kingsolver et al., 2009; Bozinovic et al., 2011; Manenti et al., 2014). We found a signal of the 20 generations of experimental evolution as flies maintained in constant, predictable or unpredictable fluctuating environments showed independent genomic differentiation. Natural populations are exposed, and likely adapted, to thermal fluctuations. Thus, as temperatures in natural habitats are not predictable, the unpredictable regime could be argued to represent the most natural condition (Colinet et al., 2015). Our genomic data suggests that the selection pressure in the unpredictable fluctuating regime is distinct from the constant and predictable fluctuating thermal environments. This result corroborates the induced response (i.e., results from the proteomic analysis presented here) and results from an earlier study where the transcriptome responses of the same selection lines showed similar patterns of a stronger response to unpredictable temperature fluctuations (Manenti et al., 2018). Furthermore, previous studies have documented that exposure to unpredictable temperature fluctuation was more stressful compared to a predictable fluctuating temperature regime with the same mean temperature (as measured by decreased performance in several life history and stress tolerance traits) (Manenti et al., 2014), and that performance of the lines selected in the unpredictable fluctuating regime had evolved enhanced stress tolerance (Manenti et al., 2015).
The finding that the unpredictable fluctuations impose a distinct selection pressure (even if the amplitude was smaller than that of the predictable fluctuating regime) suggests that the unpredictable selection regime does not resemble natural thermal profiles. If our unpredictably fluctuating regime did represent the natural environment, it could be hypothesized to represent the least novel environment and show the least change from the base population. It is often hypothesized that keeping up with environmental change by induction of plastic responses in variable environments can be costly (Dewitt et al., 1998; for an empirical example, see Kristensen et al., 2008). In this study (and in Manenti et al., 2018), predictability rather than amplitude seems to impose the strongest effect on both plastic and evolutionary responses. This points to that the cost of thermal fluctuations (and maybe plasticity generally) is related to the costs of evaluating the environment rather than the amount of regulation needed (Dewitt et al., 1998).
We looked for SNPs that show similar selective responses across selection regimes by comparing p-values of each SNP between each pair of the three selection regimes, and for each chromosome separately (Figure 3 and Supplementary Figure 3). There was not a strong pattern that SNPs with low p-values [high –log10(p)] in one selection regime also had low p-values in the other selection regimes, suggesting that in general the loci under selection are unique to each selection regime. We do note, however, that we did identify an overlap in a small number of SNPs across pairwise comparisons of the three selection regimes. This could represent a weak signal of a common evolutionary response to thermal regimes or shared laboratory conditions, not including temperature, that affect the same loci. While laboratory adaptation can be prominent, for thermal tolerance it does not seem to be a general concern (Maclean et al., 2018), and in this study did not overshadow the effect of the individual regimes.
Our findings support the conclusion that independent selection has occurred in response to the three different selection regimes. This underlines that fluctuating thermal regimes are affecting organism in a very complex way. Thus, the effect of natural fluctuations can be proposed to be associated both with amplitude (e.g., Terblanche et al., 2010; Bozinovic et al., 2011; Ma et al., 2015), but independently also by a marked effect of predictability, possibly mediated by the cost of continuously evaluating and adjusting to the present conditions (Dewitt et al., 1998).
Among the significantly differentially expressed proteins several heat shock and Turandot genes were found. Qualitative comparison to the study of the evolved transcriptome by Manenti et al. (2018) suggests a substantial overlap in functional groups, e.g., ribosomal genes, ubiquitin conjugating enzymes, Turandot genes, odorant binding proteins, but surprisingly no overlap with the heat shock genes found in this study. The Turandot and heat shock genes expressed in flies exposed to fluctuations (and particular to unpredictable fluctuations) indicate a heat stress response (Ekengren and Hultmark, 2001; Sørensen et al., 2003). The modest expression levels and a previous transcriptomic study suggest that the up-regulation due to temperature fluctuations are mainly constitutive, rather than tracking the temperature fluctuations (Manenti et al., 2018). The three heat shock proteins found here (Hsp83, Hsc70-5, and Hsp60) support this notion, as they have relatively high constitutive expression and are not expressed at much higher levels by stress (Feder and Hofmann, 1999; Sørensen et al., 2005). The other proteins detected as differentially expressed have no known connection to thermal tolerance, but include proteins important for efficient transcription and translation which might serve to maintain functioning of the cellular machinery allowing animals to survive and reproduce. The low number of proteins found to be differentially expressed is likely due to the relative benign extreme temperatures and the resulting low fold-change induced, but the low number of differentially expressed proteins does also indicate that regulation of quite few proteins is adequate for maintaining cellular function at variable temperatures. A recent study compared heat tolerance and the transcriptomic response to developmental acclimation at different mean temperatures (15 or 25°C), and either fluctuating or constant temperatures (Sørensen et al., 2016b). They found that fluctuations affected heat tolerance markedly, despite of a low number of transcripts (a few hundred) being differentially expressed between the constant and fluctuating regimes differentially expressed. In contrast, a large number of transcripts responded differences in mean temperatures (6000–8000 different expressed) (Sørensen et al., 2016b). This suggests that while some stress response proteins were induced by fluctuations, the molecular underpinnings of benign fluctuations might differ substantially from the pathways known to be involved with more extreme thermal acclimation or hardening.
