
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 27 February 2020
Sec. Livestock Genomics
Volume 10 - 2019 | https://doi.org/10.3389/fgene.2019.01406
This article is part of the Research Topic Genetic Dissection of Important Traits in Aquaculture: Genome-scale Tools Development, Trait Localization and Regulatory Mechanism Exploration View all 38 articles
 Eva Vallejos-Vidal1
Eva Vallejos-Vidal1 Sebastián Reyes-Cerpa2,3
Sebastián Reyes-Cerpa2,3 Jaime Andrés Rivas-Pardo2,3
Jaime Andrés Rivas-Pardo2,3 Kevin Maisey4
Kevin Maisey4 José M. Yáñez5
José M. Yáñez5 Hector Valenzuela4Pablo A. Cea6Victor Castro-Fernandez6
Hector Valenzuela4Pablo A. Cea6Victor Castro-Fernandez6 Lluis Tort1
Lluis Tort1 Ana M. Sandino4
Ana M. Sandino4 Mónica Imarai4*
Mónica Imarai4* Felipe E. Reyes-López1*
Felipe E. Reyes-López1*Single-nucleotide polymorphisms (SNPs) are single genetic code variations considered one of the most common forms of nucleotide modifications. Such SNPs can be located in genes associated to immune response and, therefore, they may have direct implications over the phenotype of susceptibility to infections affecting the productive sector. In this study, a set of immune-related genes (cc motif chemokine 19 precursor [ccl19], integrin β2 (itβ2, also named cd18), glutathione transferase omega-1 [gsto-1], heat shock 70 KDa protein [hsp70], major histocompatibility complex class I [mhc-I]) were analyzed to identify SNPs by data mining. These genes were chosen based on their previously reported expression on infectious pancreatic necrosis virus (IPNV)-infected Atlantic salmon phenotype. The available EST sequences for these genes were obtained from the Unigene database. Twenty-eight SNPs were found in the genes evaluated and identified most of them as transition base changes. The effect of the SNPs located on the 5’-untranslated region (UTR) or 3’-UTR upon transcription factor binding sites and alternative splicing regulatory motifs was assessed and ranked with a low-medium predicted FASTSNP score risk. Synonymous SNPs were found on itβ2 (c.2275G > A), gsto-1 (c.558G > A), and hsp70 (c.1950C > T) with low FASTSNP predicted score risk. The difference in the relative synonymous codon usage (RSCU) value between the variant codons and the wild-type codon (ΔRSCU) showed one negative (hsp70 c.1950C > T) and two positive ΔRSCU values (itβ2 c.2275G > A; gsto-1 c.558G > A), suggesting that these synonymous SNPs (sSNPs) may be associated to differences in the local rate of elongation. Nonsynonymous SNPs (nsSNPs) in the gsto-1 translatable gene region were ranked, using SIFT and POLYPHEN web-tools, with the second highest (c.205A > G; c484T > C) and the highest (c.499T > C; c.769A > C) predicted score risk possible. Using homology modeling to predict the effect of these nonsynonymous SNPs, the most relevant nucleotide changes for gsto-1 were observed for the nsSNPs c.205A > G, c484T > C, and c.769A > C. Molecular dynamics was assessed to analyze if these GSTO-1 variants have significant differences in their conformational dynamics, suggesting these SNPs could have allosteric effects modulating its catalysis. Altogether, these results suggest that candidate SNPs identified may play a crucial potential role in the immune response of Atlantic salmon.
Genetic variation occurs within and among populations, leading to polymorphisms that could be associated with genetic trait or also a phenotype in the presence of an environmental stimulus (Brookes, 1999; Rebbeck et al., 2004; Hirschhorn and Daly, 2005). A single-nucleotide polymorphism (SNP) is a single genetic code variation (i.e., polymorphic). Although multiallelic SNPs do exist, the SNPs are usually biallelic (two alternative bases occur) and require a minimum frequency (>1%) in the population (Wang et al., 1998). The SNPs are the most common form of variation in the genome and they are extensively used to study genetic differences between individuals and populations. These SNPs may contribute to changes in the genomic sequence, either in the coding (exons), intergenic, or noncoding (introns) region (Dijk et al., 2014; Ahmad et al., 2018).
SNPs are considered the most useful biomarkers for disease diagnosis or prognosis due to their common frequency, ease of analysis, low genotyping costs, and the possibility to carry out association studies based on statistical and bioinformatics tools (Srinivasan et al., 2016). Thus, SNPs have gained importance as major drivers in disease-association studies in the recent era. In mammals, on the past decade it has been seen an enormous progress in identifying hundreds of thousands SNPs to identify associations with complex clinical conditions and phenotypic traits associated with hundreds of common diseases (Welter et al., 2014; Wijmenga and Zhernakova, 2018).
Furthermore, SNPs may also have a great influence on the immune response towards pathogenic challenges and diseases outcome, contributing in a range of susceptibility to infections among the individuals. Thus, the SNP may have a protective role, may influence the rate of diseases progression or even the type of cellular immune response evoked by pathogens (Hill, 2001; Skevaki et al., 2015). In this regard, polymorphisms on several immune-related genes have been associated with susceptibility to infections including pattern recognition receptors (prr) and downstream signaling molecules (Skevaki et al., 2015), mannose- binding lectin 2 (mbl2) and toll-interleukin 1 receptor domain containing adaptor protein (tirap) (Gowin et al., 2018), c-c chemokine receptor type 5 (ccr5) (Martin et al., 1998; Salkowitz et al., 2003; Fellay et al., 2009; Chapman and Hill, 2012), interleukin 6 (il-6) (Zhang et al., 2012), and il-22 (Zhang et al., 2011), among others.
In species related to aquaculture, SNPs are especially important because they may be associated with different phenotypic traits with economical implications. Therefore, this increase of information has a direct impact on the accuracy of selection for these traits, improving the rate of genetic gain and production efficiency (Hayes et al., 2007b). The ubiquity of SNPs across the genomes examined to date, has allowed their use as markers for a wide range of applications including quantitative trait locus (QTL) mapping, pedigree analysis, association studies and population genetics, among others. Conversely, whereas the effect on gene mutations in mammals has been well documented, such information in teleost species is still limited. However, several efforts have been made to provide information regarding the consequences of genetic alterations in immune response-related genes and may influence susceptibility to diseases in fish (Kongchum et al., 2011). In this context, SNP variations were found on il-1β of Cyprinus pellegrini and C. carpio that can be helpful in understanding differential resistance to koi herpesvirus (KHV) and Aeromonas hydrophila, respectively (Jia et al., 2015; Wenne, 2018). On the other hand, three SNPs were identified in the leukocyte cell-derived chemotaxin-2 (lect2) gene to be associated with resistance to the big belly disease on Latis calcarifer (Fu et al., 2014a; Wenne, 2018). Three SNPs in the mast cell protease 8 (mcp-8) gene were also significantly associated with resistance of tilapia to Streptococcus agalactiae (Fu et al., 2014b; Wenne, 2018).
In Atlantic salmon, some studies have reported SNPs associated with a resistance genotype against infectious pancreatic necrosis (IPN) virus (IPNV). IPN is a highly transmissible disease with worldwide distribution that occurs both at the initial stage of rearing in freshwater and in post-smolts in seawater (Bruno, 2004). Importantly, asymptomatic carriers (McAllister et al., 1993), establishment of viral persistence (Reyes-Cerpa et al., 2014), IPN-resistance phenotype (Reyes-López et al., 2015), and vertical transmission through eggs (Bootland et al., 1991) have been reported. In this matter, several segregating SNP markers linked to a major QTL associated with resistance against infectious pancreatic necrosis virus (IPNV) in Atlantic salmon from an Scotland commercial breeding program have been reported (Houston et al., 2012; Wenne, 2018). Similarly, a QTL in Norwegian salmon has been employed in marker-assisted selection in breeding companies from Norway and Scotland, which resulted in 75% reduction in the number of IPN-outbreaks in the salmon farming industry. This QTL has been located on the SNP-based linkage map and identified as the epithelial cadherin (cdh1-1) gene with a functional involvement in viral attachment and entry of IPNV (Moen et al., 2009; Moen et al., 2015; Wenne, 2018). Based on these reports, it seems that the strategy to detect SNPs in immune-related genes could provide a set of candidate polymorphisms that could explain the correlation between the pathogen and the disease phenotype. Particularly, the relevance to identify SNPs in the coding sequence of immunity-related genes could explain directly (causal) the variability upon a specific phenotype evaluated (Carlson et al., 2003; Cheng et al., 2004; Hirschhorn and Daly, 2005). In this context, most of the knowledge about fish immune response is based on large-scale expressed sequence tag (EST) sequencing that has helped to identify immune-related genes in teleosts. Undoubtedly, the EST sequencing based on tissues that play a central role on immune response contribute to detect gene sequences that are directly related to host defense functions. The identification of a set of splenic leukocytes immune-related genes from Atlantic salmon IPNV-infected using EST analysis has been reported (Cepeda et al., 2011; Cepeda et al., 2012). Importantly, some of these genes were detected differentially expressed when the IPN-susceptible and IPN-resistant phenotypes were compared (Reyes-López et al., 2015). Thus, the search and identification of SNPs on these immune-related genes using a data mining strategy may contribute to provide a set of candidate polymorphisms that could help in the progress to establish a link between the possible causes of this differential expression pattern and the IPN-phenotype variability. Hayes et al. (Hayes et al., 2007b), described for first time an in silico detection of 2,507 putative SNPs in Atlantic salmon from the alignment of 100,866 EST. Despite the large number of SNPs identified, there is no gene directly associated with immune function.
Therefore, the aim of this study was the identification of SNPs (in silico) by data mining upon a set of immune-related genes whose expression has been previously reported in response to the infection with IPNV in Atlantic salmon. For this purpose, a set of immune-related genes (cc motif chemokine 19 precursor [ccl19], integrin β2 (itβ2, also named cd18), glutathione transferase omega-1 [gsto-1], heat shock 70 KDa protein [hsp70], major histocompatibility complex class I [mhc-I]) were selected as target for the in silico SNP search and identification using as template the EST sequences for these genes obtained from the Unigene database. Based on their nucleotide sequence, the SNPs were located in the 5’/3’-UTR or in the translated region. In the case of those nucleotide variations located in the translated region, the SNP were classified as synonymous (sSNPs) or nonsynonymous (nsSNPs) based on the change provoked in the predicted amino acid sequence. While for sSNPs a codon usage analysis was conducted, for those nsSNPs a homology modeling analysis was carried out in order to evaluate whether they could have an effect on the predicted tridimensional protein structure. In addition, on those nsSNP in which a significant change in the three-dimensional protein structure was observed by homology modeling, an analysis of such variants over the time was performed by molecular dynamics (MD) simulation. This study provide a set of identified candidate SNPs that may help to determine potential correlations between the immunity gene expression pattern in Atlantic salmon and their response against the pathogens they are exposed under aquaculture conditions.
The immune-related genes analyzed (cc motif chemokine 19 precursor [ccl19], integrin β2 [itβ2], glutathione transferase omega-1 [gsto-1], heat shock 70 KDa protein [hsp70], major histocompatibility complex class I [mhc-I]) were selected based on their previously reported expression on splenic leukocytes isolated from IPNV-infected Atlantic salmon and whose expression was also differentially modulated between the IPN-susceptible and IPN-resistant phenotypes (Reyes-López et al., 2015). The EST sequences for the previously above-mentioned selected candidate genes were downloaded from the Unigene database (NCBI). The detail regarding all the EST sequences analyzed in this study to identify SNPs are indicated on Supplementary Tables 1–5.
The identification of SNPs by data mining was carried out in order to identify possible functional effects in the sequences related to defense and immune response in Atlantic salmon challenged with IPNV. All the sequences including in the analysis were first preanalyzed in order to remove any vector sequences or repetitive elements using Cross-Match (Ewing and Green, 1998) and RepeatMasker (Tarailo-Graovac and Chen, 2009), respectively. The search for SNPs was carried out based on multiple sequence alignment including the total number of sequences collected for each evaluated gene using the HaploSNPer web-based tool (Tang et al., 2008). From the alignment analysis, each nucleotide variant detected was considered as putative SNP and it was defined according to the nucleotide position in the gene sequence. The sequences were then filtered to exclude possible sequencing errors and noninformative polymorphisms (variants with a frequency lower than 1%). The nonfiltered SNPs were chosen as the most probable or reliable SNPs. The nucleotide variations were described according to the nomenclature suggested by Dunnen and Antonarakis (Den Dunnen and Antonarakis, 2001).
In order to determine the SNP location onto the gene region (5’-UTR, coding region, 3’-UTR), the nucleotide sequence was obtained based on their Unigene annotation from Nucleotide database (NCBI) and compared them by alignment with BioEdit sequence alignment editor (version 7.0.5.3). In addition, the nucleotide sequence was also used as template to get the predicted amino acid sequence using ORF Finder tool (NCBI) to confirm by protein BLAST (NCBI) the annotation for the unigene sequence analyzed.
