 Tong Wang1†
Tong Wang1† Hui Gao
Hui Gao Wei Li
Wei Li Chao Liu
Chao Liu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Genet., 08 October 2019
Sec. Epigenomics and Epigenetics
Volume 10 - 2019 | https://doi.org/10.3389/fgene.2019.00962
This article is part of the Research TopicEpigenetic Biomarker and Personalized Precision MedicineView all 33 articles
Spermiogenesis is a complex cellular differentiation process that the germ cells undergo a distinct morphological change, and the protamines replace the core histones to facilitate chromatin compaction in the sperm head. Recent studies show the essential roles of epigenetic events during the histone-to-protamine transition. Defects in either the replacement or the modification of histones might cause male infertility with azoospermia, oligospermia or teratozoospermia. Here, we summarize recent advances in our knowledge of how epigenetic regulators, such as histone variants, histone modification and their related chromatin remodelers, facilitate the histone-to-protamine transition during spermiogenesis. Understanding the molecular mechanism underlying the modification and replacement of histones during spermiogenesis will enable the identification of epigenetic biomarkers of male infertility, and shed light on potential therapies for these patients in the future.
Spermatogenesis is the process of male gamete production with successive cellular differentiation, which can be subdivided into spermatogonial mitosis, spermatocytic meiosis and spermiogenesis (Roosen-Runge, 1962; Hess and Renato De Franca, 2008). During spermatogenesis, SSC (spermatogonial stem cells) undergo self-renewal and differentiate into spermatogonia that perform meiosis to generate haploid germ cells and ensure the genetic diversity through meiotic recombination (Rathke et al., 2014; Bao and Bedford, 2016). Then, the haploid germ cells undergo spermiogenesis with a distinct morphological change and chromatin compaction in the sperm nuclei to prevent the paternal genome from mutagenesis and damage (Govin et al., 2004; Bao and Bedford, 2016). During the nuclear chromatin re-organization in spermiogenesis, the majority of the somatic histones are firstly replaced by testis-specific histone variants, and transition proteins (TPs) are subsequently incorporated in the nuclei of spermatids, protamines (PRMs) further replace TPs in the late spermatids to pack the genome into the highly condensed sperm nucleus (Rathke et al., 2014; Bao and Bedford, 2016). During the histone-to-protamine transition, the histone variants and specific histone modifications play essential roles by modulating the chromatin compaction and higher-order chromatin structure (Table 1) (Boskovic and Torres-Padilla, 2013; Bao and Bedford, 2016; Hada et al., 2017; Hao et al., 2019). Defects in either the replacement or the modification of histones might result in azoospermia, oligospermia or teratozoospermia, which leads to male infertility (Table 2). The focus of this review is on recent advances in our knowledge of how epigenetic regulators, such as histone variants, histone modification and their related chromatin remodelers, regulate the highly orchestrated chromatin re-organization and facilitate the histone-to-protamine transition during spermiogenesis.
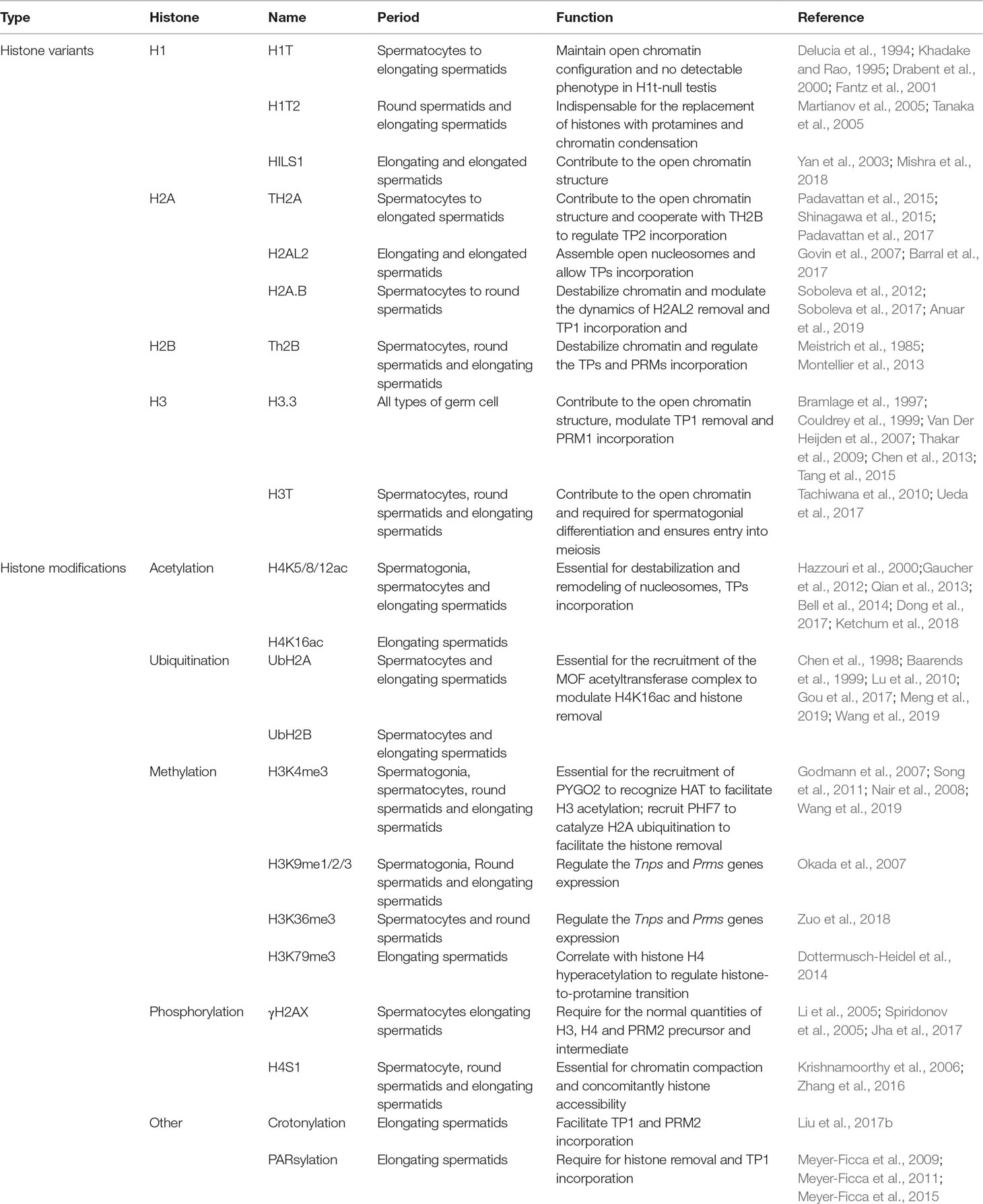
Table 1 The main histone variants and modifications during the histone-to-protamine transition.

Table 2 Mouse models related with the histone-to-protamine transition.
In eukaryotes, nucleosomes are the packing units of DNA, which contain four types of canonical histones (H2A, H2B, H3, and H4) and the linker histone H1 (Talbert and Henikoff, 2010; Kowalski and Palyga, 2012). While canonical histone expression is typically coupled to DNA replication, some non-canonical histones (histone variants) that are distinct form their canonical paralogues in amino acid sequence, are constitutively expressed and have roles in a wide range of processes (Talbert and Henikoff, 2010). Many histones variants are expressed during spermiogenesis and modulate the chromatin structure to facilitate the histone-to-protamine replacement (Mccarrey et al., 2005; Govin et al., 2007). Here, we summarize the recent advances in our understanding of the role of histone variants during the histone-to-protamine transition.
Linker histones contribute to form and stabilize the higher-order chromatin structure (Bednar et al., 1998). In mammals, there are about 11 different subtypes of histone H1 (Happel and Doenecke, 2009). Among these, H1T, H1T2, and HILS1 are testis-specific H1 variants (Figure 1) (Happel and Doenecke, 2009).
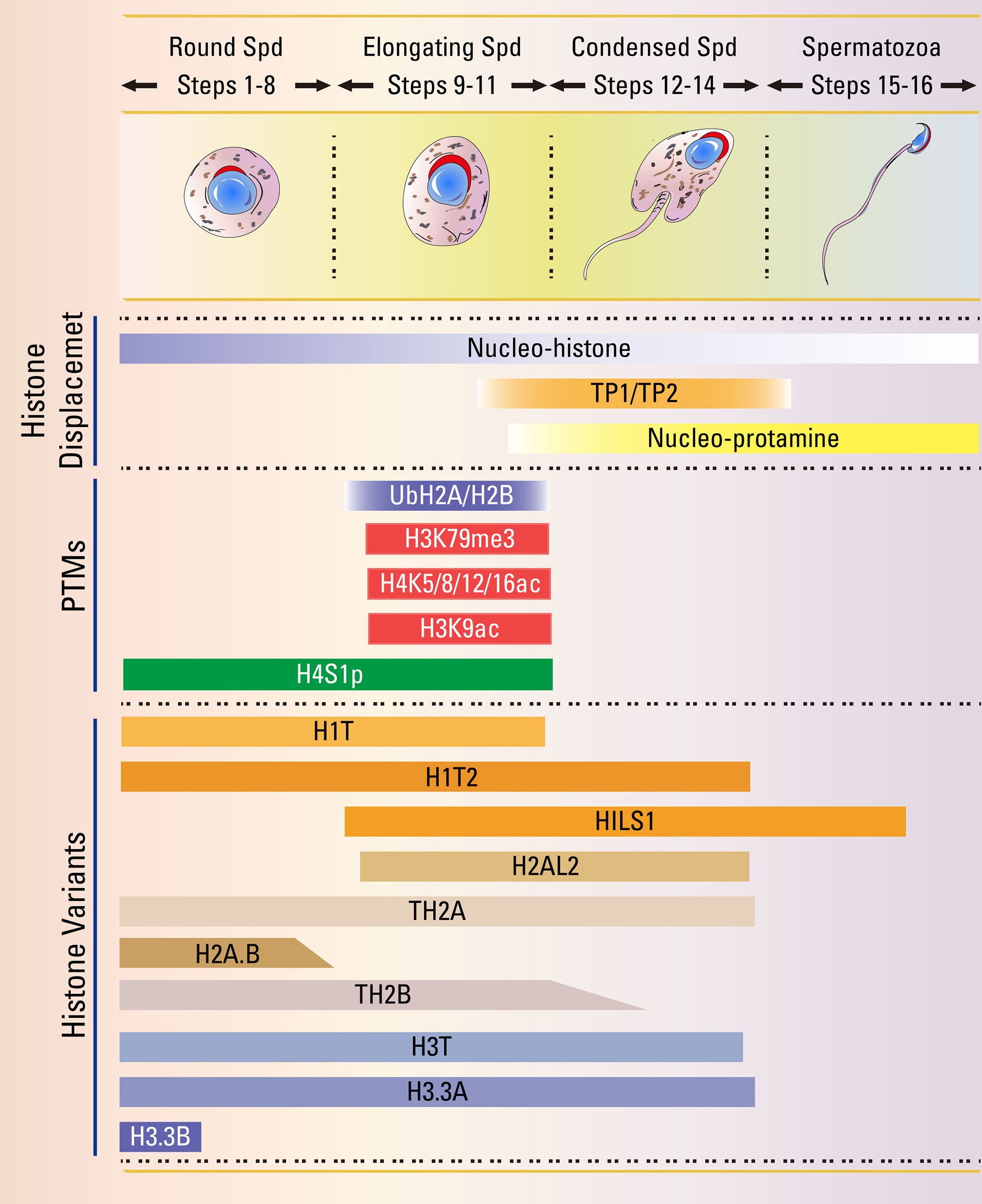
Figure 1 Summary of critical time points and epigenetic events during the histone-to-protamine transition. The haploid germ cells undergo a dramatic morphological change, and nuclear chromatin re-organization starts from round spermatid (Spd) to mature spermatozoa. Detailed studies of the indicated histone variants and histone modification might establish the precise epigenetic events of spermiogenesis.
