 Xuying Zhang
Xuying Zhang Hermann H. Swalve
Hermann H. Swalve René Pijl3
René Pijl3 Bertram Brenig
Bertram Brenig
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Genet. , 13 November 2019
Sec. Livestock Genomics
Volume 10 - 2019 | https://doi.org/10.3389/fgene.2019.01157
Bovine interdigital hyperplasia (IH) is a typical disease of the foot with varying prevalence depending on age, breed, and environmental factors resulting in different degrees of lameness. In studies based on assessments of claw health status at time of hoof trimming and applying genetic-statistical models to analyze this data, IH consistently exhibits high estimates of heritability in the range of 0.30–0.40. Although some studies have identified chromosomal regions that could possibly harbor causative genes, a clear identification of molecular causes for IH is lacking. While analyzing the large database of claw health status as documented at time of hoof trimming, we identified one herd with extreme prevalence of IH of > 50% of affected Holstein dairy cows. This herd subsequently was chosen as the object of a detailed study. A total of n = 91 cows was assessed and revealed a prevalence of 59.3% and 38.5% for IH cases, documented as “one-sided” or “two-sided”, respectively. Cows were genotyped using the BovineSNP50 BeadChip. A genome wide association study revealed two significantly associated chromosomal positions (-log10P = 5.57) on bovine chromosome 8 (BTA8) located in intron 5 and downstream of the receptor tyrosine kinase-like orphan receptor 2 (ROR2) gene. As ROR2 plays a key role in ossification of the distal limbs and is associated with brachydactylies in humans, it was a reasonable candidate for IH. A comparative sequencing of the ROR2 gene between cases and controls revealed two missense variants in exon 1 (NC_037335.1:g.85,905,534T > A, ARS-UCD1.2) and exon 9 (NC_037335.1:g.86,140,379A > G, ARS-UCD1.2), respectively. Genotyping of both variants in the cohort of 91 cattle showed that the exon 1 variant (rs377953295) remained significantly associated with IH (p < 0.0001) as a risk factor of the disease. This variant resulted in an amino acid exchange (ENSBTAP00000053765.2:p.Trp9Arg) in the N-terminal region of the ROR2 signal peptide which is necessary for proper topology of the polypeptide during translocation. Quantification of ROR2 mRNA and ROR2 protein showed that the variant resulted in a significant suppression of ROR2 expression in homozygous affected compared to wild type and carrier cows.
Foot diseases are a major and increasing health problem in dairy cattle and show rather high prevalence (Gernand et al., 2012). Often over 70% cows in dairy herds experience one or more front or hind claw lesions or deformities during their life (Smits et al., 1992; van der Waaij et al., 2005; Fjeldaas et al., 2006; van der Linde et al., 2010). Due to milk production losses and veterinary costs as well as culling of severe cases, feet and leg are of economical importance (Bruijnis et al., 2010; Schopke et al., 2013). Besides disorders like double sole, sole ulcer, sole hemorrhage, white line separation, and digital dermatitis, interdigital hyperplasia (IH) plays an important role with a prevalence of around 5%–14.2% (Smits et al., 1992; Somers et al., 2003; van der Waaij et al., 2005; van der Linde et al., 2010). Normally IH begins with a small and painless protrusion of the interdigital skin which sometimes can already be diagnosed at calf age. In the final stage clinical signs of IH are firm tumor like masses found in the interdigital space with skin lesions that can result in deep necrosis and phlegmonous inflammations.
Infectious processes are possibly implicated in the pathogenesis of IH. Tissues affected with IH exhibit decreased microbial richness and diversity compared to healthy skin (Bay et al., 2018). An increased susceptibility for IH has been described depending on the number of parities and lactations. From first parity to later parities, IH frequency and estimates of heritability increase (van der Spek et al., 2015). In a longitudinal study in which cows that had been affected with IH were analyzed with respect to the point in time when they were first diagnosed with IH, it was found that about 50% of all cows that were susceptible were affected in their first lactation, another 25% in their second lactation and another 10%–15% in third lactation while the remaining cows only exhibited IH very late in life (Swalve et al., 2017).
Already in very early literature, it has been hypothesized that a genetic predisposition is the main cause for the development of IH (Götze, 1952). In more recent studies using data collected at time of hoof trimming, rather elevated estimates of heritabilities are found (Gernand et al., 2012; van der Spek et al., 2013; Pijl and Swalve, 2017) and these even amount to magnitudes of 0.30–0.40 when threshold models are used or estimates from a linear estimation are converted to the underlying scale (Swalve et al., 2011; Pijl et al., 2013; van der Spek et al., 2013; Perez-Cabal and Charfeddine, 2015). In a study estimating the odds ratio for cows to be affected by IH when comparing the status of female ancestors, it was found that the risk to be affected increases 8.5-fold when comparing cows with affected dams and grand-dams vs. cows with non-affected ancestors (Pijl and Swalve, 2017). Up to recently, evidence for a genetic predisposition on a molecular level has been scarce and inconclusive (Koenig et al., 2005; Gernand et al., 2012; van der Spek et al., 2013). In a very recent study, a number of chromosomal regions putatively involved in the etiology of IH have been identified (Croue et al., 2019).
Aim of the present study was to identify genes that might play a role in the development of IH by means of a genome-wide association study based on a case-control design. As a further step, genes in the identified regions were subject to sequencing for detection of single nucleotide polymorphisms. Finally, polymorphic sites were further analyzed with respect to differences in gene expression. A herd of dairy cows exhibiting an extreme prevalence of IH served as a basis for the study.
Clinical inspections and sampling of cattle were done during routine hoof trimming on farm with written owner consent. Samples were taken exclusively by local veterinarians. The collection of samples was approved by the Lower Saxony State Office for Consumer Protection and Food Safety (33.19-42502-05-17A196) according to §8a Abs. 1 Nr. 2 of the German Animal Protection Law.
Clinical investigations were completely connected to the hoof trimming routine as practiced on the study farm. The hoof trimming routine consisted on visits of the hoof trimmer at intervals of approximately 3 months. At each visit, around half of the cows of the herd were subject to hoof trimming, i.e., two consecutive visits were supposed to cover the entire herd. The herd therefore was inspected at two consecutive hoof trimming events in October, 2016, and January, 2017. Another hoof trimming event in January 2018 was used to take 2mm fine needle biopsies (FNB) (Tru-Punch Sterile Disposable Biopsy Punch, Sklar Instruments, VWR, Germany) from hyperplastic interdigital skin of eight affected cows for RNA and protein analysis. Healthy control samples were collected from an abbatoir.
A total of 110 cattle was assessed at visits 1 (V1) and 2 (V2) (Table 1). Excluding animals that had been presented twice, i.e., at V1 and V2, n = 91 animals remained. During the visits, individuals were inspected visually, phenotypes were recorded and pictures were taken for documentation. Blood samples were taken for DNA extraction during V1 and V2 (n = 91).
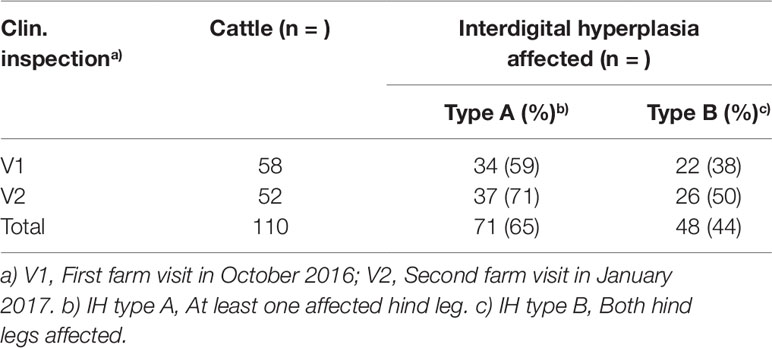
Table 1 Frequency of interdigital hyperplasia (IH) affected cows during two farm visits.
Genotyping of the 91 samples was performed using the Illumina BovineSNP50 BeadChip. Raw data were processed using GenomeStudio V2011.1 (Illumina, San Diego, USA). Final reports were imported into SVS 8.8.3 (Golden Helix, Bozeman, USA) and low quality SNPs were filtered if call rates < 90%, MAF <0.01 and Fisher´s HWE p < 0.0001 (based on controls). LD pruning was performed using default parameters. Samples were filtered with call rates < 0.95. Mitochondrial DNA and sex chromosomes were excluded from the analysis. After filtering 70 samples and 45,232 SNPs remained in the analysis. Genome-wide associations were calculated under an additive and dominant model (Duenk et al., 2017). Associations were regarded as genome-wide statistically significant above a threshold of -log10P = 5.47 (p = 0.05) (Benjamini and Yekutieli, 2001). Associations of markers (-log10P-value, y-axis) were plotted against their chromosomal positions (UMD3.1.1, x-axis).
All n = 91 cows from V1 and V2 were genotyped for rs43572154 and rs377953295. Genotype counts, genotype, and allele frequencies were calculated for both variants. Hardy-Weinberg test was calculated according to Rodriguez et al. (Rodriguez et al., 2009). Frequency distribution for alleles and genotypes vs. types of definitions of disease status (Type A IH, Type B IH) was calculated using both Chi-squared test and Fisher’ s Exact Test.