The functional impact of the SNP was assessed depending on the gene region (5’-UTR, coding region, 3’-UTR) on which the nucleotide variant was located. For those SNP located in the noncoding region (5’-UTR, 3’-UTR) the predictive nucleotide variant effect upon motifs associated to transcription factor binding sites was evaluated with TFSearch webtool (http://diyhpl.us/~bryan/irc/protocol-online/protocol-cache/TFSEARCH.html) (Heinemeyer et al., 1998). Furthermore, the predictive SNP effect onto possible exon splicing enhancer [ESEfinder (http://krainer01.cshl.edu/cgi-bin/tools/ESE3/esefinder.cgi?process=home) (Cartegni et al., 2003); RESCUE-ESE (http://hollywood.mit.edu/burgelab/rescue-ese/) (Fairbrother et al., 2004)] and exon splicing silencer [FASS-ESS (http://genes.mit.edu/fas-ess/) (Wang et al., 2004)] was also evaluated. Based on these results, the SNP functional effect was predicted according to FASTSNP in order to assign a FASTSNP score (Yuan et al., 2006). A FASTSNP score between 0 and 5 was assigned to each individually SNP evaluated (representing from 0 to 5 the minimum to maximum functional SNP effect, respectively). On the other hand, for those SNP located in the coding region the nucleotide variant was first individually evaluated with ORF Finder tool (NCBI) in order to compare the predicted unigene amino acid sequence with the SNP-containing unigene sequence by BioEdit sequence alignment editor (version 7.0.5.3). In the case of the nucleotide variations located in untranslated region (5’-UTR; 3’-UTR) and those did not provoke any change in the predicted amino acid sequence (sSNP; synonymous amino acid change), the SNP effect was evaluated with the FASTSNP decision tree in order to assign a FASTSNP score (Yuan et al., 2006). In addition, on each sSNP identified a codon usage analysis was performed according to Sharp et al. (M.Sharp and Li, 1987). The difference in the relative synonymous codon usage (RSCU) value between the variant codons and the wild-type codon (ΔRSCU = RSCU mutant - RSCU wild-type) was calculated for all the SNPs identified based on the codon usage database (http://www.kazusa.or.jp/codon) for Salmo salar. On the other hand, in those nucleotide variants in which a change in the amino acid predicted sequence was detected (nsSNP; nonsynonymous amino acid change), the nucleotide variation impact was analyzed combining the score obtained from Sorting Intolerant From Tolerant (SIFT) (applied to human variant databases) (http://sift.bii.a-star.edu.sg/) (Ng and Henikoff, 2003) and POLYPHEN web-based resources (Flanagan et al., 2010). The results were ranked according to the protocol established by Bhatti et al. (Bhatti et al., 2006) with some modifications in order to homogenize the significance of all scores obtained for the SNP obtained in this study (lower score = minimum effect; higher score = maximum effect). Thus, the scores obtained for SIFT were ranked from I (tolerated) to IV (intolerant); meanwhile the scores obtained for POLYPHEN were ranked from A (benign) to E (probably damaging). The combination of the SIFT and POLYPHEN analysis give a score whose value was ranked from 1 (minimum effect) to 4 (maximum effect). The implications of the nonsynonymous SNPs reported at protein tertiary structure level was also evaluated in silico through homology modeling analysis.
In order to evaluate whether the nonsynonymous SNPs reported in this study have an effect on the predicted tridimensional protein structure, a homology modeling analysis was carried out. For this, the nucleotide sequence was used as target to obtain the predicting tridimensional protein structure with the CPHModels-3.0 webserver (Nielsen et al., 2010). The higher bit score alignment obtained was chosen as template for GSTO-1 (PDB ID: 1EEM; score = 278; E-value = 4e-75). Alternatively, the predicted protein structure for CCL19 (PDB ID: 2HCI; score = 55; E-value = 2e-08), ITB2 (PDB ID: 2KCN; score = 42; E-value = 2e-04), HSP70 (PDB ID: 1YUW; score = 1036; E-value = 0) and MHC class I (PDB ID: 1KTL; score = 182; E-value = 4e-46) were also obtained. The tridimensional protein structure for each PDB ID match was obtained to then be used as template for the homology modeling of the above mentioned gene sequences using the MODELLER webtool (Swiss-Model) (Sali and Blundell, 1993). The stereochemical quality of the modeled tridimensional protein structure was evaluated with Procheck webtool (Swiss-Model) (Laskowski et al., 1996).
The protonation state of residues at physiological conditions (pH = 7) was assigned using the Propka software included in the Maestro Suite (Olsson et al., 2011). The models were solvated in TIP3P truncated octahedron with an extension of 12 Å over the protein surface, including 4 Cl- atoms to maintain the net charge neutrality within the system. Parameters for calculations were derived from the AMBER ff14SB forcefield (Maier et al., 2015). Energy minimization was carried out in four different stages, in each one of them, 5,000 steps of steepest descent followed by 5,000 steps of a conjugated gradient were performed. In the first stage, the solute was fixed with a positional restraint of 500 kcal/mol Å2, and only the accommodation of the solvent was allowed. Then, the hydrogen atoms of the protein were freed from the restraint to allow their relaxation. In the third stage, the minimization was done imposing a lighter restriction of 10 kcal/mol Å2 on the heavy atoms of the protein. Lastly, a minimization without restraint was conducted. After that, the systems were equilibrated under NVT conditions, heating up from 0 K to 298.15 K in a window of 200 ps and maintaining the final temperature for 100 ps, using the Langevin thermostat with a collision frequency of 2 ps-1. Then, the systems were equilibrated for 10 ns under NPT conditions at 298.15 K and 1 atmosphere using the weak-coupling Berendsen barostat. Three production runs of 100 ns under the NPT conditions with random velocities seeds were performed for each protein system. All the MD calculations were performed with periodic boundary conditions with a time step of 2 fs, a 10 Å direct space cutoff for PME and constraining hydrogen atoms with SHAKE algorithm. All the simulations were performed using Amber18 with GPU acceleration (Salomon-Ferrer et al., 2013) and the trajectories were analyzed using cpptraj (Roe and Cheatham, 2013).
A set of immune-related genes (gsto-1, ccl19, itβ2, hsp70, mhc-I) were chosen based on their previous reported role on the immune response in Atlantic salmon (Cepeda et al., 2011; Cepeda et al., 2012; Reyes-López et al., 2015). The identification of SNPs was carried out upon these genes based on data mining analysis. A total of 310 EST sequences obtained from the Unigene database (NCBI) were analyzed to identify SNPs on ccl19, itβ2, gsto-1, hsp70, and mhc-I. Twenty-eight SNPs were found, broken down into 18 transitions (7 A > G; 11 C > T) and 10 transversions (7 A > C; 1 A > T; 1 C > G; 1 G > T) as the most probable or reliable nucleotide variants (Table 1). From them, 3 SNPs were found for ccl19, 2 SNPs for itβ2, 15 SNPs for gsto-1, 4 SNPs for hsp70, and 4 SNPs for mhc-I (Table 1).
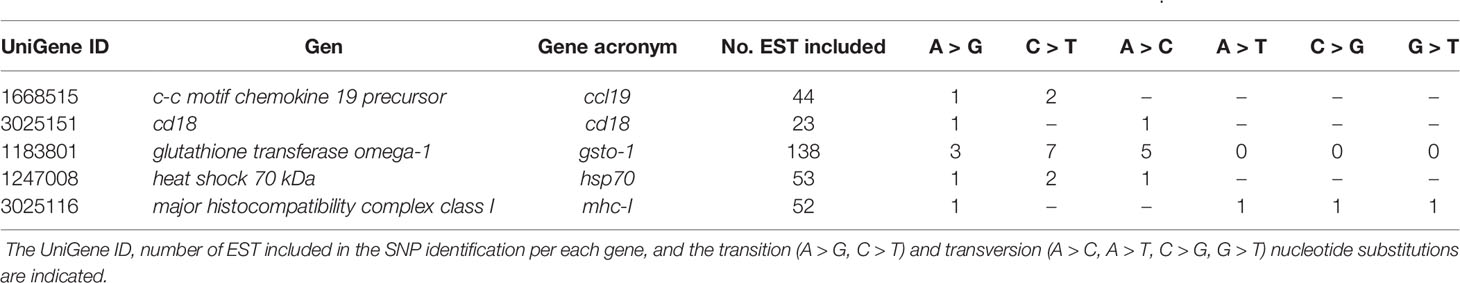
Table 1 Summary of single-nucleotide polymorphisms (SNPs) identified in a set of immune-related genes expressed in Salmo salar.
From the nucleotide variations identified, the 21.43% of the SNPs were found in the 5’-UTR region and 53.57% in the 3’-UTR region. At untranslated region level, mhc-I showed only nucleotide modifications at 5’-UTR. In the case of ccl19, itβ2, and hsp70 genes, the modifications were registered on the 3’-UTR (Table 2). Only in the case of gsto-1 the SNPs were found in both 5’- and 3’-UTR regions. To evaluate the effect of these SNPs located in the UTR regions, the FASTSNP decision tree was used in order to assign a FASTSNP score risk for each of the nucleotide variations found. The results showed that some SNPs on 5’-UTR (gsto-1: c.48C > T, c.89A > G; mhc-I: c.17G > T) and 3’-UTR (ccl19: c.933T > C, c.996C > T; gsto-1: c.1068A > C, c.1177A > C; hsp70: c.2102C > T, c.2139A > G) obtained the minimum FASTSNP score (FASTSNP score = 0), indicating that these nucleotide variations did not have any predicted consequence. However, the majority of the SNPs detected on these regions had a FASTSNP score of 1–3, which indicates that the SNP generates a low to medium impact (Table 2). Based on this SNP functional score effect, the results suggest that the nucleotide variations evaluated could be involved in the regulation of the immune-related genes evaluated.
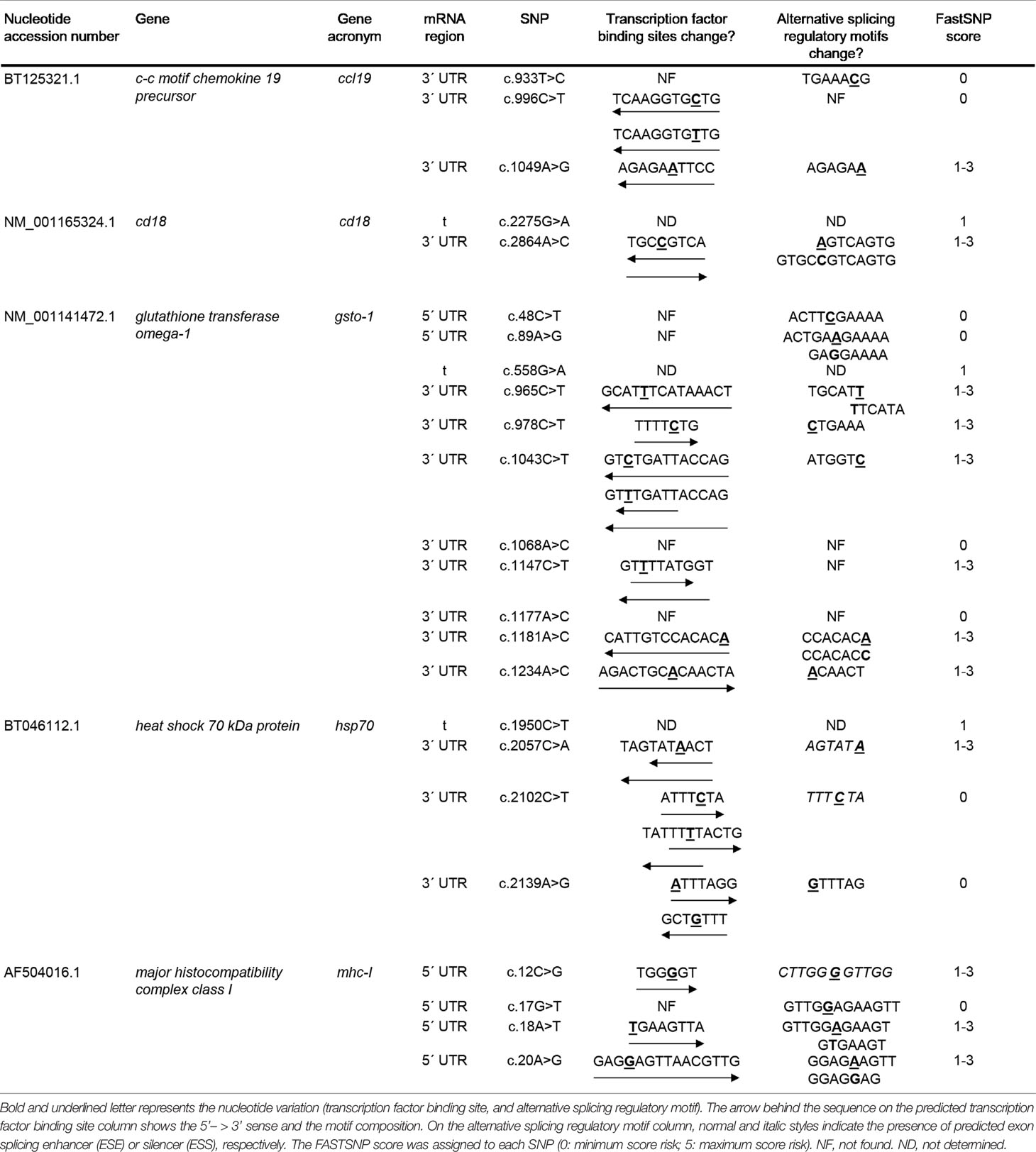
Table 2 Evaluation of nucleotide variations located in untranslated region (5’-UTR; 3’-UTR) and synonymous single-nucleotide polymorphisms (SNPs) in the translated (t) region on immune-related genes expressed in Salmo salar.
From the total number of SNPs found, the 25% of the nucleotide substitutions were located in the coding region. At functional level, three of them (10.71%) were identified as synonymous SNP since no variation in the predicted amino acid sequence for itβ2 (c.2275G > A), gsto-1 (c.558G > A), and hsp70 (c.1950C > T) was determined. Thus, a low-risk ranking was assigned (FASTSNP score = 1) (Table 2). In terms of codon usage, a positive RSCU value was observed for itβ2 (c.2275G > A) and gsto-1 (c.558G > A) (Table 3). By contrast, hsp70 (c.1950C > T) showed a negative RSCU value. This antecedents indicate that these sSNPs may have an impact in the local rate of translation elongation. On the other hand, the remaining 14.29% from total nucleotide variations were identified in the coding region as nonsynonymous SNPs. Importantly, all these nucleotide variations were only found into the gsto-1 sequence (Table 4). Two of these variations (c.205A > G, c.484T > C) obtained the highest risk score (SIFT+POLYPHEN score = 5) for nonsynonymous SNPs and whose nucleotide variations represented the modifications in the predicted amino acid sequence of serine (polar, uncharged R group) by glycine (nonpolar, flexible and the smallest R group; S26G) and by proline (nonpolar, cyclic, and rigid R group; S119P), respectively. The other two nucleotide substitutions (c.499T > C, c.769A > C) were ranked in the next risk score level (SIFT+POLYPHEN score = 4), representing in the predicted amino acid sequence a change of tyrosine (polar and aromatic R group) by histidine (polar and sometimes positively charged imidazole R group; Y124H), and threonine (polar, uncharged R group) by proline (T214P), respectively (Table 4). Taken together, these results indicate that the SNPs found in the gsto-1 could have a relevant impact at functional protein structural level.

Table 3 Evaluation of relative synonymous codon usage (RSCU) in immune-relevant genes in Salmo salar.