H1T is exclusively detected as early as mid- to late pachytene spermatocytes, and maintains high expression levels in the elongating spermatids (Figure 1) (Drabent et al., 1996; Drabent et al., 2003). Biochemical and biophysical studies found that, distinct from other somatic H1 variants, H1T binds less tightly to H1-depleted nucleosomes, suggesting it may maintain a relatively open chromatin configuration to facilitate histone replacement during spermiogenesis (Delucia et al., 1994; Khadake and Rao, 1995). Unexpectedly, H1t-null mice are fertile and exhibit no spermatogenesis abnormalities, and the histone-to-protamine transition in H1t-deficient testis is normal (Drabent et al., 2000; Fantz et al., 2001). Although the expression of some canonical subtypes, including H1.1, H1.2, and H1.4, is enhanced in H1t-null mice, elevated levels of H1.1 or H1.2 could not be observed in the H1t-deficient spermatids (Drabent et al., 2003), indicating some other types of H1 variants may play redundant roles in the histone-to-protamine transition.
H1T2 selectively localizes at the apical pole in the nucleus of round and elongating spermatids but not in mature spermatozoa (Figure 1) (Martianov et al., 2005). Distinct from H1T, H1T2 is critical for spermiogenesis, as homozygous H1t2-mutant males are infertile due to delayed nuclear condensation and aberrant elongation of spermatids. Further analysis shows the protamine levels are substantially reduced in H1t2-null spermatozoa (Martianov et al., 2005; Tanaka et al., 2005), indicating H1T2 is necessary for the incorporation of protamines, and proper chromitin condensation during the histone-to-protamine transition.
HILS1 is strongly expressed in the nuclei of elongating and elongated spermatids (Figure 1) (Yan et al., 2003). HILS1 is the least conserved H1 variant, and a poor condenser of chromatin compared with somatic H1, demonstrating the idea that HILS1 may have a distinct role in the histone-to-protamine transition (Yan et al., 2003; Mishra et al., 2018). In Drosophila, Mst77F encodes a linker histone-like protein that is similar with the mammalian HILS1 protein and expressed in elongating spermatids (Raja and Renkawitz-Pohl, 2005). The disruption of Mst77F cause male sterile as producing spermatozoa with malformed heads. Although the histone-to-protamine transition occurs independently of Mst77F, the nuclei of spermatid fail to properly condense after the histone-to-protamine replacement in Mst77F mutant male (Kimura and Loppin, 2016). However, the functional roles of HILS1 in mammalian spermiogenesis need further investigation.
Multiple testis-specific H2A variants have been identified in mammals, including TH2A, H2AL1, H2AL2, H2AL3 and H2A.B (Trostleweige et al., 1982; Govin et al., 2007; Soboleva et al., 2012).
TH2A is present and actively synthesized in early primary spermatocytes and gradually disappears during condensation of spermatid nuclei (Figure 1) (Shires et al., 1976; Trostleweige et al., 1982). TH2A could contribute to the open chromatin structure, as crystal structures of nucleosome core particles (NCPs) with TH2A show the H-bonding interactions between the TH2A/TH2A′ L1 loops are lost and the histone dimer-DNA contacts are dramatically decreased (Padavattan et al., 2015; Padavattan et al., 2017). Although a Th2a-knockout mouse model has yet to be established, mice with knockouts of the testis-specific H2B variants Th2a and Th2b exhibit male infertility with few sperm in the epididymis (Shinagawa et al., 2015). In this double-knockout mouse, impaired chromatin incorporation of transition protein 2 (TP2) and elevated H2B could be observed in the mutant testis, suggesting the TH2A and TH2B may regulate the function in chromatin dynamics or the total histone levels to facilitate the histone replacement during spermatogenesis (Shinagawa et al., 2015). As the Th2b-null male mice show normal spermatogenesis and fertility (Montellier et al., 2013), the histone replacement defect in Th2a/Th2b double-knockout male mice is probably caused by the depletion of Th2a or their synergistic effect.
In late-developing post-meiotic male germ cells, H2AL2 is specifically expressed in condensing spermatids that correlates with the expression of TPs (Figure 1) (Govin et al., 2007). By comparing H2al2-null mice to wild-type mice, H2AL2 was demonstrated to be required to load TPs onto the nucleosome and for efficient PRMs assembly during the histone-to-protamine transition. Additionally, the nucleosome reconstitution assays revealed that the incorporation of H2A.L.2 can drastically modulate the nucleosome structure to facilitate TPs invading the nucleosomes and further transformation (Barral et al., 2017). Thus, H2AL2 could assemble open nucleosomes and allow TPs invading, which further promotes protamine processing and sperm genome compaction.
H2A.B is spatially and temporally regulated during spermatogenesis and detectable from the pachytene stage to the round spermatids (Figure 1) (Soboleva et al., 2012; Soboleva et al., 2017). In vitro studies show that H2A.B is able to destabilize chromatin and has unfolding properties to chromatin (Soboleva et al., 2012), indicating H2A.B might promote chromatin reorganization and further histones displacement by TPs. Male H2a.b-null male mice are subfertile due to the production of abnormal spermatozoa and clogged seminiferous tubules (Anuar et al., 2019). In H2a.b-null elongating spermatids, H2AL2 could not be detected in pericentric heterochromatin, and the replacement of TP1 by protamines appears to be delayed (Anuar et al., 2019). These results indicate H2A.B might modulate the dynamics of H2AL2 and TP1 chromatin incorporation and removal to participate in the histone-to-protamine transition.
The testis-specific histone variant TH2B is one of the earliest histone variants identified in testis (Shires et al., 1975). TH2B massively replaces somatic H2B during meiosis and remains the main type of H2B in round and elongating spermatids (Meistrich et al., 1985; Montellier et al., 2013), suggesting TH2B might be indispensable for meiotic and post-meiotic germ cells. The crystal structure analysis shows the TH2B could not form the water-mediated hydrogen bonds with H4R78 (Urahama et al., 2014), which may affect the stability of the TH2B nucleosome and facilitate histone replacement during spermiogenesis. In a Th2b mutant mouse, which contains modified C-terminus of the TH2B protein and causes a dominant-negative effect, males were infertile and severe abnormalities were seen in the elongating spermatids, which affected subnucleosomal transitional states during histone replacement (Boskovic and Torres-Padilla, 2013; Montellier et al., 2013). In contrast, Th2b-null mice are fertile and show normal spermatogenesis process, indicating a compensatory mechanism that rescues deficiency of TH2B in the histone-to-protamine transition. Indeed, in Th2b-null testis, the expression of somatic H2B was significantly increased and elevated methylation of H4R35, H4R55, H4R67, and H2BR72 could be detected in Th2b-null spermatids. As H4R35, H4R55, H4R67, and H2BR72 participate in the interactions of histone–DNA and histone–histone, and their methylation may impair these intranucleosomal interactions (Hoghoughi et al., 2018). Thus, the elevated somatic H2B and histone modification in Th2b-null spermatids might rescue the Th2b deficiency in testis (Montellier et al., 2013; Bao and Bedford, 2016).
In humans, H2BFWT is a testis-specific histone, is synthesized and aggregated in testes, and single nucleotide polymorphisms (SNPs) in this gene is highly associated with male infertility (Churikov et al., 2004; Lee et al., 2009; Ying et al., 2012; Rafatmanesh et al., 2018; Teimouri et al., 2018). And spermatid-specific H2B (ssH2B) and H2BL1 have been identified and are strongly enriched in round or elongating spermatids, similar to that of TPs and protamines (Moss and Orth, 1993; Unni et al., 1995; Govin et al., 2007). However, the functional roles of these H2B variants in the histone-to-protamine transition still need to be further elucidated.
In addition to the two canonical histones H3.1 and H3.2, three additional H3 variants have been identified and expressed in mammal testes, including H3.3, H3T and H3.5 (Rathke et al., 2014; Bao and Bedford, 2016).
H3.3 differs from canonical H3.1 with five amino acids, is expressed throughout mouse seminiferous tubules, and accumulates in the XY body of spermatocytes (Bramlage et al., 1997; Van Der Heijden et al., 2007). Biochemical and biophysical studies show that H3.3 contributes to an open chromatin configuration and promotes transcription through disrupting the higher-order chromatin structure (Thakar et al., 2009; Chen et al., 2013). H3.3 could be encoded by two gene paralogs in mammal, H3f3a and H3f3b, and the depletion of either H3f3a or H3f3b causes male infertility. The disruption of H3f3a produces abnormal spermatozoa (Couldrey et al., 1999; Tang et al., 2015), and the loss of H3f3b leads to growth defects and death at birth, with surviving H3f3b-null males showing complete infertility (Yuen et al., 2014). In H3f3b-null germ cells, the TP1 is abnormally deposited in elongating spermatids while PRM1 could not be observed in in elongated spermatids and mature spermatozoa, indicating that H3f3b is required for chromatin reorganization and the histone-to-protamine transition (Yuen et al., 2014). H3T (H3.4) is exclusively expressed in the spermatocyte and diminishes in the elongating spermatids (Ueda et al., 2017). Biochemical studies clearly indicate that, in the H3T nucleosome, the DNA around the entry-exit regions shows more flexible than that of the H3.1-containing nucleosome, and that the H3T-containing polynucleosome could formed more open configuration than that of H3.1 (Tachiwana et al., 2010). However, the disruption of H3T leads to sterile males with azoospermia, as spermatocyte and spermatids are absent in the H3t-null testes (Ueda et al., 2017). Thus, the function of H3T in the later stage of spermatogenesis need further investigated by using spatially and temporally specific knockout mouse models.
H3.5 is highly expressed in human testis and specifically observed in spermatogonia and spermatocytes (Shiraishi et al., 2017). In vitro studies reveal that the H3.5-specific L103 residue, reduces the hydrophobic interaction with histone H4 in the H3.5-containing nucleosome, which corresponds to the H3.3 Phe104 residue (Urahama et al., 2016). H3.5 is significantly reduced in non-obstructive azoospermia (NOA) patients (Shiraishi et al., 2017), whereas the precise roles of H3.5 in spermatogenesis remain largely unknown.
Covalent conjugation of different post-translational modification of histones has a dramatic effect on the chromatin conformation by affecting the stability of the nucleosome and the histone-DNA interaction (Bao and Bedford, 2016). Many types of histone modifications have been identified to facilitate the histone-to-protamine transition, including acetylation, ubiquitination, methylation, and phosphorylation (Luense et al., 2016).
Hyperacetylated histones could facilitate histone eviction, and the acetylation of H2A, H2B, H3, H4 and histone variants have been detected in mammal testis (Grimes and Henderson, 1984a; Grimes and Henderson, 1984b; Oliva and Mezquita, 1986; Oliva et al., 1987). In Drosophila, inactivation of histone acetyltransferases by anacardic acid prevents the histones degradation and further a protamine incorporation during spermiogenesis (Awe and Renkawitz-Pohl, 2010), suggesting that histone acetylation is essential for the histone-to-protamine replacement.
