
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 22 January 2020
Sec. Livestock Genomics
Volume 10 - 2019 | https://doi.org/10.3389/fgene.2019.01287
 Xinghua Li1Changsheng Nie1Yuchen Liu1Yu Chen2Xueze Lv2Liang Wang2Jianwei Zhang2Weifang Yang2Kaiyang Li2Chuanwei Zheng3Yaxiong Jia4*Zhonghua Ning1*Lujiang Qu1*
Xinghua Li1Changsheng Nie1Yuchen Liu1Yu Chen2Xueze Lv2Liang Wang2Jianwei Zhang2Weifang Yang2Kaiyang Li2Chuanwei Zheng3Yaxiong Jia4*Zhonghua Ning1*Lujiang Qu1*New-born chicks are vulnerable to bacterial infections and not good at regulating body temperature. There is a close relationship between thermal regulation and immunity, however, the underlying mechanism is not well understood. Salmonella Pullorum (SP) is a major concern in developing countries and causes significant economic losses in poultry industry. Early body temperature (EBT) has previously shown to be correlated with host immunity and resistance to pullorum disease. In this study, we challenged three independent chick populations (Beijing You, Dwarf and Rhode Island Red) with SP at 4 days of age, and rectal temperature was measured before and after the SP attack from 2 to 7 days of age. Host defense to SP was evaluated by survival and spleen SP carrier status. The results showed that chicks with higher EBT before SP infection tend to have higher resistance to later SP attack in two populations (Dwarf and Beijing You). The association between EBT before SP attack and SP resistance was non-significant in Rohde Island Red population (P = 0.06), but the trend was consistent with the other two populations. We also found low to moderate heritability in all three populations for EBT before and after the SP attack ranging from 0.14 to 0.20. Genome-wide association studies identified several genomic regions and biological pathways determining EBT before SP attack, which provides candidate functional genes of this trait. Our results reveal the genetic determination of EBT, and the relationship between EBT and SP resistance, providing an alternative strategy for improving SP resistant activities in chicken.
Pullorum disease, an acute and systematic chick infectious disease caused by Salmonella Pullorum (SP), occurs frequently and is a major economic concern for chicken farms in developing countries (Barrow et al., 2012). SP can cause high mortality rates in chicks under 20 days old and yet few symptoms in adult chickens. In the poultry industry, disease resistance is an important trait with substantial economic value and disease control is drawing more and more public concerns (Cheng et al., 2013). Existing disease management is not enough to prevent disease outbreaks and genetic selection for resistant birds has been regarded as a promising complementary strategy. Difficulty in measuring disease resistance phenotypes remains a major obstacle in genetic research and breeding. This difficulty arises since it is demanding and costly to do large-scale challenge tests in farm settings. Additionally, different infection models have been used in the study of Salmonella resistance, complicating the interpretation of results (Calenge et al., 2010). Despite these problems, it is important to identify traits associated with SP resistance to aid in breeding and selection strategies.
Body temperature is an important physiological trait and indicator of health status (Sund-Levander et al., 2002). Young animals are unable to regulate their own body temperature and generally susceptible to various pathogen infections. There is a pronounced rise in the body temperature of chicks during the first week after hatching, particularly during the first four days (Lamoreux and Hutt, 1939). There are also breed differences of thermoregulation in newly hatched chicks (Dunnington and Siegel, 1984), indicating a genetic determination of early body temperature (EBT). Interestingly, chicken resistance to SP has been shown to be positively correlated with rapid rise of body temperature in the first 10 days post-hatch (Roberts and Cards, 1935). A two-generation selection experiment further confirmed the association between EBT and SP resistance (Hutt and Crawford, 1960a), suggesting the possibility of breeding resistant chicks based on EBT instead of pathogen exposure. In fact, many studies have reported the close relationship between body temperature and immunity regulation (Hori et al., 1991; Watanabe et al., 2008; Fisher et al., 2010; Morrison and Nakamura, 2019). However, the genetic basis of EBT has not been systematically investigated and its correlation with resistance to SP needs more detailed demonstration.
In the present study, we orally challenged three independent chicken breeds/lines with SP at 4th day post-hatch, including a highly selected commercial line (Rhode Island Red, RIR), a Chinese local line (Beijing You, BY), and a synthetic layer line (dwarf, DW) to achieve disease phenotypes. We recorded the body temperature of chicks before and after the SP challenge to better understanding the relationship between EBT and SP infection. We conducted variance component analyses to estimate genetic parameters. We also carried out genome-wide association studies to identify genomic regions and candidate genes correlated EBT before SP infection.
All experiments were approved by the Animal Care and Use Committee of China Agricultural University (Approval ID: XXCB-20090209). All the animals were fed and handled according to the regulations and guidelines established by this committee, and all efforts were made to minimize suffering.
The details about the chicken populations and challenge test procedure can be found in (Li et al., 2018). Briefly, 621 DW, 586 RIR, and 482 BY chicks were orally inoculated with 4.8×107 CFU of SP culture at 4 days post-hatch. 40 chicks from each line were randomly selected as control group, which was mock-challenged with the same volume (0.5 mL) of phosphate buffer saline. After 36 days, all chicks still alive were killed by neck dislocation, and their spleens were removed aseptically and immediately plated onto MacConkey agar media with an inoculation loop in order to check for the presence of SP. Chicks were classed as either susceptible (dead), carriers (with detectible SP load in the spleen) or clear (without detectible SP load in the spleen).
Chick temperature was estimated with clinical electronic thermometers. Thermometer probes were dipped in glycerin to facilitate insertion into the cloaca, and held in the chick rectum at 1.5 cm for 15 s before recording temperature. All temperature measurements were conducted between 13:00-16:00 from 2 to 7 days post-hatch. Early body temperature (EBT) was defined as the average temperature of 2 to 4 days post-hatch, because chick body temperature experiences a dramatic rise during the first week after hatching (Lamoreux and Hutt, 1939; Hutt and Crawford, 1960a). Temperature measurement at 4 days post-hatch was taken before SP inoculation. Fever was determined as the average temperature of 5 to 7 days.
Chicks in each population were designated as either susceptible, carrier or clear, which represents different degrees of SP resistance. The differential analysis among different resistance groups was carried out using PROC GLM of SAS (version 9.2; SAS Institute Inc., Cary, NC).
For our genetic analysis, SP resistance was defined as a three-state categorical trait (dead, carrier, clear), and temperature as a continuous trait. Variance and covariance components were estimated using the DMU software package (Madsen and Jensen, 2008). Bivariate analyses of resistance-EBT were performed using a bivariate threshold model, which was fitted using the RJMC module in the DMU software package. Heritabilities of EBT were estimated with AI-REML in a linear model of the DMU software package. Both models included the fixed effect of the cage and the random effects of additive genetics, as shown in matrix notation:
where y is a vector of phenotypic value; β and a are the vectors of fixed effects and random additive effects, respectively; X and Z are appropriate incidence matrices of fixed effects and random additive effects, respectively; and e is the random residuals.
