
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. For. Glob. Change , 16 May 2024
Sec. Forest Disturbance
Volume 7 - 2024 | https://doi.org/10.3389/ffgc.2024.1383672
This article is part of the Research Topic Biotic Pest Disturbance - Risk, Evaluation, and Management in Forest Ecosystems View all 15 articles
 Nataliya Korolyova1,2*
Nataliya Korolyova1,2* Jaromír Bláha3
Jaromír Bláha3 Jaromír Hradecký3Jaroslav Kašpar4
Jaromír Hradecký3Jaroslav Kašpar4 Barbora Dvořáková3
Barbora Dvořáková3 Rastislav Jakuš1,2
Rastislav Jakuš1,2This study investigates the efficacy of combined treatment strategy, incorporating pheromones for bark beetle I. typographus (IT) and attractant of its natural enemy T. formicarius (TF), along with anti-attractants for IT (containing 1-hexanol, 1-octen-3-ol, 3-octanol, eucalyptol, trans-thujanol, and trans-conophthorin), to enhance protection methods for Picea abies against biotic disturbances. Two field experiments—trapping experiment and tree protection experiment—were conducted in June 2023 in managed spruce-dominated beetle-affected stands in Czechia. We anticipated higher catches of IT in traps baited with IT pheromone (containing s-ipsdienol, s-cis-verbenol, and 2-methyl-3-buten-2-ol) and TF attractant compared to traps using IT pheromone alone, since compounds intrinsic to IT pheromone, namely 2-methyl-3-buten-2-ol, ipsenol, and ipsdienol, are integral components of the attractant designed for TF. We hypothesized that application of TF attractant and IT anti-attractant would enhance the treatment's protective properties, assuming that attracted TF would function as a predator, reducing bark beetle population and increasing tree survival rates. Semiochemical composition declared by the producers was verified using gas chromatography-mass spectrometry analysis. In the trapping experiment, EcoTrap-type traps were baited with six combinations of lures and anti-attractant. In the tree protection experiment, 28 mature Norway spruce trees situated at newly created forest edges underwent four treatment types: TF attractant, IT anti-attractant, their combination, and no treatment (“control”). Traps baited solely with TF attractant did not capture either beetle, whereas traps lured with IT pheromone, TF attractant and anti-attractant showed no captures of IT but recorded the highest numbers of TF, suggesting significant potential for combined treatment efficacy. Surprisingly, tree mortality was observed exclusively among trees treated only with TF attractant and in their vicinity, suggesting unique bark beetles' response to the mixture of predator's attractant and host tree kairomones, a phenomenon that was not previously reported. Application of anti-attractant and TF treatment effectively prevented tree mortality, demonstrating the repellent potential of IT anti-attractant against bark beetles. However, mortality rates showed no significant differences among control trees, those treated with anti-attractants, or those treated with the combination of anti-attractants and TF attractant, underscoring necessity for further research to optimize treatment efficacy.
The escalating frequency and severity of drought waves and windstorms have induced widespread bark beetle disturbances, significantly impacting vast forested regions (Millar and Stephenson, 2015). In Europe, the preeminent economic pest, the bark beetle Ips typographus (L., 1758), has inflicted damage on tens of millions of Norway spruce trees in recent decades, resulting in pronounced ecological, economic, and social consequences (Senf and Seidl, 2018). Forest owners employ a spectrum of measures to protect their stands and mitigate the bark beetle outbreaks. These strategies encompass the timely removal of infested trees, the implementation of pheromone traps, and the application of anti-attractants (Fettig and Hilszczański, 2015). Nevertheless, the limited cost-effectiveness of such measures over extensive areas during severe outbreaks necessitates a continued search for efficient, economically viable, and environmentally friendly methods to control I. typographus population densities.
