 B. Karamdoost Marian1†
B. Karamdoost Marian1† A. Alijanpour
A. Alijanpour S. Sasanifar
S. Sasanifar P. Álvarez-Álvarez
P. Álvarez-Álvarez- 1Department of Forestry, Faculty of Natural Resources, Urmia University, Urmia, Iran
- 2Department of Organisms and Systems Biology, Polytechnic School of Mieres, University of Oviedo, Mieres, Asturias, Spain
Introduction: This study examines the effects of the single-tree selective harvesting method on ecosystem services in a mixed temperate broadleaf forest in Iran. Key indicators such as carbon sequestration, tree species diversity, soil nutrient cycling, and stand volume are analyzed, emphasizing their significance for sustainable forest management.
Methods: The research was conducted in four districts, each comprising two parcels: one managed using the single-tree selective harvesting method and the other serving as a control. Data on ecological and biodiversity parameters were collected, and statistical analyses, including two-way ANOVA and Principal Component Analysis (PCA), were performed to assess the impact of management practices.
Results: The findings reveal that the single-tree selective harvesting method significantly influences regulating and supporting ecosystem services. Carbon storage varied with elevation, affecting both soil and forest floor litter. Tree species diversity increased, with more species present and a reduction in dominance by certain species. However, this method also resulted in reduced stand volume in the managed areas. Elevation significantly impacted diversity indices, litter carbon storage, available potassium, and stand volume. Additionally, the interaction between management and elevation was significant for soil carbon storage, richness, diversity, total nitrogen, available potassium, and stand volume.
Discussion: The single-tree selective harvesting method appears to be a viable forest management strategy for preserving ecosystem services in mixed temperate broadleaf forests, maintaining ecosystem health without significant negative effects on soil. However, careful consideration of site-specific ecological conditions and trade-offs between provisioning and regulating services is crucial. These findings have important implications for sustainable forest management in Iran and similar forest ecosystems globally.
1 Introduction
Ecosystem services are the conditions created by natural ecosystems that enhance the richness and diversity of plant and animal species, ultimately benefiting human societies (Daily, 1998). The Millennium Ecosystem Assessment (2005) categorizes ecosystem services into four groups: regulating services, supporting services, provisioning services, and cultural services (Wallace, 2007). Regulating services, such as carbon sequestration, depend on the natural processes of an ecosystem (Platon et al., 2015). Supporting services, including soil formation, originate from natural ecosystems and are essential for accessing other ecosystem services (Platon et al., 2015). Evaluating the management of natural ecosystems using ecosystem service indicators is now a critical and practical aspect of sustainable natural resource management (Grammatikopoulou and Vačkářová, 2021).
Carbon dioxide separation from the atmosphere is necessary for reducing global warming (Tong et al., 2020; Panja, 2021). Increasing forest vegetation (which plays a vital role in reducing atmospheric carbon dioxide concentrations) (Trumper, 2009; Lal, 2004) tends to be the only simple and inexpensive way to control carbon in the atmosphere.
Plant diversity is a widely used component of biodiversity in vegetation studies and an important indicator of ecological sustainability of ecosystems in environmental assessments (Pecl et al., 2017; Pollastrini et al., 2014). Forest biodiversity also affects forest growth and biodiversity mechanisms and processes, which can lead to increased biomass productivity (Zhang et al., 2010; Zou et al., 2024). This composition of tree species in forest stands plays an important role in determining the quality and quantity of litter and organic matter inputs into the forest soil, which in turn affects the nutrient cycle (Zhang et al., 2018; Carmona-Yáñez et al., 2023).
Stand volume is a basis characteristic, especially in the forest production sector (Sasanifar et al., 2019). While this characteristic is primarily considered in industrial forests for wood production, its correlation with density, tree dimensions, biomass, and habitat fertility appear to be a reliable indicator of structure in managed forest stands (Lorenz, 2010; Vafaei et al., 2017).
The success or failure of forest management methods can be determined by evaluating the rate of forest change following the implementation of various management programs (Wheeler et al., 2016; Oettel and Lapin, 2021). Therefore, in this study, the effectiveness of single-tree selective harvest method on some studied indicators of ecosystem services from temperate broadleaf forests has been investigated. On the other hand, the results will show us that in 20 years, to what extent this type of management has affected the studied characteristics of the forest. Therefore, the goals of this research were advanced by collecting data from the studied forests.
Hyrcanian forests are renowned as one of the world’s oldest surviving forests, with a history spanning millions of years. These ancient forests, belonging to the Third Age of geology, harbor unique species that cannot be found anywhere else due to the limited impact of past glaciations (Sagheb Talebi et al., 2014). During the Pleistocene (Ice Age) period, all but a few broadleaf forests were lost. The Hyrcanian forests with a temperate climate were protected and survived from freezing because they were located between the mountains and the Caspian Sea. On the other hand, these forests are located in latitude belts less than 40 degrees north and at a maximum height of 5,000 m. Accordingly, the cold climate has had a more moderate effect than other forests of high latitudes. As a result, the frosts had the least effect on them. Therefore, the Hyrcanian forests can be considered the “mother” of young forests in Europe (Sagheb Talebi et al., 2014). These forests are composed of mixed deciduous trees that play an effective role in the formation of soil horizons, humus, and soil fertility through the return of leaves to the soil surface in the autumn (Sohrabi et al., 2022).
Due to being close to nature, the single-tree selective harvest method, has been used as a silviculture method for the sustainable management of forests around the world (Sagheb Talebi et al., 2014).
Numerous studies have been conducted on different management methods in Hyrcanian forests. This study specifically focuses on managing these forests using the single-tree selective harvest method. In recent years, this method has gained popularity in the Hyrcanian region of Iran, aiming to preserve the natural characteristics of the forests and maintain their asynchronous form.
The objectives of this study are, (1) to investigate the effects of single-tree selective harvest method on ecosystem services, including carbon storage, tree species diversity, the soil nutrient cycle, and stand volume in a mixed temperate broadleaf forest in Iran, and (2) Evaluating the consequences of the single-tree selective harvest method for Iran’s forests and investigating its effectiveness in similar forest ecosystems.
2 Materials and methods
2.1 Study area
Shafarood Forest Company was established in 1973 with the purpose of meeting Iran’s cellulosic industry’s needs. It initially covered 400,000 hectares of forests in Gilan province, which is part of the Hyrcanian temperate broadleaf forests. Currently, the company manages approximately 150 thousand hectares of forests, spread across 72 districts.
In the past, the management of these forests followed the shelterwood silviculture method (from 1989 to 1998), with regenerative cutting and the first light cutting being implemented over a 10-year period. But because this cutting was done only once in light form and about 30 years ago, it did not cause any significant change in the natural structure of the forests. However, from 1996 to 2016, the management approach shifted to the single-tree selective harvest method.
2.2 Data collection
In this study, four districts were chosen from four different elevational classes within the Shafarood forests: less than 600 (0–599), 600–1,000 (600–999), 1,000–1,500 (1,000–1,499), and more than 1,500 (1,500 to the final limit of forest growth in the study area) meters above sea level (m a.s.l.). Within each district, two parcels were selected: one managed with the single-tree selective harvest method, and one serving as a control parcel. The environmental and ecological conditions, such as aspect, average slope percentage, elevational class, and tree species type, were kept similar in both parcels to ensure comparability.
The control parcels are areas where no exploitation or forest management has taken place. These parcels have not undergone any human intervention and have naturally progressed through succession.
To establish sample plots, a 100 by 200-meter inventory grid was utilized, based on previous research conducted by Khanalizadeh et al. (2020). The grid was designed using ArcGIS and overlaid onto the parcels using Google Earth. GPS coordinates for the centers of the sample plots were then recorded within the designated parcels of each district. During the field implementation of the sample plots, the centers of each parcel were determined using the GPS coordinates obtained in the previous stage.
At each sample plot center, an original plot with a circular shape and an area of 10 acres was established. A total of 30 sample plots for each district were collected (15 samples from the managed parcels and 15 samples from the control parcels). Sample collection in the study districts and parcels is presented in Table 1.
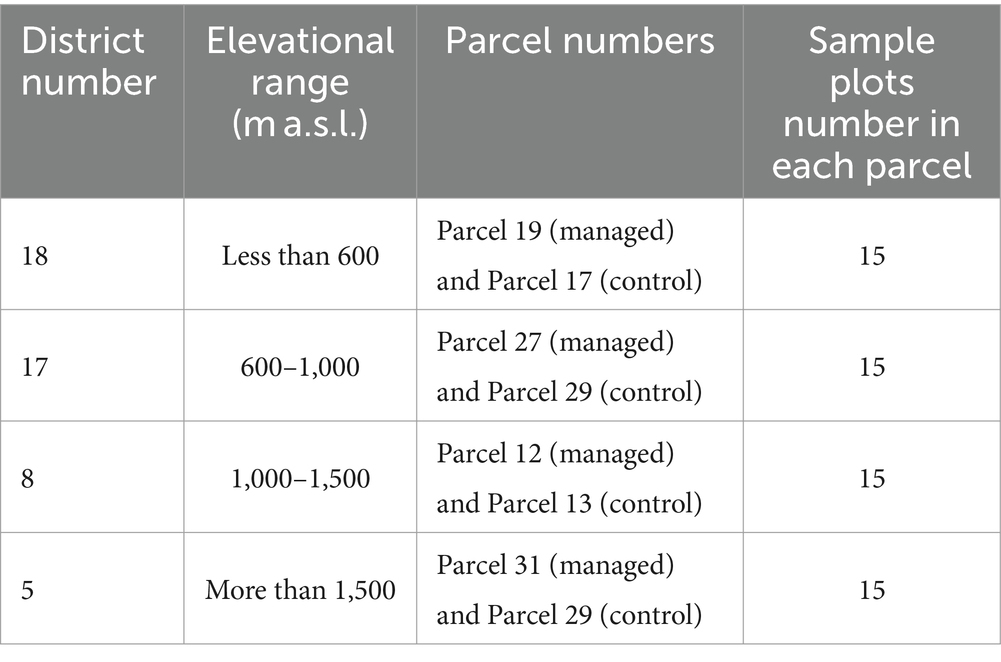
Table 1. Measured plots in each district and parcel.
In each plot, the tree species and diameter at breast height (DBH) were measured using a caliper, with DBH classes recorded in centimeters. Furthermore, the total height of five selected trees within the sample plot was measured. These included the four closest trees in the four main directions from the plot center, as well as the thickest tree within the plot.
For soil sampling, five plots were randomly chosen from all the plots within each parcel. Soil samples were collected at a depth of 0–30 cm, following the methodology described by Barnes et al. (1997). One sample was taken from the center of each plot, while four additional samples were collected from the corners. These individual soil samples were then mixed together to create a composite sample for each selected plot. The composite samples were transferred to the laboratory for further analysis.
Similarly, within the selected plots for soil sampling, intact soil samples were extracted using a cylinder at a depth of 0–30 cm. These samples were used to determine the soil bulk density.
Within the plots designated for soil sampling, a representative portion of the litter layer was collected. The litter was carefully collected from an untouched area within each plot, following the dimensions of 50 cm by 50 cm, as described in the study by Berenguer et al. (2018). All the data of this study were collected in the summer of 2021.
2.3 Studied ecosystem services
In this study, we focused on analyzing two categories of ecosystem services: regulating services, specifically carbon storage, and supporting services, including the soil nutrient cycle, woody species biodiversity, and woody stem yield. We examined these services for both the managed and control parcels, as shown in Table 2. The indicators of ecosystem services were selected based on the classification outlined in the Millennium Ecosystem Assessment (2005).
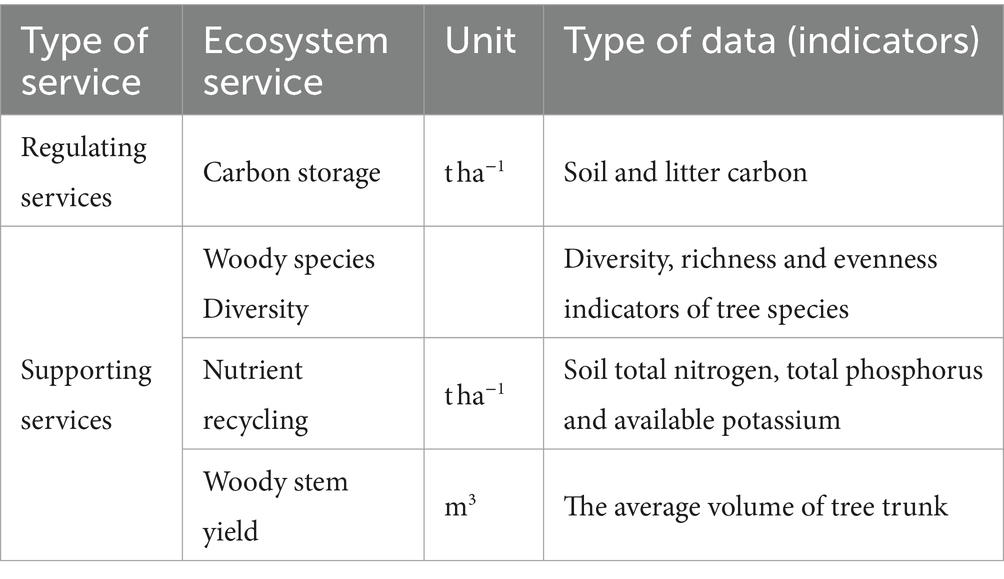
Table 2. Ecosystem service indicators and type of data.