As the proportion of the genetic variance ideally should be estimated from a large sample to be drawn from the entire population, an approximation was used for the evaluation of the relative importance of each SNP in terms of the variance accounted for. A completely random threshold model was applied using a logit-link function with SNP taken as random and applying PROC Glimmix in SAS. With the residual variance under this model commonly taken as a fixed value of pi²/3 equal to 3.29, the proportion of the variance accounted for by each single SNP was estimated as [SNP variance/(SNP variance + residual variance)] * 100.
Based on the number of cattle analyzed here, a true population wide LD estimation between pairwise SNP cannot be performed. As an approximation, based on the genotypes for putatively functional SNP as identified after sequencing and the respective genotypes for SNP included in the array, a pairwise analysis was done by i) code the genotypes as 0, 1, 2 for both SNP in question and ii) estimate the correlation between the resulting variables and square it.
DNA was extracted from blood samples using MagNA Pure LC DNA Isolation Kit I (Roche Diagnostics, Mannheim, Germany) or a modified salting out procedure (Miller et al., 1988). For Sanger sequencing of ROR2, primers were designed to amplify exons and intron–exon boundaries (Supplementary Table S1) (Sanger et al., 1977). PCR products were purified with Rapid PCR Cleanup Enzyme Set (New England Biolabs GmbH, Frankfurt am Main, Germany) and sequenced using the BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Fisher Scientific GmbH, Schwerte, Germany) on an ABI PRISM 3130xl Genetic Analyzer (Life Technologies, Foster City, United States) according to the manufacturers´ protocols. The same primers were used for sequencing PCR products. SeqMan Pro software [version 15.0.0 (160) Intel, DNASTAR] was used for sequence alignments. Genotyping of variants NC_037335.1:g.85,905,534T > A and NC_037335.1:g.86,140,379A > G (ARS-UCD1.2) was done by diagnostic sequencing. To identify IH associated variants, ROR2 was comparatively sequenced using eight IH affected (IHA) and eight healthy control cattle. Allele and genotype frequencies of variant rs377953295 were determined in 3,093 random Holstein cattle samples.
For RNA and protein analysis samples were collected during the visits at the farm. A total of eight samples (3 x A_A, 3 x A_T, 2 x T_T) were collected for RNA and seven samples (3 x A_A, 3 x A_T, 1 x T_T) for protein analysis corresponding to the genotypes at variant rs77953295. Harvested tissue was immediately immersed in RNAlater (Ambion, ThermoFisher Scientific, Dreieich, Germany) and stored at 4°C. Tissue samples were homogenized in the Qiazol Lysis Reagent (Qiagen, Hilden, Germany). According to the user guide of TRIzol Reagent, total RNA was extracted and eluted in 30 µl RNase-free water and the concentration and purity were measured on a NanoDrop (ThermoFisher Scientific, Dreieich, Germany). 0.2 µg of RNA was converted to cDNA using Maxima H Minus First Strand cDNA Synthesis Kit with dsDNase (ThermoFisher Scientific, Dreieich, Germany).
Protein was extracted and solubilized in an optimized EDTA lysis buffer (Kopec et al., 2017). Protein concentrations were measured using the Bradford assay (Bradford, 1976). RNA and protein samples were stored at –80° until further use.
Potential ROR2 isoforms were analyzed in tissue samples of the interdigital region, lung and spleen of healthy cattle using primer pairs ROR2_cDNA_1/2_fwd (5´-CAGCCCTGTTCCAACTCTGA-3´), ROR2_cDNA_1/2_rev (5´-CCGTATTCCGTCTTGCGGAT-3´), ROR2_cDNA_2_fwd (5´-GGCATGGAGTACCTGTCCAG-3´), and ROR2_cDNA_2_rev (5´-GGCCAGGTCTTTGTGGACCA-3´).
Real-time quantitative PCR was performed using cDNA synthesized from RNA isolated from FNB of IHA cattle and controls. Statistical testing for mRNA expression normalized to GAPDH (glyceraldehyde 3-phosphate dehydrogenase) and β-actin was determined by the 2-ΔΔCT method using Microsoft Excel for Mac 2011 (Livak and Schmittgen, 2001). GAPDH was amplified using primers GAPDH_cDNA_fwd (5´-CCACTCCCAACGTGTCTGTT-3´) and GAPDH_cDNA_rev (5´-GCTTCACCACCTTCTTGATCTCATC-3´) (Macabelli et al., 2014). β-actin was amplified using primers ACTB_cDNA_fwd (5´-GTCATCACCATCGGCAATGAG-3´) and ACTB_cDNA_rev (5´-AATGCCGCAGGATTCCATG-3´) (Huang et al., 2013). Selection of the suitable reference gene was done according to the MIQE guidelines (Bustin et al., 2009). When comparing relative ROR2 expression in 10 different tissue samples of a healthy Holstein cow, GAPDH showed the best invariant expression levels and was used as normalizer. When comparing relative ROR2 expression only in interdigital skin tissue β-actin showed the best invariant expression levels and was therefore used as normalizer.
Protein samples (10 µl) were mixed with 4X Bolt LDS Sample Buffer (Novex, ThermoFisher Scientific, Dreieich, Germany) supplemented with 10% 2-mercaptoethanol, and incubated at 70°C for 10 mins. Electrophoresis was performed on 8% Bolt Bis-Tris Plus Gels (Novex, Thermo Fisher Scientific) in Bolt MES SDS Running Buffer (Novex, ThermoFisher Scientific, Dreieich, Germany) at 165 V for 57 mins. Proteins were transferred to nitrocellulose membrane (ThermoFisher Scientific, Dreieich, Germany) at 15 V for 1 h in transfer buffer. After blocking overnight at 4°C, immunoblots were incubated with primary anti-ROR2 antibody (1:500, ABIN2706970, Cohesion Biosciences, Aachen, Germany) and anti-β-actin (1:10,000, A5441, Sigma Aldrich, Darmstadt, Germany) at room temperature for 1 h. Incubation with the secondary antibodies (1:5000 for #1706515, 1:10,000 for #1706516, Bio-Rad, Munich, Germany) was done at room temperature for 1 h. Immunoblots were developed with Western ECL (GERPN2109, Sigma Aldrich, Darmstadt, Germany). Images from Western blots were captured and quantification was performed with ImageJ software (Schneider et al., 2012).
During V1 and V2 a total of 110 cattle was clinically inspected (Figure 1) and blood samples drawn (Table 1). After removing duplicates an initial set of 91 samples remained. After quality filtering of genotyping data 70 cattle were used for the genome-wide association study. Depending on the type of IH 41 cases/29 controls (type A) or 29 cases/41 controls (type B) were analyzed using an additive and/or dominant genetic model. For both IH types and genetic models markers on BTA8, i.e., ARS-BFGL-NGS-64395, ARS-BFGL-NGS-69582, showed highest -log10P values above a chromosome-wide significance threshold of -log10P = 5.47 (p = 0.05) (Figures 2A, B). ARS-BFGL-NGS-64395 is located in intron 5 and ARS-BFGL-NGS-69582 downstream of the ROR2 gene (tyrosine-protein kinase transmembrane receptor) (Figure 2B). DNA sequence comparison revealed two variants in the coding region of ROR2 in exon 1 (NC_037335.1g.85905534T > A; rs377953295; ARS-UCD1.2) and exon 9 (NC_037335.1g.86140379A > G; rs43572154; ARS-UCD1.2). Estimates of r² for pairwise comparisons of rs377953295 with significant SNP included in the BeadChip were r² = 0.973 (for ARS-BFGL-NGS-64395), r² = 0.920 (for ARS-BFGL-NGS-69582) and r² = 0.652 (for Hapmap39516-BTA-82096). It is especially notable that, with the exception of one animal, rs377953295 and ARS-BFGL-NGS-64395 appeared to be in perfect linkage.