In the population control strategy of bark beetles, their natural enemies play a significant role (Wermelinger, 2004; Wegensteiner et al., 2015). One of the key predators of the critical forest pest I. typographus is the clerid beetle Thanasimus formicarius (L., 1758). T. formicarius has been observed in association with numerous species of bark beetles inhabiting both coniferous and deciduous trees (Wehnert and Müller, 2012). The species demonstrates an affinity for bark beetle pheromone components and host tree volatiles (Rudinský et al., 1971; Bakke and Kvamme, 1981; Schroeder and Lindelöw, 1989; Hulcr et al., 2006). Adult predators target adult bark beetles before they bore into host trees and lay their eggs in bark crevices of recently infested trees (Schroeder, 1999). The flight season of T. formicarius usually begins in March or April and lasts for several months (Schroeder, 2003). T. formicarius exhibits flight patterns similar to those of I. typographus, except in early spring. During this period, T. formicarius preys on other bark beetle species whose flight periods begin earlier than the I. typographus one (Schroeder, 1996; Wehnert and Müller, 2012). Adult individuals predominantly feed on adult bark beetles and their larvae, while the larvae of T. formicarius move within the galleries of bark beetles, hunting for bark beetle larvae (Koçoglu and Özcan, 2018). The density of the T. formicarius population positively correlates with the population density of bark beetles. Concurrently, elevated levels of bark beetle population density result in elevated larval mortality due to predation by T. formicarius (Weslien, 1994). Meshkova et al. (2021) demonstrated that the experimental realize of T. formicarius into pine stands resulted in a faster decline of Ips sexdentatus outbreaks compared to the control stands.
The field of chemical ecology pertaining to the interactions between T. formicarius and I. typographus witnessed significant progress since the mid-1980s (Bakke and Kvamme, 1981; Hansen, 1983), with the predominant focus of further research directed toward the examination of pheromone compounds produced by I. typographus, that elicit attraction in T. formicarius (Hulcr et al., 2006; Etxebeste et al., 2012). The chemical composition of I. typographus pheromone encompasses several compounds. The primary examples among them are 2-methyl-3-buten-2-ol (MB) and cis-verbenol (cV), which collectively serve as the principal aggregation pheromones for I. typographus (Vité et al., 1972; Bakke, 1977; Bakke and Kvamme, 1981; Birgersson et al., 1984, 1988). MB specifically influences the orientation of I. typographus at short distances, and has been observed to enhance the likelihood of their landing (Schlyter et al., 1987). Ipsdienol (Id), present in modest quantities in males prior to mating, augments the overall attractiveness of the I. typographus pheromone (Vité et al., 1972; Bakke, 1977; Bakke and Kvamme, 1981; Schlyter et al., 1992). Scientists supposed that 2-phenylethanol could be a minor component of the aggregation pheromone of I. typographus (Birgersson et al., 1984; Sun et al., 2006; Xie and Lv, 2013). Following copulation, males produce ipsenol (Ie), an anti-aggregation pheromone which, in conjunction with verbenone, assumes a pivotal role in regulating the density of bark beetle galleries under the bark. Additionally, at elevated concentrations, these two substances can redirect bark beetle attacks toward neighboring trees (Bakke and Kvamme, 1981; Hansen, 1983; Birgersson et al., 1984, 1988; Sun et al., 2006).
Previous studies have shown that T. formicarius, attracted by the aggregation pheromone of I. typographus, is specifically drawn to cV. MB, the second component of the aggregation pheromone, neither attracts T. formicarius nor enhances the attractiveness of cV when added to the mixture. In contrast, Id is highly attractive for T. formicarius, even more so than the aggregation pheromone of I. typographus (Hulcr et al., 2006). Etxebeste et al. (2012) found that Ie is also a highly attractive semiochemical for T. formicarius. The combination of Id and Ie was reported to be the most attractive mixture for T. formicarius (Bakke and Kvamme, 1981; Hulcr et al., 2006). Hansen (1983) concluded that T. formicarius has olfactory receptors for all four compounds (cV, MB, Ie, Id). Considering that MB has no attractive effect, the kairomonal response of T. formicarius to the mixture of I. typographus pheromonal compounds, cV, Id, and Ie is evident (Hansen, 1983). Tømmerås (1985) found that T. formicarius has highly specialized olfactory receptors on its antennae, specifically tuned to bark beetle pheromones. That author describes receptors that are capable of detecting (+)-ipsdienol, (–)-ipsdienol, (S)-cis-verbenol, (–)-ipsenol, (+)-lineatin, and (–)-verbenon, suggesting that the predator T. formicarius can distinguish among various species of bark beetles. However, it remains unclear how the commercial attractant developed for T. formicarius influences the trap catches of I. typographus and, more specifically, how it modifies the host colonization behavior of I. typographus in natural forest settings.