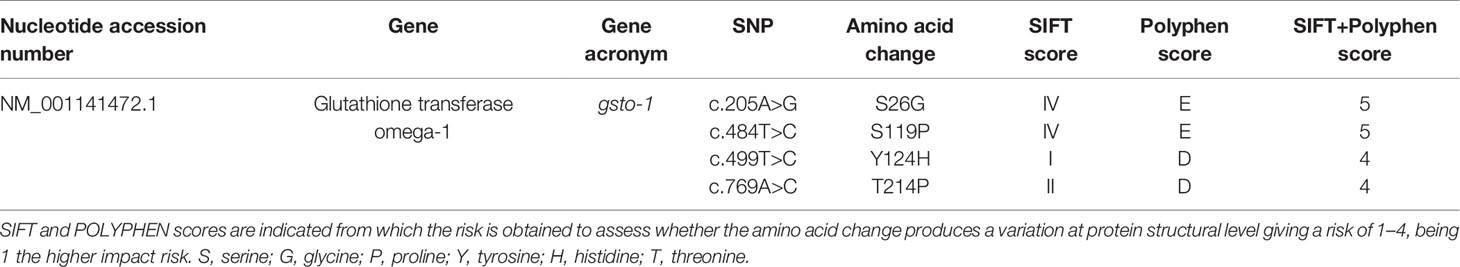
Table 4 Effect of nonsynonymous single-nucleotide polymorphisms (SNPs) in glutathione transferase omega-1 coding region of Salmo salar.
In order to evaluate whether the predicted SIFT+POLYPHEN score on the nonsynonymous nucleotide variations for gsto-1 have an impact at the protein structure level, a homology modeling was performed. To do this, the predicted tridimensional protein structure was obtained, on which the effect of such nucleotide variations determined for gsto-1 were analyzed individually.
Using the CPHModels-3.0 webserver, the higher score alignment (score = 278; E-value = 4e-75) obtained for the S. salar gsto-1 predicted amino acid sequence was the Homo sapiens Glutathione-S-transferase omega 1 (GSTO-1; PDB ID: 1EEM) and thus chosen as the best template structure.
The overall comparison between the human GSTO-1 structure (Figure 1A) and the modeled Atlantic salmon GSTO-1 (Figure 1B) showed high similarity, evidencing that the salmon predicted modeled protein does not present serious problems with the structural restrictions dictated by the template. Only some local differences were found by comparing in detail the human and salmon protein model of GSTO-1 at the helixes α4a, α4b, α7, and α8 (Figures 1B, C). To further validate the model, a stereochemical evaluation was performed by generating a Ramachandran plot to assess the Phy and Psi dihedral angles distribution. The stereochemical quality of the modeled tridimensional protein structure showed that the amino acids of the predicted sGSTO-1 structure were found mainly within the most favored (89.4%) and the additional allowed (10.1%) energy regions, meanwhile only the 0.5% were at the disallowed regions (Figure 1D). This antecedent indicates the good quality of the predicted sGSTO-1 structure obtained.

Figure 1 Homology modeling for predicted S. salar GSTO-1 based on H. sapiens GSTO-1 structure. (A) Tridimensional H. sapiens GSTO-1 (hGSTO-1) structure. (B) Predicted tridimensional S. salar GSTO-1 (sGSTO-1) structure. The differences on the secondary structure in the sGSTO-1 compared to hGSTO-1 are shown (arrow lines). (C) hGSTO-1 and sGSTO-1 overlay. (D) Ramachandran plot for the predicted sGSTO-1 structure. The amino acid distribution into most favored regions (A,B,L), additional allowed regions (a,b,l,p), generously allowed regions (~a,~b,~l,~p), and disallowed regions (GLU83) is indicated. In the protein structures, α-helices (α1- α8), β-sheets (β1-4) are indicated. The reduced glutathione (GSH) molecule is represented.
The effect of the gsto-1 nonsynonymous nucleotide variations detected were individually evaluated by homology modeling of S. salar GSTO-1. The model of the variant c.205A > G (S26G on the predicted amino acid sequence; sGSTO-1 S26G) suggests that glycine would increase the conformational freedom on the β2 sheet making it shorter (Figures 2A–C). The stereochemical quality of the sGSTO-1 S26G protein structure kept the good quality of the model, with only a slight increase in the number of the amino acids in the most favored energy regions (90.3%) compared to the predicted sGSTO-1 structure (Figure 2D).
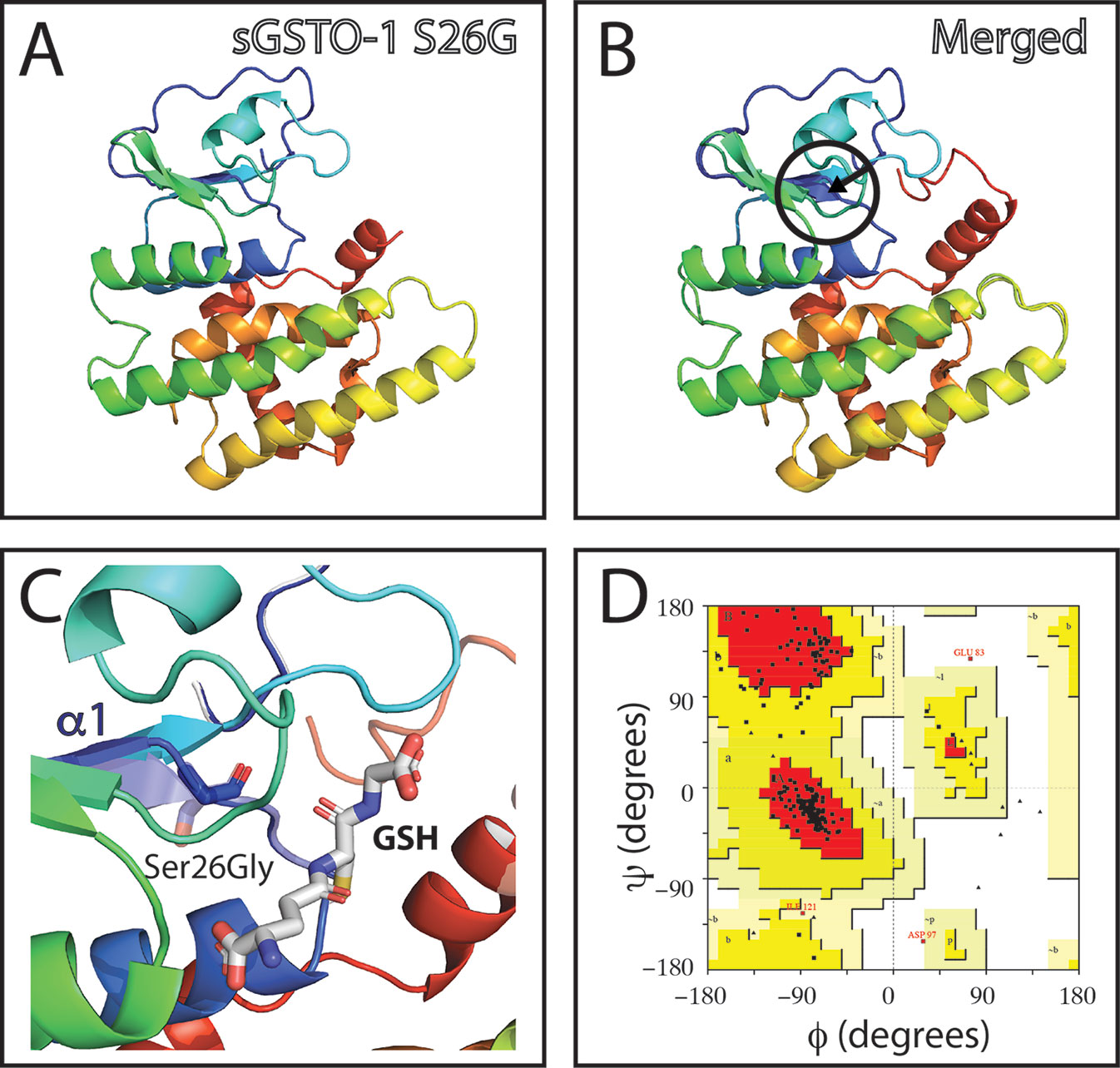
Figure 2 Evaluation of single-nucleotide polymorphism (SNP) c.205A > G effect on the predicted S. salar GSTO-1 (sGSTO-1) structure by homology modeling. (A) Predicted tridimensional sGSTO-1 including the S26G substitution (sGSTO-1 S26G). (B) sGSTO-1 and sGSTO-1 S26P overlay. The punctual amino acid substitution (dotted arrow line) and the region of the secondary structure affected (β2 sheet, circle) are indicated. (C) Enlarged view of the sGSTO-1 S26P substitution shown in (B). sGSTO-1 (transparent) and sGSTO-1 (colored) are shown. The reduced glutathione (GSH) molecule is represented. (D) Ramachandran plot for the predicted sGSTO-1 S26G structure. The amino acid distribution into most favored regions (A,B,L), additional allowed regions (a,b,l,p), generously allowed regions (~a,~b,~l,~p), and disallowed regions (GLU83, ASP97) is indicated.
The SNP c.484T> C (responsible of the modification S119P in the predicted amino acid sequence; sGSTO-1 S119P) is located in the α4 helix kink region (Figure 3). The model with the S119P substitution suggests that proline would makes the helix kink longer (Figures 3B, C). This modification may generate as consequence that the α4 helix are no longer composed by the α4a and α4b helix but rather independent two different new α-helices. Compared to the sGSTO-1 predicted structure, the sGSTO-1 S119P stereochemical quality showed a slight decrease in the number of the amino acids in the most favored energy regions (87.9%) and an increase on those located in the additional allowed regions (11.6%) compared to the predicted sGSTO-1 structure (Figure 3D).
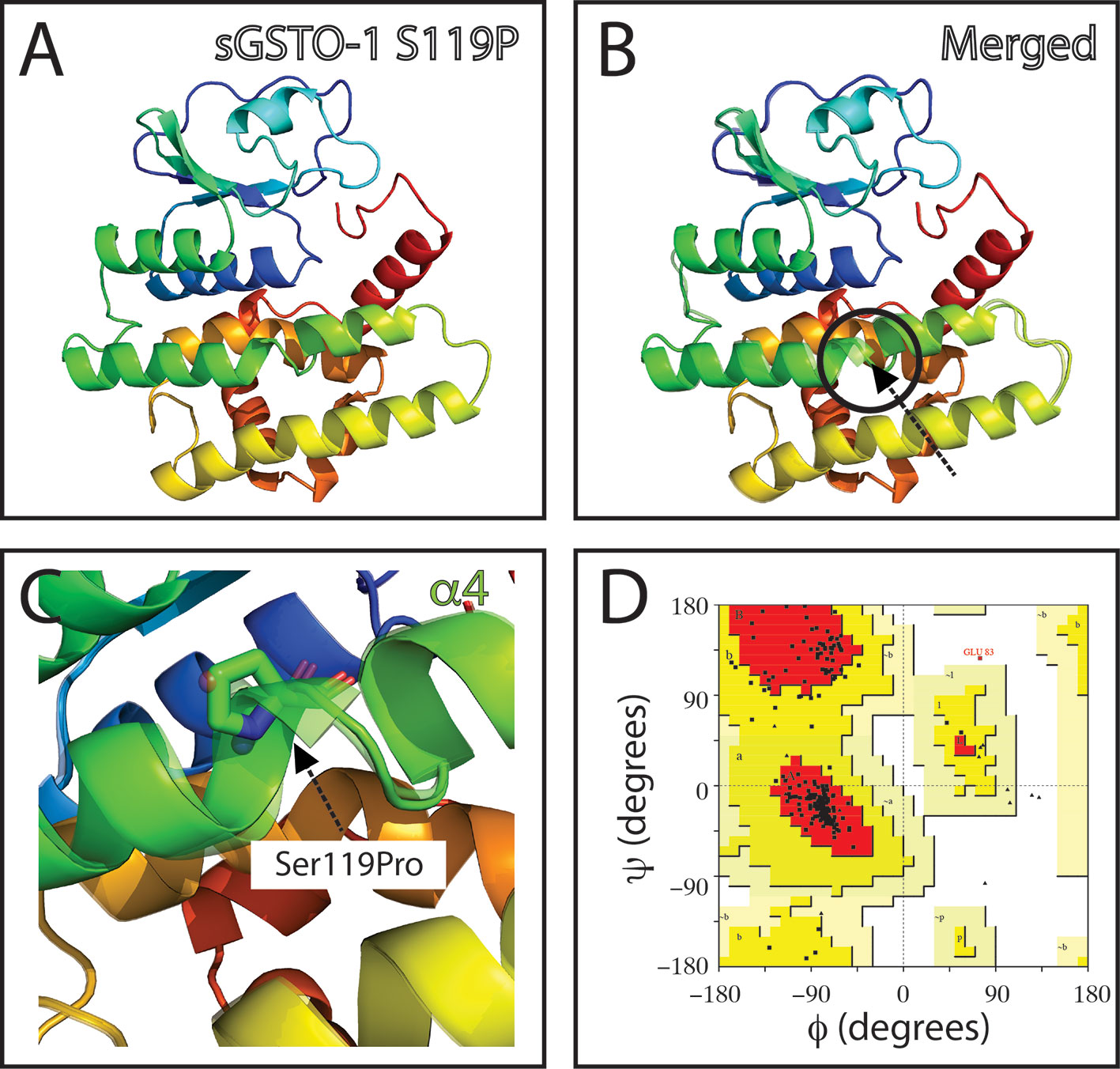
Figure 3 Evaluation of single-nucleotide polymorphism (SNP) c.484T > C effect on the predicted S. salar GSTO-1 (sGSTO-1) structure by homology modeling. (A) Predicted tridimensional sGSTO-1 including the S119P substitution (sGSTO-1 S119P). (B) sGSTO-1 and sGSTO-1 S119P overlay. The punctual amino acid substitution (dotted arrow line) and the structural region (α4 helix kink, circle) are indicated. (C) Enlarged view of the sGSTO-1 S119P substitution shown in (B). sGSTO-1 (transparent) and sGSTO-1 (colored) are shown. (D) Ramachandran plot for the predicted sGSTO-1 S119P structure. The amino acid distribution into most favored regions (A,B,L), additional allowed regions (a,b,l,p), generously allowed regions (~a,~b,~l,~p), and disallowed regions (GLU83) is indicated.