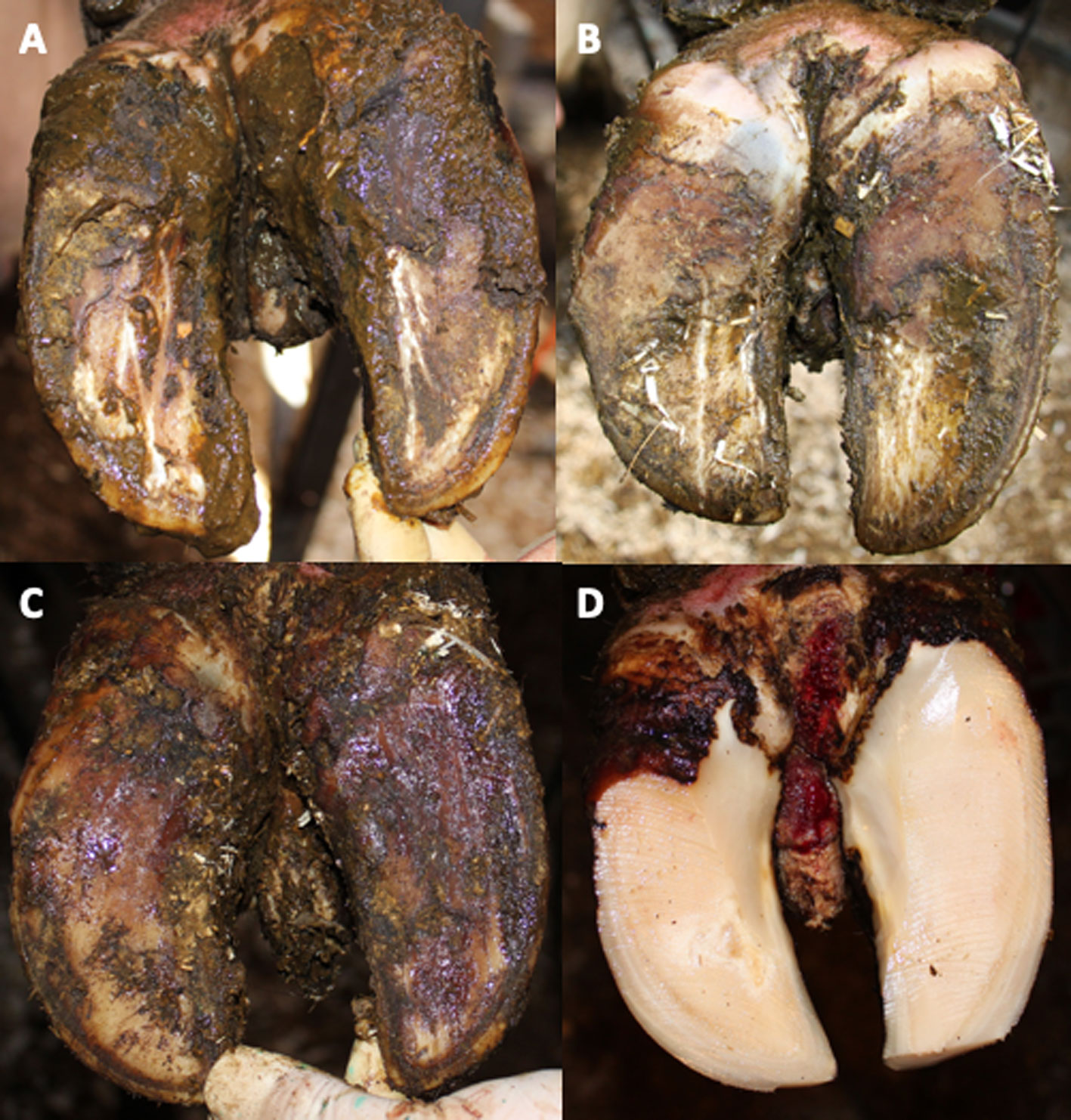
Figure 1 Clinical signs of bovine interdigital hyperplasia. (A–D) show different developmental stages with increasing size of dermal hyperplasia found in the interdigital space with skin final skin lesions (D). (D) Interdigital hyperplasia after hoof trimming.
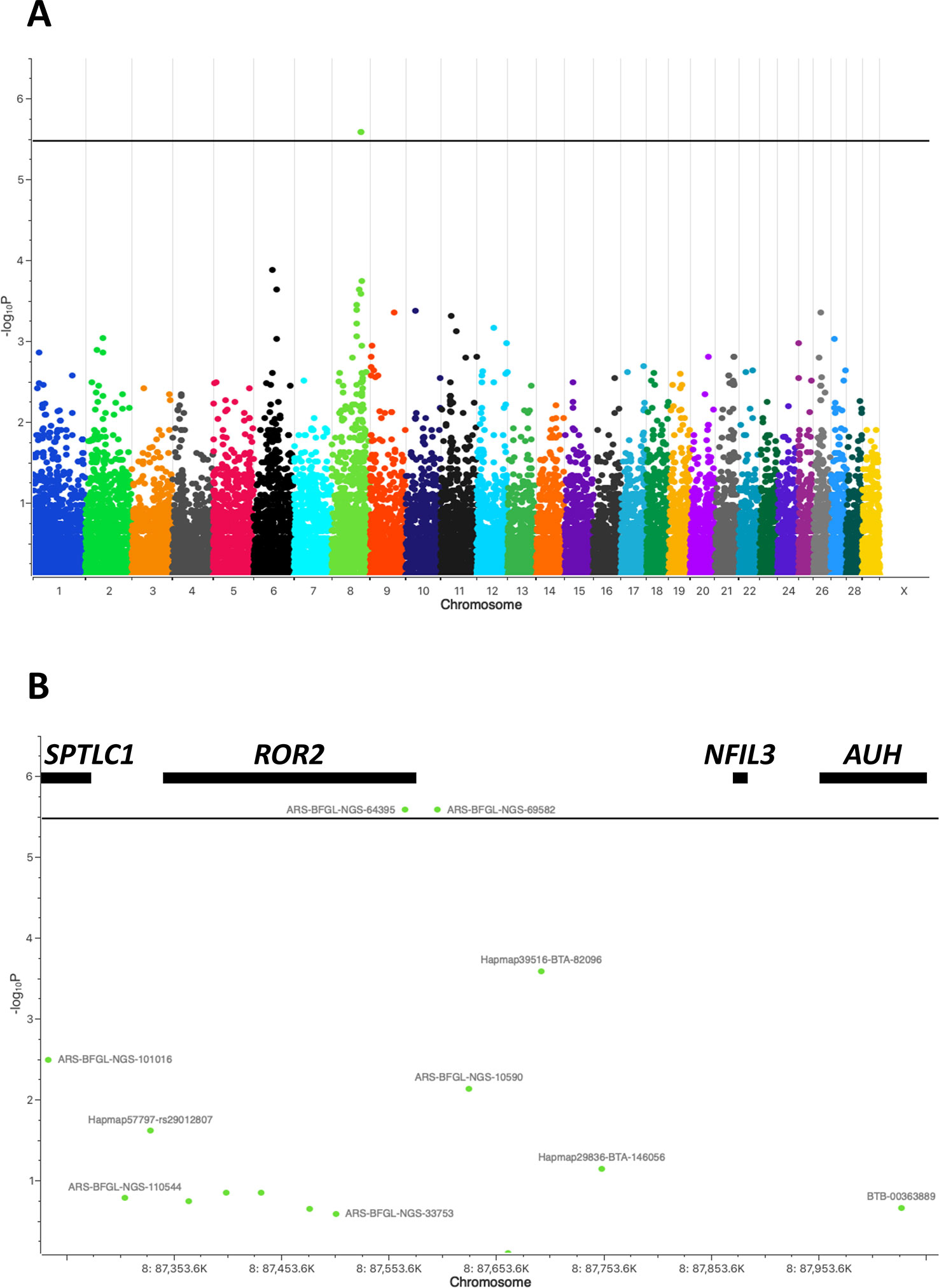
Figure 2 Manhattan plot of genome-wide association study for bovine interdigital hyperplasia. (A) Marker associations are plotted as negative log-transformed P values against the position in the bovine genome (UMD3.1.1). Two markers ARS-BFGL-NGS-64395 and ARS-BFGL-NGS-69582 exceed the genome-wide significance threshold of -log10P = 5.47 (black line). (B) Enlargement of the associated chromosomal region on BTA8 flanking markers ARS-BFGL-NGS-64395 and ARS-BFGL-NGS-69582. Genes located within this region, i.e., SPTLC1, ROR2, NFIL3, and AUH, are shown with black bars at their approximate positions.
The variant in exon 1 resulted in an amino acid exchange from tryptophan to arginine (ENSBTAP00000053765.2:p.Trp9Arg) located in the N-terminal signal peptide. The variant in exon 9 caused an amino acid exchange from methionine to valine (ENSBTAP00000053765.2:p.Met901Val) at the C-terminal intracellular end outside of any functional domain (Rebagay et al., 2012).
To determine the genotype frequencies of rs377953295 (exon 1) and rs43572154 (exon 9) in IHA and control cattle (IHF) both variants were genotyped in all clinically inspected cattle. Results are summarized in Table 2. Although for both SNPs the analyzed cohorts were in Hardy-Weinberg equilibrium, it was striking that the A_A-genotype (rs377953295) was not detected in healthy individuals. Fisher´s exact test using a 2x3 contingency table showed that the genotype distribution for this SNP was significantly deviating from expectation (p = 4.9e-4), whereas the distribution of genotypes for rs43572154 (exon 9) did not deviate. SNP rs43572154 (exon 9) was therefore excluded as potential causative variant. Association of rs377953295 (exon 1) was further investigated comparing the frequency distribution of the three genotypes with the different clinical IH types in a total of 94 cattle. The allele and genotype frequency of rs377953295 was also determined in 3,093 random Holstein cattle samples. The genotype count in this cohort was 2,276 (T_T), 622 (A_T), and 195 (A_A) resulting in allele frequencies of 0.84 (T-allele) and 0.16 (A-allele). The proportion of the total variance accounted for by SNP rs377953295 was estimated to be 32.2% for type A IH and 20.1% for type B IH.
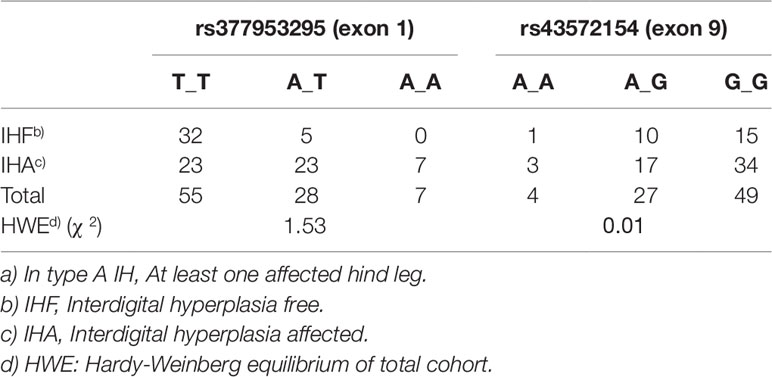
Table 2 Genotype frequencies of the receptor tyrosine kinase-like orphan receptor 2 (ROR2) variants rs377953295 (exon 1) and rs43572154 (exon 9) in interdigital hyperplasia (IH) type Aa) affected and free (= healthy) cattle.
