 Barbora Dvořáková
Barbora Dvořáková Jaroslav Holuša
Jaroslav Holuša Jakub Horák
Jakub Horák- Faculty of Forestry and Wood Sciences, Czech University of Life Sciences, Prague, Czechia
Introduction: As a result of climate change and bark beetle outbreaks in forests, extensive salvage, and sanitary cutting have been undertaken in Europe, resulting in many clear-cuts with coarse stumps. The populations of pine weevils (Hylobius abietis and Hylobius pinastri) are steadily increasing. A high abundance of pine weevils feeding on seedlings typically results in significant economic losses and prolongs the forest establishment period. However, information on these species spatial distribution and habitat selection patterns, necessary for assessing their harmfulness and subsequently accurately estimating their threat to reforested areas, is lacking. To determine which factors influence spatial selection patterns and seedling type preference, this study investigated which clear-cut factors increase pine weevil abundance and which seedling types (species and age) are the most preferred.
Methods: The experiment was carried out on 20 clear-cuts in the central Czech Republic. We evaluated soil moisture level, average stump distance and diameter, proportion of other conifers, and whether the stumps were mulched. We detected the abundance of pine weevils using pitfall traps. We determined the feeding scar intensity on the first 10 cm of seedling stems on commonly used seedlings for reforestation in central Europe: 1-year-old Scots pine (Pinus sylvestris), European larch (Larix decidua), and 3-year-old Norway spruce (Picea abies), with 1,200 seedlings in total. The individual seedling types alternated in rows. The results were evaluated using generalized linear mixed-effect models (GLMMs). As dependent variables we used total numbers of H. abietis and H. pinastri. The soil moisture level, mulching, proportion of other conifers, average stump diameter, and distance were considered independent variables.
Results: We found that Norway spruce was the least attractive to pine weevils. We found a higher abundance of H. abietis females in moist clear-cuts, confirming that dry clear-cuts are less suitable for oviposition.
Conclusion: According to our findings, if foresters want to plant coniferous seedlings, it seems that planting older spruce is a better option than planting larch and pine. These findings provide valuable insights for forest management and reforestation strategies, equipping foresters with the knowledge to mitigate the threat of pine weevils and ensure successful forest establishment.
1 Introduction
1.1 Pine weevil abundance increases as a result of bark beetle outbreaks
Ongoing climate change has increased water deficits, stressing trees and making them more vulnerable to insect pests (Holuša et al., 2018). Widespread bark beetle outbreaks have recently occurred across Europe. The most effective defense against the spread of bark beetles is to cut the infested trees and remove them from the forest before the beetle offspring leave the trunks (Mezei et al., 2017). The high number of newly emerging clear-cuts with stumps from felled trees provides suitable conditions for the spread and successful reproduction of the pine weevil (Hylobius abietis) (Linnaeus, 1758) (Coleoptera: Curculionidae) (Långström and Day, 2004). The weevils are attracted to places where fresh stumps emit volatiles (monoterpenes and ethanol) (Nordlander, 1987). Freshly made clear-cuts are suitable habitats for the pine weevil to breed (Solbreck, 1980). Therefore, the population of this species is expected to increase due to extensive clear-cut reforestation after bark beetle outbreaks (Doležal et al., 2021). The pine weevil is a widespread and severe pest associated with regenerated coniferous forests in northern and Central Europe (CAB International, 2003; Skrzecz, 2017). Forest management practices strongly influence the extent and severity of pine weevil damage (Jactel et al., 2009).
1.2 Pine weevil spread
Adult weevils, flying up to 10 km, use both smell and vision to greatly enhance their orientation when seeking a suitable breeding site (Örlander et al., 2000; Björklund, 2004). H. abietis may respond differently to primary attractants emitted by stumps depending on the season and the state of its reproductive development (Nordenhem and Eidmann, 1991). For example, the orientation of adult weevils involves responses to light and humidity, and their responses vary at different stages of the life cycle; e.g., older reproductive weevils are strongly photonegative (Havukkala and Selander, 1976; Havukkala, 1979). Nordlander et al. (2023) found that harvested stump species (coniferous vs. deciduous) and the age of clear-cuts affect pine weevil abundance and damage (Nordlander et al., 2023).
1.3 Pine weevil life cycle
The life cycle of H. abietis usually lasts 1 to 3 years. Adult pine weevils disperse to new clear-cuts during late spring and early summer (Nordenhem and Nordlander, 1994), where they lay eggs in stumps or soil on newly created clear-cuts (Nordlander et al., 1997; Tan et al., 2011). The larvae develop in the bark on the underground roots of dying stumps, where they eat the phloem and pass through four larval molts before pupation. Microclimatic conditions influence the length of the life cycle. Pupation and emergence can happen the same year (at the end of summer), but usually 1 year after egg laying or in bad microclimatic conditions, it can be postponed until the late summer of the following year, or adults may emerge more than 2 years after occupying a clear-cut (Inward et al., 2012; Wainhouse et al., 2014).
The host tree species is essential for larval development, although larvae develop on all conifers (Véle, 2022). However, pine (Pinus sylvestris L.) is the preferred tree species for larval development over Norway spruce, Picea abies (L.) Karst. (Munro, 1928; Leather et al., 1994). Larvae develop faster and have lower mortality in pine stumps (Bejer-Petersen et al., 1962; Doom and Frenken, 1980; Thorpe and Day, 2002). Most individuals (ca. 75%) in Europe develop a little longer than 1 year (ca 14 months) (Bejer-Petersen et al., 1962).
Microclimate conditions, therefore, influence the length of the life cycle and determine how long reforested clear-cuts are attractive for repeated infestations (Leather et al., 1999; Inward et al., 2012). Therefore, fallow periods are the best option for integrated pest management (IPM), which varies in Europe according to geographical location. In southern and central Europe, postponing reforestation for 2 years is sufficient, whereas 4 years or more are necessary in Nordic countries (Bejer-Petersen et al., 1962; Bakke and Lekander, 1965; Bejer-Petersen, 1975; Moore et al., 2004).
Adults can live for up to 4 years (Eidmann, 1979). While larvae feed on stumps, adults typically feed on the root collar of seedlings that are planted in clear-cuts and other sources of thin conifer bark, such as twigs in crowns of mature trees and roots of larger trees (Örlander et al., 2000; Wallertz et al., 2006; Fedderwitz et al., 2018). Adults feeding begins when adults emerge from hibernation, i.e., in spring when temperatures are 8–9°C (Munro, 1928; Nordenhem, 1989), continues throughout the entire vegetative season, when oviposition occurs, and into autumn, when the first generation of beetles may appear (Moore et al., 2004). After this damage, the seedlings lose growth potential or die (Thorpe and Day, 2002; Day et al., 2004).
1.4 Food sources for pine weevils
H. abietis is a polyphagous species (Munro, 1928; Toivonen and Viiri, 2006); adults feed on many species of conifers (e.g., P. sylvestris, Larix decidua Mill., P. abies, Pseudotsuga menziesii (Mirb.) Franco) (Leather et al., 1999; Månsson and Schlyter, 2004). Researchers who studied the feeding preferences of pine weevils for specific tree species have reported no difference between Norway spruce and Scots pine (Toivonen and Viiri, 2006) but higher damages on Douglas fir (Pseudotsuga menziesii) and Sitka spruce [Picea sitchensis (Bong.) Carr.] compared to Norway spruce (Wallertz et al., 2014). A higher preference for Douglas fir over Norway spruce was also confirmed by Doležal et al. (2021). However, pine weevil feeding scars have also been recorded on deciduous trees, such as Fraxinus excelsior L., Alnus glutinosa (L.) Gaertn., Fagus sylvatica (L.) Gaertn., Quercus robur L., Salix L. spp., Betula L. spp. and others (Toivonen and Viiri, 2006; Manlove et al., 2013).