H4 acetylation (H4K5ac, H4K8ac, H4K12ac, and H4K16ac) shows a spatial distribution pattern during spermatogenesis and is indispensable for the histone-to-protamine transition (Bao and Bedford, 2016; Ketchum et al., 2018). H4K5ac, H4K8ac and H4K12ac are expressed in spermatogonia and pre-leptotene spermatocytes, disappear in leptotene to pachytene spermatocytes, reappeared in elongating spermatids, and finally disappeared in condensing spermatids (Figure 1) (Hazzouri et al., 2000; Ketchum et al., 2018). In contrast, H4K16ac could only be detected in elongating spermatids (Figure 1) (Ketchum et al., 2018). In vitro analysis shows that H4 acetylation is essential for destabilization and remodeling of nucleosomes, and the incorporation of H4K16ac into nucleosomes prevents the formation of compact chromatin fibers and influence chromatin forming cross-fiber interactions (Tse et al., 1998; Shogren-Knaak et al., 2006; Kan et al., 2009). These findings indicate that H4 acetylation modulates higher order chromatin structure to facilitate the histone-to-protamine transition. EPC1 (Enhancer Of Polycomb Homolog 1) and TIP60 (Tat-interactive protein, 60 kDa), which are two components for the mammalian NuA4 (nucleosome acetyltransferase of H4) complexes (Figure 2) (Doyon et al., 2004), are co-localized to the nuclear periphery near the acrosomes in both round spermatids and elongating spermatids (Dong et al., 2017). The depletion of either Epc1 or Tip60 perturbs histone hyperacetylation, especially H4 acetylation, and affects histone replacement during spermiogenesis (Dong et al., 2017). Another gene that may play a role in acetylation is SIRT1 (Sirtuin 1), a member of the NAD+-dependent deacetylase. Germ cell-specific Sirt1 knockout mice display reduced male fertility due to decreased spermatozoa number and increased proportion of abnormal spermatozoa (Bell et al., 2014; Liu et al., 2017a). In Sirt1-null elongating and elongated spermatids, acetylation levels of H4K5, H4K8 and H4K12 are decreased and TP2 could not co-localize in the nucleus, leading to a chromatin condensation defect in Sirt1-null spermatozoa (Bell et al., 2014). Thus, SIRT1 may modulate other factors to promote H4 acetylation and the histone-to-protamine transition.
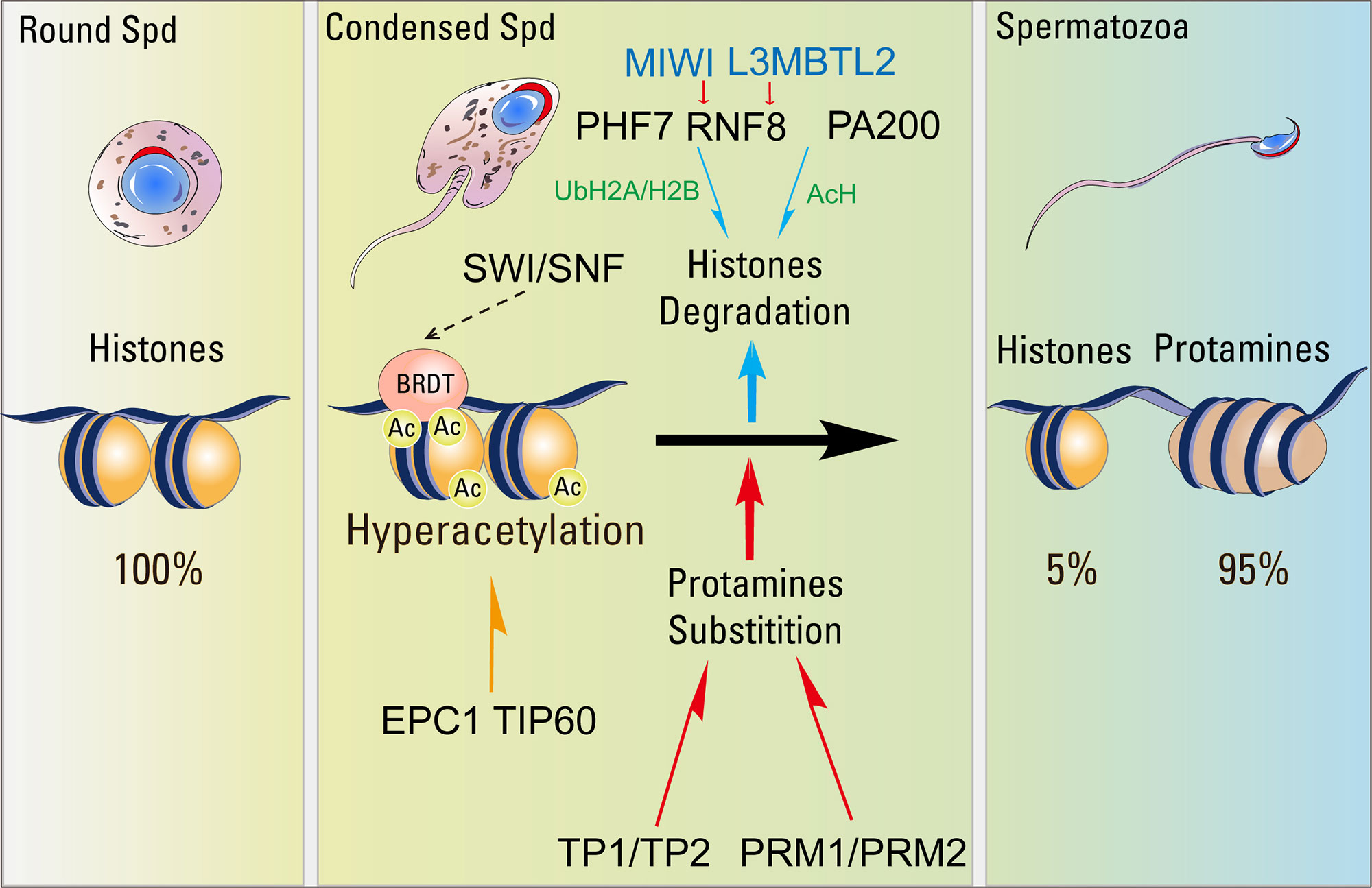
Figure 2 The key factors related to the histone-to-protamine transition. Global incorporation of various H2A, H2B and H3 histone variants creates highly unstable nucleosomes, which then undergo histone hyperacetylation by EPC1/TIP60 or some other nucleosome acetyltransferase complexes. Acetylation at critical lysines further destabilizes the nucleosomes, while tail acetylation generates a platform for the recruitment of BRDT. BRDT interacts with the SWI/SNF family protein then starts the process of histone eviction and replacement by TPs. Evicted acetylated histones would then be recognized by PA200 and degraded by proteasomes during spermatogenesis. RNF8 could catalyze the ubiquitination of H2A. Ubiquitinated H2A and H2B control H4K16ac by regulating the association of MOF to the chromatin and facilitates histone removal in elongating spermatids. MIWI binds to RNF8 in the cytoplasm of early spermatids (Spd) through a Piwi-interacting RNA (piRNA)-independent manner, and promotes the nuclear translocation of RNF8 in late spermatids to catalyze histone ubiquitination and trigger histone removal. L3MBTL2 could interact with RNF8 and facilitate RNF8-dependent histone ubiquitination-related histone removal. PHF7 could recognize the H3K4me3/me2 and catalyze H2A ubiquitination to facilitate histone removal in elongating spermatids.
The histone acetylation might be recognized by some chromatin remodelers to confer downstream signaling, and the double bromodomain and extra-terminal domain (BET) proteins have been identified to be critical epigenetic readers binding to acetylated histones and modulating changes in chromatin structure and organization during spermiogenesis (Berkovits and Wolgemuth, 2013). BRDT is a testis-specific BET member protein, which is expressed specifically in spermatocytes and spermatids, and contains two bromodomains that specifically recognize acetylated lysine residues (Shang et al., 2007; Dhar et al., 2012; Manterola et al., 2018). BRDT binds the hyperacetylated histone H4 tail and co-localizes with acetylated H4 in elongating spermatids (Pivot-Pajot et al., 2003; Govin et al., 2006). Remodeling assays have shown BRDT regulated the chromatin reorganization dependent acetylation in round spermatids (Dhar et al., 2012). In mice, the disruption of the first bromodomain in BRDT resulted in male sterility by producing the morphologically abnormal spermatids (Shang et al., 2007). In elongating spermatids with BRDT containing a knockout of bromodomain 1 (BD1), TPs and protamines remained in the cytoplasm and histone replacement did not occur, suggesting BRDT is required for the histone-to-protamine transition by mediating the replacement of acetylated histones (Figure 2) (Gaucher et al., 2012). Furthermore, BRDT was found to bind with the N-terminus of SMARCE1 (SWI/SNF-related matrix-associated actin-dependent regulator of chromatin subfamily E member 1), a member of the SWI/SNF family of ATP-dependent chromatin remodeling complexes (Dhar et al., 2012), indicating BRDT may cooperate with SMARCE1 to facilitate the histone-to-protamine transition during spermiogenesis (Figure 2).
Proteasomes catalyze ATP- and polyubiquitin-dependent protein degradation, and they are made up of a 20S catalytic core particle (CP) and regulatory particle (RP). The 20S CP could be activated by cooperation with various RPs, such as PA700/19S, PA28α/β, PA28γ, and PA200 (Stadtmueller and Hill, 2011). PA200 is highly expressed in the testis, and the disruption of PA200 results in male infertility and severe defects in spermatogenesis (Ustrell et al., 2005; Khor et al., 2006). During spermiogenesis, PA200 regulatory could directly recognize acetylated histones through a bromodomain-like module and promote their ubiquitin-independent degradation. In Pa200-null spermatids, results showed that H2B, H3 and elevated H4K16ac could be detected at the end of the elongation stage (Qian et al., 2013). Thus, PA200 specifically recognizes acetylated histones and mediates the core histones for acetylation dependent degradation through proteasomes during spermatogenesis (Figure 2).
Ubiquitin is a 76 amino acid protein that is attached to target proteins to regulate several cellular processes, such as protein degradation, cell signaling, autophagy, DNA damage responses and so on (Hershko and Ciechanover, 1998; Pickart, 2001; Welchman et al., 2005; Komander and Rape, 2012). Ubiquitinated H2A and H2B are enriched in spermatocytes and elongating spermatids (Chen et al., 1998; Baarends et al., 1999). RNF8 is an ubiquitin E3 ligase that participates in DDR (DNA damage repair) by catalyzing the ubiquitination of H2A to promote the recruitment of some DNA damage response factors on the damage sites (Ma et al., 2011). The disruption of Rnf8 causes significant late-stage developmental defects in spermatids due to problematic histone-to-protamine replacement, with the canonical histones being detectable in Rnf8-deficient mature spermatozoa (Lu et al., 2010). In Rnf8-null mice, both ubiquitinated H2A and H2B are decreased in the testes and H4K16ac is dramatically decreased as well (Lu et al., 2010). Further studies showed that ubiquitinated H2A and H2B were essential for the efficient recruitment of the MOF (males absent on the first) acetyltransferase complex, which is highly expressed in elongating spermatids and responsible for H4K16 acetylation in the chromatin (Akhtar and Becker, 2000; Lu et al., 2010). Thus, RNF8 catalyzed histone ubiquitination could modulate H4K16ac by regulating the localization of MOF on the chromatin and facilitate histone removal in the elongating spermatids.
