 Pedro Góes Mesquita
Pedro Góes Mesquita Laiza Magalhaes de Araujo
Laiza Magalhaes de Araujo Francisco de Assis Rocha Neves
Francisco de Assis Rocha Neves Maria de Fátima Borin
Maria de Fátima Borin- Laboratory of Molecular Pharmacology, Department of Pharmacy, Faculty of Sciences Health, University of Brasilia, Brasilia, Brazil
Diabetes mellitus is a metabolic disorder that affects millions of people worldwide and is linked to oxidative stress and inflammation. Thiazolidinediones (TZD) improve insulin sensitization and glucose homeostasis mediated by the activation of peroxisome proliferator-activated receptors γ (PPARγ) in patients with type 2 diabetes. However, their use is associated with severe adverse effects such as loss of bone mass, retention of body fluids, liver and heart problems, and increased risk of bladder cancer. Partial PPARγ agonists can promote the beneficial effects of thiazolidinediones with fewer adverse effects. Endophytic fungi colonize plant tissues and have a particularly active metabolism caused by the interaction with them, which leads to the production of natural products with significant biological effects that may be like that of the colonized plant. Here, we identify seven endophytic fungi isolated from Bauhinia variegata leaves that have antioxidant activities. Also, one of the extracts presented pan-agonist activity on PPAR, and another showed activity in PPARα and PPARβ/δ. A better understanding of this relationship could help to comprehend the mechanism of action of antioxidants in treating diabetes and its complications. Moreover, compounds with these capabilities to reduce oxidative stress and activate the receptor that promotes glucose homeostasis are promising candidates in treatment of diabetes.
1. Introduction
Diabetes mellitus is a metabolic disorder characterized by chronic hyperglycemia with disturbance in the metabolism of carbohydrates, lipids, and proteins. It results from a deficiency in insulin secretion, insulin action, or both, causing glucose to be unavailable in the muscles and adipose tissue and increasing gluconeogenesis in the liver (Alberti and Zimmet, 1998). Chronic hyperglycemia causes major vascular complications and is involved in developing heart disease, retinopathy, kidney disease, and circulatory problems. The development of diabetes can occur by the autoimmune destruction of beta cells, leading to a deficiency in insulin production, or by other factors that can result in tissue resistance to the action of insulin. In type 2 diabetes, insulin secretion and action deficiency are usually present in the same individual, which is the leading cause of hyperglycemia (Ada, 2014). When there is a clinical manifestation, individuals usually have insulin resistance, insulin deficiency, or both. The specific causes of type 2 diabetes are not yet defined; however, most patients are obese or have a high concentration of abdominal fat. Obesity causes a continuous elevation in plasma levels of free fatty acids, which induces hyperglycemia and impairs insulin sensitivity. The elevation of insulin secretion is a compensatory mechanism for maintaining glycemia. However, very high insulin levels do not allow compensation for the deficiency in glucose uptake and lead to insulin resistance and diabetes. Furthermore, obesity, recent weight gain, and the length of time one have been obese influence the increased risk of insulin resistance (Alberti and Zimmet, 1998; Felber and Golay, 2002; Ada, 2014).
In obesity, weight gain occurs when calorie intake exceeds energy expenditure when there is no energy balance, and the excess is stored in adipose tissue in the form of fat (lipids). The influence of adipose tissue on metabolic syndrome is central. First regarded as an energy storage tissue, it is now known that adipose tissue plays a fundamental role as an endocrine organ, capable of secreting molecules such as leptin and influencing insulin resistance, inflammation, eating behavior, and energy expenditure. It can communicate with other tissues in the body, such as muscle, liver, and the hunger center in the brain, and is in visceral and subcutaneous deposits. The accumulation of visceral adipose tissue (VAT) is linked, as well as type 2 diabetes, to other pathologies, such as heart disease, kidney, lung, and liver dysfunction, dyslipidemia, and hyperglycemia. In subcutaneous adipose tissue (SAT), it is associated with a low risk of these diseases (Majka et al., 2011).
Chronic systemic inflammation in adipose tissue is directly associated with the development of insulin resistance and type 2 diabetes, a feature of the chronic hyperglycemic state (Xu et al., 2003; Oguntibeju, 2019). Treatment usually is performed with diet, physical exercise, and antihyperglycemic agents using the following classes of drugs: biguanides, which suppress the production of glucose in the liver; glitazones, which improve insulin sensitivity by binding to PPARγ and increasing adiponectin signaling pathways; sulfonylureas and glinides, which stimulate insulin secretion; GLP1 receptor agonists (glucagon-like peptide 1), which mimic incretins, potentiating glucose-stimulated insulin secretion, decreasing glucagon secretion, promoting a feeling of satiety and decreasing gastric emptying; gliptins that potentiate the action of incretins and prevent the degradation of GLP1 by inhibiting DDP-4 (dipeptidyl peptidase 4); and alpha-glucosidase inhibitors that decrease glucose absorption in the intestine (Marín-Peñalver et al., 2016).
Oxidation and free radicals are part of the natural physiological process in multicellular organisms. Free radicals are molecules, ions, or atoms that have an unpaired electron, increasing their ability to react with other molecules. There is a very fragile balance between the production of radical compounds and their neutralization by antioxidant compounds. An imbalance known as oxidative stress occurs, which usually leads to physiological dysfunction (Halliwell, 2011). Oxidative stress is present in most processes involved in the development of diabetes. In the pancreas, the increase in reactive oxygen species (ROS) production by insulin-producing β-cells can occur through several pathways, including increasing the excessive concentration of glucose and free fatty acids (King and Loeken, 2004; Evans et al., 2005; Lupi et al., 2007; Zunino, 2009). The regulatory mechanisms involved in the pathological processes of diabetes and atherosclerosis in a correlation between oxidative stress and activation of peroxisomal proliferating receptors have been shown (Devchand et al., 2004; Hwang et al., 2004; Teissier et al., 2004; Jiang et al., 2009; Martin, 2010; Guellich et al., 2013). Polyphenols are the most abundant antioxidant substances in the diet and can be produced by plants and fungi (Manach et al., 2004). Flavonoids are polyphenols with diverse physiological and pharmacological activities, such as estrogenic, antitumor, antimicrobial, antiallergic, anti-inflammatory, and the well-known antioxidant and metal chelation (Cotelle, 2001; Reiterer et al., 2004; Das and Rosazza, 2006; Nazari et al., 2011). These substances have shown a beneficial effect concerning diabetes mellitus, acting in several ways, such as regulating glucose biochemical pathways, glucose tolerance, lipid profile, glycogen biosynthesis, glucose uptake, and insulin release (Volpato et al., 2008; Da Cunha et al., 2009). In addition, a meta-analysis published showed that dietary flavonoid intake was associated with a decrease in type 2 diabetes (Liu et al., 2014).
New drugs with better efficacy in diabetes treatment would benefit a large population. In this endeavor, endophytic fungi are promising. They are microorganisms belonging to several orders of fungi, which colonize the living and internal tissues of plants, intracellularly or extracellularly, in a relationship that varies from symbiosis to the borderline of pathogenicity (Saikkonen et al., 1998; Gallo et al., 2007). These organisms usually produce a wide variety of organic compounds due to the close metabolic interaction with their hosts, which results in a particularly active secondary metabolism (Gallo et al., 2007). In some cases, the host plant and the fungus can produce the same metabolites, as shown by the endophytic fungus Taxomyces andreanae, capable of producing the anticancer taxol (Paclitaxel®), also produced by its host, Taxus brevifolia (Stierle et al., 1993).
Bauhinia sp. plants are used in traditional and folk medicine and are reported to possess antidiabetic and anti-inflammatory activities (Zakaria et al., 2007; Prabhakar and Doble, 2008; Frankish et al., 2010). Here, we identified seven endophytic fungi isolated from Bauhinia variegata leaves and evaluated their ethyl acetate (EtOAc) extract activities over PPARα, PPARβ/δ, and PPARγ receptors and their antioxidant profile.
2. Material and methods
2.1. Endophytic fungi isolation and culture
Bauhinia variegata leaves were collected in Brasília, DF, in July. Leaves were cleaned and disinfected, and endophytic fungi were isolated according to the methodology described by Larran e coworkers (2007). Isolated fungi were grown at Sabouraud-dextrose agar medium in slants. After five days of incubation at 30°C, cultures were transferred to 500 mL flasks containing 50 mL of pre-fermentative medium described by Jackson et al. (1993), incubated at 28°C, on an orbital shaker incubator at 150 rpm for 48 h. After this period, cultures were transferred to 2000 mL flasks containing 500 mL of fermentative medium and cultured for 72 h, at 28°C, with an orbital stirring of 150 rpm.
2.2. Extract production
The mycelial mass was separated from the submerged culture by filtration under a vacuum. The fluids of the cultures were then submitted to a liquid-liquid extraction using ethyl acetate as solvent (3 times the volume) to extract the fungi metabolites. The extract was then concentrated by evaporation under a vacuum and dissolved in 1 mL of methanol. These extracts were used to evaluate antioxidant activity and polyphenols and flavonoid contents.
2.3. DNA extraction and amplification of the ITS1-5,8S-ITS2 rRNA sequences
Endophytic fungi isolated from Bauhinia variegata leaves were identified by their DNA ribosomal sequences after amplification by polymerase chain reaction (PCR) technique.
Fungi were cultured in Yeast Peptone Dextrose medium, as previously described by (White et al., 1990) at 30°C for 48 hours in a rotating shaker at 120 rpm. Two grams of the cultured fungi were submerged in liquid nitrogen, powdered, and transferred to microtubes. The samples were added with 500 µL Tris buffer, pH 8, containing 10 mmol.L-1 EDTA and 2% SDS, followed by vortexing. Genomic DNA was isolated by the phenol/chloroform method. After that, isolated DNA was solubilized in deionized water and quantified according to the method described by Sambrook et al. (1989). The portion 5.8S of the ribosomal RNA was amplified using the primers of the conserved regions ITS1-ITS2 and ITS3-ITS4 (White et al., 1990). Amplification was carried out using a final solution reagent volume of 50 µL, containing 1 µL of genomic DNA (500 ng), 2.5 µL DNTP (4 mmol.L-1), 0.5 µL Taq polymerase (0.5 U.µL-1), 5 µL of Tris/HCl buffer (100 mmol.L-1 pH 8.0), 2 µL 50 mmol.L-1 MgCl2, 1 µL of each universal primer, ITS1-ITS2 or ITS3-ITS4 (21 pmol.L-1), and 39 µL of deionized water. Thermocycler was programmed with initial DNA denaturing cycle at 94°C for 2 minutes, followed by cycling at 94°C for 1 min, 54°C for 1 min, 72°C for 1 min, and ending with a final step at 72°C for 10 min (Guimarães et al., 2008). PCR products were purified using a QIAquick® PCR Purification Kit, and their sequences were analyzed by Macrogen Inc. in the Republic of Korea. Either genomic DNA or PCR amplification products were analyzed in agarose 1% electrophoresis to determine the chain size (Möller et al., 1992), using 1kb plus DNA ladder (Life Thecnologies®) as a marker.
2.4. Phylogenetic analysis
PCR amplicons sequences were edited at BioEdit and compared, via BLAST, to sequences deposited in the GenBank database of the NCBI (National Center for Biotechnology Information). The sequences obtained and selected from GenBank (NCBI) were submitted to multiple alignments using the Clustal-W tool in the MEGA 11.0.13 software. The similarity was analyzed by grouping the neighbor-joining method with 1,000 bootstrapping repetitions to construct the phylogenetic tree (Tamura et al., 2021).
2.5. Determination of total polyphenolic content
Total polyphenols were quantified by the method described by Kumazawa et al. (2002) with minor modifications. For that, 76.8 µg of each extract was added to 250 µL of 10% sodium carbonate and 250 µL of Folin-Ciocalteu 1N. The final reaction solution volume was completed to 2.5 mL with distilled water, and the solution was vortexed. Phenols react with the Folin-Ciocalteu reagent producing a bluish-grey complex. After 60 minutes, absorbance was measured in a spectrophotometer at 760 nm. Gallic acid was used as standard, and results were expressed as gallic acid equivalents by extract mass (EAG.mg-1).
2.6. Determination of total flavonoid content
Determination of total flavonoid content was carried out according to the method described by Chang et al. (2002), based on forming an acid-stable yellow complex between aluminum chloride and flavones/flavonoids. In brief, 125 µL of 5% aluminum chloride in a methanol solution was added to 20 µL of the sample. The final volume was adjusted to 2.5 mL with methanol and vortexed. After 30 minutes, the absorbance was measured in a spectrophotometer at 425 nm. Quercetin was used as standard, and results were expressed as quercetin equivalents by mass of extracts (EQ.mg-1).