As shown in Table 3 the presence of the A_A-genotype significantly associated with the different clinical forms of IH. Differences between type A and B IH are detectable in the distribution of genotypes T_T and A_T. Furthermore, it is evident that heterozygous individuals are more often affected which suggests that the A-allele at rs377953295 (exon 1) seems to be associated with an increased risk to develop IH.
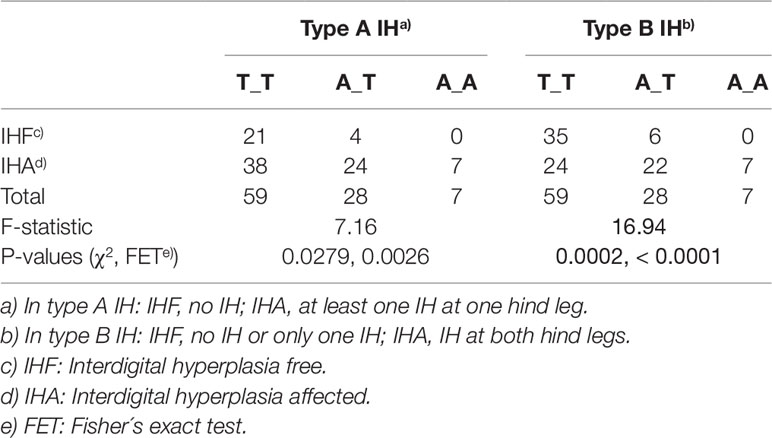
Table 3 Statistical evaluation of SNP rs377953295 (exon 1) as causative variant for type A and type B interdigital hyperplasia (IH).
In summary, the genome-wide association study identified a significantly associated chromosomal region on BTA8 harboring a potential candidate gene with a missense variant that is significantly overrepresented in IHA cattle. As the variant rs377953295 (exon 1) results in an amino acid exchange at position nine of the ROR2 signal peptide it was hypothesized that this could influence expression and finally an insufficient translocation of ROR2 into the plasma membrane.
ROR2 is a classical housekeeping gene harboring a GC-rich and TATA-less promoter (Yella and Bansal, 2017). Therefore, it was expected that ROR2 is widely expressed as shown in humans (Thul and Lindskog, 2018). However, as ROR2 has also been shown to be mainly expressed in early and expression abates during later embryonal development, it was first of all important to demonstrate ROR2 expression in tissues of adult cattle (Matsuda et al., 2001; Yoda et al., 2003). Therefore, 10 different tissues/organs, i.e., interdigital skin, lung, spleen, brain, heart, liver, stomach, intestine, urinary bladder, and bone marrow, were collected at an abattoir and analyzed. As shown in Figure 3A expression was detected at different levels in all analyzed samples. Importantly, ROR2 was expressed in the skin of the interdigital region further supporting the genomic data that it could be involved in the development of IH. In addition, it has been reported that two isoforms of ROR2 (ROR2-201, ROR2-202) exist differing in the usage of two alternative first exons. Only ROR2-201 harbors the exon with variant rs377953295 and therefore it was necessary to prove that this isoform was expressed in the interdigital skin. Using an isoform specific PCR it was possible to show that both isoforms are expressed in the interdigital skin (Figure 3B). Finally, the association of the three genotypes of variant rs377953295 with the expression of ROR2 in the interdigital skin was tested. For this purpose samples of the hyperplastic interdigital skin of cattle harboring the T_T-, A_T- and A_A-genotypes at rs377953295 were compared using interdigital skin of healthy cattle as normalizer. As depicted in Figure 4 the different genotypes within the IHA samples showed significant reductions between genotype T_T vs A_A as well as A_T vs A_A. The difference between T_T vs A_T was not significant. The differences in ROR2 mRNA levels were also reflected in the ROR2 protein levels (Figure 5). Note, that for the protein extraction of the T_T genotype only one hyperplastic skin sample was available and therefore a statistical verification was omitted. For reasons of animal welfare we refrained from additional sample collections. However, the ROR2 protein amount in the T_T sample was clearly above the other two genotypes.
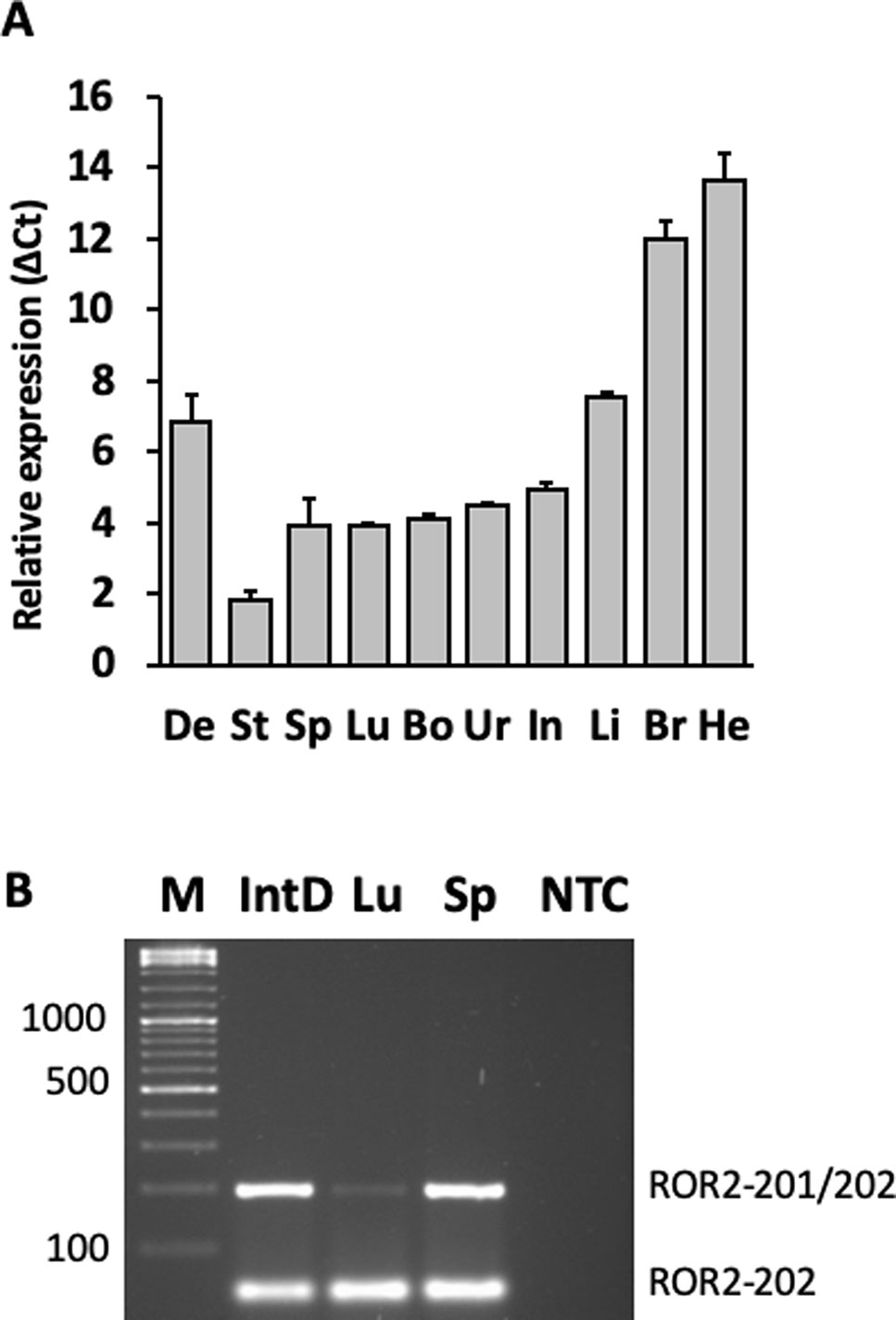
Figure 3 Detection of the receptor tyrosine kinase-like orphan receptor 2 (ROR2) transcripts and isoforms in different bovine organs and tissues. (A) Organ and/or tissue samples (De, interdigital dermis, He, heart, Br, brain, Li, liver, Int, intestine, Ur, urinary bladder, Bo, bone marrow, Lu, lung, Sp, spleen, St, stomach) were collected from healthy cattle and RNA extracted. Relative expression levels were calculated using the average Δ Ct-values of three biological replicates with GAPDH as normalizer. (B)ROR2 isoform specific primers were used to amplify RNA from IntD, interdigital dermis, Lu, lung and Sp, spleen (NTC, non template control). Two amplicons corresponding to isoform ROR2-202 and isoforms ROR2-201/202 together can be seen. Amplicons were separated on a 2% agarose gel and visualized using Ethidium bromide staining.
