 Simon Lebel Desrosiers
Simon Lebel Desrosiers Alexandre Collin
Alexandre Collin Nicolas Bélanger
Nicolas Bélanger- 1Data Science Laboratory, Université du Québec (TÉLUQ), Montréal, QC, Canada
- 2Département des sciences naturelles, Institut des Sciences de la Forêt tempérée (ISFORT), Université du Québec en Outaouais (UQO), Ripon, QC, Canada
Red oak is an important species within the North American landscape, with climate change projections indicating a potential northward shift in its distribution. However, understanding the factors influencing its regeneration success at the northern limit remains limited. Site conditions and seed provenance adaptability may play critical roles. To bridge this knowledge gap, we conducted a seed transfer study in two northern red oak stands in Quebec. We firstly investigated stand regeneration history through dendrochronological characterization. Then, we monitored the survival and growth of saplings for four red oak provenances across a south-to-north gradient in field and greenhouse settings, with varying soil nutrient levels due to fertilization, and with or without protection from large herbivores. Results indicated that stands have similar age structures with red oak establishment coinciding with the last major fire disturbance in the early 1920s. However, tree species composition and density differed, suggesting differences in fire disturbance regime or ecological succession status prior to fire. Site had the largest influence on red oak regeneration, with the highest tree density and soil water availability site exhibiting a 29% higher survival rate. Protection against large herbivores also significantly impacted red oak seedling performance, leading to a 16% higher survival rate. Germination, survival and growth also significantly differed between provenances. The local (northernmost) provenance exhibited the poorest overall performance with 28 to 42% lower germination, survival and growth rates, while the two southernmost provenances exhibited superior germination and sprout survival. An increase in soil nutrient availability was beneficial to red oak in the greenhouse, but only marginally benefited survival and growth in the field, suggesting that this factor is of less importance than other factors (e.g., water and light availability) for red oak early regeneration. The findings of this study suggest that silvicultural efforts to favor red oak should focus on site and provenance selection, and that water availability is currently limiting red oak regeneration success at acorn and early seedling life stages (i.e., 1 year old seedlings) near its northern distribution limit.
1 Introduction
Red oak (Quercus rubra L.) is native of North America and is characterized by a wide distribution, ranging from the upper midwestern United States (Minnesota) to the Canadian Maritimes (Nova Scotia) to the southeastern (Mississippi) and south-central (Oklahoma) states. Its establishment was mostly attributed to the warmer and dryer climate of the early Holocene (Abrams, 1992) and was also likely favored by First Nations and European settlements (Dey and Guyette, 2000; Munoz and Gajewski, 2010) and fire disturbance (Shumway et al., 2001; Brose et al., 2014). Prescribed burning studies suggest that fire can enhance red oak regeneration, although fire frequency and severity determine red oak regeneration success or failure (Signell et al., 2005; Knapp et al., 2015; Greenler et al., 2020). Red oak has fire-resistant physiological adaptations such as post-fire sprouting (Perala, 1974; Sander, 1990), bark thickness (Crow, 1988, Dey and Schweitzer, 2018) and wood compartmentalization after fire injury (Shigo and Shortle, 1979; Brose et al., 2014). These adaptations lead to the idea that fire is a primary driver of red oak regeneration. The benefits of fire disturbance on red oak regeneration include the suppression of late-successional, shade tolerant competing species (Brose et al., 1998; Dey and Guyette, 2000) and soil amendments with wood ash and charcoal, but this may also benefit other species such as red maple (Acer rubrum L.) (Green et al., 2010; Granger et al., 2018).
The production of wood ash during a fire improves the acid–base status of the forest floor, e.g., pH and exchangeable Ca, Mg and K, whereas the effects of ash on the mineral soil appear several years or decades after deposition, and the amplitude of its effect varies as a function of the amount of ash produced and soil types, among other factors (Thiffault et al., 2007; Reid and Watmough, 2014). By increasing soil pH, wood ash can also affect N mineralization, thus generating overall improved soil conditions for plant growth (Raison, 1979; Brais et al., 2015). Additionally, with its high surface area, charcoal can improve soil conditions for plant growth by adsorbing allelopathic compounds such as tannins and phenols, which have been recognized as adversely impacting red oak acorn germination and seedling radicle growth (Lodhi, 1978; Hanson and Dixon, 1987). This effect varies with the types and sources of allelopathic substances (Nilsen et al., 1999). Adsorption of phenols can also increase soil microbial activity and nitrification (DeLuca et al., 2006; Lehmann et al., 2011) and thus further improve soil nutrient availability (Jacoby et al., 2017; Zhou et al., 2020). However, fire can negatively impact soils by decreasing organic matter levels and increasing bulk density of surface soil (Phillips et al., 2000).
While red oak regeneration has been limited in its native North American distribution in the last century (Dey et al., 2008; Loftis and McGee, 1993), the species has been highly competitive and has established many areas very successfully in Europe since its introduction in the 18th century (Major et al., 2013; Nicolescu et al., 2020). The fast expansion in Europe where fire disturbance is not prevalent suggests that red oak regeneration is not entirely dependent of fire disturbance, and that other factors (e.g., climate) may play a large role in its failure to regenerate in North America. Other important factors affecting red oak regeneration include its 2–5 years masting cycle (Sander, 1990; Abrams and Johnson, 2013), acorn predation (Schnurr et al., 2004; Lombardo and McCarthy, 2009), herbivory (Ward et al., 2000; Blossey et al., 2019), acorn size and quality, e.g., free of infections (Kormanik et al., 1998; Yi et al., 2015), genotypes with highly specific adaptations (McGee, 1974; Kriebel et al., 1988; Sork et al., 1993; Aldrich et al., 2005) and site intrinsic conditions including soil physicochemical properties as well as light and water availability (Phares, 1971; Kolb et al., 1990; Bauweraerts et al., 2013). This study tested the effect of multiple factors on red oak regeneration of four northern provenances, including fire and site-specific effects along with fertilization with ash and biochar as well as large herbivores. Furthermore, this study combines two experiments, i.e., in the greenhouse and in the field under red oak stands, to more fully elucidate the potential role of each of these factors under both controlled and uncontrolled environments.
Red oak is an important tree species to maintain in the North American landscape as it is a source of high-value lumber and is a significant food source and shelter for fauna (Sork et al., 1983; Pekins and Mautz, 1988). Models project that climate change should be beneficial to the expansion of red oak in northeastern North America and a northern shift in the distribution of the species is expected (Iverson et al., 2019; Prasad et al., 2020). However, such models are generic and they generally overlook specific conditions and environmental factors that control regeneration at finer spatial scales. As such, studies on the regeneration dynamics of red oak at its northern distribution limit have value because this environment is likely to become more prevalent for red oak under climate change. A better understanding of the early ecology of the species at these latitudes will guide management strategies to maintain the species locally and perhaps to increase its distribution northward under climate change.
Two mature red oak stands were studied at the northern distribution limit of the species in Quebec, a rare occurrence at these latitudes at present. The first objective of this study was to assess tree species composition and age structure as a means to assess red oak regeneration history in the stands. We hypothesized that both stands exhibited similar age structure and that their occurrence closely matches the last major fire event in the area (early 1920s, Savage, 2001). The second objective was to assess the influence of seed provenance on seedling regeneration and growth, in a controlled environment (greenhouse) and in the two red oak stands, to isolate and test critical factors such as site intrinsic conditions, herbivory and soil fertility. We hypothesized that (1) intrinsic site conditions and provenances significantly affect germination, survival and growth of red oak, (2) large herbivores have important adverse impacts, and (3) increasing soil nutrient availability by adding wood ash and biochar has benefits.
2 Materials and methods
2.1 Study site
The study was conducted at the Station de Biologie des Laurentides (SBL) of Université de Montréal in St. Hippolyte, Quebec (45°98′93″ N and 74°00′61″ W, Figure 1). The site is at the northern limit of the maple-yellow birch (Betula alleghaniensis Britt.) bioclimatic domain of the lower Laurentians. It is mostly composed of maple stands under mesic conditions and coniferous stands [Abies balsamea (L.) Mill. or Thuja occidentalis L.] under hydric and sub hydric conditions (Savage, 2001). The site is characterized by a warm and humid summer. The mean annual temperature, precipitation, days without frost and degree-days simulated with the BioSIM model (Régnière and Bolstad, 1994) between 2003 and 2013 were, respectively, 4.9°C, 1270 mm (with 30% falling as snow), 153 and 2,845 (Bélanger et al., 2021a). Soils are sandy and developed from glacial till made up of anorthosite (Morin series) and felsic rocks (Bélanger et al., 2012) and are classified as Orthic Ferro-Humic and Humo-Ferric Podzols (Soil Classification Working Group, 1998). The site is part of the Grenville geological province, which is in turn part of the Canadian Precambrian Shield.
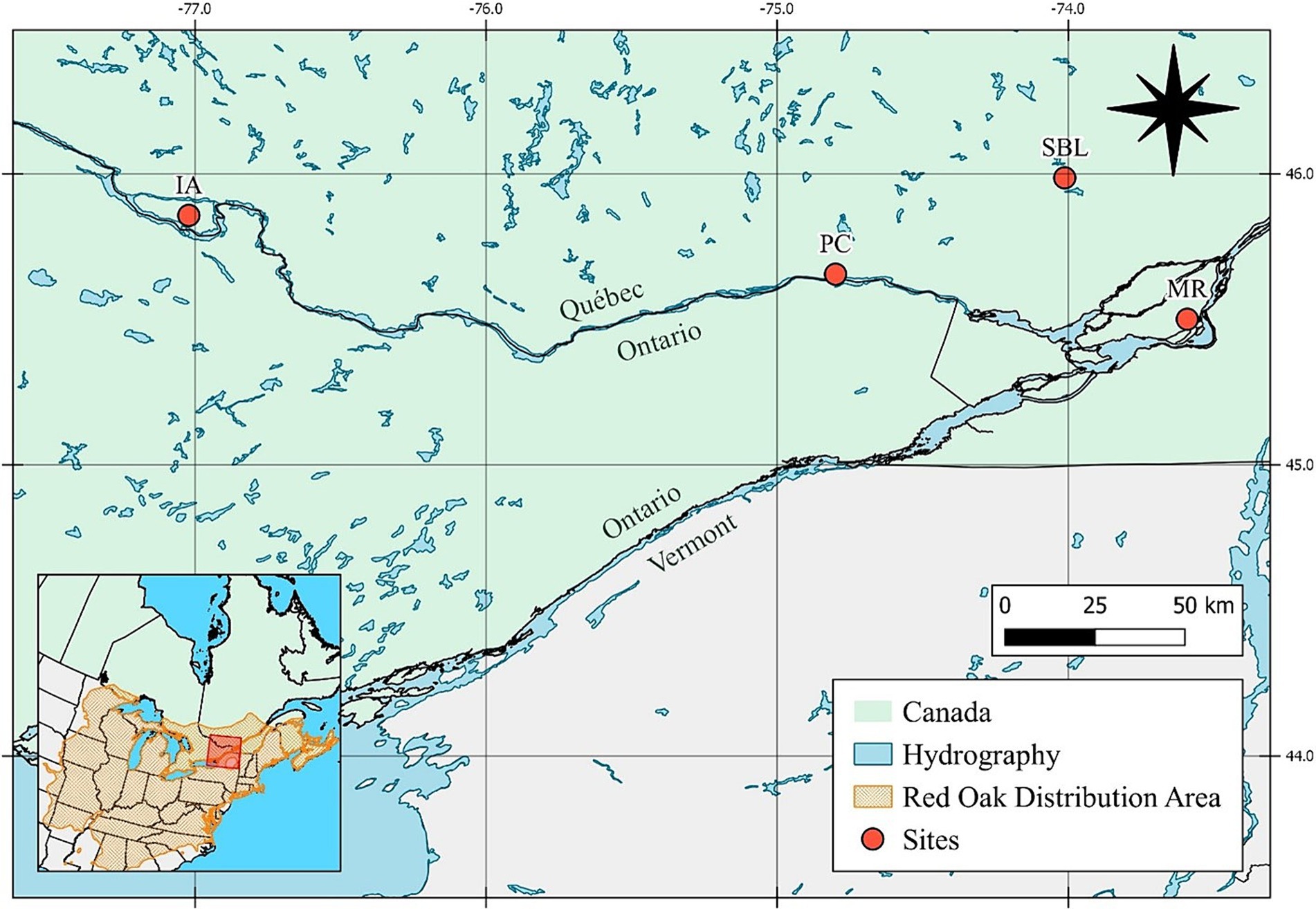
Figure 1. Site locations of red oak acorn provenances used for the 2019 field experiment at Station de biologie des Laurentides (SBL). IA is Isle-aux-Allumettes, PC is Pointe-au-Chêne and MR is Mont-Royal. Data was gathered from Government of Quebec (2019) and map projection is NAD Quebec Albers (EPSG:6624).
The only two red oak stands at SBL, i.e., Lac en Coeur (site 1; 45°58′20″ N and 73°59′51″ W) and Lac Corriveau (site 2; 45°58′52″ N and 73°59′29″ W) were selected for this study. They present mature and dominant red oak trees, are situated on well-drained hilltops with south-southeast facing slope aspects and exhibit limited red oak regeneration. They also offer diversity in terms of species composition (Table 1). Both sites are characterized by the dominance of red oak and sugar maple. The tree species composition in site 1 is less diversified than site 2. In site 1, red oak and sugar maple have an identical relative abundance and make up for 92.8% of the sampled trees, whereas only 3 other species were inventoried. Site 2 exhibits a more diversified composition as 11 species were inventoried in the stand. Red oak and sugar maple also make up the bulk of the trees in site 2 (i.e., relative abundance of 89%), but in contrast to site 1, red oak trees are substantially less abundant than sugar maple trees. Red oak and sugar maple trees have an average DBH of 54.7 cm and 32.6 cm in site 1 and of 37.9 cm and 23.1 cm in site 2, respectively (Supplementary material A). Due to their larger diameters, basal area of red oak at sites 1 and 2 is, respectively, about 25 and 5 m2 ha-1 greater than that of sugar maple. Tree density is higher at site 2 than site 1, with an average of 566 and 382 trees per hectare, respectively (results not shown). Many red oak trees show fused stems within the first 100 cm, which is characteristic of a post-fire regeneration pattern, i.e., multiple stems growing from the same stool. Fire scars on these main bases are often visible, but they were not investigated further to assess the year of fire occurrence more precisely.