On the other hand, no appreciable variations in the predicted tridimensional S. salar GSTO-1 structure were observed for the SNP c499T > C (Y124H substitution; sGSTO-1 Y214H) (Figure 4). By contrast, in the case of the SNP c.769C > C (responsible of the modification T214P in the predicted amino acid sequence; sGSTO-1 T214P), a change in the tridimensional conformation affecting the C-terminus region is observed on the α7 helix of the model (Figure 5). Punctually, the amino acid substitution makes the α7 helix shorter, thus lengthening the α7 helix kink (Figures 5B, C). This region is the special relevance in the protein structure because the residues of the C-terminal end are involved in the formation of the H site, a binding site that accommodates a hydrophobic motif adjacent to the glutathione binding site (Board et al., 2000). The stereochemical quality of the sGSTO-1 T214P was similar to sGSTO-1 (89.9% of the amino acid in the most favored regions) (Figure 5D). Thus, the predicted salmon GSTO-1 sequence analysis suggests the high-impact of T214P on the protein functionality.
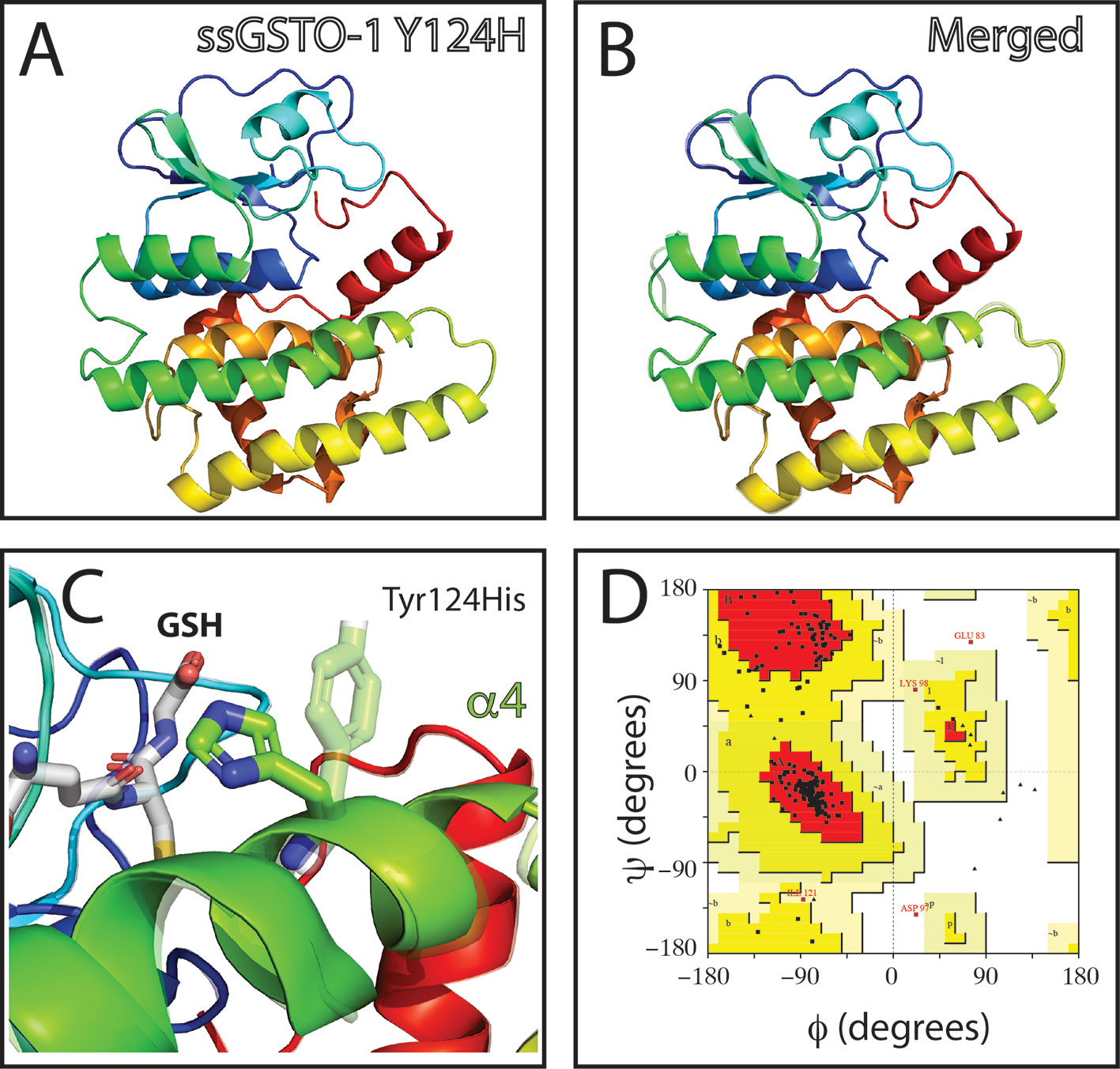
Figure 4 Evaluation of single-nucleotide polymorphism (SNP) c.499T > C effect on the predicted S. salar GSTO-1 (sGSTO-1) structure by homology modeling. (A) Predicted tridimensional sGSTO-1 including the Y124H substitution (sGSTO-1 Y124H). (B) sGSTO-1 and sGSTO-1 Y124H overlay. The punctual amino acid substitution (dotted arrow line) is indicated. (C) Enlarged view of the sGSTO-1 Y124H substitution shown in (B). sGSTO-1 (transparent), sGSTO-1 (colored), and the reduced glutathione (GSH) molecule are shown. (D) Ramachandran plot for the predicted sGSTO-1 Y124H structure. The amino acid distribution into most favored regions (A,B,L), additional allowed regions (a,b,l,p), generously allowed regions (~a,~b,~l,~p), and disallowed regions (GLU83, ASP97) is represented.
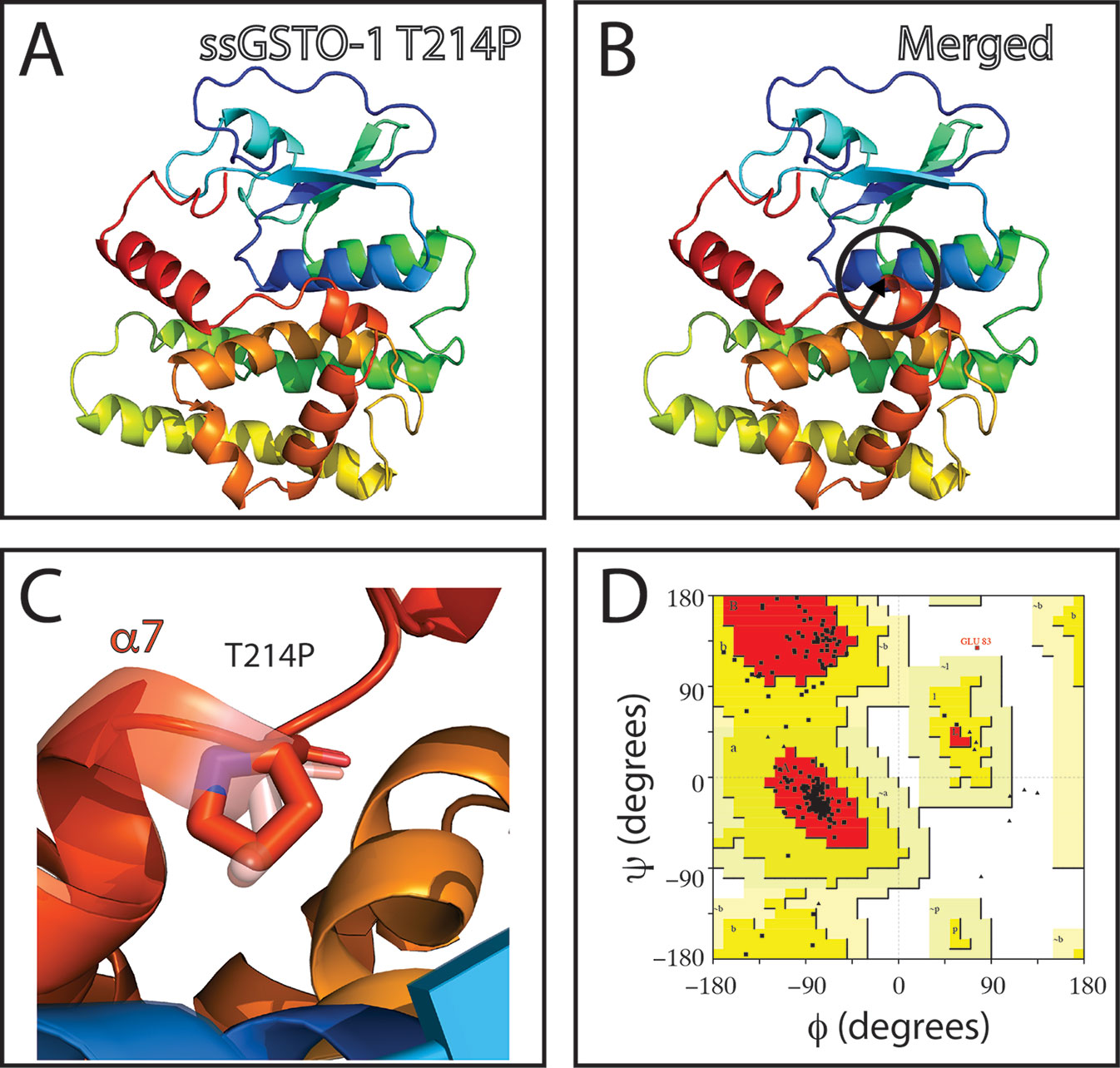
Figure 5 Evaluation of single-nucleotide polymorphism (SNP) c.769A > C effect on the predicted S. salar GSTO-1 (sGSTO-1) structure by homology modeling. (A) Predicted tridimensional sGSTO-1 including the T214P substitution (sGSTO-1 T214P). (B) sGSTO-1 and sGSTO-1 T214P overlay. The punctual amino acid substitution (dotted arrow line) and the region of the secondary structure affected (α7 helix, circle) are indicated. (C) Enlarged view of the sGSTO-1 T214P substitution shown in (B). (D) Ramachandran plot for the predicted sGSTO-1 structure. The amino acid distribution into most favored regions (A,B,L), additional allowed regions (a,b,l,p), generously allowed regions (~a,~b,~l,~p), and disallowed regions (GLU83) is indicated. In the protein structures, α-helices (α1- α8), β-sheets (β1-4) are indicated. The reduced glutathione (GSH) molecule is represented.
The predicted protein structure for S. salar CCL19 (PDB ID: 2HCI; score = 55; E-value = 2e-08), ITB2 (PDB ID: 2KCN; score = 42; E-value = 2e-04), HSP70 (PDB ID: 1YUW; score = 1036; E-value = 0), and MHC class I (PDB ID: 1KTL; score = 182; E-value = 4e-46) are also shown (Supplementary Figures 1–4).
Since homology modeling provides only restricted information about side-chain orientations, three of the GSTO-1 variants that evidenced to be more affected in their structure (S26G; S119P; T214P) were further studied by all-atom MD simulations. Based on that the homology models showed a good stereochemical quality, they were further used as initial coordinates for MD simulations. In Figure 6 is showed the global structure of the GSTO-1 protein and highlights the location of each mutation evaluated. Analysis of three replicas of 100 ns of simulation for each protein model (GSTO-1 wild-type, S26G; S119P and T214P) showed that the progression of the root mean square deviation (RMSD) of the Cα atoms as a function of time kept values below 3 Å, suggesting they were stable under the time windows explored (Supplementary Figure 5).
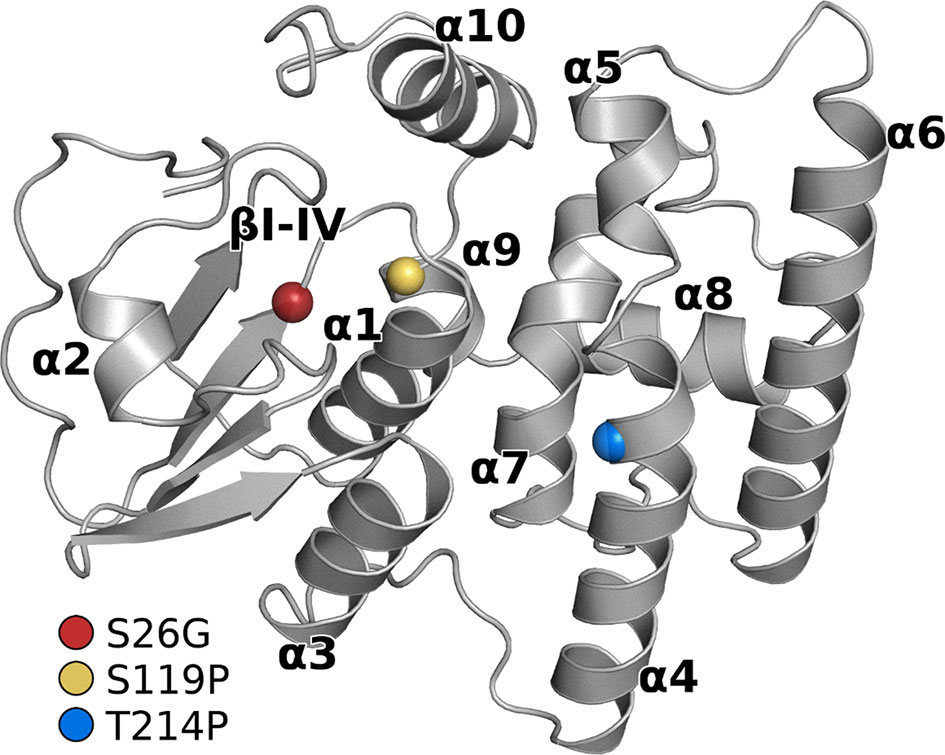
Figure 6 Structure of the GSTO-1 homology model with labeled secondary structure elements. Colored spheres show the position of each punctual variant: red (S26G), yellow (S119P), blue (T214P).
To assess if the GSTO-1 variants have significant differences in their conformational dynamics, we calculated the root mean square fluctuation (RMSF) over the whole trajectories (300 ns) comparing them with the wild-type trajectories. Importantly, even though the GSTO-1 variants are located far apart within the protein structure, all of them seem to produce similar effects on conformational flexibility (Figure 7). Compared to the wild-type GSTO-1 (Figure 7A), in the three sGSTO-1 variants explored [S26G (Figure 7B), S119P (Figure 7C), and T214P (Figure 7D)] the α10 helix located at the C-terminus of the protein is notoriously more rigid. Moreover, the loop that connects the helixes α6 and α7 is significantly rigidized in the sGSTO-1 S26G and S119P compared to the wild-type, whereas for the variant sGSTO-1 T214P this loop is only slightly more rigid than the wild-type GSTO-1. The detailed fluctuational profile for each GSTO-1 variant compared to the WT is shown on Supplementary Figure 6. Altogether, the differences observed in the local flexibility of secondary structure elements suggests that the SNP variations detected have an impact on the GSTO-1 protein structure and, in consequence, a potential impairment in the protein functionality. Nevertheless, further studies are needed to unravel the precise biophysical consequences of these GSTO-1 variants over the function of the enzyme and their consequences at physiological level.