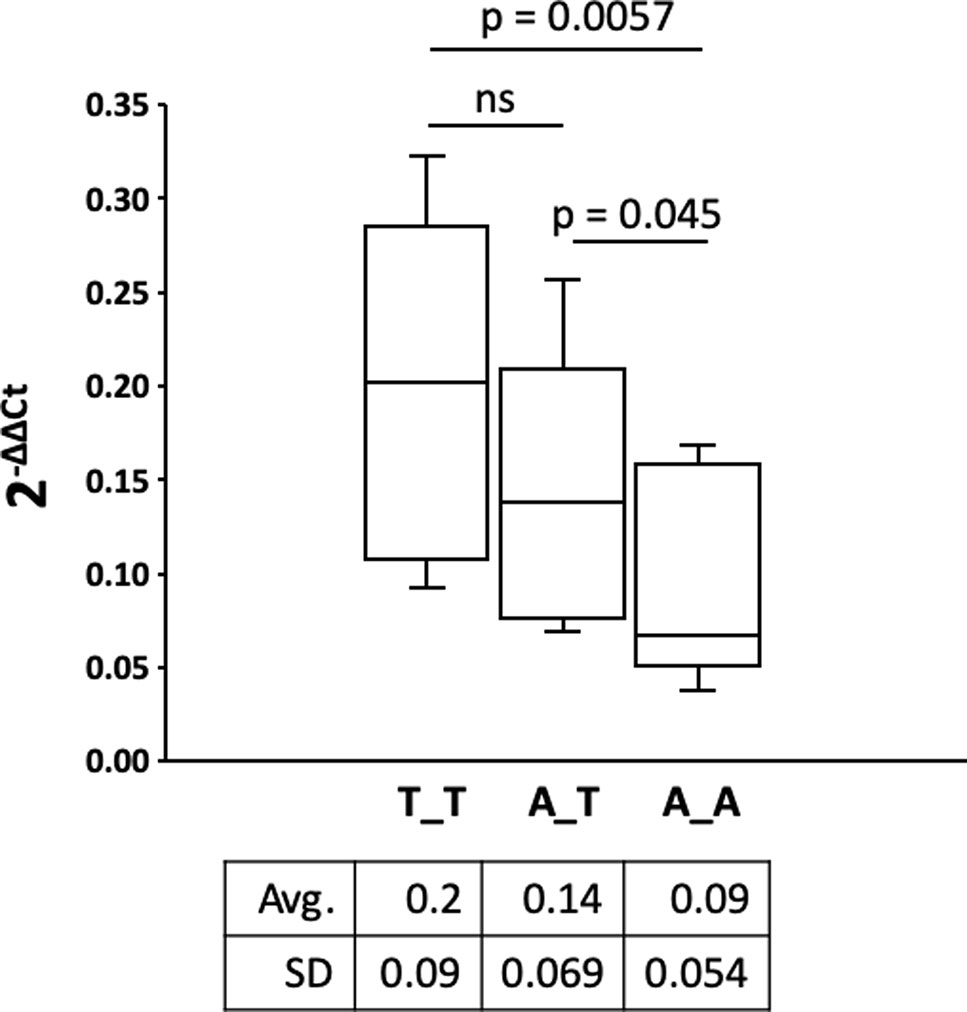
Figure 4 Comparison of the receptor tyrosine kinase-like orphan receptor 2 (ROR2) expression in hyperplastic interdigital skin tissue. Interdigital skin tissues were collected from 3 A_A, 3 A_T, 2 T_T cattle using fine needle biopsies (FNB). Genotypes A_A, A_T and T_T correspond to the missense variant in exon 1 (rs377953295). Two samples were taken from healthy control cattle as reference. Expression of ROR2 and internal control was done by real-time quantitative PCR. Calculation of ROR2 expression fold change in the IH affected cattle was done using the 2-ΔΔCt method with β-actin as normalizer and the average expression of ROR2 in the healthy controls as reference (Livak and Schmittgen, 2001). ROR2 expression fold change of the three genotypes is shown as box-and-whisker plot (whiskers indicate minima and maxima). The vertical line depicts the median. Significance was calculated using a one-tailed t-test with p < 0.05.
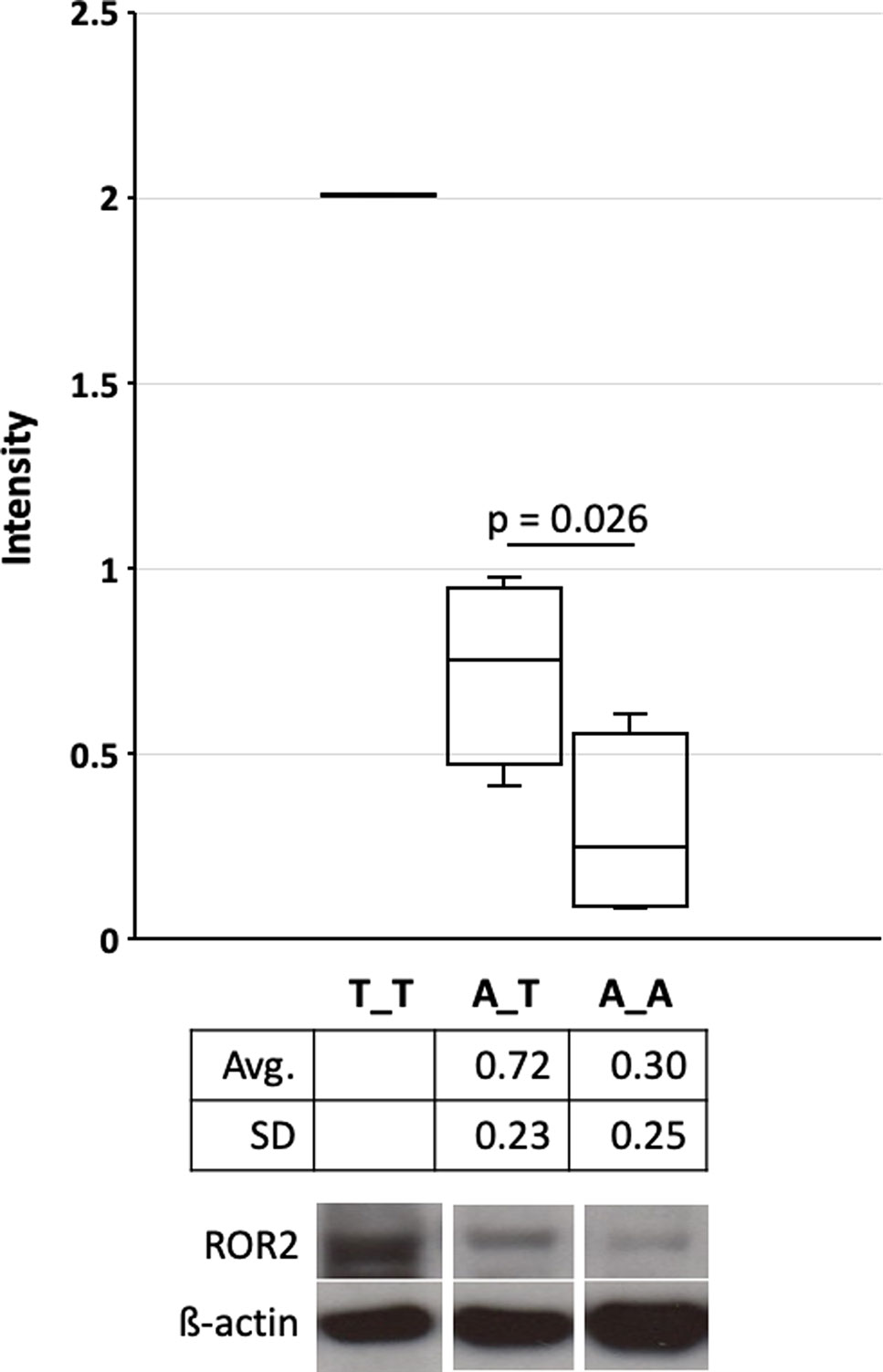
Figure 5 Quantification of the receptor tyrosine kinase-like orphan receptor 2 (ROR2) protein in hyperplastic interdigital skin tissue. Interdigital skin tissues were collected from 3 A_A, 3 A_T, 1 T_T cattle using fine needle biopsies (FNB) (see also Figure 4). Genotypes A_A, A_T and T_T correspond to the missense variant in exon 1 (rs377953295). Two samples were taken from healthy control cattle as reference. Protein extracts were separated on 8% Bolt Bis-Tris Plus gels and transferred to nitrocellulose membranes. Membranes were incubated with primary anti-ROR2 antibody and anti-β-actin. Blots were developed with Western ECL. Images were captured and intensity quantification was performed with ImageJ software (Schneider et al., 2012). Significance was calculated using a one-tailed t-test with p < 0.05. As only one samples for genotype T_T were available statistical significance, average and standard deviation is not depicted. Whiskers indicate minima and maxima. The vertical line depicts the median.
In summary, the functional analyses are in agreement with the genomic data that a reduction of ROR2 expression on RNA and protein level seems to be associated with the risk to develop IH and is highest in cattle with the A_A genotype at rs377953295.
IH is a serious health issue in cattle production. Besides the clinical and animal welfare relevance it also has tremendous effects on the general performance of the animal. Therefore, it is of great importance to elucidate the molecular genetics of IH and find potential causative variants that could be used for selection in future breeding programs.