1.5 Damage effects
The pine weevil severely damages all conifer species planted in large amounts in various parts of Europe, e.g., Norway spruce and Scots pine in Scandinavia (e.g., Johansson et al., 2015), Sitka spruce and Corsican pine in the UK (e.g., Wainhouse et al., 2009), and maritime pine and radiata pine in Spain (e.g., López-Villamor et al., 2019). In Europe, H. abietis causes annual damage of almost 120 million EUR (Lalík et al., 2021). Foresters have noted that damage intensity to the European larch (L. decidua) has increased recently. There are concerns that this increase may be greater than damage to Central Europe's other main economic seedling types.
1.6 Study aims
Based on these findings, we investigated the following questions: (I) Can different factors at clear-cuts increase the attractiveness of these sites for occupation by more pine weevil individuals, and do different management practices after logging also influence the pine weevil population? We included factors that may affect the quality (soil moisture level; see Carpenter et al., 1988) and the amount of breeding substrate (average distance between stumps and average diameter of stumps,) as well as a factor representing an increase in the emission of volatiles (presence of mulch). Recently, the amount of coarse logging residue harvested from clear-cuts, including stumps, has increased (Saarinen, 2006; Rahman et al., 2018), and we speculate that the odor of clear-cut mulch may attract more adult pine weevils (Nordlander, 1987; Brattli et al., 1998). Furthermore, we wanted to determine (II) if the attractiveness of European larch, Scots pine, and Norway spruce varies in relation to H. abietis.
2 Materials and methods
2.1 Study localities and design
The first part of the experiment investigated whether clear-cut factors affect the abundance of pine weevils in the chosen clear-cut samples. The second part of the experiment considered the feeding scar intensity on the three main economic seedling species of Europe: Norway spruce (P. abies), Scots pine (P. sylvestris), and European larch (L. decidua). The experiments were conducted at the same sites and time periods. The study localities were in the Czech Republic in the upper lands of central Bohemia in the Brdy Mts. In this region, rainfall varies between 550 and 600 mm, with average annual temperature ranging from 7 to 8°C, which is higher than the average temperatures observed in northern regions, making it a relatively dry area. The average monthly precipitation during the study period (May to August) was 70 mm. The tree species composition is dominated by Norway spruce (74%). It includes a mixture of Scots pine (15%), European larch (5%), European silver fir (Abies alba Mill.) (0.6%), oak (Quercus sp.) (2.5%), European beech (Fagus sylvatica L.) (2.1%) and others (0.8%).
The soils in the clear-cuts were dry Ranker Cambisol (vegetation coverage up to 10%), moderately moist mesobasic Cambisol (vegetation coverage 10–50%) with a cover of low grasses <20 cm tall, and moist gley or pseudogley Cambisol (vegetation coverage above 50%) with wetland grasses taller than 1 m. The grasses represented a part of the moisture levels. The soil moisture level was divided into three levels: dry, moderately moist, and wet, how soil types are affected by water (Viewegh et al., 2003). There was no slash on the clear-cuts.
In total, we selected 20 clear-cuts (localities) from large-scale experiments, covering a total of 35 × 15 km in large spruce-dominated conifer stands with a maximum of 5% pine, which were harvested in October–December 2022 and reforested in March–spring 2023. The altitudes of the areas ranged from 420 to 700 m above sea level. The study clear-cuts were planted with Scots pine, Norway spruce, and European larch at a rate of 3,200 seedlings/ha. The individual tree species alternated in rows. The spruce seedlings were 1-year-old, the larch and pine seedlings were 1-year-old, the average height of the pine seedlings was 35 (±6) cm, that of the larch seedlings was 44 (±9) cm, and that of the spruce seedlings was 36 (±8) cm.
2.2 Calculated clear-cut factors
Five clear-cut factors that could affect the abundance of pine weevils were considered: the soil moisture level, the average distance between stumps, the average diameter of stumps, the proportion of other conifers, and whether the stumps were mulched (harvested). For the average diameter of the stumps of each tree species and the distances between them, ten stumps were measured in a line passing through each locality. Mulching was performed in March before the seedlings were planted by harvesting the stumps via a tractor with a cutter and rotavator. Woodchips of ca. 3 × 5 cm were evenly dispersed over the clear-cut study area, forming a layer of ca. 5 cm. To evaluate the impact of mulching on pine weevil abundance, only the presence or absence of mulch was considered.
2.3 Trapping H. abietis
The number of weevils caught in the pitfall traps determined the total abundance of adult weevils. Five pitfall traps were placed diagonally at a distance of 10 m from each other across each clear-cut area to catch pine weevil adults; thus, the traps were as representative as possible of the total abundance in the locality. Each pitfall trap was made from a 1-L plastic bucket containing bait in the form of a small 50-ml bottle filled with 70% ethanol with an ~5 cm-long stick of pine attached. At 2 cm below the edge, 8-mm-diameter holes were made so that pine weevil beetles, not larger beetles or small vertebrates, could be caught (see Lalík et al., 2019; Figure 1). The stick was regularly replaced every seven days to ensure that the concentration of the emitted volatiles did not decrease. Two inches of 4% formaldehyde were poured on the bottom of the pitfall traps to maintain the freshness of the trapped insects. Pine weevil collection was performed every 14 days from mid-May to early September 2023.

Figure 1. A ground trap with formaldehyde from a 1-L plastic bucket with 50 ml of ethanol bait and a pine twig 5 inches thick and 5 cm long was used for catching pine weevils in the Czech Republic.
Stereomicroscopy in the laboratory was used to determine the sex ratio of H. abietis weevils by the morphological depression (dimple) in the last body segment and to determine the number of H. pinastri (Gyllenhal, 1813) weevils based on the red coloration of the legs and the white coloration of the hairs (Brosset, 2017).
2.4 Characterization of feeding scars on seedlings
On each clear-cut we selected 60 seedlings (20 for each species) which we observed. We selected a transect in the middle of the clear-cuts, which contained seedlings of varying heights and overall vitality. These seedlings were marked and fenced to prevent them from being sprayed with insecticides, which are used to protect them from pine weevil damage (Figure 2). The study plots of the seedlings were always located in the middle of the clear-cut plots. The evaluation of feeding scar intensity was performed at the beginning of September. We evaluated the feeding scar intensity on the collar of the seedlings up to 10 cm from the ground, i.e., how many 1 cm sections were debarked; values ranged from 0 to 10 cm. We also recorded the height of the highest feeding scar (hereafter referred to as the “top feeding scar”) and the total height of the seedlings. The mortality of the seedlings due to debarking was also recorded.

Figure 2. Experimental clear-cuts with stumps left and colored pins fencing the study plots with untreated pine, larch, and spruce seedlings; dry (a), moist (b), and mulched (c).
