 Xiaoxuan Du
Xiaoxuan Du Xinyi Li
Xinyi Li Jiangfeng Wang2†
Jiangfeng Wang2† Jie Gao
Jie Gao- 1College of Marine and Bioengineering, Yancheng Institute of Technology, Yancheng, Jiangsu, China
- 2Key Laboratory for the Conservation and Regulation Biology of Species in Special Environments, College of Life Science, Xinjiang Normal University, Urumqi, China
The management mode of forests has significant impacts on soil nutrients. However, with global changes, there is scant evidence to suggest whether the soil nutrients in planted and natural forests have a consistent response mechanism to environmental changes. Utilizing soil nutrient data from 263 planted forests and 434 natural forests in China, collected through field surveys of 298 forests and literature searches from 2005 to 2020, this study explores the differences in soil nutrients between natural and planted forests and their controlling factors. The results indicate that the soil available phosphorus content in natural forests is significantly higher than in planted forests (p < 0.001), and the soil pH is significantly lower than in planted forests (p < 0.001), while there is no significant difference in soil nitrogen content between the two (p > 0.05). With increases in Mean Annual Temperature (MAT) and Mean Annual Precipitation (MAP), soil available phosphorus content significantly increased, and soil pH significantly decreased (p < 0.001). Stand factors (such as stand age and stand density) have a greater influence on soil nutrients in natural forests than in planted forests. Climate factors contribute the most to the spatial variability of soil nutrients in both planted and natural forests. Compared with climate factors, stand factors and forest key leaf traits (such as leaf area, specific leaf area, leaf nitrogen and phosphorus content) had relatively little effect on soil nutrients in planted and natural forests. Climate factors directly or indirectly affect the soil nutrients of planted and natural forests by influencing stand factors and key leaf functional traits, and their direct effects are greater than their indirect effects. The results of this study demonstrate that forest soil nutrients of different types respond to global change in distinct patterns. In future forest management, special attention should be paid to the differences between artificial forests and natural forests.
1 Introduction
Soil is the direct living environment for plants. Soil contains a large amount of nutrients as the material basis for plant growth (Wang Z. et al., 2022), which plays a key role in plant growth and development. Nitrogen (N) and available phosphorus (AP) are two key nutrients that limit plant growth and development in terrestrial ecosystems (Elser et al., 2007; Reich and Oleksyn, 2004). Soil pH is also a critical soil nutrient factor that regulates biogeochemical processes, significantly affecting the structure and function of forest ecosystems. From 1990 to 2020, the global area of planted forests increased by 123 million hectares (FAO, 2020). Planted forests are playing an increasingly important ecological role in global change. However, due to differences in forest origins, it remains unclear whether planted and natural forests play the same role in soil nutrient retention. Exploring the differences in soil nutrients between planted and natural forests and their controlling factors at a macro scale is crucial for understanding the impact of global change on the ecosystem functions of different types of forests (planted vs. natural forests), such as surface carbon storage, soil conservation, and water conservation (Hua et al., 2022).
Climatic factors, especially Mean Annual Temperature (MAT) and Mean Annual Precipitation (MAP), have a significant impact on soil nutrients. Research indicates that reduced precipitation can limit soil weathering as well as the production and mineralization of organic matter, slow down the release of phosphorus from primary substances, decrease the availability of soil nitrogen and available phosphorus, and thereby reduce soil nutrient content (He et al., 2014). Excessive precipitation may also lead to leaching of soluble nutrients in the soil (especially nitrogen and available phosphorus, as well as acidic substances like ammonium ions), which may lead to the loss of nutrients such as secondary minerals phosphorus in the soil, reducing soil nitrogen and available phosphorus content (Rashmi et al., 2017), resulting in an increase in soil pH. Temperature affects the activity of soil microbes, with higher temperatures generally accelerating microbial metabolic processes, promoting the decomposition of organic matter and enhancing the activity of nitrate nitrogen and available phosphorus in soil (Li et al., 2023), and thus speeding up the release of nutrients such as nitrogen and available phosphorus (St Clair and Lynch, 2010). Temperature also affects nitrogen mineralization (conversion of organic nitrogen to inorganic nitrogen) and nitrification (conversion of ammonium to nitrate), significantly impacting soil nitrogen, available phosphorus content, and pH (Gao et al., 2015). Additionally, the combined effects of temperature and precipitation (evaporation) on soil nutrients are significant. Under drought conditions, microbial activity is reduced due to water limitation, slowing down the mineralization of organic nitrogen and promoting nitrification, converting ammoniacal nitrogen to nitrate nitrogen (Grzyb et al., 2021). Meanwhile, soil available phosphorus mainly exists in a fixed form, and evaporation affects soil moisture dynamics, indirectly influencing phosphorus solubility and plant availability, leading to soil surface salinity accumulation and thereby increasing soil pH (Naorem et al., 2023).
Stand characteristics, such as stand density, stand age, and key leaf traits, are also crucial factors affecting forest soil nutrients. Studies have shown that under higher stand densities, intense competition for limited soil nutrients occurs among tree roots, leading to a reduction in the availability of nitrogen and available phosphorus in the soil (Bo et al., 2018). High stand density also affects the accumulation and decomposition rate of surface litter, further influencing the release and cycling of nutrients like nitrogen and available phosphorus. This may result in the accumulation of more organic litter, whose decomposition produces acidic or alkaline substances affecting soil pH (Prescott and Vesterdal, 2021; Wang L. et al., 2022). In the early stages of forest development (younger stand ages), trees generally grow faster and have a higher demand for nutrients such as nitrogen and available phosphorus, leading to a rapid decrease in soil nutrients. As the forest matures, biomass accumulation increases, as does the amount of surface litter, leading to the accumulation of nutrients (especially nitrogen) in the soil (Chen X. et al., 2023). Species with higher Specific Leaf Area (SLA) and lower Leaf Dry Matter Content (LDMC) typically have thin, fast-growing leaves, higher photosynthetic efficiency, and faster nutrient cycling, more rapidly cycling soil nitrogen and available phosphorus elements (Banerjee et al., 2017). Plants with high nitrogen and phosphorus content in their leaves usually exhibit higher productivity, accelerating nutrient cycling, resulting in higher soil nitrogen and available phosphorus content in litter (Ren et al., 2019). Such plants release nitrogen and available phosphorus more quickly during decomposition, thus affecting soil nutrient content (Hobbie, 2015).
With global change, atmospheric nitrogen deposition also has a significant impact on soil nutrients. Atmospheric nitrogen deposition directly increases the supply of nitrogen in the soil, especially in nitrogen-limited ecosystems, altering their nutrient balance and promoting plant growth (Cheng et al., 2019). Certain forms of nitrogen deposition, such as nitrates (NO3−) and sulfates (SO42−), can lead to soil acidification, lowering the soil’s pH (Chen C. et al., 2023). In temperate forest soils where nitrogen does not limit microbial growth, nitrogen deposition can hinder the decomposition of organic matter, resulting in an increase in soil organic matter and higher soil nutrients (Janssens et al., 2010). Studies have shown that nitrogen deposition has increased the total nitrogen content in the leaves and roots of both planted and natural forests (Ma et al., 2018; Tian et al., 2018). However, other research has found that on a macro scale, nitrogen deposition significantly increases the total nitrogen content in the topsoil of planted forests but has no significant effect on the total nitrogen content in the topsoil of natural forests (Song et al., 2017).
Forests can be classified into natural and planted forests based on their origin. Preliminary studies have found that stand type has a significant impact on soil nutrient characteristics (Ni et al., 2022). Natural forests are mostly mixed forests with a variety of tree species and complex stand structures. Dozens of plant species alternate and depend on each other, possessing strong soil and water conservation abilities that can prevent soil erosion and water pollution. Planted forests, due to their monoculture planting and shallower root systems, tend to have relatively poor soil and water conservation effects (Hou et al., 2009). However, some studies have found that certain planted forests have a higher soil nutrient retention effect than natural forests (Brockerhoff et al., 2008; Lugo et al., 1990). At a macro scale, there is still no consensus on the soil nutrient retention capabilities of planted versus natural forests.
Based on soil data collected through field surveys and literature from 2005 to 2020, this study aims to explore the key biotic and abiotic factors affecting the distribution of soil nutrients in natural and planted forests at a macro scale, and to compare the soil nutrient retention capabilities of planted and natural forests. To address the questions above, we propose the following hypotheses: (1) The soil nitrogen and available phosphorus content in natural forests is significantly higher than in planted forests, while the pH is significantly lower than in planted forests. (2) Climatic factors are the key factors dominating the spatial distribution of soil nutrients in planted and natural forests at a macro scale, with stand factors also playing a non-negligible role. (3) Climatic factors can not only directly affect soil nutrients but can also influence soil nutrients in planted and natural forests through their effects on stand factors, with direct effects being greater than indirect effects.