We isolated individual genomic DNA from blood or muscle samples by the classical phenol-chloroform procedure. The DNA integrity was verified by agarose gel electrophoresis and the purity was checked by A260/280 ratio using the NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific™). Not all the chicks were used for genotyping. Only the individuals with conclusive symptom and high DNA quality were genotyped. Some individuals in the clearance group were also randomly abandoned to match the corresponding phenotype proportion. A total of 842 samples were sent for genotyping. A total of 842 qualified individual genomic DNA samples were genotyped using the Affymetrix 600 K chicken high-density array (Affymetrix, Inc. Santa Clara, CA, USA).
The software Axiom Analysis Suite 3.1 was used for single-nucleotide polymorphism (SNP) calling and initial quality control with the Best Practices Workflow setting from the raw data in the form of CEL files. Only samples with dish quality control (DQC) of 0.82 or better and call rate >95% were used for the downstream analyses. The SNP QC metrics were set to default values recommended by Affymetrix, except that only “PolyHighResolution” SNPs were used in our analysis. Furthermore, we excluded SNPs with unknown or repeated physical positions with an R script. SNPs on sex chromosomes were also discarded because the current statistical methods are not powerful enough to detect association between phenotypes and sex-related genotypes. After these QC steps, 818 samples and 452,291 SNPs remained. To increase the power of association analysis, we dropped 85,140 SNPs due to low variation among subpopulations (monomorphic in any one of the three breeds), 62,450 variants deviating from Hardy-Weinberg equilibrium (HWE) test (P < 1 × 10-5) and 1,774 SNPs with minor allele frequency (MAF) < 0.05 using PLINK v1.90 (Purcell et al., 2007). Missing genotypes were imputed based on information of the remaining SNP genotypes from each subpopulation separately, according to the software Beagle Version 4 (Browning and Browning, 2009). A total of 818 samples and 302,927 SNPs were included in the subsequent genome-wide association analysis. The detailed sample composition used in the GWAS is presented in the Supplementary Table 1.
Population structure and relatedness are major sources of confounding effects in genetic association studies (Astle and Balding, 2009). The most popular method for GWAS containing related individuals is the linear mixed model (LMM) method due to its effectiveness in controlling population stratification bias and reducing the inflation from polygenic background (Yu et al., 2006; Kang et al., 2008; Kang et al., 2010; Zhang et al., 2010; Lippert et al., 2011; Zhou and Stephens, 2012; Loh et al., 2015). In this study, we assessed the population structure through a principal component analysis (PCA) implemented in PLINK package. Considering that SNP clusters in high linkage disequilibrium may bias the PCA results, we first pruned the full SNP set to 23,870 independent SNPs using the -indep-pairwise 25 5 0.2 command parameters in PLINK. These unlinked SNPs were then used to calculate the top three PCs used as covariates in the mixed model. Furthermore, a pairwise kinship matrix was built using the pruned SNPs.
A single-marker univariate linear mixed model was used for testing associations between early body temperature and effective SNPs. The disease phenotype was analyzed using the following model:
In this formula, y denotes the trait values of body temperature; W refers to a matrix of covariates (fixed effects that contain the top three principal components, genotyping batch and a column of 1s) controlling for population structure and batch effect; α is a vector of corresponding coefficients that includes the intercept; x is a vector of the marker genotypes; β is the corresponding marker’s effect; u is a vector of random polygenic effects with a covariance structure as u~N (0, KVg), where K represents a genetic relatedness matrix derived from independent SNPs and Vg is the polygenic additive variance; and ϵ is a vector of random errors. The association analysis was performed using the GEMMA v0.96 software (Zhou and Stephens, 2012). The Wald test statistic AGB=Bleaf+Bstem+Bpaniclewas used to test the null hypothesis β = 0 for each SNP. We used the “qqman” package in R to draw the Manhattan and quantile-quantile (Q-Q) plots. Moreover, the genomic inflation factor λ was estimated using “GenABEL” package to evaluate the control for population stratification. (Aulchenko et al., 2007).
The P-value threshold of genome-wide significance was calculated through the simpleM method implemented in a R script for multiple testing correction (Gao et al., 2010). Through the simpleM calculation, a total of 72,648 independent effective tests were obtained. The genomewide and suggestive significance values were then calculated as 6.88e-7 (0.05/72,648) and 1.38e-5 (1.00/72,648), respectively.
The gene position information was annotated using BioMart in the Ensembl database (Ensembl Genes 96). Candidate genes were retrieved within 1 Mb of the significant SNPs. To provide insight into the functional enrichment of candidate genes, we carried out GO (Gene Ontology) and Pathway analysis using Metascape (Zhou et al., 2019). Metascape is an integrated portal that combines functional enrichment, interactome analysis, gene annotation, and membership search to leverage over 40 independent databases. As the knowledge base for human is the most comprehensive, we converted the chicken genes to their human orthologs for enrichment analysis.
Mortality rates after SP inoculation were 25.1% for RIR, 8.3% for BY and 22.7% for DW. For RIR, 17.9% of chicks were classed as carriers, compared with 0.6% for BY and 15.8% for DW. Finally, 57% of RIR completed cleared the infection, compared with 91.1% for BY and 61.5% for DW.
Normal temperature variation of the control group from 2 to 7 days post-hatch is shown in Figure 1. EBT increased over time in all three lines, and is particularly sharp during the first two days of measurement, consistent with earlier studies (Hutt and Crawford, 1960b).At 7 post-hatch, the average temperatures of three populations were not significantly different. The mean, standard deviation (SD), coefficient of variation (CV), minimum and maximum values of average early temperature are shown in Table 1. We found that the average EBT for DW chicks was 0.3–0.4°C less than the other two lines(p-value < 0.01).

Table 1 Early body temperature in three study lines.