2.7. In vitro antioxidant activity determination
2.7.1. DPPH· assay
The antioxidant activity of the fungi extract was determined using DPPH· (2,2-diphenyl-1-picrylhydrazyl) assay, as described by Blois (1958). Negative control was obtained by replacing the sample with the same volume of methanol in the reaction solution. Antioxidant activity, determined by measurement of the percentage of reduction of DPPH·, considering the absorbance of the negative control as 100%, was estimated based on the formula below:
Where:
AA: antioxidant activity (%)
ABScontrol: absorbance of the negative control at 517 nm.
ABSsample: absorbance of the sample at 517 nm.
The percentage of antioxidant activity was evaluated by comparing the results obtained for different concentrations of extracts plotted in a graphic and calculating the IC50 (concentration of the extract that decreases by 50% the oxidation of DPPH·) (Dinis et al., 1994).
2.7.2. Evaluation of the capability of the extracts in inhibiting lipid peroxidation
To evaluate the capability of the extract to inhibit lipid peroxidation, the method described by Lingnert et al. (1979) was used with some modifications. Briefly, linoleic acid was emulsified with the same volume of Tween® 20 in 0.1 M phosphate buffer, pH 7.0. For 2 mL of linoleic acid/buffer emulsion, 100 µL of samples were added, and the reaction solution was incubated for 8 hours at 37°C, hidden from the light. Then, 100 µL of this reaction solution was solubilized into 1 mL of methanol and added with 3 mL of 60% methanol solution, stirred. The absorbance was determined at 234 nm in a spectrophotometer (Shimadzu UV/VIS Model-1800).
2.7.3. Superoxide anion radical scavenging activity
The superoxide anion radical () scavenging activity by extracts was measured by determination of the inhibition of the NBT (Nitro Blue Tetrazolium) reduction. The xanthine/xanthine oxidase system was used as a superoxide anion radical source. The reaction was monitored by spectrophotometry at 560 nm for 10 minutes, modifying the method described by Robak and Gryglewski (1988). Results were expressed as sample concentration (mg.mL-1) able to inhibit 50% of the NBT oxidation rate acquired for the control reaction solution (IC50).
2.8. Cell culture
For the reporter gene assays, human mesangial cells donated by the Discipline of Nephrology at Escola Paulista de Medicina, Universidade Federal de São Paulo (UNIFESP) were used. Cells were cultured according to the method described by Chiellini and collaborators (Chiellini et al., 1998).
These cells were grown in Dulbecco’s Modified Eagle Medium (DMEM), containing 10% fetal bovine serum, 2 nmol.L-1 glutamine, 50 IU.mL-1 penicillin, and 55 IU.mL-1 streptomycin, and kept in 148 cm2 Corning® plates in an incubator at 37° C with 5% CO2.
2.9. In vitro evaluation of the cytotoxicity of extracts on mesangial cells
Cell viability and cytotoxicity of the extracts were determined according to the 3-(4,5-dimethylthiazol-2- yl) -2,5-diphenyl-tetrazolium (MTT) method. The tetrazolium ring present in the yellowish-colored MTT bromide salt is cleaved by dehydrogenases present in the mitochondria of viable cells, forming blue-colored (Mosmann, 1983). Therefore, it is considered that the amount of viable cells is proportional to the amount of formazan crystals formed. In the assay, plates were seeded with 30,500 human mesangial cells per well in the DMEM culture medium, except for the well used as a reaction blank, which contained only the culture medium. The final reaction volume was 100 µL. After 24 hours of incubation at 37°C and 5% CO2, 10 µL of MTT [5 mg.mL-1 in phosphate-buffered saline (PBS)] were added to all wells of the plates, and they were incubated for 4 hours, at 37°C and 5% CO2. For the dissolution of the formazan salt, the culture medium was removed from the wells, and 100 µL of MTT developer solution (0.04 mol.L-1 HCl in isopropanol) was added. The plates were left under agitation for 15 minutes at room temperature, and the absorbance was measured at 570 nm with a plate reader (Beckman Coulter DTX 800). The interpretation of the results was made by comparing the absorbance values obtained for cells treated with the extracts and those obtained in the controls.
2.10. PPAR receptor agonist activity – reporter gene assay
The reporter gene assay employing transient transfection was used in the research and identification of ligands for nuclear receptors (PPARα, PPARβ/δ, and PPARγ). This assay consisted of transfection by electroporation of expression and reporter plasmids inside the nucleus of human mesangial cells, followed by treatment of the cells with a known ligand (positive control) or with the extracts under study. In the presence of agonist substances, the reporter gene (luciferase) had its transcription increased and functioned as an indicator of the receptor’s transcriptional activity.
Transfection assay, cells were collected by centrifugation at 760.1 x g for 5 minutes and resuspended in PBS solution containing 0.1% dextrose and 0.1% calcium chloride to obtain a suspension containing 3x107 cells.mL-1. To each 500 µL of this solution of resuspended cells, 1.5 μg of the PPARα, PPARβ/δ, or PPARγ expression vector and 3 μg of the PPAR response element (PPRE) reporter plasmid vector were added. Cells were transferred to cuvettes and subjected to electroporation at 350 mV and 700 F using a Bio-Rad® Gene Pulser II pulse generator. After electroporation, the cells were transferred to the culture medium, distributed in 12-well Corning® plates (1 mL/well), and treated with vehicle, positive control, or different concentrations of the extracts based on the cytotoxicity assay.
After 24 hours of incubation at 37°C and 5% CO2, cells were separated by centrifugation at 760.1 x g and lysed with 1X lysis buffer (Promega®). To determine luciferase activity, 20 μL of luciferin (Kit Promega Corp®) was added to 20 μL of cell lysate. The light emission generated by the enzymatic reaction between luciferin and luciferase was quantified in a luminometer GloMax® 20/20, and the result was expressed in relative light units. All assays were performed in triplicate (Ribeiro et al., 2001). Data were expressed as the average transcription activation rate to the vehicle (DMSO:MeOH, 1:3) from at least four different experiments. Rosiglitazone 10-5 mol.L-1 was used as a positive control for assays using PPARγ, and bezafibrate at a concentration of 3x10-4 mol.L-1 was used as a positive control for assays performed with PPARα and PPARβ/δ. The results obtained were submitted to analysis of variance (one-way ANOVA) followed by the Newman-Keuls multiple comparison test.
2.11. Adipocyte differentiation assay
To assess whether the extracts produced by fungi BvFII and BvFVII were able to promote PPARγ-dependent 3T3-L1 preadipocyte differentiation and whether they interfered with differentiation promoted by PPARγ activation or with lipid accumulation within cells after differentiation, mouse embryonic fibroblasts (NIH 3T3-L1) were cultured in DMEM medium supplemented with 10% fetal bovine serum, 2 mmol.L-1 glutamine, 50 IU.mL-1 of penicillin and 50 IU.mL-1 of streptomycin and incubated at 36°C and 5% CO2. After the second day of confluent growth, cells were induced to differentiate by treatment with DMEM supplemented with 10% neonatal bovine serum, 1 μg.mL-1 insulin (Sigma, St Louis, USA), 0.5 mmol.L-1 3-Isobutyl-1-methylxanthine (Sigma, St Louis, USA) and 1 mmol.L-1 dexamethasone (Sigma, St Louis, USA) for two days. After this period, the cells were treated with DMEM supplemented with 10% neonatal bovine serum and 1 μg.mL-1 of insulin and incubated at 37°C and 5% CO2 for an additional 2 days. After that, the cells were maintained in cultivation with DMEM supplemented with 10% fetal bovine serum. After two days of incubation, the cells were stained with Oil Red O as described by Janderová et al. (2003), to observe the differentiation. The vehicle, 10-5 M rosiglitazone, fungus BvFII extract, or fungus BvFVII extract were added to the culture medium during the differentiation induction period to observe their interference in the cell differentiation process.
2.11.1. Extraction of total RNA from cells
After the treatment for the differentiation of adipocytes, the total RNA of the cells was extracted using the TRIzol reagent®, according to the protocol of the manufacturer Invitrogen™ (CA, USA).
2.11.2. Quantitative evaluation of the expression of the gene encoding FABP4 by RT-qPCR
RNA samples extracted from adipocyte cultures were evaluated for the quantitative determination of the expression of the gene encoding the adipocyte protein fatty acid binding protein 4 (FABP4). FABP4 is regulated by PPARγ and is essential in adipocyte fatty acid uptake, transport, and metabolism. For the assay, reverse transcription and quantitative PCR amplification (RT-qPCR) was performed in a single step using the Power SYBR® Green RNA to CT 1-Step kit (Applied Biosystems, Foster City, CA, USA), following the manufacturer’s instructions. The amplification product of each sample was determined by the SYBR® Green system. Reactions were prepared in 96-well plates (MicroAmp Optical, Applied Biosystems) to a final volume of 10 μL, containing 0.08 μL of RT Enzyme Mix (125x), 5 μL of SYBR® Green RT-PCR Power Mix (2x), 0.2 μL (100 nmol.L-1) of each primer (aP2 - Forward: CCATCTAGGGTTATGCTCTTCA and Reverse: ACACCGAGATTTCCTTCAAACTG; and 18S - Forward: CGGACAGGATTGAGAGATTG and Reverse: CAAATCGCTGCACCAACTAA) to obtain an initial concentration of 5 μmol,L-1, and 1 to 5 ng of RNA from the samples. The reaction conditions were the same for all RT-qPCR experiments: 30 minutes at 48°C to carry out the reverse transcription reaction; 10 minutes at 95°C for activation of the DNA polymerase enzyme; 40 cycles of 15 seconds at 95°C for denaturation; 1 minute at 60°C for annealing and extension on StepOnePlusTMReal-Time PCR Systems (Applied Biosystems) and the data obtained were analyzed with the software StepOne v2.1 (Bustin et al., 2009). The relative quantification of the expression of the gene encoding aP2 was performed using the equation 2-ΔΔCt, where ΔΔCt represents the difference between ΔCtsample and ΔCtcontrol, and ΔCt of the sample or control expresses the difference between Cq of the gene that encodes aP2 (FABP4) and the reference gene (18S) obtained for the samples. The statistical test applied was the analysis of variance (one-way ANOVA) followed by the Bonferroni test.
2.11.3. Western blotting assay
Differentiated 3T3-L1 cells were harvested from the culture plates, resuspended in 0.01 M phosphate buffer, pH 7.4, containing 0.9% NaCl (PBS), and centrifuged at 760.1 xg for 5 min at 4°C. The pellets were homogenized in lysis buffer [50 mmol.L-1 Tris buffer pH 7.5 with 150 M NaCl, 1% Triton X-100 and a diluted protease inhibitor cocktail (Sigma Aldrich, St. Louis, MO, USA). 1:200] and slowly stirred for one hour at 4°C. The total protein concentration was determined by the Bradford method (Bio-Rad Protein Assay®). To evaluate aP2 (FABP4) protein expression, 12.5 µg aliquots of protein were separated by SDS-PAGE, transferred to a PVDF membrane in a semi-dry vat (Bio-Rad, CA, USA), incubated with antibodies, and then immunodetected by chemiluminescence (ECL prime kit, GE Healthcare, Buckinghamshire, UK). The membrane was incubated overnight at 4°C with the anti-aP2 polyclonal antibody (195657, Abcam, Cambridge, USA) and the polyclonal antibody anti-GAPDH (9483, Abcam, Cambridge, USA) diluted in water containing 7% powdered milk. Immunostains were visualized using a secondary antibody conjugated to anti-goat peroxidase IgG-horseradish peroxidase conjugate (HRP) (sc-2354, Santa Cruz, CA, USA), diluted 1:7000 in water containing 7% powdered milk. The default marker used was the BenchMark™ pre-stained protein Ladder (Life Technologies, Grand Island, USA) according to the methodology described by Burnette (1981). The quantification of bands was performed with the ImageJ program version 1.46r (National Institute of Health, Bethesda, MD, USA) and expressed as arbitrary units (UA/GAPDH).
2.12. PPARγ nuclear receptor protection assay against protease digestion
The ligand-binding domain (LBD) of human PPARγ was expressed in bacteria (Escherichia coli, strain BL21) as a polyhistidine-tagged recombinant protein (His-LBD-PPARγ) and purified by cobalt affinity chromatography. The recombinant protein was incubated with rosiglitazone, with the extract produced by fungus BvFII, with the extract produced by fungus BvFVII, or with vehicle dimethyl sulfoxide (DMSO) in PBS buffer (0.01 mol.L-1, pH 7.4) for 30 minutes at room temperature. Then, it was incubated with distilled water or trypsin solution (100, 250, or 350 μg.mL-1) for 60 minutes, also at room temperature. The digestion reaction was stopped by adding denaturing sample buffer (100 mmol.L-1 Tris buffer, pH 6.8, containing 2% β-mercaptoethanol, 4% SDS, 4 mmol.L-1 EDTA, 20% glycerol, and bromophenol blue) and heating at 95°C for 10 minutes. The LBD proteolytic pattern of the protein was then analyzed by electrophoresis on a 12.5% polyacrylamide gel under denaturing conditions (Laemmli, 1970), followed by staining of the gel with the protein-dye Coomassie blue (Allan et al., 1992).