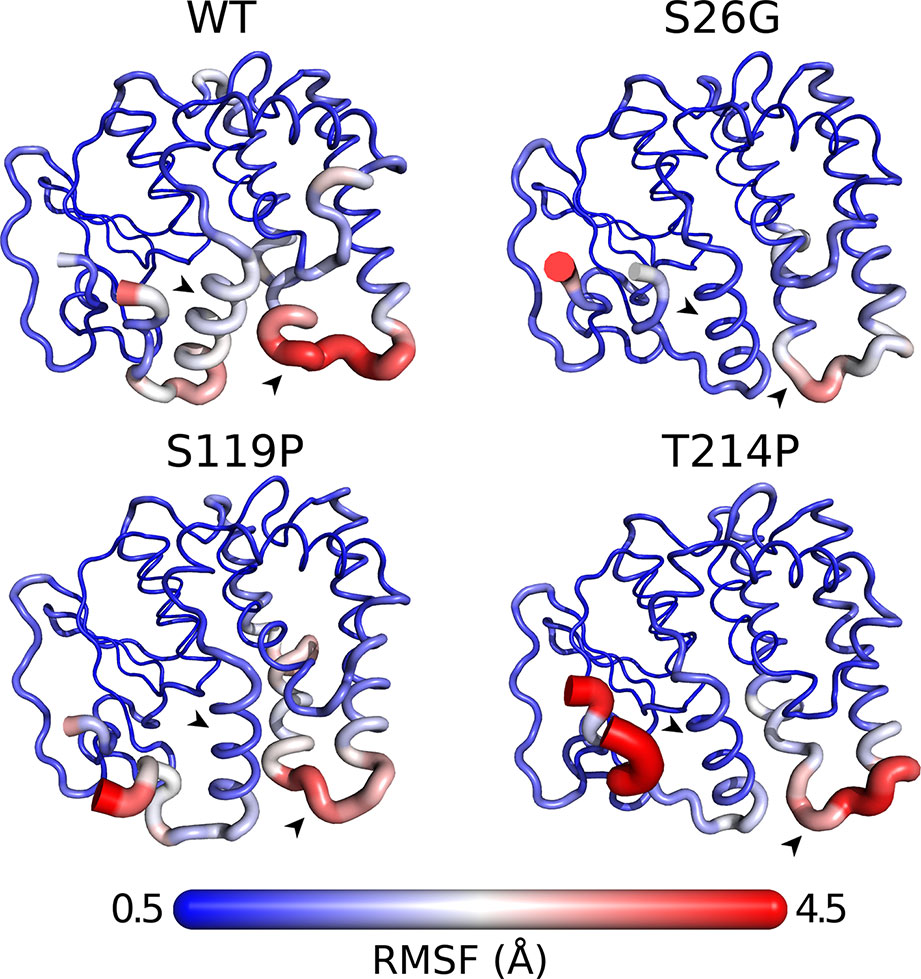
Figure 7 Differences in conformational dynamics between GSTO-1 wild-type (WT), and the variants S26G, S119P, and T214P. The radius of the secondary protein structure and the gradient color (from 0.5 to 4.5 Å) are functions of the RMSF values calculated. As the most affected protein regions, arrowheads highlight the helix α10 (black) and the loop between helixes α6 and α7 (white), respectively.
In this study, we identified a set of SNP located for a group of immune-related genes (ccl19, itβ2, gsto-1, hsp70, mhc-I) with differential gene expression in Salmo salar challenged with IPNV (Cepeda et al., 2011; Cepeda et al., 2012; Reyes-López et al., 2015). These nucleotide variations were localized in both untranslated regions (5’-UTR; 3’-UTR). In the translated region, synonymous (itβ2, gsto-1, hsp70) and nonsynonymous (gsto-1) SNPs were found. The potential impact of the nonsynonymous SNPs was also evaluated based on the predicted tridimensional S. salar GSTO-1 structure obtained through homology modeling. To our knowledge, currently there are no previous studies in Atlantic salmon in which the SNP functional effect had been evaluated at predicted tridimensional protein structure level. These results open the possibility that these candidate SNPs found on genes with immune function could be associated to the inter-individual gene expression variability in Atlantic salmon and their response against pathogens they are exposed at aquaculture rearing conditions.
The interest in identifying SNPs is given by the large amount of information that exists in model organisms that relate these nucleotide variations with susceptibility to diseases (Mooney et al., 2010), which may allow to identify a series of candidate genes as targets for therapies (Voisey and Morris, 2008). In salmon, the association between polymorphism and susceptibility has been suggested (Correa et al., 2017; Holborn et al., 2018). For this reason, the identification of polymorphisms including SNPs in genes of immunological relevance may help to establish associations between polymorphism and susceptibility to diseases affecting the productive sector. In this way, the identification of SNPs and its analysis through the use of different bioinformatics resources (including those intended for the evaluation of risk scores and estimating its consequences in the predicted three-dimensional protein structure from the predicted amino acid sequence) appears as an interesting alternative to select more accurately those candidate SNPs from a set of SNPs identified with a predicted impact.
Different strategies have been used to identify SNP including small (single-strand conformation polymorphism (SSCP), heteroduplex analyses, random shotgun, polymerase chain reaction (PCR) product sequencing, EST analysis) (Liu and Cordes, 2004), and high scale strategies (next generation sequencing (NGS) and whole genome sequencing) (Abdelrahman et al., 2017; Kumar and Kocour, 2017). Particularly, the EST sequence analysis-based SNP identification is not new although it has not been extensively explored. In this ambit, the EST sequences available on data repositories allow to exploit the redundancy existent in these collections making feasible the analysis on transcripts of the same gene from multiple individuals to identify potential true SNPs minimizing the sequencing errors (Hayes et al., 2007a). Such volume of information becomes an excellent database for the search of candidate polymorphisms through the use of bioinformatics approaches (Irizarry et al., 2000; Guryev et al., 2004). In Atlantic salmon previous reports have been carried out to identify SNPs based on ESTs data (Hayes et al., 2007b) with a high percentage (72.4%) of successfully validated SNPs from the putative identified SNPs obtained (Hayes et al., 2007a). Altogether, the search of SNP from the EST database in Atlantic salmon could contribute to the identification of a set of candidate SNP with a high percentage of success, and also allows to extract the maximum possible benefit from the large amount (and often little exploited) of available information in the data repositories. The use of integrated bioinformatics resources to evaluate the potential consequences of those SNPs (in terms of risk factor and predicted protein three-dimensional structure) could favor the selection of certain candidate SNPs for their validation (both genetic and functional) as well as could shed lights of their effects from a physiological context point of view.
According to Hayes et al. (Hayes et al., 2007b), our results showed a higher number of transition (A > G, C > T) than transversion (A > C, A > T, C > G, G > T) nucleotide substitutions. At the same time, our results showed that 25% of the functional nucleotide variants were associated to synonymous and nonsynonymous modifications. Importantly, a previous study reported a total of 23.8% of the SNPs found with these functional variations (Hayes et al., 2007b). These antecedents suggest that the methodology used in this study may result in the identification of a set of candidate SNPs using in silico approaches based on information that is currently contained from public databases.
Previous reports have assessed the use of the bioinformatics tools used in our current study (Mohamed et al., 2008; Capasso et al., 2009; Liu et al., 2009; Wei et al., 2010; Rajasekaran and Sethumadhavan, 2010). Several reported have used FASTSNP as a tool to evaluate the SNP significance in genes associated to disease susceptibility in mammals (Ali Mohamoud et al., 2014; Panda and Suresh, 2014; Phani et al., 2014; Liu et al., 2016; Yu et al., 2017). For SNP identification, we used the FASTSNP decision tree to evaluate whether the nucleotide variation on noncoding region for ccl19, itβ2, gsto-1, hsp70, and mhc-I could have an predictive effect over gene processes including transcription binding sites or splicing regulation (Yuan et al., 2006). Most of these nucleotide substitutions were ranked with a score 1-3, assigned by the predictive change upon transcription factor binding sites. Furthermore, changes affecting predicted exonic splicing enhancer or exonic splicing silencer motif may also compromise the gene function (Fairbrother et al., 2002; Cartegni et al., 2003; Fairbrother et al., 2004; Wang et al., 2004). Importantly, the link between alternative splicing on immune-related genes in Atlantic salmon has been previously proposed using IPNV (Maisey et al., 2011), suggesting that SNPs affecting these regulatory processes could play a relevant role on the host immune response. Alterations in the immune response associated to SNP located on untranslated regions have been reported in mammals including ccl19 (Muinos-Gimeno et al., 2009; Elton et al., 2010; Cai et al., 2014), itβ2 (Bachmann et al., 1997; Mueller et al., 2004; Zhao et al., 2013), and mhc-I (Cree et al., 2010). By contrast, in teleost there is a clear lack of information about polymorphisms located on the coding region of these genes, being only reported previously on hsp70 (Miichthys miiuy) and particularly in mhc-I of Atlantic salmon (Grimholt et al., 2002; Grimholt et al., 2003; Miller et al., 2004; Glover et al., 2007; Wynne et al., 2007), probably because its relevant role on the antigen presentation and the subsequent immune response activation. In regard to the modulation of these genes on Atlantic salmon against pathogens, in IPNV-infected salmon the expression of all the genes evaluated in this current study have been identified and annotated from splenic leukocytes cDNA library obtained from IPNV-infected Atlantic salmon (Cepeda et al., 2011; Cepeda et al., 2012). In addition, the expression for ccl19, itβ2, hsp70, and mhc-I was modulated in head kidney Atlantic salmon full-sibling families (Reyes-López et al., 2015). In the case of ccl19, its expression is upregulated in trout head kidney leucocytes infected with IPNV (Montero et al., 2009), suggesting a role in inflammation and immune response mediated by the recruitment of lymphocytes (Reyes-López et al., 2015). In the same study, we detected a differential expression pattern between IPN-susceptible and IPN-resistant families in the expression of ccl19 and itβ2 (cd18), noting in the susceptible families an abrupt high expression at 1 day post-infection (dpi) to then drop drastically at 5 dpi, meanwhile the expression in the IPN-resistant families remained constant (Reyes-López et al., 2015). In the case of hsp70 and mhc-I a similar upregulation was observed in the susceptible and resistant families to IPNV, but differences were found at 5 dpi when expression dropped to basal levels in susceptible families but not in those resistant, suggesting that an impaired antigen presentation can contribute to the IPN-susceptible phenotype of salmon (Reyes-López et al., 2015). Further studies are needed to elucidate if there is a correlation between the candidate SNPs found from the SNP mining analysis on this current study and IPN-resistance.
Synonymous SNPs have been often called “silent” SNPs because they do not induce a change in the protein amino acid composition due to the degenerating genetic code (more than one codon translate the same amino acid). In the last years, sSNPs are attracting more attention in mammals since these silent mutations appear to be linked to a large list of diseases (Sauna and Kimchi-Sarfaty, 2011). However, in fish no attention has been paid to these mutations so far. In human, these sSNPs can lead to disease by different mechanisms: disrupting splicing signals (Parmley and Hurst, 2007) altering regulatory binding sites (Stergachis et al., 2013); and by changing the secondary structure of the mRNA affecting protein expression (Brest et al., 2011). It has been also described that a change in the codon usage may compromise the translation elongation by introducing a rarer codon (associated to a negative ΔRSCU value). This leads to the use of a less abundant tRNA that may decrease the rate of local elongation and, in consequence, a lower protein synthesis levels and/or protein missfolding. By contrast, the opposite above-mentioned effect may take place in those nucleotide variants with a positive ΔRSCU value (Sauna and Kimchi-Sarfaty, 2011). In our study, one negative ΔRSCU value was assign (hsp70 c.1950C > T), and two positive ΔRSCU values (itβ2 c.2275G > A; gsto-1 c.558G > A). These values suggest that these sSNPs may be associated to differences in the local rate of elongation. Nonetheless, it is not clear if these nucleotide variants may cause a change at physiological level (Sauna and Kimchi-Sarfaty, 2011). Further studies are needed to elucidate the physiological consequences of these polymorphisms.
We obtained the score for the nonsynonymous SNPs based on the cutoffs for SIFT and POLYPHEN because they have previously deemed appropriate for those nucleotide variations that in mammals may play a role on infective processes (Xi et al., 2004; Bhatti et al., 2006). The ranking scheme strategy used in our study has the purpose to evaluate in silico the nucleotide variations on sequences of interest in order to provide semiquantitative information and primarily intended for its use in the absence of biochemical characterization. The nonsynonymous SNPs were found only in the gsto-1 gene sequence. GSTO-1 is an enzyme involved in biotransformation of compounds including toxic substances and oxidative stress products, transport of ligands, and regulation of signaling pathways (Burmeister et al., 2008). The GSTO-1 enzyme is responsible for catalyzing the reaction that results from glutathione conjugates generated by GSH in response to damage signals, thus influencing the relationship between reduced glutathione and oxidized glutathione (GSH/GSSG) modulating the expression of cytokines related to the Th1 immune response (Dobashi et al., 2001), thus playing a key role on the host immune response modulation. To date, very few studies have identified the effect of SNP into the gsto-1 sequence (He et al., 2013; Zmorzynski et al., 2015), including teleost species (Liao et al., 2017). The analysis of the four gsto-1 nonsynonymous SNPs found by SIFT and POLYPHEN showed two of them with the second (c.205A > G; c484T > C) and the highest (c.499T > C; c.769A > C) score possible. These results indicate that the protein would be affected on its tridimensional structure.
Despite the relevance and progress made on the SNP identification, there is a clear lack of studies in which would be proposed predictive models that could help to visualize the consequence of those SNPs located on the translated region and ranked with a high-risk score. In fact, one of the limitations of these tools is there is no reliable proposal about the changes that could take place in the protein structure. Hence, it is not possible to estimate at the structural level if the high impact score for the nucleotide modifications evaluated may have an effect in essential sectors for the protein functionality such as the active site or other fundamental interactions involved in the stabilization of its structure. To carry out this kind of analysis is necessary to characterize the mutant in vitro and determine the protein structure by, for instance, X-ray crystallography. Regrettably, the knowledge about salmon tridimensional protein structures is still very limited, with no protein structural information for GSTO-1. Therefore, in order to evaluate the nonsynonymous SNP effect at protein structure level it is strictly necessary the generation of the predicted tridimensional protein structure using bioinformatics tools. Thus, the homology modeling strategy could be used to predict the tridimensional structure of the protein and, with it, to be able to establish a relationship between the nonsynonymous SNPs-provoked structural protein changes and the relevance of them (SIFT+POLYPHEN score).