Numerous compounds that have been proven to deter I. typographus have been previously identified. Verbenone, the first compound, is synthesized either from the host compound α-pinene or by converting cis-verbenol, the primary pheromone component for I. typographus (Birgersson and Leufvén, 1988). Another category encompasses non-host volatiles, such as trans-conophthorin and green leaf alcohols, e.g., 1-hexanol and (Z)-3-hexen-1-ol, commonly found in species like birch and aspen (Zhang et al., 1999). Additionally, C8 alcohols emitted from the barks of these trees act as deterrents. Eucalyptol, a relatively new compound, has demonstrated field effectiveness, exhibiting better precision than verbenone by inhibiting cis-verbenol at the single-sensillum level (Andersson et al., 2010; Binyameen et al., 2014). Recently, oxygenated monoterpenes derived from host trees, such as trans-thujan-4-ol, have been reported to possess anti-attractant properties (Kalinová et al., 2014; Blažytė-Čereškienė et al., 2016; Schiebe et al., 2019; Jirošová et al., 2022). These compounds have been incorporated into dispensers for tree protection, yielding various degrees of success (Jakuš et al., 2003, 2022, 2024; Schiebe et al., 2011; Deganutti et al., 2023). A novel dispenser, developed by Jakuš et al. (2024) and devoid of beetle-derived compounds, has exhibited promising efficiency. However, anti-attractants are still not widely used in practical forest protection measures due to their limited effectiveness and relatively high costs in tree protection.
Zuhlke and Mueller (2008) proposed a method for controlling bark beetle population density by attracting their predators, such as T. formicarius, to their habitats. The authors explored the concept of selective attraction using attractants that contain only some components of the bark beetle's attractant bouquet. They found that even with only one, two or three components, these attractants effectively lured predators of the target bark beetle species. However, if certain specific components are missing, these attractants fail to attract or only minimally attract the target beetles, even in areas where they are abundant (Zuhlke and Mueller, 2008). The potential of using the combined treatment comprising the attractant for T. formicarius and anti-attractant for I. typographus in Norway spruce protection against biotic disturbances has not been experimentally studied in field conditions. Investigating these questions could contribute to the improvement of tree protection measures and strategies involved in forest management pest control.
The aim of this study is to assess the efficacy of the attractant that has been developed for T. formicarius in capturing T. formicarius and I. typographus in pheromone traps and mitigating spruce tree mortality. Additionally, the paper examines the feasibility of using a combined treatment, that consists of the attractant for T. formicarius and anti-attractant for I. typographus, to protect trees against I. typographus attacks. We also aim to develop a basis for further reinforcing the effectiveness of our tree protection method based on the use of anti-attractants (Jakuš et al., 2024). We anticipated observing the highest I. typographus catches in traps baited with the pheromone of I. typographus and attractant developed for T. formicarius. As compounds that are intrinsic to the I. typographus pheromone constitute components of the attractant that was designed for T. formicarius (MB, Ie, Id), we hypothesized that the traps baited with I. typographus pheromone and attractant of T. formicarius would exhibit the highest catches of I. typographus. We also hypothesized that application of both an attractant for T. formicarius and an anti-attractant for I. typographus on spruce trees would enhance the treatment's protective properties, assuming that attracted T. formicarius would prey on potential pioneer bark beetles. We anticipated that the attractant of T. formicarius would not attract I. typographus. If any I. typographus lands on a tree, it would be killed by elevated numbers of T. formicarius, which function as predators, consequently mitigating tree mortality by reducing the bark beetle population density.
Field trapping experiments were established in a forest near the town of Kostelec nad Černými Lesy in Central Bohemia (coordinates 49.9146136° N, 14.8780744° E, altitude 460 m above sea level). The 90-year old forest stand predominantly consisted of P. abies (70%) with a mixture of L. decidua (20%) and P. sylvestris (10%). A recent bark beetle calamity led to the clearing of a gap in the middle of the stand, where a trapping experiment was conducted. The study plot is situated within the area managed by the School Forest Enterprise (SLP) near the town of Kostelec nad Černými Lesy in the Central Bohemian Region of the Czech Republic. The SLP spans approximately 5,700 ha of forest land and is administrated by the Czech University of Life Sciences Prague (CZU). The region experiences mild winters, with average annual temperatures ranging from 7.0 to 7.5°C. Annual precipitation averages 650 mm, and the vegetation season typically lasts from 150 to 160 days (Tolasz et al., 2007). Currently, the area is affected by an I. typographus outbreak, which began after the drought in 2018 (Pirtskhalava-Karpova et al., 2024).