2.3.1 Regulating services
2.3.1.1 Soil carbon storage
In the laboratory, we measured the percentage of soil organic carbon using the Walkley and Black method (Nelson and Sommers, 1996). Subsequently, we utilized Equation 1 to calculate the amount of organic carbon stored in the soil, expressed in tons per hectare.
Where, OC represents organic carbon, Bd is the bulk density of soil, and e shows the depth of sampling soil in centimeters.
2.3.1.2 Litter carbon storage
The organic carbon content of the litter samples was determined using the combustion oven method, as described by Nilsson et al. (1999). To perform this analysis, the dried litter samples were first weighed. Subsequently, the samples were placed in an electric oven and subjected to temperatures ranging from 400 to 500°C for a duration of 4 h. After the incineration process, the samples were re-weighed to determine the weight of ash. The litter’s carbon content for each area was then calculated by considering the ratio of organic carbon to organic matter (54%) and using the weight of ash and the initial weight of the samples, as outlined by MacDicken (1997).
2.3.2 Supporting service indices
2.3.2.1 Woody species diversity
Richness, diversity, and evenness indices for the woody species in the plots were calculated (with Past 4.8).
Whittaker richness presents the number of species in a plant community and is obtained by counting the number of plant species per unit (Humphries et al., 1995). The value is calculated based on Equation 2.
where N is the total number of species in the reagent location.
Another indicator that is widely used in studies is the Shannon (diversity) index. The Shannon function is the most common measure of species diversity (Shannon, 1949). Equation 3 is used to calculate the Shannon-Wiener index.
where H′ shows the Shannon-Wiener index, and Pi is the relative abundance of the i number of species in the sample.
The Pielou species (evenness index) was introduced in 1969 (Pielou, 1979) and varied from 0 to 1. To calculate this index, all species of a population must be identified. Equation 4 is suggested to measure this index.
Where p_i is the ratio of species i in the population, and S is the total number of species in the sample.
2.3.2.2 Nutrient recycling
We conducted laboratory measurements of three indices: total nitrogen, using the Kjeldahl method (Bremner and Mulvaney, 1982), available phosphorus, using the Olsen method (Olsen, 1954), and available potassium, using the saturation extract method (Mehlich, 1978).
2.3.2.3 Woody stem yield
In the context of Shafaroud forests, we utilized tree-volume tariff tables to accurately assess and redefine the volume of tree trunks. This was achieved by employing recorded Diameter at Breast Height (DBH) and height values from the designated plot.
2.4 Statistical analyses
2.4.1 Comparing managed and control areas in terms of ecosystem service indices
We assessed the normality of the data distribution for each ecosystem service index using the Kolmogorov–Smirnov test. To investigate the main effects of management treatments (managed and control) and elevational ranges (less than 600, 600–1,000, 1,000–1,500, and more than 1,500 m a.s.l.), as well as their interactions, we employed a two-way ANOVA assuming normal distribution. For comparing the mean indices at the management level, we used an independent t-test, and for comparing the means among the height levels, a Duncan test was conducted (with SPSS 24).
Principal Component Analysis, or PCA, helps us reduce the dimensionality of the data while preserving its essential characteristics. Principal Component Analysis is a statistical method that transforms high-dimensional data into a lower-dimensional form while preserving the most important information. It accomplishes this by identifying new axes, called principal components, along which the data varies the most. These components are orthogonal to each other, meaning they are uncorrelated, making them a powerful tool for dimensionality reduction. PCA is most commonly firstly used when many of the variables are highly correlated with each other and it is desirable to reduce their number to an independent set. Secondly it is perfumed to distinguish the sensible variable varying among the observation. Finally, it is utilized for separation and fragmentation of various phenomena and treatments differing in view point of their measured variables (Hasan and Abdulazeez, 2021).
In this study, to identify the most significant index affected by the single-tree selective harvest method in both regulating and supporting services, we performed a principal component analysis (PCA) (with CANOVO 5). This allowed us to determine the key indicator within each service category.
3 Results
Based on the data presented in Figure 1, both the managed and control areas exhibit a J-shaped tree distribution curve. In the managed area, there is a higher number of trees per hectare in the middle diameter at breast height (DBH) classes compared to the control area. However, this trend is reversed in the larger DBH classes. Figure 1 provides a graphical representation of the number of trees per hectare in 5 cm DBH classes for both the managed and control areas.
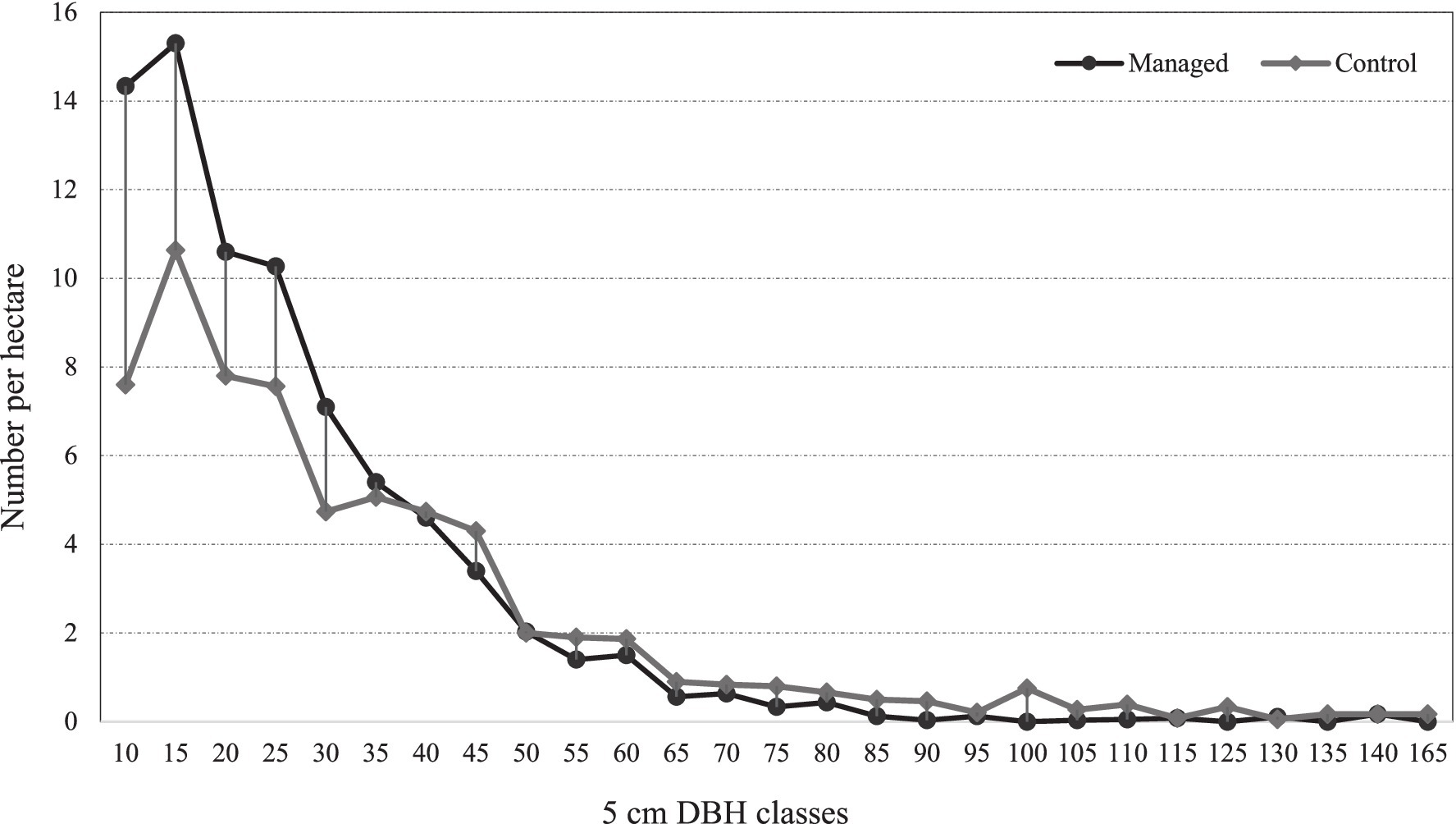
Figure 1. Number of trees per hectare in 5 cm DBH classes in the managed and control area.
3.1 Regulating services
3.1.1 Carbon storage
In terms of carbon storage, soil, and litter carbon storage were calculated for both regions. The two-way ANOVA results of soil and litter carbon storage indices are presented in Table 3.

Table 3. Variance analysis of changes in carbon sequestration under management and elevational range factors.
The interaction between management and elevation factors has a significant impact on soil carbon storage and the elevation factor plays a more influential role on litter carbon storage. It seems that forest management had a greater effect on soil carbon than litter carbon storage. Table 4 provides a comparison of the average soil carbon storage indices resulting from the interaction of management and elevation factors.
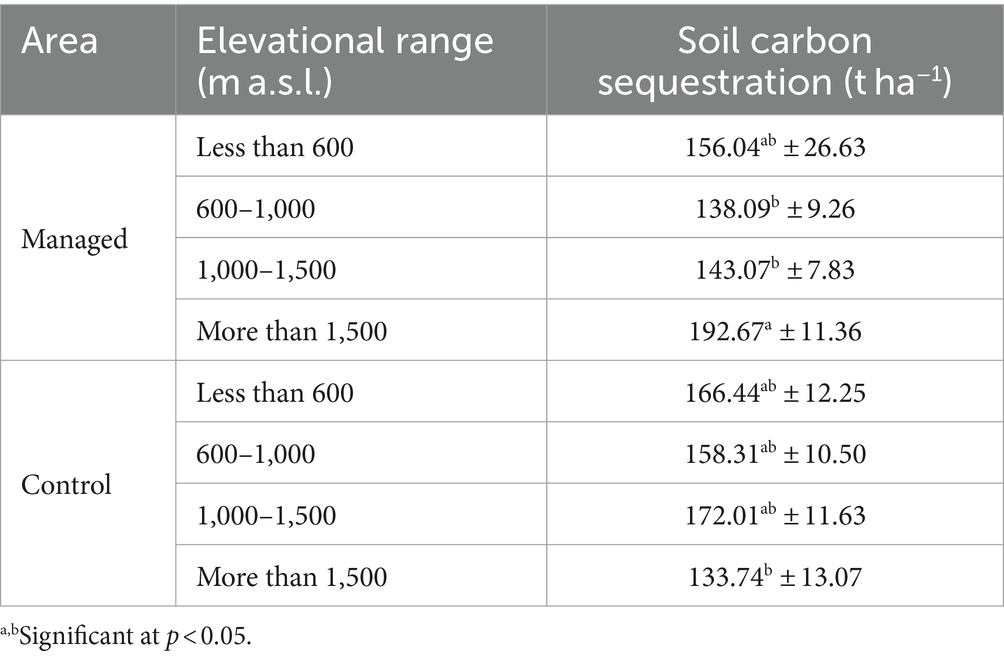
Table 4. Comparison of mean soil carbon related to the interaction effects of management and elevation factors using Duncan test (mean ± SE).
According to the data presented in Table 4, the trend of changes in soil carbon storage is not the same with changes in altitude above sea level in both regions. Additionally, Table 5 provides the average index of litter carbon storage associated with various elevation effects.

Table 5. Comparison of the average litter carbon sequestration related to the main effects of elevation factors, by independent t-test (mean ± SE).
As highlighted in Table 5, our results reveal a large increase in litter carbon storage at an altitude of 1,000–1,500 m a.s.l., which may be caused by the effect of climate or combination of tree species at this altitude.
3.2 Supporting services
3.2.1 Woody species diversity
The woody species in both managed and controlled areas are presented in Table 6. The forest stands studied in both parts are mixed broad-leaved stands. In the higher elevation, the stands are dominated by the Beech-Hornbeam mixed stands. At the lower elevation, the mixed stands of Carpinus-Alnus, are the dominant type. The results of two-way ANOVA for the richness, diversity, and evenness of woody species in managed and control areas are presented in Table 7.
