
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. For. Glob. Change , 02 April 2025
Sec. Forest Soils
Volume 8 - 2025 | https://doi.org/10.3389/ffgc.2025.1548485
This article is part of the Research Topic Nature-Based Solutions for Managing Soil Erosion and Enhancing Soil Stability View all 3 articles
 Laicong Luo1
Laicong Luo1 Kang Lin1Lingjian Tao1Chao Luo1Jianming Wang1Tianze Duan1Yubing Liu2Xiaobin Fu1Siqin Guo3Yuanqiu Liu1*
Kang Lin1Lingjian Tao1Chao Luo1Jianming Wang1Tianze Duan1Yubing Liu2Xiaobin Fu1Siqin Guo3Yuanqiu Liu1*Stand structure plays a crucial role in forest management, directly influencing the physicochemical properties of forest soils and, consequently, forest health and productivity. Cryptomeria japonica plantations are widely distributed in the mountainous regions of Japan and China and hold an important ecological status. This study aims to investigate the effects of different stand structures and soil depths on the physicochemical properties of soils in Cryptomeria japonica plantations in the Lushan region of China. The study was conducted in the Lushan National Nature Reserve. Stand structure was classified into three categories—good, medium, and poor—based on canopy closure (<0.5, 0.5–0.7, >0.7), understory vegetation cover (>0.8, 0.6–0.8, <0.5), and stand density (<650, 650–900, >900 trees per hectare). Soil samples were collected from plots representing different stand structures at four depth intervals (0–10 cm, 10–20 cm, 20–30 cm, and 30–40 cm) and analyzed for soil bulk density, porosity, water-holding capacity, as well as for the contents of carbon (C), nitrogen (N), phosphorus (P), and their stoichiometric ratios. The results indicated that both stand structure and soil depth significantly affected the physical properties and stoichiometric characteristics of the soil. Compared with poor stand structures, good stand structures significantly reduced soil bulk density, increased porosity, and enhanced water-holding capacity. Moreover, soils in stands with good structure exhibited higher organic carbon and total nitrogen contents, particularly in the surface soil layer (0–10 cm), while the C:N, C:P, and N:P ratios gradually decreased with increasing soil depth. The study demonstrated that maintaining a good stand structure—characterized by low canopy closure, high understory vegetation cover, and moderate stand density—can significantly improve soil porosity, water-holding capacity, and nutrient cycling efficiency. These findings provide a scientific basis for sustainable forest management, suggesting that optimizing stand structure can enhance soil health and overall ecosystem functionality.
In forest management, stand structure is a key measure of forest diversity, ecosystem functions, and the impacts of management practices. A well-structured stand (classified by low canopy closure <0.5, high understory vegetation cover >0.8, and moderate stand density < 650 trees/ha) not only enhances understory vegetation diversity but also improves water and nutrient retention, thus increasing timber production efficiency and facilitating the management of large-diameter, high-quality timber (Kermavnar et al., 2019). Different stand structures can influence the composition, abundance, and distribution of understory vegetation, as well as the physico-chemical properties of the soil, due to environmental changes in factors such as light availability, temperature, and humidity. However, well-structured stands are particularly effective in optimizing these environmental factors, thereby promoting more favorable conditions for soil health and understory vegetation diversity (Botequim et al., 2021). Moreover, these structural characteristics also affect nutrient cycling, including carbon (C), nitrogen (N), and phosphorus (P), all of which are essential for forest health and sustainability (Li et al., 2020; Xiao et al., 2022).
Soil water-holding capacity is a key indicator of its ability to retain and regulate water, which is essential for water retention and water-use efficiency in forest ecosystems (Huang and Shao, 2019). Various soil physicochemical properties, including bulk density, porosity, soil structure, and organic matter content, interact to affect water-holding capacity. For example, organic matter accumulation can improve soil structure by increasing porosity, enhancing soil aggregation, and subsequently boosting water retention capacity (Yang et al., 2014; Manns et al., 2016; Franko and Schulz, 2020). A well-structured forest stand can further improve soil water-holding capacity by increasing porosity and reducing soil density, enabling the soil to retain more water and continuously supply it to the ecosystem (Ilek et al., 2014; Dlapa et al., 2020). This is primarily due to increased porosity and reduced soil density, which enable the soil to retain more water and provide a continuous supply to the forest (Tague and Moritz, 2019; Sekucia et al., 2020).
Cryptomeria japonica, widely planted across Japan and China, plays a crucial role in forest restoration and sustainable land management due to its rapid growth, drought resistance, and adaptability. Its extensive use in plantation forestry is driven by its ability to conserve soil, retain water, and purify air (Farahnak et al., 2019, 2020), making it a key species for maintaining ecosystem stability and supporting forest-based ecosystem services (Xiao et al., 2022). However, the specific effects of different stand structures on the physicochemical properties of soil in these plantations remain underexplored (Duan et al., 2019).
Soil stoichiometry is the study of the balance and interaction of chemical elements, including carbon (C), nitrogen (N), and phosphorus (P), in the soil matrix (Yu et al., 2019). It examines the dynamic equilibrium and interactions of these elements, which significantly influence plant growth (Dibar et al., 2020). Enhancing forest stand structure greatly influences the accumulation and decomposition of organic matter in the soil, thereby affecting the content and proportions of carbon, nitrogen, and phosphorus (Shi et al., 2016; Rossi et al., 2025). This, in turn, promotes greater soil organic matter inputs through increased litter and root exudates, further enhancing the organic carbon pool in the soil (Manzoni et al., 2020). Additionally, forest stand structure indirectly influences soil microflora and fauna activities by modifying micro-environmental conditions, such as moisture and temperature, which in turn regulate the transformation and cycling of soil nitrogen and phosphorus (Joergensen and Scheu, 1998). For instance, areas with high stand density tend to have reduced light transmittance and disrupted temperature regimes, which can inhibit microbial decomposition and, subsequently, affect soil nutrient availability. Recent studies indicate that stand structure indirectly affects soil stoichiometric properties by influencing the soil’s physical structure and organic matter decomposition rate, thereby impacting nutrient cycling dynamics and forest ecosystem productivity (Zhang et al., 2013).
While previous studies explored the impact of forest stand structure on soil properties, the effects of different stand structures on soil characteristics in C. japonica plantations, particularly with respect to soil depth variations, remain under-researched. Soil depth is a crucial factor influencing nutrient distribution, microbial activity, and organic matter accumulation. However, research on the interaction between stand structure and soil depth and its effects on soil physicochemical properties and nutrient stoichiometry in C. japonica plantations remains limited (Xiao et al., 2022). Given the widespread distribution of C. japonica plantations in Mount Lushan and their crucial role in maintaining ecological balance and soil conservation, this study aims to thoroughly examine the specific impact of different stand structures and soil depths on soil physicochemical properties. This study focuses on C. japonica plantations with different stand structures (good, medium, poor) and analyzes soil bulk density, porosity, water-holding capacity, and the content and stoichiometric ratios of carbon (C), nitrogen (N), and phosphorus (P) at various soil depths (0–10 cm, 10–20 cm, 20–30 cm, and 30–40 cm). The main objective is to investigate how stand structure and soil depth collectively influence the physical properties and stoichiometric characteristics of the soil. Hypothesis: Forest stands with higher structural quality (good) will exhibit lower soil bulk density, higher porosity, and better water-holding capacity compared to those with medium or poor structures. We further hypothesize that these structural differences will significantly influence soil nutrient cycling, with good stands exhibiting higher organic carbon and nitrogen content and more favorable stoichiometric ratios (C:N, C:P, N:P) compared to poorer stands. Additionally, this study addresses two secondary objectives: (1) to explore how soil depth modulates the effects of stand structure on soil physicochemical properties, hypothesizing that surface layers (0–10 cm) will exhibit stronger responses to structural variations due to higher organic matter input and microbial activity; and (2) to investigate the interaction between stand structure and vertical nutrient distribution, positing that the decline in nutrient content and stoichiometric ratios with depth will be less pronounced in well-structured stands due to enhanced root penetration and organic matter stabilization. This study enhances the understanding of soil characteristics in C. japonica plantations and provides scientific evidence to optimize plantation management strategies by integrating structural and vertical soil dynamics.
This study was conducted in Lushan National Nature Reserve, located in Jiujiang City, Jiangxi Province, China (29°25′ ~ 29°41′N, 115°52′ ~ 116°06′E), covering an area of approximately 30.2 km2 (Figure 1). The region lies within the subtropical monsoon climate zone, with an average annual temperature of 11.6°C and precipitation of 2070 mm. Lushan experiences frost for about 150 days annually and is fog-covered for around 191 days. It is bordered by the Yangtze River to the north and Poyang Lake to the east. The region’s complex topography and unique climate have fostered diverse ecological niches and rich biodiversity. The Afforestation in this ecosystem are mainly composed of coniferous species, including Cryptomeria, Taiwan pine, and Chinese fir. The soils in Lushan include mountain brown soil, yellow soil, and red soil, though they are generally nutrient-poor and contain a high proportion of rocky material. With a peak elevation of 1,446 meters, Lushan’s geography and climate provide favorable conditions for diverse ecological habitats (Figure 1).