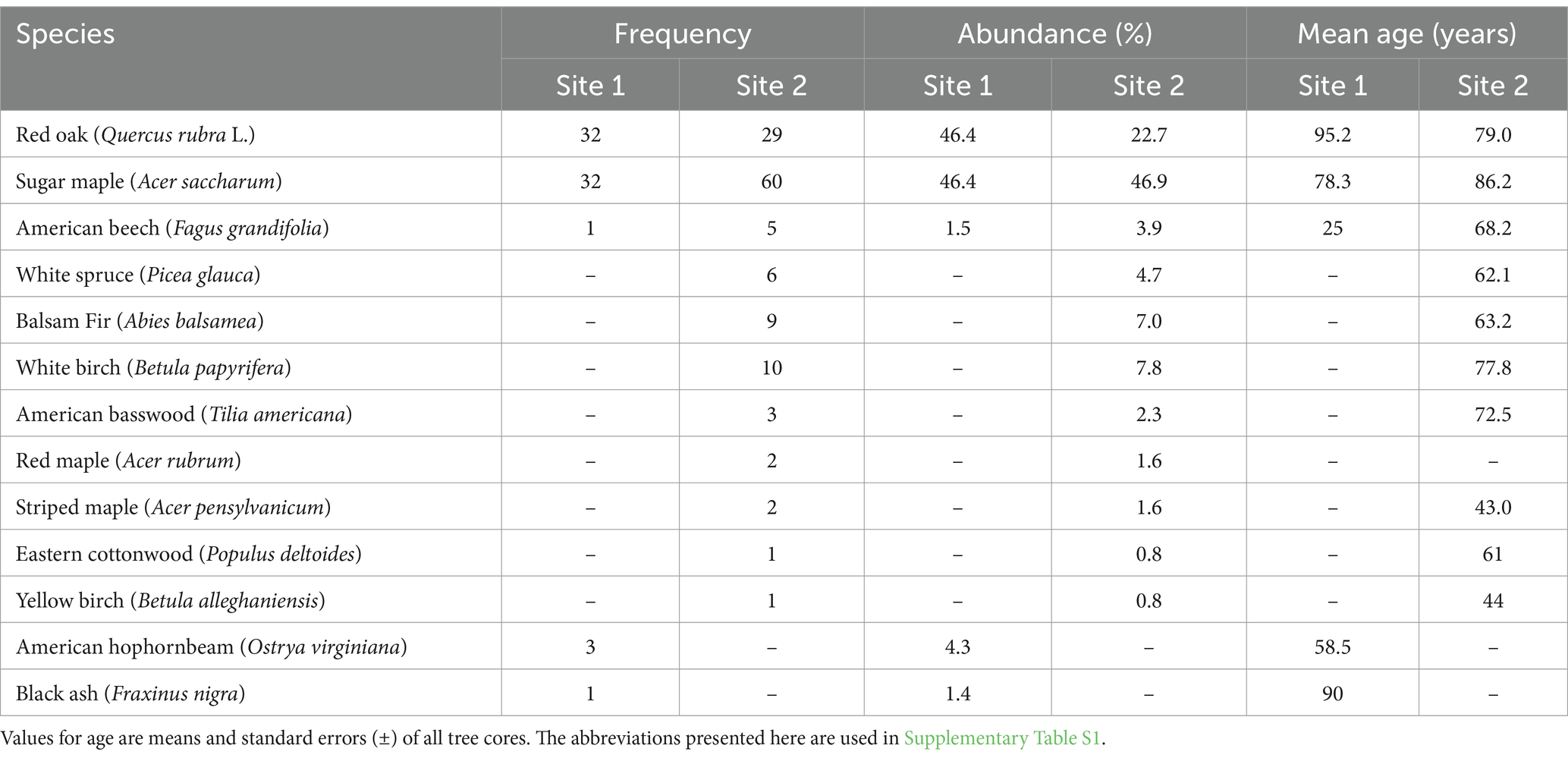
Table 1. Tree species frequencies, relative abundances and average age in the two study sites.
2.2 Site dendroecological characterization
Data were obtained from four and five 452 m2 circular plots (12 m radius) randomly selected in sites 1 and 2, respectively, in the fall of 2018 (Supplementary material G). Within each plot, all trees with a diameter at breast height (DBH) > 10 cm were identified by species and cored using a Haglof 5.1 mm diameter increment borer. For each plot, the percent contribution of each tree species to the total number of trees (abundance) was computed. Collected tree cores were prepared and sanded using standard methods (Payette and Filion, 2018) before counting growth rings under a M80 binocular microscope (Leica, Germany).
2.3 Acorn collection and preparation
Red oak acorns were directly collected in pristine forests at three different sites in the fall of 2017. Acorn collection sites included SBL as well as Mont-Royal (MR) and Pointe-au-Chêne (PC), all within the province of Quebec (Figure 1). Acorns of a fourth provenance, i.e., Isle-aux-Allumettes (IA) were obtained in winter of 2018 from a nursery operated by the ministère des Forêts, de la Faune et des Parcs of the Quebec Government. For the SBL provenance, acorns were specifically collected at the two surveyed sites (1 and 2). The sites were selected to capture a latitudinal gradient that mainly encompassed differences in climate, but it also holds differences in stand compositions, geology and soils. The MR provenance is the most southern (45°30′23″ N), followed by the IA provenance (45°51′24″ N), the PC provenance (45°65′45″ N) and the SBL (local) provenance (45°98′93″ N). The MR, PC, and IA sites are warmer and have less precipitation than SBL. The MR, PC, IA, and SBL, respectively, present a 6.8, 6.1, 5.0, and 4.3°C mean annual temperature and 1,000, 1,009, 853, and 1,193 mm mean annual precipitation (1981–2010 period, Government of Canada, 2020). The MR site is within the sugar maple-bitternut hickory bioclimatic domain, whereas the PC and IA sites are located near the northern limit of the sugar maple-bitternut hickory bioclimatic domain, with sugar maple-basswood and sugar maple-yellow birch just north of the PC and IA sites, respectively (Saucier et al., 2009). Like SBL, the PC site belongs to the Grenville geological province, and its soils were developed from glacial till composed of gneiss and marble (Ministère de l’Énergie et des Ressources naturelles, 2020). The MR site is part of the Monteregian Hills complex and is characterized by a magmatic intrusion formed during the late Mesozoic (~125 Ma), unearthed by the differential erosion of the glaciers’ over the last 2 million years. Soils are mostly formed from melanocratic and leucocratic gabbro (Amórtegui et al., 2010). The soils at the AI site are derived from alkaline sedimentary rocks of the Trenton group, including shale, limestone and dolomite (Thériault and Beauséjour, 2012). The upper mineral soil (B horizon) was sampled at five locations at all sites and analyzed for pH, total C and N as well as bulk geochemical composition (see Soil analysis section below for details). As a whole, soils at the MR and PC sites are of greater quality (i.e., higher pH and levels of nitrogen, calcium, magnesium and phosphorus) than the SBL and IA sites (Supplementary material B).
Acorn stratification was completed following the protocol described by Nature-Action Québec (2015). The acorns were first washed with a diluted NaClO solution (5%) of the original concentrated bleach solution (3%). This allowed to kill pathogens and/or mold, thus maximizing the conservation of seeds from the start-to-end of the stratification process throughout the winter period. Acorns were then selected based on the criterion of non-viable vs. viable acorns. More specifically, acorns that presented visible outside damages (mold, fissures, holes, etc.) were discarded as non-viable. Also, acorns that floated during the bleaching process were assumed to have an affected core (e.g., weevil, Curculo glandium M.) and were thus also discarded. Sorting of the acorns was done to allow optimal quality and germination rates. Selected acorns were then stored in the refrigerator at 2°C for 3 months in large plastic bags prepared with a vermiculite bed to absorb the excess initial moisture. Distilled water was sprayed in each bag on a weekly basis to maintain relative humidity at about 25% to prevent drying. Bags were also opened each week to oxygenate acorns and prevent mold development. Finally, acorns were soaked in water for 24 h one day prior to transplantation in the pots or field plots.
2.4 Greenhouse experiment
A first experiment was carried out for 10 weeks in a greenhouse during summer 2018 with the objective of characterizing germination rates, growth and survival of the red oak provenances under a soil richness gradient. One acorn was planted at the soil surface in 100 cm3 pot. A total of 170 acorns were planted for SBL and 230 acorns were planted for MR and PC each. The IA provenance was not tested as seeds were not yet available. The pots were placed in 21 rows containing 30 pots each (Supplementary material H). Each row was comprised of three sets of 10 pots of each provenance, placed randomly along the rows. Soil richness treatments were randomly distributed along the rows in series of three pots. This ensured that both the provenances and treatments were distributed throughout the room, thus reducing possible bias induced by unaccounted environmental variations in the greenhouse.
The potting soils were prepared from a base of sand mixed with rich Agro Mix® N7 organic soil (Fafard, Canada). The soil richness treatments were produced from mixing the sand and organic soils in varying proportions. The sand and organic soil mixture corresponded to 9:1, 5:5, and 1:9 volume ratio for the low, average and high soil richness treatments, respectively. The sand and organic soil mixtures were prepared in a concrete mixer, which we ran for 10 min to obtain a fully homogenized soil. Salifu and Jacobs (2006) studied red oak seedling growth under different fertilization treatments and suggested that a fertilization of 15 N-15P-15 K of 25 mg N plant-1 season-1 maximized dry mass production, 100 mg N plant-1 season-1 led to optimum N and P uptake, and 150 mg N plant-1 season-1 induced N and P toxicities. Based on this information, we applied a water soluble 20 N-20P-20 K fertilizer (Miracle-Gro™, Scotts, Canada) to the pots to enhance differences between the soil richness treatments. The low soil richness treatment received a total 3.75 mg N of fertilizer season-1, while the average soil richness treatment received 15 mg N season-1 and the high soil richness treatment received 75 mg N season-1. Equivalents using a 15 N-15P-15 K fertilizer would be 5 mg, 20 mg, and 100 mg N for the low, average and high soil richness treatments, respectively. Air temperature was set at 24°C during the day and 20°C at night, whereas light was provided for 16 h d-1. Pots were watered using a tubing system with individual outlets planted in each pot. The watering system was activated two times per week for a duration of 1 min (equivalent to 300 mL of water per pot). This kept the soil evenly moist but not saturated. Excess water could easily drain at the bottom of the perforated pots.
To assess the performance of red oak provenances as a function of soil richness, growth stages were measured throughout the experiment. Stage 1 corresponded to a successful germination, stage 2 to stem development, stage 3 to the onset of leaf development, and stage 4 to multiple/full leaf development. Germination and sprout survival rates were also monitored. Further total mortality rate per provenance was calculated. This number corresponded to the number of dead sprouts as well as the seeds that did not germinate. A subsample of seedlings (n = 32, 40, and 36 for SBL, MR and PC, respectively) was selected at end of the experiment for destructive measurements. Response variables measured were stem height (cm), stem mass (g), number of leaves, dry leaf mass (g), leaf area (cm2), support root mass (g), and fine root mass (g). Specifically, leaf area was assessed by scanning leaves as.tiff files on a white paper sheet as background and then estimating leaf area with the LeafArea R package (Katabuchi, 2015), which acts as an interface for the image processing program “ImageJ.” All mass measurements were carried out on dried samples (40°C for 48 h), except for leaf area which was measured on air-dried leaves. Other variables derived from direct measurements were mean leaf mass (g leaf-1), mean leaf foliar area (cm2 leaf-1), aboveground biomass (g), belowground biomass (g), total biomass (g), and root:shoot mass ratio.
Potting soils from the three soil richness treatments were sampled at the beginning and end of the experiment (i.e., weeks 0 and 10). Soils were representative of substrates without and with the weekly addition of a fertilizer. Three replicates were sampled for each treatment, for a total of 18 soil samples to be processed in the laboratory for chemical analysis.
2.5 Field experiment
A second experiment was carried out in the field at SBL. The experimental split-split-plot design included ten blocks distributed evenly among sites. Each block contained twenty-four 900 cm2 micro-plots separated by 10 cm wide buffers. Micro-plots were identified with stake flags. Half of the micro-plots were set up within a 1.83 m high galvanized metal fence (exclosures) in order to protect red oak seedlings from large herbivores (e.g., deer). The other half of the micro-plots was left unprotected from large herbivores. Each half-block was subdivided randomly into 3 provenances and 4 soil fertilization treatments. The layout was randomly distributed based on the protection from large herbivores (factor 1), followed by the soil fertilization treatments (factor 2), and finally by the random distribution of the 3 provenances within a soil fertilization treatment (factor 3). The soil fertilization treatments included a control, wood ash, biochar, and a combination of ash and biochar. The factor hierarchy is shown in Supplementary material I, whereas an example of an established experimental block is available in Supplementary material J. Wood ash and biochar extractible cations concentrations were estimated via the Mehlich-III acid extraction method. They, respectively, yielded 3,574 and 21.1 mg Ca/L, 150 and 10.5 mg Mg/L, 1,557 and 32.4 mg K/L and 322 and 7.60 mg Na/L. In the fall of 2017, biochar and wood ash were applied at the soil surface at a rate of 1 Mg (dry) ha-1 each, i.e., the lowest rate of biochar at which plant growth (maize) was observed (Glaser et al., 2014) and wood ash at which the acid–base status of the soil was significantly (positively) affected (Brais et al., 2015). The soil treatments are thus believed to be conservative, whereas the combination of wood ash and biochar was used to more completely emulate the expected effect of forest fires on soils. Leaf litter was removed before applying the material and then placed back. Wood ash has a water content of 40%, a pH of approximately 12.3 and a neutralizing capacity of 52%, whereas the biochar was prepared from coarsely grinding wood charcoal prepared from eastern hardwoods using a leaf grinder. Wood ash was sieved at 2 mm before being applied, whereas biochar was applied without sieving. Fertilization was done prior to the next growing season for the soil to react before red oak acorns were planted.
During the last week of May 2018, 12 acorns per micro-plot were planted in the first 5 cm of forest floor. Again, to do so, leaf litter was removed and placed back. The saplings from the greenhouse experiment were then placed in dormancy in the fall of 2018 by progressively reducing photoperiod and temperatures in the greenhouse. As a whole, seedlings had 4 months of growth. These were later transferred and buried under snow in early December 2018 at SBL. Micro-plots were re-fertilized at the same rate prior to the snow. The saplings were then transplanted in the micro-plots in early June of 2019. The IA provenance was also added to the experiment. The acorns were stratified from December 2018 to April 2019 (as described in the previous sections), germinated in pots in May 2019 and then transplanted on the sites in June 2019. The seedlings were transplanted mid-June. Three SBL, four MR, and five PC and IA provenances were transplanted in the micro-plot. In the end, the 3 original provenances were planted in one block in site 1 and one block in site 2, for a total of 48 plots (16 micro-plots per provenance) and 48 SBL seedlings, 64 MR seedlings and 80 PC seedlings, whereas the IA seedlings were planted in two blocks in site 1 and two blocks in site 2, for a total of 32 plots and 160 seedlings.
The effect of provenances and soil fertilization treatments on germination, survival and growth rates were assessed following a schedule with decreasing frequency (from 2 times a week, to once a week and to twice in August). We also monitored herbivory from any signs of browsing damage and activity/movement of small animals within the blocks using a high-resolution bird-watching camera equipped with a motion detection sensor (Wingscapes Birdcam Pro, Moultrie, United States). Measured response variables were survival (%), growth stage, number of leaves and leaf area (cm2). Seedling survival was estimated from the presence/absence of leaves and buds and the stem dryness level. Growth stage was estimated based on the size of the largest leaf of the seedling, ranging from 1 to 5 cm2 (e.g., stage 5 corresponded to the largest leaf at 5 cm2). Number of leaves and total leaf area of each seedling were estimated once in mid-July. Leaf size was assessed using the same template as for the greenhouse experiment. Other response variables derived from data were longevity (i.e., number of weeks alive), mean leaf area (cm2 leaf-1), maximum growth stage, final growth stage and growth stage regression (i.e., difference between maximum and final growth stages).