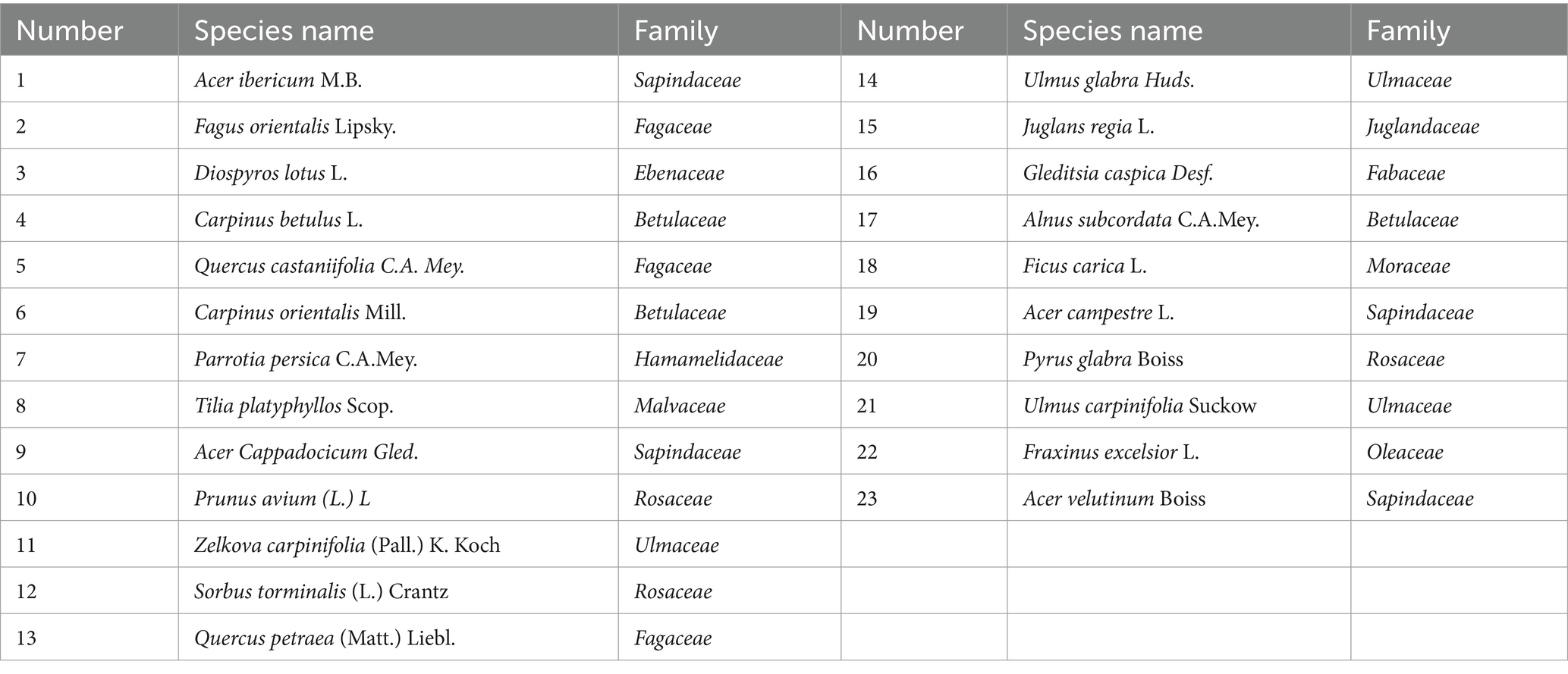
Table 6. Woody species registered in study area.
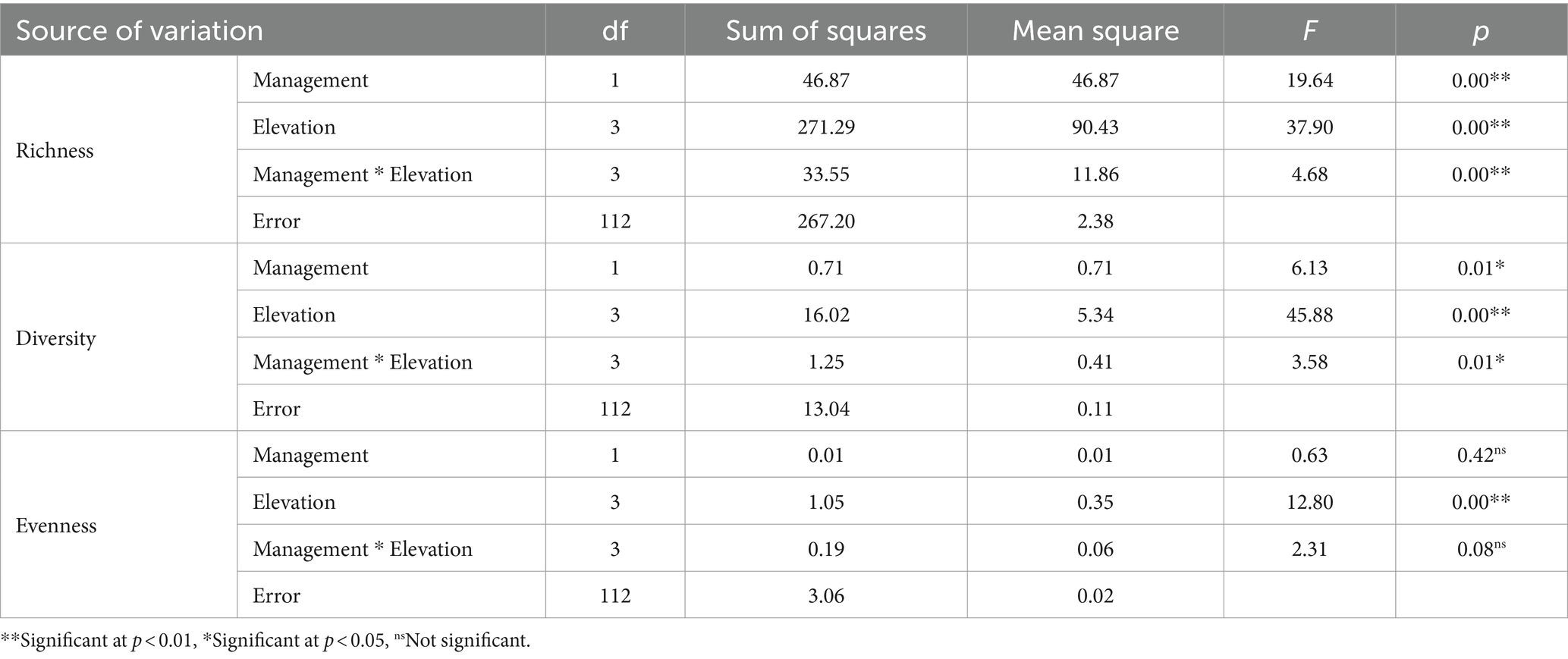
Table 7. Variance analysis of changes in woody species diversity indicators under management and elevational range factors.
Table 7 reveals that management exerted a predominant influence on richness and diversity metrics, whereas elevation exhibited substantial effects on richness, diversity, and evenness. Furthermore, the interaction between management and elevation demonstrated a considerable impact on richness and diversity indices. Figure 2 visually represents the average fluctuations in diversity indices of woody species attributable to the effects of management.
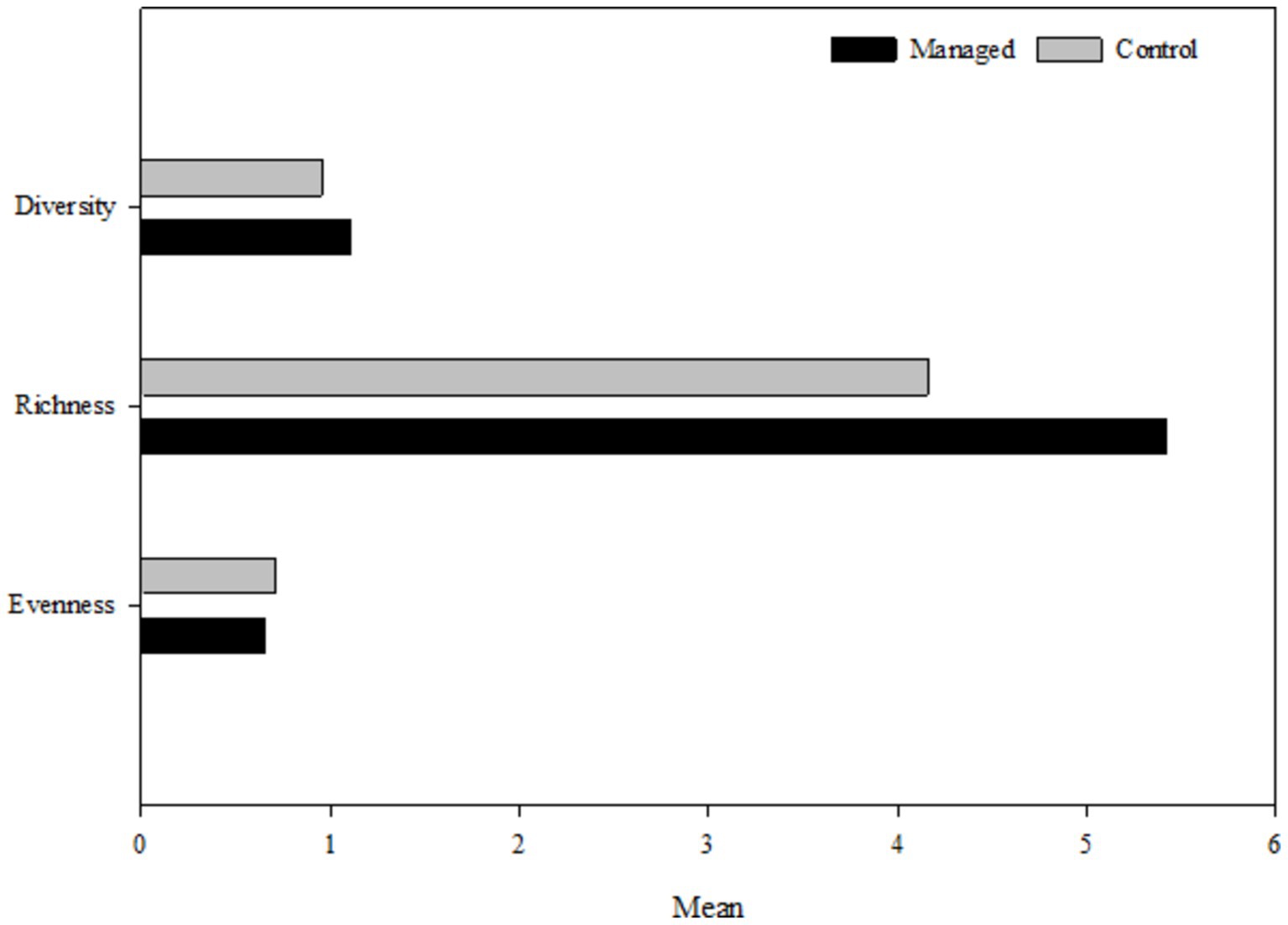
Figure 2. Diversity and richness indices for woody species in two managed and control area.
Figure 2 provides evidence that there are more species with higher abundance in the managed area than in the control area. Furthermore, the same conditions can be observed at an altitude of less than 600 m a.s.l. (Figure 3). The results of the interactive effects of the two studied factors are presented in Table 8.