The tree protection experiment was conducted in 40-year old Norway spruce-dominated stands, with spruce comprising 90% of stand composition, situated in the Potštát Forest district, near Vojenský Újezd Libavá1 (VU Libavá) in the Olomouc District, in the north-eastern sector of the Czech Republic (coordinates 49.670319° N, 17.545289° E). The area encompasses the Libavá administrative district, which is designated for military forestry and agricultural activities, and has functioned as the training grounds for the Czech army since 1946. The topography of the region is undulating, with elevations ranging between 500 and 650 m. The average annual air temperature is in the range of 5–6°C, with the average daily temperature during the growing season (April–September) not surpassing 12°C in VU Libavá. Annual precipitation averages between 700 and 800 mm/year (Tolasz et al., 2007). The prevalent monoculture of spruce trees, characterized by a low static stability, renders them susceptible to frequent wind-induced damage. The windstorm of 1991 provoked a sequence of enduring bark beetle infestations. Exacerbated by the impacts of climate change, this infestation induced a significant decline in the forested area. The region's military training activities impose constraints on the implementation of conventional forest management and pest control practices in the Potštát Forest district. In 2018, bark beetles proliferated over extensive spruce stand areas. Simultaneously, a severe drought, affecting the entire Central European region (Buras et al., 2020), induced large-scale forest dieback that persisted into 2019. Preceding the commencement of the experiment, a sanitary felling initiative was undertaken, involving the removal of deceased and infested trees within the stands that were designated for this study.
A commercial pheromone lure, Pheroprax A (BASF GmbH, Germany), was used as attractant for I. typographus. According to the material safety data sheet (MSDS), the dispenser contains s-ipsdienol, s-cis-verbenol, and 2-methyl-3-buten-2-ol. The second attractant dispenser used in our study was ThanasiWit® (Witasek PflanzenSchutz GmbH, Feldkirchen in Kärnten, Austria), that was designed for T. formicarius, which contains 2-methyl-3-buten-2-ol, ipsenol, ipsdienol, and phenylethanol. As an anti-attractant for I. typographus, a pouch dispenser containing green leaf and non-host volatiles 1-hexanol, 1-octen-3-ol, 3-octanol, eucalyptol, trans-thujanol, and trans-conophthorin was produced by Synergy Semiochemicals Corp. (British Columbia, Canada), in accordance with the formula published in Jakuš et al. (2024).
To check the composition of the dispensers, solid phase microextraction (SPME) from the headspace over the dispenser was used. Freshly opened specimen from the dispenser was placed into a 5 L glass jar, which was then sealed using aluminum foil and a lid. Volatile sampling was conducted at room temperature, 5 min after a 5-min incubation. Compound separation was performed using a two-dimensional gas chromatograph coupled with a time-of-flight mass spectrometer (GC × GC-TOF–MS) (Leco Pegasus 4D, LECO Corp., Michigan, USA). The hot split/splitless injector (275°C) was operated in a split mode (100:1 split ratio). Separation of the compounds was performed on two chromatographic columns connected in a consumable-free modulator. The HP-5 MS UI column (0.25 mm i.d., 0.25 μm film thickness) and the VF-17 MS column (1.5 m, 0.1 mm i.d., 0.1 μm film thickness) were employed for the first- and second-dimension separations, respectively. Both columns were manufactured by Agilent Technologies (USA). The temperature programme for separation started at 40°C with a hold time of 2 min, followed by a gradient of 10°C min−1 to 120°C, and then at 20°C min−1 to 300°C with a hold time of 2 min. The secondary oven and the modulator had temperature offsets of 5 and 15°C, respectively. A 5-s modulation period was used. The separated compounds underwent ionization in the ion source of MSD at 70 eV, and full spectral (35–500 Da) information was acquired at 100 Hz. The compounds were identified using mass spectral similarity, and confirmed via retention index comparison. For comparative analyses, mass spectra and retention indexes were referenced from the NIST Mass Spectral Libraries (Mass Spectrometry Data Center, NIST, USA), except in the case of trans-conophthorin, for which the mass spectrum was sourced from Zhao et al. (2019).
Six EcoTrap-type traps were installed in a clearing of a stand that had previously been affected by bark beetles. The traps were situated along the edges of the spruce stand, which featured larch and pine admixture, spaced 15 m apart and positioned 20 m from the forest edge. The traps were baited with the pheromone lure for I. typographus Pheroprax (PhI), attraction lure for T. formicarius Thanasiwit (TA), a lure with a customized mixture of anti-attractants for I. typographus (AI), and their combinations (PhI; TA; AI; PhI + TA; AI + TA; PhI + TA + AI, Table 1). The experiment took place in June 2023, with traps being inspected at intervals of 2–3 days. Bait rotation was implemented using the Latin Square method. After each beetle collection, the count of T. formicarius was recorded, and the estimation of I. typographus numbers was derived from the volume of catches.