To better characterize growing conditions during the study period, soil temperature was measured using type-K thermocouples that were manufactured in our laboratory. Six thermocouples were placed within each block (3 in the unprotected area and 3 in the high-fenced exclosures) at a 12 cm depth within the buffers. Temperature was recorded using a thermocouple digital thermometer (Treacable®, Cole-Palmer, United States). Soil volumetric water content (VWC) was measured with time-domain reflectrometry using a FieldScout TDR 300 Soil Moisture Meter (Spectrum Technologies, United States) equipped with 12.2 cm probe. Measurements were repeated 3 times around each thermocouple. Leaf area index (LAI) was measured during full canopy in July to characterize the light environment within each block using a CI-110 Plant Canopy Imager (CID Bio-Science, United States). Measurements were conducted at 1.5 m above the soil surface.
To avoid soil disturbance where acorns and seedlings were planted, side plots with and without fertilization were established as a means to sample soils, i.e., forest floor and upper podzolic B horizon, and thus assess the effects of ash and biochar on soil chemical composition. These side plots were established as one pair of the control, wood ash, biochar and wood ash+biochar treatments beside each experimental block. Forest floor and upper podzolic B samples were collected from each treatment, for a total of 8 soil samples in each block or 80 samples in total.
2.6 Soil analysis
Soil samples collected in the field were quickly brought back to the laboratory where they were immediately dried in an oven at 50°C over 48 h. A portion of the dried samples was used for measuring pH and exchangeable Ca, Mg, K and P, whereas the rest was ground to <60 μm (PM 400 Planetary Ball Mill, Retsch, Germany) for analyzing the bulk chemical composition, including total C and N. Soil pH in water was measured using soil:water ratio of 1:10 for the forest floor and 1:5 for the mineral soil (Hendershot et al., 2008). Exchangeable Ca, Mg, K and P were measured following a Mehlich 3 extraction (Ziadi and Tran, 2007). Calcium, Mg and K concentrations were measured by atomic absorption spectroscopy (55 AA analyzer, Agilent Technologies, United States), whereas P concentrations were measured using the molybdenum blue colorimetric method (SmartChem 200 Discrete Analyzer, AMS Alliance, United States). Total C and N concentrations were measured by combustion at 1040°C and infrared and thermal conductivity detection, respectively (EA 1108 CHNS-O Elemental Analyzer, Thermo Fisons, United States). Ground samples were also pressed in pellets (13 mm diameter × 10 mm thickness) using a 25 tons hydraulic press (Reflex Instruments, Australia).
Plant Root Simulator (PRS) probes (Western Ag Innovations, Canada) were used to assess solution ionic activity of the different soil richness levels in the greenhouse experiment pots following a paste preparation protocol for which the sample paste was placed on the probes for 3 h (Qian et al., 2008). Four pairs of cation and anion probes were prepared for each soil richness level. The paste was then removed and the probes were cleaned with deionized H2O and stored in the fridge in zipseal bags until analysis. Elution of the probes was done for 1 h with 0.5 M HCl. NH4-N and NO3-N were determined colorimetrically by continuous flow analysis (Autoanalyser III, Bran & Luebbe, United States), whereas other ions (H2PO4-, Ca2+, Mg2+, K+, Al3+, Fe3+, and Mn2+) were determined by inductively coupled plasma atomic emission spectroscopy (Optima 3,000-DV, PerkinElmer, United States).
Bulk chemical composition of the upper B horizons collected at the PC, MR, IA and SBL was assessed using a X-ray fluorescence spectrometer (Vanta M series, Olympus, Japan) equipped with a rhodium tube of 50 kV and a silica drift detector. Results are reported using the Geochem calibration mode, with one built-in beam filter operating at 40 kV for heavier elements and another operating at 10 kV for lighter elements. Scanning time was set to 30 s per beam such that one whole scan was completed in 60 s. Analyzer drift was verified every 25 pellets with a silica blank. Soil pH in water and total C and N were also assessed on these samples using the methods indicated above.
2.7 Statistical analysis
All statistical tests were performed in the R environment (R Core Team, 2019). Parametric analysis of variance (ANOVA) tests and multiple regression trees (MRT) were used to test the effects of various factors on response variables. For the field experiment, response variables were converted into percent values due to unbalanced number of individuals between plots. For the greenhouse experiment, parametric ANOVAs and permutational ANOVAs were used to categorize the effects of provenances and soil richness. For the field experiment, ANOVAs were used on the four nested factor levels, i.e., site, protection from large herbivores (with or without high-fenced exclosures), soil fertilization and provenances. In the ANOVA, both provenance and soil richness/fertilization were used as the fixed factors. For the field study, repeated-measures ANOVA was used to compare measured soil temperature and volumetric water content (VWC) between sites in the field study. A Tukey post-hoc test was used to identify factor level differences at an alpha significance threshold of 0.05 for the greenhouse experiment and 0.1 for the field experiment. In the latter case, a higher alpha level was preferred because of the low number of denominator degrees of freedom. We proceeded in this manner by keeping in mind that the consequence of failing to detect a difference which did occur in nature (type II error), which is necessarily bound to the alpha level, is likely more serious than the consequence of detecting a difference which did not occur (type I error) (Peterman, 1990). When the conditions of independence of the observations, normality of the residuals and homoscedasticity were not met, data were transformed with square root or natural logarithm.
In ecology, MRT are used to describe and predict relationships between species data and environmental parameters (De’ath, 2002). In this research, the specific MRT approach that was used is the conditional inference trees variant, which is a non-parametric class of regression trees embedding tree-structured regression models into a well-defined theory of conditional inference procedures (Hothorn et al., 2006). The MRT were produced by the ctree function within the party package (Hothorn et al., 2006) in R. Other R packages used to compute results or produce graphical representations included dplyr (Wickham et al., 2020), ggplot2 (Wickham, 2016), gridExtra (Auguie, 2017), multcomp (Hothorn et al., 2008), and splines (R Core Team, 2019).
3 Results
3.1 Age structure
Red oak and sugar maple were the oldest tree species at both sites (Table 1). One Fraxinus nigra M. tree was the exception in site 1, with a similar age (90 years) to the red oak. Many F. nigra trees were found near the south edge of site 1 (at a lower position on the hillslope) and the old F. nigra tree may thus originate from another stand with a similar disturbance history and composition that borders the southern limit of site 1. Our results suggests that red oak trees in site 1 established sooner than sugar maple trees by an average of 18 years. Conversely, red oak trees at site 2 are on average 7 years younger than sugar maple trees. As a whole, our results suggests that red oak is relatively close in age to sugar maple, and perhaps older at site 1, but our data do not account for possible prolonged suppression of sugar maple seedlings in the understory.
Very few trees were older than 1906. At that point in time, there was a substantial increase in tree recruitment rates at both sites. Tree recruitment at site 1 mainly occurred during two periods, i.e., 1910 to 1924 and 1951 to 1965. At site 2, recruitment dynamics were not as well defined in time. Recruitment rates were highly variable between 1907 and 1980, shifting from high to low, and they were nearly zero thereafter. Red oak recruitment occurred mainly between 1910 and 1933 at site 1, with a record year in 1918. Sugar maple recruitment was high between 1918 and 1930 and it was irregular and more largely spaced in time between 1930 and 1978. No more recruitment of sugar maple occurred thereafter at site 1. There was steady but small recruitment of red oak at site 2 between 1900 and 1931, whereas high recruitment occurred between 1947 and 1960. Sugar maple recruitment at site 2 mostly occurred between 1907 and 1947. Recruitment of sugar maple was negligible thereafter and completely absent from 1970 and onward.
3.2 Greenhouse experiment
Ionic activities of the soil substrates as measured by paste extracts on PRS probes confirm a strong increase in the availability of macronutrients for red oak seedlings from the low to average to high soil richness treatments (Supplementary material C). The only exception was for NH4-N which suggests relatively homogeneous supply rates between the 3 treatments (Figure 2).
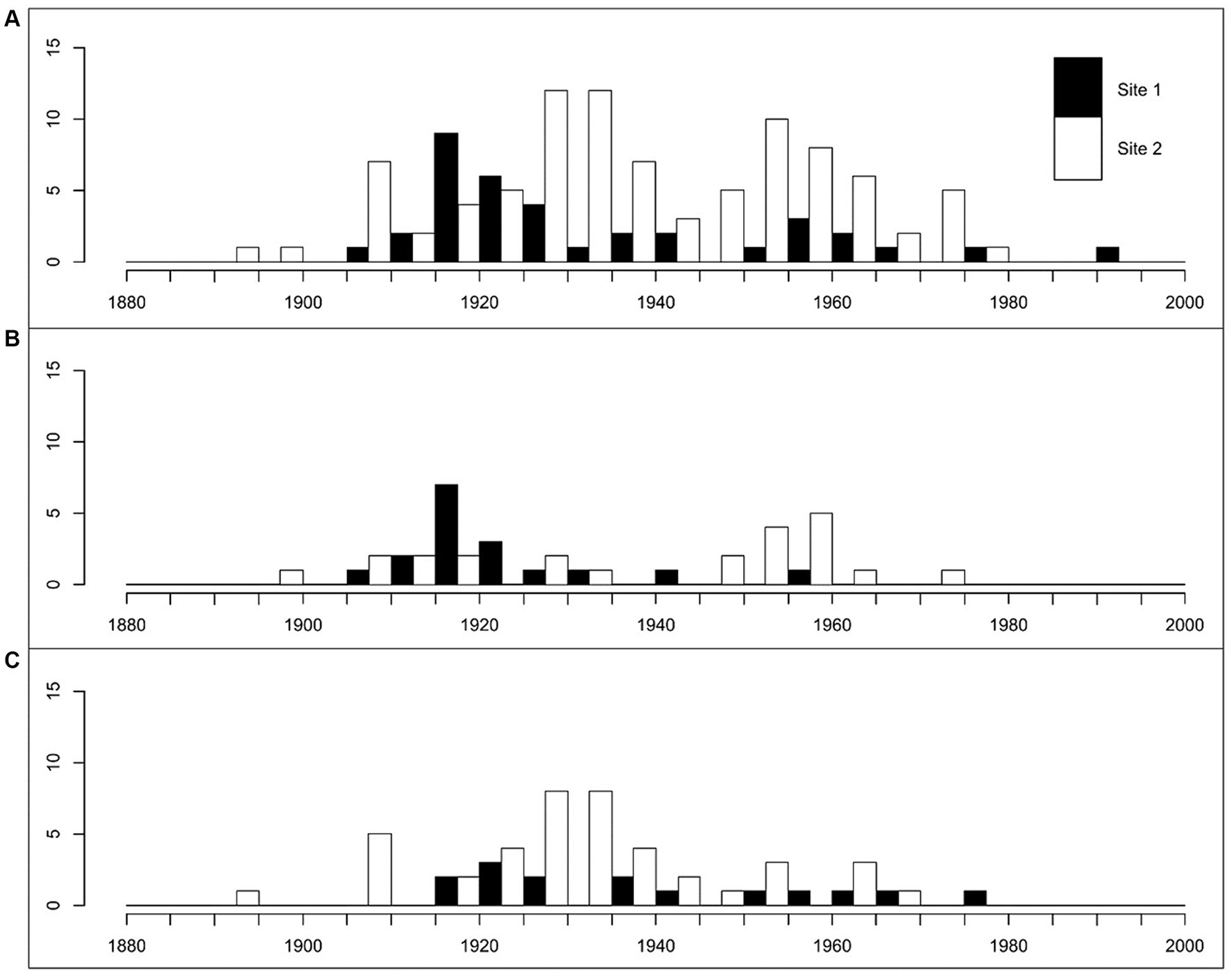
Figure 2. Age structure of all cored trees (A), red oak (B) and sugar maple (C) at the two study sites. Data are only accounting trees for which growth rings could be counted in whole. Trees with rotten or empty cores are not accounted for.
Germination rates of all red oak acorns increased by about 12% from the low soil to the high soil richness treatments (Table 2). Germination rates also increased by provenance in the following order: SBL < MR < PC (Table 2). Sprout survival rates of all red oak provenances were highest under average soil richness and lowest under low soil richness. Sprout survival rates were similar between SBL and MR, whereas PC showed higher rates (Table 2). The MR and PC provenances showed an increase in germination rates with increasing soil richness. On the other hand, the SBL provenance exhibited the greatest germination rate under low soil richness (i.e., 50%), which was a lower germination rates than that observed for the MR and PC provenances under low soil richness. The total mortality rate for the SBL provenance was highest under high soil richness, whereas mortality rates for the MR and PC provenances were highest under low soil richness and lowest under high soil richness. The only provenance that did not suffer post-germination mortality up to the 10th week of experimentation was PC within the average treatment (Table 2). The MR provenance also displayed the largest differences in germination, sprout survival and total mortality rates between the low and high soil richness treatments.
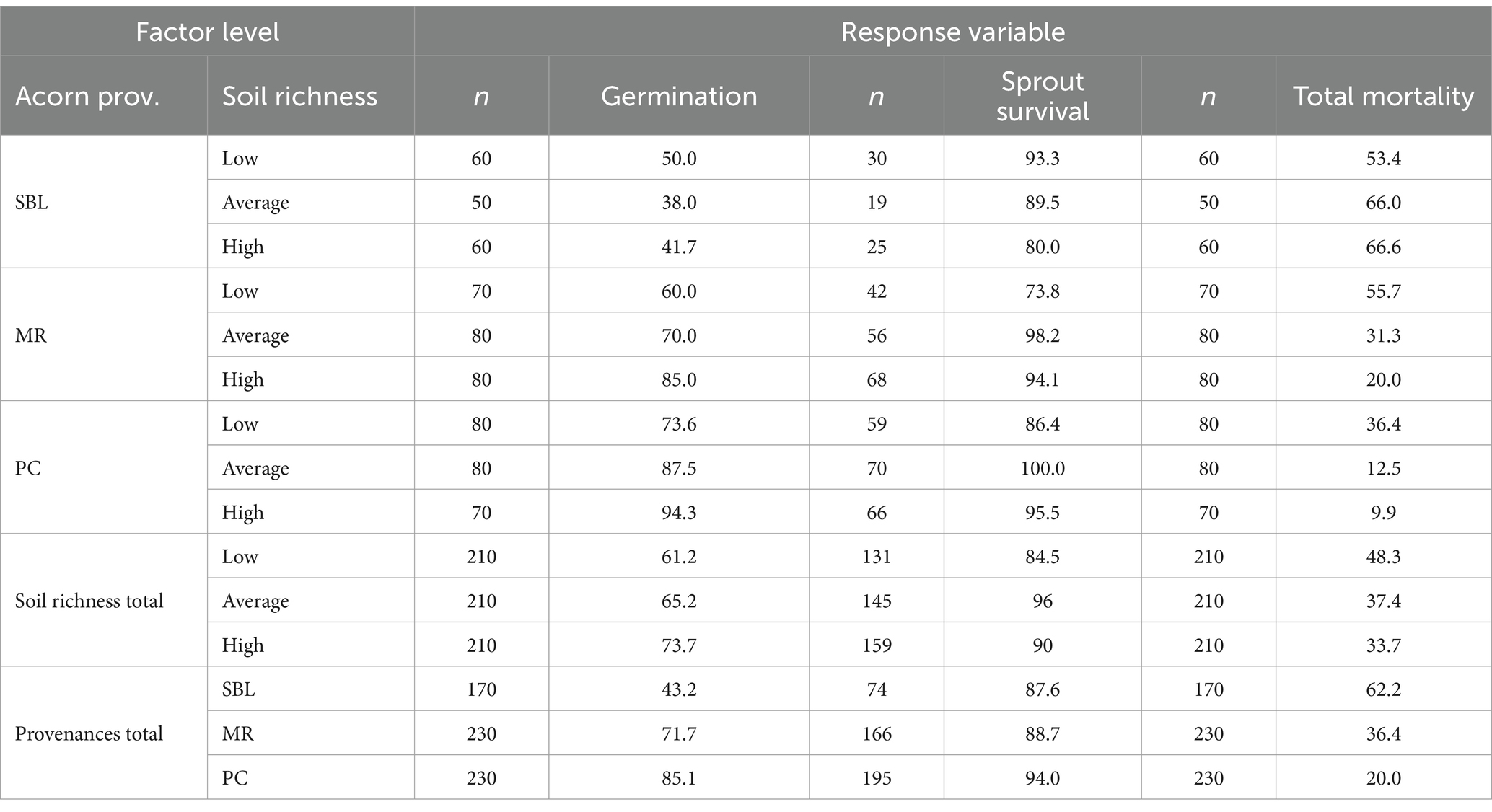
Table 2. Germination, sprout survival and total mortality rates (%) of red oak provenances as a function of soil richness over 10 weeks in the greenhouse.