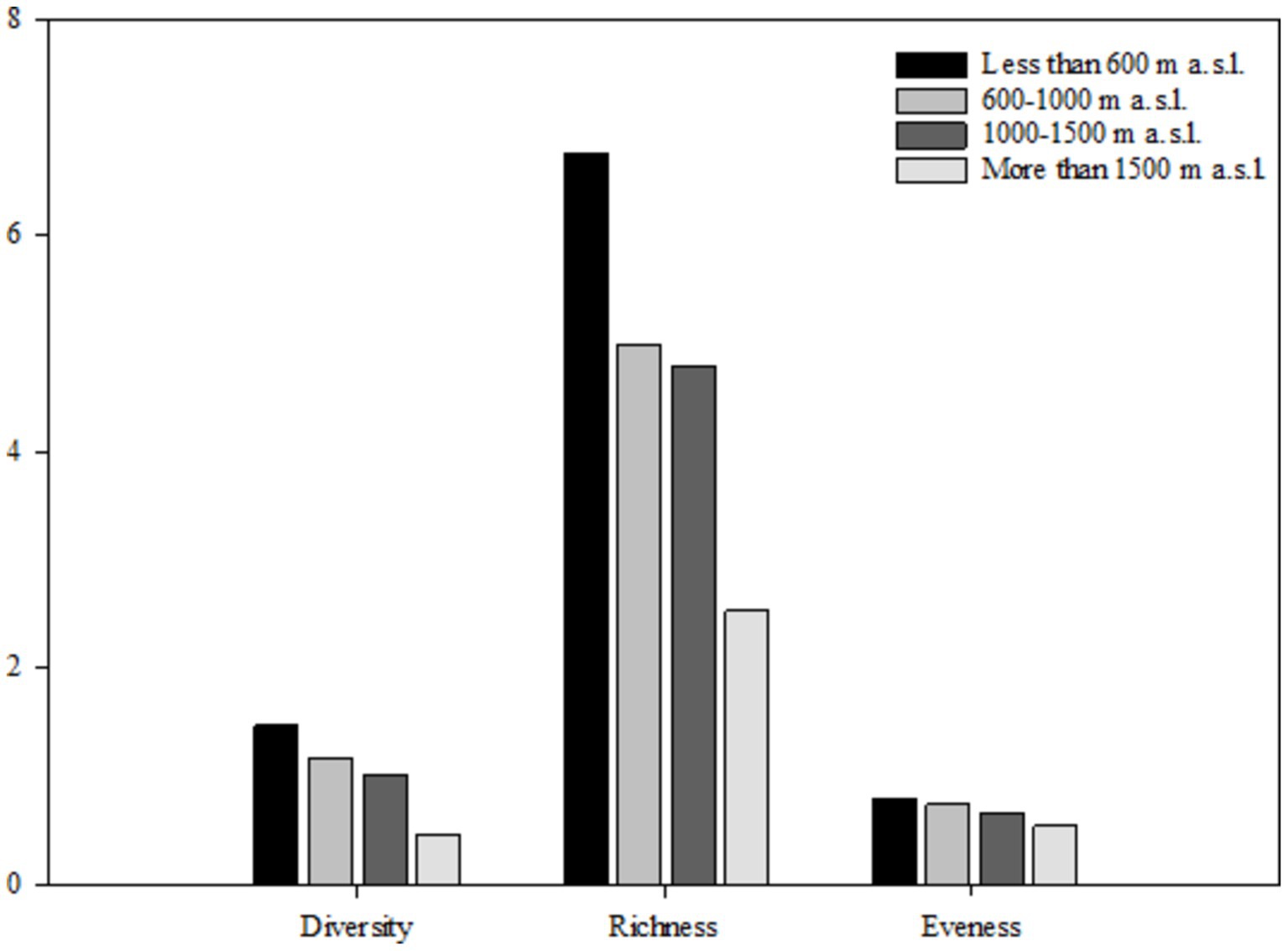
Figure 3. Comparison of the in woody species diversity indicators related to the main effects of elevational range, using Duncan test (a,bSignificant at p < 0.05).
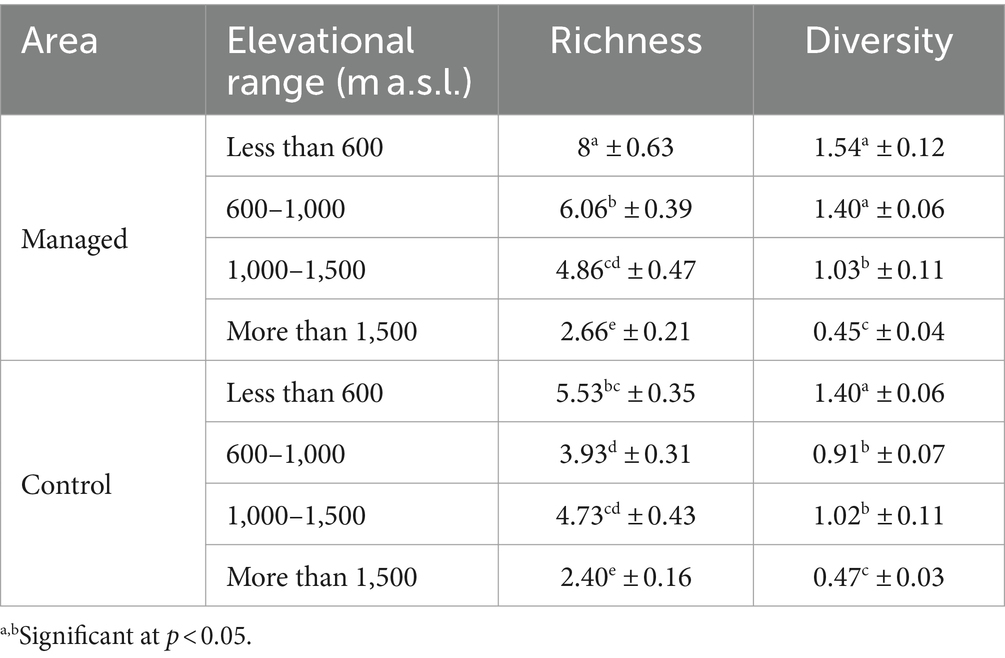
Table 8. Comparison of woody species diversity related to the interaction effects of management and elevational range using Duncan test (mean ± SE).
As Table 8 clarifies, the increasing trend of the studied indices with decreasing elevation factor is regular in the managed area, while there is minor irregularity in the control area.
3.2.2 Nutrient recycling
Table 9 displays the two-way ANOVA results for total Nitrogen, available phosphorus and potassium in the managed and control areas.
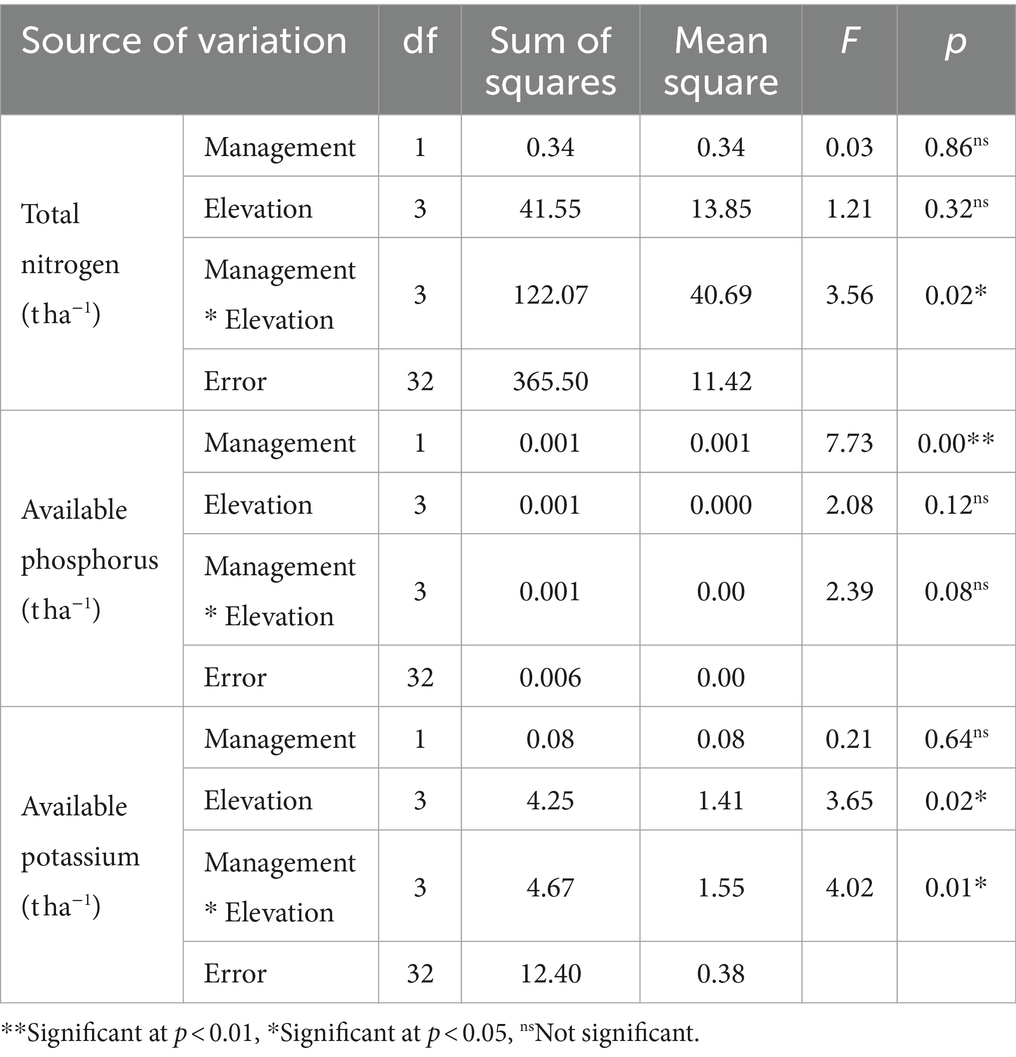
Table 9. Analysis of variance of total nitrogen, available phosphorus and potassium indicators for management and differences in elevation.
Based on the results reported in Table 9, it can be inferred that the interaction between management and elevation exerts a notable influence on the total nitrogen index. It seems that management has only affected available phosphorus index. Moreover, the elevation factor and the interaction between the investigated factors significantly affect the available potassium. Figure 4 graphically presents the average fluctuations in the available phosphorus index resulting from the effects of the management factor. Most likely, the lack of tree harvesting in the control area has led to an increase in available phosphorus reserves in the soil of this area. In Figure 5, the variations in the available potassium index are depicted, highlighting the influence of the elevation factor.
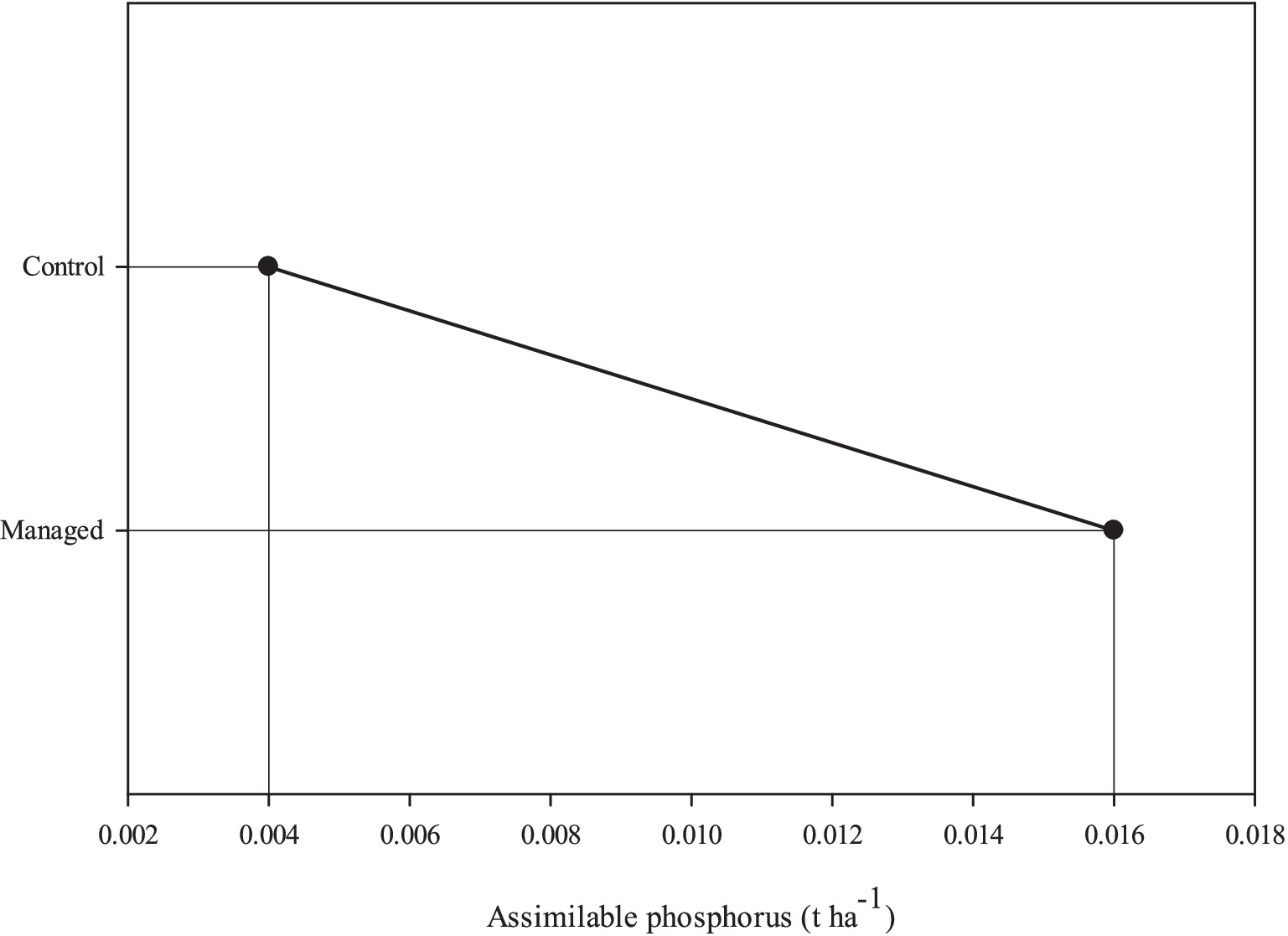
Figure 4. Assimilable phosphorus indices in two managed and control area.