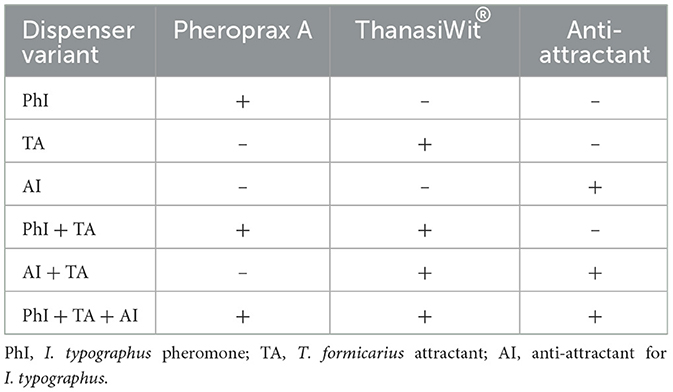
Table 1. Experimental variants (treatment types) used in the trapping experiment.
On 2 June 2023, in the Libavá military forest study area, we conducted a tree protection experiment on 28 mature Norway spruce trees using T. formicarius attractant and I. typographus attractant. We selected visually healthy spruces that were upper-canopy or mid-canopy individuals. To achieve homogeneous experimental conditions, our seven plots and selected trees were situated alongside the extended recently created southern-oriented forest edge, which resulted from salvage cutting immediately preceding the experiment. The average diameter at breast height of the experimental trees was 17 cm, with an average height of 16 m, as indicated in the forest management plan. The goal of the experiment was to investigate the efficiency of using T. formicarius attracatant in protecting Norway spruce trees against I. typographus colonization. We also aimed to test the viability of employing an attractant for T. formicarius, coupled with an anti-attractant for I. typographus, as a preventive measure against mass attacks of I. typographus. In each plot, four trees were treated with four different treatment variants: (A) anti-attractant for I. typographus (AI); (B) attractant for T. formicarius (TA); (C) anti-attractant for I. typographus and attractant for T. formicarius (AI+TA; (D) control (no treatment). The minimal inter-tree distance was 12 m, in order to prevent the potential transfusion of olfactory signals among the treated trees that could potentially obfuscate the results (Schlyter et al., 1987). The plots were spaced 50 m apart. We monitored the statuses of the treated trees (beetle-killed vs. non-attacked) for 4 months.
To compare I. typographus and T. formicarius catches among six different trap treatments (variants) in the Kostelec nad Černými lesy part of the study area, and bark beetle-caused tree mortality among four treatment types in the Libavá military forest part of the study area, we used a one-way ANOVA. The normality of the distribution of residuals in the ANOVA model was checked using the Shapiro-Wilk test from the stats package in R (R development Core Team 2023). The Levene test of the equality of variances from the car package in R was also used. If assumptions regarding the ANOVA model were not met, we used a non-parametric Kruskal-Wallis rank sum test from the stats package in R to compare I. typographus and T. formicarius catches and tree mortality among different treatment types. Dunn's test of multiple comparisons (i.e., a post-hoc test) (FSA package in R) was used to identify the pairs of treatments, for which the catches of I. typographus and T. formicarius, and tree mortality were significantly different. We used Holm's method for the adjustment of p-values regarding multiple comparisons (Holm, 1979). To visualize the results of pairwise comparisons, we quantified a compact letter display at 0.05 significance level using a cldList() function from the rcompanion package in R. To compare the total trap catches between two bark beetle species, we used a non-parametric Mann-Whitney U-test for two independent non-normally distributed groups. All analyses were performed in R.
The results of GCMS analysis, performed for the pheromone designed for I. typograpus, the attractant designed for T. formicarius, and I. typographus anti-attractant revealed 11 compounds detected in the tested dispensers. The identified compounds corresponded to the components declared by the producers of semiochemicals. Particularly, the attractant designed for T. formicarius (ThanasiWit®) incorporated 2-methyl-3-buten-2-ol, ipsenol, ipsdienol, and phenylethanol. Apart from 2-methyl-3-buten-2-ol and ipsdienol, a commercial pheromone lure developed for I. typographus (Pheroprax A) contained verbenol. Anti-attractant components comprised green leaf and non-host volatiles 1-hexanol, 1-octen-3-ol, 3-octanol, eucalyptol, trans-thujanol, and trans-conophthorin (Supplementary Table S1). Two-dimensional chromatographs derived for I. typographus pheromone, T. formicarius attractant, and I. typographus anti-attractant are depicted in the form of contour plots (Figure 1).