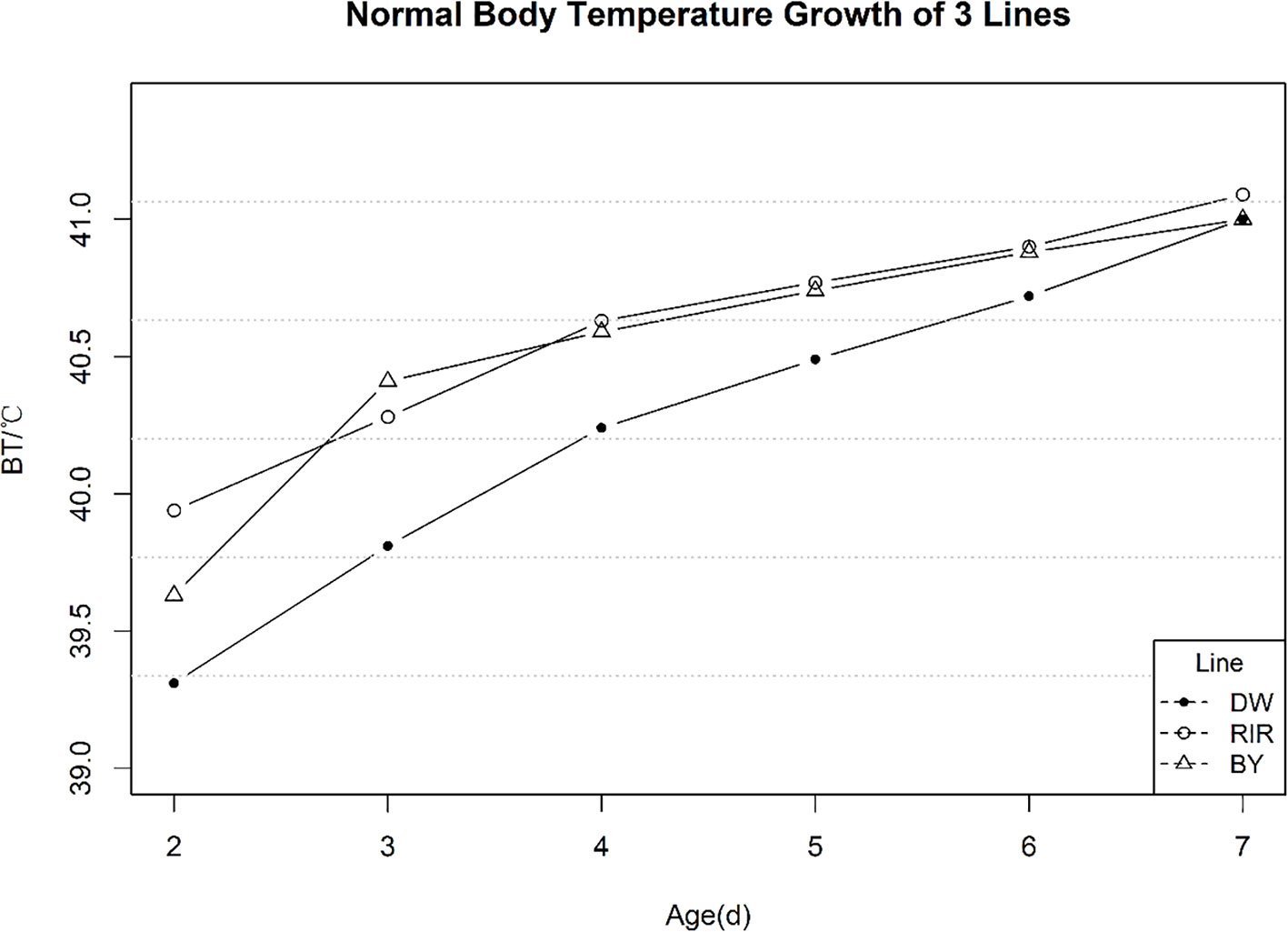
Figure 1 Normal Early Temperature of 3 Lines.
Differences in EBT for chicks with different SP outcomes is shown in Figure 2. After infection, EBT for challenged chicks increased significantly compared to the control group (p-value < 0.01).
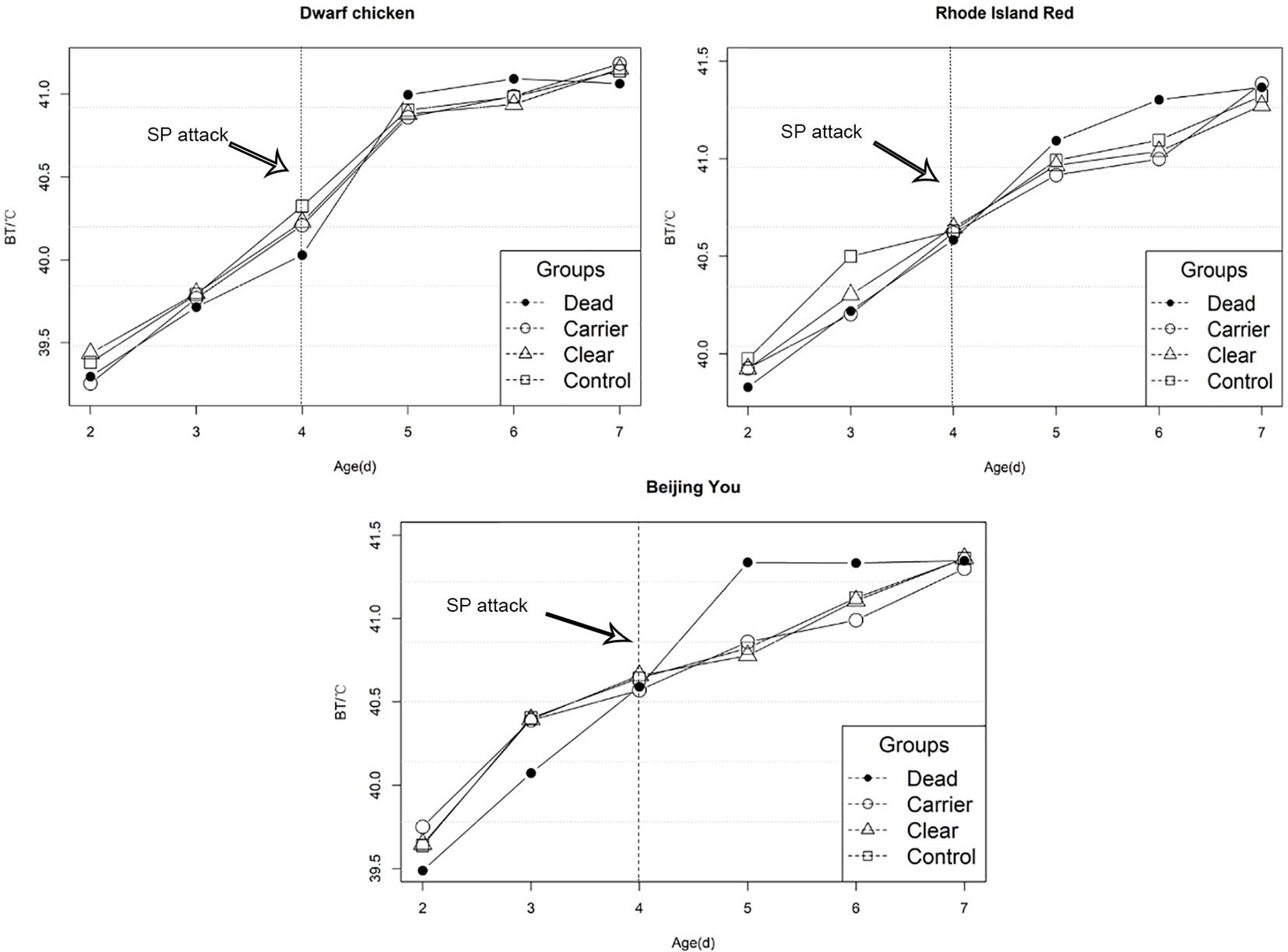
Figure 2 Early body temperature variation before and after salmonella infection. Chicks were challenged with SP at 4 days of age.