The RNF8-dependent histone ubiquitination during spermiogenesis could also be modulated by PIWI protein, which is specifically expressed during germline development and enlists piRNAs (Piwi-interacting RNAs) to repress TE (transposable elements) and protect the germ cell genome integrity (Juliano et al., 2011; Siomi et al., 2011; Gou et al., 2017). In mice, Miwi, Mili, and Miwi2, the Piwi paralogs, have been identified in the testis and are required for male fertility (Deng and Lin, 2002; Kuramochi-Miyagawa et al., 2004; Carmell et al., 2007). During spermiogenesis, MIWI binds to RNF8 in the cytoplasm of early spermatids through a piRNAs-independent manner, and APC/C mediated MIWI degradation in late spermatids is essential for nuclear translocation of RNF8, which catalyzes histone ubiquitination and further facilitates histone removal (Gou et al., 2017). In both humans and mice, mutations in the conserved destruction box (D-box) of HIWI and MIWI proteins, which lead to their stabilization, cause male infertility due to impaired histone ubiquitination and histone-to-protamine transition (Gou et al., 2017). Except MIWI, L3MBTL2 (Lethal 3 malignant brain tumor like 2), a member of the MBT-domain proteins that is implicated in chromatin compaction, could also interact with RNF8. The depletion of L3mbtl2 in germ cells affected male fertility by producing abnormal spermatozoa and the decrease of sperm counts. L3mbtl2 deficiency also caused the reduction of in levels of the RNF8 and histone ubiquitination in elongating spermatids, which further influenced the PRM1 deposition and chromatin condensation during spermiogenesis (Meng et al., 2019).
PHF7 (PHD Finger Protein 7), which contains PHD (plant homeodomain) and RING finger domain, has been identified as a novel H2A ubiquitination E3 ligase in mouse testis (Hou et al., 2012; Wang et al., 2019). PHF7 is specifically located in the elongating spermatid nuclei, and the disruption of Phf7 led to male mouse infertility as reduction of sperm count and the increased proportion of abnormal spermatozoa (Wang et al., 2019). PHF7 could recognize the H3K4me3/me2 through its PHD domain and catalyze H2A ubiquitination by its RING domain. In Phf7-null spermatids, the H2A ubiquitination was dramatically decreased that resulted in the histone retention and protamine replacement defect (Figure 2) (Wang et al., 2019). Therefore, PHF7 has dual roles during the histone-to-protamine transition that works as an epigenetic reader by recognizing H3K4me3/me2 and as an epigenetic writer through catalyzing H2A ubiquitination to promote histone removal.
Multiple histone methylation have been identified in elongating spermatids, for instance H3K4me2, H3K4me3, H3K9me2, H3K9me3, H3K27me3, H3K79me2, and H3K79me3 (Godmann et al., 2007; Song et al., 2011; De Vries et al., 2012; Dottermusch-Heidel et al., 2014). Among them, the methylation of H3K4 and plus acetylation might help to achieve a more-open chromatin configuration, whereas H3K9 and H3K27 methylation are known to be associated with a more-repressed chromatin configuration (Rathke et al., 2014), indicating a balance of “opened” and “closed” chromatin regions during the histone-to-protamine transition. As some histone methyltransferases and demethylases are detectable during spermiogenesis (Godmann et al., 2007; Liu et al., 2010; Ushijima et al., 2012), the histone methylation may be dynamically regulated in testis. Although few mouse models exist that allow precise detection of methylation activity that directly regulates histone replacement during spermiogenesis, some studies have revealed that histone methylation may modulate the histone-to-protamine transition through some other ways. PYGO2 (Pygopus homolog 2) comprises a C-terminal PHD finger, which can recognize the H3K4me3 and is specifically located in the elongating spermatid nuclei. In mice, the reduction of Pygo2 influenced the Tnp, Prm genes expression and caused the abnormal nuclear condensation, which further led to male sterility (Nair et al., 2008). Furthermore, PYGO2 associates with a histone acetyltransferase (HAT) activity, and the acetylation of H3 is disrupted in Pygo2 reduced elongating spermatids (Nair et al., 2008), indicating PYGO2 may recognize H3K4me3 through its PHD domain and could recruit HAT to facilitate H3 acetylation and further histone-to-protamine transitions. As described before, PHF7 could recognize the H3K4me3/me2 and catalyze H2A ubiquitination to facilitate the histone-to-protamine transitions (Wang et al., 2019). The predominant histone methyltransferase SETD2 (SET domain–containing 2) catalyzes the H3K36me3, and knocking out Setd2 in mouse germ cells causes aberrant spermiogenesis, resulting in complete male infertility. Moreover, the disruption of SETD2 causes complete loss of H3K36me3 and impaired activation of Tnp and Prm genes (Zuo et al., 2018), indicating H3K36me3 may regulate the histone-to-protamine transition by activating Tnp and Prm genes expression. Contrarily, JHDM2A (JmjC-domain-containing histone demethylase 2A) is an H3K9me2/1-specific demethylase. The loss of Jhdm2a in mice exhibits post-meiotic chromatin condensation defects and leads to male infertility. Although global H3K9 methylation has no effect in Jhdm2a-null testis, JHDM2A directly binds to and controls H3K9 methylation at the promoter of Tnp1 and Prm1 genes, which further regulates the sperm genome packaging and chromatin condensation (Okada et al., 2007).
Histone phosphorylation is involved in various cellular processes (Rossetto et al., 2012; Bao and Bedford, 2016), and dynamic histone phosphorylation have been observed during spermatogenesis (Govin et al., 2010; Bao and Bedford, 2016). The phosphorylation of histone H2AX at residue Ser139 (γH2AX) plays important roles in many biological processes, such as meiotic recombination and male sex chromosome inactivation in germ cells (Li et al., 2005). γH2AX is detectable in elongating spermatids, and TSSK6 has been identified to be responsible for the H2AX phosphorylation during spermiogenesis (Jha et al., 2017). In mice, targeted deletion of Tssk6 leads to male sterility caused by the impairment in morphology and motility of spermatozoa (Spiridonov et al., 2005). In spermatozoa, the loss of TSSK6 blocks γH2AX formation, resulting in elevated H3, H4 and the precursor and intermediate of PRM2 (Jha et al., 2017). These results indicate that TSSK6 may mediate γH2AX to participate in the histone-to-protamine transition. H4S1 phosphorylation is highly expressed in mouse spermatocyte, round and elongating spermatids (Krishnamoorthy et al., 2006; Zhang et al., 2016). H4S1 phosphorylation has been found to be essential for chromatin compaction and concomitantly histone accessibility (Krishnamoorthy et al., 2006; Wendt and Shilatifard, 2006), suggesting that H4S1 phosphorylation is required for histone replacement during spermiogenesis. Outside the canonical histones, many phosphorylated residues have been identified, using mass spectrometry analyses, that exist on different testis-specific histone variants, such as H1T, HILS1, TH2A, TH2B (Sarg et al., 2009; Pentakota et al., 2014; Mishra et al., 2015; Luense et al., 2016; Hada et al., 2017). Although many core histones and histone variants phosphorylation have been identified in germ cells, their physiological roles need further investigation.
A variety of histone lysine modifications have been identified, including butyrylation, crotonylation, malonylation, propionylation, and succinylation (Tan et al., 2011; Sabari et al., 2017). Kcr (Lysine crotonylation) is a newly identified histone modification and is detectable in elongating spermatids, which regulated testis-specific genes activation in post-meiotic germ cells (Tan et al., 2011). The CDYL (chromodomain Y-like) protein, which contains a C-terminal CoAP domain that interacts with CoA to achieve its crotonyltransferase activity, may suppress the histone Kcr by converting crotonyl-CoA to β-hydroxybutyryl-CoA. Accordingly, Cdyl-deficient male mice show reduced fertility, decreased epididymal sperm count and sperm cell motility, and dysregulated histone Kcr (Liu et al., 2017b). In the Cdyl-deficient mouse testes, further analysis showed that the elevated TP1 and PRM2 were localized in a chromatin-free regions (Liu et al., 2017b), suggesting that histone crotonylation is essential for the histone-to-protamine transition during spermiogenesis.
Poly-ADP-ribosylation (PARsylation) is a common protein PTM (post-translational modification) observed in higher eukaryotes and involved in many different fundamental cellular functions. All of core histones and the linker histone H1 can be ADP-ribosylated (Gagne et al., 2006; Messner and Hottiger, 2011), which could be catalyzed by poly(ADP-ribose) polymerases, such as PARP1 and PARP2, and resolved by PARG (PAR glycohydrolase) (Gibson and Kraus, 2012). The PARP1, PARP2 and PARsylation proteins are specifically detected in elongating spermatids (Meyer-Ficca et al., 2005), and the perturbed PARsylation causes reduced male fertility with abnormal retention of core histones, H1T and HILS1 in mature spermatozoa (Meyer-Ficca et al., 2009; Meyer-Ficca et al., 2011; Meyer-Ficca et al., 2015). Thus, PARsylation is essential for the histone-to-protamine replacement, yet the precise PARsylation histone sites need further characterization.
Between histone eviction and protamine incorporation in the nuclei of spermatids, about ninety percent of the chromatin components consist of TPs, which are arginine- and lysine-rich proteins encoded by Tnp1 and Tnp2 (Meistrich et al., 2003). However, the functional roles of each TP are still controversial (Rathke et al., 2014). TP1 could reduce the melting temperature of DNA and relax the DNA from core particles of nucleosome, whereas TP2 tends to compact the nucleosomal DNA by increasing its melting temperature, indicating TP2 may promote DNA condensation while TP1 facilitates the eviction of the histones (Singh and Rao, 1988; Akama et al., 1998; Kolthur-Seetharam et al., 2009; Rathke et al., 2014). However, a separate study that shown that neither TP1 nor TP2 leads to the conformation changes in supercoiled DNA (Levesque et al., 1998). These differences might reveal their unique roles during mammal spermiogenesis, as single knockout of either Tnp1 or Tnp2 leads to little morphological alteration of spermatozoa in mouse models. Elevated TP2 and TP1 proteins could be observed in Tnp1-null and Tnp2-null spermatids, respectively (Yu et al., 2000; Zhao et al., 2001). Thus, TP1 and TP2 may compensate for each other in vivo. Indeed, Tnp1 and Tnp2 double-knockout mice show severe abnormal spermiogenesis with a general decrease in sperm motility and abnormal sperm morphology (Shirley et al., 2004). The chromatin condensation is perturbed in the Tnp1 and Tnp2 double-knockout mice as severe histones retention is detectable, indicating TPs function redundantly yet have unique roles in the histone-to-protamine transition (Shirley et al., 2004; Zhao et al., 2004; Bao and Bedford, 2016).
Protamines are basic proteins that replace TPs in late spermatids (Rathke et al., 2014; Bao and Bedford, 2016). Two protamine genes (Prm1 and Prm2) localize on the same chromosome in both humans and mice (Balhorn, 2007). Protamines tightly interact with DNA via a central arginine-rich DNA-binding domain (Balhorn, 2007). Unlike Tnp genes, the disruption of either Prm1 or Prm2 leads to the male infertility (Cho et al., 2001). Protamines have multiple PTM sites, and a total of 11 PTMs have been identified on the protamines of mouse spermatozoa, including acetylation, phosphorylation and methylation (Brunner et al., 2014). One site of interest is PRM2 S55, which is a candidate phosphorylated substrate residue of CAMK4 (Ca2+/calmodulin-dependent protein kinase IV) (Wu et al., 2000). Targeted Camk4 knockout male mice are infertile, and the transition protein displacement by PRM2 is perturbed as a specific loss of PRM2 and prolonged retention of TP2 in Camk4-null spermatids. In vitro, PRM2 could be phosphorylated by CAMK4, implicating CAMK4 mediated PRM2 phosphorylation is required for the protamine incorporation during spermiogenesis (Wu et al., 2000). Thus, the specific post-translational modifications on protamines may also be essential for the histone-to-protamine transition.