In the study described here a herd with a high prevalence of IH was identified which was followed over a longer period of time allowing a repeated thorough clinical inspection and sampling during professional hoof trimming. A GWAS was performed showing two genome-wide significantly associated SNP loci at around 87.5Mb on BTA8. In a previous study, 17 suggestive associations (p < 0.20) spreading across the bovine genome had been detected for IH in Holstein cattle (van der Spek et al., 2015). Although five of them were also located on BTA8 (8Mb, 24Mb, 25MB, 43Mb), the IH associated region in this study was located further downstream. Fortunately, only four genes were located in the direct chromosomal proximity of the associated SNPs (Figure 2B). Genes flanking ROR2, i.e., SPTLC1 (long-chain base subunit 1 of serine palmitoyltransferase) (Suh et al., 2014; Ho and Jerath, 2018), NFIL3 (interleukin 3-regulated nuclear factor) (Kobayashi et al., 2011; Kobayashi et al., 2014), and AUH (3-methylglutaconyl-CoA hydratase) (Ijlst et al., 2002; Ly et al., 2003) were excluded as potential candidates as they have been reported in humans or mice to be causative for hereditary sensory and autonomic neuropathy (type 1A), susceptibility to inflammatory bowel disease and 3-methylglutaconic aciduria (type 1), respectively. On the other hand, as ROR2 had been associated with terminal limb malformations in humans including cutaneous syndactyly, it seemed to be a reasonable candidate for further analysis (Oldridge et al., 2000; Schwabe et al., 2000; Afzal et al., 2000; van Bokhoven et al., 2000; Bacchelli et al., 2003; Tufan et al., 2005; Ali et al., 2007; Brunetti-Pierri et al., 2008). Additionally, one of the associated SNPs was directly located within the gene (intron 5). A DNA sequence comparison of ROR2 in IHA and healthy cattle revealed two missense variants in exon 1 and exon 9, respectively. However, only the variant in exon 1 (rs377953295) resulting in an amino acid exchange in the signal peptide of ROR2 (ENSBTAP00000053765.2:p.Trp9Arg) remained significantly associated with IH after screening the cohort of cattle at the farm. So far ROR2 variants have not been associated with any other disorder or trait in cattle and therefore the identification of the missense variant was a novel finding.
But also regarding its biological function, ROR2 was obviously an interesting candidate. ROR2 belongs to the receptor tyrosine kinases (RTK), a large superfamily of transmembrane glycoproteins. Previous studies have shown that ROR2 is important for the formation of the distal limbs (Takeuchi et al., 2000; Matsuda et al., 2001). Molecular genetic analyses in humans have revealed that mutations in ROR2 cause dominant Brachydactyly type B and recessive Robinow Syndrome, with terminal limb malformations as common symptoms (Oldridge et al., 2000; Schwabe et al., 2000; Afzal et al., 2000; van Bokhoven et al., 2000; Bacchelli et al., 2003; Tufan et al., 2005; Ali et al., 2007; Brunetti-Pierri et al., 2008; Kjaer et al., 2009). Defects of the distal limbs have also been observed in ROR2-/- mice, mainly due to abnormal cartilage and growth plate development, as well as ossification (Takeuchi et al., 2000; DeChiara et al., 2000). Hence, the reported functional role, determined positional association and identification of a missense variant in ROR2 in IHA cattle were convincing enough to extend the analyses.
Therefore, ROR2 expression was analyzed in the interdigital skin. Although earlier studies indicated that ROR2 expression in adults was restricted to parathyroid, testis, and uterus, it is now known that its expression may be more widespread than originally thought (Katoh and Katoh, 2005; Thul and Lindskog, 2018). During embryogenesis ROR2 expression was identified in heart, lungs, face, and limbs (Matsuda et al., 2001). Our data show that different amounts of ROR2 mRNA can be detected in interdigital skin, lung, spleen, brain, heart, liver, stomach, intestine, urinary bladder, and bone marrow of adult cattle. In addition, two ROR2 isoforms (ROR2-201, ROR2-202) were detected differing in the usage of an alternative first exon. Especially isoform ROR2-201 harboring the exon with the missense variant was detectable in the interdigital skin. Although compared to other organs and/or tissues expression of ROR2 in the interdigital skin was intermediate, the identification of ROR2 transcripts was important to support its assumed role in IH development. Furthermore, the relative amount of ROR2 transcripts significantly differed between the three genotypes in correlation with the IH status. Homozygous A_A IHA cattle showed significantly reduced ROR2 expression in the hyperplastic interdigital skin, which is also reflected in the amount of ROR2 protein. In addition, ROR2 expression in hyperplastic compared to normal interdigital skin showed a significant reduction, which explains the observation that hyperplastic skin alterations were also present in T_T or A_T cattle. The down-regulation of ROR2 could be explained by a general mechanism of transcriptional quality control. For instance, when mutant signal sequences fail to bind to the signal recognition particle at the ribosome exit site, the nascent chain instead contacts Argonaute2 and the mutant mRNAs are specifically degraded (Karamyshev et al., 2014). Specific mRNA degradation preemptively regulates aberrant protein production (Karamyshev et al., 2014). However, other mechanisms resulting in the general down-regulation of ROR2 in hyperplastic interdigital skin have to be taken into account (Roman-Gomez et al., 2007; Lara et al., 2010).
In the etiology of IH, the role of ROR2 in signal transduction seems to be important. The main histological alterations in hyperplastic interdigital skin are proliferating fibroblasts, multiplex papilliferous epidermal ridges as well as increased cellularity in the stratum granulosum and stratum spinosum (Amstel and Shearer, 2006; Kashyap et al., 2017). In this respect, it is important to note that Wnt signaling pathways play crucial roles in the regulation of skin development and epidermal stem cells behavior (Clevers et al., 2014; Veltri et al., 2018). Sustained epidermal activation of Wnt/β-catenin signaling not only stimulates fibroblast proliferation, but also causes structural remodeling of the entire dermis (Collins et al., 2011). Binding of Wnt isoforms to Fz-LRP complex (frizzled-low density lipoprotein receptor-related protein) generates β-catenin signaling, whereas binding to the atypical receptor tyrosine kinase ROR2 can inhibit this activity (Gordon and Nusse, 2006). The significant down-regulation of ROR2 could affect the inhibition of the canonical Wnt pathway, resulting in the abnormal cellular processes related to aberrant epidermal development. This potential biochemical effect is consistent with our finding that ROR2 seems to be an IH suppressor and that decreased expression level of ROR2 leads to IH development. Noteworthy, disease suppressive relationships of ROR2 have been described in hepatocellular cancer, colon cancer, and hematological malignancies (Roman-Gomez et al., 2007; Lara et al., 2010; Yuan et al., 2011; Geng et al., 2012). The canonical Wnt pathway has a pro-tumorigenic effect, leading to a series of cellular processes including proliferation, differentiation, polarity, migration, invasion, adhesion, and survival (Chien et al., 2009; Ford et al., 2013). Through inhibiting this canonical Wnt signaling as a gatekeeper, ROR2 has been proposed to play a role in tumor suppression (Ford et al., 2013). However, further studies will be required to clarify the exact molecular mechanism caused by down-regulation of ROR2 expression in IH development.
Publicly available datasets were analyzed in this study. This data can be found here: https://www.ncbi.nlm.nih.gov/genome/gdv/browser/?context=genome&acc=GCF_002263795.1, https://www.ensembl.org/Bos_taurus/Gene/Summary?g=ENSBTAG00000005092;r=8:85905346-86141520;t=ENSBTAT00000061589, https://www.proteinatlas.org, http://www.ncbi.nlm.nih.gov/sites/entrez?db=OMIM, http://www.informatics.jax.org/marker/MGI:1347521.
The animal study was reviewed and approved by Lower Saxony State Office for Consumer Protection and Food Safety (33.19-42502-05-17A196). Written informed consent was obtained from the owners for the participation of their animals in this study.