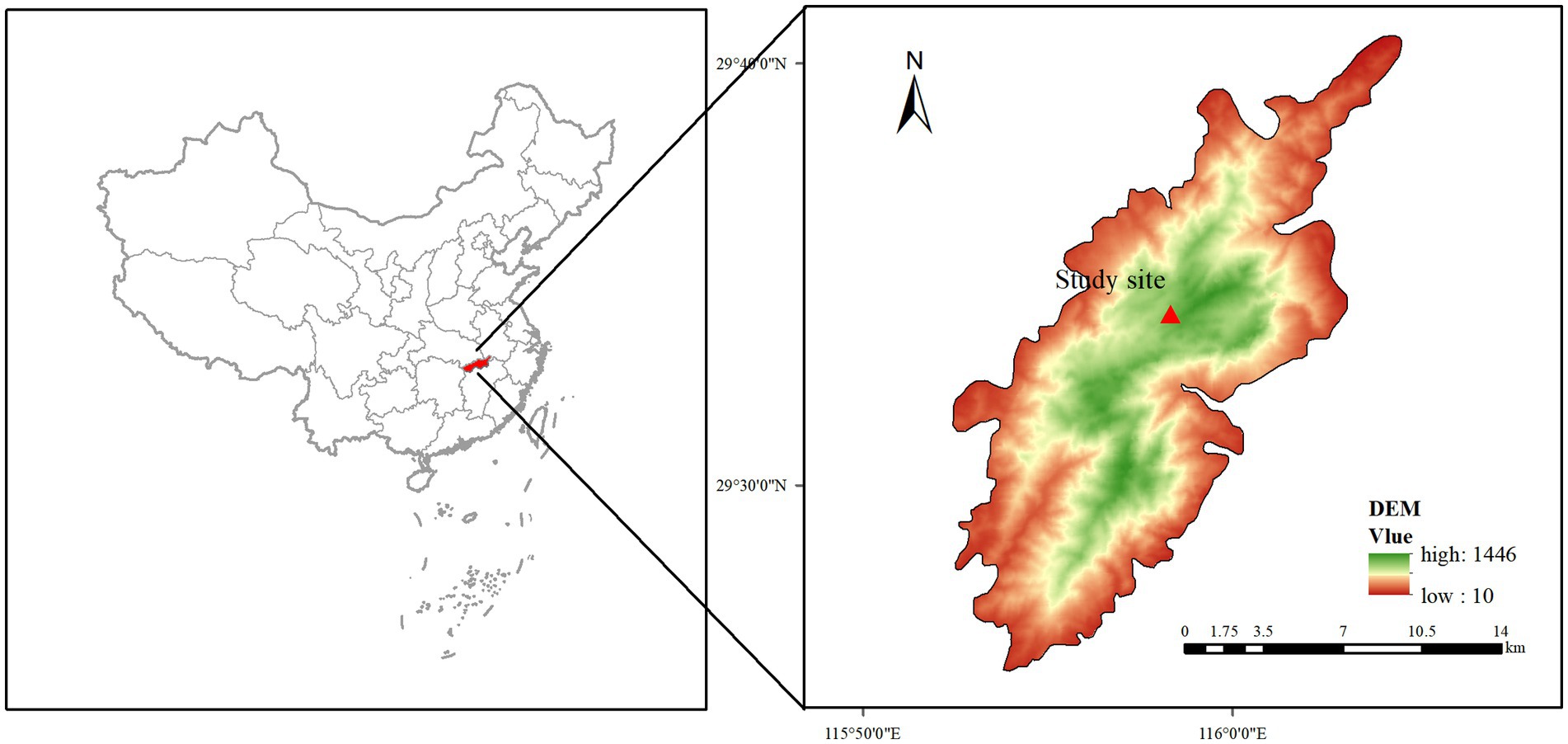
Figure 1. Location of the study site.
Based on field survey data collected in this study (Table 1), we classified C. japonica plantations into three stand structure categories: good, moderate, and poor. This classification was determined through a comprehensive survey that measured indicators such as tree height, diameter at breast height (DBH), crown width, tree density, canopy closure, understory vegetation cover, and litter biomass. Using these data, we categorized stand structure based on three key parameters: canopy closure, understory vegetation cover, and tree density. Stand structure was classified based on canopy closure, understory vegetation cover, and tree density. Specifically, the Good stand structure is defined as having low canopy closure (<0.5), high understory vegetation cover (>0.8), and low stand density (<650 trees per hectare). The Moderate stand structure is defined as having moderate canopy closure (0.5–0.7), moderate understory vegetation cover (0.6–0.8), and moderate stand density (650–900 trees per hectare). The Poor stand structure is characterized by high canopy closure (>0.7), low understory vegetation cover (<0.5), and high tree density (>900 trees per hectare). Sample plots were selected following a random sampling principle to ensure representativeness. Each plot was rectangular in shape, measuring 15 m × 15 m with a total area of 225 m2. Five sample plots were designated for each stand category, yielding a total of 15 plots. Within each plot, three sampling locations along the diagonal were selected, and soil samples were taken at four depth intervals: 0–10 cm, 10–20 cm, 20–30 cm, and 30–40 cm. Two methods were used to collect soil samples: undisturbed samples for physical property measurements were obtained using a soil auger, and surface soil was collected and dried for nutrient analysis. During analysis, stones, litter, and other non-soil components were removed to ensure accuracy. Soil moisture content was determined using the drying method. Soil bulk density, porosity, and water retention characteristics, including maximum water holding capacity, capillary water holding capacity, and field capacity, were determined using the soil auger method. Total nitrogen (TN) and total phosphorus (TP) content were determined using the H2SO4-HClO4 digestion method and analyzed with an automated chemical analyzer (Smart Chem 200, Rome, Italy). Soil organic carbon (SOC) content was measured using the potassium dichromate oxidation method with additional heating.
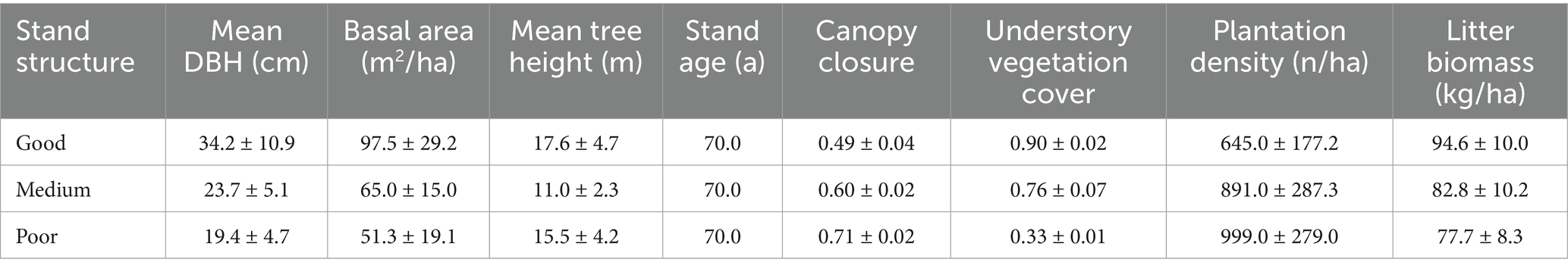
Table 1. Information on sample plots of C. japonica plantations with different stand structures.
The data were analyzed using IBM SPSS 25.0 software. One-way analysis of variance (ANOVA) and Tukey’s post hoc test (α = 0.05) were used to assess the effects of stand structure and soil depth on soil bulk density, porosity, water retention characteristics (maximum water holding capacity, capillary water holding capacity, and field capacity), nutrient content (carbon, nitrogen, and phosphorus), and stoichiometric characteristics. Pearson correlation analysis was performed to explore relationships among soil properties.
In the 10–20 cm soil layer, bulk density in areas with good stand structure was lower than in medium stand areas and in poor stand areas (by 7.85 and 25.89%) (Figure 2A). Total porosity, saturated water content, field capacity, and capillary water holding capacity in the 0–10 cm layer were 67.21%, 637.88 g·kg−1, 432.33 g·kg−1, and 528.98 g·kg−1, respectively, significantly higher than deeper layers (Figure 2). Soil physical properties, such as bulk density, porosity, saturated water content, field capacity, and capillary water holding capacity, were significantly affected by stand structure, soil depth, and their interaction (p < 0.05) (Table 2). Soils with good stand structure consistently had lower bulk densities at all depths than those with medium or poor stand structures (Figure 2A). However, an inconsistency was observed, as soils in areas with good stand structure had higher bulk densities than those in medium and poor stand areas (Figure 2A). A similar trend was observed for saturated water content, field capacity, and capillary water holding capacity (Figures 2C–E). Within each stand structure, bulk density increased with soil depth (Figure 2A).