2 Materials and methods
2.1 Experimental sites and soil nutrients data
The data collection comes partly from literature search and partly from field measurement. We searched relevant peer-reviewed published journal articles from 2005 to 2020 from Web of Science, Google Scholar, and CNKI. The key words used in the search were: “natural forest,” “planted forest,” “soil physical and chemical properties.” 182 relevant papers were retrieved. Then the data was screened in the literature using the following criteria: (1) The study should provide the latitude and longitude of the plot, and whether the forest in the plot is a natural forest or a planted forest. (2) The study should provide complete site and forest information, including the plot location, tree species, age of the stand, tree DBH, and tree density. (3) The study should provide or allow the calculation of the mean value, standard deviation, or standard error of the data on the physical and chemical properties of the soil in the plot. (4) The study presents the results of field studies rather than retrospective or simulation studies. For the articles meeting our standards, soil nitrogen, soil available phosphorus and soil pH indexes in the physical and chemical properties of 0–20 cm soil surface were extracted. If a study has multiple sampling depths from 0 to 20 cm at the same site, we treat these observations as independent samples. In these articles, stand characteristics such as age, stand density and DBH of each plot were collected.
Based on field surveys, 21 sites were selected to conduct actual measurements of soil nutrients data in 298 forests. At each site, at least four 30 × 30 m forest plots were selected, each representing the typical zonal vegetation of the area. We recorded the latitude, longitude, elevation and slope of each site for comprehensive analysis, and recorded the site location, tree species, age, DBH and tree density in real time. In the actual sampling, we used the same method as in the literature to measure the physical and chemical properties of soil. In each forest plot, a five-point sampling method was adopted. Using a soil corer, soil samples were collected from different directions at depths of 0–30 cm. The soil samples were air-dried at room temperature. After air drying, roots, stones, and other debris were removed. The samples were then gently ground only and sieved through a 2 mm mesh, and preserved for the determination of soil N and available P contents as well as pH. The soil N content was determined by the Kjeldahl method. KCl solution was added to the soil samples to extract NH4+ from the soil. At the same time, NO3− in the soil is extracted by adding saturated CaSO4 solution (Verma and Sagar, 2020). Based on the off-line column extraction method, the soil samples were extracted using HCl and MH2SO4 solutions, and then the available phosphorus content in the soil samples was analyzed by using the FI amperometric method (Jakmunee and Junsomboon, 2009). And soil pH was measured in a 1:2.5 soil and deionized water mixture for each sample using a pH meter (PHS-3C, Lei ci). Before measurement, the soil solution was shaken for 30 min and then boiled for 5 min. The Soil Inorganic Carbon (SIC) was determined using the 08.53 calculator (M1.08.53.E, Eijkelkamp). Here, SIC refers to the carbonate value measured by the amount of carbon dioxide (CO2) emission during digestion with strong acid (Hong et al., 2019).
2.2 Data sources and processing
2.2.1 Climate factors data
The data for the Mean Annual Temperature (MAT), Mean Annual Temperature of the Coldest Month (MACT), Mean Annual Temperature of the Hottest Month (MAHT), and Mean Annual Precipitation (MAP) were extracted from the WorldClim (version 2.0) global climate database with a spatial resolution of 1 km.1 The data for the Average Sunshine Duration (ASD) were sourced from the China Meteorological Data Service Centre,2 and the Mean Annual Evaporation (MAE) was also obtained from the China Meteorological Data Service Centre (see text footnote 2).
2.2.2 Leaf functional traits data
We selected four leaf functional traits to define resource utilization strategies for natural and planted forests in our dataset: Specific Leaf Area (SLA), Leaf Dry Matter Content (LDMC), Leaf nitrogen (LN), and Leaf phosphorus (LP). Trait values were obtained from the TRY.3 For sites that are actually measured in the field, we measured the fresh single leaf area of leaves without petioles using a Japanese Cano Scan LIDE 110 portable leaf area meter. The fresh leaf weight was measured with an electronic balance (precision of 0.0001 g), and the leaves were then dried in an oven at 105°C before the temperature was lowered to 60°C. After drying, the leaf dry weight was measured with a 1/10,000 electronic balance. SLA (m2/kg) is calculated as leaf area/dry leaf weight, and LDMC (g/g) as leaf dry weight/fresh leaf weight. For the analysis of leaf nutrient LN, LP, the Kjeldahl method is used to determine the total nitrogen content of the leaves, and the Mo-Sb colorimetric method is used to determine the total phosphorus content of the leaves (Dibar et al., 2020). The functional traits in this study refer to community-level functional traits in forest ecosystems.
2.2.3 Total nitrogen deposition data
The total nitrogen deposition includes both dry nitrogen deposition and wet nitrogen deposition. The data on total nitrogen deposition were extracted from the Resource Sharing Platform of the China National Ecosystem Science Data Center, which has a spatial resolution of 1 km.4
2.3 Statistical analysis
A t-test at a significance level of 0.05 was used to examine the differences in soil N, available P, and pH between plantation and natural forests. Significance analysis of differences was conducted in the R software package (version 4.3.1) “agricolae.” Principal Component Analysis (PCA) for dimensionality reduction of leaf functional traits data was performed using the R package “pcaMethods,” with the two principal components (PC1 and PC2) obtained representing key leaf functional traits (Homeier et al., 2021). Among them, PC1 is the first principal component extracted from leaf functional traits, and PC2 is the second principal component. Climate factors include mean annual temperature (MAT), mean temperature of the coldest month (MACT), mean temperature of the hottest month (MAHT), mean annual precipitation (MAP), mean annual evaporation (MAE), and mean annual sunshine duration (ASD). Stand factors include stand age, mean diameter at breast height, and stand density.
A linear mixed-effects model was employed to explore the impacts of climate factors, stand factors, key leaf traits, and nitrogen deposition on soil nutrients in plantation and natural forests, while fully considering the effect of random effects on the outcomes. This analysis was conducted using the R package “lme4” (Gurevitch et al., 2018). The R package “linkET” was used to test for multicollinearity among various potential influencing factors, and the correlations between these factors and soil N, available P, pH were visualized as heatmaps (Zhao et al., 2014).
Variance decomposition analysis was used to explore the contribution (%) of various influencing factors to the spatial variability of soil nutrients in plantation and natural forests, and this analysis was completed using the “vegan” package in R language (Kong et al., 2022). The boosted regression analysis method was employed to quantify the independent contribution of each influencing factor to the spatial variability of soil nutrients in plantation and natural forests, and this analysis was conducted using the “randomForest” package in R language (Yang X. et al., 2021).
To delve deeper into the mechanisms and pathways influencing the spatial variability of soil nutrients in plantation and natural forests, a structural equation model (SEM) was constructed to examine the direct and indirect effects of key influencing factors on soil N, available P, and pH. Observed variables were initially divided into composite variables and incorporated into structural equation models (SEMs). To validate the reliability of the relationship between key ecological factors and soil N, available P, pH, piecewiseSEM was employed to clarify the random effects of sampling points and provide the contributions of environmental predictive factors. These analyses were conducted using the “piecewiseSEM,” “nlme,” and “lme4” packages (Yang L. et al., 2021). The goodness of fit of the modeling results was assessed using the Fisher C test. Based on a significance level of p < 0.05 and satisfactory model fit (0 ≤ Fisher’s C/df ≤ 2 and 0.05 < p < 1.00), the model was progressively adjusted through a stepwise modification process (Figure 1).
Figure 1. Graphic abstract on the significant differences in soil nutrients between natural forests and planted forests, and how climate factors, stand factors and leaf functional traits affect the distribution of soil nutrients in natural and planted forests at the macro scale.
3 Results
3.1 Comparison of the significant differences in soil nutrients between planted and natural forests
The soil available P content in natural forests is approximately 9–15 mg/kg, while in planted forests, it is about 2–12 mg/kg. The available P content in the soil of natural forests is significantly higher than that in planted forests (p < 0.001), with the soil available P content in natural forests being about 71.4% higher than in planted forests. The soil pH range in natural forests is approximately 3–6, whereas in planted forests, it ranges from about 5–7.5. The soil pH in natural forests is significantly lower than that in planted forests (p < 0.001). However, there is no significant difference in soil N content between natural and planted forests (p > 0.05) (Figure 2).