2.13. Evaluation of the inhibition of trypsin proteolytic activity by the extracts of fungi BvFII and BvFVII
To evaluate the ability to inhibit the proteolytic activity of trypsin on gelatin by extracts of fungi BvFII and BvFVII, a polyacrylamide gel (12.5%) containing 0.25% gelatin was prepared. Then, 100 µL of reaction medium with 10 µL of trypsin (200 µg.mL-1) in 80 µL of PBS buffer (0.01 mol.L-1, pH 7.4), and 10 µL of vehicle (DMSO), fungus BvFII extract (400 µg.mL-1), or fungus BvFVII extract (400 µg.mL-1) were incubated for 1 h at room temperature. After the incubation period, 20 µL of each sample were soaked in 6 mm diameter filter paper discs deposited on the gel and incubated for 60 minutes at room temperature. Proteolytic patterns were determined by the difference in intensity of the gel staining by Coomassie blue. After the decolorization of the gel, the intensity of the bands was measured using the Image J program, version 1.46r (National Institutes of Health, Bethesda, MD, USA) and expressed as a rate of inhibition regarding trypsin activity (Chiang et al., 2012).
2.14. Statistical analysis
All the experiments were performed in at least three different sets, and data were expressed as mean ± standard deviation (SD) of at least three measurements. IC50 values were calculated using Graphpad Prism® version 9.0.0. Data obtained in the experiments were analyzed through the one-way analysis of variance (ANOVA) test followed by Bonferroni’s multiple comparison tests. All statistical tests were performed using the software Graphpad Prism®.
3. Results
3.1. Submerged culture of the fungi and extract obtained
The mass of the extracts produced was determined, and the yield was calculated to the volume of fluid culture extracted and the mycelial mass produced. Fungus BvFI produced the highest amount of mycelia mass (wet weight 62.19 g) and the BvFII the lowest amount of mycelial mass (wet weight 6.32 g) after submerged liquid culture. However, the higher yield of metabolites extracts were acquired from the cultures of fungi BvFII and BVFIX that produced 35.2 mg and 89.5 mg of extract, respectively.
3.2. Molecular identification of fungi
3.2.1. Conserved regions of the ribosomal RNA gene (ITS - Internal transcribed spacer) from the fungal genomic DNA were amplified
Conserved regions of the ribosomal RNA gene (ITS - Internal transcribed spacer) from the fungal genomic DNA were amplified for molecular identification. The polymerase chain reaction amplification products were sequenced and compared with sequence data from species described in GenBank (National Institute of Health).
The alignment performed in GenBank using the advanced Blast tool showed identity above 98% in all analyzes performed. The e-value (expected value) is the probability of an alignment occurring by chance and is calculated based on the quality of the alignment performed. The lower the e-value, the higher the quality of the alignment performed.
As shown in the results, fungi BvFII, BvFVII and BvFIX belong to the same genus, Phomopsis sp., but the strains were kept separately because of their differences. Fungus BvFI showed a maximum identity of 94.20% with Colletotrichum sp. in the alignment performed. The fungus BvFV, with 100% identity, was characterized as Colletotrichum sp. Fungus BvFIII obtained greater identity with the fungi of the Phoma sp family, with a maximum identity of 99.64%. Fungus BvFVIII has a maximum identity of 100% with Pestalotiopsis mangiferae. The similarity was analyzed by grouping the neighbor-joining method with 1,000 bootstrapping repetitions to construct the phylogenetic tree (Figure 1).
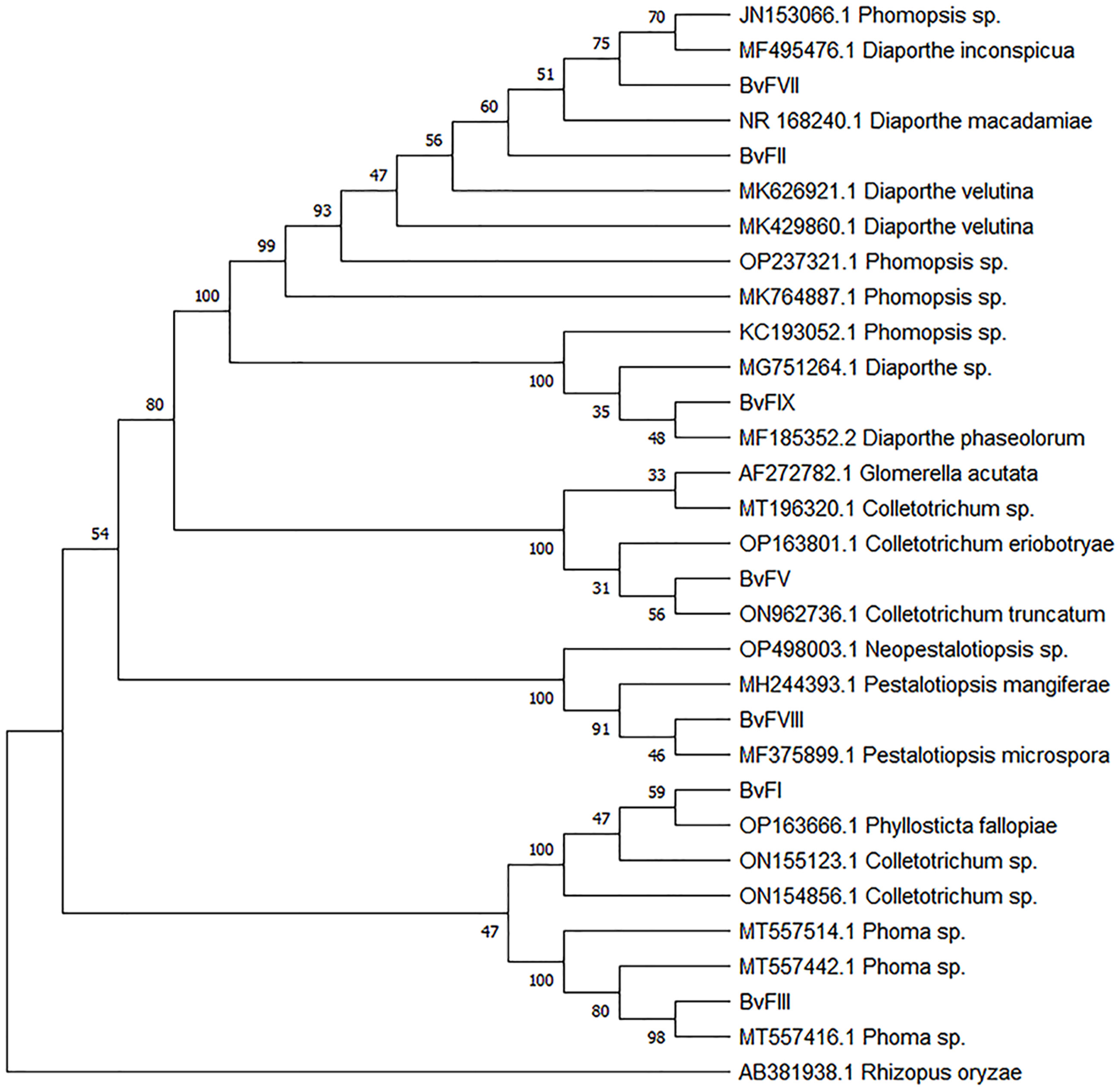
Figure 1 Phylogenetic tree constructed for PCR amplicons of the ITS1-5.8S-ITS2 rDNA of the endophytic fungi by genetic similarity comparison with data acquired with data from GenBank (NCBI). Phylogenetic tree constructed on the basis of the similarity of the PCR amplicons of the internal transcribed spacer 1 and 2 rDNA regions and sequences found in GenBank (NCBI), analyzed by the grouping neighbor-joining method, with 1,000 bootstrapping repetitions. The number of ramifications presents the bootstrap value (Felsenstein, 1985). The analysis involved 31 FASTA sequences. Rhizopus oryzae was used to root the phylogenetic tree. All positions containing gaps and missing data were eliminated. Evolutionary analyses were conducted in MEGA11 (Tamura et al., 2021).
3.3. Polyphenols and flavonoid contents
Obtained extracts were analyzed to determine their polyphenols and flavonoid contents using, respectively, Folin-Ciocalteu and aluminum chloride methods, as previously described. Calibration curves were constructed using gallic acid and quercetin as standards. Results are shown in Figure 2A.
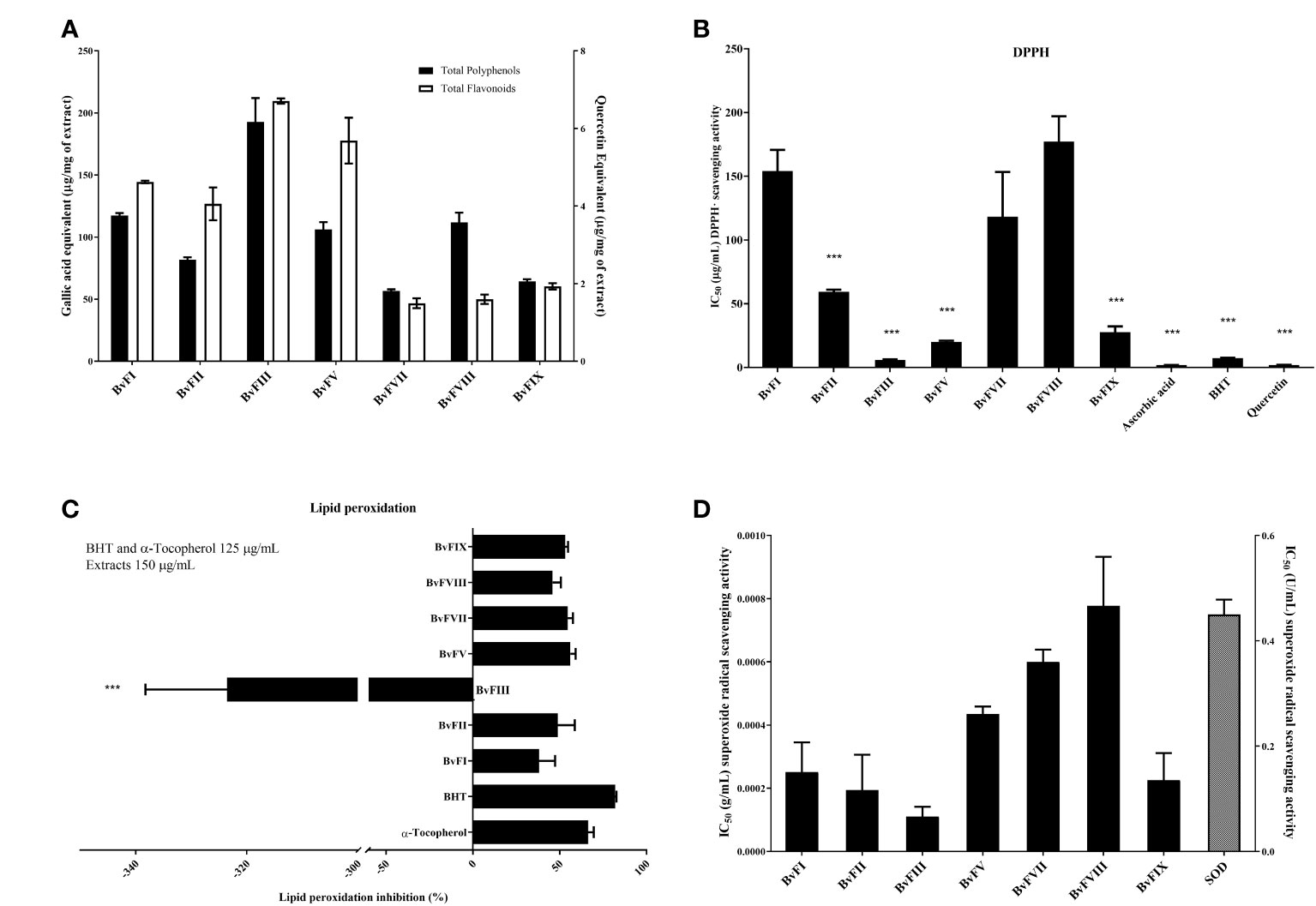
Figure 2 Polyphenols and flavonoids content and antioxidant effects. (A) Polyphenols and flavonoids contents in the extracts produced by the endophytic fungi isolated from Bauhinia variegata leaves. (B) Hydrogen-donating ability of the fungi extracts (BvFI, BvFII, BvFIII, BvFV, BvFVII, BvFVIII, and BvFIX) was evaluated by the DPPH· method. The concentration range of the extracts used in the reaction medium was 0.036 to 150.6 µg.mL-1. Standard compounds, ascorbic acid, BHT, and quercetin concentrations in the reaction medium also were used in the same concentrations. (C) Evaluation of the ability to inhibit lipid peroxidation by extracts of fungi. The emulsion of linoleic acid with polysorbate 20 in the presence and absence of extracts (150 µg.mL-1) and standards with the antioxidant activity known [tocopherol (125 µg.mL-1) and BHT (125 µg.mL-1)]. (D) Endophytic fungi extract ability to scavenge superoxide anion radicals. Extracts’ ability in scavenge superoxide anion radicals was evaluated using the xanthine/xanthine oxidase system as the source of the superoxide. produced reduces NBT to formazan detected by spectrophotometry at 560 nm. Antioxidants scavenge superoxide anion radicals, decreasing the reduction of NBT and, consequently, formazan production. Results were expressed as the percentage of inhibition of the lipid peroxidation process, as mean ± standard deviation of three different assays. The statistical test applied was the analysis of variance (one-way ANOVA) followed by the Newman-Keuls multiple comparison test, ***, significantly different, p<0.0001 vs. all compounds.