During the histone-to-protamine transition, many epigenetic regulators work together to facilitate paternal genome re-organization and packaging into the highly condensed nuclei of spermatozoa, through histone variation, specific histone modification and their related chromatin remodelers. Any defects during the histone-to-protamine transition would lead to male infertility (Bao and Bedford, 2016). While the morphological changes during spermiogenesis are well characterized, the precise molecular mechanisms underlying the chromatin re-organization, in particular the transition from histones to protamines, are still unclear. It’s difficult to characterize the dynamic processes that occur during histone eviction, transition protein incorporation and protamine insertion. Moreover, 10% of the spermatozoa population in the epididymis has not yet completed the histone-to-protamine transition (Yoshida et al., 2018). These problems may be ascribed to a lack of experimental methods, which could fully recapitulate germ cell development in vitro. Further physiological insights may be gained by developing an in vitro germ-cell culture system that more accurately recapitulates the in vivo histone-to-protamine transition.
Many histone variants modulate histone replacement by regulating the chromatin structure; therefore, nucleosomes containing these histone variants often maintain a relatively decondensed and open chromatin configuration, facilitating histone replacement during spermiogenesis. The redundant function of histone variants in modulating chromatin configuration ensures that defects in some histone variants have a limited effect on spermatogenesis. Indeed, some mutant histone variants in mouse models are dispensable for male fertility, and mice may show elevated levels of compensatory histones or histone variants. However, the redundant function of histone variants makes it difficult to explore the precise role of each histone variant in histone replacement.
Although many histone modifications have been identified during the histone-to-protamine transition, many studies are descriptive and correlative. The direct manipulation of histone modification sites to reveal function is still urgently needed. With the development of gene editing tools, for example the CRISPR/Cas9 system, mouse models disrupting these histone modifications may be generated and used to elucidate function and in vivo relevance in the future. The following open-ended questions still need to be answered to provide in-depth investigation in the field.
1. In addition to the histone variants and modifications mentioned above, what other novel histone variants and modifications participate in the histone-to-protamine transition? How can we identify them?
2. How and where do histone variants replace canonical histones? What signal is needed to initiate replacement?
3. As histone variants and modifications are identified that participate in the histone-to-protamine transition, how do we establish an epigenetic modulating network for this process? Which type of histone code is the initiating code?
4. Histone hyperacetylation works as a determining event during the histone-to-protamine transition. Is this histone hyperacetylation an initial signal or an indirect consequence of prior events?
5. What’s the relationship between these histone variants and modifications? What’s the mechanism underlying the cross talk between them?
6. Chromatin assembly is modulated by histone chaperones or other chromatin remodelers. What’s the functional role of histone chaperones during the histone-to-protamine transition? Which histone variants or modifications send a signal to the chaperones?
7. (7) How do the transition proteins replace the histone? How do protamines replace the transition proteins? What are the detailed functional roles of transition proteins?
8. (8) Are there still post-translational modifications that need to be discovered in order to more accurately describe how histone modification plays a role in spermatozoa maturation?
These questions and their underlying ideas need further investigation and refining to help us more thoroughly understand the complex molecular relationships and exact regulating mechanisms of the histone-to-protamine transition.
WT and HG wrote the manuscript and drew the figures; CL and WL proposed the idea and revised the manuscript. All authors listed have made a substantial, direct and intellectual contribution to the work and approved it for publication.
This work was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (Grant No. XDA16020701), the National Key R&D Program of China (grant 2016YFA0500901, 2018YFC1004202), the National Natural Science Foundation of China (grants 31771501, 91649202) and the Youth Innovation Promotion Association CAS (2018109).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Tracey Baas for critical reading of the manuscript.
Akama, K., Sato, H., Hasegawa, S., Shimada, I., Nakano, M. (1998). Transition protein 1 from boar late spermatid nuclei having DNA-melting activity is a dimeric protein. Biochem. Mol. Biol. Int. 44, 315–323. doi: 10.1080/15216549800201332
Akhtar, A., Becker, P. B. (2000). Activation of transcription through histone H4 acetylation by MOF, an acetyltransferase essential for dosage compensation in Drosophila. Mol. Cell 5, 367–375. doi: 10.1016/S1097-2765(00)80431-1
Anuar, N. D., Kurscheid, S., Field, M., Zhang, L., Rebar, E., Gregory, P., et al. (2019). Gene editing of the multi-copy H2A.B gene and its importance for fertility. Genome Biol. 20, 23. doi: 10.1186/s13059-019-1633-3
Awe, S., Renkawitz-Pohl, R. (2010). Histone H4 acetylation is essential to proceed from a histone- to a protamine-based chromatin structure in spermatid nuclei of Drosophila melanogaster. Syst. Biol. in Reprod. Med. 56, 44–61. doi: 10.3109/19396360903490790
Baarends, W. M., Hoogerbrugge, T. W., Roest, H. P., Ooms, M., Vreeburg, J., Hoeijmakers, J. H. J., et al. (1999). Histone ubiquitination and chromatin remodeling in mouse spermatogenesis. Dev. Biol. 207, 322–333. doi: 10.1006/dbio.1998.9155
Balhorn, R. (2007). The protamine family of sperm nuclear proteins. Genome Biol. 8, 1–8. doi: 10.1186/gb-2007-8-9-227
Bao, J., Bedford, M. T. (2016). Epigenetic regulation of the histone-to-protamine transition during spermiogenesis. Reproduction 151, R55–R70. doi: 10.1530/REP-15-0562
Barral, S., Morozumi, Y., Tanaka, H., Montellier, E., Govin, J., De Dieuleveult, M., et al. (2017). Histone Variant H2A.L.2 Guides transition protein-dependent protamine assembly in male germ cells. Mol. Cell 66, 89–101 e108. doi: 10.1016/j.molcel.2017.02.025
Bednar, J., Horowitz, R. A., Grigoryev, S. A., Carruthers, L. M., Hansen, J. C., Koster, A. J., et al. (1998). Nucleosomes, linker DNA, and linker histone form a unique structural motif that directs the higher-order folding and compaction of chromatin. Proc. Natl. Acad. Sci. U. S. A. 95, 14173–14178. doi: 10.1073/pnas.95.24.14173
Bell, E. L., Nagamori, I., Williams, E. O., Del Rosario, A. M., Bryson, B. D., Watson, N., et al. (2014). SirT1 is required in the male germ cell for differentiation and fecundity in mice. Development 141, 3495–3504. doi: 10.1242/dev.110627
Berkovits, B. D., Wolgemuth, D. J. (2013). The role of the double bromodomain-containing BET genes during mammalian spermatogenesis. Gametogenesis 102, 293–326. doi: 10.1016/B978-0-12-416024-8.00011-8
Boskovic, A., Torres-Padilla, M. E. (2013). How mammals pack their sperm: a variant matter. Genes Dev. 27, 1635–1639. doi: 10.1101/gad.226167.113
Bramlage, B., Kosciessa, U., Doenecke, D. (1997). Differential expression of the murine histone genes H3.3A and H3.3B. Differentiation 62, 13–20. doi: 10.1046/j.1432-0436.1997.6210013.x
Brunner, A. M., Nanni, P., Mansuy, I. M. (2014). Epigenetic marking of sperm by post-translational modification of histones and protamines. Epigenet. Chromatin 7, 1–12. doi: 10.1186/1756-8935-7-2
Carmell, M. A., Girard, A., Van De Kant, H. J. G., Bourc’his, D., Bestor, T. H., De Rooij, D. G., et al. (2007). MIWI2 is essential for spermatogenesis and repression of transposons in the mouse male germline. Dev. Cell 12, 503–514. doi: 10.1016/j.devcel.2007.03.001
Chen, H. Y., Sun, J. M., Zhang, Y., Davie, J. R., Meistrich, M. L. (1998). Ubiquitination of histone H3 in elongating spermatids of rat testes. J. Biol. Chem. 273, 13165–13169. doi: 10.1074/jbc.273.21.13165
Chen, P., Zhao, J. C., Wang, Y., Wang, M., Long, H. Z., Liang, D., et al. (2013). H3.3 actively marks enhancers and primes gene transcription via opening higher-ordered chromatin. Genes Dev. 27, 2109–2124. doi: 10.1101/gad.222174.113
Cho, C., Willis, W. D., Goulding, E. H., Haesook, J. H., Choi, Y. C., Hecht, N. B., Eddy, E. M. (2001). Haploinsufficiency of protamine-1 or-2 causes infertility in mice. Nat. Genet. 28, 82–86. doi: 10.1038/ng0501-82
Churikov, D., Siino, J., Svetlova, M., Zhang, K. L., Gineitis, A., Bradbury, E. M., et al. (2004). Novel human testis-specific histone H2B encoded by the interrupted gene on the X chromosome. Genomics 84, 745–756. doi: 10.1016/j.ygeno.2004.06.001
Couldrey, C., Carlton, M. B. L., Nolan, P. M., Colledge, W. H., Evans, M. J. (1999). A retroviral gene trap insertion into the histone 3.3A gene causes partial neonatal lethality, stunted growth, neuromuscular deficits and male sub-fertility in transgenic mice. Hum. Mol. Genet. 8, 2489–2495. doi: 10.1093/hmg/8.13.2489
De Vries, M., Ramos, L., Housein, Z., De Boer, P. (2012). Chromatin remodelling initiation during human spermiogenesis. Biol. Open 1, 446–457. doi: 10.1242/bio.2012844
Delucia, F., Faraonemennella, M. R., Derme, M., Quesada, P., Caiafa, P., Farina, B. (1994). Histone-induced condensation of rat testis chromatin—testis-specific H1t versus somatic H1 variants. Biochem. Biophys. Res. Commun. 198, 32–39. doi: 10.1006/bbrc.1994.1005
Deng, W., Lin, H. F. (2002). Miwi, a murine homolog of Piwi, encodes a cytoplasmic protein essential for spermatogenesis. Dev. Cell 2, 819–830. doi: 10.1016/S1534-5807(02)00165-X
Dhar, S., Thota, A., Rao, M. R. S. (2012). Insights into role of bromodomain, testis-specific (Brdt) in acetylated histone H4-dependent chromatin remodeling in mammalian spermiogenesis. J. Biol. Chem. 287, 6387–6405. doi: 10.1074/jbc.M111.288167
Dong, Y., Isono, K., Ohbo, K., Endo, T. A., Ohara, O., Maekawa, M., et al. (2017). EPC1/TIP60-mediated histone acetylation facilitates spermiogenesis in mice. Mol. Cell. Biol. 37, e00082-17. doi: 10.1128/MCB.00082-17
Dottermusch-Heidel, C., Klaus, E. S., Gonzalez, N. H., Bhushan, S., Meinhardt, A., Bergmann, M., et al. (2014). H3K79 methylation directly precedes the histone-to-protamine transition in mammalian spermatids and is sensitive to bacterial infections. Andrology 2, 655–665. doi: 10.1111/j.2047-2927.2014.00248.x
Doyon, Y., Selleck, W., Lane, W. S., Tan, S., Cote, J. (2004). Structural and functional conservation of the NuA4 histone acetyltransferase complex from yeast to humans. Mol. Cell. Biol. 24, 1884–1896. doi: 10.1128/MCB.24.5.1884-1896.2004
Drabent, B., Benavente, R., Hoyer-Fender, S. (2003). Histone H1t is not replaced by H1.1 or H1.2 in pachytene spermatocytes or spermatids of H1t-deficient mice. Cytogenet. Genome Res. 103, 307–313. doi: 10.1159/000076818
Drabent, B., Bode, C., Bramlage, B., Doenecke, D. (1996). Expression of the mouse testicular histone gene H1t during spermatogenesis. Histochem. Cell Biol. 106, 247–251. doi: 10.1007/BF02484408
Drabent, B., Saftig, P., Bode, C., Doenecke, D. (2000). Spermatogenesis proceeds normally in mice without linker histone H1t. Histochem. Cell Biol. 113, 433–442.