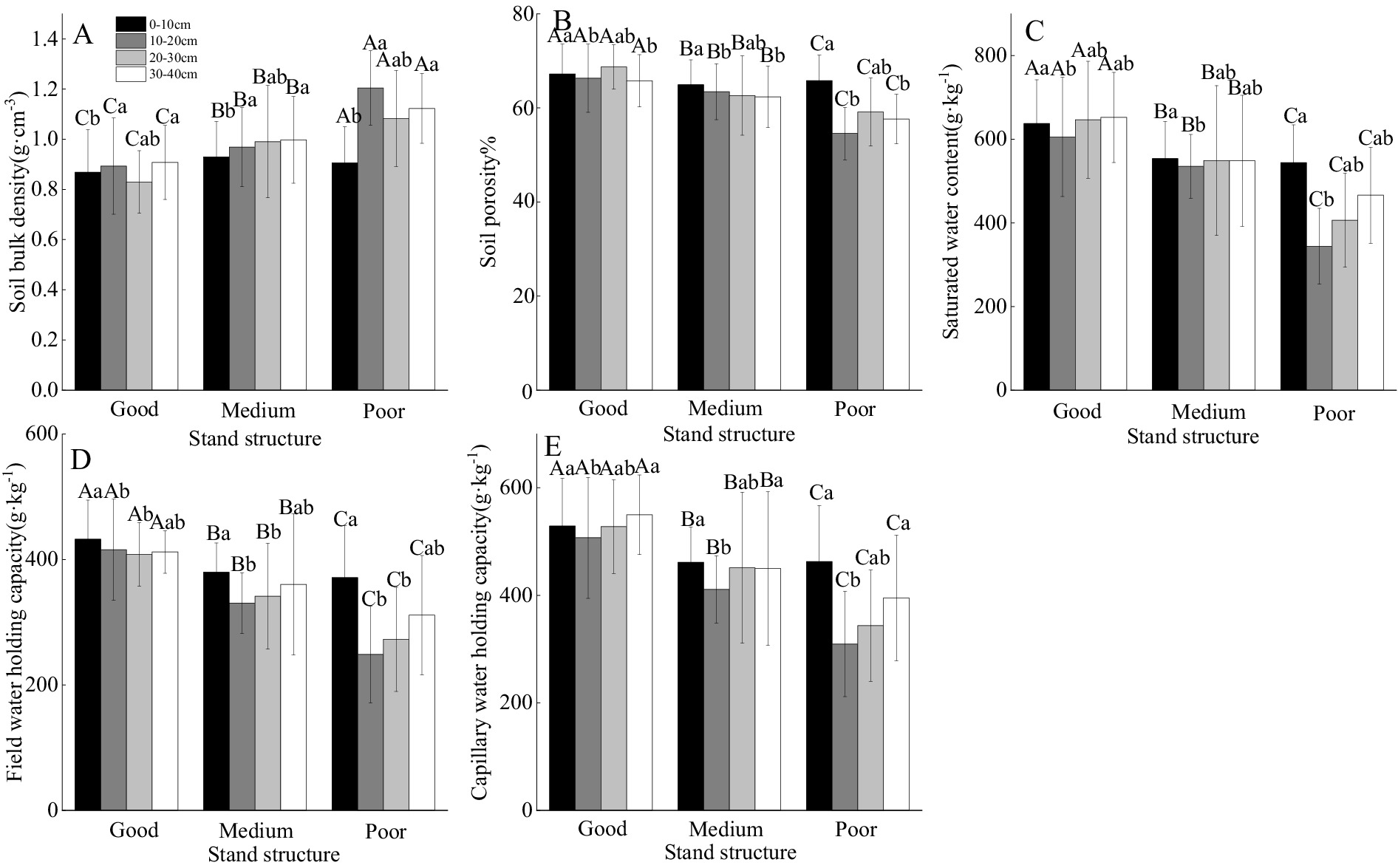
Figure 2. The impact of different stand structures and soil depths on soil physical [(A) Soil bulk density, (B) Soil porosity, (C) Saturated water content, (D) Field water holding and (E) Capillary water holding capacity]. Capital letters indicate comparisons among different forest stand structures, while lowercase letters indicate comparisons among different soil depths. Different letters indicate statistically significant differences between treatments (p < 0.05).
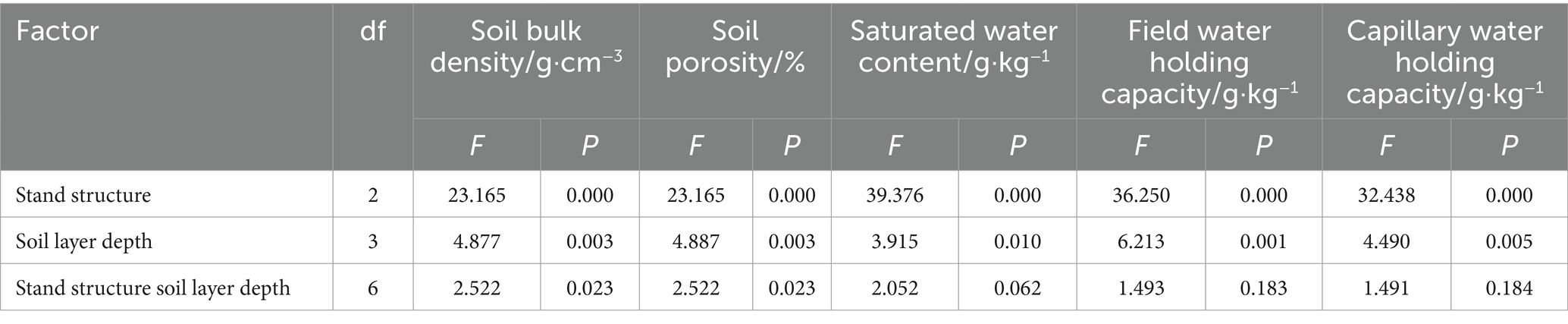
Table 2. Effects of different stand structures and soil depths on soil physical properties.
In the 0–10 cm layer, total phosphorus content in soils of areas with medium stand structure was significantly higher than in those with good and poor structures (by 33.56 and 34.66%) (Figure 3C). In areas with medium stand structure, total phosphorus content in the 0–10 cm layer was significantly higher than in the 20–30 cm layer (by 36.57%) (Figure 3C). Soil carbon (C), nitrogen (N), and phosphorus (P) contents were significantly influenced by stand structure, soil depth, and their interaction (p < 0.05) (Table 3). Within each stand structure, soil organic carbon and total nitrogen levels significantly decreased with increasing soil depth (Figures 3A,B). At the same soil depth, soils with good stand structure had significantly higher total nitrogen content than those with medium or poor structures (Figure 3B). Furthermore, medium stand areas had higher organic carbon and total nitrogen contents than poor stand areas (Figures 3A,B).