The MRT analysis on germination rates suggests a first partition within the provenance factor, with the SBL provenance being separated from the MR and PC provenances (Figure 3). A second partition is suggested within the treatment factor for the MR and PC provenances, with the low soil richness treatment being separated from the average and high soil richness treatments. A third partition separates the MR provenance from the PC provenance, whereas a final partition separates the average soil richness treatment from the high soil richness treatment for the MR provenance (Figure 3). From the resulting MRT analysis, it can be assessed that SBL differs most from MR and PC, regardless of fertility treatments, and that the difference between low and average soil richness treatments had a higher impact on MR and PC survival than the difference between average and high soil richness treatments. The final partitions show that PC survival is mostly unaffected by an increase in soil nutrient availability beyond the average treatment, while MR shows a slightly improved survival rate.
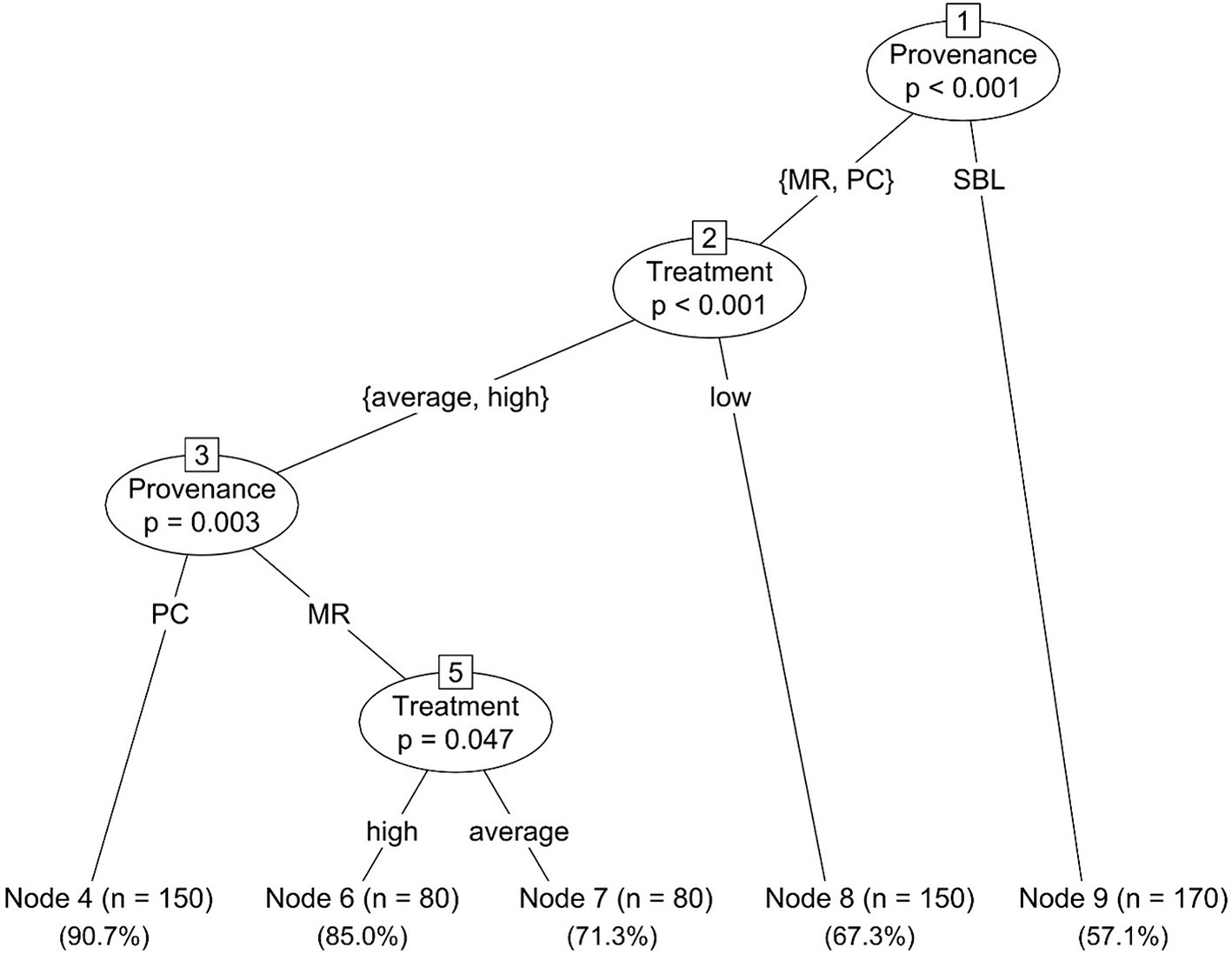
Figure 3. Multiple regression tree of red oak germination rates (as a yes/no binary variable) as function of soil richness and acorn provenance over 10 weeks of greenhouse experimentation. Associated germination rates are specified under each node.
Morphological variables are presented in Supplementary material D. As a whole, the high soil richness was associated with greater height, foliar surface and leaf, stem and root mass, whereas the low soil richness treatment exhibited the lowest values. The average soil richness treatment did exhibit slightly higher stem mass than the high soil richness treatment. Root/shoot ratio were about 0.8 under low and average soil richness, while the ration was about 0.7 under high soil richness. Two-way parametric and permutational ANOVA tests suggest that total leaf area and mass as well as stem mass were significantly different among soil richness treatments, among provenances, and that there was a significant interaction between the two factors (Supplementary material E). Boxplot analysis of these variables (Figure 4) indicate that this interaction is the result of the SBL provenance. While performance of the MR and PC provenances generally increased with soil richness, the SBL provenance performed at its maximum under average soil richness and performance substantially decreased under high soil richness. ANOVA tests also suggest a significant difference in stem height between provenances, in support root mass between soil richness treatments and between provenances, and in ratio of fine roots mass/support roots mass and fine roots mass/leaf mass between treatments (Supplementary material E).
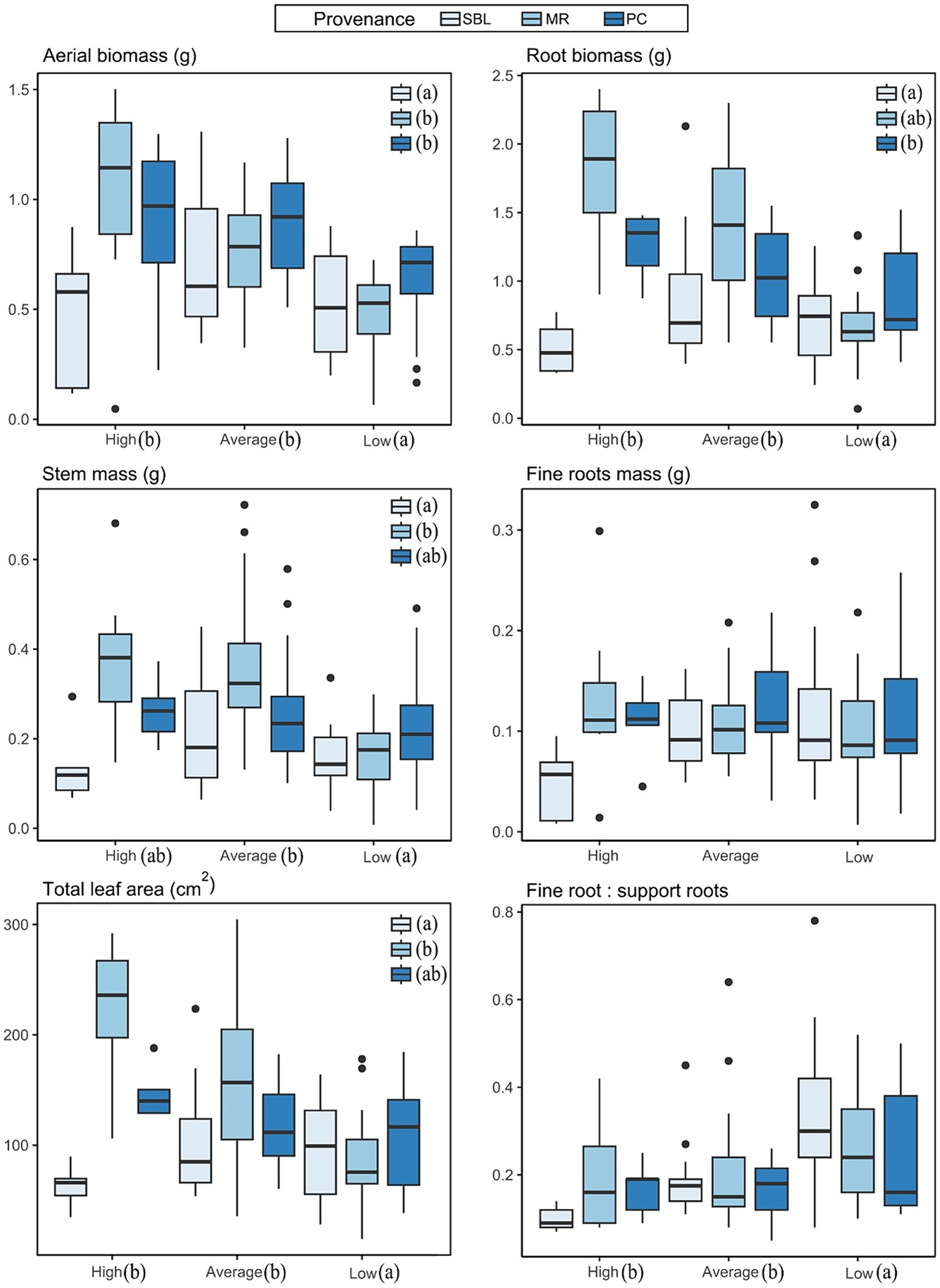
Figure 4. Boxplots of response variables measured in the greenhouse (see Supplementary material D for full data table). Letters indicate significant differences (p < 0.05) between treatments and between provenances.
3.3 Field experiment
For the 2018 field experiment, we observed 100% mortality of planted acorns after a two-week period. Mostly heavy rodent predation (mainly eastern chipmunk, Tamias striatus L.) but also very dry conditions were fatal to all planted red oak acorns. No other testing/results are thus reported here.
Chemical analyses show that forest floor composition is generally similar between the sites (Supplementary material B). However, site 1 has higher mineral soil total C and N concentrations as well as higher extractible Ca, Mg and K concentrations, whereas soil pH is lower. Ash and biochar fertilization treatments had a substantial positive effect on forest floor Ca, Mg and K concentrations and pH but had no detectable effect on the mineral soil. Wood ash application led to a substantial increase in forest floor pH and Ca, Mg and K concentrations. Biochar only slightly increased forest floor pH and Ca, Mg and K, whereas the combination of ash and biochar showed the largest increase (Supplementary material B). There is an inverse relationship between soil VWC and temperature, i.e., VWC decreasing with an increase in temperature (Supplementary material F). Soil temperature and VWC were, respectively, highest and lowest in August. Site 1 showed slightly higher soil temperatures and lower soil VWC than site 2 throughout most of the summer. Leaf area index averaged 1.9 and 2.4 at sites 1 and 2, respectively. This difference is likely associated with variation in tree density (as expressed in basal area) between the sites (Supplementary material A).
For the 2019 field experiment, there was a large difference in the overall (i.e., all provenances) planted seedling survival rate between the two sites throughout the experiment (Figure 5A). At the last survey, seedling survival rate was 45% in site 1 and 74% in site 2. The difference in survival rate widened between the two sites as early as June 16th and averaged 30.5% for the remainder of the survey. Seedling survival rate also displayed a similar overall pattern at both sites. Independent of sites, seedlings planted in high-fenced exclosures (protected) showed a final survival rate of 65.8% while unprotected seedlings showed a survival rate of 53.2% (Figure 5B). The difference between the protected and unprotected seedlings widened on June 20th but it stabilized at the next survey, i.e., June 26th. Thereafter, survival rate between protected and unprotected seedlings followed a similar pattern and the difference between the two averaged 11.9% for the remainder of the survey.