As shown in Figure 3, we found that average EBT for SP-susceptible birds was lower in all three chicken lines compared to carrier and resistant birds (p < 0.05 in DW and BY). EBT was also lower for RIR; however, the results were not statistically significant (p-value = 0.06). There was no significant difference in EBT between carrier and clear birds (p-value > 0.05).
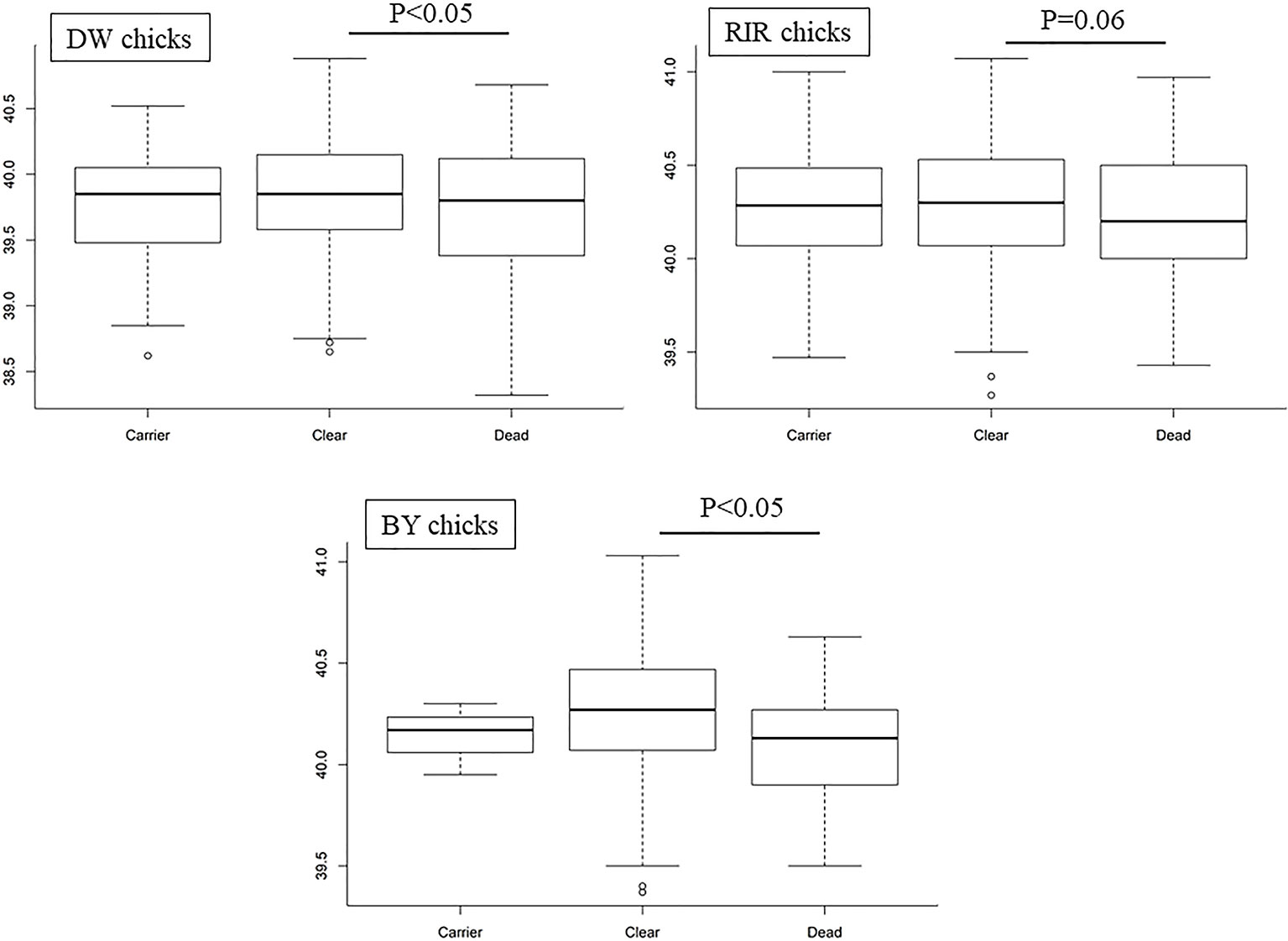
Figure 3 Boxplots of temperature from different Salmonella Pullorum resistance; P represents the p-value between Dead and Clear groups.
Genetic parameters are important references for the selection of infection-resistant animals in breeding. Higher heritability means that the trait could be improved through direct phenotype selection. Most disease trait showed heritabilities lower than 0.1, which means the selection could not work very well. However, we can use other traits to carry an indirect selection. The estimated heritability of EBT is shown in Table 2. EBT before and after SP infection shows low to moderate heritability (0.14–0.20). The genetic correlation between resistance and EBT is 0.23 in DW, 0.09 in RIR and 0.04 in BY, however the reduced correlation in BY is likely due in large part to the low overall mortality in this population. Our results suggest that selection for increased early EBT in some populations would be a useful tool to increase SP resistance.
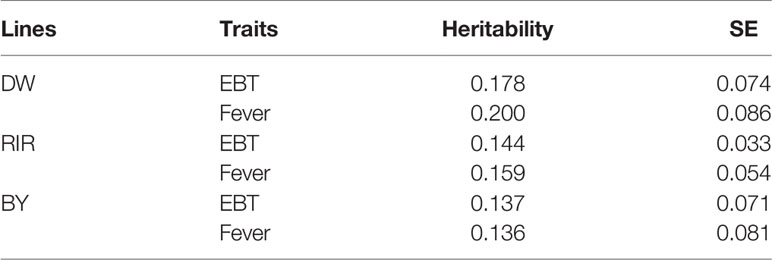
Table 2 Heritability of early body temperature and fever.
Since all of the GWASs were carried out within a pure breed population, the population stratification was not a major confounding effect in our study. The inflation factors (λ) were estimated to be 0.9996, 1.0267 and 1.0093 in BY, DW and RIR respectively. The Manhattan plot and the Q-Q plot for the GWAS results are displayed in Figure 4. In the BY population, the single-marker association analysis identified 2 SNPs above the genome-wide significant threshold on chromosome 15. In the DW population, 3 SNPs crossing the suggestive genomewide significant threshold were identified on chromosomes 2 and 4. However, no significant SNP was identified in the RIR population. Details of the significant markers identified are presented in Table 3. One of the five SNPs are upstream SNP, and the other four are intron SNP.