Fantz, D. A., Hatfield, W. R., Horvath, G., Kistler, M. K., Kistler, W. S. (2001). Mice with a targeted disruption of the H1t gene are fertile and undergo normal changes in structural chromosomal proteins during spermiogenesis. Biol. Reprod. 64, 425–431. doi: 10.1095/biolreprod64.2.425
Gagne, J. P., Hendzel, M. J., Droit, A., Poirier, G. G. (2006). The expanding role of poly(ADP-ribose) metabolism: current challenges and new perspectives. Curr. Opin. Cell Biol. 18, 145–151. doi: 10.1016/j.ceb.2006.02.013
Gaucher, J., Boussouar, F., Montellier, E., Curtet, S., Buchou, T., Bertrand, S., et al. (2012). Bromodomain-dependent stage-specific male genome programming by Brdt. EMBO J. 31, 3809–3820. doi: 10.1038/emboj.2012.233
Gibson, B. A., Kraus, W. L. (2012). New insights into the molecular and cellular functions of poly(ADP-ribose) and PARPs. Nat. Rev. Mol. Cell Biol. 13, 411–424. doi: 10.1038/nrm3376
Godmann, M., Auger, V., Ferraroni-Aguiar, V., Di Sauro, A., Sette, C., Behr, R., et al. (2007). Dynamic regulation of histone h3 methylation at lysine 4 in mammalian spermatogenesis. Biol. Reprod. 77, 754–764. doi: 10.1095/biolreprod.107.062265
Gou, L. T., Kang, J. Y., Dai, P., Wang, X., Li, F., Zhao, S., et al. (2017). Ubiquitination-deficient mutations in human Piwi cause male infertility by impairing histone-to-protamine exchange during spermiogenesis. Obstet. Gynecol. Survey 72, 540–541. doi: 10.1097/OGX.0000000000000482
Govin, J., Caron, C., Lestrat, C., Rousseaux, S., Khochbin, S. (2004). The role of histones in chromatin remodelling during mammalian spermiogenesis. Eur. J. Biochem. 271, 3459–3469. doi: 10.1111/j.1432-1033.2004.04266.x
Govin, J., Dorsey, J., Gaucher, J., Rousseaux, S., Khochbin, S., Berger, S. L. (2010). Systematic screen reveals new functional dynamics of histones H3 and H4 during gametogenesis. Genes Dev. 24, 1772–1786. doi: 10.1101/gad.1954910
Govin, J., Escoffier, E., Rousseaux, S., Kuhn, L., Ferro, M., Thevenon, J., et al. (2007). Pericentric heterochromatin reprogramming by new histone variants during mouse spermiogenesis. J. Cell Biol. 176, 283–294. doi: 10.1083/jcb.200604141
Govin, J., Lestrat, C., Caron, C., Pivot-Pajot, C., Rousseaux, S., Khochbin, S. (2006). Histone acetylation-mediated chromatin compaction during mouse spermatogenesis. Cancer Ther. 57, 155–15+. doi: 10.1007/3-540-37633-X_9
Grimes, S. R., Henderson, N. (1984a). Acetylation of rat testis histones H2b and Th2b. Dev. Biol. 101, 516–521. doi: 10.1016/0012-1606(84)90165-9
Grimes, S. R., Jr., Henderson, N. (1984b). Hyperacetylation of histone H4 in rat testis spermatids. Exp. Cell Res. 152, 91–97. doi: 10.1016/0014-4827(84)90232-5
Hada, M., Masuda, K., Yamaguchi, K., Shirahige, K., Okada, Y. (2017). Identification of a variant-specific phosphorylation of TH2A during spermiogenesis. Sci. Rep. 7, 1–13. doi: 10.1038/srep46228
Hao, S. L., Ni, F. D., Yang, W. X. (2019). The dynamics and regulation of chromatin remodeling during spermiogenesis. Gene 706, 201–210. doi: 10.1016/j.gene.2019.05.027
Happel, N., Doenecke, D. (2009). Histone H1 and its isoforms: contribution to chromatin structure and function. Gene 431, 1–12. doi: 10.1016/j.gene.2008.11.003
Hazzouri, M., Pivot-Pajot, C., Faure, A. K., Usson, Y., Pelletier, R., Sele, B., et al. (2000). Regulated hyperacetylation of core histones during mouse spermatogenesis: involvement of histone-deacetylases. Eur. J. Cell Biol. 79, 950–960. doi: 10.1078/0171-9335-00123
Hershko, A., Ciechanover, A. (1998). The ubiquitin system. Annu. Rev. Biochem. 67, 425–479. doi: 10.1146/annurev.biochem.67.1.425
Hess, R. A., Renato De Franca, L. (2008). Spermatogenesis and cycle of the seminiferous epithelium. Adv. Exp. Med. Biol. 636, 1–15. doi: 10.1007/978-0-387-09597-4_1
Hoghoughi, N., Barral, S., Vargas, A., Rousseaux, S., Khochbin, S. (2018). Histone variants: essential actors in male genome programming. J. Biochem. 163, 97–103. doi: 10.1093/jb/mvx079
Hou, X., Zhang, W., Xiao, Z., Gan, H., Lin, X., Liao, S., et al. (2012). Mining and characterization of ubiquitin E3 ligases expressed in the mouse testis. BMC Genomics 13, 495. doi: 10.1186/1471-2164-13-495
Jha, K. N., Tripurani, S. K., Johnson, G. R. (2017). TSSK6 is required for gamma H2AX formation and the histone-to-protamine transition during spermiogenesis. J. Cell Sci. 130, 1835–1844. doi: 10.1242/jcs.202721
Juliano, C., Wang, J. Q., Lin, H. F. (2011). Uniting germline and stem cells: the function of Piwi proteins and the piRNA pathway in diverse organisms. Ann. Rev. Genet. 45, 447–469. doi: 10.1146/annurev-genet-110410-132541
Kan, P. Y., Caterino, T. L., Hayes, J. J. (2009). The H4 tail domain participates in intra- and internucleosome interactions with protein and DNA during folding and oligomerization of nucleosome arrays. Mol. Cell. Biol. 29, 538–546. doi: 10.1128/MCB.01343-08
Ketchum, C. C., Larsen, C. D., Mcneil, A., Meyer-Ficca, M. L., Meyer, R. G. (2018). Early histone H4 acetylation during chromatin remodeling in equine spermatogenesis. Biol. Reprod. 98, 115–129. doi: 10.1093/biolre/iox159
Khadake, J. R., Rao, M. R. S. (1995). DNA-condensing and chromatin-condensing properties of rat testes Hla and hit compared to those of rat-liver Hlbdec—Hlt is a poor condenser of chromatin. Biochemistry 34, 15792–15801. doi: 10.1021/bi00048a025
Khor, B., Bredemeyer, A. L., Huang, C. Y., Turnbull, I. R., Evans, R., Maggi, L. B., et al.(2006). Proteasome activator PA200 is required for normal spermatogenesis. Mol. Cell. Biol. 26, 2999–3007. doi: 10.1128/MCB.26.8.2999-3007.2006
Kimura, S., Loppin, B. (2016). The Drosophila chromosomal protein Mst77F is processed to generate an essential component of mature sperm chromatin. Open Biol. 6, 1–12. doi: 10.1098/rsob.160207
Kolthur-Seetharam, U., Pradeepa, M. M., Gupta, N., Narayanaswamy, R., Rao, M.R.S. (2009). Spatiotemporal organization of AT- and GC-rich DNA and their association with transition proteins TP1 and TP2 in rat condensing spermatids. J. Histochem. Cytochem. 57, 951–962. doi: 10.1369/jhc.2009.953414
Komander, D., Rape, M. (2012). The ubiquitin code. Ann. Rev. Biochem. 81, 203–229. doi: 10.1146/annurev-biochem-060310-170328
Kowalski, A., Palyga, J. (2012). Linker histone subtypes and their allelic variants. Cell Biol. Int. 36, 981–996. doi: 10.1042/CBI20120133
Krishnamoorthy, T., Chen, X., Govin, J., Cheung, W. L., Dorsey, J., Schindler, K., et al. (2006). Phosphorylation of histone H4 Ser1 regulates sporulation in yeast and is conserved in fly and mouse spermatogenesis. Genes Dev. 20, 2580–2592. doi: 10.1101/gad.1457006
Kuramochi-Miyagawa, S., Kimura, T., Ijiri, T. W., Isobe, T., Asada, N., Fujita, Y., et al. (2004). Mili, a mammalian member of piwi family gene, is essential for spermatogenesis. Development 131, 839–849. doi: 10.1242/dev.00973
Lee, J., Park, H. S., Kim, H. H., Yun, Y. J., Lee, D. R., Lee, S. (2009). Functional polymorphism in H2BFWT-5'UTR is associated with susceptibility to male infertility. J. Cell. Mol. Med. 13, 1942–1951. doi: 10.1111/j.1582-4934.2009.00830.x
Levesque, D., Veilleux, S., Caron, N., Boissonneault, G. (1998). Architectural DNA-binding properties of the spermatidal transition proteins 1 and 2. Biochem. Biophys. Res. Commun. 252, 602–609. doi: 10.1006/bbrc.1998.9687
Li, A., Eirin-Lopez, J., Ausio, J. (2005). H2AX: tailoring histone H2A for chromatin-dependent genomic integrity. Biochem. Cell Biol. 83, 505–515. doi: 10.1139/o05-114
Liu, C., Song, Z. H., Wang, L. N., Yu, H. Y., Liu, W. X., Shang, Y. L., et al. (2017a). Sirt1 regulates acrosome biogenesis by modulating autophagic flux during spermiogenesis in mice. Development 144, 441–451. doi: 10.1242/dev.147074
Liu, S. M., Yu, H. J., Liu, Y. Q., Liu, X. H., Zhang, Y., Bu, C., et al. (2017b). Chromodomain protein CDYL acts as a crotonyl-CoA hydratase to regulate histone crotonylation and spermatogenesis. Mol. Cell 67, 853–85+. doi: 10.1016/j.molcel.2017.07.011
Liu, Z. L., Zhou, S. L., Liao, L., Chen, X., Meistrich, M., Xu, J. M. (2010). Jmjd1a Demethylase-regulated histone modification is essential for cAMP-response element modulator-regulated gene expression and spermatogenesis. J. Biol. Chem. 285, 2758–2770. doi: 10.1074/jbc.M109.066845
Lu, L. Y., Wu, J. X., Ye, L., Gavrilina, G. B., Saunders, T. L., Yu, X. C. (2010). RNF8-dependent histone modifications regulate nucleosome removal during spermatogenesis. Dev. Cell 18, 371–384. doi: 10.1016/j.devcel.2010.01.010
Luense, L. J., Wang, X. S., Schon, S. B., Weller, A. H., Shiao, E. L., Bryant, J. M., et al.(2016). Comprehensive analysis of histone post-translational modifications in mouse and human male germ cells. Epigenet. Chromatin 9, 1–15. doi: 10.1186/s13072-016-0072-6