Although there is no previous antecedents in fish, the SNP effect on the protein structure has been evaluated on genes of immunologic interest (Jang et al., 2017; Majumdar et al., 2017; Melzer and Palanisamy, 2018), including human glutathione-S-transferase superfamily (Kitteringham et al., 2007). The structure of GSTO-1 is composed mainly of an N-terminal domain of the thioredoxin type which consists of four central β-sheets surrounded at each end by α-helices, and a C-terminal end which is mainly made of α-helices (Board et al., 2000). The latter helices, fold over the N-terminal domain generating a network of hydrogen bonds which define a continuous surface (β/α structure) (Board et al., 2000). This β/α structure could be destabilized by the SNP c.205A > G (S26 amino acid substitution) located in β2 sheet, which could introduce a structural conformational freedom either in the β1 sheet as in the adjacent one (β2 sheet). In addition, on the N-terminal end there is the glutathione binding site, also called G-site, in which the cysteine 32 (located at the N-terminal end of the α1 helix) forms a disulfide bond in the presence of reduced glutathione (GSH) (Rossjohn et al., 1998). It has been described in other GST proteins that a H-bond between the GSH sulfur and the OH of the tyrosine or serine stabilizes the GSH thiolate anion (Kortemme and Creighton, 1995). Mutations of these serine or tyrosine sites lead to a substantial or complete protein inactivation (Stenberg et al., 1991; Board et al., 1995). Based on the predicted tridimensional S. salar GSTO-1, the S26 is located in the proximity of the G-site. Taken together, the S26G substitution could affect the protein functionality, although more studies are needed to establish the consequence at protein level of this candidate nonsynonymous SNP.
The predicted protein structure for sGSTO-1 S119P showed a modification on the α4 helix kink region, rendering it longer compared with the wild-type. This probably because the proline substitution prevents the H-bonds network within the alpha helix. In a previous report has been described that proline modifies the secondary protein structure, suggesting that the replacement of Ser119Pro probably interferes with the α4 helix formation (Chiu et al., 2013). On the other hand, despite the high SIFT+POLYPHEN score obtained for the SNP c.499T > C, no appreciable changes in terms of the secondary structure were observed in the predicted sGSTO-1 Y124H model. However, the physicochemical change may have impacts that are not easily predicted from a structural point of view.
The homology model for the SNP c.769A > C (sGSTO-1 T214P) showed a change in the tridimensional conformation at the C-terminal end of the α7 helix of the protein. The α7 helix is directly involved in the H site of the protein, which has a hydrophobic motif and is adjacent to the G binding site. This site is composed of both the N-terminal and the C-terminal ends and has the putative function of being a binding site for GSH or other target molecules. In the sGSTO-1, the threonine 214 (hydrophilic nature) forms part of a hydrophobic motif, contributing to the formation of H bonds, thus allowing the union of a greater variety of substrates (Board et al., 2000). Therefore, the T214 substitution could decrease the efficiency of the protein, since having an exclusively apolar character decreases the range of substrates that can bind to this active site.
MD simulations of those SNPs with a relevant impact based on the predicted GSTO-1 structure (S26G; S119P; T214P) were also assessed. The results show that these variations affect the dynamics of common regions within the GSTO-1 structure, turning the overall structure of the enzyme more rigid than the wild-type variant. From the three variants above-mentioned assayed, only T214P is located directly in the main affected helix, while the other two (S26G; S119P) are located closer to the N-terminal region. Since the protein structure is a complex network of covalent and noncovalent interactions, the result obtained by MD simulations implies that the individual local effect of each one of these substitutions is being transduced throughout the protein structure and affecting regions that are likely to be determining in the stability or the catalytic capabilities of the enzyme. Since conformational dynamics are an essential aspect of a protein function, these differences are probably impacting the functioning of sGSTO-1, and consequently the fitness of the organism (Bhabha et al., 2013). Enhanced rigidity is a trait commonly associated with more stable proteins (Radestock and Gohlke, 2011). Therefore, it is possible that these SNPs could alter the stability of GSTO-1. On the other hand, although these variations are not located at the binding site of the enzyme, it is possible that these SNPs could have allosteric effects modulating the catalysis. This because conformational dynamics can play a role in multiple aspects of enzymatic activity like accessibility of the substrate to the active site, product release rate and the probability of finding catalytically active populations within the conformational ensemble (Henzler-Wildman et al., 2007; Kokkinidis et al., 2012).
Altogether, and based on the modeled sGSTO-1 structure, most of the candidate nonsynonymous SNP identified showed changed in the secondary structure dynamics. Therefore, the strategy to evaluate nonsynonymous SNP based on homology modeling and all-atom MD simulations provide additional evidence that may help to rank them in order to subsequently validate those that are most relevant according to their structural effect. Further studies should be focused in the in vitro characterization of these enzymes to generate a complete biophysical picture of the effects of these SNPs in these genes associated to immune function.
The conceptualization of the study was performed by EV-V, MI, and FER-L. The methodology was originally proposed by EV-V, KM, AMS, MI, and FER-L. The SNP search and identification was carried out by EV-V, SR-C, JY, and FER-L. The codon usage analysis was carried out by EV-V, HV, and FER-L. The homology modeling was conducted by EV-V, JAR-P, KM, and FER-L. The molecular dynamics simulations were performed by JAR-P, PC, VC-F, and FER-L. EV-V, SR-C, JAR-P, KM, JY, HV, PC, VC-F, LT, AMS, MI, and FER-L participated actively in the data analysis and interpretation. EV-V, SR-C, and FER-L wrote the original draft. All the authors corrected, read, and approved the final manuscript.
This study was supported by INNOVA-CORFO (No. 09MCSS-6691 and 09MCSS-6698), FONDECYT (No. 1161015; 11150807; 11180705; 11181133), DICYT- USACH, VRIDEI-USACH (USA1899 VRIDEI 021943IB-PAP), and Universidad Mayor startup funds (No. OI101205; SR-C). The authors also thank to the grants from CONICYT-BCH (Chile) Postdoctoral fellowship (No. 74170091; EV-V), International postdoctoral stay 2019 (Universidad de Chile, UCH1566; EV-V) and VRIDEI-USACH (FR-L). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2019.01406/full#supplementary-material
Figure S1 | Homology modeling for predicted S. salar CCL19 based on tridimensional H. sapiens CCL19 structure. (A) Tridimensional H. sapiens CCL19 (hCCL19) structure. (B) Predicted tridimensional S. salar CCL19 (sCCL19) structure. (C) hCCL19 and sCCL19 overlay. (D) Ramachandran plot for the predicted sCCL19 structure. The amino acid distribution into most favored regions (A,B,L), additional allowed regions (a,b,l,p), generously allowed regions (~a,~b,~l,~p) is indicated.
Figure S2 | Homology modeling for predicted S. salar ITB2 based on tridimensional H. sapiens ITB2 structure. (A) Tridimensional H. sapiens ITB2 (hITB2) structure. (B) Predicted tridimensional S. salar ITB2 (sITB2) structure. (C) hITB2 and sITB2 overlay. (D) Ramachandran plot for the predicted sITB2 structure. The amino acid distribution into most favored regions (A,B,L), additional allowed regions (a,b,l,p), generously allowed regions (~a,~b,~l,~p) is represented.
Figure S3 | Homology modeling for predicted S. salar HSP70 based on tridimensional H. sapiens HSP70 structure. (A) Tridimensional H. sapiens HSP70 (hHSP70) structure. (B) Predicted tridimensional S. salar HSP70 (sHSP70) structure. (C) hHSP70 and sHSP70 overlay. (D) Ramachandran plot for the predicted sHSP70 structure. The amino acid distribution into most favored regions (A,B,L), additional allowed regions (a,b,l,p), generously allowed regions (~a,~b,~l,~p) and disallowed regions (GLN83 and ALA540) is represented.
Figure S4 | Homology modeling for predicted S. salar MHC class I based on tridimensional H. sapiens MHC class I structure. (A) Tridimensional H. sapiens MHC class I (hMHC class I) structure. (B) Predicted tridimensional S. salar MHC class I (sMHC class I) structure. (C) hMHC class I and sMHC class I overlay. (D) Ramachandran plot for the predicted sMHC class I structure. The amino acid distribution into most favored regions (A,B,L), additional allowed regions (a,b,l,p), generously allowed regions (~a,~b,~l,~p) and disallowed regions (ASP39 and ILE40) is represented.
Figure S5 | Molecular dynamics simulation for the wild-type GSTO-1 (A) and the variants S26G (B), S119P (C), and T214P (D). The root mean square deviation (RMSD) of the Cα atoms on a time windows explored of 100 ns is shown for each replicate.
Figure S6 | Root mean square fluctuation (RMSF) profiles for each mutant analyzed (S26G; S119P; T214P) compared to the GSTO-1 wild-type.
Abdelrahman, H., ElHady, M., Alcivar-Warren, A., Allen, S., Al-Tobasei, R., Bao, L., et al. (2017). Aquaculture genomics, genetics and breeding in the United States: current status, challenges, and priorities for future research. BMC Genomics 18, 191. doi: 10.1186/s12864-017-3557-1
Ahmad, T., Valentovic, M. A., Rankin, G. O. (2018). Effects of cytochrome P450 single nucleotide polymorphisms on methadone metabolism and pharmacodynamics. Biochem. Pharmacol. 153, 196–204. doi: 10.1016/j.bcp.2018.02.020
Ali Mohamoud, H. S., Manwar Hussain, M. R., El-Harouni, A. A., Shaik, N. A., Qasmi, Z. U., Merican, A. F., et al. (2014). First comprehensive in silico analysis of the functional and structural consequences of SNPs in human GalNAc-T1 gene. Comput. Math. Methods Med. 2014, 904052. doi: 10.1155/2014/904052
Bachmann, M. F., McKall-Faienza, K., Schmits, R., Bouchard, D., Beach, J., Speiser, D. E., et al. (1997). Distinct roles for LFA-1 and CD28 during activation of naive T cells: adhesion versus costimulation. Immunity 7, 549–557. doi: 10.1016/S1074-7613(00)80376-3
Bhabha, G., Ekiert, D. C., Jennewein, M., Zmasek, C. M., Tuttle, L. M., Kroon, G., et al. (2013). Divergent evolution of protein conformational dynamics in dihydrofolate reductase. Nat. Struct. Mol. Biol. 20, 1243–1249. doi: 10.1038/nsmb.2676
Bhatti, P., Church, D. M., Rutter, J. L., Struewing, J. P., Sigurdson, A. J. (2006). Candidate single nucleotide polymorphism selection using publicly available tools: a guide for epidemiologists. Am. J. Epidemiol. 164, 794–804. doi: 10.1093/aje/kwj269
Board, P. G., Coggan, M., Wilce, M. C., Parker, M. W. (1995). Evidence for an essential serine residue in the active site of the Theta class glutathione transferases. Biochem. J. 311, 247–250. doi: 10.1042/bj3110247
Board, P. G., Coggan, M., Chelvanayagam, G., Easteal, S., Jermiin, L. S., Schulte, G. K., et al. (2000). Identification, characterization, and crystal structure of the Omega class glutathione transferases. J. Biol. Chem. 275, 24798–24806. doi: 10.1074/jbc.M001706200
Bootland, L. M., Dobos, P., Stevenson, R. M. W. (1991). The IPNV carrier state and demonstration of vertical transmission in experimentally infected brook trout. Dis. Aquat. Organ. 10, 13–21. doi: 10.3354/dao010013
Brest, P., Lapaquette, P., Souidi, M., Lebrigand, K., Cesaro, A., Vouret-Craviari, V., et al. (2011). A synonymous variant in IRGM alters a binding site for miR-196 and causes deregulation of IRGM-dependent xenophagy in Crohn’s disease. Nat. Genet. 43, 242–245. doi: 10.1038/ng.762
Bruno, D. W. (2004). Changes in prevalence of clinical infectious pancreatic necrosis among farmed Scottish Atlantic salmon, Salmo salar L. between 1990 and 2002. Aquaculture 235, 13–26. doi: 10.1016/j.aquaculture.2003.11.035
Burmeister, C., Luersen, K., Heinick, A., Hussein, A., Domagalski, M., Walter, R. D., et al. (2008). Oxidative stress in Caenorhabditis elegans: protective effects of the Omega class glutathione transferase (GSTO-1). FASEB J. 22, 343–354. doi: 10.1096/fj.06-7426com