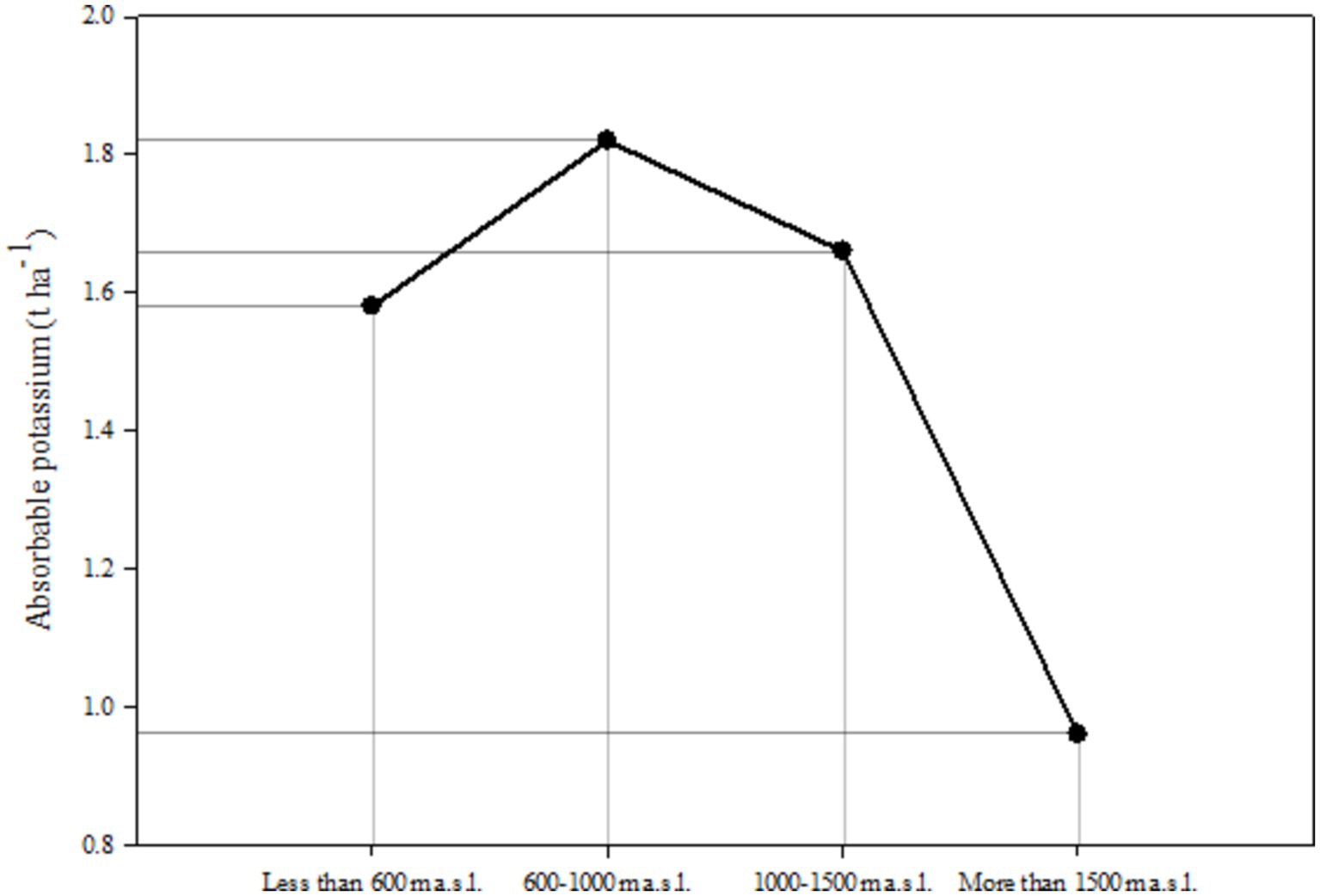
Figure 5. Assimilable potassium indices in elevational range.
According to Figure 5, the process of changes in available phosphorus in relation to changes in height above sea level is not a regular process. The results of the interactive effects of the two studied factors on total Nitrogen and available phosphorus are presented in Table 10.
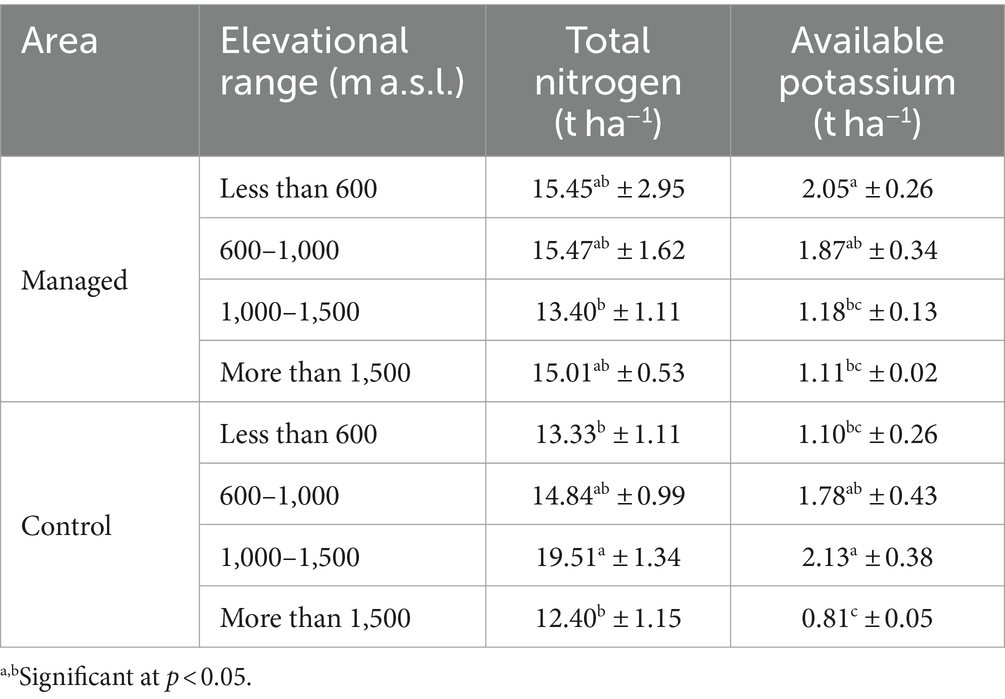
Table 10. Comparison of mean total nitrogen and available potassium indices at different elevations in managed and control areas, using Duncan (mean ± SE).
It can be seen that the trend of changes in total nitrogen and available potassium in both regions is decreasing in relation to the increase in altitude above sea level (Table 10).
3.2.3 Woody stem yield
The two-way ANOVA results on the mean stand volume of trees are presented in Table 11.
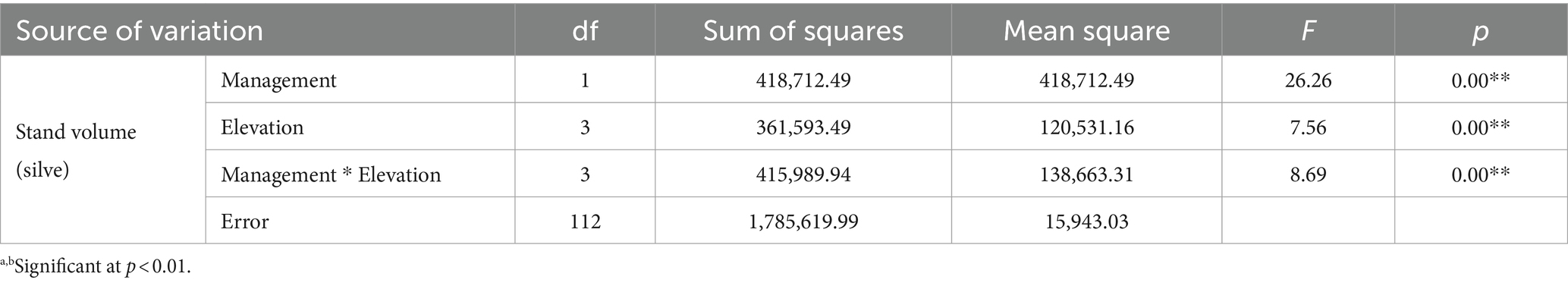
Table 11. Analysis of variance of stand volume for management and differences in elevation.
According to Table 11, it can be seen that stand volume has been affected by all three factors of management, height above sea level and mutual effects of both. In Figure 6, we show the mean differences in the stand volume index due to the effects of the management factor. It is understandable that the average stand volume in the control area is higher than the managed area due to the lack of wood harvesting.
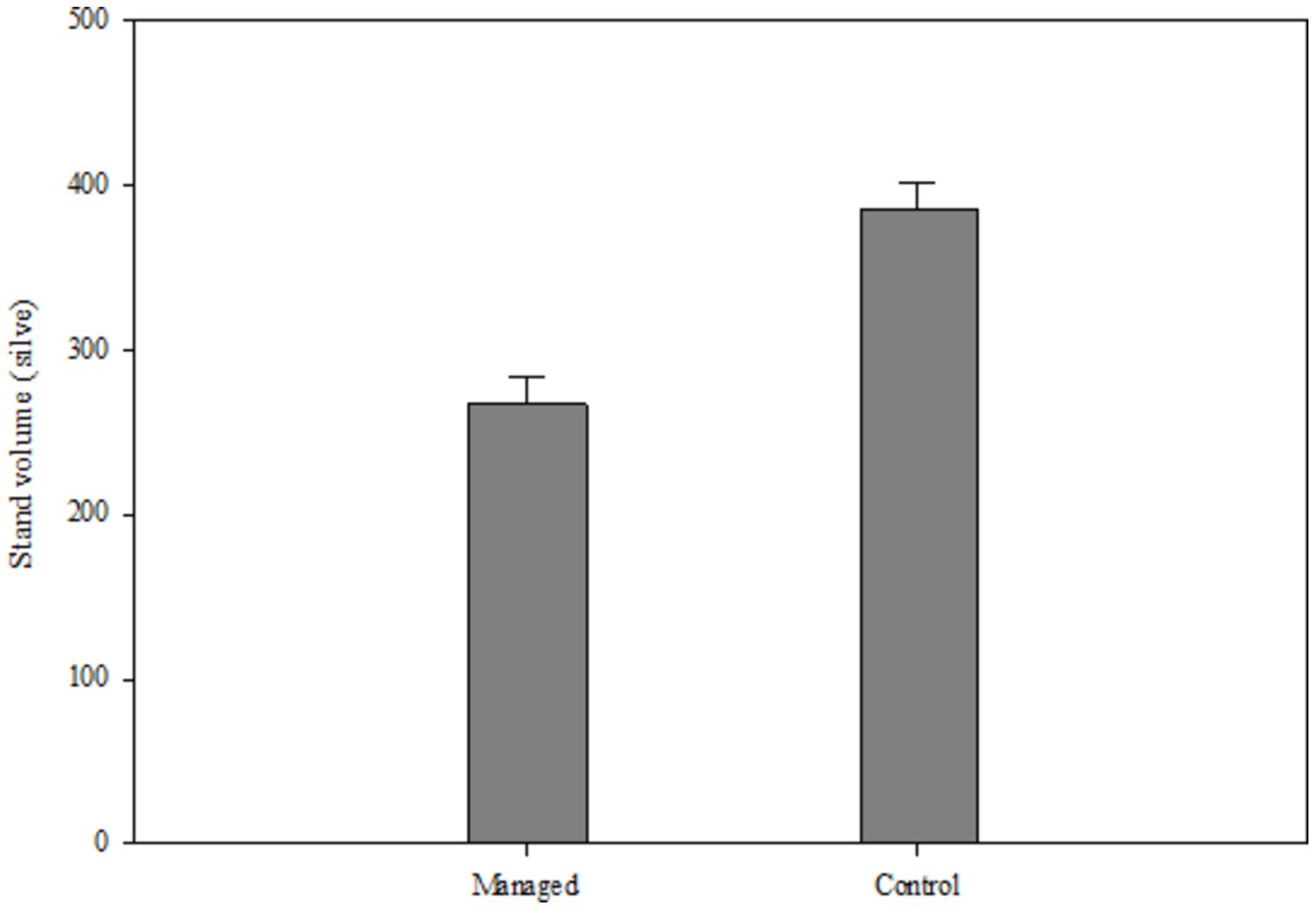
Figure 6. Stand volume indices in two managed and control area.
Figure 7 demonstrate the mean differences in the stand volume index due to the effects of the elevation factor. At higher altitudes, for reasons such as difficult access, difficult transportation, and harsher weather, less wood is harvested, which shows that the average stand volume is the highest at an altitude of more than 1,500 m a.s.l. The results of the interactive effects of the two studied factors on the stand volume index are presented in Table 12.