3.4. In vitro antioxidant activity determination
3.4.1. DPPH· assay
The antioxidant activity of the extracts was evaluated by DPPH· method and compared with compounds with recognized antioxidant activity, ascorbic acid, Butylhydroxytoluene (BHT), and quercetin. Results are reported in Figure 2B. Extracts of the fungi BvFIII, BvFV, BvFIX, and BvFII showed the higher DPPH· scavenging activity, the lower IC50 values in crescent order. The IC50 of the metabolites of fungus BvFIII was between those found to the standard antioxidants used as control, exhibiting a high antioxidant acitivity.
3.4.2. Lipid peroxidation inhibition assay
Linoleic acid is a polyunsaturated fatty acid necessary for producing arachidonic acid. It is present in the cell membrane’s phospholipid and is involved in signaling as a lipid second messenger. Both lipids are PPAR ligands (Forman et al., 1997). To evaluate the influence of endophytic fungi extracts on lipid peroxidation, an emulsion of linoleic acid (5 mmol.L-1) and polysorbate 20 (Tween®20) in saline phosphate buffer (0.1 mol.L-1 and pH 7.0) was incubated for 8 hours at 37°C in the presence or absence of extracts and positive controls tocopherol and BHT. In this way, linoleic acid undergoes auto-oxidation, and its product can be measured by spectrophotometry at the absorbance of 234 nm. The results obtained in this test are shown in Figure 2C. Excepting by extract of fungus BvFIII, all the extracts were able to inhibit lipid peroxidation. The weakest effect was observed for the extract of fungus BvFI, with 38% inhibition of lipid peroxidation. The highest effect was observed for the extract of fungus BvFV, which inhibits 56.1% of the oxidation process. BHT and α-tocopherol used as positive controls presented 89.9% and 66.6% of lipid peroxidation inhibition, respectively. Extracts of fungi BvFII, BvFVII, BvFVIII, and BvFXI presented percentages of lipid peroxidation of 48.8%, 54.6%, 45.9%, and 53.2%, respectively. Results also suggested that the fungus BvFIII extract improved lipid peroxidation by more than 2 and a half times (-262.4%).
3.4.3. Sequestration of the superoxide radical anion ()
Extracts’ ability in scavenge superoxide anion radical was evaluated using the xanthine/xanthine oxidase (Robak and Gryglewski, 1988) system as the source of the superoxide radical anion (O2-∙) produced reduced NBT to formazan detected by spectrophotometry at 560 nm. Antioxidants scavenge superoxide anion radicals, decreasing the reduction of NBT and, consequently, formazan production. Endophytic fungi extracts scavenged superoxide anion radicals (Figure 2D). Extract of fungus BvFVII (IC50 0.80 mg.mL-1), fungus BvFVIII (IC50 0.147 mg.mL-1) and BvFV (IC50 0.083mg.mL-1) showed lowest ability in scavenge superoxide. Extracts of fungus BvFI (IC50 0.208 mg.mL-1) and BvFIX (IC50 0.199 mg.mL-1) presented similar activity and extracts of fungus BvFII and BvFIII, with IC50 0.147 mg.mL-1 and 0.083 mg.mL-1, respectively, were the extracts showing highest ability in scavenge superoxide anion radical.
3.5. Adipocyte differentiation assay
As can be seen in Figure 3A, mouse embryonic fibroblasts (NIH 3T3L1) showed different patterns of oil red O staining after differentiation. Cells treated with rosiglitazone (10-5 M) showed the most accumulation of lipids. The fungus BvFII extract, at a concentration of 20 µg.mL-1, also stimulated the differentiation of these cells to a degree slightly higher than that presented by the vehicle and lower than that induced by rosiglitazone. At the concentration of 25 µg.mL-1 of extract, there was no significant change in the staining of cells to the control. The fungus BvFVII extract did not promote cell differentiation compared to the vehicle.
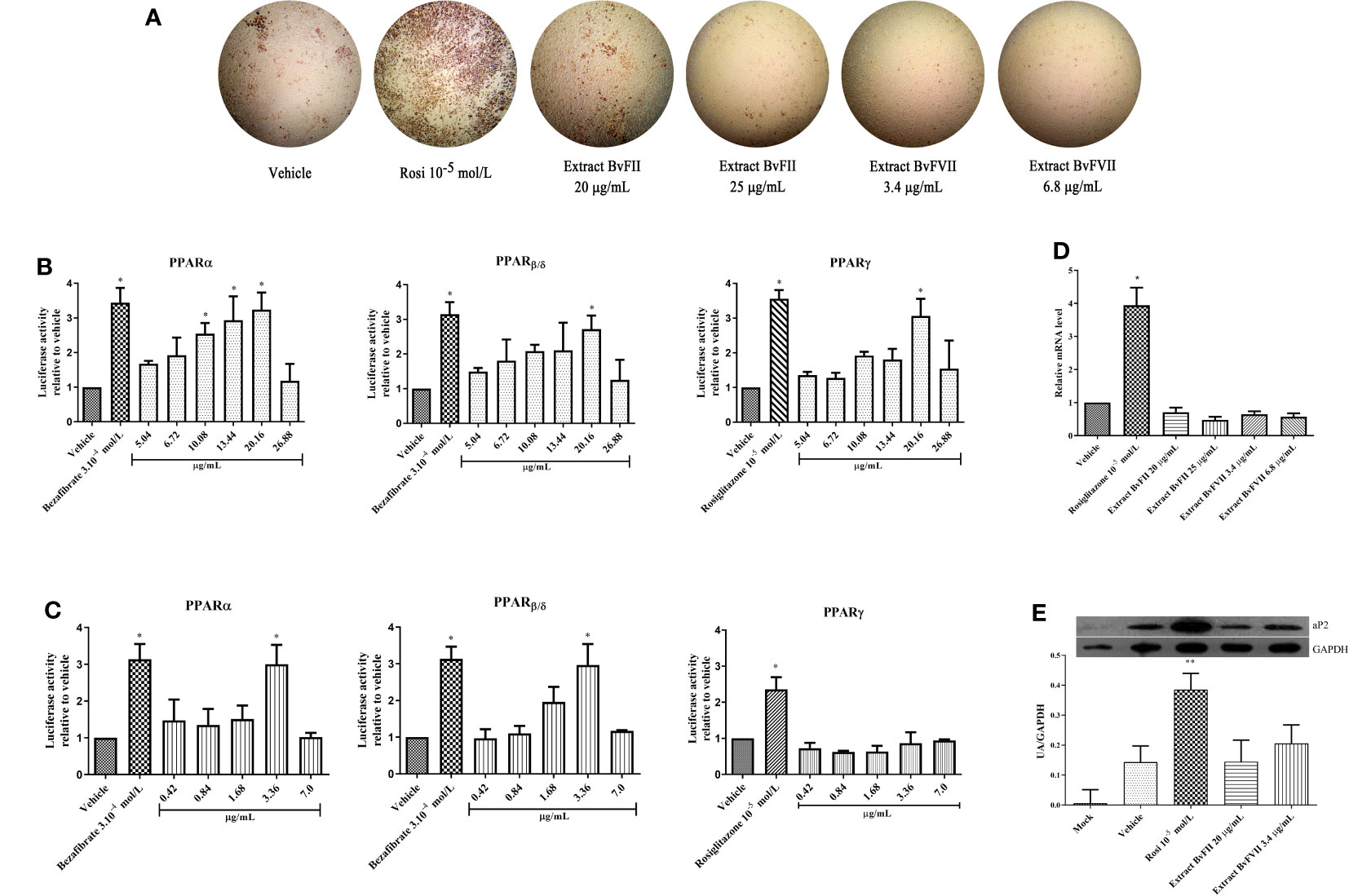
Figure 3 Extracts BvFII and BvFVII do not promote adipogenesis in 3T3-L1. Cells were induced to differentiate with insulin and exposed to vehicle (DMSO), Rosi 10-5 mol.L-1, Extract BvFII 20 or 25 µg.mL-1 and Extract BvFVII 3.4 or 7.8 µg.mL-1. (A) After 14 days, the cells were fixed, stained with oil red O, and photo documented. (B, C) Human mesangial cells were co-transfected with the expression vectors for PPARα or PPARγ, and the plasmid containing the luciferase reporter gene and treated with vehicle (DMSO:MeOH, 1:3), bezafibrate 3.10-4 mol.L-1for PPARα, β and rosiglitazone 10-5 M for PPARγ (positive controls) or increasing concentrations of extracts of fungi BvFII (B) and BvFVII (C). (D) Real-time quantitative PCR to evaluate the expression of Fabp4 (aP2) after 8 days of differentiation. (E) Western blot to evaluate the expression of FABP4 (aP2) protein after 14 days of differentiation. Data are presented as the mean (SD) of three independent experiments conducted in triplicate and expressed as activation relative to transcript levels in vehicle samples (DMSO). *p ≤ 0.05; **p ≤ 0.001.
3.6. Evaluation of agonist activity of fungi extracts in PPAR α, β and γ on reporter gene assay
The cultured extracts were submitted to the evaluation of their agonist capacity of the PPARα, PPARβ, and PPARγ using the gene reporter assay and treated using the concentration obtained on MTT assay. The results of the coactivation obtained are shown in Figures 3B, C. The extract of fungus BvFII was able to activate all PPAR isoforms, and this activation was similar to positive controls at a concentration of 20.16 µg.mL-1 (Figure 3B). The extract of fungus BvFVII also showed agonist activity, but only on PPARα and PPARβ receptors (Figure 3C). The other extracts didn’t show any activation (data not shown).
3.7. Evaluation of the relative gene expression of adipocyte protein 2 (aP2 or FABP4) by quantitative PCR
3T3-L1 cells were differentiated for 8 days with a complete medium and treated with vehicle (DMSO), rosiglitazone (10-5 mol.L-1), fungus BvFII extract (20 or 25 µg.mL-1), or fungus BvFVII extract (3.4 or 7.2 µg.mL-1) during differentiation. The difference in ap2 protein gene expression (FABP4) was not significant between the vehicle (DMSO) and the fungal extracts. There was only a significant difference between protein expression in cells treated with rosiglitazone and the other treatments (Figure 3D).
3.7.1. Western blotting
To evaluate the influence of extracts produced by fungi BvFII and BVFVII on the expression of aP2 in differentiated 3T3-L1 cells, the total protein of the cells was extracted, quantified, and analyzed by Western Blotting assay (Figure 3E). The concentrations of 20 µg.mL-1 for the extract of the fungus BvFII and 3.4 µg.mL-1 for the extract of the fungus BvFVII were chosen to evaluate the expression of the aP2 protein because they showed the highest activation over PPARγ in the reporter gene assay (Figures 3B, C).
3.8. Protease digestion protection assay
The binding of agonists to the LBD of nuclear receptors determines conformational changes critical for transcriptional activation by these receptors (Allan et al., 1992). These alterations are reflected in a modification of the accessibility of proteolytic sites on the surface of the receptor and, therefore, in an increase in the resistance of the receptor’s LBD to digestion by proteases. Thus, the protease digestion protection assay is one of the instruments used to study PPARγ conformational changes in the presence of ligands. From the results obtained, shown in Figure 4A, it is possible to observe that there was greater protection against digestion exerted by trypsin both for the samples treated with the extract of fungus BvFII and for those treated with the extract of the fungus BvFVII when compared to the treatment with rosiglitazone, even for the trypsin dose of 350 µg.mL-1.