Cai, W., Tao, J., Zhang, X., Tian, X., Liu, T., Feng, X., et al. (2014). Contribution of homeostatic chemokines CCL19 and CCL21 and their receptor CCR7 to coronary artery disease. Arter. Thromb. Vasc. Biol. 34, 1933–1941. doi: 10.1161/ATVBAHA.113.303081
Capasso, M., Ayala, F., Russo, R., Avvisati, R. A., Asci, R., Iolascon, A. (2009). A predicted functional single-nucleotide polymorphism of bone morphogenetic protein-4 gene affects mRNA expression and shows a significant association with cutaneous melanoma in Southern Italian population. J. Cancer Res. Clin. Oncol. 135, 1799–1807. doi: 10.1007/s00432-009-0628-y
Carlson, C. S., Eberle, M. A., Rieder, M. J., Smith, J. D., Kruglyak, L., Nickerson, D. A. (2003). Additional SNPs and linkage-disequilibrium analyses are necessary for whole-genome association studies in humans. Nat. Genet. 33, 518–521. doi: 10.1038/ng1128
Cartegni, L., Wang, J., Zhu, Z., Zhang, M. Q., Krainer, A. R. (2003). ESEfinder: A web resource to identify exonic splicing enhancers. Nucleic Acids Res. 31, 3568–3571. doi: 10.1093/nar/gkg616
Cepeda, V., Cofre, C., González, R., MacKenzie, S., Vidal, R. (2011). Identification of genes involved in immune response of Atlantic salmon (Salmo salar) to IPN virus infection, using expressed sequence tag (EST) analysis. Aquaculture 318, 54–60. doi: 10.1016/j.aquaculture.2011.04.045
Cepeda, V., Cofre, C., González, R., MacKenzie, S., Vidal, R. (2012). Corrigendum to “Identification of genes involved in immune response of Atlantic salmon (Salmo salar) to IPN virus infection, using expressed sequence tags (EST) analysis” [Aquaculture 318 (2011) 54–60]. Aquaculture 338–341, 314. doi: 10.1016/j.aquaculture.2012.01.002
Chapman, S. J., Hill, A. V. S. (2012). Human genetic susceptibility to infectious disease. Nat. Rev. Genet. 13, 175–188. doi: 10.1038/nrg3114
Cheng, Q., Wu, B., Kager, L., Panetta, J. C., Zheng, J., Pui, C. H., et al. (2004). A substrate specific functional polymorphism of human gamma-glutamyl hydrolase alters catalytic activity and methotrexate polyglutamate accumulation in acute lymphoblastic leukaemia cells. Pharmacogenetics 14, 557–567. doi: 10.1097/01.fpc.0000114761.78957.7e
Chiu, C. C., Singh, S., De Pablo, J. J. (2013). Effect of proline mutations on the monomer conformations of amylin. Biophys. J. 105, 1227–1235. doi: 10.1016/j.bpj.2013.07.029
Correa, K., Lhorente, J. P., Bassini, L., López, M. E., Di Genova, A., Maass, A., et al. (2017). Genome wide association study for resistance to Caligus rogercresseyi in atlantic salmon (Salmo salar L.) using a 50k SNP genotyping array. Aquaculture 472, 61–65. doi: 10.1016/j.aquaculture.2016.04.008
Cree, B. A. C., Rioux, J. D., McCauley, J. L., Pierre-Antoine, F. D. G., Goyette, P., McElroy, J., et al. (2010). A major histocompatibility class I locus contributes to multiple sclerosis susceptibility independently from HLA-DRB1*15:01. PloS One 5, e11296. doi: 10.1371/journal.pone.0011296
Den Dunnen, J. T., Antonarakis, E. (2001). Nomenclature for the description of human sequence variations. Hum. Genet. 109, 121–124. doi: 10.1007/s004390100505
Dijk, E. L. van, Auger, H., Jaszczyszyn, Y., Thermes, C. (2014). Ten years of next-generation sequencing technology. Trends Genet. 30, 418–426. doi: 10.1016/j.tig.2014.07.001
Dobashi, K., Aihara, M., Araki, T., Shimizu, Y., Utsugi, M., Iizuka, K., et al. (2001). Regulation of LPS induced IL-12 production by IFN-gamma and IL-4 through intracellular glutathione status in human alveolar macrophages. Clin. Exp. Immunol. 124, 290–296. doi: 10.1046/j.1365-2249.2001.01535.x
Elton, T. S., Sansom, S. E., Martin, M. M. (2010). Cardiovascular disease, single nucleotide polymorphisms; and the renin angiotensin system: is there a MicroRNA connection? Int. J. Hypertens. 2010, 281692. doi: 10.4061/2010/281692
Ewing, B., Green, P. (1998). Base-Calling of automated sequencer traces using phred. ii. error probabilities. Genome Res. 8, 175–185. doi: 10.1101/gr.8.3.175
Fairbrother, W. G., Yeh, R., Sharp, P. A., Burge, C. B. (2002). Predictive identification of exonic splicing enhancers in human genes. Science 297 (5583), 1007–1013. doi: 10.1126/science.1073774
Fairbrother, W. G., Yeo, G. W., Yeh, R., Goldstein, P., Mawson, M., Sharp, P. A., et al. (2004). RESCUE-ESE identifies candidate exonic splicing enhancers in vertebrate exons. Nucleic Acids Res. 32, 187–190. doi: 10.1093/nar/gkh393
Fellay, J., Ge, D., Shianna, K. V., Colombo, S., Ledergerber, B., Cirulli, E. T., et al. (2009). Common genetic variation and the control of HIV-1 in humans. PloS Genet. 5, e1000791. doi: 10.1371/journal.pgen.1000791
Flanagan, S. E., Patch, A. M., Ellard, S. (2010). Using SIFT and PolyPhen to predict loss-of-function and gain-of-function mutations. Genet. Test Mol. Biomarkers 14, 533–537. doi: 10.1089/gtmb.2010.0036
Fu, G. H., Bai, Z. Y., Xia, J. H., Liu, X. J., Liu, F., Wan, Z. Y., et al. (2014a). Characterization of the LECT2 gene and its associations with resistance to the big belly disease in asian seabass. Fish Shellfish Immunol. 37, 131–138. doi: 10.1016/j.fsi.2014.01.019
Fu, G. H., Wan, Z. Y., Xia, J. H., Liu, F., Liu, X. J., Yue, G. H. (2014b). The MCP-8 gene and its possible association with resistance to Streptococcus agalactiae in tilapia. Fish Shellfish Immunol. 40, 331–336. doi: 10.1016/j.fsi.2014.07.019
Glover, K. A., Grimholt, U., Bakke, H. G., Nilsen, F., Storset, A., Skaala, O. (2007). Major histocompatibility complex (MHC) variation and susceptibility to the sea louse Lepeophtheirus salmonis in Atlantic salmon Salmo salar. Dis. Aquat Organ 76, 57–65. doi: 10.3354/dao076057
Gowin, E., Świątek-Kościelna, B., Kałużna, E., Strauss, E., Wysocki, J., Nowak, J., et al. (2018). How many single-nucleotide polymorphisms (SNPs) must be tested in order to prove susceptibility to bacterial meningitis in children? analysis of 11 SNPs in five genes involved in the immune response and their effect on the susceptibility to bacterial meningitis in children. Innate Immun. 24, 163–170. doi: 10.1177/1753425918762038
Grimholt, U., Drabløs, F., Jørgensen, S. M., Høyheim, B., Stet, R. J. M. (2002). The major histocompatibility class I locus in Atlantic salmon (Salmo salar L.): Polymorphism, linkage analysis and protein modelling. Immunogenetics 54, 570–581. doi: 10.1007/s00251-002-0499-8
Grimholt, U., Larsen, S., Nordmo, R., Midtlyng, P., Kjoeglum, S., Storset, A., et al. (2003). MHC polymorphism and disease resistance in Atlantic salmon (Salmo salar); facing pathogens with single expressed major histocompatibility class I and class II loci. Immunogenetics 55, 210–219. doi: 10.1007/s00251-003-0567-8
Guryev, V., Berezikov, E., Malik, R., Plasterk, R. H. A., Cuppen, E. (2004). Single nucleotide polymorphisms associated with rat expressed sequences. Genome Res. 14, 1438–1443. doi: 10.1101/gr.2154304
Hayes, B. J., Nilsen, K., Berg, P. R., Grindflek, E., Lien, S. (2007a). SNP detection exploiting multiple sources of redundancy in large EST collections improves validation rates. Bioinformatics 23, 1692–1693. doi: 10.1093/bioinformatics/btm154
Hayes, B., Laerdahl, J. K., Lien, S., Moen, T., Berg, P., Hindar, K., et al. (2007b). An extensive resource of single nucleotide polymorphism markers associated with Atlantic salmon (Salmo salar) expressed sequences. Aquaculture 265, 82–90. doi: 10.1016/j.aquaculture.2007.01.037
He, Q.-Y., Ding, L.-J., Sun, H.-X., Yu, B., Dai, A.-Y., Zhang, N.-Y., et al. (2013). FSH modulates the expression of inhibin-alpha and the secretion of inhibins via orphan nuclear receptor NUR77 in ovarian granulosa cells. Mol. Reprod. Dev. 80, 734–743. doi: 10.1002/mrd.22206
Heinemeyer, T., Wingender, E., Reuter, I., Hermjakob, H., Kel, A. E., Kel, O. V., et al. (1998). Databases on transcriptional regulation: TRANSFAC, TRRD and COMPEL. Nucleic Acids Res. 26, 362–367. doi: 10.1093/nar/26.1.362
Henzler-Wildman, K. A., Lei, M., Thai, V., Kerns, S. J., Karplus, M., Kern, D. (2007). A hierarchy of timescales in protein dynamics is linked to enzyme catalysis. Nature 450, 913–916. doi: 10.1038/nature06407
Hill, A. V. S. (2001). The genomics and genetics of human infectious disease susceptibility. Annu. Rev. Genomics Hum. Genet. 2, 373–400. doi: 10.1146/annurev.genom.2.1.373
Hirschhorn, J. N., Daly, M. J. (2005). Genome-wide association studies for common diseases and complex traits. Nat. Rev. Genet. 6, 95–108. doi: 10.1038/nrg1521
Holborn, M. K., Ang, K. P., Elliott, J. A. K., Powell, F., Boulding, E. (2018). Genome wide association analysis for bacterial kidney disease resistance in a commercial North American Atlantic salmon (Salmo salar) population using a 50 K SNP panel. Aquaculture 495, 465–471. doi: 10.1016/j.aquaculture.2018.06.014
Houston, R. D., Davey, J. W., Bishop, S. C., Lowe, N. R., Mota-Velasco, J. C., Hamilton, A., et al. (2012). Characterisation of QTL-linked and genome-wide restriction site-associated DNA (RAD) markers in farmed Atlantic salmon. BMC Genomics 13, 244. doi: 10.1186/1471-2164-13-244
Irizarry, K., Kustanovich, V., Li, C., Brown, N., Nelson, S., Wong, W., et al. (2000). Genome-wide analysis of single-nucleotide polymorphisms in human expressed sequences. Nat. Genet. 26, 233–236. doi: 10.1038/79981
Jang, J.-H., Wei, J.-D., Kim, M., Kim, J.-Y., Cho, A. E., Kim, J.-H. (2017). Leukotriene B4 receptor 2 gene polymorphism (rs1950504, Asp196Gly) leads to enhanced cell motility under low-dose ligand stimulation. Exp. Mol. Med. 49, e402. doi: 10.1038/emm.2017.192
Jia, Z.-Y., Ge, H.-Z., Bai, Y.-Y., Li, C.-T., Shi, L.-Y. (2015). Characterization and SNP variation of the interleukin-1 β gene of bighead carp (Cyprinus pellegrini Tchang, 1933) and five strains of common carp [Cyprinus carpio Linnaeus (1758)] in China. J. Appl. Ichthyol. 31, 1102–1106. doi: 10.1111/jai.12862
Kitteringham, N. R., Palmer, L., Owen, A., Lian, L. Y., Jenkins, R., Dowdall, S., et al. (2007). Detection and biochemical characterisation of a novel polymorphism in the human GSTP1 gene. Biochim. Biophys. Acta 1770, 1240–1247. doi: 10.1016/j.bbagen.2007.05.001
Kokkinidis, M., Glykos, N. M., Fadouloglou, V. E. (2012). Protein flexibility and enzymatic catalysis Adv. Protein Chem. Struct. Bio. 87, 181–218. doi: 10.1016/B978-0-12-398312-1.00007-X
Kongchum, P., Sandel, E., Lutzky, S., Hallerman, E. M., Hulata, G., David, L., et al. (2011). Association between IL-10a single nucleotide polymorphisms and resistance to cyprinid herpesvirus-3 infection in common carp (Cyprinus carpio). Aquaculture 315, 417–421. doi: 10.1016/j.aquaculture.2011.02.035
Kortemme, T., Creighton, T. E. (1995). Ionisation of cysteine residues at the termini of model alpha-helical peptides. Relevance to unusual thiol pKa values in proteins of the thioredoxin family. J. Mol. Biol. 253, 799–812. doi: 10.1006/jmbi.1995.0592
Kumar, G., Kocour, M. (2017). Applications of next-generation sequencing in fisheries research: a review. Fish. Res. 186, 11–22. doi: 10.1016/j.fishres.2016.07.021
Laskowski, R. A., Rullmannn, J. A., MacArthur, M. W., Kaptein, R., Thornton, J. M. (1996). AQUA and PROCHECK-NMR: programs for checking the quality of protein structures solved by NMR. J. Biomol NMR 8, 477–486. doi: 10.1007/BF00228148
Liao, Z., Wan, Q., Shang, X., Su, J. (2017). Large-scale SNP screenings identify markers linked with GCRV resistant traits through transcriptomes of individuals and cell lines in Ctenopharyngodon idella. Sci. Rep. 7, 1184. doi: 10.1038/s41598-017-01338-7
Liu, Z. J., Cordes, J. F. (2004). DNA marker technologies and their applications in aquaculture genetics. Aquaculture 238, 1–37. doi: 10.1016/j.aquaculture.2004.05.027
Liu, Y. H., Li, C. G., Zhou, S. F. (2009). Prediction of deleterious functional effects of non-synonymous single nucleotide polymorphisms in human nuclear receptor genes using a bioinformatics approach. Drug Metab. Lett. 3, 242–286. doi: 10.2174/187231209790218145
Liu, J. M., Cui, Y. Z., Zhang, G. L., Zhou, X. Y., Pang, J. X., Wang, X. Z., et al. (2016). Association between dentin matrix protein 1 (Rs10019009) polymorphism and ankylosing spondylitis in a Chinese Han population from Shandong province. Chin. Med. J. (Engl). 129, 657–664. doi: 10.4103/0366-6999.177972
Maier, J. A., Martinez, C., Kasavajhala, K., Wickstrom, L., Hauser, K. E., Simmerling, C. (2015). ff14SB: improving the accuracy of protein side chain and backbone parameters from ff99SB. J. Chem. Theory Comput. 11, 3696–3713. doi: 10.1021/acs.jctc.5b00255
Maisey, K., Toro-Ascuy, D., Montero, R., Reyes-López, F. E., Imarai, M. (2011). Identification of CD3ϵ, CD4, CD8β splice variants of Atlantic salmon. Fish Shellfish Immunol. 31, 815–822. doi: 10.1016/j.fsi.2011.07.022