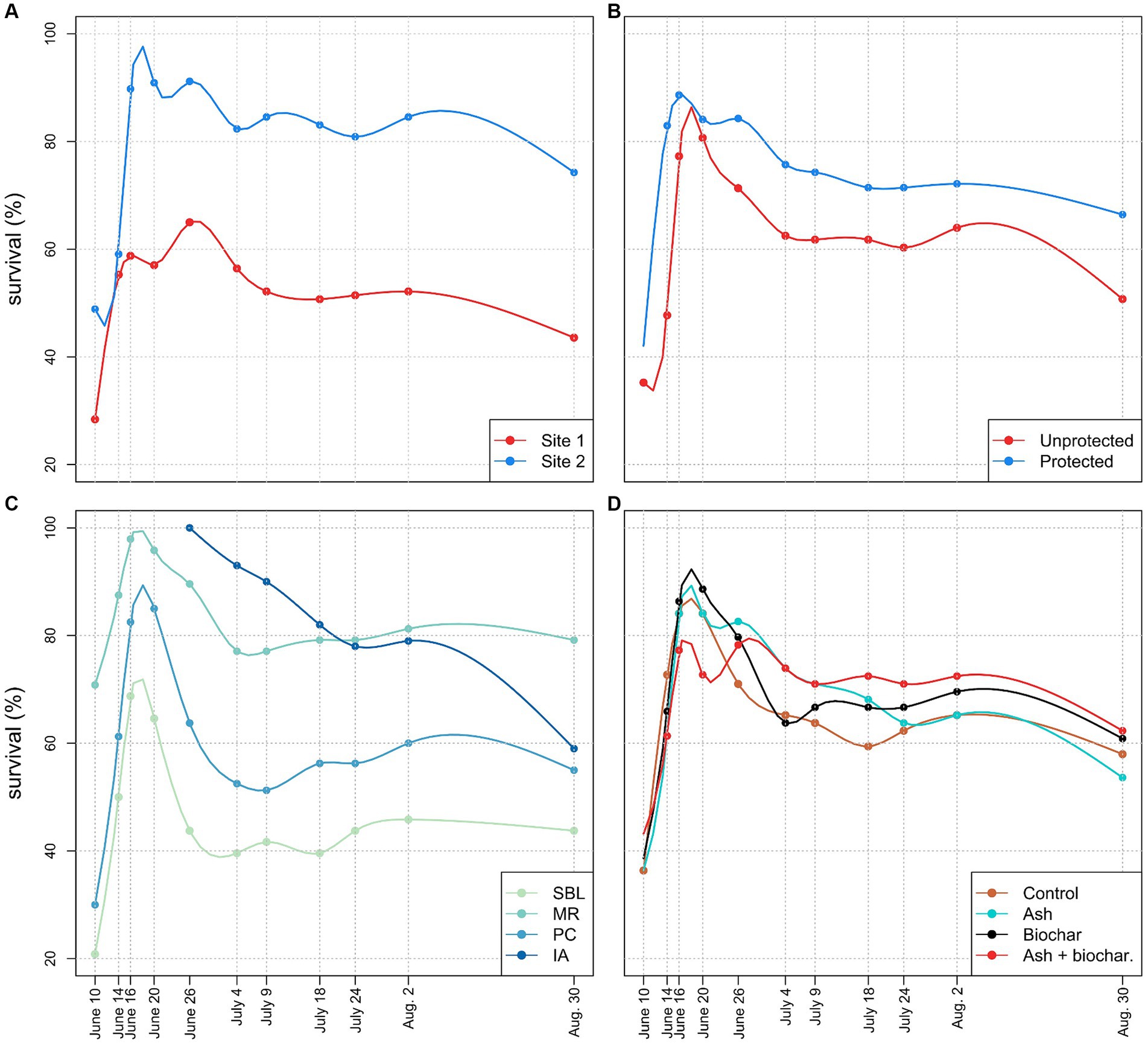
Figure 5. Red oak seedling survival rates in the experimental plots at SBL during the 2019 survey as a function of site (A), protection against large herbivores (B), acorn provenance (C) and soil fertilization (D). The initial rise in survival rates is the result of some seedlings showing vitality (i.e., leaf development) several days after transplantation.
Seedling survival rate largely varied as a function of acorn provenances (Figure 5C). At the end of the survey, seedling survival rate was highest for the MR provenance (79.2%), followed by IA (59.7%), PC (55.0%), and SBL (43.8%). Survival rate of the MR, PC, and SBL provenances widened early in the survey (between June 14th and 16th) and differences were then maintained for the rest of the survey. While seedling survival rates stabilized by July 4th, survival rate of the IA provenance decreased steadily throughout the survey. However, a short respite in mortality was observed between July 24th and August 2nd. Seedling survival rates displayed smaller variation among soil fertilization treatments (Figure 5D). The highest survival rate was observed under the ash + biochar treatment at 63.6%, followed by the biochar treatment at 61.6%, the control treatment at 58.8% and the ash treatment at 54%. Overall, seedling survival patterns were similar among fertilization treatments. Although the wood ash treatment initially performed strongly, i.e., 84.1% survival rate between June 16th and 20th, it underperformed relative to the other treatments thereafter. Conversely, the ash + biochar treatment exhibited the lowest survival rate early in the survey, but it was ultimately the treatment with the highest survival rate at the end of the experiment.
The one-factor ANOVA tests show that site had a significant effect on the seedling survival of the SBL, MR, and PC provenances as well as all other growth development variables measured (Tables 3, 4). All data indicate that seedlings at site 2 outperformed seedlings at site 1 in regard to survival and growth development. Protection against large herbivores led to higher seedling survival, mean area per leaf and stage regression. A higher stage regression was observed for protected seedlings, which means that seedlings exhibit a larger difference between the maximum and final growth stage achieved. The IA seedlings at site 2 also outperformed seedlings at site 1 in regard to mean area per leaf, final growth stage reached and survival time (Tables 3, 4). Seedling survival rate was marginally significant at p = 0.1 (i.e., 14.0% higher at site 2 vs. site 1). The IA provenance showed higher but non-significant survival rates in protected plots than in unprotected ones, while significantly higher maximum and final stages reached highlight the effectiveness of the high-fenced exclosures for IA seedling growth. Seedling protection against large herbivores led to significantly lower maximum and final growth stages reached, which is contrary to SBL, MR, and PC seedlings.
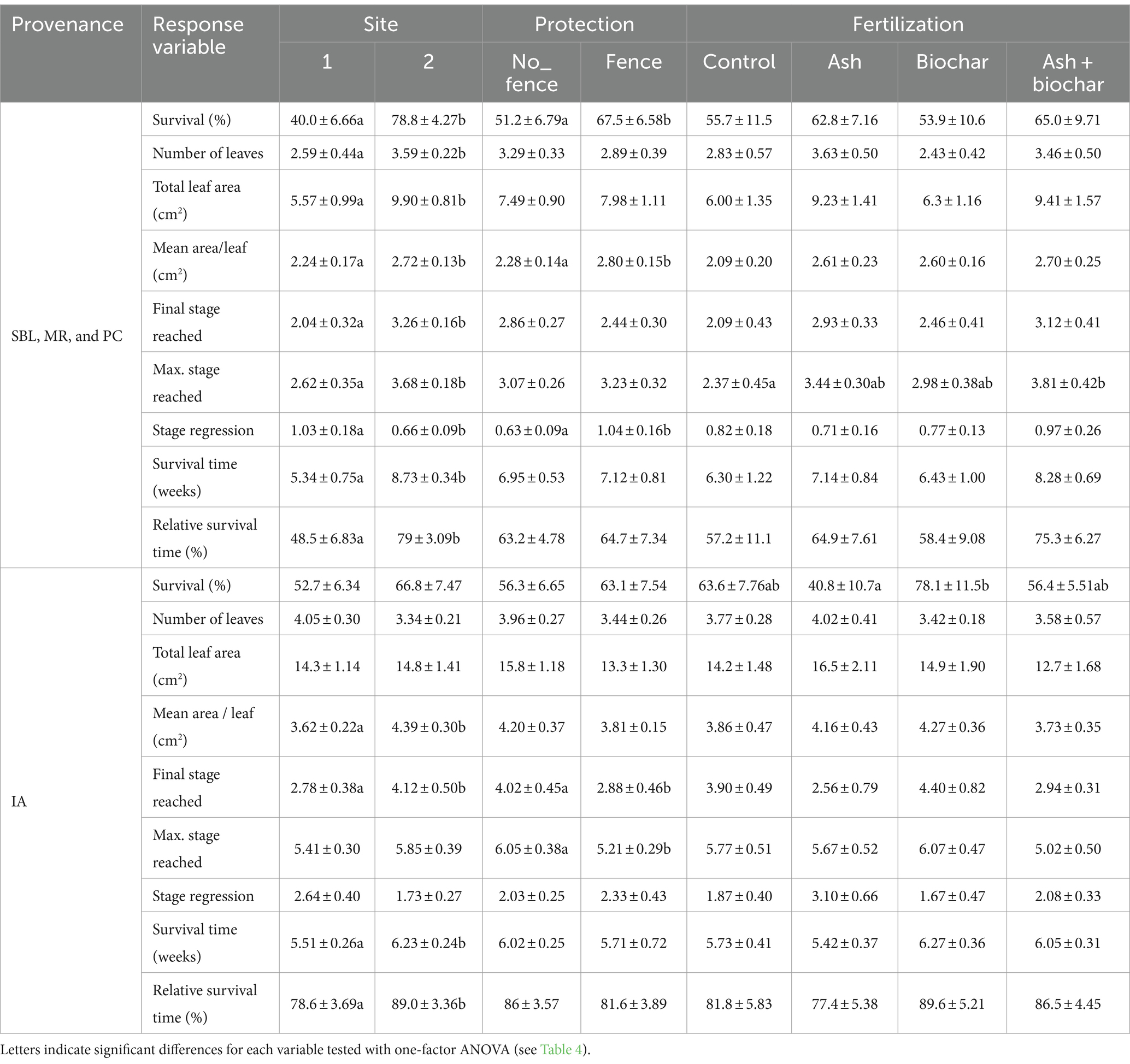
Table 3. Mean ± standard errors of seedling survival and growth development variables measured as a function of site, protection against large herbivores and soil fertilization.
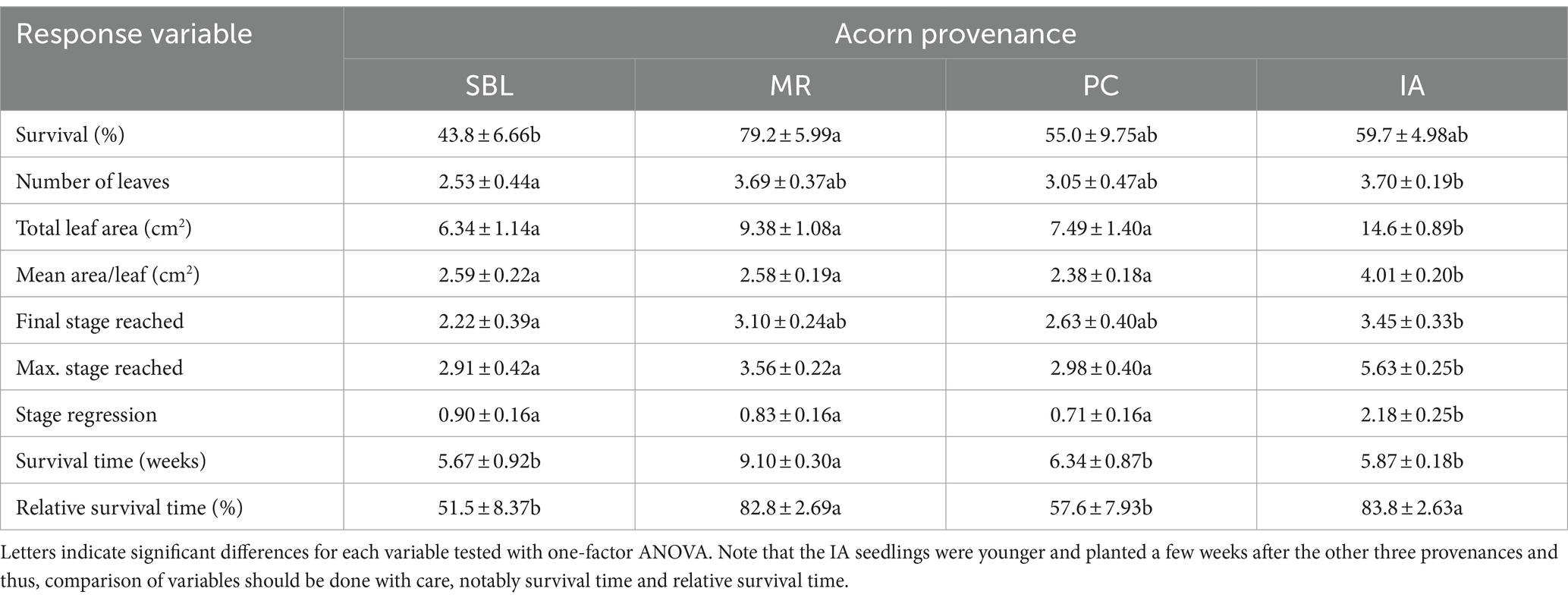
Table 4. Mean ± standard errors of seedling survival and growth development variables measured as a function of acorn provenance.
The SBL seedlings exhibited the lowest values for most response variables (e.g., survival rate, number of leaves, final growth stage reached and relative survival time) measured compared to the other provenances (Table 3). The MR provenance exhibited greater performance than the SBL and PC provenances in terms of survival, number of leaves, total leaf area, final and maximum stage reached, survival time and relative survival time, but only survival time and relative survival time were significantly higher than the SBL and PC provenances (Table 3). The IA provenance exhibited a relative survival time similar to the MR provenance, whereas survival rate was 59.7% and not statistically different from the SBL and PC provenances. An increase in mortality of the IA provenance occurred near the end of the survey (Figure 5C). Yet, the IA seedlings exhibited significantly greater total leaf area and final and maximum growth stage reached (Table 3).
Soil fertilization significantly affected the maximum stage achieved by the SBL, MR, and PC seedlings, with an average of 2.37 under the control, 3.44 under ash, 2.98 under biochar and 3.81 under ash + biochar (Tables 3, 4). Soil fertilization had a significant effect on survival rates of IA seedlings, with lower (40.8%) and higher (78.1%) survival under the ash and biochar treatments, respectively (Tables 3, 4).
The MRT analyses captured survival and growth development stages of all four provenances simultaneously (Figure 6). The regression tree of survival rate highlights the effects of site and provenance (Figure 6A). A first separation was made between sites, with site 2 exhibiting higher survival rate than site 1. A second separation was made at site 1 between the provenance groups of MR–IA and SBL–PC. The MR–IA groups had higher survival rate than the SBL–PC group. The regression tree of relative survival time also displays the effects of site and provenance (Figure 6B). However, a first separation was made between the acorn provenance groups of IA–MR and SBL–PC. The IA–MR group exhibited higher relative survival time than the SBL–PC group. A second separation distinguished the two groups into site 1 and site 2. Relative survival time was higher for both groups at site 2. The regression tree of maximum growth stage reached (Figure 6C) and total leaf area (Figure 6D) led to a first separation between the IA provenance and the provenance group SBL–MR–PC. The SBL–MR–PC group was then subdivided into site 1 and site 2. The IA provenance exhibited the highest maximum stage reached and total leaf area. For the SBL–MR–PC group, maximum growth stage reached and total leaf area were higher at site 2.