2.5 Statistical analysis
We split the study into two objectives (a determination of clear-cut factors and weevil abundance and a comparison of the damage to three seedling types (tree species and age of seedlings) because the relationship between the number of pine weevils and damaged seedlings is very weak, as evidenced by several studies (Wilson and Day, 1994; Örlander et al., 1997; Von Sydow, 1997; López-Villamor et al., 2019; Nordlander et al., 2023). The level of damage to seedlings cannot be related to the number of weevils caught in traps, as seedlings planted at the locality provide only part of the available food source for pine weevils (Nordlander et al., 2023). Adults naturally feed on branches in the crowns of mature trees and their roots or logging residues (Nordlander et al., 2003; Hansen et al., 2005; Fedderwitz et al., 2018). The following analyses were conducted in R Statistical Software (v. 4.2.1.).
The numbers of H. abietis females, males, and all adults of H. pinastri were used as dependent variables to evaluate the influence of the considered clear-cut factors. Adults were caught and summit from 5 pitfall traps on each of 20 clear-cuts (e.g., N = 20). The soil moisture level, mulching of stumps, proportion of other conifers, the average diameter of stumps, and the average distance between stumps were considered independent variables. In total, five models were analyzed. First, we tested for multicollinearity among the independent variables using the HH package (Heiberger, 2019) with the following criterion for excluding variables: VIF > 2. The exclusion of multicollinearity from statistical models enhances reliability and interpretation (Graham, 2003). The DHARMa package (Hartig, 2021) was used to assess the suitability of the GLM by testing the distribution of residuals; Gaussian, Poisson, and negative binomial distributions were considered for the dependent variables. If a negative binomial distribution was fit, the resulting Theta value was used in the final models. If none of these distributions were suitable, a quasi-Poisson distribution was used. We used generalized linear mixed-effect models (GLMMs) constructed using the R package MASS (Venables and Ripley, 2002) and generalized linear models (GLM). The numbers of H. abietis females, males, and all adults of H. pinastri were used as dependent variables. Clear-cut was used as a random factor. The results were visualized using the package visreg (Breheny and Burchett, 2017).
The same procedure was used to evaluate the effects of clear-cut factors on pine weevil (H. abietis and H. pinastri) feeding on seedlings. The feeding scar intensity was used as the dependent variable, and soil moisture level, mulching, proportion of other conifers, average stump diameter, and average distance of stumps were considered independent variables.
GLMM was used to evaluate the effects of seedling type on the intensity of Hylobius abietis and Hylobius pinastri feeding scars. The feeding scar intensity was used as a dependent variable, and seedling types (Species) and their heights were used as independent variables.
To test the correlation between H. pinastri and H. abietis abundance, we used the Shapiro–Wilk test combined with a histogram to test the normality of the data distribution. Due to the non-Gaussian distribution, the Spearman correlation test was used.
The influence of seedling types on the height of the top feeding scar GLMM was used. The height of the top feeding scar was used as a dependent variable, and seedling types and their height were used as independent variables.
GLM was also used to evaluate the probability of survival of all seedling types. Survival was the dependent variable, and seedling type, plant height, and the feeding scar intensity were the independent variables.
3 Results
3.1 Clear-cut factors and presence of weevils
A total of 6,172 weevils were caught in all 20 clear-cuts. Of these, 5,918 were H. abietis and 254 were H. pinastri. There were 2,849 males and 3,069 females of the H. abietis species. Only the soil moisture level influenced the total number of H. abietis (p = 0.05) and female abundance (p = 0.041) on clear-cuts (Table 1). None of the other factors considered had a statistically significant effect on the total number of males or females of H. abietis caught in the pitfall traps (p > 0.05). H. pinastri weevils accounted for 4.2% of all the caught weevils.
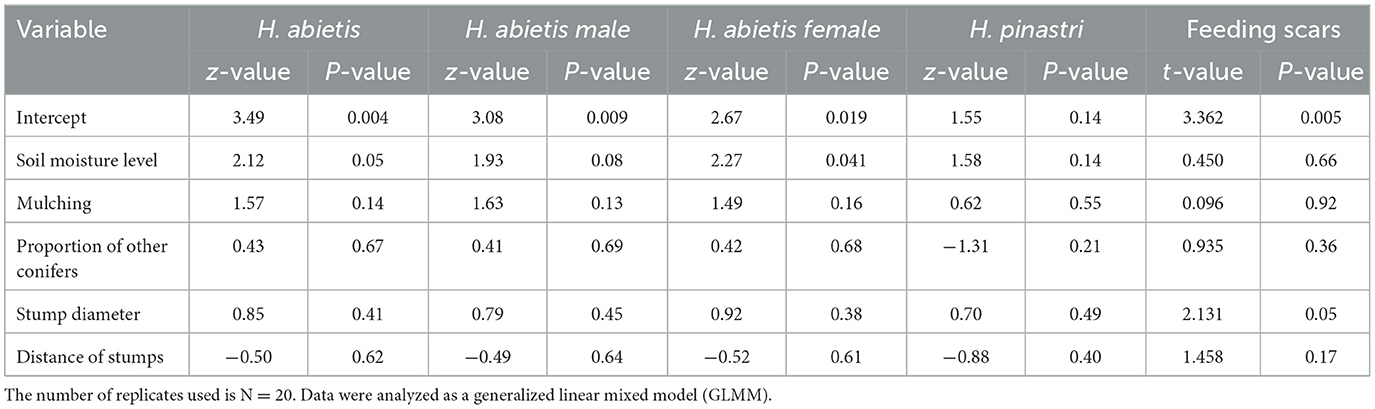
Table 1. The influence of clear-cut environmental variables (factors) on the total abundance of Hylobius abietis, its males, females, and H. pinastri beetles and the intensity of Hylobius abietis feeding scars in the Czech Republic.
We found that the number of H. pinastri caught was strongly and significantly correlated with the number of H. abietis caught for both males and females [RS (%)= 93.52].
3.2 Feeding on seedlings
A comparison of pine weevil feeding intensity on the collar of individual trees showed that the tree seedling type had a statistically significant effect on the feeding scar intensity. Plant height didn't have any effect (Table 2). Pines and larches were significantly more debarked than spruces (Supplementary Figure 1). Besides the seedling parameters, we also evaluated how clear-cut factors affected feeding scar intensity. We found that only the average diameter of stumps influenced the feeding scar intensity (P = 0.05; Table 1).
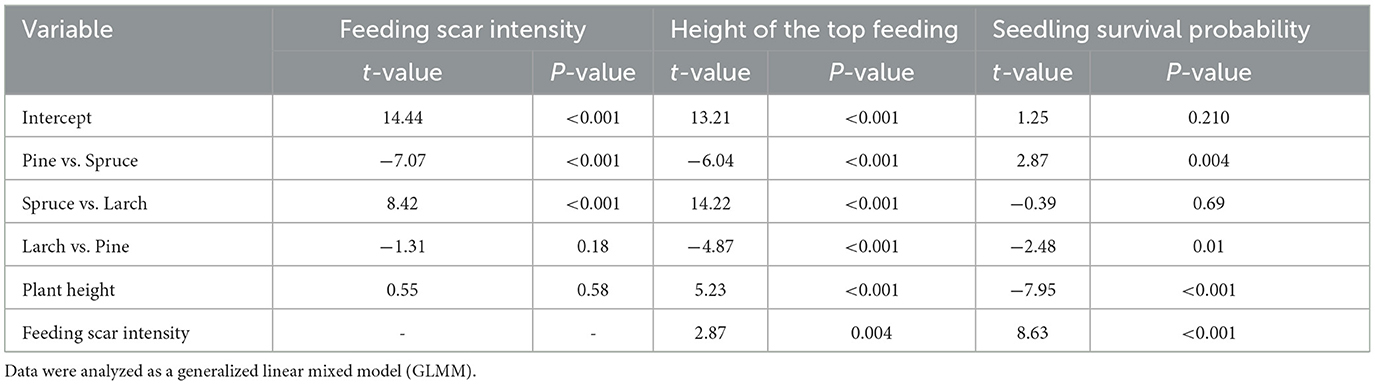
Table 2. The influence of seedling types on the intensity of Hylobius abietis feeding scars, the height of the top feeding scar, and the function of seedling survival probability in clear-cuts in the Czech Republic.