Ma, T., Keller, J. A., Yu, X. C. (2011). RNF8-dependent histone ubiquitination during DNA damage response and spermatogenesis. Acta Biochim. Biophys. Sin. 43, 339–345. doi: 10.1093/abbs/gmr016
Manterola, M., Brown, T. M., Oh, M. Y., Garyn, C., Gonzalez, B. J., Wolgemuth, D. J. (2018). BRDT is an essential epigenetic regulator for proper chromatin organization, silencing of sex chromosomes and crossover formation in male meiosis. PLoS Genet. 14, 1–30. doi: 10.1371/journal.pgen.1007209
Martianov, I., Brancorsini, S., Catena, R., Gansmuller, A., Kotaja, N., Parvinen, M., et al. (2005). Polar nuclear localization of H1T2, a histone H1 variant, required for spermatid elongation and DNA condensation during spermiogenesis. Proc. Natl Acad. Sci. U. S. A. 102, 2808–2813. doi: 10.1073/pnas.0406060102
Mccarrey, J. R., Geyer, C. B., Yoshioka, H. (2005). Epigenetic regulation of testis-specific gene expression. Testicular Cell Dyn. Endocr. Signal. 1061, 226–242. doi: 10.1196/annals.1336.025
Meistrich, M. L., Bucci, L. R., Trostleweige, P. K., Brock, W. A. (1985). Histone variants in rat spermatogonia and primary spermatocytes. Dev. Biol. 112, 230–240. doi: 10.1016/0012-1606(85)90137-X
Meistrich, M. L., Mohapatra, B., Shirley, C. R., Zhao, M. (2003). Roles of transition nuclear proteins in spermiogenesis. Chromosoma 111, 483–488. doi: 10.1007/s00412-002-0227-z
Meng, C., Liao, J., Zhao, D., Huang, H., Qin, J., Lee, T. L., et al. (2019). L3MBTL2 regulates chromatin remodeling during spermatogenesis. Cell Death Differ. 13, 1–14. doi: 10.1038/s41418-019-0283-z
Messner, S., Hottiger, M. O. (2011). Histone ADP-ribosylation in DNA repair, replication and transcription. Trends Cell Biol. 21, 534–542. doi: 10.1016/j.tcb.2011.06.001
Meyer-Ficca, M. L., Ihara, M., Bader, J. J., Leu, N. A., Beneke, S., Meyer, R. G. (2015). Spermatid head elongation with normal nuclear shaping requires ADP-ribosyltransferase PARP11 (ARTD11) in Mice. Biol. Reprod. 92, 1–13. doi: 10.1095/biolreprod.114.123661
Meyer-Ficca, M. L., Ihara, M., Lonchar, J. D., Meistrich, M. L., Austin, C. A., Min, W., et al. (2011). Poly(ADP-ribose) metabolism is essential for proper nucleoprotein exchange during mouse spermiogenesis. Biol. Reprod. 84, 218–228. doi: 10.1095/biolreprod.110.087361
Meyer-Ficca, M. L., Lonchar, J., Credidio, C., Ihara, M., Li, Y., Wang, Z. Q., et al.(2009). Disruption of poly(ADP-ribose) homeostasis affects spermiogenesis and sperm chromatin integrity in mice. Biol. Reprod. 81, 46–55. doi: 10.1095/biolreprod.108.075390
Meyer-Ficca, M. L., Scherthan, H., Burkle, A., Meyer, R. G. (2005). Poly(ADP-ribosyl)ation during chromatin remodeling steps in rat spermiogenesis. Chromosoma 114, 67–74. doi: 10.1007/s00412-005-0344-6
Mishra, L. N., Gupta, N., Rao, S. M. R. (2015). Mapping of post-translational modifications of spermatid-specific linker histone H1-like protein, HILS1. J. Proteomics 128, 218–230. doi: 10.1016/j.jprot.2015.08.001
Mishra, L. N., Shalini, V., Gupta, N., Ghosh, K., Suthar, N., Bhaduri, U., et al.(2018). Spermatid-specific linker histone HILS1 is a poor condenser of DNA and chromatin and preferentially associates with LINE-1 elements. Epigenet. Chromatin 11, 1–21. doi: 10.1186/s13072-018-0214-0
Montellier, E., Boussouar, F., Rousseaux, S., Zhang, K., Buchou, T., Fenaille, F., et al. (2013). Chromatin-to-nucleoprotamine transition is controlled by the histone H2B variant TH2B. Genes Dev. 27, 1680–1692. doi: 10.1101/gad.220095.113
Moss, S. B., Orth, J. M. (1993). Localization of a spermatid-specific histone-2b protein in mouse spermiogenic cells. Biol. Reprod. 48, 1047–1056. doi: 10.1095/biolreprod48.5.1047
Nair, M., Nagamori, I., Sun, P., Mishra, D. P., Rheaume, C., Li, B., et al. (2008). Nuclear regulator Pygo2 controls spermiogenesis and histone H3 acetylation. Dev. Biol. 320, 446–455. doi: 10.1016/j.ydbio.2008.05.553
Okada, Y., Scott, G., Ray, M. K., Mishina, Y., Zhang, Y. (2007). Histone demethylase JHDM2A is critical for Tnp1 and Prm1 transcription and spermatogenesis. Nature 450, 119–11+. doi: 10.1038/nature06236
Oliva, R., Bazett-Jones, D., Mezquita, C., Dixon, G. H. (1987). Factors affecting nucleosome disassembly by protamines in vitro. Histone hyperacetylation and chromatin structure, time dependence, and the size of the sperm nuclear proteins. J. Biol. Chem. 262, 17016–17025.
Oliva, R., Mezquita, C. (1986). Marked differences in the ability of distinct protamines to disassemble nucleosomal core particles in vitro. Biochemistry 25, 6508–6511. doi: 10.1021/bi00369a025
Padavattan, S., Shinagawa, T., Hasegawa, K., Kumasaka, T., Ishii, S., Kumarevel, T. (2015). Structural and functional analyses of nucleosome complexes with mouse histone variants TH2a and TH2b, involved in reprogramming. Biochem. Biophys. Res. Commun. 464, 929–935. doi: 10.1016/j.bbrc.2015.07.070
Padavattan, S., Thiruselvam, V., Shinagawa, T., Hasegawa, K., Kumasaka, T., Ishii, S., et al. (2017). Structural analyses of the nucleosome complexes with human testis-specific histone variants, hTh2a and hTh2b. Biophys. Chem. 221, 41–48. doi: 10.1016/j.bpc.2016.11.013
Pentakota, S. K., Sandhya, S., Sikarwar, A. P., Chandra, N., Rao, M. R. S. (2014). Mapping post-translational modifications of mammalian testicular specific histone variant TH2B in tetraploid and haploid germ cells and their implications on the dynamics of nucleosome structure. J. Proteome Res. 13, 5603–5617. doi: 10.1021/pr500597a
Pickart, C. M. (2001). Mechanisms underlying ubiquitination. Annu. Rev. Biochem. 70, 503–533. doi: 10.1146/annurev.biochem.70.1.503
Pivot-Pajot, C., Caron, C., Govin, M., Vion, A., Rousseaux, S., Khochbin, S. (2003). Acetylation-dependent chromatin reorganization by BRDT, a testis-specific bromodomain-containing protein. Mol. Cell. Biol. 23, 5354–5365. doi: 10.1128/MCB.23.15.5354-5365.2003
Qian, M. X., Pang, Y., Liu, C. H., Haratake, K., Du, B. Y., Ji, D. Y., et al. (2013). Acetylation-mediated proteasomal degradation of core histones during DNA repair and spermatogenesis. Cell 153, 1012–1024. doi: 10.1016/j.cell.2013.04.032
Rafatmanesh, A., Nikzad, H., Ebrahimi, A., Karimian, M., Zamani, T. (2018). Association of the c.-9C > T and c.368A > G transitions in H2BFWT gene with male infertility in an Iranian population. Andrologia 50, 1–6. doi: 10.1111/and.12805
Raja, S. J., Renkawitz-Pohl, R. (2005). Replacement by Drosophila melanogaster protamines and Mst77F of histones during chromatin condensation in late spermatids and role of sesame in the removal of these proteins from the male pronucleus. Mol. Cell. Biol. 25, 6165–6177. doi: 10.1128/MCB.25.14.6165-6177.2005
Rathke, C., Baarends, W. M., Awe, S., Renkawitz-Pohl, R. (2014). Chromatin dynamics during spermiogenesis. Biochim. Biophys. Acta 1839, 155–168. doi: 10.1016/j.bbagrm.2013.08.004
Roosen-Runge, E. C. (1962). The process of spermatogenesis in mammals. Biol. Rev. Camb Philos. Soc. 37, 343–377. doi: 10.1111/j.1469-185X.1962.tb01616.x
Rossetto, D., Avvakumov, N., Cote, J. (2012). Histone phosphorylation A chromatin modification involved in diverse nuclear events. Epigenetics 7, 1098–1108. doi: 10.4161/epi.21975
Sabari, B. R., Zhang, D., Allis, C. D., Zhao, Y. M. (2017). Metabolic regulation of gene expression through histone acylations. Nat. Rev. Mol. Cell Biol. 18, 90–101. doi: 10.1038/nrm.2016.140
Sarg, B., Chwatal, S., Talasz, H., Lindner, H. H. (2009). Testis-specific linker histone H1t is multiply phosphorylated during spermatogenesis identification of phosphorylation siTES. J. Biol. Chem. 284, 3610–3618. doi: 10.1074/jbc.M805925200
Shang, E. Y., Nickerson, H. D., Wen, D. C., Wang, X. Y., Wolgemuth, D. J. (2007). The first bromodomain of Brdt, a testis-specific member of the BET sub-family of double-bromodomain-containing proteins, is essential for male germ cell differentiation. Development 134, 3507–3515. doi: 10.1242/dev.004481
Shinagawa, T., Huynh, L. M., Takagi, T., Tsukamoto, D., Tomaru, C., Kwak, H. G., et al. (2015). Disruption of Th2a and Th2b genes causes defects in spermatogenesis. Development 142, 1287–1292. doi: 10.1242/dev.121830
Shiraishi, K., Shindo, A., Harada, A., Ohkawa, Y., Kurumizaka, H., Kimura, H., et al.(2017). Roles of histone H3.5 in human spermatogenesis and spermatogenic disorders. J. Urol. 197, E85–E85. doi: 10.1016/j.juro.2017.02.277
Shires, A., Carpenter, M. P., Chalkley, R. (1975). New histones found in mature mammalian testes. Proc. Natl. Acad. Sci. U. S. A. 72, 2714–2718. doi: 10.1073/pnas.72.7.2714
Shires, A., Carpenter, M. P., Chalkley, R. (1976). Cysteine-containing H2b-like histone found in mature mammalian testis. J. Biol. Chem. 251, 4155–4158.