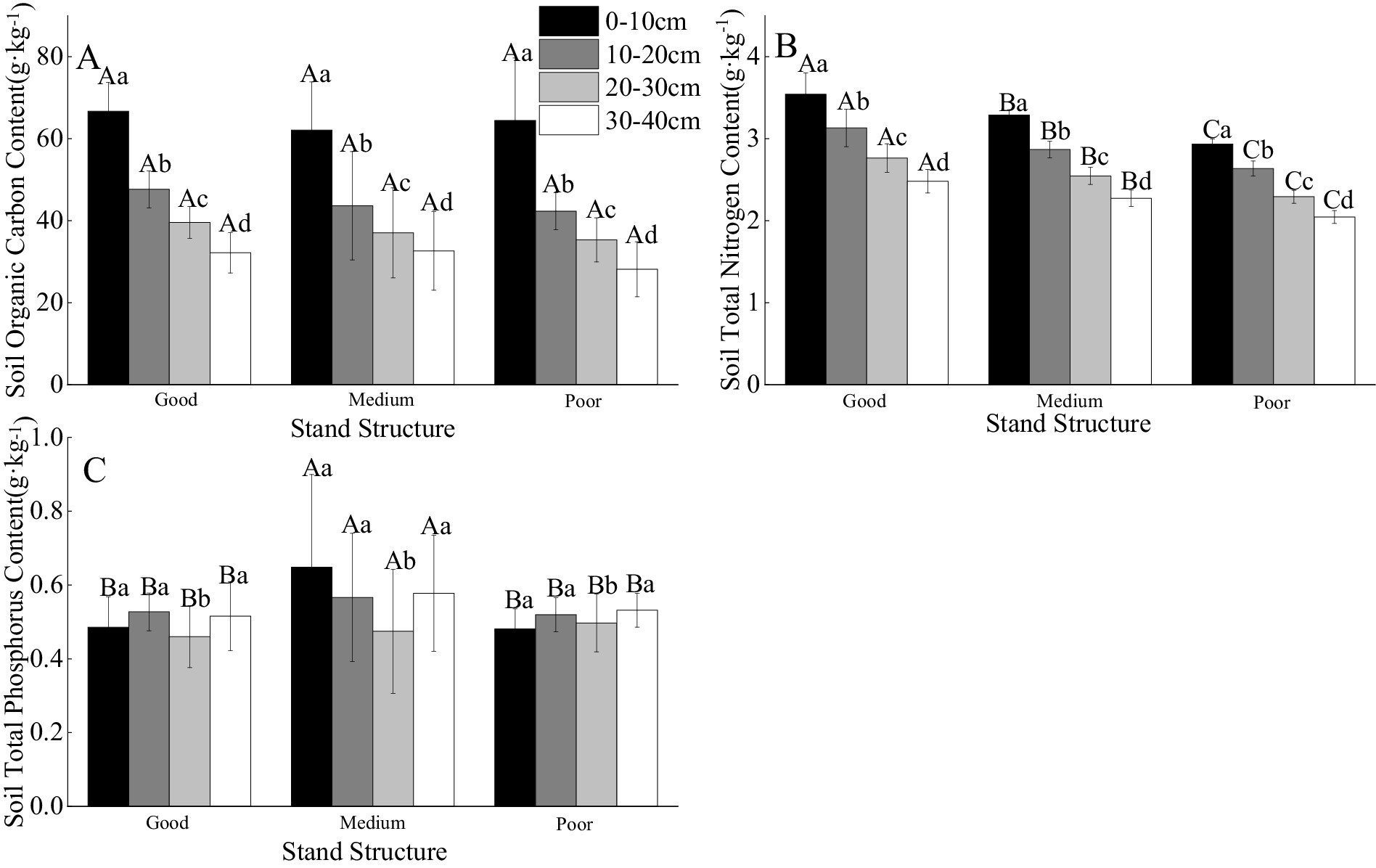
Figure 3. The impact of different stand structures and soil depths on the contents of soil C, N, and P [(A) Soil organic carbon, (B) Soil total nitrogen and (C) Soil total phosphorus]. Capital letters indicate comparisons among different forest stand structures, while lowercase letters indicate comparisons among different soil depths. Different letters indicate statistically significant differences between treatments (p < 0.05).
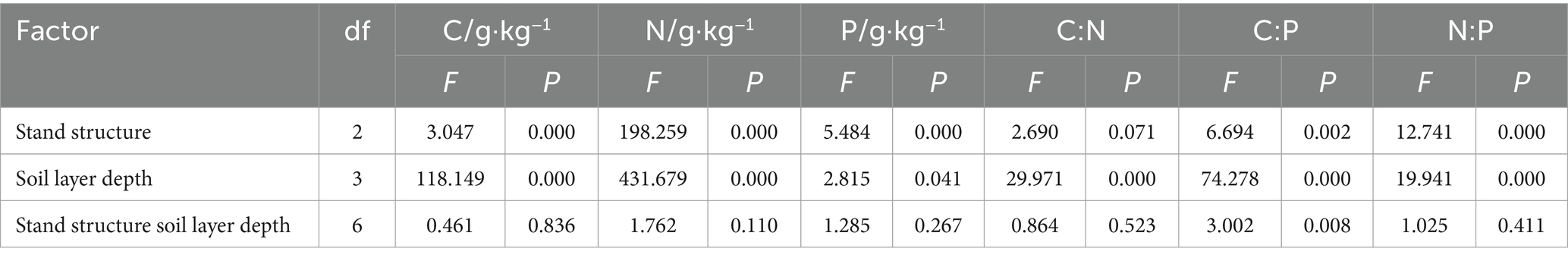
Table 3. The impact of different stand structures and soil depths on soil chemical properties.
In the 0–10 cm layer, soils in areas with medium stand structure had significantly lower C:P ratios than those in areas with good or poor structures (by −25.5% and − 20.8%, respectively) (Figure 4B). Stand structure, soil depth, and their interaction significantly influenced soil C:N, C:P, and N:P ratios (p < 0.05) (Table 3). Within each stand structure, soil C:N ratios decreased with increasing soil depth (Figure 4A). In the 0–10 cm layer, soils in areas with poor stand structure had higher C:N ratios than those with good or medium structures (Figure 4A). Within the same stand structure, soil C:P ratios decreased with increasing soil depth (Figure 4B). In areas with poor stand structure, soil N:P ratios decreased with increasing soil depth (Figure 4C). At all depths, soils in areas with good stand structure had higher N:P ratios than those in areas with medium or poor structures (Figure 4C).
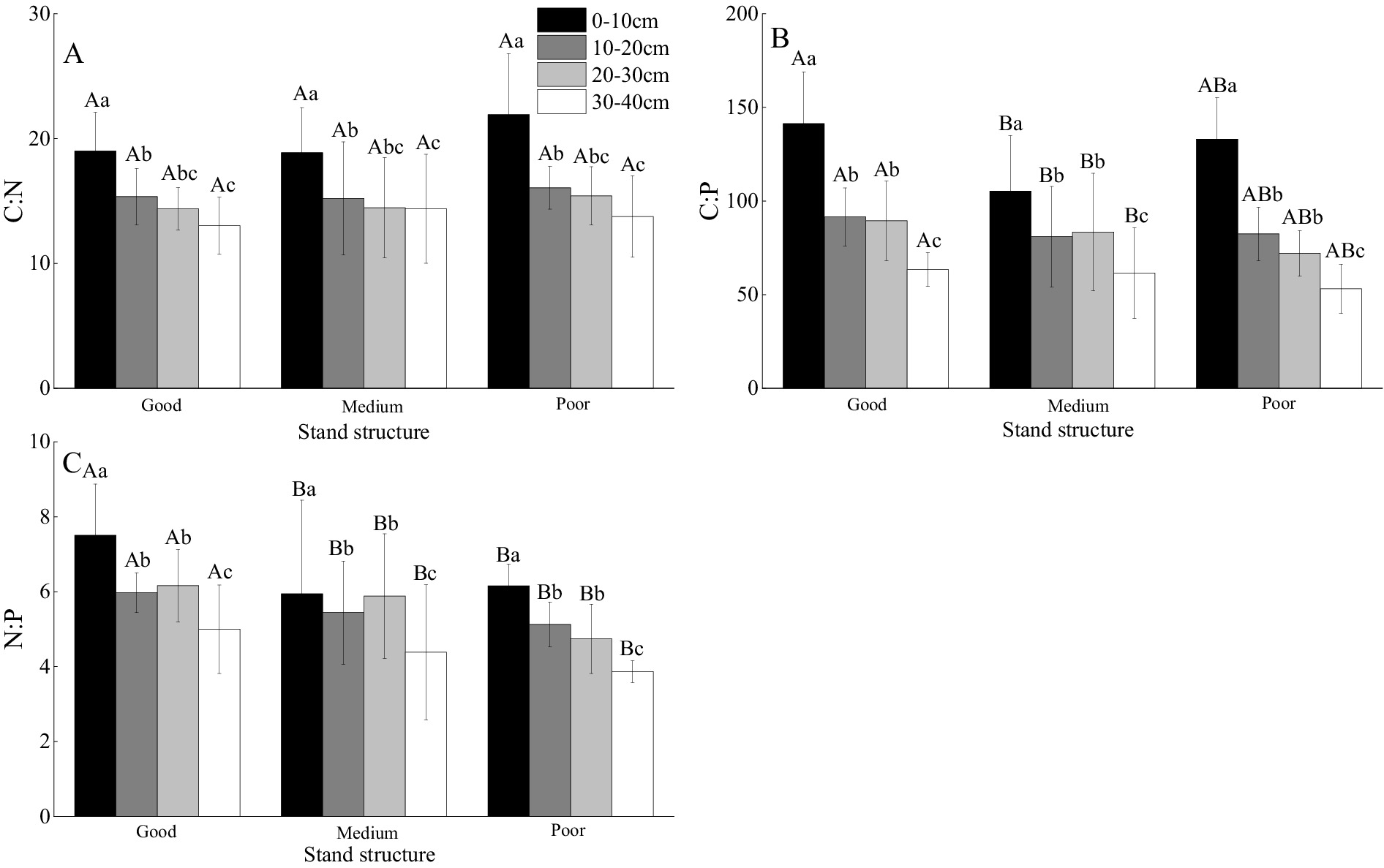
Figure 4. The impact of different stand structures and soil depths on soil C:N, C:P, and N:P ratios. Capital letters indicate comparisons among different forest stand structures, while lowercase letters indicate comparisons among different soil depths. Different letters indicate statistically significant differences between treatments (p < 0.05).