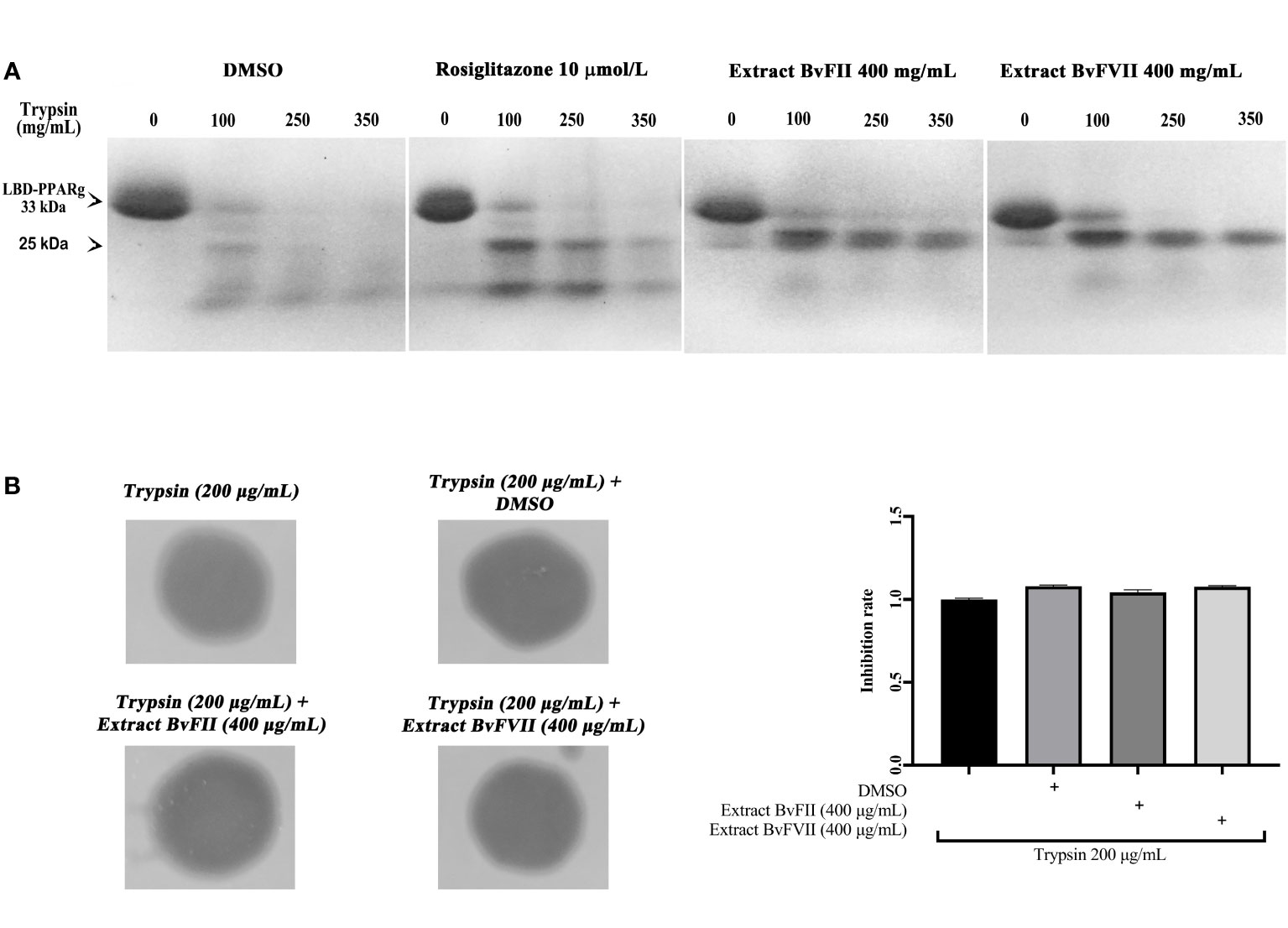
Figure 4 LBD proteolysis pattern of PPARγ by trypsin after treatment with different ligands. (A) Trypsin activity on PPARγ LBD was evaluated by using the recombinant protein his-LBD-hPPARγ, which was incubated with vehicle (DMSO), 10 µmol.L-1 rosiglitazone, fungus BvFII extract, or fungus BvFVII extract and with trypsin (0, 100, 250 and 350 μg.mL-1) for 60 minutes. LBD proteolytic patterns of PPARγ were analyzed by polyacrylamide gel electrophoresis under denaturing conditions, followed by staining the gel with Coomassie blue. (B) The proteolytic activity of trypsin on gelatin was evaluated after incubation of the enzyme with vehicle (DMSO) or with extracts of fungus BvFII (400 µg.mL-1) or fungus BvFVII (400 µg.mL-1). Polyacrylamide gel with 0.25% gelatin was left in contact with filter paper discs soaked in 20 µL of trypsin solution (200 µg.mL-1) for 1 h. Proteolytic patterns were analyzed by staining the gel with Coomassie blue. Dark areas show degraded gelatin. The quantification of the inhibition rate of vehicle and extracts BvFII and BvFVII is shown on the right. The data presented represent the mean ± standard deviation of the mean. The statistical test applied was the analysis of variance (one-way ANOVA) followed by the Newman-Keuls multiple comparison test, with no statistically significant difference.
3.9. Evaluation of the inhibition of trypsin proteolytic activity by extracts of fungi BvFII and BvFVII
Samples of the extract of fungus BvFII or fungus BvFVII were incubated with trypsin in PBS buffer to assess whether the extracts interfered with the proteolytic activity of the enzyme. Trypsin proteolytic activity was evaluated in a polyacrylamide gel containing gelatin (Figure 4B). As a negative control, trypsin was incubated with DMSO alone. The results suggest that extracts from fungus BvFII (400 µg.mL-1) or fungus BvFVII (400 µg.mL-1) do not interfere with the proteolytic activity of trypsin on gelatin.
4. Discussion
The potential of drugs from the microbiota of the Cerrado biome is little explored, and numerous studies have shown that fungi, especially endophytic fungi, emerge as a new perspective source of natural products with biological activity (Strobel et al., 2004; Zhang et al., 2006; Arnold, 2007; Rodriguez et al., 2009; Zhou et al., 2010; Blackwell, 2011; Mousa and Raizada, 2013).
Bauhinia variegata is a plant well adapted to the Cerrado and produces phenolic compounds, such as tannins and flavonoids (Duarte et al., 2007), and has anti-inflammatory (Mohamed et al., 2009), hepatoprotective activity (Bodakhe and Ram, 2007), antihyperglycemic (Azevedo et al., 2006; Kumar et al., 2012; Kulkarni and Garud, 2015) in addition to activating PPARγ (Silva, 2014).
After the publication of the work describing the ability of an endophytic fungus to produce the same active metabolite as its host plant (Stierle et al., 1993), the prospect was opened that natural products found in medicinal plants could be produced by endophytic fungi that colonize them (Newman and Cragg, 2015).
In this sense, by producing substances with hypoglycemic activity (Azevedo et al., 2006; Frankish et al., 2010; Kumar et al., 2012; Kulkarni and Garud, 2015) and substances that activate PPARγ (Costa, 2005; Silva, 2014), Bauhinia variegata was chosen for the isolation of endophytic fungi that colonize it, to evaluate the ability of these fungi to produce metabolites that could activate PPAR (α, β/δ and, γ) and that had antioxidant activity, to search for new bioactive compounds that could be used in the treatment of diabetes.
Three of the seven isolated endophytic fungi, BvFII, BvFVII, and BvFIX, were identified as fungi from the genus Phomopsis (Table 1). This genus corresponds to the asexual (anamorph) form of the ascomycete Diaporthe, which encompasses more than 900 species of fungi, and is characterized as a major producer of secondary metabolites, including antidiabetic, anti-inflammatory, and antioxidant compounds (Xu et al., 2021). The fungi BvFI and BvFV showed higher similarity with Colletotrichum sp. on the blast analysis. Tianpanich et al. (2011) isolated six substances with antioxidant activity from a Colletotrichum sp. that were characterized as five isocoumarin derivatives and a phthalide, of which four showed cytotoxic activity. In another work, Li et al. (2022) demonstrated that diterpene lactones obtained from Colletotrichum sp. showed strong antibacterial and also antioxidant activity. The fungus BvFI was related to the Guignardia (synonym: Phyllosticta) (Figure 1), an endophytic/phytopathogenic fungus. Some species of the genus Guignardia, such as Guignardia citricarpa and Guignardia psidii cause diseases such as citrus black spot and guava fruit rot. Other species like Guignardia mangiferae, P. capitalensis, and P. citribraziliensis are considered endophytic because they were extracted from plants without signs of infection. Guignardia mangiferae is an important producer of bioactive substances, such as terpenes (Yuan et al., 2010) and acetylcholinesterase inhibitors. The fungus BvFIII was characterized as Phoma sp. This genus is known as an important plant pathogen, and it is also found in the marine environment; it produces hundreds of secondary metabolites already described (Rai et al., 2018), among them substances with antifungal and cytotoxic action against tumor cell lines have already been described (Li et al., 2005; Wang et al., 2012). These previous works corroborate the results obtained in this work, which show the antioxidant activity of the metabolites of these fungi isolated from B. variegata (Figure 2).
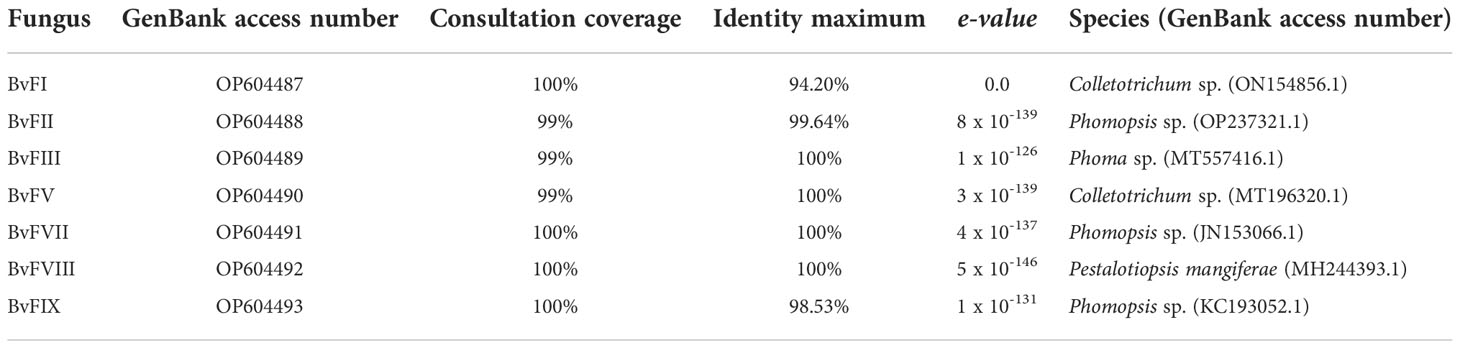
Table 1 Strains of endophytic fungi isolated from Bauhinia variegata.
Oral antihyperglycemic thiazolidinediones favor insulin sensitization mediated by PPARγ activation. They have long been used for the treatment of type 2 diabetes, but several serious adverse effects, such as loss of bone density, retention of body fluids, troglitazone-related liver problems, and heart problems related to rosiglitazone consumption (Singh et al., 2007). However, PPARγ ligands remain an effective option in the treatment of type 2 diabetes. They can improve insulin sensitivity in all tissues by producing increased glucose uptake in peripheral tissues. Partial agonists may have more positive effects as PPARγ ligands in diabetes by decreasing adverse effects caused by the activation of unwanted transcription, favored by the binding of full agonists, and by preventing Cdk5-mediated PPARγ phosphorylation, improving insulin sensitivity. ERK can also phosphorylate serine 273 of PPARγ and can be suppressed by Cdk5 via MEK activation (Banks et al., 2015), and, in this way, PPARγ has its function controlled. Thus, the treatment of type 2 diabetes with ERK/MEK axis inhibitors could be used to improve insulin resistance.
To assess whether the extracts showed agonist activity on the different PPAR isoforms, we performed a gene reporter assay, followed by treatment with the different extracts at maximum concentrations established by the results of the cytotoxicity assay performed previously. Among the tested extracts, two of them, extracts BvFII and BvFVII, showed PPAR agonist activity (Figures 2B, C). Extract BvFII at a concentration of 20.16 µg.mL-1 (Figure 2B) activated all three PPAR isoforms (α, β/δ and γ), showing to be a pan-agonist. In turn, extract BvFVII activated only PPARα and PPARβ, but showed low activation on chimeric PPARγ (LBD-PPARγ/DBD-GAL4).