We also evaluated the differences in the feeding scar intensity among seedling types (tree species and age of seedlings) with no influence of clear-cut factors (Figure 3). Larch and pine were significantly more damaged than spruce, but no difference was found between them (Table 2). On dead seedlings, the feeding scar intensity was greater. The average (±se) damage intensity in dead seedlings was 7, and those that survived were 4.5 (Z = −8.49, p ≤ 0.001) (Figure 4).
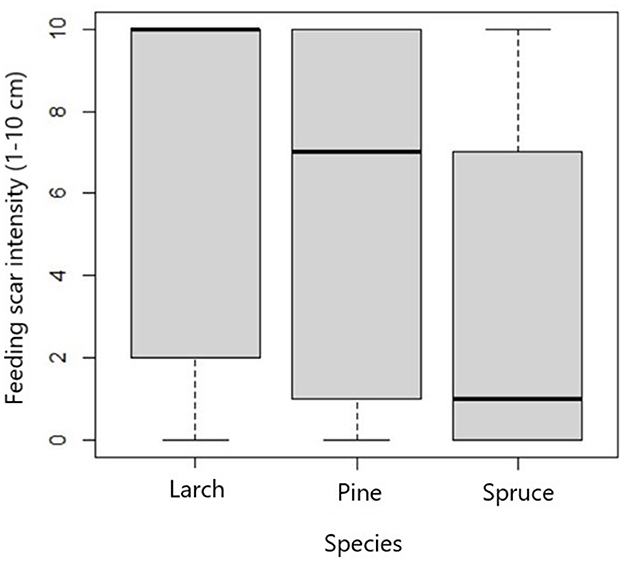
Figure 3. The feeding scar intensity caused by Hylobius abietis and Hylobius pinastri on three different types of seedlings expressed as the number of parts with feeding on 10 cm of the studied stem section on experimental clear-cuts in the Czech Republic.
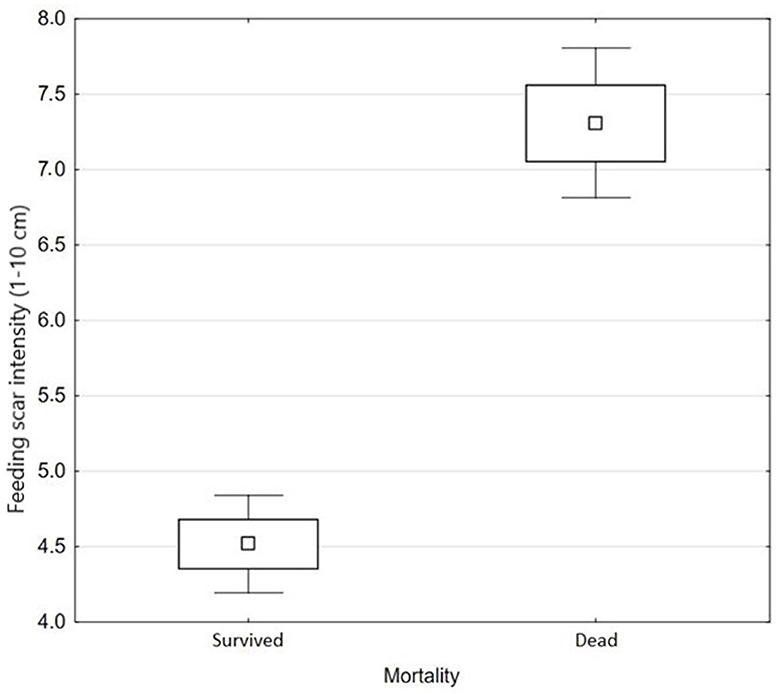
Figure 4. The average (±SE) damage intensity (on a scale of 1–10) in seedlings that died and survived.
In addition to the feeding scar intensity, the height of the top feeding scar was evaluated. The effects of seedling type and height had a statistically significant effect on the top feeding scar (Table 2). The top feeding scar on larch was the highest, while that on spruce was the lowest (Supplementary Figure 2). The number of dead seedlings was greater than the number of surviving ones.
The average mortality rate was 22.7% (SE = 2.3) for larch, 22.4% (SE = 2.6%) for pine, and 23.2% (SE = 2.4%) for spruce. Pine is more likely to survive the damage than spruce and larch (Table 2).
4 Discussion
Our study focused on five factors that may affect the abundance of pine weevils and which seedling types they prefer as food sources. Only the soil moisture level affected the pine weevil abundance. None of the other factors we examined, e.g., average stump diameter, the proportion of other conifers, the average distance between stumps, and the mulching of stumps, affected the overall number of pine weevils caught. We found that the weevils preferred 1-year-old Scots pine (P. sylvestris) and 1-year-old European larch (L. decidua) compared to 3-year-old Norway spruce (P. abies).
Because it is impossible to relate the number of beetles caught in traps and feeding damage on seedlings (Nordlander et al., 2023), we evaluated the influence of clear-cut factors on the feeding scar intensity caused by pine weevil adults. We didn't find any of the factors influencing feeding scar intensity. Only one factor, the average diameter of stumps tends to be significant but shows a higher feeding intensity with decreasing diameter. We would expect the opposite effect. We assumed that bigger stumps would emit more volatiles and allow more beetle breeding space. This finding needs to be studied more in the future.
4.1 Hylobius species
Among the Hylobius weevils caught, H. abietis was the most abundant, with a male-to-female ratio of 1:1.08, which is consistent with the findings of other studies (Nordenhem and Eidmann, 1991; Bylund et al., 2004; Voolma and Sibul, 2006; Heber et al., 2024). The proportion of H. pinastri in the total population was less than 5%. The abundance of H. pinastri is generally much lower and thus poses less of a threat of economic losses (Långström, 1982; Von Sydow and Örlander, 1994; Örlander et al., 1997; Voolma, 2001). The proportion of H. pinastri caught was similar in some studies (e.g., Nordlander, 1990) but significantly greater in others. In Estonia, H. pinastri accounted for a quarter of the caught weevils, but H. abietis was the dominant species (Luik and Voolma, 1989).
We found a high correlation between the number of H. abietis and H. pinastri weevils captured in each clear-cut area, suggesting that similar environmental factors suit these two species. The difference between these two species is that H. pinastri is less attracted to α-pinene than H. abietis (Von Sydow and Örlander, 1994). Unlike H. abietis, H. pinastri prefers to feed on spruce rather than pine (Viiri and Miettinen, 2013). Another difference in ecological niches may be that H. pinastri prefers damper sites more than H. abietis (Örlander et al., 2000).