Majumdar, I., Nagpal, I., Paul, J. (2017). Homology modeling and in silico prediction of Ulcerative colitis associated polymorphisms of NOD1. Mol. Cell. Probes 35, 8–19. doi: 10.1016/j.mcp.2017.05.009
Martin, M. P., Dean, M., Smith, M. W., Winkler, C., Cerrard, B., Michael, N. L., et al. (1998). Genetic acceleration of AlDS. Progression by a promoter variant of ccr5. Science 282 (5395), 1907–1911. doi: 10.1126/science.282.5395.1907
McAllister, P. E., Schill, W. B., Owens, W. J., Hodge, D. L. (1993). Determining the prevalence of infectious pancreatic necrosis virus in asymptomatic brook trout Salvelinus fontinalis : a study of clinical samples and processing methods. Dis. Aquat. Organims 15, 157–162. doi: 10.3354/dao015157
Melzer, A. M., Palanisamy, N. (2018). Deleterious single nucleotide polymorphisms of protein kinase R identified by the computational approach. Mol. Immunol. 101, 65–73. doi: 10.1016/j.molimm.2018.05.026
Miller, K. M., Winton, J. R., Schulze, a D., Purcell, M. K., Ming, T. J. (2004). Major histocompatibility complex loci are associated with susceptibility of Atlantic salmon to infectious hematopoietic necrosis virus. Environ. Biol. Fishes 69, 307–316. doi: 10.1023/B:EBFI.0000022874.48341.0f
Moen, T., Baranski, M., Sonesson, A. K., Kjoglum, S. (2009). Confirmation and fine-mapping of a major QTL for resistance to infectious pancreatic necrosis in Atlantic salmon (Salmo salar): population-level associations between markers and trait. BMC Genomics 10, 368. doi: 10.1023/B:EBFI.0000022874.48341.0f
Moen, T., Torgersen, J., Santi, N., Davidson, W. S., Baranski, M., Ødegård, J., et al. (2015). Epithelial cadherin determines resistance to infectious pancreatic necrosis virus in Atlantic salmon. Genetics 200, 1313–1326. doi: 10.1534/genetics.115.175406
Mohamed, M. F., Frye, R. F., Langaee, T. Y. (2008). Interpopulation variation frequency of human inosine 5’-monophosphate dehydrogenase type II (IMPDH2) genetic polymorphisms. Genet. Test 12, 513–516. doi: 10.1089/gte.2008.0049
Montero, J., Chaves-Pozo, E., Cuesta, A., Tafalla, C. (2009). Chemokine transcription in rainbow trout (Oncorhynchus mykiss) is differently modulated in response to viral hemorrhagic septicaemia virus (VHSV) or infectious pancreatic necrosis virus (IPNV). Fish Shellfish Immunol. 27, 661–669. doi: 10.1016/j.fsi.2009.08.003
Mooney, S. D., Krishnan, V. G., Evani, U. S. (2010). Bioinformatic tools for identifying disease gene and SNP candidates. In: Barnes, M., Breen, G. (eds) Genetics Variation . Methods Mol. Biol. 628, 307–319. Totowa, NJ: Humana Press. doi: 10.1007/978-1-60327-367-1_17
Mueller, K. L., Daniels, M. A., Felthauser, A., Kao, C., Jameson, S. C., Shimizu, Y. (2004). Cutting edge: LFA-1 integrin-dependent T cell adhesion is regulated by both ag specificity and sensitivity. J. Immunol. 173, 2222–2226. doi: 10.4049/jimmunol.173.4.2222
Muinos-Gimeno, M., Guidi, M., Kagerbauer, B., Martin-Santos, R., Navines, R., Alonso, P., et al. (2009). Allele variants in functional MicroRNA target sites of the neurotrophin-3 receptor gene (NTRK3) as susceptibility factors for anxiety disorders. Hum. Mutat. 30, 1062–1071. doi: 10.1002/humu.21005
Ng, P. C., Henikoff, S. (2003). SIFT: Predicting amino acid changes that affect protein function. Nucleic Acids Res. 31, 3812–3814. doi: 10.1093/nar/gkg509
Nielsen, M., Lundegaard, C., Lund, O., Petersen, T. N. (2010). CPHmodels-3.0–remote homology modeling using structure-guided sequence profiles. Nucleic Acids Res. 38, 576–581. doi: 10.1093/nar/gkq535
Olsson, M. H. M., Søndergaard, C. R., Rostkowski, M., Jensen, J. H. (2011). PROPKA3: Consistent Treatment of Internal and Surface Residues in Empirical pKa predictions. J. Chem. Theory Comput. 7, 525–537. doi: 10.1021/ct100578z
Panda, R., Suresh, P. K. (2014). Computational identification and analysis of functional polymorphisms involved in the activation and detoxification genes implicated in endometriosis. Gene 542, 89–97. doi: 10.1016/j.gene.2014.03.058
Parmley, J. L., Hurst, L. D. (2007). Exonic splicing regulatory elements skew synonymous codon usage near intron-exon boundaries in mammals. Mol. Biol. Evol. 24, 1600–1603. doi: 10.1093/molbev/msm104
Phani, N. M., Acharya, S., Xavy, S., Bhaskaranand, N., Bhat, M. K., Jain, A., et al. (2014). Genetic association of KCNJ10 rs1130183 with seizure susceptibility and computational analysis of deleterious non-synonymous SNPs of KCNJ10 gene. Gene 536, 247–253. doi: 10.1016/j.gene.2013.12.026
Radestock, S., Gohlke, H. (2011). Protein rigidity and thermophilic adaptation. Proteins Struct. Funct. Bioinforma. 79, 1089–1108. doi: 10.1002/prot.22946
Rajasekaran, R., Sethumadhavan, R. (2010). Exploring the structural and functional effect of pRB by significant nsSNP in the coding region of RB1 gene causing retinoblastoma. Sci. China Life Sci. 53, 234–240. doi: 10.1007/s11427-010-0039-y
Rebbeck, T. R., Spitz, M., Wu, X. (2004). Assessing the function of genetic variants in candidate gene association studies. Nat. Rev. Genet. 5, 589–597. doi: 10.1038/nrg1403
Reyes-Cerpa, S., Reyes-López, F., Toro-Ascuy, D., Montero, R., Maisey, K., Acuña-Castillo, C., et al. (2014). Induction of anti-inflammatory cytokine expression by IPNV in persistent infection. Fish Shellfish Immunol. 41, 172–182. doi: 10.1016/j.fsi.2014.08.029
Reyes-López, F. E., Romeo, J. S., Vallejos-Vidal, E., Reyes-Cerpa, S., Sandino, A. M., Tort, L., et al. (2015). Differential immune gene expression profiles in susceptible and resistant full-sibling families of Atlantic salmon (Salmo salar) challenged with infectious pancreatic necrosis virus (IPNV). Dev. Comp. Immunol. 53, 210–221. doi: 10.1016/j.dci.2015.06.017
Roe, D. R., Cheatham, T. E. (2013). PTRAJ and CPPTRAJ: Software for processing and analysis of molecular dynamics trajectory data. J. Chem. Theory Comput. 9, 3084–3095. doi: 10.1021/ct400341p
Rossjohn, J., Polekhina, G., Feil, S. C., Allocati, N., Masulli, M., De Illio, C., et al. (1998). A mixed disulfide bond in bacterial glutathione transferase: functional and evolutionary implications. Structure 6, 721–734. doi: 10.1016/S0969-2126(98)00074-4
Sali, A., Blundell, T. L. (1993). Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 234, 779–815. doi: 10.1006/jmbi.1993.1626
Salkowitz, J. R., Bruse, S. E., Meyerson, H., Valdez, H., Mosier, D. E., Harding, C. V., et al. (2003). CCR5 promoter polymorphism determines macrophage CCR5 density and magnitude of HIV-1 propagation in vitro. Clin. Immunol. 108, 234–240. doi: 10.1016/S1521-6616(03)00147-5
Salomon-Ferrer, R., Case, D. A., Walker, R. C. (2013). An overview of the Amber biomolecular simulation package. Wiley Interdiscip. Rev. Comput. Mol. Sci. 3, 198–210. doi: 10.1002/wcms.1121
Sauna, Z. E., Kimchi-Sarfaty, C. (2011). Understanding the contribution of synonymous mutations to human disease. Nat. Rev. Genet. 12, 683–691. doi: 10.1038/nrg3051
Sharp, M. P., Li, W.-H. (1987). The codon adaptation index - a measure of directional synonymous codon usage bias, and its potential applications. Nucleic Acids Res. 15, 1281–1295. doi: 10.1093/nar/15.3.1281
Skevaki, C., Pararas, M., Kostelidou, K., Tsakris, A., Routsias, J. G. (2015). Single nucleotide polymorphisms of Toll-like receptors and susceptibility to infectious diseases. Clin. Exp. Immunol. 180, 165–177. doi: 10.1111/cei.12578
Srinivasan, S., Clements, J. A., Batra, J. (2016). Single nucleotide polymorphisms in clinics: Fantasy or reality for cancer? Crit. Rev. Clin. Lab. Sci. 53, 29–39. doi: 10.3109/10408363.2015.1075469
Stenberg, G., Board, P. G., Mannervik, B. (1991). Mutation of an evolutionarily conserved tyrosine residue in the active site of a human class Alpha glutathione transferase. FEBS Lett. 293, 153–155. doi: 10.1016/0014-5793(91)81174-7
Stergachis, A. B., Haugen, E., Shafer, A., Fu, W., Vernot, B., Reynolds, A., et al. (2013). Exonic transcription factor binding directs codon choice and affects protein evolution. Science (80), 1367–1372. doi: 10.1126/science.1243490
Tang, J., Leunissen, J. A., Voorrips, R. E., van der Linden, C. G., Vosman, B. (2008). HaploSNPer: a web-based allele and SNP detection tool. BMC Genet. 9, 23. doi: 10.1186/1471-2156-9-23
Tarailo-Graovac, M., Chen, N. (2009). Using RepeatMasker to identify repetitive elements in genomic sequences. Curr. Protoc. Bioinforma. 4.10.1–4.10.14. doi: 10.1002/0471250953.bi0410s25
Voisey, J., Morris, C. P. (2008). SNP technologies for drug discovery: a current review. Curr. Drug Discovery Technol. 5, 230–235. doi: 10.2174/157016308785739811
Wang, D. G., Fan, J., Siao, C., Berno, A., Young, P., Sapolsky, R., et al. (1998). Large-scale identification, mapping, and genotyping of single-nucleotide polymorphisms in the human genome. Science 280 (5366), 1077–1082. doi: 10.1126/science.280.5366.1077
Wang, Z., Rolish, M. E., Yeo, G., Tung, V., Mawson, M., Burge, C. B. (2004). Systematic identification and analysis of exonic splicing silencers. Cell 119, 831–845. doi: 10.1016/j.cell.2004.11.010
Wei, Q., Wang, L., Wang, Q., Kruger, W. D., Dunbrack, R. L. (2010). Testing computational prediction of missense mutation phenotypes: functional characterization of 204 mutations of human cystathionine beta synthase. Proteins 78, 2058–2074. doi: 10.1002/prot.22722
Welter, D., MacArthur, J., Morales, J., Burdett, T., Hall, P., Junkins, H., et al. (2014). The NHGRI GWAS Catalog, a curated resource of SNP-trait associations. Nucleic Acids Res. 42, 1001–1006. doi: 10.1093/nar/gkt1229
Wenne, R. (2018). Single nucleotide polymorphism markers with applications in aquaculture and assessment of its impact on natural populations. Aquat. Living Resour. 31, 2. doi: 10.1051/alr/2017043
Wijmenga, C., Zhernakova, A. (2018). The importance of cohort studies in the post-GWAS era. Nat. Genet. 50, 322–328. doi: 10.1038/s41588-018-0066-3
Wynne, J. W., Cook, M. T., Nowak, B. F., Elliott, N. G. (2007). Major histocompatibility polymorphism associated with resistance towards amoebic gill disease in Atlantic salmon (Salmo salar L.). Fish Shellfish Immunol. 22, 707–717. doi: 10.1016/j.fsi.2006.08.019
Xi, T., Jones, I. M., Mohrenweiser, H. W. (2004). Many amino acid substitution variants identified in DNA repair genes during human population screenings are predicted to impact protein function. Genomics 83, 970–979. doi: 10.1016/j.ygeno.2003.12.016
Yu, Y., Chen, Y., Wang, F.-L., Sun, J., Li, H.-J., Liu, J.-M. (2017). Cytokines Interleukin 4 (IL-4) and Interleukin 10 (IL-10) gene polymorphisms as potential host susceptibility factors in virus-induced encephalitis. Med. Sci. Monit. 23, 4541–4548. doi: 10.12659/MSM.904364
Yuan, H. Y., Chiou, J. J., Tseng, W. H., Liu, C. H., Liu, C. K., Lin, Y. J., et al. (2006). FASTSNP: an always up-to-date and extendable service for SNP function analysis and prioritization. Nucleic Acids Res. 34, 635–641. doi: 10.1093/nar/gkl236
Zhang, G., Chen, X., Chan, L., Zhang, M., Zhu, B., Wang, L., et al. (2011). An SNP selection strategy identified IL-22 associating with susceptibility to tuberculosis in Chinese. Sci. Rep. 1, 20. doi: 10.1038/srep00020
Zhang, G., Zhou, B., Wang, W., Zhang, M., Zhao, Y., Wang, Z., et al. (2012). A functional single-nucleotide polymorphism in the promoter of the gene encoding interleukin 6 is associated with susceptibility to tuberculosis. J. Infect. Dis. 205, 1697–1704. doi: 10.1093/infdis/jis266
Zhao, R., Song, Z., Dong, R., Li, H., Shen, C., Zheng, S. (2013). Polymorphism of ITGB2 gene 3’-UTR+145C/A is associated with biliary atresia. Digestion 88, 65–71. doi: 10.1159/000352025
Keywords: single-nucleotide polymorphism, immune response, synonymous SNP, nonsynonymous SNP, homology modeling, 3D protein structure, molecular dynamics simulation, Salmo salar
Citation: Vallejos-Vidal E, Reyes-Cerpa S, Rivas-Pardo JA, Maisey K, Yáñez JM, Valenzuela H, Cea PA, Castro-Fernandez V, Tort L, Sandino AM, Imarai M and Reyes-López FE (2020) Single-Nucleotide Polymorphisms (SNP) Mining and Their Effect on the Tridimensional Protein Structure Prediction in a Set of Immunity-Related Expressed Sequence Tags (EST) in Atlantic Salmon (Salmo salar). Front. Genet. 10:1406. doi: 10.3389/fgene.2019.01406
Received: 04 December 2018; Accepted: 24 December 2019;
Published: 27 February 2020.
Edited by:
Gen Hua Yue, Temasek Life Sciences Laboratory, SingaporeReviewed by:
Vladimir M. Milenkovic, University Medical Center Regensburg, GermanyCopyright © 2020 Vallejos-Vidal, Reyes-Cerpa, Rivas-Pardo, Maisey, Yáñez, Valenzuela, Cea, Castro-Fernandez, Tort, Sandino, Imarai and Reyes-López. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mónica Imarai, bW9uaWNhLmltYXJhaUB1c2FjaC5jbA==; Felipe E. Reyes-López, RmVsaXBlLlJleWVzQHVhYi5jYXQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.