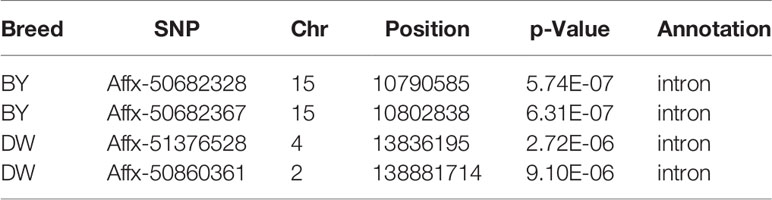
Table 3 List of significant SNPs of genome-wide association studies.
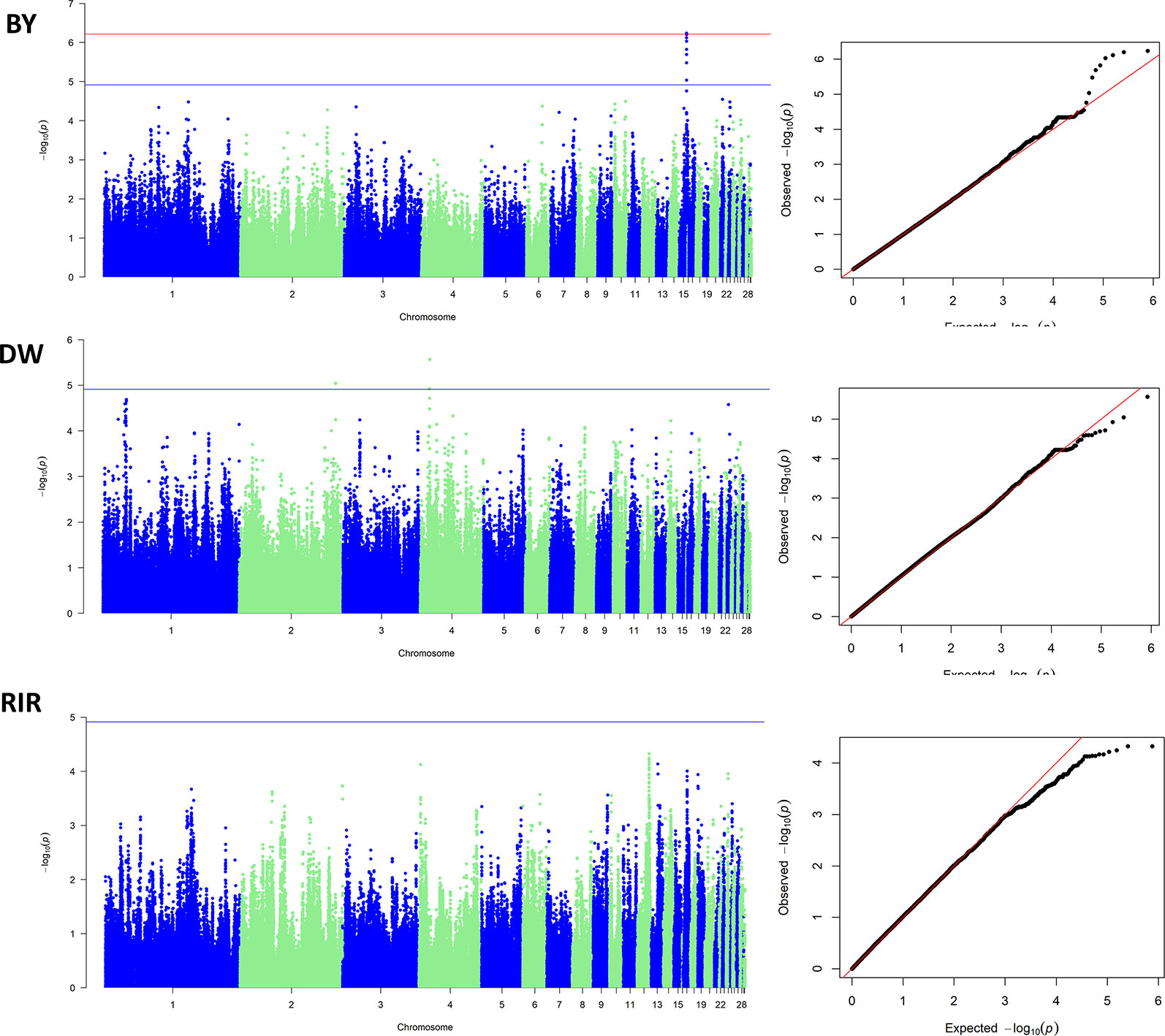
Figure 4 Manhattan plots and Q-Q plots of GWAS results.
There were three significant regions detected to be associated with EBT in total. With the help of BioMart in the Ensembl database, we achieved 162 genes located within 1 Mb of the significant SNPs. These gene IDs were converted into their human orthologs, and we finally obtained 118 gene IDs which were sent to Metascape for enrichment analysis. GO and Pathway analysis were performed to determined the biological functions of these genes. 57 GO biological processes were detected, such as oxidative phosphorylation, ubiquitin-dependent protein catabolic process and MAPK family signaling cascades. Seven KEGG pathway were found, including Alzheimer’s disease, Arginine and proline metabolism, Amyotrophic lateral sclerosis (ALS) Oxidative phosphorylation, Parkinson’s disease, Non-alcoholic fatty liver disease (NAFLD), and Cardiac muscle contraction. Five Reactome gene sets were identified, including MAPK family signaling cascades, Neddylation, S Phase, MAPK6/MAPK4 signaling and Platelet homeostasis. Heatmap of top 13 clusters with their representative enriched terms is presented in Figure 5.