4.2 Clear-cut factors
Our first objective was to identify the effect of clear-cut factors on the abundance of pine weevils. We did not include the size of the clear-cuts among our factors because the weevils fly in and then spread out, so the density is even throughout the clear-cut (Hansen et al., 2005). According to Örlander et al. (2000), weevils migrate by flight and land in the crowns of mature trees near clear-cut edges, where they feed for a short period before they spread over the clear-cut for reproduction (Örlander et al., 2000; Skrzecz et al., 2021). The pitfall traps were equally dispersed throughout the clear-cut, so the number of caught weevils represented population density.
Neither stump diameters nor stump distances per unit area affected our experiment's abundance of pine weevils. Also, López-Villamor et al. (2019) reported that only two factors influenced the level of seedling damage: the age of the clear-cuts and the amount of logging residues left in the clear-cuts. The average stump diameter and basal area of the cut did not affect the damage (López-Villamor et al., 2019; Véle, 2022). Nordlander et al. (2023) reported that 2-year-old clear-cuts were more damaged than 1-year-old. We studied only 1-year-old clear-cuts because the pine weevil development in Central Europe is shorter compared to Scandinavia (Bejer-Petersen et al., 1962).
Deciduous tree stumps in clear-cuts could reduce beetle abundance, as pine weevils use conifer stumps for reproduction (Björkman et al., 2015). Our clear-cuts did not include any stumps of deciduous trees; we tested only the species composition with domination of spruce stumps and only a small proportion of pine and larch, which did not exceed 10%. Therefore, pine and larch stumps did not affect the number of pine weevils. We did not find sufficient data supporting a significant correlation between the amount of substrate suitable for egg laying, e.g., the number of stumps and the number of pine weevils.
We hypothesized that stumps in damper habitats would emit more volatiles because they could be infested with fungi (Carpenter et al., 1988) and thus attract more pine weevils. This hypothesis was supported by the result that higher H. abietis female abundance was found on clear-cuts with higher soil moisture levels.
Adult pine weevils are attracted to volatile compounds emitted by stumps and woody residues in logged areas (Tan et al., 2011); we hypothesized that weevil abundance would be greater in mulched clear-cuts due to high concentrations of emitted volatiles, as was the case in a study with slash (López-Villamor et al., 2019). Mulched clear-cuts did not attract significantly more beetles; however, only three of the studied clear-cuts were mulched. This hypothesis requires further study.
4.3 Feeding on seedlings
For the experiment, we chose 1-year-old clear-cuts because these are the areas where seedlings are most threatened in central Europe. Over time, their vulnerability decreases (Örlander et al., 1997; Örlander and Nilsson, 1999; Nordlander et al., 2011; Paraschiv, 2020; Galko et al., 2022). The clear-cuts contained seedlings of European forests' three main economic tree species. We observed damage to the 1-year-old seedlings of P. sylvestris, 1-year-old L. decidua, and 3-year-old P. abies, which are commonly used in such seedling stages for reforestation in central Europe. The seedlings were planted evenly (alternating rows) in clear-cuts. We predicted that pine seedlings would be the most damaged as they are the primary tree species for pine weevil development and have more larvae than spruce (Von Sydow and Birgersson, 1997). Véle (2022) found that adult beetles hatching from spruce stumps are larger, but the abundance is smaller. This leads us to think that pine is more suitable for them (Véle, 2022).
In our study, the feeding scar intensity on pine seedlings did not differ from feeding scars on larch. Larch was also found to be attractive by Doležal et al. (2021), but in their laboratory experiment, larch was preferred for almost the entire experimental period, while pine weevils started to prefer pine only during the last week of his experiment. However, it is interesting why the pine weevil prefers to eat larch, as female pine weevils feeding on larch lay far fewer eggs than females feeding on spruce or pine (Doležal et al., 2021).
However, pine was reported as the most preferred tree species in most other studies. Notably, pine was compared only with spruce (Örlander et al., 2000; Véle, 2022) or deciduous seedlings or spruce, pine, and ash (Leather et al., 1994). H. abietis likely prefers Scots pine than Norway spruce because spruce in central Europe is usually planted as a bare-root seedling, which is thicker than pine and, therefore, probably less palatable. One reason may be that older spruce seedlings have a higher concentration of (-)-limonene than pine (Wibe et al., 1998), which may outweigh the smell of attractive compounds (Nordlander, 1990, 1991; Lindgren et al., 1996). Wallertz et al. (2014) found no differences between spruce, pine, or hybrid larch. A possible reason why the pine weevil might prefer larch to spruce could be the smaller size of larch seedlings than spruce seedlings. Larger seedlings, e.g., those with larger stem diameters, are known to survive the infestation of pine weevils better (Thorsen et al., 2001).
Certain seedlings may be more palatable to the pine weevil, so they return to the seedling and debark the entire stem. If pine weevils bite a seedling, resin, and volatiles are emitted and attract other weevils, which may be why the whole stem becomes debarked. We compared the height of damage to the seedlings because several studies have shown that larger seedlings are more resistant than thinner seedlings (Wainhouse et al., 2008). Because seedling size also influences weevil feeding, older seedlings are less preferred by pine weevils than younger ones (Thorsen et al., 2001); for larger seedlings with thicker bark, we believe that weevils are more likely to climb to taller heights and seek more palatable bark, which is why taller seedlings have more top feeding scars.
4.4 Implications for forest management
The protection of reforested areas from the pine weevil (H. abietis) is one of the largest problems in forest management in many European countries (Nordlander et al., 2011; Willoughby et al., 2017; Tudoran et al., 2021; Galko et al., 2022). Our experiment showed that in the coniferous forests of central Europe, all clear-cuts with fresh conifer stumps could be impacted by many pine weevils. If a clear-cut is reforested with conifer seedlings in the first year after logging, a high level of damage caused by pine weevils must be considered. Pine weevils preferred 1-year-old larch and pine over 3-year-old spruce. We would recommend foresters to plant deciduous trees.
However, when growing all three main coniferous species, e.g., pine, larch, and spruce, foresters must be prepared for pine weevil damage and the necessary use of IPM against them. Foresters may plant already treated seedlings and cause weevil mortality (with insecticide) or protect the seedlings (treating them with wax, sand, or collars); alternatively, foresters may have to monitor the seedlings and spray them when the economic damage threshold is exceeded. We advise foresters to use the fallow period, which in central Europe consists of postponing reforestation for 2 years while the amount of volatile compounds emitted from fresh stumps decreases, to decrease the attractiveness of clear-cuts to pine weevils (Rahman et al., 2018).
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
BD: Writing – review & editing, Writing – original draft, Methodology, Investigation, Conceptualization. JHol: Writing – review & editing, Validation, Supervision, Methodology, Conceptualization. JHor: Writing – review & editing, Data curation. JHra: Writing – review & editing. MB: Writing – review & editing, Methodology. MZ: Writing – review & editing, Methodology.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research was supported by the Czech University of Life Sciences Prague project No. IGA A_01_23.
Acknowledgments
We thank the Colloredo-Mannsfeld Forestry Company for providing the clear-cuts with seedlings. The fieldwork was conducted in the Svatá Anna and Lhota forest districts. We thank Martin Káš for his help in the fieldwork.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2024.1399405/full#supplementary-material
Supplementary Figure 1. Residuals of feeding scar frequencies on a 10-cm section of seedlings among different types of seedlings (pine, larch, spruce) in clear-cuts in the Czech Republic. Generalized linear mixed-effect models (GLMMs) were constructed using the R package MASS.