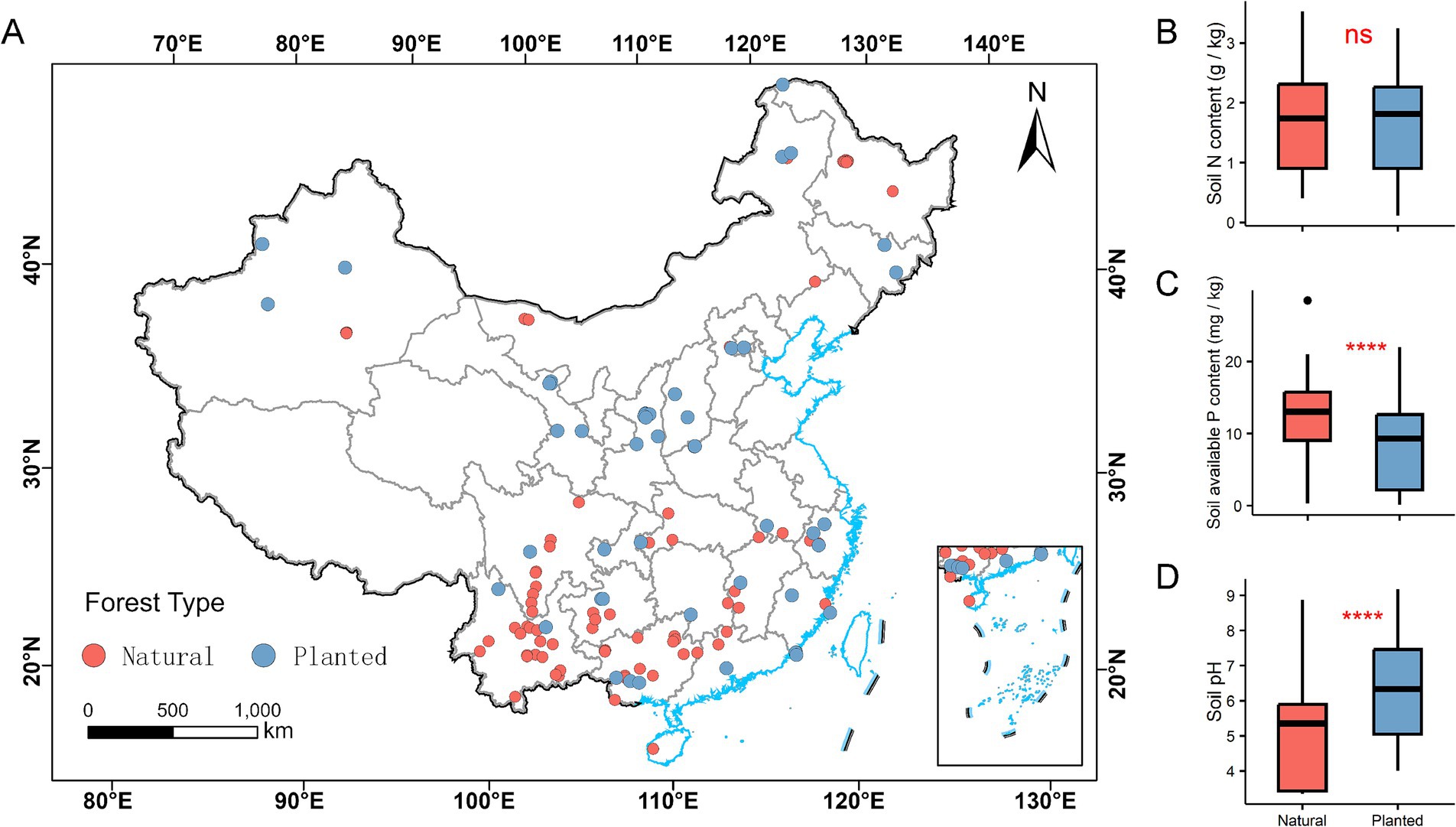
Figure 2. Location of natural and planted forests sample sites in China (A) and comparison of soil nitrogen content (B), soil available phosphorus content (C), and pH (D) differences. Differences were compared using a t-test with significance at the 0.05 level. ***p < 0.001, ns p > 0.05.
3.2 Correlations between potential influencing factors and soil nutrients
With the increase in Mean Annual Temperature (MAT) and Mean Annual Precipitation (MAP), soil available phosphorus (AP) content significantly increased in both natural and planted forests (p < 0.001), while soil pH significantly decreased. The soil nitrogen (N) content in planted forests did not significantly change with the increase in MAT and MAP, whereas in natural forests, soil N content significantly decreased (p < 0.001). Additionally, under the combined effects of MAT and MAP, Mean Annual Evaporation (MAE) showed a positive correlation with soil N content and pH, but a negative correlation with soil available P content (Supplementary Figures S1–S3). With the increase in nitrogen deposition, the soil N content in both planted and natural forests increased, but the change was not significant; however, soil pH significantly decreased. The soil available P content in planted forests significantly increased with the increase in nitrogen deposition (p < 0.001), while there was no significant change in the soil P content in natural forests (p > 0.05) (Figure 3).
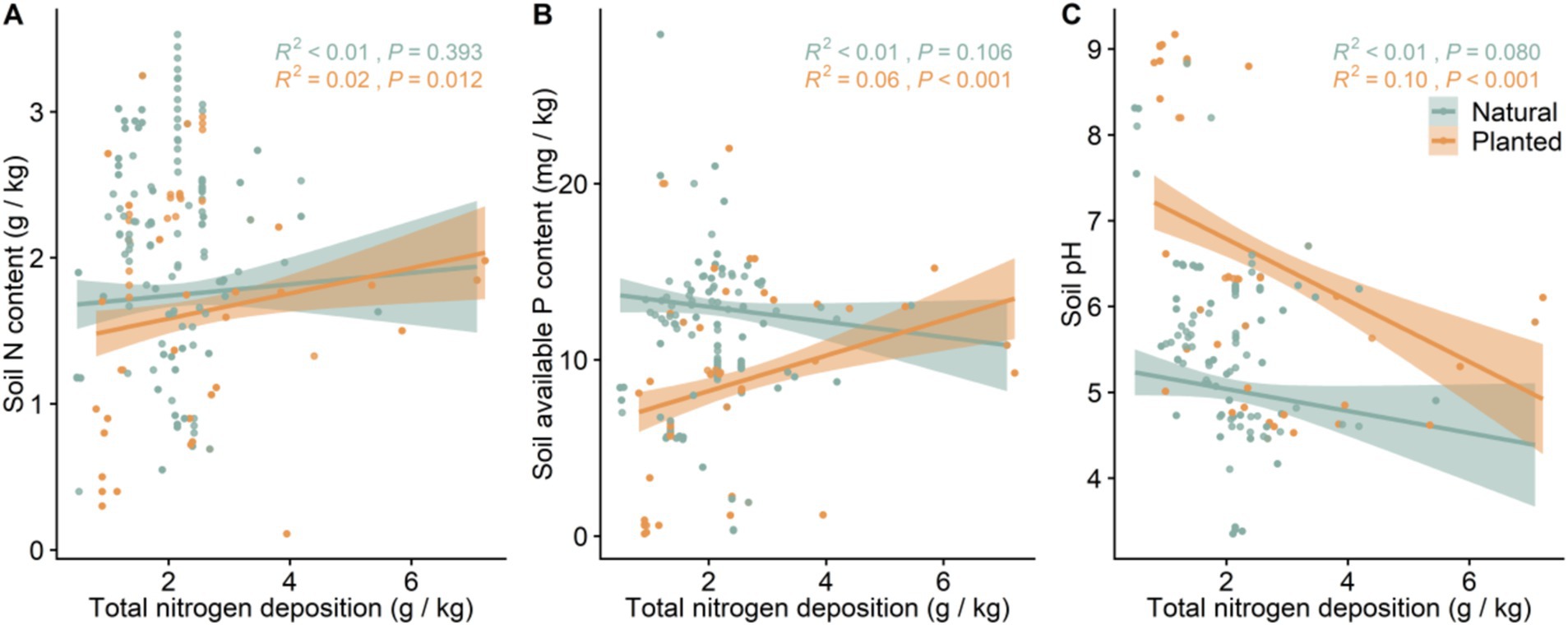
Figure 3. The linear relationship between soil nutrients and total nitrogen deposition. Soil nutrients include: (A) soil nitrogen content; (B) soil available phosphorus content; (C) soil pH. R2 indicates the model fit, and the p-value represents significance.
Overall, with the increase in stand density, average diameter at breast height, and stand age, the soil nutrients (N, available P, and pH) in planted forests did not show a significant trend of change (p > 0.05). In natural forests, soil nitrogen (N) content and pH significantly increased (p < 0.001), while soil available phosphorus (AP) content significantly decreased. The impact of stand factors on soil nutrients in natural forests was greater than in planted forests (Supplementary Figure 4).
Compared to climate factors and stand factors, key leaf traits have a relatively minor impact on both planted and natural forests. Overall, the explanatory power of key leaf traits on the spatial variability of soil nutrients is stronger in planted forests than in natural forests (Supplementary Figure 5).
3.3 Direct and indirect effects of key factors on soil nutrients
The results of variance decomposition indicate that soil nutrients in both planted and natural forests exhibit strong environmental plasticity (explained R2 ranging from 0.51 to 0.941). Compared to stand factors and key leaf trait factors, climate factors contribute the most to the spatial variability of soil nutrients in both planted and natural forests. This finding is further validated in the gbm (Gradient Boosting Machine) model—that is, the independent contribution of each climate factor is significantly greater than that of other types of influencing factors (Figure 4).