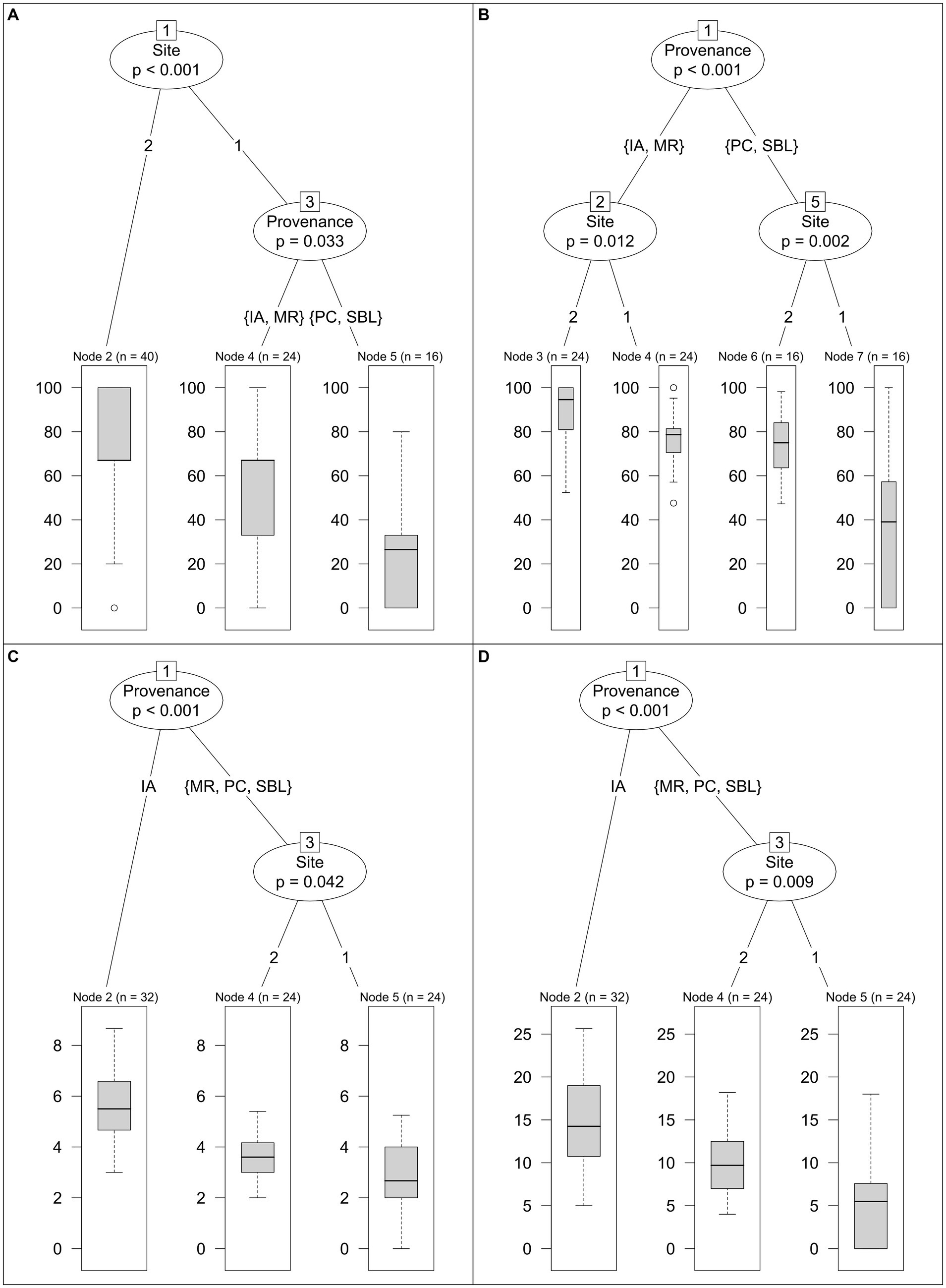
Figure 6. Multiple regression tree of red oak survival rate (A, in %), relative survival time (B, in %), maximum stage reached (C, classes 1–8) and total leaf area (D, in cm2) as function of site and acorn provenance. Other factors tested such as protection against large herbivores, soil fertilization and blocking within site were not significant at p < 0.05 and were thus excluded from the analyses.
4 Discussion
The series of experiments conducted in this study have helped portray the main factors responsible for red oak regeneration at SBL. Based on the age of red oaks that match the last known fire occurrence, its seems likely that fire has played a significant role on oak regeneration in the past. Fire suppression from the landscape in the last 50 years may also have led to a series of regeneration failures of red oak. The oldest trees at sites 1 and 2 date back to 1920, which points to a wildfire affecting much of the SBL area (Savage, 2001; Bélanger et al., 2004). While both red oak and sugar maple are the dominant tree species at the sites, the cohort recruitment pattern at each site are different. On the one hand, site 1 shows a greater dominance in red oak, mostly established between 1920 and 1940, with some subsequent regeneration from competing sugar maple and greater overall red oak biomass. On the other hand, site 2 exhibits a more concurrent and longer establishment period of red oak and sugar maple trees, the former showing a lower frequency but demonstrating a clear growth advantage from the initial cohort establishment. The red oak trees are, however, smaller in size in comparison to site 1, thus also resulting in lower biomass. Fire disturbance severity, frequency and timing may have played a role in the resulting tree species composition and age structure of these red oak stands (Crow, 1988; Abrams, 1992). Brose (2010) suggested that red oak regeneration is dominant after a medium or high intensity burn applied during either spring or summer, while red maple and yellow poplar regeneration dominate plots after low intensity burns, regardless of timing. In this regard, observed differences in stand tree species composition and density between sites 1 and 2 are likely the result of a divergence in fire severity. This is also in alignment with the idea that the regenerative strategy of red oak is mainly derived from its sprouting ability (Sander, 1990) rather than a physical resistance to fire (in comparison to other oak and hardwood species), with higher fire severity advantaging red oak regeneration because it eliminates mature trees of competing species.
Establishment success of red oak in Europe in the absence of fire as well as situations where the species is considered invasive (Riepšas and Straigytė, 2008; Langmaier and Lapin, 2020) suggest that many other factors govern red oak regeneration. Explanations for this include lower predation from local fauna and higher shade tolerance than indigenous oak species (Major et al., 2013; Nicolescu et al., 2020). In the prolonged absence of fire at SBL, our study highlights that site conditions, namely water availability, genetics (provenance) and control of herbivory are key factors governing (in confounding ways) germination, survival and early growth of red oak.
4.1 Site characteristics
Site was found to be the most significant factor controlling planted red oak seedlings survival and growth differences in the field experiment. Seedlings at site 2 performed better than site 1 for all response variables tested. In this respect, stand characteristics may well provide insights as to the main drivers of red oak regeneration at the sites. Site 2 exhibits much greater tree species diversity and density but lower basal area. Fei et al. (2003) found a strong relationship between cover species and density of red oak regeneration, with a steep decrease under any other species than itself. This somewhat differs from this study’s results. As such, it could be argued that the diversified canopy cover at site 2 was beneficial to red oak seedling survival and growth during periods of low water availability.
Although red oak saplings and mature trees are resistant to water stress, seedlings are still vulnerable to such pressures, especially prior to extensive root system development (Kolb et al., 1990; Bauweraerts et al., 2013). For the most part of the 2019 summer, soils at site 1 were warmer and drier than soils at site 2. This difference may be explained by higher density of American beech regeneration at site 2, which is associated with lower light transmission (Collin et al., 2017, 2018), resulting in lower soil temperature and higher soil moisture (Bélanger et al., 2021a). During field measurements, symptoms of water stress such as dried leaves were often observed on experimental seedlings before their death, notably at site 1, which concurs with the explanation of red oak seedling mortality due to water constraints. Kleiner et al. (1992) have also proposed that red oak seedling shoot growth can be negatively impacted by water stress. The 2019 summer at SBL was slightly warmer and drier than normal, especially in May and June (Ministère de l’Environnement et de la Lutte contre les changements climatiques, 2021a), suggesting that water stress may have affected survival and growth that specific year. Germination and seedling establishment will likely be a critical phase under climate change, but red oak seedlings should become less vulnerable to water stress as they develop their root system (Crow, 1988; Sander, 1990). Repeated measures ANOVA suggest that soil temperatures and water volumetric contents measured in 2019 changed significantly throughout the whole season, although no significant difference was detected between sites. In this respect, it is not possible to conclude with high certainty that water stress was responsible for the differences in seedling performance between sites 1 and 2. Other overstory effects could also have been important and acting in interaction with soil water availability.
Measurements of LAI suggest 20.6% more leaf surface area in site 2. We suspect tree species composition, notably higher American beech regeneration, is responsible for this difference. Messier and Bellefleur (1988) have shown that spectral irradiance of the forest floor (W/m2) is lower under American beech than under sugar maple and yellow birch at climax stage. Similar results were reported by Collin et al. (2018) at SBL. Furthermore, Canham et al. (1994) found that light is at <2% of full sun under American beech canopy, while it is at >5% under red oak canopy. Our results at site 2 thus suggest that lower light availability does not negatively impact the performance of red oak seedlings. Conversely, Phares (1971) observed that while red oak seedlings accumulate nutrients under heavy shade (e.g., 10% of light availability), height growth is triggered only at a light availability threshold of 30% light or more, and the highest biomass increments were measured at full sunlight. Brose and Rebbeck (2017) showed similar red oak seedling survival rates (15–75% from lowest to highest available sunlight), although height growth and root collar development were substantially impaired below 40% available sunlight. This implies that higher light availability at site 1 is most likely not responsible for poor seedling performance and corroborates the idea that soil water availability or other overstory effects on microclimate may have played a more significant role in early seedling survival and growth.
4.2 Genetic adaptation
Provenance had a significant impact on germination and growth in the greenhouse as well as seedling survival and growth in the field. This is especially important to highlight as acorn and seedling stock quality are significant factors to consider for red oak seedling development (Ward et al., 2000; Lombardo and McCarthy, 2009).
Contrary to the MR and PC provenances, the SBL provenance underperformed under the higher soil richness level in the greenhouse. Interestingly, the SBL provenance also underperformed in the field experiment compared to the other provenances, whether fertilized or not. McGee (1974) found that seed source elevation can affect seedling budbreak, bud and leaf development and height. Furthermore, Abrams (1994) found that adjacent red oak stands can exhibit distinct genotypes as a result of contrasting microenvironments. Notably, they observed that differences in water availability and drought effects within a limited geographical range created differences in net photosynthetic rates and leaf water potential within the red oak provenances studied.
Two explanations are thus proposed for the poor performance of the SBL provenance. First, of the acorns collected for the germination and seedling studies, acorn infection rates by curculio weevils were highest for the SBL provenance at 77%, whereas the MR and PC provenances had much lower infection rates at 25 and 22%, respectively. Furthermore, damage to the cotyledon was shown to negatively impact red oak acorn germination rate and seedling performance (Yi et al., 2019) because germination and early growth of red oak seedlings are highly dependent on acorn nutrient reserves (Yi and Wang, 2015) since SBL had the smallest average acorn size (3.29 ± 0.07 cm3 for SBL, 3.47 ± 0.06 cm3 for PC and 4.10 ± 0.06 cm3 for MR). It is thus possible that a lower performance of the SBL provenance seedlings in the experiments could be due to an overall lower quality. Second, the poor performance of SBL acorns in the greenhouse could indicate that this provenance was genetically selected to perform better under more acidic and nutrient-poor soil conditions. Acorn collection sites for MR and PC red oaks show higher overall soil nutrient richness (N, P, Ca, Mg) and pH in comparison to SBL and IA, which corroborates with the idea that provenances adapt to specific soil conditions.
In addition, the MR provenance, which is the southernmost provenance, exhibited the highest survival rate in the field experiment by as much as 33% in comparison to the other provenances. For one, the MR provenance has performed better possibly because of greater acorn nutrient reserves that would have been inherited from the highest soil quality at that site. Also, southern provenances are expected to grow sooner in the spring, and this timing would have corresponded to more suitable growing conditions at SBL (e.g., cooler and wetter). Its better performance may also have been favored compared to other provenances by the specific meteorological conditions prevailing during the 2019 summer (i.e., warmer and drier than normal, Ministère de l’Environnement et de la Lutte contre les changements climatiques, 2021a). Specifically, the MR provenance was possibly able to deal better with water stress despite limited root development. Our results in respect to the MR provenance are in support with the general idea that there will be a northern shift in the distribution of red oak in northeastern North America (Iverson et al., 2019; Prasad et al., 2020).
Red oaks, despite spanning a vast geographic range in North American forest landscapes, experience habitat fragmentation due to increasing selection pressures driven by climate change (Iverson and Prasad, 2002). As a result, this species may benefit from assisted migration to mitigate the negative effects of population fragmentation and regeneration failure on certain sites (Rauschendorfer et al., 2022). In a study investigating the effectiveness of assisted migration at the northern edge of red oak distribution, Etterson et al. (2020) found that southern ecotypes exhibited both greater survival and growth compared to their northern counterparts. This finding aligns with the results of our study, which also support the concept of selecting appropriate provenances based on climate analogs (Dessai et al., 2005; Salinger et al., 2005) within the context of assisted migration (Pedlar et al., 2012; Williams and Dumroese, 2013). However, conducting similar research over multiple planting years will still be necessary to further elucidate and understand how ecotypes will cope in the long term under future climate conditions.
4.3 Predation and herbivory
Acorn predation and leaf herbivory had significant impacts on red oak regeneration. The most severe impact was with acorn predation. We believe the failed germination study in 2018 was mostly due to high predation. Regardless of provenance, acorns were almost entirely consumed by local fauna, mainly chipmunks (Tamias striatus), when planted in May 2018 as we observed an abundance of empty acorn residues on the ground. Many pictures of eastern chipmunks, white-tailed deer (Odocoileus virginianus Zimm.), squirrels (Sciurus spp. and Glaucomys spp.) and North American porcupine (Erethizon dorsatum L.) foraging red oak acorns and seedlings were captured in the plots (Lebel-Desrosiers, 2021). The 2018 summer was also one of the warmest in record (Ministère de l’Environnement et de la Lutte contre les changements climatiques, 2021b), and this has likely participated in the desiccation of acorns. Second, browsing impacted seedling survival for SBL, MR and PC provenances, but only affected overall growth marginally. Blossey et al. (2019) showed a substantial difference in red oak seedling growth rate induced by browsing, whereas Buckley et al. (1998) found that the removal of plant competitors can enhance the pressure from deer browsing on red oak seedlings. Thus, browsing can be of significant importance for red oak regeneration, and its apparent effect on growth may increase after several growing seasons. This idea is also corroborated by Long et al. (2012) who observed a positive and increasing effect of deer browsing protection on height and root collar development of red oak seedlings over 5 years.
4.4 Soil nutrient availability
Soils are often identified as a main descriptor of site quality and suitability for given species (Hulshof et al., 2020) and red oak specifically (Nowacki et al., 1990; Nowacki and Abrams, 1992). The literature regarding red oak describes the species as thriving on nutrient poor, acidic, uphill mesic sites (Crow, 1988; Sander, 1990), a description that suits both study sites at SBL. As a whole, sites 1 and 2 exhibited similar acid–base status in the forest floor, but the mineral soil at site 1 showed slightly higher total N and exchangeable base cation concentrations. These results suggest that soil nutrients had minimal effect on red oak seedling metrics measured in the field experiment, which may be explained by the presence of too many confounding factors that hold greater limiting effects on its regeneration. For one, water limitations in 2019 likely overshadowed the effect of soil nutrient supply and fertilization on red oak seedlings. The fact that ash and biochar fertilization have, for the most part, not shown a significant increase in seedling survival and growth at either site reinforces the idea that soils with higher pH and nutrient availability, as seen for the forest floor at both sites following ash and biochar application, are not more conducive to red oak survival and growth. This is opposite to the results of the greenhouse experiment that show a positive effect of soil nutrient availability on germination, growth and survival of red oak seedlings. However, the greenhouse experiment was conducted under ideal growing conditions regarding light and water availability as well as air temperature, and without predation and disease.
It should be considered, however, that the absence of a response of red oak seedlings to ash and biochar fertilization could be due to a delay for nutrients to be absorbed by the roots. Several studies show a delayed response of plants to ash fertilization (Reid and Watmough, 2014; Bélanger et al., 2021b). Root length and radius, or more specifically total root surface area, influence plant nutrient uptake (Barber and Silberbush, 1984). As such, the underdeveloped root system of the planted seedlings likely had a low capacity for absorbing the nutrients added from the ash. This could also stem from the fact that while the benefits of ash fertilization on the forest floor occurs quickly, the response of the mineral soil is slower as it takes more time for the nutrients to migrate further down and react with soil exchangeable surfaces (Reid and Watmough, 2014, Brais et al., 2015). Schuler and Robison (2010) showed that fertilization had very little positive effects on red oak seedling survival and growth, even in the case where trees competed with other plants for nutrients and other resources. Furthermore, Kolb et al. (1990) showed that, at higher soil fertility and in the absence of light or water stress, yellow poplar performs better than red oak. Thus, since red oak is outpaced by late-successional or more shade-tolerant competing species, regeneration of the species may not benefit much from higher soil nutrient availability if other resources are limited. Fertilization may provide more benefits in soils with marginal fertility where competing plant species are potentially suppressed by the intrinsically low nutrient supply.