Supplementary Figure 2. Residuals of top feeding scar heights caused by Hylobius abietis and Hylobius pinastri on pine, larch, and spruce seedlings in the Czech Republic. Generalized linear mixed-effect models (GLMMs) were constructed using the R package MASS.
Abbreviations
IPM, Integrated pest management; Mts, Mountains; GLM, General linear model; GLLM, Generative large language model; VIF, Variance inflation factor.
References
Bakke, A., and Lekander, B. (1965). Studies on Hylobius abietis L. II. The influence of exposure on the development and production of Hylobius abietis, illustrated through one Norwegian and one Swedish experiment. Meddelelser fra Det norske Skogsforsøksvesen 20, 117–135.
Bejer-Petersen, B. (1975). Length of Development and Survival of Hylobius abietis as Influenced by Silvicultural Exposure to Sunlight. Koebenhavn: Arsskrift, Kongelige Veterinaer og Landbohoeejskole.
Bejer-Petersen, B., Juutinen, E., Bakke, A., Butovitsch, V., Eidman, H., Heqvist, K. J., et al. (1962). Studies on Hylobius abietis L. 1. Development in the Nordic countries. Acta Entomol. Fenn. 17, 1–107.
Björklund, N. (2004). Movement behaviour and resource tracking in the pine weevil Hylobius abietis [Doctoral thesis]. Uppsala: Swedish University of Agricultural Sciences.
Björkman, C., Bylund, H., Nilsson, U., Nordlander, G., and Schroeder, M. (2015). “Effects of new forest management on insect damage risk in a changing climate,” in Climate Change and Insect Pests, eds. C. Björkman and P. Niemelä (Wallingford: CABI), 248–266.
Brattli, J. G., Andersen, J., and Nilssen, A. C. (1998). Primary attraction and host tree selection in deciduous and conifer living Coleoptera: Scolytidae, Curculionidae, Cerambycidae and Lymexylidae. J. Appl. Entomol. 122, 345–352. doi: 10.1111/j.1439-0418.1998.tb01511.x
Breheny, P., and Burchett, W. (2017). Visualization of regression models using visreg. R J. 9, 56–71. doi: 10.32614/RJ-2017-046
Brosset, A. (2017). Effects of methyl jasmonate and other types of damage to seedlings on pine weevil Hylobius abietis L. (Coleoptera: Curculionidae) feeding [Master thesis]. Uppsala: Sveriges Iantbruksuniversitet.
Bylund, H., Nordlander, G., and Nordenhem, H. (2004). Feeding and oviposition rates in the pine weevil Hylobius abietis (Coleoptera: Curculionidae). Bull. Entomol. Res. 94, 307–317. doi: 10.1079/BER2004304
CAB International. (2003). Distribution Maps of Plant Pests, No. 643. Hylobius abietis. Wallingford: CABI Publishing.
Carpenter, S. E., Harmon, M. E., Ingham, E. R., Kelsey, R. G., Lattin, J. D., Schowalter, T. D., et al. (1988). Early patterns of heterotroph activity in conifer logs. Proc. R. Soc. Edinb. B. Biol. Sci. 94, 33–43. doi: 10.1017/S0269727000007089
Day, K. R., Nordlander, G., Kenis, M., and Halldorson, G. (2004). “General biology and life cycles of bark weevils,” in Bark and Wood Boring Insects in Living Trees in Europe, A Synthesis, eds. F. Lieutier, K.R. Day, A. Battisti, J.C. Grégoire and H.F. Evans (Dordrecht: Springer), 331–349.
Doležal, P., Kleinová, L., and Davídková, M. (2021). Adult feeding preference and fecundity in the large pine weevil, Hylobius abietis (Coleoptera: Curculionidae). Insects 12:473. doi: 10.3390/insects12050473
Doom, D., and Frenken, G. W. P. (1980). Postponement of replanting as a silvicultural method to prevent damage by the large pine weevil Curculio Hylobius abietis. Ned. Bosb. Tijdschr. 52, 217–227.
Eidmann, H. H. (1979). Integrated Management of Pine Weevil (Hylobius abietis L.) Populations in Sweden. Uppsala: USDA Forest Service General Technical Report WO.
Fedderwitz, F., Björklund, N., Ninkovic, V., and Nordlander, G. (2018). Does the pine weevil (Hylobius abietis) prefer conifer seedlings over other main food sources? Silva Fenn. 52:9946. doi: 10.14214/sf.9946
Galko, J., Lalík, M., Rell, S., Nikolov, C., Barta, M., Pittner, J., et al. (2022). Comprehensive comparison of treatments for controlling the large pine weevil (Hylobius abietis) in Central Europe. Sci. Rep. 12, 9673–9673. doi: 10.1038/s41598-022-13729-6
Graham, M. H. (2003). Confronting multicollinearity in ecological multiple regression. Ecology 84, 2809–2815. doi: 10.1890/02-3114
Hansen, L. W., Ravn, H. P., and Geldmann, J. (2005). Within- and between-stand distribution of attacks by pine weevil [Hylobius abietis (L.)]. Scand. J. For. Res. 20, 122–129. doi: 10.1080/02827580510008284
Hartig, F. (2021). Package ‘DHARMa'. Available online at: https://ftp.onet.pl/pub/mirrors/CRAN/web/packages/DHARMa/DHARMa.pdf (accessed June 3, 2024).
Havukkala, I. (1979). The humidity reactions of the large pine weevil, Hylobius abietis (Coleoptera, Curculionidae), during three stages of its life cycle. Ann. Ent. Fenn. 42, 59–64.
Havukkala, I., and Selander, J. (1976). Reactions of the large pine weevil, Hylobius abietis L. (Col., Curculionidae), to various light and humidity stimuli during three stages of its life cycle. Ann. Ent. Fenn. 42, 54–62.
Heber, T., Helbig, C. E., Osmers, S., Larquette, M., and Müller, M. G. (2024). Evaluation of trap type and attractant composition for potential mass trapping of Hylobius abietis. Agric. For. Entomol. 26, 1–17. doi: 10.1111/afe.12593
Heiberger, R. M. (2019). Package ‘HH'. Available online at: https://mran.microsoft.com/snapshot/2019–11-29/web/packages/HH/HH.pdf (accessed November 29, 2019).
Holuša, J., Lubojacký, J., Curn, V., Tonka, T., Lukášová, K., Horák, J., et al. (2018). Combined effects of drought stress and Armillaria infection on tree mortality in Norway spruce plantations. For. Ecol. Manag. 427, 434–445. doi: 10.1016/j.foreco.2018.01.031
Inward, D. J. G., Wainhouse, D., and Peace, A. (2012). The effect of temperature on the development and life cycle regulation of the pine weevil Hylobius abietis and the potential impacts of climate change. Agric. For. Entomol. 14, 348–357. doi: 10.1111/j.1461-9563.2012.00575.x
Jactel, H., Nicoll, B. C., Branco, M., Gonzalez-Olabarria, J. R., Grodzki, W., Långström, B., et al. (2009). Influences de la sylviculture sur le risque de dégâts biotiques et abiotiques dans les peuplements forestiers. Ann. For. Sci. 66, 701–701. doi: 10.1051/forest/2009054
Johansson, K., Hajek, J., Sjölin, O., and Normark, E. (2015). Early performance of Pinus sylvestris and Picea abies – a comparison between seedling size, species, and geographic location of the planting site. Scand. J. For. Res. 30, 388–400. doi: 10.1080/02827581.2014.987808
Långström, B. (1982). Life Cycles and Shoot-Feeding of the Pine Shoot Beetles (No. 163). Alnarp: Sveriges Lantbruksuniversitet.