Then, it was evaluated whether extracts from fungi BvFII and BvFVII were able to modify the structure of the LBD of PPARγ. For this, we performed the protease protection assay. The LBDs of PPARs have a large binding pocket, the largest concerning other nuclear receptors. When binding to the agonist, LBD undergoes crucial changes in its structure, reflecting the difficulty of accessing proteolytic sites on its surface and, therefore, increasing its resistance to digestion by proteases. In this case, when analyzed by SDS-PAGE, it shows a typical running pattern on the polyacrylamide gel, with evidence of a band of approximately 25 kDa that, in the presence of an agonist, it resists degradation by proteases. The result presented shows that, when in the presence of extracts of fungi BvFII and BvFVII, the LBD of PPARγ presents electrophoretic patterns like each other, suggesting that these extracts protected the receptor more efficiently than rosiglitazone (Figure 4A). This is clear since the band of approximately 25 kDa appears more protected. Berger et al. (2003) describe a partial agonist that showed a pattern of protection against trypsin activity similar to that shown in these results. An assay was carried out to evaluate the proteolytic activity of trypsin in the presence of the extracts (Figure 4B) to rule out the hypothesis that some substance present in the extracts was preventing the proteolytic function of trypsin since there are studies that describe the production of proteases by fungi (De Souza et al., 2015). The results showed that there was no difference between the degradation pattern of gelatin in the presence or absence of extracts and vehicle (DMSO). This suggests that the effect of the extracts in the protection against protease digestion assay was by direct binding to PPARγ.
So far, some studies have shown the agonist activity of metabolites from fungi on PPARγ (Sato et al., 2002; Lee et al., 2008; Lee et al., 2012; Hsu et al., 2013) and another in inhibiting its expression, but, to our knowledge, none with activity on the other α and β/δ receptor isoforms. However, works showing the agonist activity of natural products from plants on PPARγ (Wang et al., 2014) and the other PPAR isoforms (α and β/δ) are also activated by natural products. Martin et al. (2013) showed four isoflavones (polyphenolic compounds) with agonist activity on PPARα, PPARβ/δ, and PPARγ, characterizing these substances as pan-agonists. In another work, it was shown that the hexane extract of Tabebuia heptaphylla has pan-agonist activity on PPARs. It has also been shown that PPARα can be activated by a compound isolated from tomatoes, 9-oxo-10(E),12(E)-octadecadienoic acid (Kim et al., 2011), and another isolate of Cannabis sativa (Nadal et al., 2017) Green tea extract, in turn, was able to activate PPARβ/δ (Danesi et al., 2009) while coffee extract inhibited PPARγ (Aoyagi et al., 2014).
TZDs can promote the differentiation of preadipocytes, causing unfavorable effects of weight gain in patients who use them (Wilding, 2006). Activation of PPARα also promotes preadipocyte differentiation. To assess whether this effect could also be caused by extracts from fungi BvFII and BvFVII, 3T3-L1 preadipocytes were induced to differentiate in the presence or absence of extracts, rosiglitazone, or vehicle (DMSO) (Figure 3A). The extracts from fungi BvFII and BvFVII stimulated a slight accumulation of lipids in the cells, visibly lower than that promoted by rosiglitazone at a concentration of 10-5 M. This result was unexpected since full PPARγ agonists are known to be capable of inducing adipogenesis in 3T3-L1 cells (Rosen and Spiegelman, 2014), and PPARα agonists influence 3T3-L1 preadipocyte.
Therefore, it was interesting to evaluate the influence of these extracts on gene transcription of the FABP4 (aP2) protein. The FABP4 (aP2) protein is a lipid chaperone that is located in the cell cytoplasm and is regulated by PPARγ in adipocytes, where it is highly expressed and involved in lipid storage. In the real-time PCR assay (Figure 3D) and the Western Blotting (Figure 3C), it was observed that they did not influence the gene and protein expression of FABP4 (aP2), confirming the result observed in the adipocyte differentiation assay and lipid disclosure by red oil O staining (Figure 3A).
One hypothesis to explain the low induction of adipogenesis by the extracts, even with positive agonist activity and suggestion of binding to the receptor, would be that the extracts were somehow hampering the action of aP2. The aP2 gene is flanked by the PPARγ response element, and the non-functional aP2 decreases the adipogenic effect. However, to confirm this hypothesis, other experiments are necessary.
These findings may contribute to the discovery of agonist molecules that can be tested for use as antihyperglycemic agents in the treatment of type 2 diabetes and dyslipidemia. With the molecular identification and phylogenetic analysis of the fungi, it was possible to verify that the extract of the fungi BvFII and BvFVII, but not the BvFIX of the genus Phomopsis, are the ones with the highest degree of kinship (Figure 1). This result may explain the way the extract of the fungus BvFIX didn’t show activity on PPARs assays.
Oxidative stress in the cell results from the imbalance between the antioxidant defense system, which is reduced, and the production of reactive oxygen species, which is increased. This imbalance is involved in the etiology of several diseases mediated, among other pathways, by damage to DNA, lipids, and proteins, in addition to signaling for deregulated gene transcription.
Flavonoids have also been described as agonists of different PPARs (Salam et al., 2008; Quang et al., 2013; Xu et al., 2018). However, flavonoids also have antioxidant and pro-oxidant activity, depending on their concentration in the reaction medium and the pH of the medium. The pro-oxidant effect of flavonoids is, as a rule, directly proportional to the total number of hydroxyl groups and is responsible for the cytotoxic and pro-apoptotic effects of flavonoids isolated from various medicinal plants (Metodiewa et al., 1999). These characteristics of flavonoids agree with the results obtained (Figure 2), in which extracts with the lowest polyphenol content were the ones that showed low antioxidant activity when tested with the stable radical DPPH. In a review written by Hamilton et al. (2012), endophytic fungi are important in the symbiotic relationship that exists between them and the host plants, and hyphal growth occurs more easily in leaves that are in growth. Initially, when interacting with the plant, it is necessary for the fungus to produce reactive oxygen species (ROS), thus preventing the plant’s defenses from acting. It is not known for sure what defines the relationship of symbiosis or parasitism, but after infection, there is a balance in the production of ROS and antioxidant substances by both the plant and the fungus, and this relationship is, in some cases, species-specific (Tanaka et al., 2006).
To further advance the understanding of the antioxidant mechanism of action of the extracts, they were evaluated for their ability to prevent lipid peroxidation and to scavenge the superoxide radical anion. The process of lipid peroxidation of polyunsaturated fatty acids depends on the presence of oxygen. These fatty acids can activate peroxisome proliferator-activated receptors α, β/δ, and γ. Antioxidant agents can act by preventing the oxidation of these fatty acids, and thus, the action of the pro-oxidant agent, or by inactivating radical substances. High concentrations of glucose increase the concentration of reactive oxygen species (ROS) and also the activation of an enzyme called phospholipase A2, which is capable of inducing the phosphorylation of two serines (505 and 515) (Pavicevic et al., 2008) and the release of arachidonic acid and linoleic acid from the cell membrane phospholipid. After release, these fatty acids are oxidized (lipid peroxidation) by ROS producing, in addition to other products, 4-hydroxyl-2-nonenal (4-HNE), which binds to PPARβ/δ, increasing insulin secretion. When chronic hyperglycemia occurs, 4-HNE is produced in excess, and adducts with proteins, DNA, and phospholipids are formed, with consequent β-cell apoptosis in the pancreas.
In the test performed, it was evaluated whether the extracts were able to prevent the beginning of the lipid peroxidation process. As the reaction occurs in an oil-in-water emulsion, it is necessary to consider the polarity of the antioxidant agent since nonpolar substances may present higher activity because they are retained in the oil droplets, where the fatty acid is located, in the case of the test, linoleic acid. Both BHT (2,6-bis(1,1-dimethylethyl)-4-methylphenol) and tocopherol are nonpolar substances and were used as positive controls in the assay. Except for the fungus BvFIII extract, all the extracts tested were able to prevent linoleic acid peroxidation (Figure 2C). The fact that extracts from fungi BvFII and BvFVII show agonist activity in PPARβ/δ and, in addition, are antioxidants, may represent an interesting activity when we think that this could prevent β-cell apoptosis and, thus, prevent the development of diabetes. The presence of low polar antioxidant substances, which were in emulsion interphase, or nonpolar miscible in the oil phase, may have been responsible for the good activity in the inhibition of lipid peroxidation. As shown in Figure 2A, the extracts have flavonoids. This class of substances has a lipophilic character and may be responsible for the antioxidant action. Regarding the fungus BvFIII extract, a hypothesis that explains its pro-oxidant activity is that the concentration of the active substance is very high, and this was the cause of the pro-oxidative activity, which resulted in the oxidation of linoleic acid. It is also known that flavonoids can have an antioxidant or pro-oxidant character, depending on the pH of the medium or the presence of a transition metal in the reaction medium, such as copper (Cu2+) or iron, in addition to oxygen (Cao et al., 1997).
To evaluate the capacity of scavenging the superoxide radical anion, (), by the extracts, the xanthine/xanthine oxidase system was used as a producer of ROS. It is known that the superoxide radical anion is present in aerobic cells, plays an essential role in oxidation reactions, and interferes with cell signaling, together with other reactive oxygen species. Furthermore, it is involved in the pathogenesis of several diseases. As in the lipid peroxidation assay, the presence of flavonoids in the extracts may explain the ability to scavenge the radical, highlighting the fungus BvFIII extract (Figure 2D), which was able to do so at a low concentration of 0.083 mg.mL-1. To understand the relevance of this result, in work published by Dong et al. (2013), the best activity in bee honey extract was observed at a concentration of 28 mg.mL-1. With this, it is possible to perceive that all extracts obtained showed excellent activity when compared to the cited work.
More studies are still needed for a better understanding of the metabolic pathways and integrated effects of the sequestration of reactive oxygen species and activation of nuclear receptors, but the results obtained can be considered promising since the same extracts showed the ability to activate PPARγ without much interference in adipocyte differentiation, and were still able to inhibit the initiation step of lipid peroxidation and scavenge the superoxide radical anion. These biological activities are promising in the search for candidates for new antihyperglycemic agents, as they would allow the development of drugs with specific activities, with fewer incidences of adverse effects.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors upon request.
Author contributions
PM performed the experiments, analyzed the data, and wrote the manuscript. LA contributed with experiments of extracts obtention. FN coordinated the master project. MB conceived and coordinated this study. All authors contributed to the manuscript and approved the submitted version.
Funding
This work was supported by the Federal District Research Support Foundation (FAPDF - processes 193.000.376/2008 and 0193.000977/2015), the National Council for Scientific and Technological Development (CNPq), the Ministry of Science, Technology, Innovation and Communications (MCTIC), and the National Fund for Scientific and Technological Development (FNDCT) - MCT/CNPq/FNDCT/FAPs/MEC/CAPES/PRO CENTRO-OESTE (processes 564506/2010-9 and 407851/2013-5) and the Coordination for the Improvement of Higher Education Personnel (CAPES). This study was also financed in part by the Coordination for the Improvement of Higher Education Personnel – Brazil (CAPES, Portuguese: Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) – Finance Code 001.