The heatmap of correlations among various soil properties shows the correlation coefficients between soil physical and chemical characteristics, including bulk density, total porosity, saturated water content, capillary water holding capacity, field capacity, soil SOC, TP, TN, C:N, C:P, and N:P. Soil bulk density exhibited significant or highly significant negative correlations with total porosity (r = −1.00, p < 0.01), saturated water content (r = −0.88, p < 0.01), capillary water holding capacity (r = −0.78, p < 0.01), field capacity (r = −0.78, p < 0.01), soil SOC (r = −0.19, p < 0.01), TP (r = −0.14, p < 0.05), and TN (r = −0.15, p < 0.01). Total porosity exhibited significant or highly significant positive correlations with saturated water content (r = +0.88, p < 0.01), capillary water holding capacity (r = +0.78, p < 0.01), field capacity (r = +0.78, p < 0.01), SOC (r = +0.19, p < 0.01), TP (r = +0.14, p < 0.05), and TN (r = +0.15, p < 0.01). Saturated water content exhibited a highly significant positive correlation with capillary water holding capacity (r = +0.90, p < 0.01), field capacity (r = +0.88, p < 0.01) and TN (r = +0.16, p < 0.05). Capillary water holding capacity exhibited significant or highly significant positive correlation with field capacity (r = +0.97, p < 0.01) and TN (r = +0.16, p < 0.05). Field capacity exhibited a highly significant positive correlation with TN (r = +0.25, p < 0.01). Soil SOC exhibited significant or highly significant positive correlations with TP (r = +0.30, p < 0.01), TN (r = +0.67, p < 0.05), C:N (r = +0.79, p < 0.05), C:P (r = +0.73, p < 0.01), and N:P (r = +0.29, p < 0.01). Soil TP exhibited significant or highly significant positive correlations with C:N (r = +0.14, p < 0.05), but highly significant negative correlations with C:P (r = −0.39, p < 0.01) and N:P (r = −0.61, p < 0.01). Soil TN exhibited highly significant positive correlations with C:N (r = +0.10, p < 0.01), C:P (r = +0.44, p < 0.01) and N:P (r = +0.54, p < 0.01). Soil C:N exhibited a highly significant positive correlation with C:P (r = +0.65, p < 0.01). Soil C:P exhibited a highly significant positive correlation with N:P (r = +0.73, p < 0.01) (Figure 5).
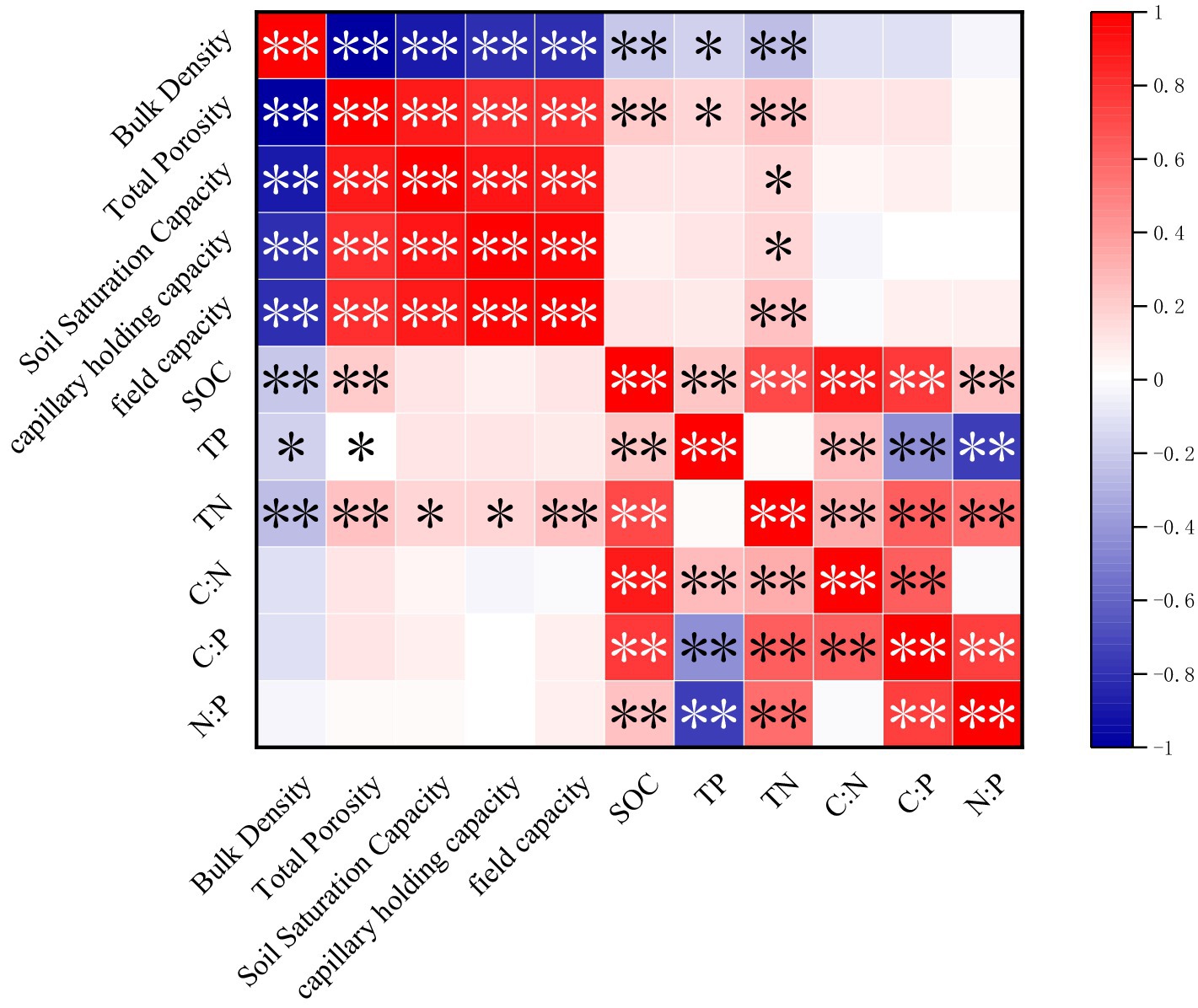
Figure 5. Pearson correlation analysis of various soil factors.
The observed differences highlight the significant influence of stand structure on soil properties, offering important implications for forest management practices. Key structural characteristics, such as stand density and canopy cover, are crucial in shaping the microenvironment within forest ecosystems (del Campo et al., 2022). Forests with a well-developed canopy tend to exhibit higher soil water retention, owing to reduced bulk density and enhanced porosity (Zuo et al., 2023; Gong et al., 2024). These findings offer valuable insights for promoting plant growth, enhancing water cycling, and improving drought resistance in forest ecosystems. Soil water retention capacity is a key indicator of its ability to supply water, directly influencing plant growth and ecosystem water cycling dynamics (Xia and Shao, 2008). A higher bulk density typically signifies more compacted soil, reduced pore space, and limited water infiltration and storage, which may hinder root expansion (Wang et al., 2024). Conversely, increased porosity is typically linked to better water retention and aeration, which promotes root proliferation and enhances microbial activity. Therefore, in forest management and soil conservation, it is essential to consider the effects of soil depth variation on physical properties, while optimizing stand structure and surface cover strategies to enhance soil structure and water retention capacity.