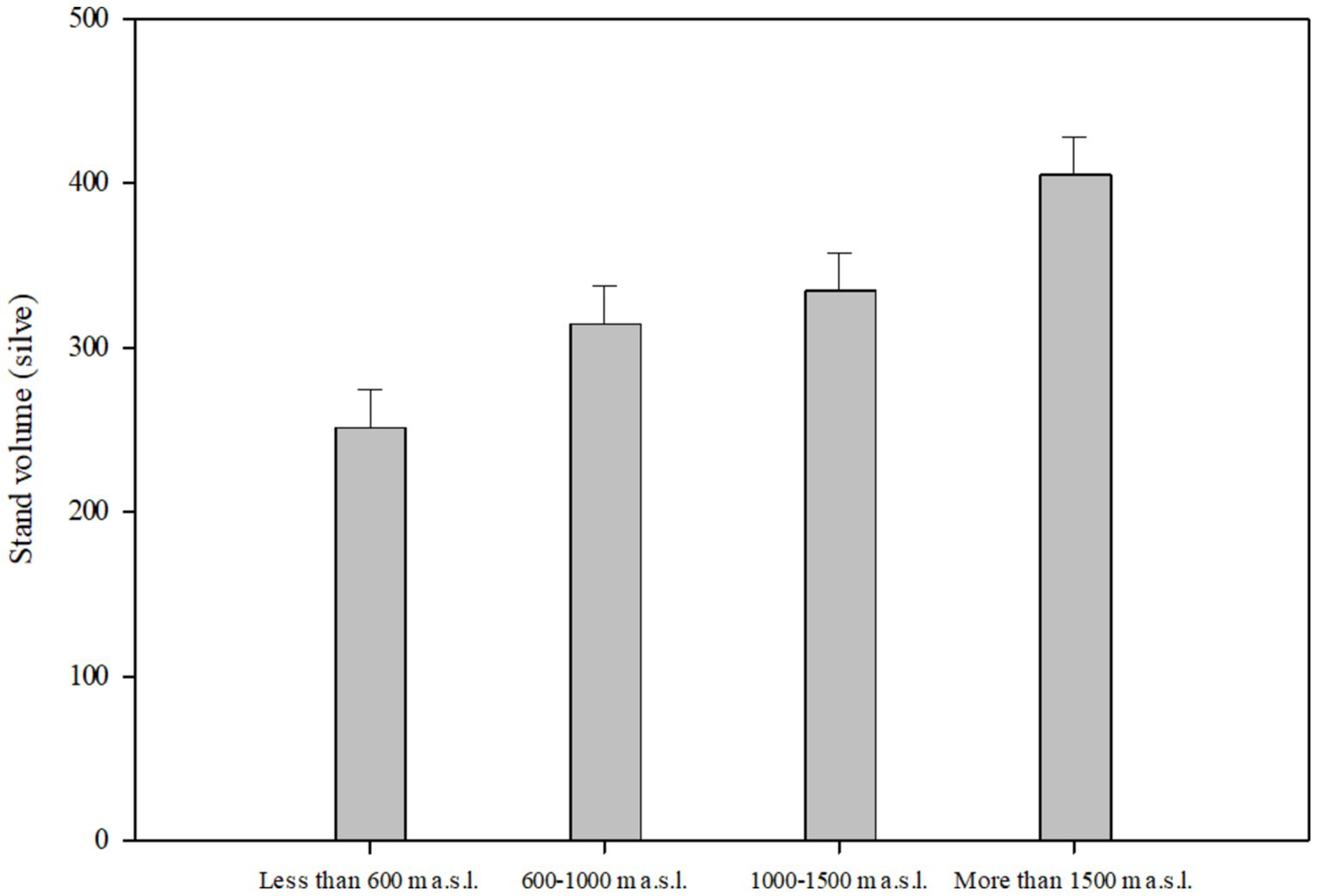
Figure 7. Stand volume indices in elevational range.
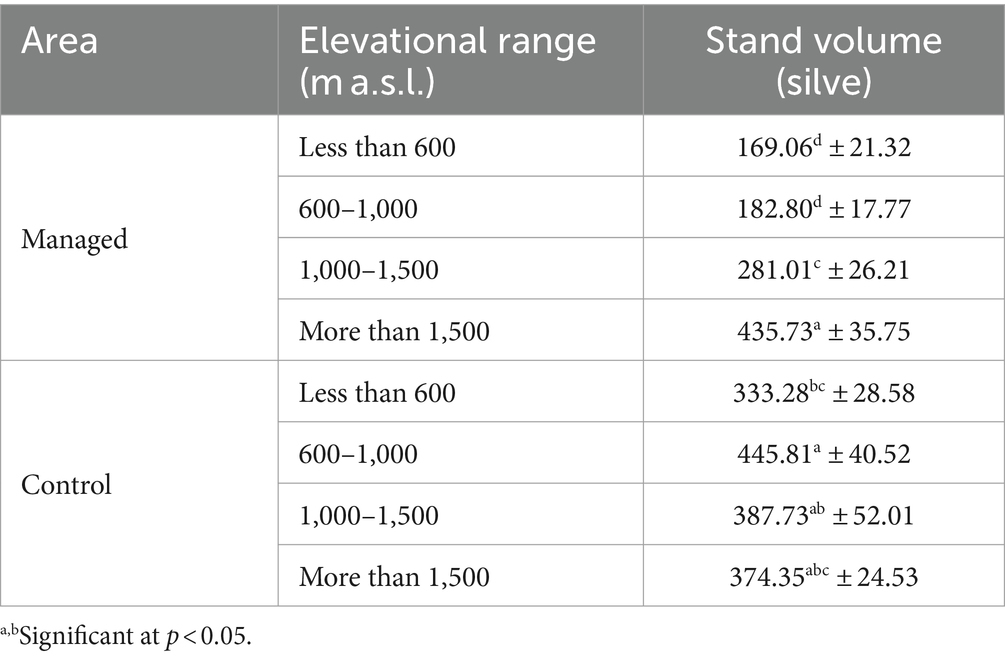
Table 12. Comparison of mean stand volume indice at different elevations in managed and control areas, using Duncan (mean ± SE).
The mean stand volume in both area increases based on the elevation, and this rising trend is considerable in the managed areas.
3.3 The most significantly modified indices
Principal component analysis (PCA) of the indicators examined reveals that the first three components account for 89.54% of the variance in the data, with the first and second components explaining 69.5% of the variance. Among the components, parameters with a factor load greater than 0.6, such as soil carbon storage, available potassium, total nitrogen, richness, diversity, and stand tree volume, were identified as the most influential variables affecting site differentiation.
The main component analysis is presented in Figure 8 to determine the most importantly modified indices in ecosystem services classes.
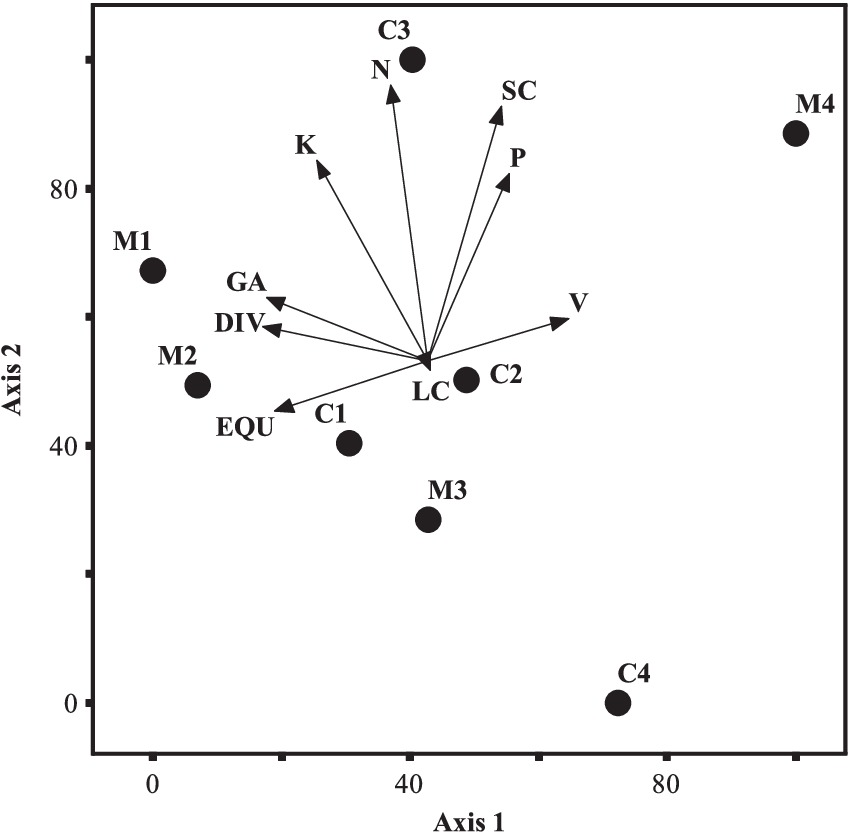
Figure 8. Ranking of indicators of regulating and supporting services in the space of the first and second components GA (richness), DIV (diversity), EQU (evenness), K (available potassium), N (total nitrogen), SC (soil carbon storage), P (available phosphorus), LC (litter carbon storage), and VO (stand volume); C1–C4 (control area in 4 sea level), M1–M4 (managed area in 4 sea level).
Figure 8 depicts various indicators including GA (richness), DIV (diversity), EQU (evenness), K (available potassium), N (total nitrogen), SC (soil carbon storage), P (available phosphorus), LC (litter carbon storage), and VO (stand volume). Additionally, C1 to C4 represent the first to fourth elevation levels in the control area, while M1 to M4 represent the first to fourth elevation levels in the managed area.
Based on the analysis of this figure, the total nitrogen, soil carbon storage, available potassium, and phosphorus indices tend to be higher in the control area. Conversely, the richness and diversity indices tend to be higher in the managed area. The volume index tends to be higher at elevations above 1,500 m a.s.l. in the managed area. Litter carbon storage and stand volume show a tendency toward the control area, while the diversity and richness indices show a tendency toward the managed area.
4 Discussion
The findings of this study indicate that both the managed and control areas exhibit a J-shaped distribution curve of trees, characteristic of uneven age forests. This outcome suggests that the single-tree selective harvest method employed in the management of the studied forests did not alter their natural structure.
A similar study conducted by Moe and Owari (2020) investigated the impact of single-cutting forest management on sustainability indices in Chinese forests. Their research revealed that 50 years of forest management using the single-cutting method resulted in improved stability indices of forest stands, including a J-shaped tree distribution curve, as well as enhanced forest regeneration.
4.1 Regulating services
The soil component within ecosystems plays a pivotal role in forest environments, significantly contributing to carbon sequestration and storage (Hashimoto et al., 2009). Recent decades have seen dedicated efforts toward utilizing this resource for mitigating climate change and managing carbon. The present study’s outcomes reveal that individual tree-cutting management exerts minimal impact on soil and litter carbon storage compared to control areas. Regarding carbon storage, it is necessary to understand that increased soil carbon does not necessarily imply a decrease in ecosystem health. Ecosystem management can optimize carbon sequestration by balancing carbon stocks between soil and biomass. Soils enriched with organic carbon can support greater biodiversity, improve nutrient cycling, and increase forest resilience, even if standing biomass is slightly reduced (Lin et al., 2024; Sakib et al., 2024). In this regard, it is noted that soil carbon storage declined most in the 41–50% thinned plots due to reduced carbon storage in the humus layer (Li et al., 2023). This approach highlights the importance of soils as long-term carbon sinks, contributing significantly to climate change mitigation (Deal, 2020). While our study demonstrates a redistribution of carbon from above-ground biomass to soil carbon storage, further research is necessary to measure the annual flux of carbon sequestration. This would help to clarify whether soil carbon uptake compensates for the reduced carbon stored in the stand volume.
This paper underscores the impact of elevation on litter carbon storage and the interplay between elevation and management on soil carbon storage. Observations regarding elevation’s effect on litter carbon storage indicate that the most substantial values occur within the 1,000–1,500 m a.s.l. range. Significantly, this elevation range exhibits higher concentrations of available potassium, indicative of an enriched soil profile that enhances the physiological traits of tree species, consequently fostering increased litter accumulation. This increased litter volume, in turn, shows a greater share of litter carbon storage.
Rai et al. (2021) conducted a study on carbon storage in protected forests in India, encompassing both net and mixed scenarios. Their findings establish a clear correlation between elevated litter volume and increased carbon storage within ecosystems, particularly notable in mixed-species compositions. Similarly, Lee et al. (2020) examined soil and litter carbon dynamics in Korean forests, concluding that needle-leaf masses exhibit greater litter carbon sequestration compared to their broadleaf counterparts, primarily due to the larger litter volume of needle-leaf species. Conversely, the proliferation of beech species at elevations ranging from 1,000–1,500 m a.s.l. could contribute to enhanced litter accumulation. Mölder et al. (2008) suggested that the gradual decomposition of beech tree foliage leads to thicker litter layers. As indicated by Abrari (2020), this gradual decomposition process potentially accounts for the noticeable accumulation of beech litter on forest surfaces.