Conflict of interest
The authors declare that the research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ada A. D. A. (2014). Diagnosis and classification of diabetes mellitus. Diabetes Care 37, S81–S90. doi: 10.2337/dc14-S081
Alberti K. G. M. M., Zimmet P. F. (1998). Definition, diagnosis and classification of diabetes mellitus and its complications. part 1: diagnosis and classification of diabetes mellitus. provisional report of a WHO consultation. Diabetic Med. 15, 539–553. doi: 10.1002/(SICI)1096-9136(199807)15:7<539:AID-DIA668>3.0.CO;2-S
Allan G. F., Leng X., Tsai S. Y., Weigel N. L., Edwards D. P., Tsai M. J., et al. (1992). Hormone and antihormone induce distinct conformational changes which are central to steroid receptor activation. J. Biol. Chem. 267, 19513–19520. doi: 10.1016/S0021-9258(18)41805-4
Aoyagi R., Funakoshi-Tago M., Fujiwara Y., Tamura H.. (2014). Coffee inhibits adipocyte differentiation via inactivation of PPARγ. Biol Pharm Bull 37 (11), 1820–5. doi: 10.1248/bpb.b14-00378
Arnold A. E. (2007). Understanding the diversity of foliar endophytic fungi: progress, challenges, and frontiers. Fungal Biol. Rev. 21, 51–66. doi: 10.1016/j.fbr.2007.05.003
Azevedo C., Maciel F., Silva L., Ferreira A., Da Cunha M., Machado O., et al. (2006). Isolation and intracellular localization of insulin-like proteins from leaves of bauhinia variegata. Braz. J. Med. Biol. Res. 39, 1435–1444. doi: 10.1590/S0100-879X2006001100007
Banks A. S., Mcallister F. E., Camporez J. P. G., Zushin P.-J. H., Jurczak M. J., Laznik-Bogoslavski D., et al. (2015). An ERK/Cdk5 axis controls the diabetogenic actions of PPAR[ggr]. Nature 517, 391–395. doi: 10.1038/nature13887
Berger J. P., Petro A. E., Macnaul K. L., Kelly L. J., Zhang B. B., Richards K., et al. (2003). Distinct properties and advantages of a novel peroxisome proliferator-activated protein γ selective modulator. Mol. Endocrinol. 17, 662–676. doi: 10.1210/me.2002-0217
Blackwell M. (2011). The fungi: 1, 2, 3 … 5.1 million species? Am. J. Bot. 98, 426–438. doi: 10.3732/ajb.1000298
Blois M. S. (1958). Antioxidant determinations by the use of a stable free radical. Nature 181, 1199–1200. doi: 10.1038/1811199a0
Bodakhe S. H., Ram A. (2007). Hepatoprotective properties of bauhinia variegata bark extract. Yakugaku Zasshi 127, 1503–1507. doi: 10.1248/yakushi.127.1503
Burnette W. N. (1981). “Western blotting”: Electrophoretic transfer of proteins from sodium dodecyl sulfate-polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein a. Analytical Biochem. 112, 195–203. doi: 10.1016/0003-2697(81)90281-5
Bustin S. A., Benes V., Garson J. A., Hellemans J., Huggett J., Kubista M., et al. (2009). The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 55, 611–622. doi: 10.1373/clinchem.2008.112797
Cao G., Sofic E., Prior R. L. (1997). Antioxidant and prooxidant behavior of flavonoids: Structure-activity relationships. Free Radical Biol. Med. 22, 749–760. doi: 10.1016/S0891-5849(96)00351-6
Chang C.-C., Yang M. H., Wen H. M., Chern J. C. (2002). Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 10, 178–182. doi: 10.38212/2224-6614.2748
Chiang H.-M., Chen H.-C., Lin T.-J., Shih I. C., Wen K.-C. (2012). Michelia alba extract attenuates UVB-induced expression of matrix metalloproteinases via MAP kinase pathway in human dermal fibroblasts. Food Chem. Toxicol. 50, 4260–4269. doi: 10.1016/j.fct.2012.08.018
Chiellini G., Apriletti J. W., Yoshihara H. A., Baxter J. D., Ribeiro R. C. J., Scanlan T. S. (1998). A high-affinity subtype-selective agonist ligand for the thyroid hormone receptor. Chem. Biol. 5, 299–306. doi: 10.1016/S1074-5521(98)90168-5
Costa M. (2005). Extrato de bauhinia variegata sobre a atividade transcricional mediada pelo receptor dos proliferadores peroxissomais-gama (PPARgama) (Universidade de Brasília: Mestrado).
Cotelle N. (2001). Role of flavonoids in oxidative stress. Curr. Top. Med. Chem. 1, 569–590. doi: 10.2174/1568026013394750
Da Cunha A. M., Menon S., Menon R., Couto A. G., Bürger C., Biavatti M. W. (2009). Hypoglycemic activity of dried extracts of bauhinia forficata link. Phytomedicine 17 (1), 37–41. doi: 10.1016/j.phymed.2009.06.007
Danesi F., Di Nunzio M., Boschetti E., Bordoni A. (2009). Green tea extract selectively activates peroxisome proliferator-activated receptor β/δ in cultured cardiomyocytes. Br. J. Nutr. 101, 1736–1739. doi: 10.1017/S0007114508145871
Das S., Rosazza J. P. N. (2006). Microbial and enzymatic transformations of flavonoids. J. Natural Prod. 69, 499–508. doi: 10.1021/np0504659
De Souza P. M., Bittencourt M. L., Caprara C. C., De Freitas M., De Almeida R. P., Silveira D., et al. (2015). A biotechnology perspective of fungal proteases. Braz. J. Microbiol. 46, 337–346. doi: 10.1590/S1517-838246220140359
Devchand P. R., Ziouzenkova O., Plutzky J. (2004). Oxidative stress and peroxisome proliferator-activated receptors: Reversing the curse? Circ. Res. 95, 1137–1139. doi: 10.1161/01.RES.0000151331.69399.b2
Dinis T. C., Maderia V. M., Almeida L. M. (1994). Action of phenolic derivatives (acetaminophen, salicylate, and 5-aminosalicylate) as inhibitors of membrane lipid peroxidation and as peroxyl radical scavengers. Arch. Biochem. Biophys. 315, 161–169. doi: 10.1006/abbi.1994.1485
Dong R., Zheng Y., Xu B. (2013). Phenolic profiles and antioxidant capacities of Chinese unifloral honeys from different botanical and geographical sources. Food Bioprocess Technol. 6, 762–770. doi: 10.1007/s11947-011-0726-0
Duarte M. R., Silva A. G., Costa R. E., Faria L. T. (2007). Bauhinia variegata: diagnose morfoanatômica e análise comparativa entre exemplares de regiões climáticas distintas. Latin Am. J. Pharm. 26, 837.
Evans J. L., Maddux B. A., Goldfine I. D. (2005). The molecular basis for oxidative stress-induced insulin resistance. Antioxid. Redox Signal 7, 1040–1052. doi: 10.1089/ars.2005.7.1040
Felber J. P., Golay A. (2002). Pathways from obesity to diabetes. Int. J. Obes. Relat. Metab. Disord. 26 Suppl 2, S39–S45. doi: 10.1038/sj.ijo.0802126
Felsenstein J. (1985). Confidence limits on phylogenies: An approach using the bootstrap. Evolution 39, 783–791. doi: 10.1111/j.1558-5646.1985.tb00420.x
Forman B. M., Chen J., Evans R. M. (1997). Hypolipidemic drugs, polyunsaturated fatty acids, and eicosanoids are ligands for peroxisome proliferator-activated receptors α and δ. Proc. Natl. Acad. Sci. 94, 4312–4317. doi: 10.1073/pnas.94.9.4312
Frankish N., De Sousa Menezes F., Mills C., Sheridan H. (2010). Enhancement of insulin release from the beta-cell line INS-1 by an ethanolic extract of bauhinia variegata and its major constituent roseoside. Planta Med. 76, 995–997. doi: 10.1055/s-0029-1240868
Gallo M. B. C., Pupo M. T., Bastos J. K., Nunes A. S., Cavalcanti B. C., Moraes M. O., et al. (2007). Atividade citotóxica de extratos de fungos endofíticos isolados de Smallanthus sonchifolius. Rev. Bras. Biociências 5, 402–404.
Guellich A., Damy T., Conti M., Claes V., Samuel J.-L., Pineau T., et al. (2013). Tempol prevents cardiac oxidative damage and left ventricular dysfunction in the PPAR-α KO mouse. Am. J. Physiol. 304, H1505–H1512. doi: 10.1152/ajpheart.00669.2012
Guimarães D. O., Borges W. S., Kawano C. Y., Ribeiro P. H., Goldman G. H., Nomizo A., et al. (2008). Biological activities from extracts of endophytic fungi isolated from viguiera arenaria and tithonia diversifolia. FEMS Immunol. Med. Microbiol. 52, 134–144. doi: 10.1111/j.1574-695X.2007.00354.x
Halliwell B. (2011). Free radicals and antioxidants - quo vadis? Trends Pharmacol. Sci. 32, 125–130. doi: 10.1016/j.tips.2010.12.002
Hamilton C. E., Gundel P. E., Helander M., Saikkonen K. (2012). Endophytic mediation of reactive oxygen species and antioxidant activity in plants: a review. Fungal Diversity 54, 1–10. doi: 10.1007/s13225-012-0158-9
Hsu W.-H., Lee B.-H., Chang Y.-Y., Hsu Y.-W., Pan T.-M. (2013). A novel natural Nrf2 activator with PPARγ-agonist (monascin) attenuates the toxicity of methylglyoxal and hyperglycemia. Toxicol. Appl. Pharmacol. 272, 842–851. doi: 10.1016/j.taap.2013.07.004
Hwang J., Kleinhenz D. J., Lassègue B., Griendling K. K., Dikalov S., Hart C. M. (2004). Peroxisome proliferator-activated receptor- ligands regulate endothelial membrane superoxide production. Am. J. Physiol. Cell Physiol. 288, C899. doi: 10.1152/ajpcell.00474.2004
Jackson M., Karwowski J., Humphrey P., Kohl W., Barlow G., Tanaka S. (1993). Calbistrins, novel antifungal agents produced by penicillium restrictum. J. Antibiotics 46, 34–38. doi: 10.7164/antibiotics.46.34
Janderová L., Mcneil M., Murrell A. N., Mynatt R. L., Smith S. R. (2003). Human mesenchymal stem cells as an in vitro model for human adipogenesis. Obes. Res. 11, 65–74. doi: 10.1038/oby.2003.11
Jiang B., Liang P., Zhang B., Song J., Huang X., Xiao X. (2009). Role of PPAR-beta in hydrogen peroxide-induced apoptosis in human umbilical vein endothelial cells. Atherosclerosis 204, 353–358. doi: 10.1016/j.atherosclerosis.2008.09.009
Kim Y.-I., Hirai S., Takahashi H., Goto T., Ohyane C., Tsugane T., et al. (2011). 9-oxo-10(E),12(E)-octadecadienoic acid derived from tomato is a potent PPAR α agonist to decrease triglyceride accumulation in mouse primary hepatocytes. Mol. Nutr. Food Res. 55, 585–593. doi: 10.1002/mnfr.201000264
King G. L., Loeken M. R. (2004). Hyperglycemia-induced oxidative stress in diabetic complications. Histochem. Cell Biol. 122, 333–338. doi: 10.1007/s00418-004-0678-9
Kulkarni Y. A., Garud M. S. (2015). Effect of bauhinia variegata Linn.(Caesalpiniaceae) extract in streptozotocin induced type I diabetic rats. Oriental Pharm. Exp. Med. 15 (3), 1–8. doi: 10.1007/s13596-015-0186-6
Kumar P., Baraiya S., Gaidhani S., Gupta M., Wanjari M. M. (2012). Antidiabetic activity of stem bark of bauhinia variegata in alloxan-induced hyperglycemic rats. J. Pharmacol. Pharmacotherapeut. 3, 64. doi: 10.4103/0976-500X.92518
Kumazawa S., Taniguchi M., Suzuki Y., Shimura M., Kwon M. S., Nakayama T. (2002). Antioxidant activity of polyphenols in carob pods. J. Agric. Food Chem. 50, 373–377. doi: 10.1021/jf010938r
Laemmli U. (1970). Most commonly used discontinuous buffer system for SDS electrophoresis. Nature 227, 680–685. doi: 10.1038/227680a0
Larran S., Perelló A., Simón M., Moreno V. (2007). The endophytic fungi from wheat (Triticum aestivum l.). World J. Microbiol. Biotechnol. 23, 565–572. doi: 10.1007/s11274-006-9266-6
Lee B.-H., Hsu W.-H., Chang Y.-Y., Kuo H.-F., Hsu Y.-W., Pan T.-M. (2012). Ankaflavin: a natural novel PPARγ agonist upregulates Nrf2 to attenuate methylglyoxal-induced diabetes in vivo. Free Radical Biol. Med. 53, 2008–2016. doi: 10.1016/j.freeradbiomed.2012.09.025
Lee I.-K., Lee J.-H., Yun B.-S. (2008). Polychlorinated compounds with PPAR-γ agonistic effect from the medicinal fungus phellinus ribis. Bioorg. Med. Chem. Lett. 18, 4566–4568. doi: 10.1016/j.bmcl.2008.07.034
Lingnert H., Vallentin K., Eriksson C. E. (1979). Measurement of antioxidative effect in model system. J. Food Process. Preservation 3, 87–103. doi: 10.1111/j.1745-4549.1979.tb00574.x
Li H., Qing C., Zhang Y., Zhao Z. (2005). Screening for endophytic fungi with antitumour and antifungal activities from Chinese medicinal plants. World J. Microbiol. Biotechnol. 21, 1515–1519. doi: 10.1007/s11274-005-7381-4
Liu Y.-J., Zhan J., Liu X.-L., Wang Y., Ji J., He Q.-Q. (2014). Dietary flavonoids intake and risk of type 2 diabetes: A meta-analysis of prospective cohort studies. Clin. Nutr 33 (1), 59–63. doi: 10.1016/j.clnu.2013.03.011
Li N., Xu D., Huang R. H., Zheng J. Y., Liu Y. Y., Hu B. S., et al. (2022). A new source of diterpene lactones from andrographis paniculata (Burm. f.) nees-two endophytic fungi of colletotrichum sp. with antibacterial and antioxidant activities. Front. Microbiol. 13, 819770. doi: 10.3389/fmicb.2022.819770
Lupi R., Del Guerra S., Mancarella R., Novelli M., Valgimigli L., Pedulli G. F., et al. (2007). Insulin secretion defects of human type 2 diabetic islets are corrected in vitro by a new reactive oxygen species scavenger. Diabetes Metab. 33, 340–345. doi: 10.1016/j.diabet.2007.03.005
Majka S. M., Barak Y., Klemm D. J. (2011). Concise review: adipocyte origins: weighing the possibilities. Stem Cells 29, 1034–1040. doi: 10.1002/stem.653
Manach C., Scalbert A., Morand C., Rémésy C., Jiménez L. (2004). Polyphenols: food sources and bioavailability. Am. J. Clin. Nutr. 79, 727–747. doi: 10.1093/ajcn/79.5.727
Marín-Peñalver J. J., Martín-Timón I., Sevillano-Collantes C., Del Cañizo-Gómez F. J. (2016). Update on the treatment of type 2 diabetes mellitus. World J. Diabetes 7, 354–395. doi: 10.4239/wjd.v7.i17.354
Martin H. (2010). Role of PPAR-gamma in inflammation. Prospects for therapeutic intervention by food components. Mutat Res 669 (1–7), 57–63. doi: 10.1016/j.mrfmmm.2009.09.009
Martin A., Doddareddy M. R., Gavande N., Nammi S., Groundwater P. W., Roubin R. H., et al. (2013). The discovery of novel isoflavone pan peroxisome proliferator-activated receptor agonists. Bioorg. Med. Chem. 21, 766–778. doi: 10.1016/j.bmc.2012.11.040
Metodiewa D., Jaiswal A. K., Cenas N., Dickancaite E., Segura-Aguilar J. (1999). Quercetin may act as a cytotoxic prooxidant after its metabolic activation to semiquinone and quinoidal product. Free Radic. Biol. Med. 26, 107–116. doi: 10.1016/S0891-5849(98)00167-1
Mohamed M. A., Mammoud M. R., Hayen H. (2009). Evaluation of antinociceptive and anti-inflammatory activities of a new triterpene saponin from bauhinia variegata leaves. Z. Für Naturforschung C. 64, 798–808. doi: 10.1515/znc-2009-11-1208
Möller E., Bahnweg G., Sandermann H., Geiger H. (1992). A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucleic Acids Res. 20, 6115. doi: 10.1093/nar/20.22.6115
Mosmann T. (1983). Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 65, 55–63. doi: 10.1016/0022-1759(83)90303-4
Mousa W. K., Raizada M. N. (2013). The diversity of anti-microbial secondary metabolites produced by fungal endophytes: an interdisciplinary perspective. Front. Microbiol. 4. doi: 10.3389/fmicb.2013.00065
Nadal X., Del Río C., Casano S., Palomares B., Ferreiro‐Vera C., Navarrete C., et al. (2017). Tetrahydrocannabinolic acid is a potent PPARγ agonist with neuroprotective activity. British Journal of Pharmacology 174 (23), 4263–4276. doi: 10.1111/bph.14019
Nazari M., Ghorbani A., Hekmat-Doost A., Jeddi-Tehrani M., Zand H. (2011). Inactivation of nuclear factor-[kappa]B by citrus flavanone hesperidin contributes to apoptosis and chemo-sensitizing effect in ramos cells. Eur. J. Pharmacol. 650, 526–533. doi: 10.1016/j.ejphar.2010.10.053
Newman D. J., Cragg G. M. (2015). Endophytic and epiphytic microbes as “Sources” of bioactive agents. Front. Chem. 3. doi: 10.3389/fchem.2015.00034
Oguntibeju O. O. (2019). Type 2 diabetes mellitus, oxidative stress and inflammation: examining the links. Int. J. Physiol. Pathophysiol. Pharmacol. 11, 45.