5 Conclusion
This study of red oak at the northern limit of its distribution showed that the species remained dominant in the two studied stands throughout the 20th century but is now showing regeneration failure. Stands appear to be shifting toward more shade-tolerant, late successional species such as American beech. Results also suggest that fire played a role in the establishment of red oak and succession of sugar maple, with red oak stand age structure dating to the last known fire disturbance occurrence. Although red oak trees were dominant in both stands, variability in tree species composition and density between stands suggests that regeneration dynamics differed substantially. This result may be due to differences in pre-fire species composition, fire frequency/severity and site physical characteristics such as slope direction and aspect. The study also displayed the importance of other factors in the regeneration process of red oak trees at this latitude. Site was found to have a significant impact on germination, survival and growth of seedlings. The site with higher LAI showed greater regeneration success, although more research is needed to assess if this was solely a result of lower water stress or other overstory effects on understory microclimate. Acorn provenance was also another important factor affecting red oak regeneration. The findings of this research therefore display the need to better characterize the response of red oak to site quality while considering the genetic signatures for optimizing success at early regeneration stages. Furthermore, parasites such as weevils and borers and foraging by small mammals, mostly rodents, were found to have large effects on regeneration at the acorn stage, while deer browsing was found to significantly affect survival, and growth to some extent, at the seedling stage. Finally, soil nutrient availability affected germination, survival and growth positively only when other factors (e.g., water and light) were not limiting. As a whole, this study highlights the need to select sites with adequate water availability, use provenances with proper future climate analogs, and protect seedlings against herbivory to maximize regeneration success, though further testing is required to fully assess the effect of water availability and associated thresholds for red oak seedling and sapling survival and growth. Fertilization with wood ash will likely only be beneficial if other conditions are met.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because observations were made using a camera only.
Author contributions
SL: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Software, Visualization, Writing – original draft, Writing – review & editing. AC: Methodology, Validation, Writing – review & editing, Data curation, Investigation, Software, Visualization. NB: Methodology, Validation, Writing – review & editing, Conceptualization, Formal analysis, Funding acquisition, Project administration, Resources, Supervision.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. Financial support was provided to N. Bélanger by the Natural Sciences and Engineering Council of Canada (NSERC Discovery grants: RGPIN 2015–03699 and 2020–04931), together with a grant from the Canada Foundation for Innovation John R. Evans Leaders Fund (35370). This project also benefited from three Undergraduate Student Research Awards (USRA) by NSERC and USRA supplements by the Fonds de recherche du Québec – Nature et technologies.
Acknowledgments
Authors would like to acknowledge Benjamin Leduc, Zachary Déziel, Charlène Mélançon and Justin Bélanger for their help in the field and laboratory. We also thank the Center for Forest Research for providing access to the greenhouse infrastructure at UQAM. Finally, we thank the staff at SBL for providing access to the sites and research facilities.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2024.1451161/full#supplementary-material
References
Abrams, M. D. (1992). Fire and the development of oak forests. Bioscience 42, 346–353. doi: 10.2307/1311781
Abrams, M. D. (1994). Genotypic and phenotypic variation as stress adaptations in temperate tree species: a review of several case studies. Tree Physiol. 14, 833–842. doi: 10.1093/treephys/14.7-8-9.833
Abrams, M. D., and Johnson, S. E. (2013). The impacts of mast years and prescribed fires on tree regeneration in oak forests at the Mohonk preserve, southeastern New York, USA. Nat. Areas J. 33, 427–434. doi: 10.3375/043.033.0405
Aldrich, P. R., Glaubitz, J. C., Parker, G. R., Rhodes, O. E., and Michler, C. H. (2005). Genetic structure inside a declining red oak community in old-growth forest. J. Hered. 96, 627–634. doi: 10.1093/jhered/esi115
Amórtegui, A., Marquis, R., and Dubé-Loubert, H. (2010). Geological overview of the Mount Royal (Montreal area) to Eustis mine (Sherbrooke area) along the highway number 10.
Auguie, B. (2017). gridExtra: miscellaneous functions for “grid” graphics. R package version 2.3. Available at: https://CRAN.R-project.org/package=gridExtra
Barber, S. A., and Silberbush, M. (1984). “Plant root morphology and nutrient uptake, chapter 4” in American Society of Agronomy Special Publication Number 49. eds. N. Roots, W. Influx, P. Growth, S. A. Barber, and D. R. Bouldin (Anaheim, CA), 65–87.
Bauweraerts, I., Wertin, T. M., Ameye, M., McGuire, M. A., Teskey, R. O., and Steppe, K. (2013). The effect of heat waves, elevated CO2 and low soil water availability on northern red oak (Quercus rubra L.) seedlings. Glob. Chang. Biol. 19, 517–528. doi: 10.1111/gcb.12044
Bélanger, N., Côté, B., Fyles, J. W., Courchesne, F., and Hendershot, W. H. (2004). Forest regrowth as the controlling factor of soil nutrient availability 75 years after fire in a deciduous forest of southern Quebec. Plant Soil 262, 363–272. doi: 10.1023/B:PLSO.0000037054.21561.85
Bélanger, N., Holmden, C., Courchesne, F., Côté, B., and Hendershot, W. H. (2012). Constraining soil mineral weathering 87Sr/86Sr for calcium apportionment studies of a deciduous forest growing on soils developed from granitoid igneous rocks. Geoderma 185–186, 84–96. doi: 10.1016/j.geoderma.2012.03.024
Bélanger, N., Collin, A., Khlifa, R., and Lebel-Desrosiers, S. (2021a). Balsam fir and American beech influence soil respiration rates in opposite directions in a sugar maple forest near its northern range limit. Front. For. Glob. Change 4:664584. doi: 10.3389/ffgc.2021.664584
Bélanger, N., Palma Ponce, G., and Brais, S. (2021b). Contrasted growth response of hybrid larch (Larix × marschlinsii), jack pine (Pinus banksiana) and white spruce (Picea glauca) to wood ash application in northwestern Quebec, Canada. iForest 14, 155–165. doi: 10.3832/ifor3597-014
Blossey, B., Curtis, P., Boulanger, J., and Davalos, A. (2019). Red oak seedlings as indicators of deer browse pressure: gauging the outcome of different white-tailed deer management approaches. Ecol. Evol. 9, 13085–13103. doi: 10.1002/ece3.5729
Brais, S., Bélanger, N., and Guillemette, T. (2015). Wood ash and N fertilization in the Canadian boreal forest: soil properties and response of jack pine and black spruce. For. Ecol. Manag. 348, 1–14. doi: 10.1016/j.foreco.2015.03.021
Brose, P. H., Van Lear, D., and Cooper, R. (1998). Using shelterwood harvests and prescribed fire to regenerate oak stands on productive upland sites. For. Ecol. Manag. 113, 125–141.
Brose, P. H. (2010). Long-term effects of single prescribed fires on hardwood regeneration in oak shelterwood stands. For. Ecol. Manag. 260, 1516–1524. doi: 10.1016/j.foreco.2010.07.050
Brose, P. H., Dey, D. C., and Waldrop, T. A. (2014). The fire-oak literature of eastern North America: Synthesis and guidelines. General Technical Report NRS-135. Delaware, OH: Northern Research Station, USDA Forest Service.
Brose, P. H., and Rebbeck, J. (2017). A comparison of the survival and development of the seedlings of four upland oak species grown in four different understory light environments. J. For. 115, 159–166. doi: 10.5849/jof.15-155
Buckley, D. S., Sharik, T. L., and Isebrands, J. G. (1998). Regeneration of northern red oak: positive and negative effects of competitor removal. Ecology 79, 65–78. doi: 10.1890/0012-9658(1998)079[0065:RONROP]2.0.CO;2
Canham, C. D., Finzi, A. C., Pacala, S. W., and Burbank, D. H. (1994). Causes and consequences of resource heterogeneity in forests: interspecific variation in light transmission by canopy trees. Can. J. For. Res. 24, 337–349. doi: 10.1139/x94-046
Collin, A., Messier, C., Kembel, S., and Bélanger, N. (2017). Low light availability associated with American beech is the main factor for reduced sugar maple seedling survival and growth rates in a hardwood forest of southern Quebec. Forests 8:413. doi: 10.3390/f8110413
Collin, A., Messier, C., Kembel, S. W., and Bélanger, N. (2018). Can sugar maple establish into the boreal forest? Insights from seedlings under various canopies in southern Quebec. Ecosphere 9:e02022. doi: 10.1002/ecs2.2022
Crow, T. R. (1988). Reproductive mode and mechanisms for self-replacement of northern red oak (Quercus rubra) - a review. For. Sci. 34, 19–40. doi: 10.1093/forestscience/34.1.19
De’ath, G. (2002). Multivariate regression tress: a new technique for modeling species-environment relatioships. Ecology 83, 1105–1117. doi: 10.1890/0012-9658(2002)083[1105:MRTANT]2.0.CO;2
DeLuca, T. H., MacKenzie, M. D., Gundale, M. J., and Holben, W. E. (2006). Wildfire-produced charcoal directly influences nitrogen cycling in ponderosa pine forests. Soil Sci. Soc. Am. J. 70, 448–453. doi: 10.2136/sssaj2005.0096
Dessai, S., Lu, X., and Risbey, J. S. (2005). On the role of climate scenarios for adaptation planning. Glob. Environ. Chang. 15, 87–97. doi: 10.1016/j.gloenvcha.2004.12.004
Dey, D. C., and Guyette, R. P. (2000). Anthropogenic fire history and red oak forests in south-Central Ontario. For. Chron. 76, 339–347. doi: 10.5558/tfc76339-2
Dey, D. C., Miller, G. W., and Kabrick, J. M. (2008). Sustaining northern red oak forests: managing oak from regeneration to canopy dominance in mature stands in R. L. Deal , ed. Integrated Restoration of Forested Ecosystems to Achieve Multiresource Benefits, Proceedings of the 2007 National Silviculture Workshop, Ketchikan, AK. General technical Report PNW-733, Pacific Northwest Research Station, Portland, OH: USDA Forest Service, 91–105.
Dey, D., and Schweitzer, C. (2018). A review on the dynamics of prescribed fire, tree mortality, and injury in managing oak natural communities to minimize economic loss in North America. Forests 9:461. doi: 10.3390/f9080461
Etterson, J. R., Cornett, M. W., White, M. A., and Kavajecz, L. C. (2020). Assisted migration across fixed seed zones detects adaptation lags in two major north American tree species. Ecol. Appl. 30:e02092. doi: 10.1002/eap.2092
Fei, S., Steiner, K. C., Finley, J. C., and McDill, M. E. (2003). Relationships between biotic and abiotic factors and regeneration of chestnut oak, white oak and northern red oak. J. W. SambeekVan, J. O. Dawson, F. Ponder, E. F. Loewenstein, and J. S. Fralish, editors. Proceedings of the 13th Central Hardwood Forest Conference, NC-234, St. Paul, MN. North Central Research Station, USDA Forest Service, 223–227.
Glaser, B., Wiedner, K., Seelig, S., Schmidt, H.-P., and Gerber, H. (2014). Biochar organic fertilizers from natural resources as substitute for mineral fertilizers. Agron. Sustain. Dev. 35, 667–678. doi: 10.1007/s13593-014-0251-4
Government of Canada. (2020). Historical climate data. Available at: https://climat.meteo.gc.ca/
Government of Quebec. (2019). Forêt ouverte. Available at: https://www.foretouverte.gouv.qc.ca/ (Retrieved Febuary 10, 2021).
Granger, J. J., Buckley, D. S., Sharik, T. L., Zobel, J. M., DeBord, W. W., Hartman, J. P., et al. (2018). Northern red oak regeneration: 25-year results of cutting and prescribed fire in Michigan oak and pine stands. For. Ecol. Manag. 429, 467–479. doi: 10.1016/j.foreco.2018.06.003
Green, S. R., Arthur, M. A., and Blankenship, B. A. (2010). Oak and red maple seedling survival and growth following periodic prescribed fire on xeric ridgetops on the Cumberland plateau. For. Ecol. Manag. 259, 2256–2266. doi: 10.1016/j.foreco.2010.02.026
Greenler, S. M., Swihart, R. K., and Saunders, M. R. (2020). Prescribed fire promotes acorn survival and seedling emergence from simulated caches. For. Ecol. Manag. 464:118063. doi: 10.1016/j.foreco.2020.118063
Hanson, P. J., and Dixon, R. K. (1987). Allelopathic effects of interrupted fern on northern red oak seedlings: amelioration by Suillus luteus L.: Fr. Plant Soil 98, 43–51. doi: 10.1007/BF02381726
Hendershot, W. H., Lalande, H., and Duquette, M. (2008). “Soil reaction and exchangeable acidity, chapter 16” in Soil sampling and analysis. eds. M. R. Carter and E. G. Gregorich. 2nd ed (Boca Raton, FL: CRC Press), 173–178.
Hothorn, T., Hornik, K., and Zeileis, A. (2006). Unbiased recursive partinionning: a conditional inference framework. J. Comput. Graph. Stat. 15, 651–674. doi: 10.1198/106186006X133933
Hothorn, T., Bretz, F., and Westfall, P. (2008). Simultaneous inference in general parametric models. Biom. J. 50, 346–363. doi: 10.1002/bimj.200810425
Hulshof, C. M., Spasojevic, M. J., and Schrodt, F. (2020). The edaphic control of plant diversity. Glob. Ecol. Biogeogr. 29, 1634–1650. doi: 10.1111/geb.13151
Iverson, L. R., and Prasad, A. M. (2002). Potential redistribution of tree species habitat under five climate change scenarios in the eastern US. For. Ecol. Manag. 155, 205–222. doi: 10.1016/S0378-1127(01)00559-X
Iverson, L. R., Peters, M. P., Matthews, S. N., Prasad, A., Hutchinson, T., Bartig, J., et al. (2019). “Adapting oak management in an age of ongoing mesophication but warming climate” in Oak symposium: sustaining oak forests in the 21st century through science-based management. General technical report SRS-237. eds. S. L. Clark and C. J. Schweitzer (Asheton, NC: Southern Research Station, USDA Forest Service), 35–45.