Långström, B., and Day, K. R. (2004). “Damage, control and management of weevil pests, especially Hylobius abietis,” in Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis, eds. F. Lieutier, K.R. Day, A. Battisti, J.C. Grégoire and H.F. Evans (Springer: Dordrecht), 415–444.
Lalík, M., Galko, J., Kunca, A., Nikolov, C., Rell, S., Zúbrik, M., et al. (2021). Ecology, management and damage by the large pine weevil (Coleoptera: Curculionidae) in coniferous forests within Europe. Cent. Eur. For. J. 67, 91–107. doi: 10.2478/forj-2021-0005
Lalík, M., Holuša, J., Galko, J., Resnerov,á, K., Kunca, A., Nikolov, C., et al. (2019). Simple is best: pine twigs are better than artificial lures for trapping of pine weevils in pitfall traps. Forests 10:642. doi: 10.3390/f10080642
Leather, S. R., Ahmed, S. I., and Hogan, L. (1994). Adult feeding preferences of the large pine weevil, Hylobius abietis (Coleoptera: Curculionidae). Eur. J. Entomol. 91:385.
Leather, S. R., Day, K. R., and Salisbury, A. N. (1999). The biology and ecology of the large pine weevil, Hylobius abietis (Coleoptera: Curculionidae): a problem of dispersal? Bull. Entomol. Res. 89, 3–16. doi: 10.1017/S0007485399000024
Lindgren, B. S., Nordlander, G., and Birgersson, G. (1996). Feeding deterrence of verbenone to the pine weevil, Hylobius abietis (L.) (Col., Curculionidae). J. Appl. Entomol. 120, 397–403. doi: 10.1111/j.1439-0418.1996.tb01627.x
López-Villamor, A., Carreño, S., López-Goldar, X., Suárez-Vidal, E., Sampedro, L., Nordlander, G., et al. (2019). Risk of damage by the pine weevil Hylobius abietis in southern Europe: effects of silvicultural and landscape factors. For. Ecol. Manag. 444, 290–298. doi: 10.1016/j.foreco.2019.04.027
Luik, A., and Voolma, K. (1989). “Some aspects of the occurrence, biology and cold-hardiness of Hylobius weevils. Insects affecting reforestation: biology and damages,” in Insects Affecting Reforestation: Biology and Damage, eds. R.I. Alfaro and S.G. Glover (Victoria, BC: Pacific and Yukon Region, Forestry Canada), 28–33.
Månsson, P. E., and Schlyter, F. (2004). Hylobius pine weevils adult host selection and antifeedants: feeding behaviour on host and non-host woody scandinavian plants. Agric. For. Entomol. 6, 165–171. doi: 10.1111/j.1461-9563.2004.00217.x
Manlove, J. D., Styles, J., and Leather, S. R. (2013). Feeding of the adults of the large pine weevil, Hylobius abietis (Coleoptera: Curculionidae). Eur. J. Entomol. 94, 153–156.
Mezei, P., BlaŽenec, M., Grodzki, W., Škvarenina, J., and Jakuš, R. (2017). Influence of different forest protection strategies on spruce tree mortality during a bark beetle outbreak. Ann. For. Sci. 74, 1–12. doi: 10.1007/s13595-017-0663-9
Moore, R., Brixey, J. M., and Milner, A. D. (2004). Effect of time of year on the development of immature stages of the Large Pine Weevil (Hylobius abietis L.) in stumps of Sitka spruce (Picea sitchensis Carr.) and influence of felling date on their growth, density and distribution. J. Appl. Entomol. 128, 167–176. doi: 10.1111/j.1439-0418.2004.00828.x
Munro, J. W. (1928). The biology and control of Hylobius abietis, L. For. Int. J. For. Res. 2, 31–39. doi: 10.1093/oxfordjournals.forestry.a063077
Nordenhem, H. (1989). Age, sexual development, and seasonal occurrence of the pine weevil Hylobius abietis (L.). J. Appl. Entomol. 108, 260–270. doi: 10.1111/j.1439-0418.1989.tb00456.x
Nordenhem, H., and Eidmann, H. H. (1991). Response of the pine weevil Hylobius abietis L. (Col., Curculionidae) to host volatiles in different phases of its adult life cycle. J. Appl. Entomol. 112, 353–358. doi: 10.1111/j.1439-0418.1991.tb01067.x
Nordenhem, H., and Nordlander, G. (1994). Olfactory oriented migration through soil by root-living Hylobius abietis (L.) larvae (Col., Curculionidae). J. Appl. Entomol. 117, 457–462. doi: 10.1111/j.1439-0418.1994.tb00762.x
Nordlander, G. (1987). A method for trapping Hylobius abietis (L.) with a standardized bait and its potential for forecasting seedling damage. Scand. J. For. Res. 2, 199–213. doi: 10.1080/02827588709382458
Nordlander, G. (1990). Limonene inhibits attraction to α-pinene in the pine weevils Hylobius abietis and H. pinastri. J. Chem. Ecol. 16, 1307–1320. doi: 10.1007/BF01021028
Nordlander, G. (1991). Host finding in the pine weevil Hylobius abietis: effects of conifer volatiles and added limonene. Entomol. Exp. Appl. 59, 229–237. doi: 10.1111/j.1570-7458.1991.tb01507.x
Nordlander, G., Björklund, N., Hellqvist, C., Nordenhem, H., Liziniewicz, M., Hjelm, K., et al. (2023). Trap catch data are poor predictors of damage caused by pine weevil (Hylobius abietis) to conifer seedlings. For. Ecol. Manag. 537:120968. doi: 10.1016/j.foreco.2023.120968
Nordlander, G., Bylund, H., Örlander, G., and Wallertz, K. (2003). Pine weevil population density and damage to coniferous seedlings in a regeneration area with and without shelterwood. Scand. J. For. Res. 18, 438–448. doi: 10.1080/02827580310001634
Nordlander, G., Hellqvist, C., Johansson, K., and Nordenhem, H. (2011). Regeneration of European boreal forests: effectiveness of measures against seedling mortality caused by the pine weevil Hylobius abietis. For. Ecol. Manag. 262, 2354–2363. doi: 10.1016/j.foreco.2011.08.033
Nordlander, G., Nordenhem, H., and Bylund, H. (1997). Oviposition patterns of the pine weevil Hylobius abietis. Entomol. Exp. Appl. 85, 1–9. doi: 10.1046/j.1570-7458.1997.00229.x
Örlander, G., and Nilsson, U. (1999). Effect of reforestation methods on pine weevil (Hylobius abietis) damage and seedling survival. Scand. J. For. Res. 14, 341–354. doi: 10.1080/02827589950152665
Örlander, G., Nilsson, U., and Nordlander, G. (1997). Pine weevil abundance on clear-cuttings of different ages: a 6-year study using pitfall traps. Scand. J. For. Res. 12, 225–240. doi: 10.1080/02827589709355405
Örlander, G., Nordlander, G., Wallertz, K., and Nordenhem, H. (2000). Feeding in the crowns of scots pine trees by the pine weevil Hylobius abietis. Scand. J. For. Res. 15, 194–201. doi: 10.1080/028275800750015000
Paraschiv, M. (2020). Risk assessment of damage by Hylobius abietis (L.) on forest sites to be planted. Rev. Silvic. Cineg. 25, 11–15.