HS and BB conceived and designed the overall study. XZ carried out the molecular experiments. HS, FR, and RP performed the clinical inspections and sample collection at the farm. XZ, HS, MW-D, and BB carried out the bioinformatics analysis. XZ and BB wrote the initial draft and subsequently all authors contributed to the presentation of the results. All authors read and confirmed the final version of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors are grateful to M. Hirschfeld, D. Niehaus, S. Bierau, S. Balzer, L. Jüttner, M. Scharfenstein, G. Kopke, M. Neubert, and S. Pach for expert technical assistance. The Association for Bioeconomy Research (Bonn), German Livestock Association (E. Feddersen), and VIT (F. Reinhardt) are thanked for support. XZ is a fellow of the Chinese Scholarship Council (CSC). We acknowledge support by the German Research Foundation and the Open Access Publications of the University of Göttingen.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2019.01157/full#supplementary-material
Afzal, A. R., Rajab, A., Fenske, C. D., Oldridge, M., Elanko, N., Ternes-Pereira, E., et al. (2000). Recessive Robinow syndrome, allelic to dominant brachydactyly type B, is caused by mutation of ROR2. Nat. Genet. 25, 419–422. doi: 10.1038/78107
Ali, B. R., Jeffery, S., Patel, N., Tinworth, L. E., Meguid, N., Patton, M. A., et al. (2007). Novel Robinow syndrome causing mutations in the proximal region of the frizzled-like domain of ROR2 are retained in the endoplasmic reticulum. Hum. Genet. 122, 389–395. doi: 10.1007/s00439-007-0409-0
Amstel, S. v., Shearer, J. (2006). Manual for treatment and control of lameness in cattle. (Ames, Iowa, USA: Blackwell Publishing Professional). doi: 10.1002/9780470344576
Bacchelli, C., Wilson, L. C., Cook, J. A., Winter, R. M., Goodman, F. R. (2003). ROR2 is mutated in hereditary brachydactyly with nail dysplasia, but not in Sorsby syndrome. Clin. Genet. 64, 263–265. doi: 10.1034/j.1399-0004.2003.00139.x
Bay, V., Griffiths, B., Carter, S., Evans, N. J., Lenzi, L., Bicalho, R. C., et al. (2018). 16S rRNA amplicon sequencing reveals a polymicrobial nature of complicated claw horn disruption lesions and interdigital phlegmon in dairy cattle. Sci. Rep. 8, 15529. doi: 10.1038/s41598-018-33993-9
Benjamini, Y., Yekutieli, D. (2001). The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 29, 1165–1188. doi: 10.1214/aos/1013699998
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. doi: 10.1006/abio.1976.9999
Bruijnis, M. R., Hogeveen, H., Stassen, E. N. (2010). Assessing economic consequences of foot disorders in dairy cattle using a dynamic stochastic simulation model. J. Dairy Sci. 93, 2419–2432. doi: 10.3168/jds.2009-2721
Brunetti-Pierri, N., Del Gaudio, D., Peters, H., Justino, H., Ott, C. E., Mundlos, S., et al. (2008). Robinow syndrome: phenotypic variability in a family with a novel intragenic ROR2 mutation. Am. J. Med. Genet. A 146A, 2804–2809. doi: 10.1002/ajmg.a.32530
Bustin, S. A., Benes, V., Garson, J. A., Hellemans, J., Huggett, J., Kubista, M., et al. (2009). The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 55, 611–622. doi: 10.1373/clinchem.2008.112797
Chien, A. J., Conrad, W. H., Moon, R. T. (2009). A Wnt Survival Guide: From Flies to Human Disease. J. Invest. Dermatol. 129, 1614–1627. doi: 10.1038/jid.2008.445
Clevers, H., Loh, K. M., Nusse, R. (2014). An integral program for tissue renewal and regeneration: Wnt signaling and stem cell control. Sci. 346, 54–5+. doi: 10.1126/science.1248012
Collins, C. A., Kretzschmar, K., Watt, F. M. (2011). Reprogramming adult dermis to a neonatal state through epidermal activation of beta-catenin. Dev. 138, 5189–5199. doi: 10.1242/dev.064592
Croue, I., Michenet, A., Leclerc, H., Ducrocq, V. (2019). Genomic analysis of claw lesions in Holstein cows: opportunities for genomic selection, quantitative trait locus detection, and gene identification. J. Dairy Sci. 102, 6306–6318. doi: 10.3168/jds.2018-15979
DeChiara, T. M., Kimble, R. B., Poueymirou, W. T., Rojas, J., Masiakowski, P., Valenzuela, D. M., et al. (2000). Ror2, encoding a receptor-like tyrosine kinase, is required for cartilage and growth plate development. Nat. Genet. 24, 271–274. doi: 10.1038/73488
Duenk, P., Calus, M. P. L., Wientjes, Y. C. J., Bijma, P. (2017). Benefits of Dominance over Additive Models for the Estimation of Average Effects in the Presence of Dominance. (Bethesda) 7, 3405–3414. doi: 10.1534/g3.117.300113
Fjeldaas, T., Sogstad, A. M., Osteras, O. (2006). Claw trimming routines in relation to claw lesions, claw shape and lameness in Norwegian dairy herds housed in tie stalls and free stalls. Prev. Vet. Med. 73, 255–271. doi: 10.1016/j.prevetmed.2005.09.004
Ford, C. E., Ma, S. S. Q., Quadir, A., Ward, R. L. (2013). The dual role of the novel Wnt receptor tyrosine kinase, ROR2, in human carcinogenesis. Int. J. Cancer 133, 779–787. doi: 10.1002/ijc.27984
Götze, R. (1952). Praktische Hinweise zur Erkennung der Erbgesundheit und Erbfruchtbarkeit aus dem Erscheinungsbild des Zuchtbullen. Tierärztl. Umsch. 7, 466–474.
Geng, M., Cao, Y. C., Chen, Y. J., Jiang, H., Bi, L. Q., Liu, X. H. (2012). Loss of Wnt5a and Ror2 protein in hepatocellular carcinoma associated with poor prognosis. World J. Gastroentero. 18, 1328–1338. doi: 10.3748/wjg.v18.i12.1328
Gernand, E., Rehbein, P., von Borstel, U. U., Konig, S. (2012). Incidences of and genetic parameters for mastitis, claw disorders, and common health traits recorded in dairy cattle contract herds. J. Dairy Sci. 95, 2144–2156. doi: 10.3168/jds.2011-4812
Gordon, M. D., Nusse, R. (2006). Wnt signaling: multiple pathways, multiple receptors, and multiple transcription factors. J. Biol. Chem. 281, 22429–22433. doi: 10.1074/jbc.R600015200
Ho, K. W. D., Jerath, N. U. (2018). V144D Mutation of SPTLC1 Can Present with Both Painful and Painless Phenotypes in Hereditary Sensory and Autonomic Neuropathies Type I. Case Rep. Genet. 2018, 1898151. doi: 10.1155/2018/1898151
Huang, Y. Z., Li, M. X., Wang, J., Zhan, Z. Y., Sun, Y. J., Sun, J. J., et al. (2013). A 5'-regulatory region and two coding region polymorphisms modulate promoter activity and gene expression of the growth suppressor gene ZBED6 in cattle. PloS One 8, e79744. doi: 10.1371/journal.pone.0079744
Ijlst, L., Loupatty, F. J., Ruiter, J. P., Duran, M., Lehnert, W., Wanders, R. J. (2002). 3-Methylglutaconic aciduria type I is caused by mutations in AUH. Am. J. Hum. Genet. 71, 1463–1466. doi: 10.1086/344712
Karamyshev, A. L., Patrick, A. E., Karamysheva, Z. N., Griesemer, D. S., Hudson, H., Tjon-Kon-Sang, S., et al. (2014). Inefficient SRP interaction with a nascent chain triggers a mRNA quality control pathway. Cell 156, 146–157. doi: 10.1016/j.cell.2013.12.017
Kashyap, D. K., Giri, D. K., Dewangan, G. (2017). Interdigital Fibroma in Fore Limb of a Male Buffalo: A Case Report. Buffalo Bull. 36, 561–564.
Katoh, M., Katoh, M. (2005). Comparative genomics on ROR1 and ROR2 orthologs. Oncol. Rep. 14, 1381–1384. doi: 10.3892/or.14.5.1381
Kjaer, K. W., Tiner, M., Cingoz, S., Karatosun, V., Tommerup, N., Mundlos, S., et al. (2009). A novel subtype of distal symphalangism affecting only the 4th finger. Am. J. Med. Genet. A 149A, 1571–1573. doi: 10.1002/ajmg.a.32905
Kobayashi, T., Matsuoka, K., Sheikh, S. Z., Elloumi, H. Z., Kamada, N., Hisamatsu, T., et al. (2011). NFIL3 is a regulator of IL-12 p40 in macrophages and mucosal immunity. J. Immunol. 186, 4649–4655. doi: 10.4049/jimmunol.1003888
Kobayashi, T., Steinbach, E. C., Russo, S. M., Matsuoka, K., Nochi, T., Maharshak, N., et al. (2014). NFIL3-deficient mice develop microbiota-dependent, IL-12/23-driven spontaneous colitis. J. Immunol. 192, 1918–1927. doi: 10.4049/jimmunol.1301819
Koenig, S., Sharifi, A. R., Wentrot, H., Landmann, D., Eise, M., Simianer, H. (2005). Genetic parameters of claw and foot disorders estimated with logistic models. J. Dairy Sci. 88, 3316–3325. doi: 10.3168/jds.S0022-0302(05)73015-0
Kopec, A. M., Rivera, P. D., Lacagnina, M. J., Hanamsagar, R., Bilbo, S. D. (2017). Optimized solubilization of TRIzol-precipitated protein permits Western blotting analysis to maximize data available from brain tissue. J. Neurosci. Methods 280, 64–76. doi: 10.1016/j.jneumeth.2017.02.002
Lara, E., Calvanese, V., Huidobro, C., Fernandez, A. F., Moncada-Pazos, A., Obaya, A. J., et al. (2010). Epigenetic repression of ROR2 has a Wnt-mediated, pro-tumourigenic role in colon cancer. Mol. Cancer 9, 170. doi: 10.1186/1476-4598-9-170
Livak, K. J., Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Ly, T. B., Peters, V., Gibson, K. M., Liesert, M., Buckel, W., Wilcken, B., et al. (2003). Mutations in the AUH gene cause 3-methylglutaconic aciduria type I. Hum. Mutat. 21, 401–407. doi: 10.1002/humu.10202
Macabelli, C. H., Ferreira, R. M., Gimenes, L. U., de Carvalho, N. A., Soares, J. G., Ayres, H., et al. (2014). Reference gene selection for gene expression analysis of oocytes collected from dairy cattle and buffaloes during winter and summer. PloS One 9, e93287. doi: 10.1371/journal.pone.0093287
Matsuda, T., Nomi, M., Ikeya, M., Kani, S., Oishi, I., Terashima, T., et al. (2001). Expression of the receptor tyrosine kinase genes, Ror1 and Ror2, during mouse development. Mech. Dev. 105, 153–156. doi: 10.1016/S0925-4773(01)00383-5
Miller, S. A., Dykes, D. D., Polesky, H. F. (1988). A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 16, 1215. doi: 10.1093/nar/16.3.1215
Oldridge, M., Fortuna, A. M., Maringa, M., Propping, P., Mansour, S., Pollitt, C., et al. (2000). Dominant mutations in ROR2, encoding an orphan receptor tyrosine kinase, cause brachydactyly type B. Nat. Genet. 24, 275–278. doi: 10.1038/73495
Perez-Cabal, M. A., Charfeddine, N. (2015). Models for genetic evaluations of claw health traits in Spanish dairy cattle. J. Dairy Sci. 98, 8186–8194. doi: 10.3168/jds.2015-9562
Pijl, R., Swalve, H. H. (2017). Proceedings of the 19th International Symposium and 11th Conference Lameness in Ruminants, Munich, Germany in Incidence Rates For Claw Disorders In Relation To Status Of Female Ancestors Based On Assessments At Routine Hoof Trimming In German Holstein Dairy Cattle. Eds. Fiedler, A., Schindhelm, K., 297.