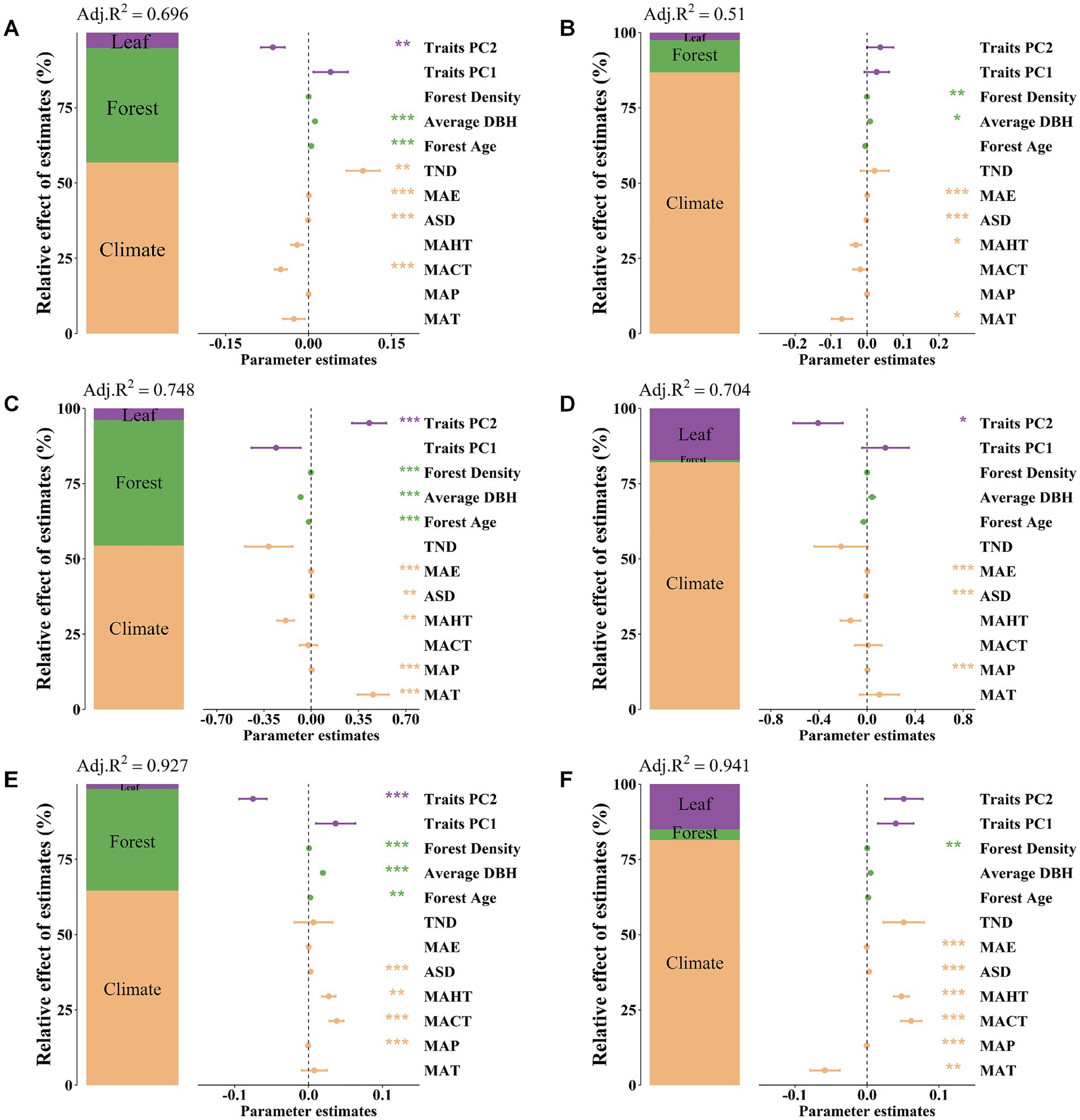
Figure 4. Using variance decomposition analysis to explore the contributions of climate factors, forest factors, and leaf functional traits to soil nitrogen content, available phosphorus content, and soil pH value in planted and natural forests. Soil nitrogen content of natural forests (A), Soil nitrogen content of planted forests (B), Soil available phosphorus content of natural forests (C), Soil available phosphorus content of planted forests (D), Soil pH of natural forests (E), Soil pH of planted forests (F). The average parameter estimates of model predictors (standardized regression coefficients), associated 95% confidence intervals, and the relative importance of each factor, expressed as a percentage of explained variance. The adjusted R2 of the average model and the p-values for each predictor are as follows: *p < 0.05; **p < 0.01; ***p < 0.001.
The results from the random forest show the significance of the average predictive value of each influencing factor within the random forest, further illustrating that climate factors are the key determinants in driving the spatial distribution of soil nutrients in planted and natural forests at macro scales (Figure 5). Heatmap analysis further explains the significant correlations between soil nutrients and various key factors (Figure 6).
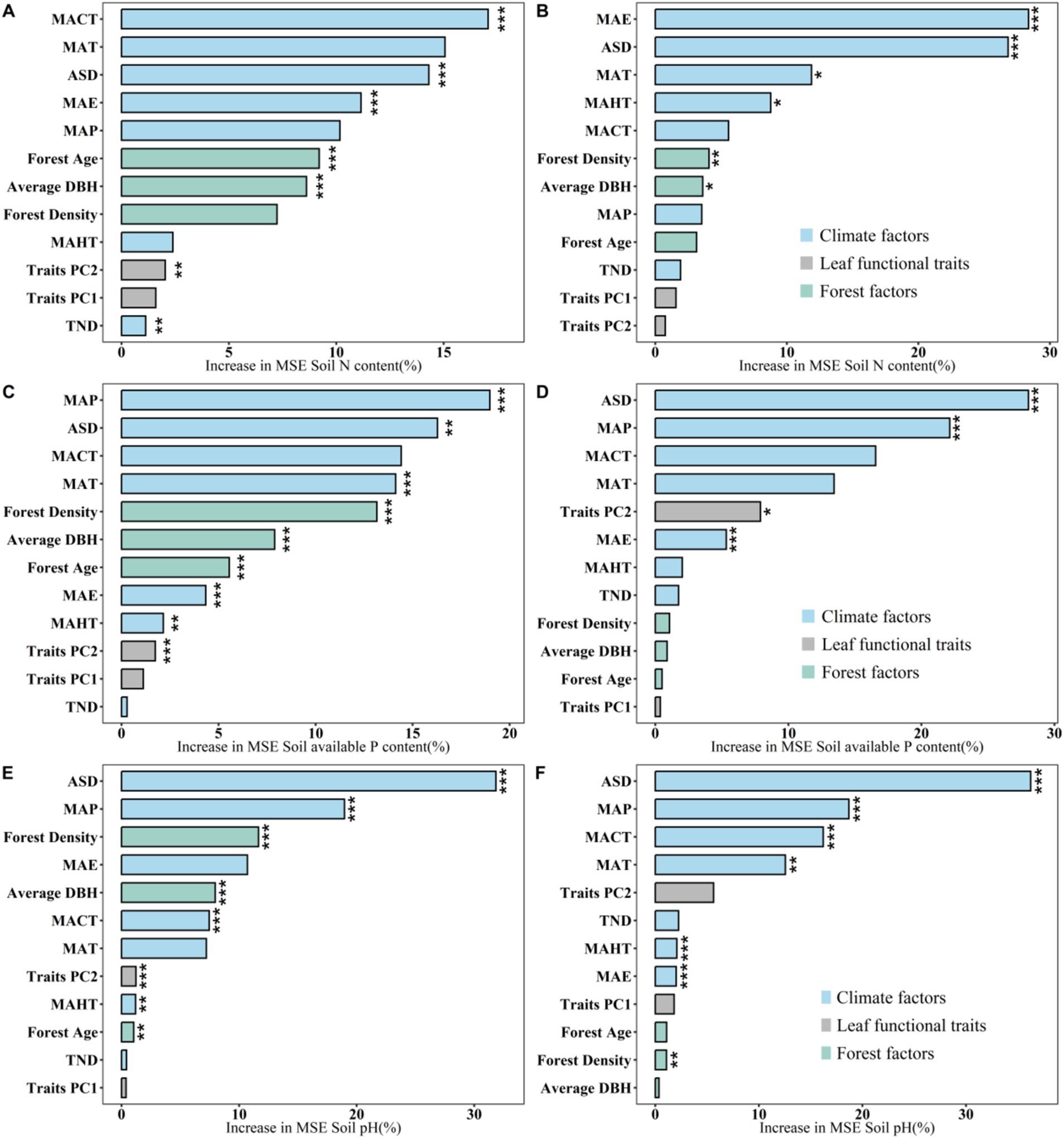
Figure 5. Exploring the independent contributions of various potential influencing factors on soil nutrients in planted and natural forests based on the random forest model. Soil nitrogen content of natural forests (A), Soil nitrogen content of planted forests (B), Soil available phosphorus content of natural forests (C), Soil available phosphorus content of planted forests (D), Soil pH of natural forests (E), Soil pH of planted forests (F). Climate factors include: Mean annual temperature [MAT]; Monthly coldest average temperature [MACT]; Monthly hottest average temperature [MAHT]; Mean annual precipitation [MAP]; Annual sunlight duration [ASD]; Mean annual evaporation [MAE]; Total nitrogen deposition [TND]. Forest factors include: Forest Age; Forest diameter at breast height [Average DBH]; Forest Density. Leaf functional traits include: Leaf functional traits PC1 [Traits PC1]; Leaf functional traits PC2 [Traits PC2]. Asterisks indicate levels of significance (***p < 0.001; **p < 0.01; *p < 0.05).