Our research unveils a significant interplay between management practices and elevation, both influencing soil carbon storage. The trends in soil carbon storage across different elevations in both study areas exhibit irregular patterns. Crucially, the managed area demonstrates its highest soil carbon storage at 1500 m a.s.l., while the control area’s peak value is observed between 1,000 and 1,500 m a.s.l. Lee et al. (2020) propose that variations in elevation and diverse management approaches impact tree growth parameters, including average crown diameter and diameter at breast height (DBH), thereby shaping carbon storage within litter and soil components. The pronounced net carbon index in both study regions likely enhances the carbon cycle between plants and soil, facilitating storage within plant and soil structures.
4.2 Supporting services
4.2.1 Woody species diversity
Biodiversity stands as a pivotal factor impacting forest ecosystem sustainability and performance, making its conservation decisive for sustainable forest management. This study highlights management’s substantial influence on species diversity, notably affecting richness and diversity indices, with managed areas displaying higher values. Raymond et al. (2018) investigated the effects of single-cutting management in Quebec, finding that enhanced diversity and richness correlated with increased light availability due to the method. Elevated cutting intensity, light, richness, and diversity fostered non-commercial species proliferation. The study infers that single-cutting management and favorable conditions for tree species regeneration bolstered species richness and diversity. Elevation significantly impacted richness, diversity, and evenness indices, with lower elevations experiencing higher values due to native interference altering light and species regeneration dynamics.
Amini et al. (2021) explored human-made and natural illumination’s effects on tree species diversity in Hyrcanian forests. Human-made illumination yielded pronounced diversity indices, possibly due to canopy gap creation. Single-cutting interventions showed minimal diversity index effects; instead, native interference and necessary cuts induced canopy changes, light alterations, and regeneration, boosting diversity indices. Management-elevation interplay influenced richness and diversity indices. Managed areas displayed increased indices with lower elevation, while control areas showed a slight rise within 1,000–1,500 m a.s.l. This corresponds to heightened indices in soil carbon storage, total nitrogen, and available potassium reported in this elevation range. These factors likely contribute to increased diversity indices in the control area at these specific elevations. Species richness and diversity all showed the highest at middle elevations in control area. In these forests, the temperature is gradually decreased and the precipitation is increased gradually along the middle altitudinal gradient. The increase in species diversity at lower elevation is due to the increase in precipitation, the decrease in that at higher elevation due to temperature limitation, and the maximum diversity at the middle elevation due to the most suitable heat and moisture conditions there (Rahbek, 1995; Colwell et al., 2004; Moradi et al., 2016). Zhang et al. (2016) stated that changes in height above sea level are one of the most important factors that affect tree species diversity indices. These researchers found the highest amount of diversity in temperate forests in middle altitudes. In this study, we observed this process in the control area, which is going through its natural sequence.
4.2.2 Nutrient recycling
The findings of this study concerning nutrient cycling and supporting service indices indicate that the management factor primarily affects soil available phosphorus, resulting in higher content in managed areas compared to control areas.
The intricate process of phosphorus retrieval and recycling faces challenges due to its transformation into sediments or organic forms. Notably, the lower base density in the control region seems to stimulate greater soil respiration, leading to heightened microbial activity. Combined with increased precipitation reaching the soil surface, these factors accelerate phosphorus mineralization and its absorption by plants. Consequently, the available phosphorus content decreases in the control area’s soil. The research by Chen et al. (2003) suggests that forests with intensified microbial activity and respiration exhibit accelerated phosphorus mineralization. Therefore, it is more simply said that plant and microbial communities at P-rich sites transfer P from soil minerals into the biogeochemical P cycle. The set of mechanisms involved in this transfer is what we term P acquiring strategy. In contrast, tight P cycling is expected at sites poor in P. That means plants and microbes use P from organic sources and minimize P losses from the biogeochemical cycle (Lang et al., 2017). Apparently, this cycle is maintained in the control area and the species have taken it out of reach by consuming available phosphorus. But this problem is in a different way in the managed area, and the use of available phosphorus from the soil has decreased due to the open harvesting of trees.
Elevation emerges as a critical determinant of available potassium levels within this study. Specifically, elevations between 600–1,000 m a.s.l. and 1,000–1,500 m a.s.l. display higher levels compared to both higher and lower elevations. Prior research establishes that total nitrogen and potassium are susceptible to alterations due to diverse management practices in natural ecosystems (Puladi et al., 2012). Middle elevations, relatively less impacted by native activities such as logging and livestock grazing, exhibit enhanced nutrient levels. This transformation enhances soil bioactivity and organic matter degradation, thus acting as a source of nutrients like nitrogen and potassium.
Regarding the combined effects of management strategies and elevation on soil nutrient indices, lower elevations contribute to increased available nitrogen and potassium levels in both regions, although some trends appear irregular. In managed areas, elevations below 600 m a.s.l. exhibit the highest nitrogen and potassium content. Similarly, this elevation range experiences peak values for tree species richness, diversity, and soil carbon storage. Conversely, the control area shows elevated total nitrogen and available potassium levels within the 1,000–1,500 m a.s.l. range, corresponding to the highest soil carbon storage, diversity and richness indexes. These patterns likely stem from the flow of organic matter into the soil, fostering degradation and subsequently elevating essential nutrient levels such as nitrogen and potassium. Earlier research supports the notion that heightened microbial activity, growth, and tree-related functions positively affect soil nitrogen and potassium content (Mahmoodi et al., 2020). In this case, it can be seen that the non-interference in the control area has caused a greater balance of forest stands to be observed in the middle altitudes and a greater reserve of nutrients in the soil.
4.2.3 Woody stem yield
In this study, the interactions between investigated factors significantly affected the stand volume of both managed and control stands. Regarding management, the control area exhibited a higher mean stand volume per hectare than the managed area. Preserving trees with substantial diameter at breast height (DBH) in the control region contributed to increased averages for DBH, basal area, and stand volume.
Regarding elevation’s influence, a notable rise in mean stand volume was observed at elevations surpassing 1,500 m above sea level (m a.s.l.). Rezaei Sangdehi et al. (2020) explored the impacts of elevational gradients on Hyrcanian forests’ quantitative traits, supporting our finding of increasing mean basal area and stand volume with ascending elevations. The augmentation in mean DBH and basal area values at higher elevations can be attributed to ecological shifts along the elevation gradient, encompassing changes in climate parameters, species composition (notably Fagus orientalis stands), and the overall Hyrcanian forest structure. As elevation rises and base density decreases, diverse species access more light and essential resources, promoting larger diameters.
Forest management, especially in single-tree selective harvest method, has been studied in relation to ecosystem health, productivity, and carbon storage. In managed forests, although a reduction in total standing volume is observed compared to unmanaged forests, this decrease is due to density regulation. Thinning practices allow growth to concentrate on a smaller number of trees, which are often of better quality and have greater growth potential. This reduction in competition among the remaining trees results in more vigorous growth, leading to better structural stability and greater resistance to diseases and pests (Li et al., 2023; Picchio et al., 2018; Lin et al., 2024). Therefore, although the total volume may be lower, productivity in terms of high-quality timber and the overall health of individual trees tend to improve (Zeller et al., 2021).
It is essential to recognize that productivity, measured in terms of standing volume, is only one indicator of ecosystem health. In managed forests, productivity manifests in other ways, such as increased species diversity, improved soil quality, and maintained carbon sequestration (Hundera et al., 2013; Nasibullina et al., 2023). Thus, even if there is a reduction in volume, the ecosystem as a whole can be considered healthier due to its balanced capacity to provide multiple ecosystem services sustainably (Ara et al., 2023). In summary, it is not suggested that soil carbon is superior to tree carbon, but rather that managed ecosystems can achieve a sustainable balance that favors both the growth of high-quality trees and soil carbon storage.
Lastly, considering combined management and elevation impacts on stand volume, the peak was identified at moderate elevations within the control zone. This aligns with the dominance of beech species in the control region, where preservation strategies supported larger diameters by avoiding extensive cutting practices. On the other hand, the presence of more suitable climatic conditions in terms of temperature and precipitation in the middle altitudes has improved the conditions of the forest in this area, increasing the diversity and richness of species, soil carbon and soil nutrients, and as a result, the trees have grown better and become stronger. All these factors have finally led to an increase in the average volume of trunk wood in this part of the control area.
5 Conclusion
This study has investigated the impact of single-tree selective harvest method on ecosystem services within an Iranian mixed temperate broadleaf forest. The results of this research showed that single-tree selective harvest method in these forests did not leave an uncontrolled impact. It was found that some characteristics of the forest, such as the indices of diversity, richness and available phosphorus, have increased in the area managed in this way. These outcomes posit that this approach could serve as an effective strategy for achieving sustainable forest management objectives.
Concerning tree species diversity, the study has demonstrated that single-tree selective harvest method contributes to heightened richness and diversity of tree species in the forest. An uncontrolled increase in the diversity and richness of tree species in forest stands is not a good thing. This study showed that after 20 years, single-tree selective harvest method in these stands has not changed in a way that would take the forest out of a stable state. It is important to note that our study does not imply that the previous management regime was unhealthy. Rather, we aim to demonstrate that the single-tree selective harvest method can maintain ecosystem services such as carbon storage, species diversity, and soil nutrient cycling, offering a sustainable approach to forest management.
Although our results demonstrate a shift in carbon storage between different forest carbon pools, this study does not evaluate the rates of carbon sequestration over time. Therefore, it is not possible to determine which forest management approach is more beneficial for climate change mitigation based solely on these findings. Future research should focus on measuring carbon accumulation rates in different forest carbon pools to provide a clearer understanding of the climate-related impacts of selective forest management. More recent studies also emphasize the importance of long-term monitoring to capture changes in carbon sequestration rates across various management regimes (Jiang et al., 2020).
Our analysis reveals a redistribution of carbon storage from above-ground biomass to below-ground biomass under different forest management regimes. While this shift indicates changes in carbon allocation, it should not be interpreted as a direct improvement in ecosystem quality. To thoroughly assess ecosystem health, further research would be required, including additional variables such as biodiversity, soil stability, and nutrient cycling, which were beyond the scope of this study.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors without undue reservation.
Author contributions
BK: Writing – original draft. AA: Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing. AB: Project administration, Writing – original draft, Writing – review & editing. SS: Formal analysis, Investigation, Writing – original draft, Writing – review & editing. PÁ-Á: Conceptualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The first author and others extend their appreciation to the Urmia University for financing the cost of her PhD thesis.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abrari, V. K. (2020). Species diversity in tree strata and its relation to some features of beech stand in Hyrcanian forests (case study: Savadkooh, Mazandaran). J. Environ. Sci. Technol. 22, 389–397.
Amini, S., Moayeri, M., Shataee, S., and Rahmani, R. (2021). Geometric indices and regeneration species diversity in natural and man-made canopy gaps. J. Wood For. Sci. Technol. 28, 1–20. doi: 10.22069/JWFST.2021.18417.1891
Ara, M., Pinno, B. D., Scaria, F., Froese, R. E., and Bokalo, M. (2023). Thinning increases individual tree growth while reducing the growth heterogeneity of Lodgepole pine. Forests 14:1091. doi: 10.3390/f14061091
Berenguer, E., Tritsch, I., Hasan, A.F., Sist, P., and Gond, V. (2018). The potential of multisource remote sensing for mapping the biomass of a degraded Amazonian forest. Forests. 9:303. doi: 10.3390/f9060303
Bremner, J., and Mulvaney, C. (1982). “Total nitrogen” in Methods of soil analysis. Part 2. eds. A. L. Page, R. H. Miller, and D. R. Keeney (Madison, WI, USA: Amer. Soc. Agron.), 595–624.
Carmona-Yáñez, M. D., Lucas-Borja, M. E., Zema, D. A., Jing, X., Kooch, Y., Garrido Gallego, P., et al. (2023). Influence of management and stand composition on ecosystem multifunctionality of Mediterranean tree forests. Trees 37, 1801–1816. doi: 10.1007/s00468-023-02462-w
Chen, C., Condron, L., Davis, M., and Sherlock, R. (2003). Seasonal changes in soil phosphorus and associated microbial properties under adjacent grassland and forest in New Zealand. For. Ecol. Manag. 177, 539–557. doi: 10.1016/S0378-1127(02)00450-4
Colwell, R. K., Rahbek, C., and Gotelli, N. J. (2004). The mid-domain effect and species richness patterns: what have we learned so far? Am. Nat. 163, E1–E23. doi: 10.1086/382056
Daily, G. (1998). Nature's services: societal dependence on natural ecosystems. Environ. Values 7, 365–367.
Deal, R. L. (2020). Integrating ecosystem services into sustainable forest management of public lands. For. Manag. Res. Partnerships 83, 83–93.