Rahman, A., Viiri, H., and Tikkanen, O. P. (2018). Is stump removal for bioenergy production effective in reducing pine weevil (Hylobius abietis) and Hylastes spp. breeding and feeding activities at regeneration sites? For. Ecol. Manag. 424, 184–190. doi: 10.1016/j.foreco.2018.05.003
Saarinen, V. M. (2006). The effects of slash and stump removal on productivity and quality of forest regeneration operations—preliminary results. Biomass Bioenerg. 30, 349–356. doi: 10.1016/j.biombioe.2005.07.014
Skrzecz, I. (2017). “Insects associated with reforestation and their management in Poland,” in Biological Control of Pest and Vector Insects, ed. V.D.C. Shields (Rijeka, Croatia: InTech), 133–168.
Skrzecz, I., Sukovata, L., Jabłoński, T., Sowińska, A., and Szmidla, H. (2021). Spatio-temporal distribution of Hylobius abietis in Scots pine stands – implications for pest monitoring. J. Pest Sci. 94, 1393–1404. doi: 10.1007/s10340-021-01339-6
Solbreck, C. (1980). Dispersal distances of migrating pine weevils, Hylobius abietis, coleoptera: curculionidae. Entomol. Exp. Appl. 28, 123–131. doi: 10.1111/j.1570-7458.1980.tb02997.x
Tan, J. Y., Wainhouse, D., Morgan, G., and Day, K. R. (2011). Interaction between flight, reproductive development and oviposition in the pine weevil Hylobius abietis. Agric. For. Entomol. 13, 149–156. doi: 10.1111/j.1461-9563.2010.00503.x
Thorpe, K. V., and Day, K. R. (2002). The impact of host plant species on the larval development of the large pine weevil Hylobius abietis L. Agric. For. Entomol. 4, 187–194. doi: 10.1046/j.1461-9563.2002.00144.x
Thorsen, Å. A., Mattsson, S., and Weslien, J. (2001). Influence of stem diameter on the survival and growth of containerized Norway spruce seedlings attacked by pine weevils (Hylobius spp.). Scand. J. For. Res. 16, 54–66. doi: 10.1080/028275801300004415
Toivonen, R., and Viiri, H. (2006). Adult large pine weevils Hylobius abietis feed on silver birch Betula pendula even in the presence of conifer seedlings. Agric. For. Entomol. 8, 121–128. doi: 10.1111/j.1461-9563.2006.00290.x
Tudoran, A., Bylund, H., Nordlander, G., Oltean, I., and Puentes, A. (2021). Using associational effects of European beech on Norway spruce to mitigate damage by a forest regeneration pest, the pine weevil Hylobius abietis. For. Ecol. Manag. 486:118980. doi: 10.1016/j.foreco.2021.118980
Véle, A. (2022). Pine or spruce? Comparison of stump suitability for the large pine weevil Hylobius abietis (L.) development. Sylwan 166, 114–122. doi: 10.26202/sylwan.2021102
Venables, W. N., and Ripley, B. D. (2002). Modern Applied Statistics With S, 4th Edn. New Yor, NYk: Springer.
Viewegh, J., Kusbach, A., and Mikeska, M. (2003). Czech forest ecosystem classification. J. Forest Sci. 49, 74–82. doi: 10.17221/4682-JFS
Viiri, H., and Miettinen, O. (2013). Feeding preferences of Hylobius pinastri Gyll. Balt. For. 19, 161–164.
Von Sydow, F. (1997). Abundance of pine weevils (Hylobius abietis) and damage to conifer seedlings in relation to silvicultural practices. Scand. J. For. Res. 12, 157–167. doi: 10.1080/02827589709355397
Von Sydow, F., and Birgersson, G. (1997). Conifer stump condition and pine weevil (Hylobius abietis) reproduction. Can. J. For. Res. 27, 1254–1262. doi: 10.1139/x97-089
Von Sydow, F., and Örlander, G. (1994). The influence of Shelterwood density on Hylobius abietis (L.) occurrence and feeding on planted conifers. Scand. J. For. Res. 9, 367–375. doi: 10.1080/02827589409382853
Voolma, K. (2001). The large pine weevil, Hylobius abietis L., in the felling areas of the Räpina forest district: a case study with baited ground traps. Metsanduslikud Uurimused 35, 172–178.
Voolma, K., and Sibul, I. (2006). The effect of collecting fluid on the catch results of Hylobius abietis in pitfall traps. Agron. Res. 4, 457–460.
Wainhouse, D., Brough, S., and Greenacre, B. (2008). Managing the Pine Weevil on Lowland Pine. Forestry Commission Practice. Note 14. Edinburgh: Forestry Commission.
Wainhouse, D., Inward, D. J. G., and Morgan, G. (2014). Modelling geographical variation in voltinism of Hylobius abietis under climate change and implications for management. Agric. For. Entomol. 16, 136–146. doi: 10.1111/afe.12043
Wainhouse, D., Staley, J. T., Jinks, R., and Morgan, G. (2009). Growth and defence in young pine and spruce and the expression of resistance to a stem-feeding weevil. Oecologia 158, 641–650. doi: 10.1007/s00442-008-1173-0
Wallertz, K., Nordenhem, H., and Nordlander, G. (2014). Damage by the pine weevil Hylobius abietis to seedlings of two native and five introduced tree species in Sweden. Silva Fenn. 48:1188. doi: 10.14214/sf.1188
Wallertz, K., Nordlander, G., and Örlander, G. (2006). Feeding on roots in the humus layer by adult pine weevil, Hylobius abietis. Agric. For. Entomol. 8, 273–279. doi: 10.1111/j.1461-9563.2006.00306.x
Wibe, A., Borg-Karlson, A. K., Persson, M., Norin, T., and Mustaparta, H. (1998). Enantiomeric composition of monoterpene hydrocarbons in some conifers and receptor neuron discrimination of α-pinene and limonene enantiomers in the pine weevil, Hylobius abietis. J. Chem. Ecol. 24, 273–287. doi: 10.1023/A:1022580308414
Willoughby, I., Moore, R., and Nisbet, T. (2017). Interim Guidance on the Integrated Management of Hylobius abietis in UK Forestry. Forest Research Research Note. Available online at: https://www.forestresearch.gov.uk/documents/607/FR_InterimguidanceonmanagementHylobiusabietis_2017.pdf (accessed May 29, 2022).
Keywords: bark beetle, clear-cuts, Hylobius pinastri, pest, seedlings
Citation: Dvořáková B, Holuša J, Horák J, Hradecký J, Bledý M and Zelenka M (2024) Pine weevil (Hylobius abietis) preferences among species of conifer seedlings planted on clear-cuts in central Europe. Front. For. Glob. Change 7:1399405. doi: 10.3389/ffgc.2024.1399405
Received: 11 March 2024; Accepted: 28 May 2024;
Published: 20 June 2024.
Edited by:
Pierluigi Bonello, The Ohio State University, United StatesReviewed by:
Manuela Branco, University of Lisbon, PortugalJuan A. Martin, Polytechnic University of Madrid, Spain
Copyright © 2024 Dvořáková, Holuša, Horák, Hradecký, Bledý and Zelenka. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Barbora Dvořáková, RHZvcmFrb3ZhMUBmbGQuY3p1LmN6