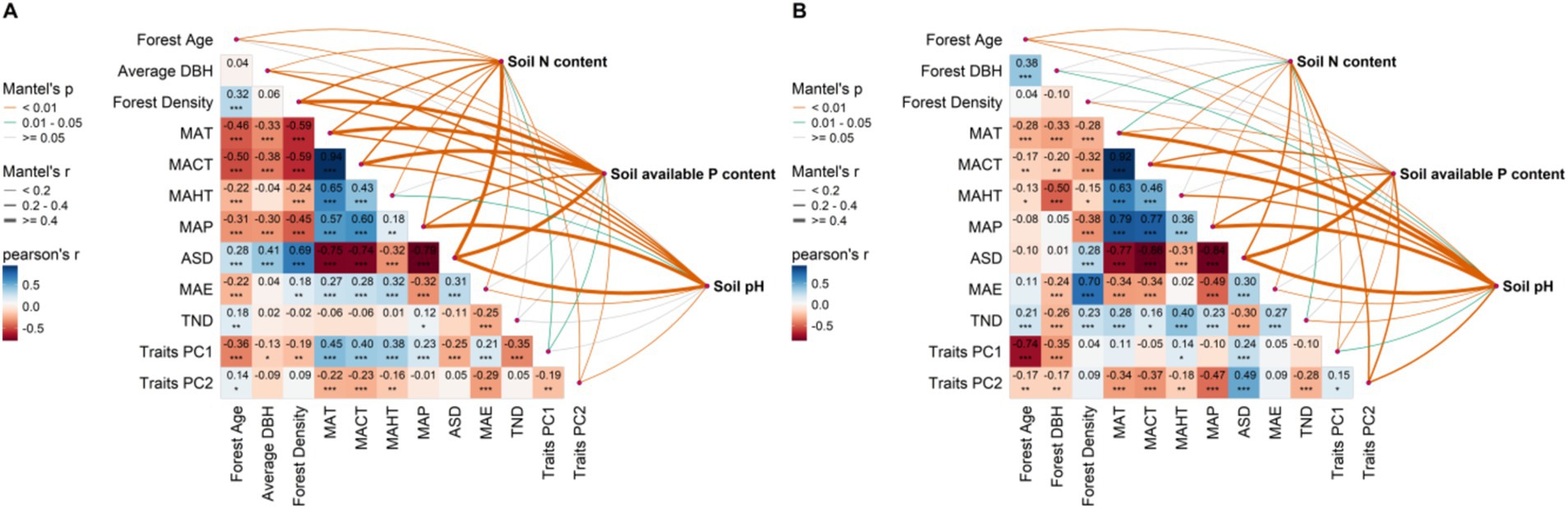
Figure 6. Multivariate correlation analysis of potential influencing factors on soil nitrogen content, soil available phosphorus content, and soil pH in natural and planted forests. (A) Natural forests; (B) Planted forests. Influencing factors include: Forest Age; Forest diameter at breast height [Average DBH]; Forest Density; Mean annual temperature [MAT]; Monthly coldest average temperature [MACT]; Monthly hottest average temperature [MAHT]; Mean annual precipitation [MAP]; Annual sunlight duration [ASD]; Mean annual evaporation [MAE]; Total nitrogen deposition [TND]; Leaf functional traits PC1 [Traits PC1]; Leaf functional traits PC2 [Traits PC2].
The structural equation model indicates that for planted forests, climate factors can not only directly affect soil nutrient factors but can also indirectly influence soil nutrient factors by affecting key leaf traits, with their direct effects being greater than their indirect effects. However, for natural forests, climate factors only have a direct impact on soil nutrients, with the indirect effects being not significant (p > 0.05) (Figure 7).
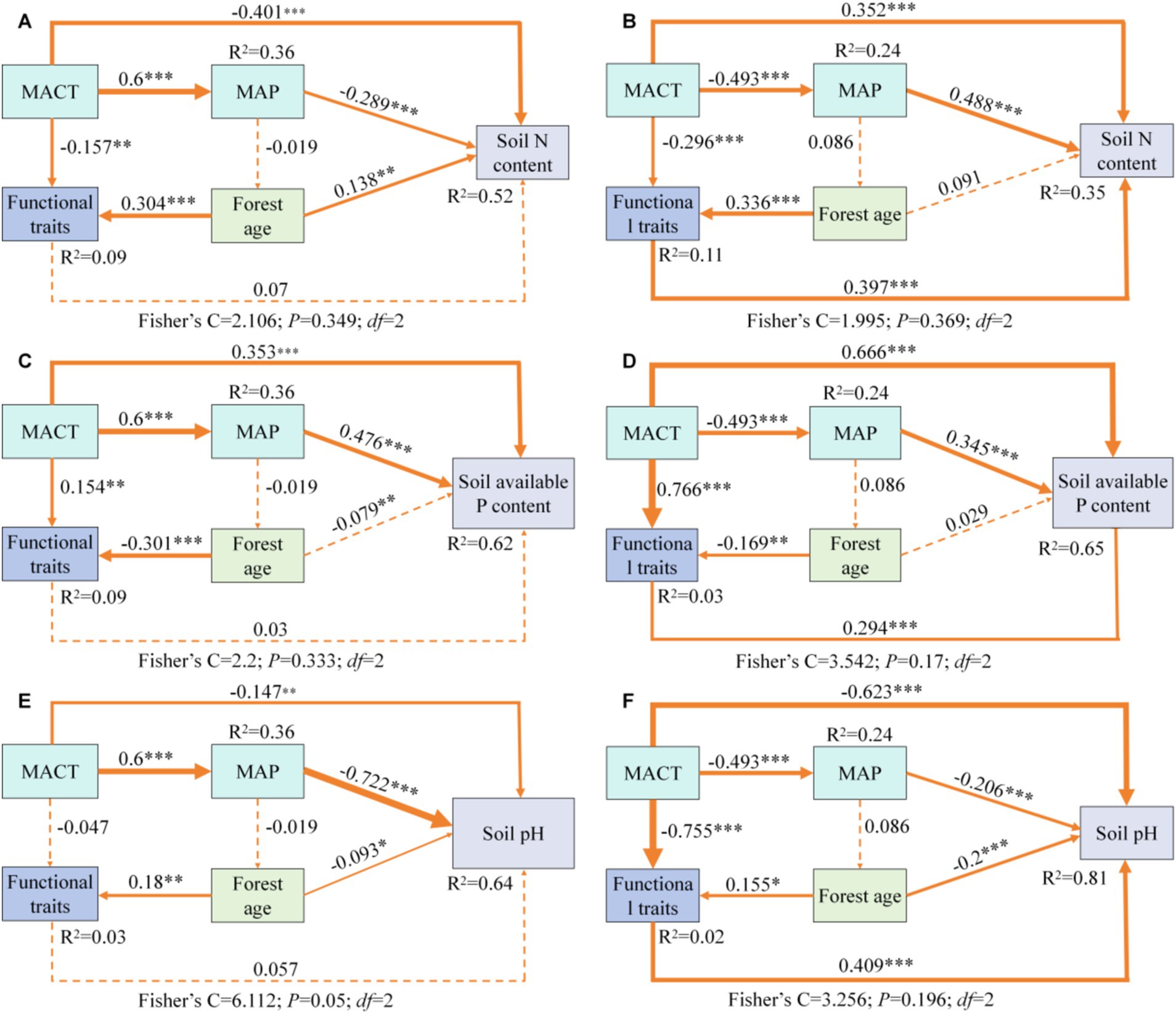
Figure 7. Exploring the impact pathways of climate factors (MACT, MAP), key leaf traits, and stand factors (stand age) on soil nutrients in planted and natural forests based on the structural equation model. Soil nitrogen content of natural forests (A), Soil nitrogen content of planted forests (B), Soil available phosphorus content of natural forests (C), Soil available phosphorus content of planted forests (D), Soil pH of natural forests (E), Soil pH of planted forests (F). The width of the arrows is proportional to the significance level, numbers beside the paths represent standardized SEM coefficients, and stars indicate significant differences (***p < 0.001; **p < 0.01; *p < 0.05). R2 indicates the goodness of fit of the generalized additive model.