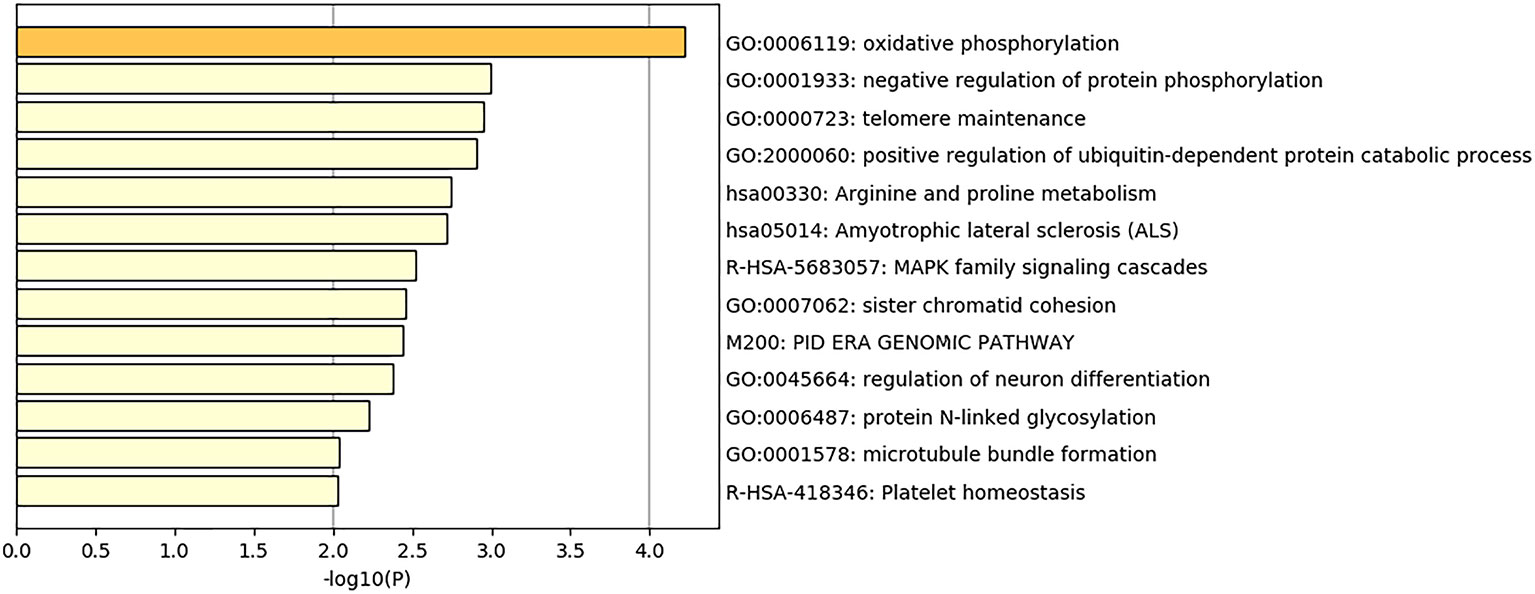
Figure 5 Heatmap of top 13 clusters with their representative enriched terms.
Thermal regulation is a basic physiological process in response to infection, which has been conserved in warm-blooded and cold-blooded vertebrates’ evolution for more than 600 million years (Evans et al., 2015). A fever response is usually beneficial for the host survival during infection, and young animals show different body temperature regulations and disease resistance. It is easy to associate these two traits which may share some genetic determination.
There has been a significant interest in the genetic basis of Salmonella resistance (Berthelot et al., 1998; Beaumont et al., 1999; Girard-Santosuosso et al., 2002). In the current study, we found major differences in genetic correlations between EBT and SP resistance among different chicken populations, implying that genetic background has an important influence in these traits. In both DW and BY chickens, we observe that higher EBT tends to be associated with greater SP resistance, consistent with previous research(Hutt and Crawford, 1960b). Differences in EBT could reflect the transition rate from poikilothermy to homiothermy in the chick, and may be related to better control of thermoregulation. Alternatively, the relationship between EBT and SP resistance may be related to immune defense mechanisms such as phagocytosis, bacteriolysis or the production of antibodies (Scholes, 1940).
We also provide a detailed description of host temperature dynamics following salmonella infection in chickens. The DW chicks with the sex-linked dwarf allele (dw) showed a lower EBT than the normal chicks, which is consistent with Dunnington’s result (Dunnington and Siegel, 1984). We found evidence of increased temperature in all populations after inoculation. After challenge, susceptible birds showed significantly greater EBT than carrier or resistant birds, suggesting that they suffered more severe infection. This is somewhat unexpected, as fever is typically associated with bacterial infection and has been shown to increase host survival (Kane and Peterson, 1976; Kluger et al., 1975). However, an acute body weight loss occurred as anticipated. Besides, we observe that both clear and carrier chicken showed longer sustained elevated EBT after infection, in contrast to susceptible chicks which showed rapid increases in EBT initially, with a decrease two days post-infection.
For the first time, we estimated the heritability of early body temperature. As showed in Table 2, EBT is a trait with low to moderate heritability, indicating genetic determination plays an important role. GWAS remains a promising method for studying genomic architecture of complex traits. Our study identified 4 significant SNPs associated with early body temperature in total. However, the significant regions do not overlap among different chicken breeds, showing that the genetic composition of EBT is relatively complex just as many disease traits. The most significant SNP, Affx-50682328 is located at the intron of OSBP2 gene on GGA15. The annotation of OSBP2 (oxysterol binding protein 2) showed it has the function of cholesterol binding and lipid transporter activity. Other positional candidate genes include several Class I MHC mediated antigen processing and presentation genes such as FBXO21, FBXO32, and FBXW8; immune-related genes such as CARD8, DTX1, and TRIB1; RNA processing genes such as SF3A1, ZMAT5, DDX54. In mice study, FBXO21 was found to facilitate Lys29-linkage and activation of ASK1 (apoptosis signal-regulating kinase 1) and promote type I interferon production upon viral infection (Yu et al., 2016). Genetic variants in CARD8 gene have been reported to be associated with many infectious diseases such as inflammatory bowel disease (McGovern et al., 2006), Crohn’s disease (Roberts et al., 2010) and tuberculosis (Pontillo et al., 2013). TRIB1 was reported to play a critical role in differentiation of tissue-resident M2-like macrophages (Satoh et al., 2013). These immune-related genes are very likely to connected early body temperature and SP resistance. In another chicken body temperature GWA study, Van Goor identified QTL for body temperature on GGA 14, 15, 26, and 27 (Van Goor et al., 2015). Though there are no overlapped genomic regions, our study shared a candidate gene (DDX54) with theirs (DDX42), both of which are putative RNA helicases and implicated in a number of cellular processes involving alteration of RNA secondary structure such as translation initiation, nuclear and mitochondrial splicing, and ribosome and spliceosome assembly (Linder, 2006). Interestingly, RNA splicing was recently found to be correlated with body temperature (Preußner et al., 2017), pointing out that candidate genes with similar functions deserve further research.
The present study focused on investigating the genetic determination of EBT, the correlation with SP resistance and providing positional candidate genes. More researches are necessary to explore the biological processes and molecular mechanism of these genes to associate immune activities and body temperature regulation.
In conclusion, our study describes early body temperature variation before and after SP infection. Controlling for genetic background, we show that elevated EBT is associated with greater SP resistance in BY population, and that EBT and SP resistance are correlated in the BY and DW population. Our results could prove useful in designing breeding programs for improvement of SP resistance in some chicken breeds, and provide positional candidate genes for genetic determination of early body temperature.