Shirley, C. R., Hayashi, S., Mounsey, S., Yanagimachi, R., Meistrich, M. L. (2004). Abnormalities and reduced reproductive potential of sperm from Tnp1- and Tnp2-null double mutant mice. Biol. Reprod. 71, 1220–1229. doi: 10.1095/biolreprod.104.029363
Shogren-Knaak, M., Ishii, H., Sun, J. M., Pazin, M. J., Davie, J. R., Peterson, C. L. (2006). Histone H4-K16 acetylation controls chromatin structure and protein interactions. Science 311, 844–847. doi: 10.1126/science.1124000
Singh, J., Rao, M. R. S. (1988). Interaction of rat testis protein, Tp, with nucleosome core particle. Biochem. Int. 17, 701–710.
Siomi, M. C., Sato, K., Pezic, D., Aravin, A. A. (2011). PIWI-interacting small RNAs: the vanguard of genome defence. Nat. Rev. Mol. Cell Biol. 12, 246–258. doi: 10.1038/nrm3089
Soboleva, T. A., Nekrasov, M., Pahwa, A., Williams, R., Huttley, G. A., Tremethick, D. J. (2012). A unique H2A histone variant occupies the transcriptional start site of active genes. Nat. Struct. Mol. Biol. 19, 25–U37. doi: 10.1038/nsmb.2161
Soboleva, T. A., Parker, B. J., Nekrasov, M., Hart-Smith, G., Tay, Y. J., Tng, W. Q., et al. (2017). A new link between transcriptional initiation and pre-mRNA splicing: the RNA binding histone variant H2A.B. PloS Genet. 13, 1–31. doi: 10.1371/journal.pgen.1006633
Song, N., Liu, J., An, S. C., Nishino, T., Hishikawa, Y., Koji, T. (2011). Immunohistochemical analysis of histone H3 modifications in germ cells during mouse spermatogenesis. Acta Histochem. Cytochem. 44, 183–190. doi: 10.1267/ahc.11027
Spiridonov, N. A., Wong, L., Zerfas, P. M., Starost, M. F., Pack, S. D., Paweletz, C. P., et al. (2005). Identification and characterization of SSTK, a serine/threonine protein kinase essential for male fertility. Mol. Cell. Biol. 25, 4250–4261. doi: 10.1128/MCB.25.10.4250-4261.2005
Stadtmueller, B. M., Hill, C. P. (2011). Proteasome Activators. Mol. Cell 41, 8–19. doi: 10.1016/j.molcel.2010.12.020
Tachiwana, H., Kagawa, W., Osakabe, A., Kawaguchi, K., Shiga, T., Hayashi-Takanaka, Y., et al. (2010). Structural basis of instability of the nucleosome containing a testis-specific histone variant, human H3T. Proc. Natl. Acad. Sci. U. S. A. 107, 10454–10459. doi: 10.1073/pnas.1003064107
Talbert, P. B., Henikoff, S. (2010). Histone variants - ancient wrap artists of the epigenome. Nat. Rev. Mol. Cell Biol. 11, 264–275. doi: 10.1038/nrm2861
Tan, M. J., Luo, H., Lee, S., Jin, F. L., Yang, J. S., Montellier, E., et al. (2011). Identification of 67 histone marks and histone lysine crotonylation as a new type of histone modification. Cell 146, 1015–1027. doi: 10.1016/j.cell.2011.08.008
Tanaka, H., Iguchi, N., Isotani, A., Kitamura, K., Toyama, Y., Matsuoka, Y., et al. (2005). HANP1/H1T2, a novel histone H1-like protein involved in nuclear formation and sperm fertility. Mol. Cell. Biol. 25, 7107–7119. doi: 10.1128/MCB.25.16.7107-7119.2005
Tang, M. C. W., Jacobs, S. A., Mattiske, D. M., Soh, Y. M., Graham, A. N., Tran, A., et al. (2015). Contribution of the two genes encoding histone variant H3.3 to viability and fertility in mice. PloS Genet. 11, 1–23. doi: 10.1371/journal.pgen.1004964
Teimouri, M., Najaran, H., Hosseinzadeh, A., Mazoochi, T. (2018). Association between two common transitions of H2BFWT gene and male infertility: a case-control, meta, and structural analysis. Andrology 6, 306–316. doi: 10.1111/andr.12464
Thakar, A., Gupta, P., Ishibashi, T., Finn, R., Silva-Moreno, B., Uchiyama, S., Fukui, et al. (2009). H2A.Z and H3.3 histone variants affect nucleosome structure: biochemical and biophysical studies. Biochemistry 48, 10852–10857. doi: 10.1021/bi901129e
Trostleweige, P. K., Meistrich, M. L., Brock, W. A., Nishioka, K., Bremer, J. W. (1982). Isolation and characterization of Th2a, a germ cell-specific variant of histone-2a in rat testis. J. Biol. Chem. 257, 5560–5567.
Tse, C., Sera, T., Wolffe, A. P., Hansen, J. C. (1998). Disruption of higher-order folding by core histone acetylation dramatically enhances transcription of nucleosomal arrays by RNA polymerase III. Mol. Cell. Biol. 18, 4629–4638. doi: 10.1128/MCB.18.8.4629
Ueda, J., Harada, A., Urahama, T., Machida, S., Maehara, K., Hada, M., et al.(2017). Testis-specific histone variant H3t gene is essential for entry into spermatogenesis. Cell Rep. 18, 593–600. doi: 10.1016/j.celrep.2016.12.065
Unni, E., Zhang, Y., Kangasniemi, M., Saperstein, W., Moss, S. B., Meistrich, M. L. (1995). Stage-specific distribution of the spermatid-specific histone 2b in the rat testis. Biol. Reprod. 53, 820–826. doi: 10.1095/biolreprod53.4.820
Urahama, T., Harada, A., Maehara, K., Horikoshi, N., Sato, K., Sato, Y., et al.(2016). Histone H3.5 forms an unstable nucleosome and accumulates around transcription start sites in human testis. Epigenet. Chromatin 9, 1–16. doi: 10.1186/s13072-016-0051-y
Urahama, T., Horikoshi, N., Osakabe, A., Tachiwana, H., Kurumizaka, H. (2014). Structure of human nucleosome containing the testis-specific histone variant TSH2B. Acta Crystallogr. Sect. F-Struct. Biol. Commun. 70, 444–449. doi: 10.1107/S2053230X14004695
Ushijima, Y., Inoue, Y. H., Konishi, T., Kitazawa, D., Yoshida, H., Shimaji, K., et al. (2012). Roles of histone H3K9 methyltransferases during Drosophila spermatogenesis. Chrom. Res. 20, 319–331. doi: 10.1007/s10577-012-9276-1
Ustrell, V., Pratt, G., Gorbea, C., Rechsteiner, M. (2005). Purification and assay of proteasome activator PA200. Methods Enzymol. 398, 321–329. doi: 10.1016/S0076-6879(05)98026-9
Van Der Heijden, G. W., Derijck, A. a. H. A., Posfai, E., Giele, M., Pelczar, P., Ramos, L., et al. (2007). Chromosome-wide nucleosome replacement and H3.3 incorporation during mammalian meiotic sex chromosome inactivation. Nat. Genet. 39, 251–258. doi: 10.1038/ng1949
Wang, X., Kang, J. Y., Wei, L., Yang, X., Sun, H., Yang, S., et al. (2019). PHF7 is a novel histone H2A E3 ligase prior to histone-to-protamine exchange during spermiogenesis. Development 146, 1–11. doi: 10.1242/dev.175547
Welchman, R. L., Gordon, C., Mayer, R. J. (2005). Ubiquitin and ubiquitin-like proteins as multifunctional signals. Nat. Rev. Mol. Cell. Biol. 6, 599–609. doi: 10.1038/nrm1700
Wendt, K. D., Shilatifard, A. (2006). Packing for the germy: the role of histone H4 Ser1 phosphorylation in chromatin compaction and germ cell development. Genes Dev. 20, 2487–2491. doi: 10.1101/gad.1477706
Wu, J. Y., Ribar, T. J., Cummings, D. E., Burton, K. A., Mcknight, G. S., Means, A. R. (2000). Spermiogenesis and exchange of basic nuclear proteins are impaired in male germ cells lacking Camk4. Nat. Genet. 25, 448–452. doi: 10.1038/78153
Yan, W., Ma, L., Burns, K. H., Matzuk, M. M. (2003). HILS1 is a spermatid-specific linker histone H1-like protein implicated in chromatin remodeling during mammalian spermiogenesis. Proc. Natl. Acad. Sci. U. S. A. 100, 10546–10551. doi: 10.1073/pnas.1837812100
Ying, H. Q., Scott, M. B., Zhou-Cun, A. (2012). Relationship of SNP of H2BFWT gene to male infertility in a Chinese population with idiopathic spermatogenesis impairment. Biomarkers 17, 402–406. doi: 10.3109/1354750X.2012.677066
Yoshida, K., Muratani, M., Araki, H., Miura, F., Suzuki, T., Dohmae, N., et al. (2018). Mapping of histone-binding sites in histone replacement-completed spermatozoa. Nat. Commun. 9, 1–11. doi: 10.1038/s41467-018-06243-9
Yu, Y. E., Zhang, Y., Unni, E., Shirley, C. R., Deng, J. M., Russell, L. D., et al. (2000). Abnormal spermatogenesis and reduced fertility in transition nuclear protein 1-deficient mice. Proc. Natl. Acad. Sci. U. S. A. 97, 4683–4688. doi: 10.1073/pnas.97.9.4683
Yuen, B.T.K., Bush, K. M., Barrilleaux, B. L., Cotterman, R., Knoepfler, P. S. (2014). Histone H3.3 regulates dynamic chromatin states during spermatogenesis. Development 141, 3483–3494. doi: 10.1242/dev.106450
Zhang, Z. H., Mu, S. M., Guo, M. S., Wu, J. L., Li, Y. Q., Zhang, H., et al. (2016). Dynamics of histone H2A, H4 and HS1ph during spermatogenesis with a focus on chromatin condensation and maturity of spermatozoa. Sci. Rep. 6, 1–11. doi: 10.1038/srep25089
Zhao, M., Shirley, C. R., Mounsey, S., Meistrich, M. L. (2004). Nucleoprotein transitions during spermiogenesis in mice with transition nuclear protein Tnp1 and Tnp2 mutations. Biol. Reprod. 71, 1016–1025. doi: 10.1095/biolreprod.104.028191
Zhao, M., Shirley, C. R., Yu, Y. E., Mohapatra, B., Zhang, Y., Unni, E., et al. (2001). Targeted disruption of the transition protein 2 gene affects sperm chromatin structure and reduces fertility in mice. Mol. Cell Biol. 21, 7243–7255. doi: 10.1128/MCB.21.21.7243-7255.2001
Keywords: spermiogenesis, histone-to-protamine transition, histone variants, histone modification, male infertility
Citation: Wang T, Gao H, Li W and Liu C (2019) Essential Role of Histone Replacement and Modifications in Male Fertility. Front. Genet. 10:962. doi: 10.3389/fgene.2019.00962
Received: 08 May 2019; Accepted: 10 September 2019;
Published: 08 October 2019.
Edited by:
Kyoko Yokomori, University of California, United StatesReviewed by:
Alexander Kouzmenko, Tokiwa Foundation, JapanCopyright © 2019 Wang, Gao, Li and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wei Li, bGV3YXlzQGlvei5hYy5jbg==; Chao Liu, bGl1Y2hhb0Bpb3ouYWMuY24=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.