4 Discussion
4.1 Differences in soil nutrients between planted and natural forests
We found that the available phosphorus content in the soil of natural forests is significantly higher than that in planted forests. In natural forests, long-term biomass accumulation and decomposition processes may lead to the accumulation of available phosphorus in the soil (Liu et al., 2012). The decomposition of fallen leaves, dead branches, animal remains, and other organic matter can increase the available phosphorus content in the soil (Liu et al., 2021). Additionally, natural forests typically have higher biodiversity, including a variety of plants, microbes, and animals (Stephens and Wagner, 2007). This diversity may promote more efficient available phosphorus cycling and utilization. We also found that the soil pH values in natural forests are significantly lower than those in planted forests. The decomposition of organic matter in natural forests may produce acidic substances, thereby reducing the soil pH value (Jin et al., 2022). These acidic substances include humic and fulvic acids. There was no significant difference in the nitrogen content of the soil between natural and planted forests, mainly because the sources of nitrogen, such as atmospheric deposition (ammonia and nitrogen oxides) and biological nitrogen fixation, are similar in both types of forests (Yang Y. et al., 2021). The nitrogen cycle in the soil is fast and easily influenced by environmental changes. Therefore, the nitrogen content is affected by various factors and does not reflect the differences in forest types as directly as the available phosphorus content does.
4.2 Impacts of climate factors on soil nutrients
Temperature is a key factor influencing soil microbial activity, and microbes play an important role in the soil nutrient cycle (Gao et al., 2015). Higher temperatures typically promote the growth and metabolic activity of soil microbes, accelerating the decomposition of organic matter and releasing more mineral nutrients (such as nitrogen, phosphorus, potassium, etc.). Precipitation directly affects soil moisture content, which in turn influences the solubility and availability of nutrients in the soil (Wang L. et al., 2022; Wang Z. et al.,2022). Moderate precipitation helps dissolve minerals and organic matter in the soil, making it easier for plants to absorb nutrients. We discovered that with the increase in temperature and precipitation, the soil available phosphorus content in both planted and natural forests significantly increased, while pH significantly decreased. The increase in temperature and precipitation not only enhances biological activity and accelerates the decomposition of organic matter, thereby releasing more phosphorus and acidic substances, but also enhances the growth of plant roots and microbial activity, promoting the mineralization of soil available phosphorus, which aids in the more efficient acquisition of phosphorus from the soil (Lu et al., 2015). The increase in precipitation may enhance the leaching of alkaline ions in the soil, thus leading to a decrease in pH value (Wu et al., 2019). In recent years, an increasing number of studies have found that nitrogen deposition significantly affects soil nutrient content, with an increasing trend in soil nitrogen content in both planted and natural forests as nitrogen deposition increases (Tian et al., 2018; Zhu et al., 2015). However, we also made the surprising discovery that the soil phosphorus content in planted forests significantly increases with the increase in nitrogen deposition, while there is no significant change in the soil available phosphorus content in natural forests. Planted forests are usually composed of single tree species, which may have specific adaptability to the absorption and utilization of nitrogen. With increased nitrogen deposition, these tree species may absorb available phosphorus from the soil more effectively through enhanced root activity (Liao et al., 2024). Furthermore, root exudates (such as organic acids) may alter soil pH, thereby increasing the availability of phosphorus (Zhang et al., 2023). Nitrogen deposition also affects the structure and function of the soil microbial community, which plays a key role in the cycling of phosphorus (Ma et al., 2022). In planted forests, the increased nitrogen may promote the activity of certain microbes that can release more available phosphorus (Xia et al., 2020).
4.3 Impacts of stand factors on soil nutrients
Overall, with the increase in stand density and stand age, no significant trend in the change of soil nutrients (N, available P, and pH) was observed in planted forests, whereas in natural forests, soil nitrogen (N) content and pH significantly increased, and soil available phosphorus (AP) content significantly decreased. A higher stand density implies increased competition among trees, which could lead to a reduction in nutrient absorption and cycling efficiency (Farooq et al., 2021). In planted forests, this competition might be mitigated by human management practices, such as thinning, thus resulting in no significant change in nutrients (Forrester, 2013). In natural forests, due to the more complex ecosystem, nutrient cycling may be more efficient, leading to a significant increase in soil N content and pH. Additionally, as stand age increases, soil microbial communities and activity may change (Kang et al., 2018), which are crucial for soil nutrient cycling. In planted forests, due to the monoculture of tree species and possible human intervention, this effect might be suppressed. In natural forests, over time, an increase in microbial diversity might promote the cycling of N and increase in pH (Wang et al., 2023).
4.4 Impacts of key leaf traits on soil nutrients
Compared to climate and stand factors, the impact of key leaf traits on soil nutrients in planted and natural forests is relatively minor. Overall, the key leaf traits in planted forests have a stronger explanatory power for the spatial variability of soil nutrients than in natural forests. Planted forests often undergo more human management, such as regular thinning and fertilization. These management practices may enhance the association between tree leaf traits (such as leaf area, chlorophyll content, photosynthetic efficiency, etc.) and soil nutrients (dos Santos et al., 2006). The growth of natural forests relies more on natural conditions, where interactions are more complex and uncertain. Additionally, in natural forests, different tree species may occupy different ecological niches, each adapting to specific soil nutrient conditions (Gu et al., 2017). This niche differentiation leads to a more diversified relationship between leaf traits and soil nutrients spatially (Sterck et al., 2011). In contrast, due to the lack of such differentiation in planted forests, the leaf traits of tree species are more likely to directly reflect uniform soil nutrient conditions.
4.5 Combined effects of biotic and abiotic on soil nutrients
The research further reveals that on a macro scale, the soil nutrients in both planted and natural forests exhibit strong environmental plasticity, with climate factors contributing more to the spatial variability of soil nutrients in planted and natural forests compared to stand factors and key leaf traits. Climate is a major driver influencing soil nutrient cycling. Temperature and precipitation directly affect soil microbial activity and the rate of organic matter decomposition, which in turn influences nutrient release and cycling (Yuan et al., 2021). Due to significant variations in climate factors on a macro scale, their impact on soil nutrients often surpasses that of local stand or vegetation characteristics. Climate conditions directly affect plant growth patterns, photosynthetic efficiency, and water use efficiency, which in turn affects the demand for and efficiency of soil nutrient utilization by plants (Xu et al., 2023). On a macro scale, variations in these factors may be more significant than differences within a stand. The major contribution of climate factors to the spatial variability of soil nutrients in planted and natural forests on a macro scale is primarily due to the direct impact of climate conditions on soil biochemical processes and plant growth, and these impacts are more significant and widespread on a large scale than the effects of stand structure and plant leaf traits (Gong et al., 2023).
The direct effects of climate factors on shaping the variability of soil nutrients in planted and natural forests are greater than their indirect effects. The direct impact of climate on soil nutrients is usually more rapid and intense. For example, changes in temperature and humidity can significantly alter microbial activity and nutrient cycling rates in a short period (Zhou et al., 2012). In contrast, the impact of climate on leaf traits is a more complex and time-delayed process (Wu et al., 2015). Changes in leaf traits first affect plant physiological processes, and then these changes indirectly affect soil nutrient cycling. While the indirect effects of climate factors on soil nutrients are also important, their impact is generally more complex, time-delayed, and more concentrated on local plant–soil systems (Faucon et al., 2017).
5 Conclusion
Using soil nutrients data from 263 planted forests and 434 natural forests in China, including soil nitrogen content, soil available phosphorus content, and soil pH, this study revealed the differences in soil nutrients between natural and planted forests and their influencing factors. The research results found that there was no significant difference in soil nitrogen content between planted forests and natural forests. However, the soil available phosphorus content of natural forests is significantly higher than that of planted forests, and the soil pH is significantly lower than that of planted forests. Overall, the distribution of soil nutrients in natural and planted forests is the result of the combined effects of climate factors, stand factors, and key leaf functional traits. Among them, climate factors are the main driving factors for the spatial variation of soil nutrients in both planted and natural forests. The impact of stand factors and key leaf functional traits on soil nutrients in planted and natural forests is smaller than that of climatic factors. Climate factors not only directly affect the soil N, available P, and pH of planted and natural forests, but also indirectly affect soil nutrients in planted and natural forests by influencing forest stands and key leaf functional traits, and the direct effect is greater than the indirect effect. Therefore, paying attention to global climate change plays a crucial role in studying the spatial variability of soil nutrients in both planted and natural forests. As the planting area of planted and natural forests continues to expand, further research into the impact of these factors on soil nutrients in natural and planted forests holds significant value in the face of future climate changes. This research is crucial for achieving carbon neutrality, mitigating climate change, improving soil nutrients, and protecting biodiversity.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors without undue reservation.