Well-structured stands exhibit higher soil organic carbon and total nitrogen content than those with moderate or poor structures. This is primarily attributed to the richer understory vegetation and biodiversity in well-structured stands (Smyčková et al., 2024), which facilitate the input and accumulation of organic matter, such as leaf litter, decomposing plant layers, and root exudates. Soil organic carbon is predominantly influenced by organic matter, which enhances nitrogen accumulation through the decomposition of organic matter into nitrogenous compounds. Well-structured stands usually have a more robust root system, which contributes to increased soil carbon and nitrogen through root growth, exudation, and decomposition processes (Smyčková et al., 2024). These roots also play a crucial role in forming and stabilizing soil aggregates, which increases soil porosity and enhances water and nutrient retention, thereby promoting organic carbon accumulation and nutrient cycling (Bodner et al., 2021). The study also underscores the importance of considering vertical variability in forest soil dynamics, as soil depth is linked to key soil properties affecting organic matter, nutrient distribution, and microbial activity (Walkiewicz et al., 2021). With increasing soil depth, organic carbon and total nitrogen decrease, while total phosphorus content remains relatively stable across depths, with a notable decline observed at 20–30 cm. The surface soil layer is the main site for organic matter input, where plant residues accumulate most, resulting in the highest levels of organic carbon and nitrogen. High metabolic activity and microbial diversity in the surface layer facilitate the rapid decomposition of organic matter, releasing plant nutrients (Condron et al., 2010). As soil depth increases, microbial abundance and activity decrease, leading to reduced organic matter decomposition in deeper layers (Muindi, 2019), which explains the gradual decline in organic carbon and nitrogen content in the soil profile. In contrast, phosphorus content shows less variation across the soil profile, but decreases significantly between 20 and 30 cm, primarily due to the behavior of phosphorus in the soil, which is influenced by mineral composition, pH, phosphorus adsorption and desorption, its mineral solubility, and root absorption processes (Muindi, 2019; Amadou et al., 2022).
The C:P ratio in soils of forests with well-structured stands is significantly higher than in those with moderate stand structures. Generally, a higher C:P ratio reflects a relative abundance of carbon over phosphorus, which may be attributed to the greater deposition of carbon-rich, phosphorus-deficient organic matter in well-structured stands, leading to higher organic carbon content. Likewise, the N:P ratio is significantly higher in soils of well-structured forests compared to those with moderate or poor stand structures. The N:P ratio is an important indicator of nutrient balance in the soil and reflects nutrient limitations within the ecosystem (Menge et al., 2012). Well-structured forests foster nitrogen-rich pools in the soil, maintaining elevated nitrogen levels through increased organic matter input, enhanced microbial activity, and improved nutrient cycling (Lucas-Borja et al., 2011). In forests where nitrogen is relatively abundant, phosphorus is often limited due to factors such as soil fixation, low bioavailability, or restricted absorption by plants and microbes, resulting in higher N:P ratios in the soil (Islam et al., 2024). As soil depth increases, the C:N, C:P, and N:P ratios typically decline. The surface soil layer typically receives the highest input of organic matter, including undecomposed or partially decomposed plant residues, resulting in higher C:N and C:P ratios. As soil depth increases, organic matter content and microbial activity decrease (Jobbagy and Jackson, 2000), causing a relative decline in carbon content compared to nitrogen and phosphorus. In the surface soil, decomposition of organic matter is facilitated by intense microbial activity, releasing nitrogen and phosphorus, while carbon is utilized as an energy source, thereby altering nutrient ratios (Chen et al., 2003; Condron et al., 2010). Decomposition of organic matter in deeper soils is slower, mainly due to limitations such as light, temperature, and moisture (Jobbagy and Jackson, 2000), which explains the reduced C:N and C:P ratios at greater depths.
This study performed a detailed Pearson correlation analysis to examine the relationships between soil physical properties and stoichiometric characteristics under different stand structures in the C. japonica plantations of Mount Lushan, aiming to clarify how these factors collectively influence forest ecosystem functionality (Zhang et al., 2013). The interactions and balance among the various physical and chemical properties of the soil matrix are essential for maintaining the vitality and functionality of forest ecosystems (Schoenholtz et al., 2000). Notably, soil physical properties such as bulk density, porosity, and water-holding capacity directly influence water retention and plant growth, thus affecting the water cycle within the ecosystem (Assouline, 2006). Additionally, the stoichiometric ratios of carbon, nitrogen, and phosphorus in the soil act as indicators of nutrient cycling dynamics and the capacity for plant nutrient absorption (Luo et al., 2020). The complex interrelationships among these factors, including the negative correlation between bulk density and porosity, and the positive correlation between porosity and water-holding capacity, offer valuable insights into soil structural integrity and its functional role.
Our findings highlight the importance of stand structure and soil depth in shaping soil properties, yet the role of forest site characteristics (e.g., topography, elevation, soil type, and rockiness) in modulating ecosystem services such as carbon sequestration, soil conservation, and biodiversity maintenance warrants further discussion.
The vertical variability in soil organic carbon and nitrogen observed in this study (e.g., higher surface-layer SOC and TN) may be amplified by topographic factors. For instance, steep slopes in mountainous regions (e.g., Mount Lu) are prone to erosion, which can accelerate nutrient leaching and reduce soil organic matter accumulation in lower slopes (Cislaghi et al., 2021). Conversely, gentle slopes or concave landforms may retain more moisture and litter inputs, enhancing carbon storage and microbial activity (Rossi et al., 2025). These dynamics align with findings in Mediterranean beech forests, where elevation-dependent temperature shifts influence species distribution and carbon sink capacity (Rossi et al., 2025).
The rocky soils of Mount Lu (classified as mountain brown and yellow soils) impose constraints on root penetration and water-holding capacity. In such environments, rock outcrops create heterogeneous soil pockets where litter accumulates locally, forming “carbon hotspots” despite lower overall bulk density (Wang et al., 2016; Cislaghi et al., 2021). However, shallow soils in rocky areas limit deep root development, reducing root reinforcement against landslides—a critical protective ecosystem service. This echoes studies in the Southern Alps, where root reinforcement was significantly lower in rocky, steep terrains despite similar stand structures (Cislaghi et al., 2021).
Optimizing stand structure for ecosystem services requires site-specific strategies (Karimi et al., 2022). In low-elevation areas with higher drought risk, promoting mixed stands (e.g., C. japonica with broadleaves) could enhance water-use efficiency and carbon sequestration through complementary root architectures (Rossi et al., 2025). On steep slopes, maintaining moderate tree density and avoiding group thinning (which drastically reduces root cohesion) is critical to prevent soil erosion, as demonstrated in Alpine protection forests (Cislaghi et al., 2021). Additionally, preserving surface organic layers in rocky soils through reduced disturbance can stabilize microhabitats, supporting understory biodiversity and nutrient cycling (Smyčková et al., 2024).
Our results corroborate and extend previous research on stand structure and soil property relationships. For instance, research in Mediterranean beech forests (Rossi et al., 2025) similarly demonstrated that reduced canopy closure and intermediate stand density facilitate greater porosity and organic C accumulation in the soil, in agreement with our findings for C. japonica stands. Structural indices such as the Shannon index and diameter differentiation have proven effective in capturing these dynamics (Pommerening, 2002). Where we differ, however, is in phosphorus dynamics: whereas our study found invariant TP concentrations with depth in well-stratified stands, research in subtropical mixed forests (Xiao et al., 2022) demonstrated pronounced vertical phosphorus stratification, perhaps a function of parent material and microbial activity variation.
The positive effects of well-structured stands for soil health concur with Alpine protection forests (Cislaghi et al., 2021), in which heterogeneous canopy cover and intermediate tree density reduced erosion and enhanced nutrient conservation. Recent studies emphasize that redundancy among structural indices (e.g., strong correlations between Shannon and Mingling indices) necessitates careful selection to avoid overinterpretation (Alterio et al., 2021). In addition, our work identifies that optimizing stand structure—by maintaining low canopy closure (<0.5) and high understory vegetation cover (>0.8)—can improve soil water-holding capacity and organic matter input. For foresters, this suggests prioritizing structural diversity in reforestation efforts, such as thinning overstocked stands to reduce competition and promoting understory vegetation through selective logging or controlled burning (Stephens et al., 2023). Additionally, incorporating mixed-species plantings (e.g., C. japonica with nitrogen-fixing broadleaves) could synergistically enhance soil fertility and resilience (Forrester et al., 2006).
This study has several limitations. First, it was conducted in a single geographic location (Mount Lu), meaning its findings may not be fully representative of C. japonica plantations in different climatic or edaphic environments. Second, the cross-sectional nature of the study limits our ability to infer long-term processes, such as how soil properties evolve with stand age or climate change. Third, while we focused on stand structure and soil depth, other important factors—such as microbial community composition, root exudate chemistry, and atmospheric deposition—were not measured, potentially overlooking key drivers of nutrient cycling.
Future research should broaden the geographic scope to include C. japonica stands across a wider range of environmental conditions, allowing for a more comprehensive assessment of structural effects (Kitagami et al., 2022). Long-term monitoring will be crucial to understanding temporal variations in soil properties and evaluating the effectiveness of different management strategies (Bissett et al., 2011). Experimental interventions, such as controlled thinning trials or litter addition/removal experiments, would further clarify the causal links between stand structure and soil processes (Baena et al., 2013; Wu et al., 2022).