Pijl, R., Alkhoder, H., Swalve, H. H. (2013). Proceedings of the 17th International Symposium and 9th International Conference on Lameness in Ruminants in Environmental effects and a genetic predisposition influence Interdigital Hyperplasia. Eds. Whay, B., Hockenhull, J. (Bristol, UK), 65.
Rebagay, G., Yan, S., Liu, C., Cheung, N. K. (2012). ROR1 and ROR2 in Human Malignancies: Potentials for Targeted Therapy. Front. Oncol. 2, 34. doi: 10.3389/fonc.2012.00034
Rodriguez, S., Gaunt, T. R., Day, I. N. (2009). Hardy-Weinberg equilibrium testing of biological ascertainment for Mendelian randomization studies. Am. J. Epidemiol. 169, 505–514. doi: 10.1093/aje/kwn359
Roman-Gomez, J., Jimenez-Velasco, A., Cordeu, L., Vilas-Zornoza, A., Jose-Eneriz, E. S., Garate, L., et al. (2007). WNT5A, a putative tumour suppressor of lymphoid malignancies, is inactivated by aberrant methylation in acute lymphoblastic leukaemia. Eur. J. Cancer 43, 2736–2746. doi: 10.1016/j.ejca.2007.10.004
Sanger, F., Nicklen, S., Coulson, A. R. (1977). DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. U. S. A 74, 5463–5467. doi: 10.1073/pnas.74.12.5463
Schneider, C. A., Rasband, W. S., Eliceiri, K. W. (2012). NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675. doi: 10.1038/nmeth.2089
Schopke, K., Weidling, S., Pijl, R., Swalve, H. H. (2013). Relationships between bovine hoof disorders, body condition traits, and test-day yields. J. Dairy Sci. 96, 679–689. doi: 10.3168/jds.2012-5728
Schwabe, G. C., Tinschert, S., Buschow, C., Meinecke, P., Wolff, G., Gillessen-Kaesbach, G., et al. (2000). Distinct mutations in the receptor tyrosine kinase gene ROR2 cause brachydactyly type B. Am. J. Hum. Genet. 67, 822–831. doi: 10.1086/303084
Smits, M. C. J., Frankena, K., Metz, J. H. M., Noordhuizen, J. P. T. M. (1992). Prevalence of digital disorders in zero-grazing dairy-cows. Livest. Prod. Sci. 32, 231–244. doi: 10.1016/S0301-6226(12)80004-2
Somers, J. G., Frankena, K., Noordhuizen-Stassen, E. N., Metz, J. H. (2003). Prevalence of claw disorders in Dutch dairy cows exposed to several floor systems. J. Dairy Sci. 86, 2082–2093. doi: 10.3168/jds.S0022-0302(03)73797-7
Suh, B. C., Hong, Y. B., Nakhro, K., Nam, S. H., Chung, K. W., Choi, B. O. (2014). Early-onset severe hereditary sensory and autonomic neuropathy type 1 with S331F SPTLC1 mutation. Mol. Med. Rep. 9, 481–486. doi: 10.3892/mmr.2013.1808
Swalve, H. H., Alkhoder, H., Pijl, R. (2011). 8th Conference on Lameness in Ruminants in Genetic background of disorders of the bovine hoof from data collected at hoof trimming(New Zealand).
Swalve, H. H., Kopke, G., Pijl, R., Mülling, C. K. W. (2017). Proceedings of the 19th International Symposium and 11th Conference Lameness in Ruminants, Munich, Germany in Interdigital Hyperplasia In Holstein Cows: A Case Study Of A Farm With High Prevalence. Eds. Fiedler, A., Schindhelm, K., 122.
Takeuchi, S., Takeda, K., Oishi, I., Nomi, M., Ikeya, M., Itoh, K., et al. (2000). Mouse Ror2 receptor tyrosine kinase is required for the heart development and limb formation. Genes Cells 5, 71–78. doi: 10.1046/j.1365-2443.2000.00300.x
Thul, P. J., Lindskog, C. (2018). The human protein atlas: A spatial map of the human proteome. Protein Sci. 27, 233–244. doi: 10.1002/pro.3307
Tufan, F., Cefle, K., Turkmen, S., Turkmen, A., Zorba, U., Dursun, M., et al. (2005). Clinical and molecular characterization of two adults with autosomal recessive Robinow syndrome. Am. J. Med. Genet. A 136, 185–189. doi: 10.1002/ajmg.a.30785
van Bokhoven, H., Celli, J., Kayserili, H., van Beusekom, E., Balci, S., Brussel, W., et al. (2000). Mutation of the gene encoding the ROR2 tyrosine kinase causes autosomal recessive Robinow syndrome. Nat. Genet. 25, 423–426. doi: 10.1038/78113
van der Linde, C., de Jong, G., Koenen, E. P., Eding, H. (2010). Claw health index for Dutch dairy cattle based on claw trimming and conformation data. J. Dairy Sci. 93, 4883–4891. doi: 10.3168/jds.2010-3183
van der Spek, D., van Arendonk, J. A., Vallee, A. A., Bovenhuis, H. (2013). Genetic parameters for claw disorders and the effect of preselecting cows for trimming. J. Dairy Sci. 96, 6070–6078. doi: 10.3168/jds.2013-6833
van der Spek, D., van Arendonk, J. A., Bovenhuis, H. (2015). Genome-wide association study for claw disorders and trimming status in dairy cattle. J. Dairy Sci. 98, 1286–1295. doi: 10.3168/jds.2014-8302
van der Spek, D., van Arendonk, J. A. M., Bovenhuis, H. (2015). Genetic relationships between claw health traits of dairy cows in different parities, lactation stages, and herds with different claw disorder frequencies. J. Dairy Sci. 98, 6564–6571. doi: 10.3168/jds.2015-9561
van der Waaij, E. H., Holzhauer, M., Ellen, E., Kamphuis, C., de Jong, G. (2005). Genetic parameters for claw disorders in Dutch dairy cattle and correlations with conformation traits. J. Dairy Sci. 88, 3672–3678. doi: 10.3168/jds.S0022-0302(05)73053-8
Veltri, A., Lang, C., Lien, W. H. (2018). Concise Review: Wnt Signaling Pathways in Skin Development and Epidermal Stem Cells. Stem Cells 36, 22–35. doi: 10.1002/stem.2723
Yella, V. R., Bansal, M. (2017). DNA structural features of eukaryotic TATA-containing and TATA-less promoters. FEBS Open Bio 7, 324–334. doi: 10.1002/2211-5463.12166
Yoda, A., Oishi, I., Minami, Y. (2003). Expression and function of the Ror-family receptor tyrosine kinases during development: lessons from genetic analyses of nematodes, mice, and humans. J. Recept. Signal Transduct. Res. 23, 1–15. doi: 10.1081/RRS-120018757
Keywords: Claw disease, cattle, interdigital hyperplasia, limax, tyloma, receptor tyrosine kinase-like orphan receptor 2
Citation: Zhang X, Swalve HH, Pijl R, Rosner F, Wensch-Dorendorf M and Brenig B (2019) Interdigital Hyperplasia in Holstein Cattle Is Associated With a Missense Mutation in the Signal Peptide Region of the Tyrosine-Protein Kinase Transmembrane Receptor Gene. Front. Genet. 10:1157. doi: 10.3389/fgene.2019.01157
Received: 13 August 2019; Accepted: 22 October 2019;
Published: 13 November 2019.
Edited by:
Jiuzhou Song, University of Maryland, College Park, United StatesReviewed by:
Bjørg Heringstad, Norwegian University of Life Sciences, NorwayCopyright © 2019 Zhang, Swalve, Pijl, Rosner, Wensch-Dorendorf and Brenig. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bertram Brenig, YmJyZW5pZ0Bnd2RnLmRl
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.