Author contributions
XD: Funding acquisition, Writing – original draft, Writing – review & editing. XL: Data curation, Investigation, Writing – original draft. JW: Conceptualization, Investigation, Writing – review & editing. JX: Formal analysis, Writing – original draft. JG: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by 2023 Yellow Sea Wetland International Cooperation Key Project (Project No. HHSDKT202303) and Basic Scientific Research Project (XJEDU2023P071).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2024.1525250/full#supplementary-material
Footnotes
References
Banerjee, A. K., Ghosh, S., and Dewanji, A. (2017). Do plants have a choice of traits to be modulated? Evidence from an invasive plant Mikania micrantha Kunth in different urban environments. Am. J. Plant Sci. 8, 835–855. doi: 10.4236/ajps.2017.84057
Bo, H., Wen, C., Song, L., Yue, Y., and Nie, L. (2018). Fine-root responses of Populus tomentosa forests to stand density. Forests 9:562. doi: 10.3390/f9090562
Brockerhoff, E., Jactel, H., Parrotta, J., Quine, C., and Sayer, J. (2008). Plantation forests and biodiversity: oxymoron or opportunity? Biodivers. Conserv. 17, 925–951. doi: 10.1007/s10531-008-9380-x
Chen, X., Taylor, A. R., Reich, P. B., Hisano, M., Chen, H. Y. H., and Chang, S. X. (2023). Tree diversity increases decadal forest soil carbon and nitrogen accrual. Nature 618, 94–101. doi: 10.1038/s41586-023-05941-9
Chen, C., Xiao, W., and Chen, H. Y. H. (2023). Mapping global soil acidification under N deposition. Glob. Chang. Biol. 29, 4652–4661. doi: 10.1111/gcb.16813
Cheng, Y., Wang, J., Chang, S. X., Cai, Z., Müller, C., and Zhang, J. (2019). Nitrogen deposition affects both net and gross soil nitrogen transformations in forest ecosystems: a review. Environ. Pollut. 244, 608–616. doi: 10.1016/j.envpol.2018.10.054
Dibar, D. T., Zhang, K., Yuan, S., Zhang, J., Zhou, Z., and Ye, X. (2020). Ecological stoichiometric characteristics of carbon (C), nitrogen (N) and phosphorus (P) in leaf, root, stem, and soil in four wetland plants communities in Shengjin Lake, China. PLoS One 15:e0230089. doi: 10.1371/journal.pone.0230089
dos Santos, U. M., de Carvalho Gonçalves, J. F., and Feldpausch, T. R. (2006). Growth, leaf nutrient concentration and photosynthetic nutrient use efficiency in tropical tree species planted in degraded areas in Central Amazonia. For. Ecol. Manag. 226, 299–309. doi: 10.1016/j.foreco.2006.01.042
Elser, J. J., Bracken, M. E. S., Cleland, E. E., Gruner, D. S., Harpole, W. S., Hillebrand, H., et al. (2007). Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 10, 1135–1142. doi: 10.1111/j.1461-0248.2007.01113.x
FAO (2020). Global forest resources assessment 2020. FAO. Available at: https://openknowledge.fao.org/handle/20.500.14283/ca8753en (Accessed November 2, 2024).
Farooq, T. H., Shakoor, A., Rashid, M. H. U., Zhang, S., Wu, P., and Yan, W. (2021). Annual growth progression, nutrient transformation, and carbon storage in tissues of Cunninghamia lanceolata monoculture in relation to soil quality indicators influenced by intraspecific competition intensity. J. Soil Sci. Plant Nutr. 21, 3146–3158. doi: 10.1007/s42729-021-00595-w
Faucon, M.-P., Houben, D., and Lambers, H. (2017). Plant functional traits: soil and ecosystem services. Trends Plant Sci. 22, 385–394. doi: 10.1016/j.tplants.2017.01.005
Forrester, D. I. (2013). Growth responses to thinning, pruning and fertiliser application in Eucalyptus plantations: a review of their production ecology and interactions. For. Ecol. Manag. 310, 336–347. doi: 10.1016/j.foreco.2013.08.047
Gao, W., Zhao, W., Yang, H., Yang, H., Chen, G., Luo, Y., et al. (2015). Effects of nitrogen addition on soil inorganic N content and soil N mineralization of a cold-temperate coniferous forest in great Xing’an mountains. Acta Ecol. Sin. 35, 130–136. doi: 10.1016/j.chnaes.2015.07.003
Gong, H., Song, W., Wang, J., Wang, X., Ji, Y., Zhang, X., et al. (2023). Climate factors affect forest biomass allocation by altering soil nutrient availability and leaf traits. J. Integr. Plant Biol. 65, 2292–2303. doi: 10.1111/jipb.13545
Grzyb, A., Wolna-Maruwka, A., and Niewiadomska, A. (2021). The significance of microbial transformation of nitrogen compounds in the light of integrated crop management. Agronomy 11:1415. doi: 10.3390/agronomy11071415
Gu, L., Gong, Z., and Li, W. (2017). Niches and interspecific associations of dominant populations in three changed stages of natural secondary forests on loess plateau, P.R. China. Sci. Rep. 7:6604. doi: 10.1038/s41598-017-06689-9
Gurevitch, J., Koricheva, J., Nakagawa, S., and Stewart, G. (2018). Meta-analysis and the science of research synthesis. Nature 555, 175–182. doi: 10.1038/nature25753
He, M., Dijkstra, F. A., Zhang, K., Li, X., Tan, H., Gao, Y., et al. (2014). Leaf nitrogen and phosphorus of temperate desert plants in response to climate and soil nutrient availability. Sci. Rep. 4:6932. doi: 10.1038/srep06932
Hobbie, S. E. (2015). Plant species effects on nutrient cycling: revisiting litter feedbacks. Trends Ecol. Evol. 30, 357–363. doi: 10.1016/j.tree.2015.03.015
Homeier, J., Seeler, T., Pierick, K., and Leuschner, C. (2021). Leaf trait variation in species-rich tropical Andean forests. Sci. Rep. 11:9993. doi: 10.1038/s41598-021-89190-8
Hong, S., Gan, P., and Chen, A. (2019). Environmental controls on soil pH in planted forest and its response to nitrogen deposition. Environ. Res. 172, 159–165. doi: 10.1016/j.envres.2019.02.020
Hou, X., Duan, C., Tang, C., and Fu, D. (2009). Nutrient relocation, hydrological functions, and soil chemistry in plantations as compared to natural forests in Central Yunnan, China. Ecol. Res. 25, 139–148. doi: 10.1007/s11284-009-0645-y
Hua, F., Bruijnzeel, L. A., Meli, P., Martin, P. A., Zhang, J., Nakagawa, S., et al. (2022). The biodiversity and ecosystem service contributions and trade-offs of forest restoration approaches. Science 376, 839–844. doi: 10.1126/science.abl4649
Jakmunee, J., and Junsomboon, J. (2009). Determination of available phosphorus in soils by using a new extraction procedure and a flow injection amperometric system. Talanta 79, 1076–1080. doi: 10.1016/j.talanta.2009.01.028
Janssens, I. A., Dieleman, W., Luyssaert, S., Subke, J.-A., Reichstein, M., Ceulemans, R., et al. (2010). Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 3, 315–322. doi: 10.1038/ngeo844
Jin, Z., Luo, D., Yu, Y., Yang, S., Zhang, J., and Cao, G. (2022). Soil pH changes in a small catchment on the Chinese loess plateau after long-term vegetation rehabilitation. Ecol. Eng. 175:106503. doi: 10.1016/j.ecoleng.2021.106503
Kang, H., Gao, H., Yu, W., Yi, Y., Wang, Y., and Ning, M. (2018). Changes in soil microbial community structure and function after afforestation depend on species and age: case study in a subtropical alluvial island. Sci. Total Environ. 625, 1423–1432. doi: 10.1016/j.scitotenv.2017.12.180
Kong, W., Wei, X., Wu, Y., Shao, M., Zhang, Q., Sadowsky, M. J., et al. (2022). Afforestation can lower microbial diversity and functionality in deep soil layers in a semiarid region. Glob. Chang. Biol. 28, 6086–6101. doi: 10.1111/gcb.16334
Li, H., Ren, R., Zhang, H., Zhang, G., He, Q., Han, Z., et al. (2023). Factors regulating interaction among inorganic nitrogen and phosphorus species, plant uptake, and relevant cycling genes in a weakly alkaline soil treated with biochar and inorganic fertilizer. Sci. Total Environ. 905:167280. doi: 10.1016/j.scitotenv.2023.167280
Liao, L., Wang, J., Dijkstra, F. A., Lei, S., Zhang, L., Wang, X., et al. (2024). Nitrogen enrichment stimulates rhizosphere multi-element cycling genes via mediating plant biomass and root exudates. Soil Biol. Biochem. 190:109306. doi: 10.1016/j.soilbio.2023.109306
Liu, L., Gundersen, P., Zhang, T., and Mo, J. (2012). Effects of phosphorus addition on soil microbial biomass and community composition in three forest types in tropical China. Soil Biol. Biochem. 44, 31–38. doi: 10.1016/j.soilbio.2011.08.017
Liu, J., Lu, H., Wu, L., Kerr, P. G., and Wu, Y. (2021). Interactions between periphytic biofilms and dissolved organic matter at soil-water interface and the consequent effects on soil phosphorus fraction changes. Sci. Total Environ. 801:149708. doi: 10.1016/j.scitotenv.2021.149708
Lu, H., Li, Z., Fu, S., Méndez, A., Gascó, G., and Paz-Ferreiro, J. (2015). Effect of biochar in cadmium availability and soil biological activity in an Anthrosol following acid rain deposition and aging. Water Air Soil Pollut. 226:164. doi: 10.1007/s11270-015-2401-y
Lugo, A. E., Cuevas, E., and Sanchez, M. J. (1990). Nutrients and mass in litter and top soil of ten tropical tree plantations. Plant Soil 125, 263–280. doi: 10.1007/BF00010665
Ma, J., Kang, F., Cheng, X., and Han, H. (2018). Response of soil organic carbon and nitrogen to nitrogen deposition in a Larix principis-rupprechtii plantation. Sci. Rep. 8:8638. doi: 10.1038/s41598-018-26966-5
Ma, X., Wang, T., Shi, Z., Chiariello, N. R., Docherty, K., Field, C. B., et al. (2022). Long-term nitrogen deposition enhances microbial capacities in soil carbon stabilization but reduces network complexity. Microbiome 10:112. doi: 10.1186/s40168-022-01309-9
Naorem, A., Jayaraman, S., Dang, Y., Dalal, R., Sinha, N., Ch, S., et al. (2023). Soil constraints in an arid environment-challenges, prospects, and implications. Agronomy 13:220. doi: 10.3390/agronomy13010220
Ni, Y., Jian, Z., Lixiong, Z., Liu, J., Lei, L., Zhu, J., et al. (2022). Climate, soil nutrients, and stand characteristics jointly determine large-scale patterns of biomass growth rates and allocation in Pinus massoniana plantations. For. Ecol. Manag. 504:119839. doi: 10.1016/j.foreco.2021.119839
Prescott, C. E., and Vesterdal, L. (2021). Decomposition and transformations along the continuum from litter to soil organic matter in forest soils. For. Ecol. Manag. 498:119522. doi: 10.1016/j.foreco.2021.119522
Rashmi, I., Shirale, A., Kartikha, K. S., Shinogi, K. C., Meena, B. P., and Kala, S. (2017). “Leaching of plant nutrients from agricultural lands” in Essential plant nutrients: uptake, use efficiency, and management. eds. M. Naeem, A. A. Ansari, and S. S. Gill (Cham: Springer International Publishing), 465–489.