The loci affected by selection were (for those that could be functionally annotated) related to regulation of the expression of the genome, rather than to genes with known functions in thermal responses or tolerance. This suggests that the evolutionary adaption to fluctuations within benign temperatures is to a large extent achieved by trans-regulation of the genome, rather than by increasing thermal stability and adaptation of proteins seen in extremophiles (Jollivet et al., 2012).
The distribution of distances between SNPs seemingly under selection and the transcriptome differences imposed by selection by the thermal regimes also showed no sign of cis-regulation (i.e., short distances). For the transcripts showing a change in expression following selection this might indicate some degree of trans-regulation. For the proteins responding to our thermal regimes this might suggest that evolved differences and the plastic response are not generally achieved by common genes. However, we acknowledge that linkage and linkage disequilibrium might contribute to our results. Unfortunately, lack of information on linkage in D. simulans and the pooled sequencing data prevents us from a more dedicated analysis of this effect. One potential cis-regulated mechanism is related to the Desat2 gene (a lipid desaturase), which could be related to membrane homeoviscous adaptation (Hazel, 1995). However, this remains a hypothesis for future testing.
Fluctuating temperature regimes can vary in terms of period, amplitude, rate of temperature change and predictability in addition to the mean temperature. Thus, even if studies with thermal fluctuations can be considered more ecologically relevant than those with constant temperatures, it is by no means simple to compare results across laboratory studies and to extrapolate findings to field conditions (Ketola and Kristensen, 2017). We found no marked overlap between loci being affected by selection and the proteins induced by the regimes suggesting that the evolutionary and plastic responses are achieved by distinct genes. The genes associated by many significant SNPs suggest that transcription, translation and post-translational modification are targets of evolutionary change. Increased expression of Turandot proteins under fluctuations support this group of genes/proteins as emerging candidates for mediating thermal acclimation to fluctuations. Based on our findings we argue that it is important to acknowledge that predictable and unpredictable (and constant) thermal environments have different impacts on fitness. While other studies have shown that constant and fluctuating temperatures induce different plastic responses and evolutionary pressures (Botero et al., 2015; Dey et al., 2016), this aspect is often ignored in functional and evolutionary studies on thermal adaptation. This is critical because climate models predict more variable and less predictable climates in the future (IPCC, 2014).
The raw genome sequencing data was uploaded to the NCBI SRA (short read archive) BioProject accession: PRJNA629467.
JS and VL acquired the financial support for the project leading to this publication. JS, TM, and VL conceived and designed the study. JS and TM performed experiments and data collection. JS, JB, and MS analyzed the data and wrote first draft of the manuscript. All authors contributed to the final draft and approved the final version of the manuscript.
The study was supported by the Graduate School of Science and Technology (GSST) at Science and Technology, Aarhus University (TM), and by a Sapere Aude: DFF-Starting Grant (JS) and a large frame grant (VL) from the Independent Research Fund Denmark, Natural Sciences.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors would like to thank Annemarie Højmark and Trine Bech Søgaard for their help with the laboratory work and Sandro Cavicchi for proving the flies.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2020.555843/full#supplementary-material
Araujo, M. B., Ferri-Yanez, F., Bozinovic, F., Marquet, P. A., Valladares, F., and Chown, S. L. (2013). Heat freezes niche evolution. Ecol. Lett. 16, 1206–1219.