The datasets generated for this study can be found in FigShare https://figshare.com/articles/EBT-GWAS_zip/10059092.
The animal study was reviewed and approved by Animal Care and Use Committee of China Agricultural University.
LQ, ZN and YJ conceived and designed the research. XLi performed the experiments, analyzed, and interpreted the data, and wrote the manuscript. CN, YJ and YL performed the experiments and interpreted the data. YC, XLv, LW, JZ, WY, CZ and KL interpreted the data and contributed reagents and materials.
This work was supported by the earmarked fund for the Beijing Innovation Team of the Modern Agro-industry Technology Research System (BAIC04–2018, BAIC04–2019), National Natural Science Foundation of China (31772581) and Chinese Agricultural Research System (CARS-41).
Author CZ was employed by company Beinongda Technology Co., Ltd, China.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We gratefully acknowledge our colleagues in the Poultry Team at the National Engineering Laboratory for Animal Breeding of China Agricultural University, for their assistance on sample collection and helpful comments on the study, and Judith Mank from University of British Columbia, for her kind help on the polish of the manuscript. We thank Beinongda Technology co., LTD, for their kind help in the preparation of chicken populations.
BY, Beijing You; DW, Dwarf Chicken; EBT, Early body temperature; GWAS, genome-wide association study; LD, linkage disequilibrium; LMM, linear mixed model; PCA, principle component analysis; RIR, Rhode Island Red; SNP, single-nucleotide polymorphism; SP, Salmonella pullorum.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2019.01287/full#supplementary-material
Astle, W., Balding, D. J. (2009). Population structure and cryptic relatedness in genetic association studies. Statist. Sci. 24, 451–471. doi: 10.1214/09-STS307
Aulchenko, Y. S., Ripke, S., Isaacs, A., van Duijn, C. M. (2007). GenABEL: an R library for genome-wide association analysis. Bioinformatics 23, 1294–1296. doi: 10.1093/bioinformatics/btm108
Barrow, P. A., Jones, M. A., Smith, A. L., Wigley, P. (2012). The long view: Salmonella – the last forty years. Avian Pathol. 41, 413–420. doi: 10.1080/03079457.2012.718071
Beaumont, C., Protais, J., Guillot, J. F., Colin, P., Proux, K., Millet, N., et al. (1999). Genetic resistance to mortality of day-old chicks and carrier-state of hens after inoculation with Salmonella enteritidis. Avian Pathol. 28, 131–135. doi: 10.1080/03079459994858
Berthelot, F., Beaumont, C., Mompart, F., Girard-Santosuosso, O., Pardon, P., Duchet-Suchaux, M. (1998). Estimated heritability of the resistance to cecal carrier state of Salmonella enteritidis in chickens. Poultry Sci. 77, 797–801. doi: 10.1093/ps/77.6.797
Browning, B. L., Browning, S. R. (2009). A unified approach to genotype imputation and haplotype-phase inference for large data sets of trios and unrelated individuals. Am. J. Hum. Genet. 84, 210–223. doi: 10.1016/j.ajhg.2009.01.005
Calenge, F., Kaiser, P., Vignal, A., Beaumont, C. (2010). Genetic control of resistance to salmonellosis and to Salmonella carrier-state in fowl: a review. Genet. Selection Evol. 42, 11. doi: 10.1186/1297-9686-42-11
Cheng, H. H., Kaiser, P., Lamont, S. J. (2013). Integrated genomic approaches to enhance genetic resistance in chickens. Annu. Rev. Anim. Biosci. 1, 239–260. doi: 10.1146/annurev-animal-031412-103701
Dunnington, E. A., Siegel, P. B. (1984). Thermoregulation in newly hatched chicks. Poult. Sci. 63, 1303–1313. doi: 10.3382/ps.0631303
Evans, S. S., Repasky, E. A., Fisher, D. T. (2015). Fever and the thermal regulation of immunity: the immune system feels the heat. Nat. Rev. Immunol. 15, 335–349. doi: 10.1038/nri3843
Fisher, D. T., Vardam, T. D., Muhitch, J. B., Evans, S. S. (2010). Fine-tuning immune surveillance by fever-range thermal stress. Immunol. Res. 46, 177–188. doi: 10.1007/s12026-009-8122-9
Gao, X., Becker, L. C., Becker, D. M., Starmer, J. D., Province, M. A. (2010). Avoiding the high Bonferroni penalty in genome-wide association studies. Genet. Epidemiol. 34, 100–105. doi: 10.1002/gepi.20430
Girard-Santosuosso, O., Lantier, F., Lantier, I., Bumstead, N., Elsen, J.-M., Beaumont, C. (2002). Heritability of susceptibility to Salmonella enteritidis infection in fowls and test of the role of the chromosome carrying the NRAMP1 gene. Genet. Selection Evol. 34, 211–219. doi: 10.1051/gse:2002004
Hori, T., Nakashima, T., Take, S., Kaizuka, Y., Mori, T., Katafuchi, T. (1991). Immune cytokines and regulation of body temperature, food intake and cellular immunity. Brain Res. Bull. 27, 309–313. doi: 10.1016/0361-9230(91)90117-3
Hutt, F. B., Crawford, R. D. (1960a). On breeding chicks resistant to pullorum disease without exposure thereto. Can. J. Genet. Cytology 2, 357–370. doi: 10.1139/g60-038
Hutt, F. B., Crawford, R. D. (1960b). On breeding chicks resistant to pullorum disease without exposure thereto. Can. J. Genet. Cytol. 2, 357–370. doi: 10.1139/g60-038
Kane, E., Peterson, R. A. (1976). Effects on body temperature produced by micro-injection of Salmonella typhimurium lipopolysaccharide into the third cerebral ventricle of the chicken. Poult. Sci. 55, 1580–1582. doi: 10.3382/ps.0551580
Kang, H. M., Zaitlen, N. A., Wade, C. M., Kirby, A., Heckerman, D., Daly, M. J., et al. (2008). Efficient control of population structure in model organism association mapping. Genetics 178, 1709–1723. doi: 10.1534/genetics.107.080101
Kang, H. M., Sul, J. H., Service, S. K., Zaitlen, N. A., Kong, S., Freimer, N. B., et al. (2010). Variance component model to account for sample structure in genome-wide association studies. Nat. Genet. 42, 348–354. doi: 10.1038/ng.548
Kluger, M. J., Ringler, D. H., Anver, M. R. (1975). Fever and survival. Science 188, 166–168. doi: 10.1126/science.188.4184.166