This study examines the effects of different stand structures and soil depths on the physical and chemical properties of soil in Japanese cedar plantations on Mount Lushan. The results show that stand structure and soil depth significantly affect both the physical properties (e.g., soil bulk density and porosity) and the chemical properties [e.g., carbon (C), nitrogen (N), phosphorus (P), and their stoichiometric ratios]. A well-structured stand reduces soil bulk density and increases porosity, thereby enhancing the soil’s water-holding capacity, which is key for improving water retention and bolstering drought resilience in forest ecosystems. Additionally, the content of soil organic carbon and total nitrogen decreases with increasing soil depth. In contrast, the surface soil, with higher organic matter input and more active microbial activity, promotes greater nutrient accumulation. In well-structured stands, the C:P and N:P ratios are generally higher, indicating a greater relative abundance of carbon and nitrogen, likely due to increased organic matter input and enhanced microbial activity. As soil depth increases, the C:N and C:P ratios generally decrease, reflecting the high initial input and rapid decomposition of organic matter in the surface soil. These findings provide a scientific foundation for understanding how changes in forest structure affect soil properties and forest ecosystem functions, with important implications for forest management and ecological restoration. Optimizing stand structure by maintaining low canopy closure and high understory vegetation cover through selective thinning and understory protection, combined with stratified soil management (e.g., preserving surface organic layers to enhance carbon accumulation and implementing deep-soil moisture regulation to sustain microbial activity), can effectively improve soil porosity, water retention, and nutrient stoichiometry. These practices, alongside mixed-species planting (e.g., integrating nitrogen-fixing broadleaf species with C. japonica), will enhance soil resource utilization across depths, thereby supporting forest ecosystem resilience and sustainable productivity.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
LL: Writing – original draft, Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Writing – review & editing. KL: Investigation, Methodology, Writing – review & editing. LT: Supervision, Writing – review & editing. CL: Investigation, Methodology, Writing – review & editing. JW: Writing – review & editing. TD: Investigation, Methodology, Writing – review & editing. YL: Software, Writing – review & editing. XF: Software, Writing – review & editing. SG: Supervision, Writing – review & editing. YL: Formal analysis, Funding acquisition, Writing – review & editing.
The author(s) declare that financial support was received for the research and/or publication of this article. This study was financially supported by the Jiangxi Provincial Forestry Innovation Project “Experimental Research and Demonstration on Close-to-Nature Transformation and Ecological Function Enhancement of Japanese Cedar and Japanese Cypress Plantations in Lushan Mountain”.
We sincerely acknowledge Jiahui Huang and Tianjun Bai for their enthusiastic assistance in the field work.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that Gen AI was used in the creation of this manuscript. Generative AI (ChatGPT) was used for language editing and enhancing the clarity of the manuscript. The author(s) confirm and take full responsibility for the use of generative AI in the preparation of this manuscript. All content and scientific analysis were independently conducted by the author(s), and the AI tool was only employed for improving the language and readability of the text.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2025.1548485/full#supplementary-material
Alterio, E., Cislaghi, A., Bischetti, G. B., and Sitzia, T. (2021). Exploring correlation between stand structural indices and parameters across three Forest types of the southeastern Italian Alps. Forests 12:1645. doi: 10.3390/f12121645
Amadou, I., Faucon, M.-P., and Houben, D. (2022). Role of soil minerals on organic phosphorus availability and phosphorus uptake by plants. Geoderma 428:116125. doi: 10.1016/j.geoderma.2022.116125
Assouline, S. (2006). Modeling the relationship between soil bulk density and the water retention curve. Vadose Zone J. 5, 554–563. doi: 10.2136/vzj2005.0083
Baena, C. W., Andrés-Abellán, M., Lucas-Borja, M. E., Martínez-García, E., García-Morote, F. A., Rubio, E., et al. (2013). Thinning and recovery effects on soil properties in two sites of a Mediterranean forest, in Cuenca Mountain (south-eastern of Spain). For. Ecol. Manag. 308, 223–230. doi: 10.1016/j.foreco.2013.06.065
Bissett, A., Richardson, A. E., Baker, G., and Thrall, P. H. (2011). Long-term land use effects on soil microbial community structure and function. Appl. Soil Ecol. 51, 66–78. doi: 10.1016/j.apsoil.2011.08.010
Bodner, G., Mentler, A., and Keiblinger, K. (2021). “Plant roots for sustainable soil structure management in cropping systems,” in The root systems in sustainable agricultural intensification. eds. Z. Rengel and I. Djalovic (Wiley, Novi Sad), 45–90.
Botequim, B., Bugalho, M. N., Rodrigues, A. R., Marques, S., Marto, M., and Borges, J. G. (2021). Combining tree species composition and understory coverage indicators with optimization techniques to address concerns with landscape-level biodiversity. Land 10:126. doi: 10.3390/land10020126
Chen, G., Zhu, H., and Zhang, Y. (2003). Soil microbial activities and carbon and nitrogen fixation. Res. Microbiol. 154, 393–398. doi: 10.1016/s0923-2508(03)00082-2
Cislaghi, A., Alterio, E., Fogliata, P., Rizzi, A., Lingua, E., Vacchiano, G., et al. (2021). Effects of tree spacing and thinning on root reinforcement in mountain forests of the European southern Alps. For. Ecol. Manag. 482:118873. doi: 10.1016/j.foreco.2020.118873
Condron, L., Stark, C., O’callaghan, M., Clinton, P., and Huang, Z. (2010). “The role of microbial communities in the formation and decomposition of soil organic matter,” in Soil microbiology and sustainable crop production. eds. G. Dixon and E. Tilston (Dordrecht: Springer), 81–118.
del Campo, A. D., Otsuki, K., Serengil, Y., Blanco, J. A., Yousefpour, R., and Wei, X. (2022). A global synthesis on the effects of thinning on hydrological processes: implications for forest management. For. Ecol. Manag. 519:120324. doi: 10.1016/j.foreco.2022.120324
Dibar, D., Zhang, K., Yuan, S., Zhang, J., Zhou, Z., and Ye, X. (2020). Ecological stoichiometric characteristics of carbon (C), nitrogen (N) and phosphorus (P) in leaf, root, stem, and soil in four wetland plants communities in Shengjin Lake, China. PLoS One 15:89. doi: 10.1371/journal.pone.0230089
Dlapa, P., Hriník, D., Hrabovský, A., Šimkovic, I., Žarnovičan, H., Sekucia, F., et al. (2020). The impact of land-use on the hierarchical pore size distribution and water retention properties in loamy soils. Water 12:339. doi: 10.3390/w12020339
Duan, A., Lei, J., Hu, X., Zhang, J., Du, H., Zhang, X., et al. (2019). Effects of planting density on soil bulk density, pH and nutrients of Unthinned Chinese fir mature stands in south subtropical region of China. Forests 10:351. doi: 10.3390/f10040351
Farahnak, M., Mitsuyasu, K., Hishi, T., Katayama, A., Chiwa, M., Jeong, S., et al. (2020). Relationship between very fine root distribution and soil water content in pre-and post-harvest areas of two coniferous tree species. Forests 11:1227. doi: 10.3390/f11111227
Farahnak, M., Mitsuyasu, K., Jeong, S., Otsuki, K., Chiwa, M., Sadeghi, S. M. M., et al. (2019). Soil hydraulic conductivity differences between upslope and downslope of two coniferous trees on a hillslope. J. Forest Res. 24, 143–152. doi: 10.1080/13416979.2019.1590967
Forrester, D. I., Bauhus, J., Cowie, A. L., and Vanclay, J. K. (2006). Mixed-species plantations of Eucalyptus with nitrogen-fixing trees: a review. For. Ecol. Manag. 233, 211–230. doi: 10.1016/j.foreco.2006.05.012
Franko, U., and Schulz, E. (2020). Carbon accumulation in a bare fallow Chernozem soil with high carbon input rates. European J. Soil Sci. 72, 265–273. doi: 10.1111/ejss.12937