Ashander, J., Chevin, L.-M., and Baskett, M. L. (2016). Predicting evolutionary rescue via evolving plasticity in stochastic environments. Proc. R. Soc. B Biol. Sci. 283:20161690. doi: 10.1098/rspb.2016.1690
Botero, C. A., Weissing, F. J., Wright, J., and Rubenstein, D. R. (2015). Evolutionary tipping points in the capacity to adapt to environmental change. Proc. Natl. Acad. Sci. U.S.A. 112, 184–189. doi: 10.1073/pnas.1408589111
Boyce, M. S., Haridas, C. V., Lee, C. T., and The Nceas Stochastic Demography Working Groups (2006). Demography in an increasingly variable world. Trends Ecol. Evol. 21, 141–148. doi: 10.1016/j.tree.2005.11.018
Bozinovic, F., Bastias, D. A., Boher, F., Clavijo-Baquet, S., Estay, S. A., and Angilletta, M. J. (2011). The mean and variance of environmental temperature interact to determine physiological tolerance and fitness. Physiol. Biochem. Zool. 84, 543–552.
Castaneda, L. E., Romero-Soriano, V., Mesas, A., Roff, D. A., and Santos, M. (2019). Evolutionary potential of thermal preference and heat tolerance in Drosophila subobscura. J. Evol. Biol. 32, 818–824. doi: 10.1111/jeb.13483
Colinet, H., Sinclair, B. J., Vernon, P., and Renault, D. (2015). Insects in fluctuating thermal environments. Annu. Rev. Entomol. 60, 123–140. doi: 10.1146/annurev-ento-010814-021017
Dewitt, T. J., Sih, A., and Wilson, D. S. (1998). Costs and limits of phenotypic plasticity. Trends Ecol. Evol. 13, 77–81.
Dey, S., Proulx, S. R., and Teotonio, H. (2016). Adaptation to temporally fluctuating environments by the evolution of maternal effects. PLoS Biol. 14:e1002388. doi: 10.1371/journal.pbio.1002388
Ekengren, S., and Hultmark, D. (2001). A family of Turandot-related genes in the humoral stress response of Drosophila. Biochem. Biophys. Res. Commun. 284, 998–1003.
Feder, M. E., and Hofmann, G. E. (1999). Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 61, 243–282. doi: 10.1146/annurev.physiol.61.1.243
Fragata, I., Lopes-Cunha, M., Barbaro, M., Kellen, B., Lima, M., Faria, G. S., et al. (2016). Keeping your options open: Maintenance of thermal plasticity during adaptation to a stable environment. Evolution 70, 195–206.
Gerken, A. R., Eller, O. C., Hahn, D. A., and Morgan, T. J. (2015). Constraints, independence, and evolution of thermal plasticity: probing genetic architecture of long- and short-term thermal acclimation. Proc. Natl. Acad. Sci. U. S. A. 112, 4399–4404. doi: 10.1073/pnas.1503456112
Gienapp, P., Teplitsky, C., Alho, J. S., Mills, J. A., and Merila, J. (2008). Climate change and evolution: disentangling environmental and genetic responses. Mol. Ecol. 17, 167–178. doi: 10.1111/j.1365-294X.2007.03413.x
Gunderson, A. R., and Stillman, J. H. (2015). Plasticity in thermal tolerance has limited potential to buffer ectotherms from global warming. Proc. R. Soc. B Biol. Sci. 282:20150401. doi: 10.1098/rspb.2015.0401
Hallsson, L. R., and Bjorklund, M. (2012). Selection in a fluctuating environment leads to decreased genetic variation and facilitates the evolution of phenotypic plasticity. J. Evol. Biol. 25, 1275–1290. doi: 10.1111/j.1420-9101.2012.02512.x
Hazel, J. R. (1995). Thermal adaptation in biological membranes: is homeoviscous adaptation the explanation? Annu. Rev. Physiol. 57, 19–42.
Hoffmann, A. A., Anderson, A., and Hallas, R. (2002). Opposing clines for high and low temperature resistance in Drosophila melanogaster. Ecol. Lett. 5, 614–618.
Hoffmann, A. A., and Parsons, P. A. (1989). An integrated approach to environmental stress tolerance and life-history variation: desiccation tolerance in Drosophila. Biol. J. Linn. Soc. 37, 117–136.