Pavicevic Z., Leslie C. C., Malik K. U. (2008). cPLA2 phosphorylation at serine-515 and serine-505 is required for arachidonic acid release in vascular smooth muscle cells. J. Lipid Res. 49, 724–737. doi: 10.1194/jlr.M700419-JLR200
Prabhakar P. K., Doble M. (2008). A target based therapeutic approach towards diabetes mellitus using medicinal plants. Curr. Diabetes Rev. 4, 291–308. doi: 10.2174/157339908786241124
Quang T. H., Ngan N. T., Minh C. V., Kiem P. V., Tai B. H., Nhiem N. X., et al. (2013). Anti-inflammatory and PPAR transactivational properties of flavonoids from the roots of sophora flavescens. Phytother. Res. 27, 1300–1307. doi: 10.1002/ptr.4871
Rai M., Gade A., Zimowska B., Ingle A. P., Ingle P. (2018). Marine-derived phoma-the gold mine of bioactive compounds. Appl. Microbiol. Biotechnol. 102, 9053–9066. doi: 10.1007/s00253-018-9329-2
Reiterer G., Toborek M., Hennig B. (2004). Quercetin protects against linoleic acid-induced porcine endothelial cell dysfunction. J. Nutr. 134, 771–775. doi: 10.1093/jn/134.4.771
Ribeiro R. C. J., Feng W., Wagner R. L., Costa C. U. H. R. M., Pereira A. C., Apriletti J. W., et al. (2001). Definition of the surface in the thyroid hormone receptor ligand binding domain for association as homodimers and heterodimers with retinoid X receptor. J. Biol. Chem. 276, 14987–14995. doi: 10.1074/jbc.M010195200
Robak J., Gryglewski R. J. (1988). Flavonoids are scavengers of superoxide anions. Biochem. Pharmacol. 37, 837–841. doi: 10.1016/0006-2952(88)90169-4
Rodriguez R. J., White J. F. Jr., Arnold A. E., Redman R. S. (2009). Fungal endophytes: diversity and functional roles. New Phytol. 182, 314–330. doi: 10.1111/j.1469-8137.2009.02773.x
Rosen E. D., Spiegelman B. M. (2014). What we talk about when we talk about fat. Cell 156, 20–44. doi: 10.1016/j.cell.2013.12.012
Saikkonen K., Faeth S., Helander M., Sullivan T. (1998). Fungal endophytes: a continuum of interactions with host plants. Annu. Rev. Ecol. Systematics 29, 319–343. doi: 10.1146/annurev.ecolsys.29.1.319
Salam N. K., Huang T. H., Kota B. P., Kim M. S., Li Y., Hibbs D. E. (2008). Novel PPAR-gamma agonists identified from a natural product library: a virtual screening, induced-fit docking and biological assay study. Chem. Biol. Drug Des. 71, 57–70. doi: 10.1111/j.1747-0285.2007.00606.x
Sambrook J., Fritsch E. F., Maniatis T. (1989). Molecular cloning (New York: Cold spring harbor laboratory press).
Sato M., Tai T., Nunoura Y., Yajima Y., Kawashima S., Tanaka K. (2002). Dehydrotrametenolic acid induces preadipocyte differentiation and sensitizes animal models of noninsulin-dependent diabetes mellitus to insulin. Biol. Pharm. Bull. 25, 81–86. doi: 10.1248/bpb.25.81
Silva C. A. D. M. (2014). Estudo químico biomonitorado de extratos das folhas de bauhinia variegata var. variegata DC. para a identificação de agonista do receptor ativado por proliferadores peroxissomais-gama (PPARg). Tese(doutorado) - Universidade de Brasília, Faculdadede Ciências da Saúde. doi: 10.26512/2014.12.T.18052
Singh S., Loke Y. K., Furberg C. D. (2007). Long-term risk of cardiovascular events with rosiglitazone. JAMA 298, 1189–1195. doi: 10.1001/jama.298.10.1189
Stierle A., Strobel G., Stierle D. (1993). Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of pacific yew. Science 260, 214–216. doi: 10.1126/science.8097061
Strobel G., Daisy B., Castillo U., Harper J. (2004). Natural products from endophytic microorganisms. J. Natural Prod. 67, 257–268. doi: 10.1021/np030397v
Tamura K., Stecher G., Kumar S. (2021). MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 38, 3022–3027. doi: 10.1093/molbev/msab120
Tanaka A., Christensen M. J., Takemoto D., Park P., Scott B. (2006). Reactive oxygen species play a role in regulating a fungus–perennial ryegrass mutualistic interaction. Plant Cell Online 18, 1052–1066. doi: 10.1105/tpc.105.039263
Teissier E., Nohara A., Chinetti G., Paumelle R., Cariou B., Fruchart J. C., et al. (2004). Peroxisome proliferator-activated receptor {alpha} induces NADPH oxidase activity in macrophages, leading to the generation of LDL with PPAR-{alpha} activation properties. Circ. Res. 95(12), 1174–82. doi: 10.1161/01.RES.0000150594.95988.45
Tianpanich K., Prachya S., Wiyakrutta S., Mahidol C., Ruchirawat S., Kittakoop P. (2011). Radical scavenging and antioxidant activities of isocoumarins and a phthalide from the endophytic fungus colletotrichum sp. J. Nat. Prod. 74, 79–81. doi: 10.1021/np1003752
Volpato G. T., Damasceno D. C., Rudge M. V., Padovani C. R., Calderon I. M. (2008). Effect of bauhinia forficata aqueous extract on the maternal-fetal outcome and oxidative stress biomarkers of streptozotocin-induced diabetic rats. J. Ethnopharmacol. 116, 131–137. doi: 10.1016/j.jep.2007.11.013
Wang L., Waltenberger B., Pferschy-Wenzig E.-M., Blunder M., Liu X., Malainer C., et al. (2014). Natural product agonists of peroxisome proliferator-activated receptor gamma (PPARγ): a review. Biochem. Pharmacol. 92, 73–89. doi: 10.1016/j.bcp.2014.07.018
Wang L.-W., Xu B.-G., Wang J.-Y., Su Z.-Z., Lin F.-C., Zhang C.-L., et al. (2012). Bioactive metabolites from phoma species, an endophytic fungus from the Chinese medicinal plant arisaema erubescens. Appl. Microbiol. Biotechnol. 93, 1231–1239. doi: 10.1007/s00253-011-3472-3
White T. J., Bruns T., Lee S., Taylor J. (1990). Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protocols: Guide Methods Appl. 18, 315–322. doi: 10.1016/B978-0-12-372180-8.50042-1
Wilding J. (2006). Thiazolidinediones, insulin resistance and obesity: Finding a balance. Int. J. Clin. Pract. 60, 1272–1280. doi: 10.1111/j.1742-1241.2006.01128.x
Xu H., Barnes G. T., Yang Q., Tan G., Yang D., Chou C. J., et al. (2003). Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Invest. 112, 1821–1830. doi: 10.1172/JCI200319451
Xu T.-C., Lu Y.-H., Wang J.-F., Song Z.-Q., Hou Y.-G., Liu S.-S., et al. (2021). Bioactive secondary metabolites of the genus diaporthe and anamorph phomopsis from terrestrial and marine habitats and endophytes: 2010–2019. Microorganisms 9, 217. doi: 10.3390/microorganisms9020217
Xu L. J., Yu M. H., Huang C. Y., Niu L. X., Wang Y. F., Wu C. Z., et al. (2018). Isoprenylated flavonoids from morus nigra and their PPAR gamma agonistic activities. Fitoterapia 127, 109–114. doi: 10.1016/j.fitote.2018.02.004
Yuan W. H., Liu M., Jiang N., Guo Z. K., Ma J., Zhang J., et al. (2010). Guignardones a–c: Three meroterpenes from guignardia mangiferae. Eur. J. Org. Chem. 2010, 6348–6353. doi: 10.1002/ejoc.201000916
Zakaria Z. A., Wen L. Y., Abdul Rahman N. I., Abdul Ayub A. H., Sulaiman M. R., Gopalan H. K. (2007). Antinociceptive, anti-inflammatory and antipyretic properties of the aqueous extract of bauhinia purpurea leaves in experimental animals. Med. Princ. Pract. 16, 443–449. doi: 10.1159/000107749
Zhang H. W., Song Y. C., Tan R. X. (2006). Biology and chemistry of endophytes. Nat. Prod. Rep. 23, 753–771. doi: 10.1039/b609472b
Zhou X., Zhu H., Liu L., Lin J., Tang K. (2010). A review: recent advances and future prospects of taxol-producing endophytic fungi. Appl. Microbiol. Biotechnol. 86, 1707–1717. doi: 10.1007/s00253-010-2546-y
Keywords: peroxisome proliferator-activated receptors, antioxidant, diabetes, endophytic fungi, Bauhinia variegata
Citation: Mesquita PG, Araujo LM, Neves FAR and Borin MdF (2022) Metabolites of endophytic fungi isolated from leaves of Bauhinia variegata exhibit antioxidant activity and agonist activity on peroxisome proliferator-activated receptors α, β/δ and γ. Front. Fungal Bio. 3:1049690. doi: 10.3389/ffunb.2022.1049690
Received: 20 September 2022; Accepted: 23 November 2022;
Published: 08 December 2022.
Edited by:
João Vicente Braga de Souza, National Institute of Amazonian Research (INPA), BrazilReviewed by:
Lijian Xu, Heilongjiang University, ChinaRamendra Pati Pandey, SRM University (Delhi-NCR), India
Copyright © 2022 Mesquita, Araujo, Neves and Borin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maria de Fátima Borin, bWZib3JpbkB1bmIuYnI=; bWZib3JpbkBnbWFpbC5jb20=