Gong, C., Tan, Q., Liu, G., and Xu, M. (2024). Positive effects of mixed-species plantations on soil water storage across the Chinese loess plateau. For. Ecol. Manag. 552:121571. doi: 10.1016/j.foreco.2023.121571
Huang, L., and Shao, M. (2019). Advances and perspectives on soil water research in China’s loess plateau. Earth Sci. Rev. 199:102962. doi: 10.1016/j.earscirev.2019.102962
Ilek, A., Kucza, J., and Szostek, M. (2014). The effect of stand species composition on water storage capacity of the organic layers of forest soils. Eur. J. Forest Res. 134, 187–197. doi: 10.1007/s10342-014-0842-2
Islam, M., Siddique, K. H. M., Padhye, L. P., Pang, J., Solaiman, Z. M., Hou, D., et al. (2024). A critical review of soil phosphorus dynamics and biogeochemical processes for unlocking soil phosphorus reserves. Adv. Agronom 185, 153–249. doi: 10.1016/bs.agron.2024.02.004
Jobbagy, E. G., and Jackson, R. B. (2000). The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 10:423. doi: 10.2307/2641104
Joergensen, R. G., and Scheu, S. (1998). Response of soil microorganisms to the addition of carbon, nitrogen and phosphorus in a forest Rendzina. Soil Biol. Biochem. 31, 859–866. doi: 10.1016/S0038-0717(98)00185-0
Karimi, Z., Abdi, E., Deljouei, A., Cislaghi, A., Shirvany, A., Schwarz, M., et al. (2022). Vegetation-induced soil stabilization in coastal area: an example from a natural mangrove forest. Catena 216:106410. doi: 10.1016/j.catena.2022.106410
Kermavnar, J., Marinšek, A., Eler, K., and Kutnar, L. (2019). Evaluating short-term impacts of Forest management and microsite conditions on understory vegetation in temperate fir-beech forests: floristic, ecological, and trait-based perspective. Forests 10:909. doi: 10.3390/f10100909
Kitagami, Y., Obase, K., Chen, C.-F., and Matsuda, Y. (2022). Effects of climatic and edaphic conditions on structuring patterns of soil nematode communities in Japanese cedar (Cryptomeria japonica) plantations. For. Ecol. Manag. 524:120518. doi: 10.1016/j.foreco.2022.120518
Li, A., DeLuca, T. H., Sun, S., Zhang, J., and Wang, G. (2020). Bryophytes impact the fluxes of soil non-carbon dioxide greenhouse gases in a subalpine coniferous forest. Biol. Fertil. Soils 56, 1151–1163. doi: 10.1007/s00374-020-01483-8
Lucas-Borja, M. E., Candel, D., Jindo, K., Moreno, J. L., Andrés, M., and Bastida, F. (2011). Soil microbial community structure and activity in monospecific and mixed forest stands, under Mediterranean humid conditions. N.A. And Soil 354, 359–370. doi: 10.1007/s11104-011-1072-8
Luo, G., Xue, C., Jiang, Q., Xiao, Y., Zhang, F., Guo, S., et al. (2020). Soil carbon, nitrogen, and phosphorus cycling microbial populations and their resistance to global change depend on soil C N P stoichiometry. Msystems 5, 10–1128. doi: 10.1128/msystems.00162-20
Manns, H. R., Parkin, G. W., and Martin, R. C. (2016). Evidence of a union between organic carbon and water content in soil. Can. J. Soil Sci. 96, 305–316. doi: 10.1139/cjss-2015-0084
Manzoni, S., Trofymow, J. A., Jackson, R. B., and Porporato, A. (2020). Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter. Ecol. Monogr. 80, 89–106. doi: 10.1890/09-0179.1
Menge, D. N. L., Hedin, L. O., and Pacala, S. W. (2012). Nitrogen and phosphorus limitation over long-term ecosystem development in terrestrial ecosystems. PLoS One 7:e42045. doi: 10.1371/journal.pone.0042045
Muindi, E. M. (2019). Understanding soil phosphorus. Int. J. Plant Soil Sci. 31, 1–18. doi: 10.9734/ijpss/2019/v31i230208
Pommerening, A. (2002). Approaches to quantifying forest structures. Forestry 75, 305–324. doi: 10.1093/forestry/75.3.305
Rossi, L. M. W., Oggioni, S. D., Brocco, S., Djacenko, S., and Vacchiano, G. (2025). Interplay of species mixture, climate change, and management regimes on carbon stocks and sinks in a Mediterranean beech forest. For. Ecol. Manag. 578:122472. doi: 10.1016/j.foreco.2024.122472
Schoenholtz, S. H., Miegroet, H. V., and Burger, J. A. (2000). A review of chemical and physical properties as indicators of forest soil quality challenges and opportunities. For. Ecol. Manag. 138, 335–356. doi: 10.1016/S0378-1127(00)00423-0
Sekucia, F., Dlapa, P., Kollár, J., Cerdá, A., Hrabovský, A., and Svobodová, L. (2020). Land-use impact on porosity and water retention of soils rich in rock fragments. Catena 195:104807. doi: 10.1016/j.catena.2020.104807
Shi, S., Peng, C., Wang, M., Zhu, Q., Yang, G., Yang, Y., et al. (2016). A global meta-analysis of changes in soil carbon, nitrogen, phosphorus and sulfur, and stoichiometric shifts after forestation. Plant Soil 407, 323–340. doi: 10.1007/s11104-016-2889-y
Smyčková, M., Koutecký, T., Ujházyová, M., Ujházy, K., Verheyen, K., Volařík, D., et al. (2024). Herb layer species richness declines with heterogeneity of the forest structure in primary beech-dominated forests while proportion of forest specialists increases. For. Ecol. Manag. 556:121728. doi: 10.1016/j.foreco.2024.121728
Stephens, S. L., Foster, D. E., Battles, J. J., Bernal, A. A., Collins, B. M., Hedges, R., et al. (2023). Forest restoration and fuels reduction work: different pathways for achieving success in the Sierra Nevada. Ecol. Appl. 34:e2932. doi: 10.1002/eap.2932
Tague, C. L., and Moritz, M. A. (2019). Plant accessible water storage capacity and tree-scale root interactions determine how Forest density reductions Alter Forest water use and productivity. Front. Forest Glob Change 2:36. doi: 10.3389/ffgc.2019.00036
Walkiewicz, A., Bieganowski, A., Rafalska, A., Khalil, M. I., and Osborne, B. (2021). Contrasting effects of Forest type and stand age on soil microbial activities: an analysis of local scale variability. Biology 10:850. doi: 10.3390/biology10090850
Wang, D., Niu, J., Yang, T., Miao, Y., Zhang, L., Chen, X., et al. (2024). Soil water infiltration characteristics of reforested areas in the paleo-periglacial eastern Liaoning mountainous regions, China. CATENA 234:107613. doi: 10.1016/j.catena.2023.107613
Wang, D., Shen, Y., Li, Y., and Huang, J. (2016). Rock outcrops redistribute organic carbon and nutrients to nearby soil patches in three karst ecosystems in SW China. PLoS One 11:e0160773. doi: 10.1371/journal.pone.0160773
Wu, F., Xu, Z., and Yang, W. (2022). Carbon and nutrient transfer via above-and below-ground litter in forests. Forests 13:2176. doi: 10.3390/f13122176
Xia, Y. Q., and Shao, M. A. (2008). Soil water carrying capacity for vegetation: a hydrologic and biogeochemical process model solution. Ecol. Model. 214, 112–124. doi: 10.1016/j.ecolmodel.2008.01.024
Xiao, T., Wang, C., Yuan, X., Tao, L., Li, P., Deng, W., et al. (2022). Effects of different Forest gap ages on soil physical properties and stoichiometric characteristics in Cryptomeria japonica plantations (L.f.) D. Don, 1839. Forests 13:1708. doi: 10.3390/f13101708
Yang, F., Zhang, G. L., Yang, J. L., Li, D. C., Zhao, Y. G., Liu, F., et al. (2014). Organic matter controls of soil water retention in an alpine grassland and its significance for hydrological processes. J. Hydrol. 519, 3086–3093. doi: 10.1016/j.jhydrol.2014.10.054
Yu, M.-F., Tao, Y., Liu, W., Xing, W., Liu, G., Wang, L., et al. (2019). C, N, and P stoichiometry and their interaction with different plant communities and soils in subtropical riparian wetlands. Environ. Sci. Pollut. Res. 27, 1024–1034. doi: 10.1007/s11356-019-07004-x
Zhang, Z.-S., Song, X.-L., Lu, X.-G., and Xue, Z.-S. (2013). Ecological stoichiometry of carbon, nitrogen, and phosphorus in estuarine wetland soils: influences of vegetation coverage, plant communities, geomorphology, and seawalls. J. Soils Sedim. 13, 1043–1051. doi: 10.1007/s11368-013-0693-3
Keywords: Cryptomeria japonica plantations, stand structure, soil layer depth, soil physical properties, soil ecological stoichiometry
Citation: Luo L, Lin K, Tao L, Luo C, Wang J, Duan T, Liu Y, Fu X, Guo S and Liu Y (2025) Effects of stand structure and soil depth on soil properties in Cryptomeria japonica plantations. Front. For. Glob. Change. 8:1548485. doi: 10.3389/ffgc.2025.1548485
Edited by:
Azade Deljouei, University of Florida, United StatesReviewed by:
Seyed Mohammad Moein Sadeghi, University of Florida, United StatesCopyright © 2025 Luo, Lin, Tao, Luo, Wang, Duan, Liu, Fu, Guo and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuanqiu Liu, bGl1eXE0MDRAMTYzLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.