Huang, D. W., Sherman, B. T., and Lempicki, R. A. (2009). Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 4, 44–57.
IPCC (2014). “Summary for Policymakers,” In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Eds C.B. Field et al. (Cambridge, NY: Cambridge University Press), 1–32.
Jollivet, D., Mary, J., Gagniere, N., Tanguy, A., Fontanillas, E., Boutet, I., et al. (2012). Proteome adaptation to high temperatures in the ectothermic hydrothermal vent Pompeii worm. PLoS One 7:e31150. doi: 10.1371/journal.pone.0031150
Käll, L., Canterbury, J. D., Weston, J., Noble, W. S., and MacCoss, M. J. (2007). Semi-supervised learning for peptide identification from shotgun proteomics datasets. Nat. Methods 4, 923–925.
Kellermann, V., Overgaard, J., Hoffmann, A. A., Fløjgaard, C., Svenning, J. C., and Loeschcke, V. (2012). Upper thermal limits of Drosophila are linked to species distributions and strongly constrained phylogenetically. Proc. Natl. Acad. Sci. U. S. A 109, 16228–16233. doi: 10.1073/pnas.1207553109
Kellermann, V. M., Hoffmann, A. A., Kristensen, T. N., Moghadam, N. N., and Loeschcke, V. (2015). Experimental evolution under fluctuating thermal conditions does not reproduce patterns of adaptive clinal differentation in Drosophila melanogaster. Am. Nat. 186, 582–593.
Ketola, T., and Kristensen, T. N. (2017). Experimental approaches for testing if tolerance curves are useful for predicting fitness in fluctuating environments. Front. Ecol. Evol. 5:129. doi: 10.3389/fevo.2017.00129
Ketola, T., Laakso, J., Kaitala, V., and Airaksinen, S. (2004). Evolution of Hsp90 expression in Tetrahymena thermophila (Protozoa, Ciliata) populations exposed to thermally variable environments. Evolution 58, 741–748. doi: 10.1111/j.0014-3820.2004.tb00407.x
Kingsolver, J. G., Ragland, G. J., and Diamond, S. E. (2009). Evolution in a constant environment: Thermal fluctuations and thermal sensitivity of laboratory and field populations of Manduca sexta. Evolution 63, 537–541. doi: 10.1111/j.1558-5646.2008.00568.x
Köcher, T., Pichler, P., Schutzbier, M., Stingl, C., Kaul, A., Teucher, N., et al. (2009). High precision quantitative proteomics using iTRAQ on an LTQ Orbitrap: A new mass spectrometric method combining the benefits of all. J. Proteome Res. 8, 4743–4752. doi: 10.1021/pr900451u
Kofler, R., Pandey, R. V., and Schlotterer, C. (2011). PoPoolation2: identifying differentiation between populations using sequencing of pooled DNA samples (Pool-Seq). Bioinformatics 27, 3435–3436. doi: 10.1093/bioinformatics/btr589
Kontopoulos, D. G., van Sebille, E., Lange, M., Yvon-Durocher, G., Barraclough, T. G., and Pawar, S. (2020). Phytoplankton thermal responses adapt in the absence of hard thermodynamic constraints. Evolution 74, 775–790. doi: 10.1111/evo.13946
Kristensen, T. N., Hoffmann, A. A., Overgaard, J., Sørensen, J. G., Hallas, R., and Loeschcke, V. (2008). Costs and benefits of cold acclimation in field-released Drosophila. Proc. Natl. Acad. Sci. U. S. A. 105, 216–221.