Reich, P. B., and Oleksyn, J. (2004). Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 101, 11001–11006. doi: 10.1073/pnas.0403588101
Ren, Y., Gao, G. L., Ding, G. D., Zhang, Y., Guo, M. S., Cao, H. Y., et al. (2019). Stoichiometric characteristics of nitrogen and phosphorus in leaf-litter-soil system of Pinus sylvestris var. mongolica plantations. Ying Yong Sheng Tai Xue Bao 30, 743–750. doi: 10.13287/j.1001-9332.201903.040
Song, L., Tian, P., Zhang, J., and Jin, G. (2017). Effects of three years of simulated nitrogen deposition on soil nitrogen dynamics and greenhouse gas emissions in a Korean pine plantation of Northeast China. Sci. Total Environ. 609, 1303–1311. doi: 10.1016/j.scitotenv.2017.08.017
St Clair, S. B., and Lynch, J. P. (2010). The opening of Pandora’s box: climate change impacts on soil fertility and crop nutrition in developing countries. Plant Soil 335, 101–115. doi: 10.1007/s11104-010-0328-z
Stephens, S. S., and Wagner, M. R. (2007). Forest plantations and biodiversity: a fresh perspective. J. For. 105, 307–313. doi: 10.1093/jof/105.6.307
Sterck, F., Markesteijn, L., Schieving, F., and Poorter, L. (2011). Functional traits determine trade-offs and niches in a tropical forest community. Proc. Natl. Acad. Sci. 108, 20627–20632. doi: 10.1073/pnas.1106950108
Tian, D., Du, E., Jiang, L., Ma, S., Zeng, W., Zou, A., et al. (2018). Responses of forest ecosystems to increasing N deposition in China: a critical review. Environ. Pollut. 243, 75–86. doi: 10.1016/j.envpol.2018.08.010
Verma, P., and Sagar, R. (2020). Effect of nitrogen (N) deposition on soil-N processes: a holistic approach. Sci. Rep. 10:10470. doi: 10.1038/s41598-020-67368-w
Wang, L., Deng, D., Feng, Q., Xu, Z., Pan, H., and Li, H. (2022). Changes in litter input exert divergent effects on the soil microbial community and function in stands of different densities. Sci. Total Environ. 845:157297. doi: 10.1016/j.scitotenv.2022.157297
Wang, Z., Huang, W., and Pang, F. (2022). Selenium in soil-plant-microbe: a review. Bull. Environ. Contam. Toxicol. 108, 167–181. doi: 10.1007/s00128-021-03386-2
Wang, L., Zhang, H., Xu, C., Yuan, J., Xu, X., Wang, J., et al. (2023). Long-term nitrogen fertilization and sweetpotato cultivation in the wheat-sweetpotato rotation system decrease alkaline phosphomonoesterase activity by regulating soil phoD-harboring bacteria communities. Sci. Total Environ. 900:165916. doi: 10.1016/j.scitotenv.2023.165916
Wu, J., Su, Y., Chen, X., Liu, L., Sun, C., Zhang, H., et al. (2019). Redistribution characteristics of atmospheric precipitation in different spatial levels of Guangzhou urban typical forests in southern China. Atmos. Pollut. Res. 10, 1404–1411. doi: 10.1016/j.apr.2019.03.012
Wu, D., Zhao, X., Liang, S., Zhou, T., Huang, K., Tang, B., et al. (2015). Time-lag effects of global vegetation responses to climate change. Glob. Chang. Biol. 21, 3520–3531. doi: 10.1111/gcb.12945
Xia, Z., Yang, J., Sang, C., Wang, X., Sun, L., Jiang, P., et al. (2020). Phosphorus reduces negative effects of nitrogen addition on soil microbial communities and functions. Microorganisms 8:1828. doi: 10.3390/microorganisms8111828
Xu, L., Gao, G., Wang, X., and Fu, B. (2023). Distinguishing the effects of climate change and vegetation greening on soil moisture variability along aridity gradient in the drylands of northern China. Agric. For. Meteorol. 343:109786. doi: 10.1016/j.agrformet.2023.109786
Yang, X., Baskin, C. C., Baskin, J. M., Pakeman, R. J., Huang, Z., Gao, R., et al. (2021). Global patterns of potential future plant diversity hidden in soil seed banks. Nat. Commun. 12:7023. doi: 10.1038/s41467-021-27379-1
Yang, Y., Liu, L., Zhang, F., Zhang, X., Xu, W., Liu, X., et al. (2021). Soil nitrous oxide emissions by atmospheric nitrogen deposition over global agricultural systems. Environ. Sci. Technol. 55, 4420–4429. doi: 10.1021/acs.est.0c08004
Yang, L., Shen, F., Zhang, L., Cai, Y., Yi, F., and Zhou, C. (2021). Quantifying influences of natural and anthropogenic factors on vegetation changes using structural equation modeling: a case study in Jiangsu Province, China. J. Clean. Prod. 280:124330. doi: 10.1016/j.jclepro.2020.124330
Yuan, M. M., Guo, X., Wu, L., Zhang, Y., Xiao, N., Ning, D., et al. (2021). Climate warming enhances microbial network complexity and stability. Nat. Clim. Chang. 11, 343–348. doi: 10.1038/s41558-021-00989-9
Zhang, F., Hou, Y., Zed, R., Mauchline, T. H., Shen, J., Zhang, F., et al. (2023). Root exudation of organic acid anions and recruitment of beneficial actinobacteria facilitate phosphorus uptake by maize in compacted silt loam soil. Soil Biol. Biochem. 184:109074. doi: 10.1016/j.soilbio.2023.109074
Zhao, S., Guo, Y., Sheng, Q., and Shyr, Y. (2014). Advanced heat map and clustering analysis using heatmap3. Biomed. Res. Int. 2014:986048. doi: 10.1155/2014/986048
Zhou, J., Xue, K., Xie, J., Deng, Y., Wu, L., Cheng, X., et al. (2012). Microbial mediation of carbon-cycle feedbacks to climate warming. Nat. Clim. Chang. 2, 106–110. doi: 10.1038/nclimate1331
Keywords: natural forest, planted forest, soil nutrients, climate change, stand age
Citation: Du X, Li X, Wang J, Xu J and Gao J (2025) Climate factors dominate the spatial variation of forest soil nutrients: a meta analysis. Front. For. Glob. Change. 7:1525250. doi: 10.3389/ffgc.2024.1525250
Edited by:
Xi Zhang, Louisiana State University Agricultural Center, United StatesReviewed by:
Li Ji, Helmholtz Association of German Research Centres (HZ), GermanyJunfu Dong, Shandong University, China
Copyright © 2025 Du, Li, Wang, Xu and Gao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaoxuan Du, aGFvYmluZ3NodWFpa2VAaG90bWFpbC5jb20=; Jie Gao, amllZ2FvNzJAZ21haWwuY29t
†These authors have contributed equally to this work