Lamoreux, W. F., Hutt, F. B. (1939). Variability of body temperature in the normal chick. Poult. Sci. 18, 70–75. doi: 10.3382/ps.0180070
Li, X., Nie, C., Zhang, Z., Wang, Q., Shao, P., Zhao, Q., et al. (2018). Evaluation of genetic resistance to Salmonella Pullorum in three chicken lines. Poult. Sci. 97, 764–769. doi: 10.3382/ps/pex354
Linder, P. (2006). Dead-box proteins: a family affair—active and passive players in RNP-remodeling. Nucleic Acids Res. 34, 4168–4180. doi: 10.1093/nar/gkl468
Lippert, C., Listgarten, J., Liu, Y., Kadie, C. M., Davidson, R. I., Heckerman, D. (2011). FaST linear mixed models for genome-wide association studies. Nat. Methods 8, 833–835. doi: 10.1038/nmeth1681
Loh, P.-R., Tucker, G., Bulik-Sullivan, B. K., Vilhjálmsson, B. J., Finucane, H. K., Salem, R. M., et al. (2015). Efficient Bayesian mixed-model analysis increases association power in large cohorts. Nat. Genet. 47, 284–290. doi: 10.1038/ng3190
Madsen, P., Jensen, J. (2008). A user’s guide to DMU. A package for analyzing multivariate mixed models. Version, 6. doi: 10.1146/annurev-physiol-020518-114546
McGovern, D. P. B., Butler, H., Ahmad, T., Paolucci, M., van Heel, D. A., Negoro, K., et al. (2006). TUCAN (CARD8) genetic variants and inflammatory bowel disease. Gastroenterology 131, 1190–1196. doi: 10.1053/j.gastro.2006.08.008
Morrison, S. F., Nakamura, K. (2019). Central Mechanisms for thermoregulation. Annu. Rev. Physiol. 81, 285–308. doi: 10.1146/annurev-physiol-020518-114546
Pontillo, A., Carvalho, M. S., Kamada, A. J., Moura, R., Schindler, H. C., Duarte, A. J. S., et al. (2013). Susceptibility to mycobacterium tuberculosis infection in HIV-positive patients is associated with CARD8 genetic variant. JAIDSJ. AcquiredImmune DeficiencySyndromes 63, 147. doi: 10.1097/QAI.0b013e31828f93bb
Preußner, M., Goldammer, G., Neumann, A., Haltenhof, T., Rautenstrauch, P., Müller-McNicoll, M., et al. (2017). Body temperature cycles control rhythmic alternative splicing in mammals. Mol. Cell 67, 433–446. e4. doi: 10.1016/j.molcel.2017.06.006
Purcell, S., Neale, B., Todd-Brown, K., Thomas, L., Ferreira, M. A. R., Bender, D., et al. (2007). PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 81, 559–575. doi: 10.1086/519795
Roberts, E. R., Cards, L. R. (1935). Inheritance of resistance to bacterial infection in animals: a genetic study of pullorum disease. Bull. Univ. Ill.Urbana-ChampaignCampusAgric. Exp. doi: 10.1038/nature11930 Stn. No. 419.
Roberts, R. L., Topless, R. K. G., Phipps-Green, A. J., Gearry, R. B., Barclay, M. L., Merriman, T. R. (2010). Evidence of interaction of CARD8 rs2043211 with NALP3 rs35829419 in Crohn’s disease. Genes Immun. 11, 351–356. doi: 10.1038/gene.2010.11
Satoh, T., Kidoya, H., Naito, H., Yamamoto, M., Takemura, N., Nakagawa, K., et al. (2013). Critical role of Trib1 in differentiation of tissue-resident M2-like macrophages. Nature 495, 524–528. doi: 10.1038/nature11930
Scholes, J. C. (1940). “The relationship between body temperature and genetic resistance to Salmonella Pullorum in the fowl,” in Thesis Cornell Univ. Available at: http://www.cabdirect.org/abstracts/19432200530.html [Accessed December 28, 2017]. doi: 10.1186/s12711-015-0176-7
Sund-Levander, M., Forsberg, C., Wahren, L. K. (2002). Normal oral, rectal, tympanic and axillary body temperature in adult men and women: a systematic literature review. ScandinavianJ. CaringSci. 16, 122–128. doi: 10.1046/j.1471-6712.2002.00069.x
Van Goor, A., Bolek, K. J., Ashwell, C. M., Persia, M. E., Rothschild, M. F., Schmidt, C. J., et al. (2015). Identification of quantitative trait loci for body temperature, body weight, breast yield, and digestibility in an advanced intercross line of chickens under heat stress. Genet. SelectionEvol. 47, 96. doi: 10.1186/s12711-015-0176-7
Watanabe, M., Tomiyama-Miyaji, C., Kainuma, E., Inoue, M., Kuwano, Y., Ren, H., et al. (2008). Role of α-adrenergic stimulus in stress-induced modulation of body temperature, blood glucose and innate immunity. Immunol. Lett. 115, 43–49. doi: 10.1016/j.imlet.2007.09.010
Yu, J., Pressoir, G., Briggs, W. H., Vroh Bi, I., Yamasaki, M., Doebley, J. F., et al. (2006). A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nat. Genet. 38, 203–208. doi: 10.1038/ng1702
Yu, Z., Chen, T., Li, X., Yang, M., Tang, S., Zhu, X., et al. (2016). Lys29-linkage of ASK1 by Skp1-Cullin 1-Fbxo21 ubiquitin ligase complex is required for antiviral innate response. Elife 5. doi: 10.7554/eLife.14087
Zhang, Z., Ersoz, E., Lai, C.-Q., Todhunter, R. J., Tiwari, H. K., Gore, M. A., et al. (2010). Mixed linear model approach adapted for genome-wide association studies. Nat. Genet. 42, 355–360. doi: 10.1038/ng.546
Zhou, X., Stephens, M. (2012). Genome-wide efficient mixed-model analysis for association studies. Nat. Genet. 44, 821–824. doi: 10.1038/ng2310
Keywords: Salmonella Pullorum, body temperature, disease resistance, heritability, genome-wide association study, chicken
Citation: Li X, Nie C, Liu Y, Chen Y, Lv X, Wang L, Zhang J, Yang W, Li K, Zheng C, Jia Y, Ning Z and Qu L (2020) The Genetic Architecture of Early Body Temperature and Its Correlation With Salmonella Pullorum Resistance in Three Chicken Breeds. Front. Genet. 10:1287. doi: 10.3389/fgene.2019.01287
Received: 25 July 2019; Accepted: 22 November 2019;
Published: 22 January 2020.
Edited by:
Jesús Fernández, Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria, SpainReviewed by:
Xiaozhu Wang, Auburn University, United StatesCopyright © 2020 Li, Nie, Liu, Chen, Lv, Wang, Zhang, Yang, Li, Zheng, Jia, Ning and Qu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yaxiong Jia, yaxiongjia@163.com; Zhonghua Ning, ningzhh@cau.edu.cn; Lujiang Qu, quluj@163.com
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.