Kristensen, T. N., Ketola, T., and Kronholm, I. (2018). Adaptation to environmental stress at different timescales. Ann. N. Y. Acad. Sci. 1476, 5–22. doi: 10.1111/nyas.13974
Lande, R. (2009). Adaptation to an extraordinary environment by evolution of phenotypic plasticity and genetic assimilation. J. Evol. Biol. 22, 1435–1446. doi: 10.1111/j.1420-9101.2009.01754.x
Latosinska, A., Vougas, K., Makridakis, M., Klein, J., Mullen, W., Abbas, M., et al. (2015). Comparative analysis of label-free and 8-Plex iTRAQ approach for quantitative tissue proteomic analysis. PLoS One 10:e0137048. doi: 10.1371/journal.pone.0137048
Lecheta, M. C., Awde, D. N., O’Leary, T. S., Unfried, L. N., Jacobs, N. A., Whitlock, M. H., et al. (2020). Integrating GWAS and transcriptomics to identify the molecular underpinnings of thermal stress responses in Drosophila melanogaster. Front. Genet. 11:658. doi: 10.3389/fgene.2020.00658
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., et al. (2009). The Sequence Alignment/Map format and SAMtools. Bioinformatics 25, 2078–2079. doi: 10.1093/bioinformatics/btp352
Logan, M. L., and Cox, C. L. (2020). Genetic Constraints, Transcriptome Plasticity, and the Evolutionary Response to Climate Change. Front. Genet. 11:538226. doi: 10.3389/fgene.2020.538226
Ma, G., Hoffmann, A. A., and Ma, C. S. (2015). Daily temperature extremes play an important role in predicting thermal effects. J. Exp. Biol. 218(Pt 14), 2289–2296. doi: 10.1242/jeb.122127
Maclean, H. J., Kristensen, T. N., Sørensen, J. G., and Overgaard, J. (2018). Laboratory maintenance does not alter ecological and physiological patterns among species: a Drosophila case study. J. Evol. Biol. 31, 530–542. doi: 10.1111/jeb.13241
Manenti, T., Loeschcke, V., Moghadam, N. N., and Sørensen, J. G. (2015). Phenotypic plasticity is not affected by experimental evolution in constant, predictable or unpredictable fluctuating thermal environments. J. Evol. Biol. 28, 2078–2087. doi: 10.1111/jeb.12735
Manenti, T., Loeschcke, V., and Sørensen, J. G. (2018). Constitutive up-regulation of Turandot genes rather than changes in acclimation ability is associated with the evolutionary adaptation to temperature fluctuations in Drosophila simulans. J. Insect. Physiol. 104, 40–47. doi: 10.1016/j.jinsphys.2017.11.008
Manenti, T., Sørensen, J. G., and Loeschcke, V. (2017). Environmental heterogeneity does not affect levels of phenotypic plasticity in natural populations of three Drosophila species. Ecol. Evol. 7, 2716–2724. doi: 10.1002/ece3.2904
Manenti, T., Sørensen, J. G., Moghadam, N. N., and Loeschcke, V. (2014). Predictability rather than amplitude of temperature fluctuations determines stress resistance in a natural population of Drosophila simulans. J. Evol. Biol. 27, 2113–2122. doi: 10.1111/jeb.12463
Merilä, J., and Hendry, A. P. (2014). Climate change, adaptation, and phenotypic plasticity: the problem and the evidence. Evol. Appl. 7, 1–14. doi: 10.1111/eva.12137
Overgaard, J., Kristensen, T. N., Mitchell, K. A., and Hoffmann, A. A. (2011). Thermal tolerance in widespread and tropical Drosophila species: Does phenotypic plasticity increase with latitude? Am. Nat. 178, S80–S96.
Perkins-Kirkpatrick, S. E., and Lewis, S. C. (2020). Increasing trends in regional heatwaves. Nat. Commun. 11:3357. doi: 10.1038/s41467-020-16970-7
Pigliucci, M. (1996). How organisms respond to environmental changes: From phenotypes to molecules (and vice versa). Trends Ecol. Evol. 11, 168–173. doi: 10.1016/0169-5347(96)10008-2
Pigliucci, M. (2001). Phenotypic Plasticity: Beyond Nature and Nurture. Baltimore: The John Hopkins University Press.
Pigliucci, M., Murren, C. J., and Schlichting, C. D. (2006). Phenotypic plasticity and evolution by genetic assimilation. J. Exp. Biol. 209, 2362–2367. doi: 10.1242/jeb.02070
Pottiez, G., Wiederin, J., Fox, H. S., and Ciborowski, P. (2012). Comparison of 4-plex to 8-plex iTRAQ quantitative measurements of proteins in human plasma samples. J. Proteome Res. 11, 3774–3781. doi: 10.1021/pr300414z
R Core Team (2019). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Radchuk, V., Reed, T., Teplitsky, C., van de Pol, M., Charmantier, A., Hassall, C., et al. (2019). Adaptive responses of animals to climate change are most likely insufficient. Nat. Commun. 10:3109. doi: 10.1038/s41467-019-10924-4
Schou, M. F., Kristensen, T. N., Kellermann, V., Schlötterer, C., and Loeschcke, V. (2014). A Drosophila laboratory evolution experiment points to low evolutionary potential under increased temperatures likely to be experienced in the future. J. Evol. Biol. 27, 1859–1868. doi: 10.1111/jeb.12436
Schreiber, S. J. (2010). Interactive effects of temporal correlations, spatial heterogeneity and dispersal on population persistence. Proc. Biol. Sci. 277, 1907–1914. doi: 10.1098/rspb.2009.2006
Sgrò, C., Terblanche, J. S., and Hoffmann, A. A. (2016). What can plasticity contribute to insect responses to climate change? Annu. Rev. Entomol. 61, 433–451.