Grammatikopoulou, I., and Vačkářová, D. (2021). The value of forest ecosystem services: a meta-analysis at the European scale and application to national ecosystem accounting. Ecosyst. Serv. 48:101262. doi: 10.1016/j.ecoser.2021.101262
Hasan, B. M. S., and Abdulazeez, A. M. (2021). A review of principal component analysis algorithm for dimensionality reduction. J. Soft Comput. Data Min. 2, 20–30. doi: 10.30880/jscdm.2021.02.01.003
Hashimoto, T., Miura, S., and Ishizuka, S. (2009). Temperature controls temporal variation in soil CO2 efflux in a secondary beech forest in Appi Highlands, Japan. J. For. Res. 14, 44–50. doi: 10.1007/s10310-008-0096-2
Humphries, C. J., Williams, P. H., and Vane-Wright, R. I. (1995). Measuring biodiversity value for conservation. Annu. Rev. Ecol. Syst. 26, 93–111. doi: 10.1146/annurev.es.26.110195.000521
Hundera, K., Aerts, R., DeBeenhouwer, M., VanOvertveld, K., Helsen, K., Muys, B., et al. (2013). Both forest fragmentation and coffee cultivation negatively affect epiphytic orchid diversity in Ethiopian moist evergreen Afromontane forests. Biol. Conserv. 159, 285–291. doi: 10.1016/j.biocon.2012.10.029
Jiang, M., Medlyn, B. E., Drake, J. E., Duursma, R. A., Anderson, I. C., Barton, C. V., et al. (2020). The fate of carbon in a mature forest under carbon dioxide enrichment. Nature 580, 227–231. doi: 10.1101/696898
Khanalizadeh, A., Rad, J. E., Amiri, G. Z., Zare, H., Rammer, W., and Lexer, M. J. (2020). Assessing selected microhabitat types on living trees in oriental beech (Fagus orientalis L.) dominated forests in Iran. Ann. For. Sci. 77, 1–13. doi: 10.1007/s13595-020-00996-4
Lal, R. (2004). Soil carbon sequestration impacts on global climate change and food security. Science 304, 1623–1627. doi: 10.1126/science.1097396
Lang, F., Krüger, J., Amelung, W., Willbold, S., Frossard, E., Bünemann, E. K., et al. (2017). Soil phosphorus supply controls P nutrition strategies of beech forest ecosystems in Central Europe. Biogeochemistry 136, 5–29. doi: 10.1007/s10533-017-0375-0
Lee, S., Lee, S., Shin, J., Yim, J., and Kang, J. (2020). Assessing the carbon storage of soil and litter from National Forest Inventory data in South Korea. Forests 11:1318. doi: 10.3390/f11121318
Li, X., Wang, R., Liu, K., Zhou, Y., and Hu, J. (2023). Effects of thinning on tree growth and soil physiochemical properties in Cunninghamia lanceolata plantation. J. For. Res. 28, 251–259. doi: 10.1080/13416979.2023.2198156
Lin, N., Feng, M., Huang, H., Qiu, Z., Ma, T., and Chen, S. (2024). Effects of thinning on carbon storage in a mixed broadleaved plantation in a subtropical area of China. Forests 15:638. doi: 10.3390/f15040638
Nasibullina, A., van der Maaten-Theunissen, M., van der Maaten, E., Fischer, H., and Wagner, S. (2023). Thinning effects on growth and occurrence of rotting in aspen stands. J. For. Sci. 69, 525–538. doi: 10.17221/103/2023-JFS
MacDicken, K.G. (1997). A guide to monitoring carbon storage in forestry and agroforestry projects. Arlington, Virginia: Winrock Internationl Institute for Agricultural Development.
Mahmoodi, M., Kooch, Y., and Kianmehr, A. (2020). Effect of broadleaved tree species on chemical and microbial soil properties in plain forest of Noor. Iranian Journal of Forest and Poplar Research. 28, 85–96. doi: 10.22092/ijfpr.2020.341983.1909
Mehlich, A. (1978). New extractant for soil test evaluation of phosphorus, potassium, magnesium, calcium, sodium, manganese and zinc. Commun. Soil Sci. Plant Anal. 9, 477–492. doi: 10.1080/00103627809366824
Millennium Ecosystem Assessment (2005). Ecosystems and human well-being: wetlands and water. Island: World Resources Institute.
Moe, K. T., and Owari, T. (2020). Sustainability of high-value timber species in mixed conifer–broadleaf forest managed under selection system in northern Japan. Forests 11:484. doi: 10.3390/f11050484
Mölder, A., Bernhardt-Römermann, M., and Schmidt, W. (2008). Herb-layer diversity in deciduous forests: raised by tree richness or beaten by beech? For. Ecol. Manag. 256, 272–281. doi: 10.1016/j.foreco.2008.04.012
Moradi, H., Naqinezhad, A., Siadati, S., Yousefi, Y., Attar, F., Etemad, V., et al. (2016). Elevational gradient and vegetation-environmental relationships in the central Hyrcanian forests of northern Iran. Nord. J. Bot. 34, 1–14. doi: 10.1111/njb.00535
Nelson, D. W., and Sommers, L. E. (1996). “Total carbon, organic carbon, and organic matter” in Methods of soil analysis. USA: Soil Science Society of America and American Society of Agronomy, Madison. Part 3 Chemical methods, vol. 5, 961–1010. doi: 10.2136/sssabookser5.3.c34
Nilsson, M.-C., Wardle, D. A., and Dahlberg, A. (1999). Effects of plant litter species composition and diversity on the boreal forest plant-soil system. Oikos 86, 16–26. doi: 10.2307/3546566
Oettel, J., and Lapin, K. (2021). Linking forest management and biodiversity indicators to strengthen sustainable forest management in Europe. Ecol. Indic. 122:107275. doi: 10.1016/j.ecolind.2020.107275
Olsen, S. R. (1954). Estimation of available phosphorus in soils by extraction with sodium bicarbonate. Washington, DC: US Government Printing office, US Department of Agriculture.
Panja, P. (2021). Deforestation, carbon dioxide increase in the atmosphere and global warming: a modelling study. Int. J. Model. Simul. 41, 209–219. doi: 10.1080/02286203.2019.1707501
Pecl, G. T., Araújo, M. B., Bell, J. D., Blanchard, J., Bonebrake, T. C., Chen, I.-C., et al. (2017). Biodiversity redistribution under climate change: impacts on ecosystems and human well-being. Science 355:eaai9214. doi: 10.1126/science.aai9214
Picchio, R., Venanzi, R., Latterini, F., Marchi, E., Laschi, A., and Lo Monaco, A. (2018). Corsican pine (Pinus laricio Poiret) stand management: medium and long-lasting effects of thinning on biomass growth. Forests 9:257. doi: 10.3390/f9050257
Pielou, E. (1979). A quick method of determining the diversity of foraminiferal assemblages. J. Paleontol. 35, 1237–1242.
Platon, V., Frone, S., and Constantinescu, A. (2015). New developments in assessing forest ecosystem services in Romania. Proc. Econ. Finance 22, 45–54. doi: 10.1016/S2212-5671(15)00225-7
Pollastrini, M., Holland, V., Brüggemann, W., Koricheva, J., Jussila, I., Scherer-Lorenzen, M., et al. (2014). Interactions and competition processes among tree species in young experimental mixed forests, assessed with chlorophyll fluorescence and leaf morphology. Plant Biol. 16, 323–331. doi: 10.1111/plb.12068
Puladi, N., Delavar, M., Golchin, A., and Koper, A.M. (2012). Effects of plantation on soil quality indicators and carbon sequestration in Safrabasteh poplar research station in Guilan province. Iranian Journal of Forest and Poplar Research. 20, 84–95. doi: 10.22092/ijfpr.2012.6103
Rahbek, C. (1995). The elevational gradient of species richness: a uniform pattern? Ecography 18, 200–205. doi: 10.1111/j.1600-0587.1995.tb00341.x
Rai, P., Vineeta, S., Shukla, G., Manohar K, A., Bhat, J. A., Kumar, A., et al. (2021). Carbon storage of single tree and mixed tree dominant species stands in a reserve forest—case study of the eastern sub-Himalayan region of India. Land 10:435. doi: 10.3390/land10040435
Raymond, P., Royo, A. A., Prévost, M., and Dumais, D. (2018). Assessing the single-tree and small group selection cutting system as intermediate disturbance to promote regeneration and diversity in temperate mixedwood stands. For. Ecol. Manag. 430, 21–32. doi: 10.1016/j.foreco.2018.07.054
Rezaei Sangdehi, S. M., Fallah, A., Oladi, J., and Latifi, H. (2020). Determination the effect of altitude gradient on quantitative characteristics of Forest stands (case study: district-3 of Sangdeh forests). J. Wood For. Sci. Technol. 27, 1–16.
Sagheb Talebi, K., Sajedi, T., and Pourhashemi, M. (2014). Forests of Iran: a treasure from the past, a Hope for the future. Netherlands: Springer. doi: 10.22069/JWFST.2019.16794.1817
Sakib, N., Poudel, T. R., Hao, Y., Roberts, N. J., Iddrisu, A.-Q., Adhikari, S., et al. (2024). Effects of different thinning intensities on carbon storage in Pinus koraiensis middle-aged plantations in Northeast China. Forests 15:738. doi: 10.3390/f15050738
Sasanifar, S., Alijanpour, A., Shafiei, A. B., Rad, J. E., Molaei, M., and Azadi, H. (2019). Forest protection policy: lesson learned from Arasbaran biosphere reserve in Northwest Iran. Land Use Policy 87:104057. doi: 10.1016/j.landusepol.2019.104057
Shannon, C. E. (1949). The mathematical theory of communication, by CE Shannon (and recent contributions to the mathematical theory of communication), W. Weaver. Champaign, IL, USA: University of Illinois Press.
Sohrabi, H., Jourgholami, M., Lo Monaco, A., and Picchio, R. (2022). Effects of forest harvesting operations on the recovery of earthworms and nematodes in the Hyrcanain old-growth forest: assessment, mitigation, and best management practice. Land 11:746. doi: 10.3390/land11050746
Tong, X., Brandt, M., Yue, Y., Ciais, P., Rudbeck Jepsen, M., Penuelas, J., et al. (2020). Forest management in southern China generates short term extensive carbon sequestration. Nat. Commun. 11:129. doi: 10.1038/s41467-019-13798-8
Trumper, K. (2009). The natural fix? The role of ecosystems in climate mitigation: a UNEP rapid response assessment. Cambridge, UK: UNEP/Earthprint.
Vafaei, S., Soosani, J., Adeli, K., Fadaei, H., and Naghavi, H. (2017). Estimation of aboveground biomass using optical and radar images (case study: Nav-e Asalem forests, Gilan). Iranian Journal of Forest and Poplar Research. 25, 320–331.
Wallace, K. J. (2007). Classification of ecosystem services: problems and solutions. Biol. Conserv. 139, 235–246. doi: 10.1016/j.biocon.2007.07.015
Wheeler, C. E., Omeja, P. A., Chapman, C. A., Glipin, M., Tumwesigye, C., and Lewis, S. L. (2016). Carbon sequestration and biodiversity following 18 years of active tropical forest restoration. For. Ecol. Manag. 373, 44–55. doi: 10.1016/j.foreco.2016.04.025
Zeller, L., Caicoya, A. T., and Pretzsch, H. (2021). Analyzing the effect of silvicultural management on the trade-off between stand structural heterogeneity and productivity over time. Eur. J. For. Res. 140, 615–634. doi: 10.1007/s10342-020-01350-z
Zhang, Q., Hakkenberg, C. R., and Song, C. (2018). Evaluating the effectiveness of forest conservation policies with multi-temporal remotely sensed imagery: a case study from Tiantangzhai township, Anhui, China. Comprehen. Remote Sens. 9, 39–58. doi: 10.1016/B978-0-12-409548-9.10435-X
Zhang, B., Li, W., and Xie, G. (2010). Ecosystem services research in China: progress and perspective. Ecol. Econ. 69, 1389–1395. doi: 10.1016/j.ecolecon.2010.03.009
Zhang, J.-T., Zhang, M., and Mian, R. (2016). Effects of elevation and disturbance gradients on forest diversity in the Wulingshan nature reserve, North China. Environ. Earth Sci. 75, 1–10. doi: 10.1007/s12665-016-5734-y
Keywords: single-tree selective harvest method, forest management, ecosystem services, carbon storage, tree species diversity
Citation: Karamdoost Marian B, Alijanpour A, Banj Shafiei A, Sasanifar S and Álvarez-Álvarez P (2024) Effects of single-tree selective harvest method on ecosystem services in a mixed temperate broadleaf forest in Iran. Front. For. Glob. Change. 7:1461996. doi: 10.3389/ffgc.2024.1461996
Edited by:
Sumit Chakravarty, Uttar Banga Krishi Viswavidyalaya, IndiaReviewed by:
David Ellison, University of Bern, SwitzerlandBrian J. Palik, Forest Service (USDA), United States
Copyright © 2024 Karamdoost Marian, Alijanpour, Banj Shafiei, Sasanifar and Álvarez-Álvarez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: A. Alijanpour, YS5hbGlqYW5wb3VyQHVybWlhLmFjLmly
†These authors have contributed equally to this work