Jacoby, R., Peukert, M., Succurro, A., Koprivova, A., and Kopriva, S. (2017). The role of soil microorganisms in plant mineral nutrition - current knowledge and future directions. Front. Plant Sci. 8:1617. doi: 10.3389/fpls.2017.01617
Katabuchi, M. (2015). LeafArea: an R package for rapid digital image analysis of leaf area. Ecol. Res. 30, 1073–1077. doi: 10.1007/s11284-015-1307-x
Kleiner, K. W., Abrams, M. D., and Schultz, J. C. (1992). The impact of water and nutrient deficiencies on the growth, gas exchange and water relations of red oak and chestnut oak. Tree Physiol. 11, 271 The role of soil microorganisms in plant mineral nutrition 287–287. doi: 10.1093/treephys/11.3.271
Knapp, B. O., Stephan, K., and Hubbart, J. A. (2015). Structure and composition of an oak-hickory forest after over 60 years of repeated prescribed burning in Missouri, U.S.A. For. Ecol. Manag. 344, 95–109. doi: 10.1016/j.foreco.2015.02.009
Kolb, T. E., Steiner, K. C., McCormick, L. H., and Bowersox, T. W. (1990). Growth response of northern red-oak and yellow-poplar seedlings to light, soil moisture and nutrients in relation to ecological strategy. For. Ecol. Manag. 38, 65–78. doi: 10.1016/0378-1127(90)90086-Q
Kormanik, P. P., Sung, S. S., Kormanik, T. L., Schlarbaum, S. E., and Zarnoch, S. J. (1998). Effect of acorn size on development of northern red oak 1-0 seedlings. Can. J. For. Res. 28, 1805–1813. doi: 10.1139/x98-152
Kriebel, H. B., Merritt, C., and Stadt, T. (1988). Genetics of Growth in Quercus rubra: provenance and family effects by the early third decade in the north Central USA. Silvae Genet. 37, 193–198.
Langmaier, M., and Lapin, K. (2020). A systematic review of the impact of invasive alien plants on forest regeneration in European temperate forests. Front. Plant Sci. 11:524969. doi: 10.3389/fpls.2020.524969
Lebel-Desrosiers, S. (2021). Facteurs écologiques régissant la régénération du chêne rouge (Quercus rubra L.) à la limite nordique de son aire de répartition au Québec. M.Sc. thesis: Département de géographie, Université de Montréal.
Lehmann, J., Rillig, M. C., Thies, J., Masiello, C. A., Hockaday, W. C., and Crowley, D. (2011). Biochar effects on soil biota – a review. Soil Biol. Biochem. 43, 1812–1836. doi: 10.1016/j.soilbio.2011.04.022
Lodhi, M. A. K. (1978). Allelopathic effects of decaying litter of dominant trees and their associated soil in a lowland forest community. Am. J. Bot. 65, 340–344. doi: 10.1002/j.1537-2197.1978.tb06076.x
Loftis, D. L., and McGee, C. E. (1993). “Oak regeneration: serious problems, practical recommendations in symposium proceedings” in General technical report SE-84 (Southeastern Forest Experiment Station: USDA Forest Service).
Lombardo, J. A., and McCarthy, B. C. (2009). Seed germination and seedling vigor of weevil-damaged acorns of red oak. Can. J. For. Res. 39, 1600–1605. doi: 10.1139/x09-079
Long, R. P., Brose, P. H., and Horsley, S. B. (2012). Responses of northern red oak seedlings to lime and deer exclosure fencing in Pennsylvania. Can. J. For. Res. 42, 698–709. doi: 10.1139/x2012-025
Major, K. C., Nosko, P., Kuehne, C., Campbell, D., and Bauhus, J. (2013). Regeneration dynamics of non-native northern red oak (Quercus rubra L.) populations as influenced by environmental factors: a case study in managed hardwood forests of southwestern Germany. For. Ecol. Manag. 291, 144–153. doi: 10.1016/j.foreco.2012.12.006
McGee, C. E. (1974). Elevation of seed sources and planting sites affects phenology and development of red oak seedlings. For. Sci. 20, 160–164.
Ministère de l’Énergie et des Ressources naturelles (2020). Système d’information géominière du Québec, Interactive map : Gouvernement du Québec Available at: https://sigeom.mines.gouv.qc.ca/signet/classes/I1108_afchCarteIntr.
Ministère de l’Environnement et de la Lutte contre les changements climatiques (2021a). Juillet 2019: le huitième mois le plus chaud au sud du Québec et le plus chaud mesuré sur Terre en plus de cent ans : Gouvernement du Québec Available at: https://www.environnement.gouv.qc.ca/climat/faits-saillants/2019/juillet.htm.
Ministère de l’Environnement et de la Lutte contre les changements climatiques (2021b). Canicule de juin-juillet 2018: une chaleur rarement ressentie de manière aussi soutenue : Gouvernement du Québec Available at: https://www.environnement.gouv.qc.ca/climat/faits-saillants/2018/canicule.htm.
Messier, C., and Bellefleur, P. (1988). Light quantity and quality on the forest floor of pioneer and climax stages in a birch - beech - sugar maple stand. Can. J. For. Res. 18, 615–622. doi: 10.1139/x88-090
Munoz, S. E., and Gajewski, K. (2010). Distinguishing prehistoric human influence on late-Holocene forests in southern Ontario, Canada. The Holocene 20, 967–981. doi: 10.1177/0959683610362815
Nature-Action Québec. (2015). Guide pratique pour la propagation d'arbres à noix: de la graine à l'arbre. Available at: http://www.noixduquebec.org/uploads/1/9/2/2/19220259/guide_pratique_pour_la_propagation_darbres_%C3%A0_noix...de_la_graine_%C3%A0_larbre.pdf
Nicolescu, V.-N., Vor, T., Mason, W. L., Bastien, J.-C., Brus, R., Henin, J.-M., et al. (2020). Ecology and management of northern red oak (Quercus rubra L. syn. Q. borealis F. Michx.) in Europe: a review. Forestry 93, 481–494. doi: 10.1093/forestry/cpy032
Nilsen, E. T., Walker, J. F., Miller, O. K., Semones, S. W., Lei, T. T., and Clinton, B. D. (1999). Inhibition of seedling survival under Rhododendron maximum (Ericaceae): could allelopathy be a cause? Am. J. Bot. 86, 1597–1605. doi: 10.2307/2656796
Nowacki, G. J., Abrams, M. D., and Lorimer, G. G. (1990). Composition, structure, and historical development of northern red oak stands along an edaphic gradient in north-Central Wisconsin. For. Sci. 36, 276–292. doi: 10.1093/forestscience/36.2.276
Nowacki, G. J., and Abrams, M. D. (1992). Community, edaphic, and historical analysis of mixed oak forests of the ridge and Valley Province in Central Pennsylvania. Can. J. For. Res. 22, 790–800. doi: 10.1139/x92-108
Payette, S., and Filion, L. (2018). La dendroécologie: Principes, méthodes et applications. PRESSES DE L’UNIVERSITÉ LAVAL.
Pedlar, J. H., McKenney, D. W., Aubin, I., Beardmore, T., Beaulieu, J., Iverson, L., et al. (2012). Placing forestry in the assisted migration debate. Bioscience 62, 835–842. doi: 10.1525/bio.2012.62.9.10
Pekins, P. J., and Mautz, W. W. (1988). Digestability and nutritional value of autumn diets of deer. J. Wildl. Manag. 52, 328–332. doi: 10.2307/3801244
Perala, D. A. (1974). “Growth and survival of northern hardwood sprouts after burning” in Research note NC-176, north central Forest experiment station (St. Paul, MN: USDA Forest Service).
Peterman, B. M. (1990). Statistical power analysis can improve fisheries research and management. Can. J. Fish. Aquat. Sci. 47, 2–15. doi: 10.1139/f90-001
Phares, R. E. (1971). Growth of red oak (Quercus Rubra L.) seedlings in relation to light and nutrients. Ecology 52, 669–672. doi: 10.2307/1934157
Phillips, D. H., Foss, J. E., Buckner, E. R., Evans, R. M., and FitzPatrick, E. A. (2000). Response of surface horizons in an oak Forest to prescribed burning. Soil Sci. Soc. Am. J. 64, 754–760. doi: 10.2136/sssaj2000.642754x
Prasad, A., Pedlar, J., Peters, M., McKenney, D., Iverson, L., Matthews, S., et al. (2020). Combining US and Canadian forest inventories to assess habitat suitability and migration potential of 25 tree species under climate change. Divers. Distrib. 26, 1142–1159. doi: 10.1111/ddi.13078
Qian, P., Schoenau, J. J., and Ziadi, N. (2008). “Ion supply rates using ion-exchange resins, chapter 13” in Soil sampling and methods of analysis. eds. M. R. Carter and E. G. Gregorich (Boca Raton, FL: CRC Press), 161–166.
R Core Team (2019). R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing Available at: http://www.R-project.org/.
Raison, R. J. (1979). Modification of the soil environment by vegetation fires, with particular reference to nitrogen transformations: a review. Plant Soil 51, 73–108. doi: 10.1007/BF02205929
Rauschendorfer, J., Rooney, R., and Külheim, C. (2022). Strategies to mitigate shifts in red oak (Quercus sect. Lobatae) distribution under a changing climate. Tree Physiol. 42, 2383–2400. doi: 10.1093/treephys/tpac090
Régnière, J., and Bolstad, P. (1994). Statistical simulation of daily air temperature patterns in eastern North America to forecast seasonal events in insect pest management. Environ. Entomol. 23, 1368–1380. doi: 10.1093/ee/23.6.1368
Reid, C., and Watmough, S. A. (2014). Evaluating the effects of liming and wood-ash treatment on forest ecosystems through systematic meta-analysis. Can. J. For. Res. 44, 867–885. doi: 10.1139/cjfr-2013-0488
Riepšas, E., and Straigytė, L. (2008). Invasiveness and ecological effects of red oak (Quercus rubra L.) in Lithuanian forests. Balt. For. 14, 122–130. Available at: https://hdl.handle.net/20.500.12259/81433
Salifu, K. F., and Jacobs, D. F. (2006). Characterizing fertility targets and multi-element interactions in nursery culture of Quercus rubra seedlings. Ann. For. Sci. 63, 231–237. doi: 10.1051/forest:2006001
Salinger, M. J., Sivakumar, M. V. K., and Motha, R. (2005). Reducing vulnerability of agriculture and forestry to climate variability and change: workshop summary and recommendations. Clim. Chang. 70, 341–362. doi: 10.1007/s10584-005-5954-8
Sander, I. L. (1990). “Northern red oak” in Silvics of North America: 2, Hardwoods: Agriculture handbook. eds. R. M. Burns and B. H. Honkala (Washington, DC: USDA Forest Service), 727–733.
Saucier, J. P., Robitaille, A., and Grondin, P. (2009). “Écologie Forestière in Manuel de Foresterie” in Ordre des ingénieurs forestier du Québec et Éditions Multimondes. 2nd ed (Montréal), 186–205.
Savage, C. (2001). Recolonisation forestière dans les basses Laurentides au sud du domaine climacique de l'érablière à boulot jaune. M.Sc. thesis, Département des sciences biologiques: Université de Montréal.
Schnurr, J. L., Canham, C. D., Ostfeld, R. S., and Inouye, R. S. (2004). Neighborhood analyses of small-mammal dynamics: impacts on seed predation and seedling establishment. Ecology 85, 741–755. doi: 10.1890/02-0644
Schuler, J. L., and Robison, D. J. (2010). Performance of northern red oak enrichment plantings in naturally regenerating southern Appalachian hardwood stands. New For. 40, 119–130. doi: 10.1007/s11056-009-9187-y
Shigo, A. L., and Shortle, W. C. (1979). Compartmentalization of discolored wood in heartwood of red oak. Phytopathology 69, 710–711. doi: 10.1094/Phyto-69-710
Shumway, D. L., Abrams, M. D., and Ruffner, C. M. (2001). A 400-year history of fire and oak recruitment in an old-growth oak forest in western Maryland, U.S.A. Can. J. For. Res. 31, 1437–1443. doi: 10.1139/cjfr-31-8-1437
Signell, S. A., Abrams, M. D., Hovis, J. C., and Henry, S. W. (2005). Impact of multiple fires on stand structure and tree regeneration in central Appalachian oak forests. For. Ecol. Manag. 218, 146–158. doi: 10.1016/j.foreco.2005.07.006
Soil Classification Working Group. (1998). The Canadian system of soil classification. 3rd ed. Agriculture and Agri-Food Canada Publication 1646, 187 pp.
Sork, V. L., Stacey, P., and Averett, J. E. (1983). Utilization of red oak acorns in non-bumper crop year. Oecologia 59, 49–53. doi: 10.1007/BF00388070
Sork, V. L., Stowe, K. A., and Hochwender, C. (1993). Evidence for local adaptation in closely adjacent subpopulations of northern red oak (Quercus rubra L.) expressed as resistance to leaf herbivores. Am. Nat. 142, 928–936. doi: 10.1086/285581
Thériault, R., and Beauséjour, S. (2012). Carte géologique du Québec, Édition 2012. DV 2012–06. Ministère des ressources naturelles : Gouvernement du Québec Available at: https://numerique.banq.qc.ca/patrimoine/details/52327/2241970?docref=ggRNiE1thqwy5YXOoPCncw.
Thiffault, E., Bélanger, N., Paré, D., and Munson, A. D. (2007). How do forest harvesting methods compare with wildfire? A case study of soil chemistry and tree nutrition in the boreal forest. Can. J. For. Res. 37, 1658–1668. doi: 10.1139/x07-031
Ward, J. S., Gent, M. P. N., and Stephens, G. R. (2000). Effects of planting stock quality and browse protection-type on height growth of northern red oak and eastern white pine. For. Ecol. Manag. 127, 205–216. doi: 10.1016/S0378-1127(99)00132-2
Wickham, H., François, R., Henry, L., and Müller, K. (2020). dplyr: a grammar of data manipulation. R package version 0.8.5. Available at: https://CRAN.R-project.org/package=dplyr
Williams, M. I., and Dumroese, R. K. (2013). Preparing for climate change: forestry and assisted migration. J. For. 111, 287–297. doi: 10.5849/jof.13-016
Yi, X., and Wang, Z. (2015). The importance of cotyledons for early-stage oak seedlings under different nutrient levels: a multi-species study. J. Plant Growth Regul. 35, 183–189. doi: 10.1007/s00344-015-9516-7
Yi, X., Zhang, J., and Wang, Z. (2015). Large and small acorns contribute equally to early-stage oak seedlings: a multiple species study. Eur. J. For. Res. 134, 1019–1026. doi: 10.1007/s10342-015-0906-y
Yi, X., Bartlow, A. W., Curtis, R., Agosta, S. J., Steele, M. A., and Heard, M. (2019). Responses of seedling growth and survival to post-germination cotyledon removal: an investigation among seven oak species. J. Ecol. 107, 1817–1827. doi: 10.1111/1365-2745.13153
Zhou, C., Heal, K., Tigabu, M., Xia, L., Hu, H., Yin, D., et al. (2020). Biochar addition to forest plantation soil enhances phosphorus availability and soil bacterial community diversity. For. Ecol. Manag. 455:117635. doi: 10.1016/j.foreco.2019.117635
Keywords: red oak, natural regeneration, ecological factors, provenance, fire disturbance, distribution area, assisted migration
Citation: Lebel Desrosiers S, Collin A and Bélanger N (2024) Factors affecting early red oak (Quercus rubra L.) regeneration near its northern distribution limit in Quebec. Front. For. Glob. Change. 7:1451161. doi: 10.3389/ffgc.2024.1451161
Edited by:
Alison D. Munson, Laval University, CanadaReviewed by:
Sergio Espinoza, Catholic University of the Maule, ChileJesús Rodríguez-Calcerrada, Polytechnic University of Madrid, Spain
Copyright © 2024 Lebel Desrosiers, Collin and Bélanger. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Simon Lebel Desrosiers, c2ltb24ubGViZWxkZXNyb3NpZXJzQHRlbHVxLmNh