Simões, P., Santos, M. A., Carromeu-Santos, A., Quina, A. S., Santos, M., and Matos, M. (2020). Beneficial developmental acclimation in reproductive performance under cold but not heat stress. J. Therm. Biol. 90:102580. doi: 10.1016/j.jtherbio.2020.102580
Sørensen, J. G., Kristensen, T. N., and Loeschcke, V. (2003). The evolutionary and ecological role of heat shock proteins. Ecol. Lett. 6, 1025–1037.
Sørensen, J. G., Kristensen, T. N., and Overgaard, J. (2016a). Evolutionary and ecological patterns of thermal acclimation capacity in Drosophila: is it important for keeping up with climate change? Curr. Opin. Insect Sci. 17, 98–104. doi: 10.1016/j.cois.2016.08.003
Sørensen, J. G., Schou, M. F., Kristensen, T. N., and Loeschcke, V. (2016b). Thermal fluctuations affect the transcriptome through mechanisms independent of average temperature. Sci. Rep. 6:30975. doi: 10.1038/srep30975
Sørensen, J. G., Nielsen, M. M., Kruhoffer, M., Justesen, J., and Loeschcke, V. (2005). Full genome gene expression analysis of the heat stress response, in Drosophila melanogaster. Cell Stress Chaperones 10, 312–328.
Sørensen, J. G., Schou, M. F., and Loeschcke, V. (2017). Evolutionary adaptation to environmental stressors: a common response at the proteomic level. Evolution 71, 1627–1642.
Spivak, M., Weston, J., Bottou, L., Käll, L., and Noble, W. S. (2009). Improvements to the percolator algorithm for Peptide identification from shotgun proteomics data sets. J. Proteome Res. 8, 3737–3745. doi: 10.1021/pr801109k
Terblanche, J. S., Nyamukondiwa, C., and Kleynhans, E. (2010). Thermal variability alters climatic stress resistance and plastic responses in a globally invasive pest, the Mediterranean fruit fly (Ceratitis capitata). Entomol. Exp. Appl. 137, 304–315.
van Heerwaarden, B., Lee, R. F. H., Overgaard, J., and Sgro, C. M. (2014). No patterns in thermal plasticity along a latitudinal gradient in Drosophila simulans from eastern Australia. J. Evol. Biol. 27, 2541–2553. doi: 10.1111/jeb.12510
Williams, B. R., van Heerwaarden, B., Dowling, D. K., and Sgro, C. M. (2012). A multivariate test of evolutionary constraints for thermal tolerance in Drosophila melanogaster. J. Evol. Biol. 25, 1415–1426. doi: 10.1111/j.1420-9101.2012.02536.x
Keywords: heat tolerance, genomics, proteomics, thermal fluctuations, Drosophila simulans
Citation: Sørensen JG, Manenti T, Bechsgaard JS, Schou MF, Kristensen TN and Loeschcke V (2020) Pronounced Plastic and Evolutionary Responses to Unpredictable Thermal Fluctuations in Drosophila simulans. Front. Genet. 11:555843. doi: 10.3389/fgene.2020.555843
Received: 04 May 2020; Accepted: 06 October 2020;
Published: 28 October 2020.
Edited by:
David Vieites, Consejo Superior de Investigaciones Científicas (CSIC), SpainReviewed by:
C.-Elisa Schaum, University of Hamburg, GermanyCopyright © 2020 Sørensen, Manenti, Bechsgaard, Schou, Kristensen and Loeschcke. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jesper G. Sørensen, amVzcGVyLnNvZXJlbnNlbkBiaW8uYXUuZGs=
†Present address: Tommaso Manenti, Laboratori Biokyma Srl, Anghiari, Italy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.