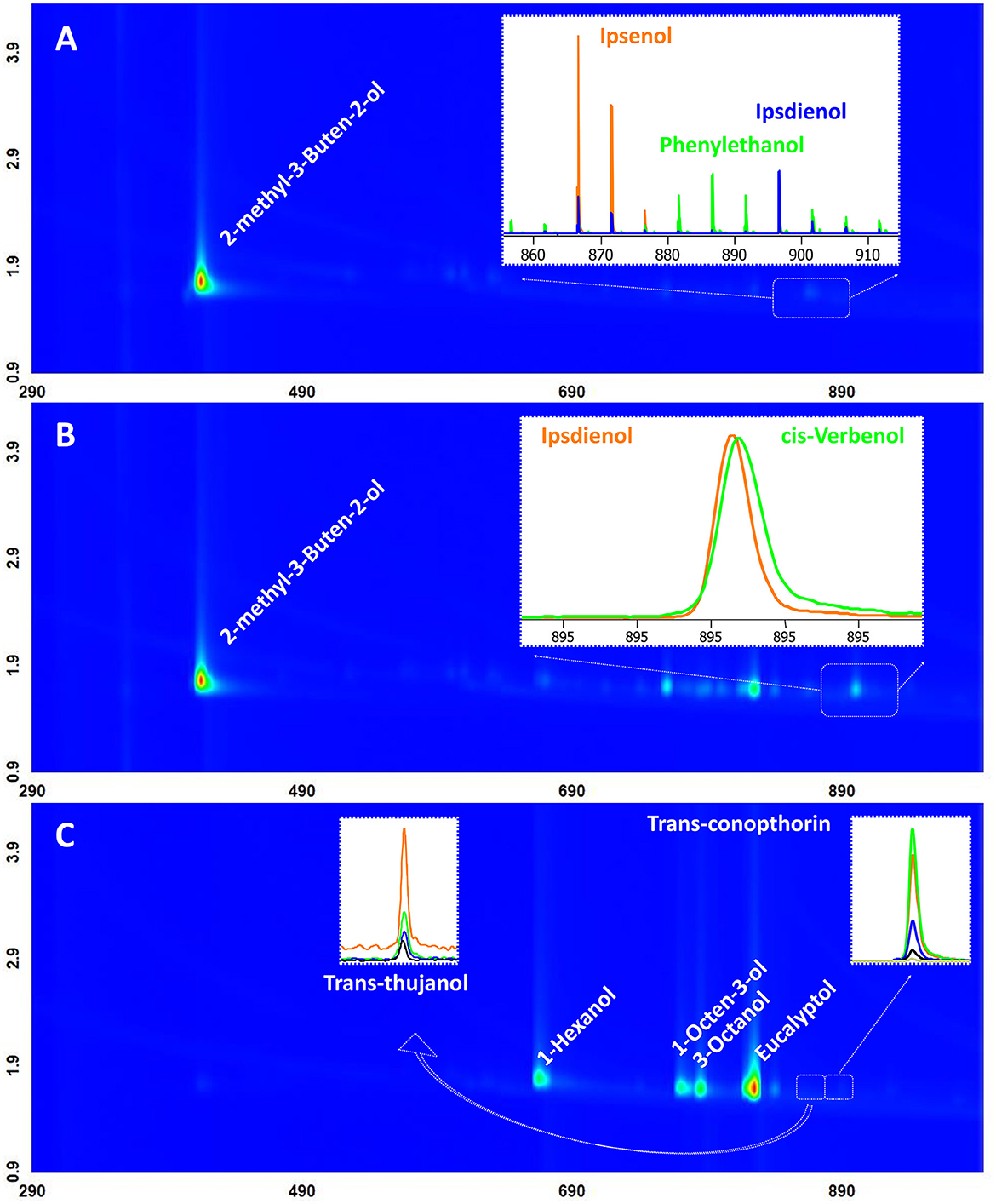
Figure 1. Two-dimensional chromatographs depicting the emissions from the tested dispensers. Panel (A) represents ThanasiWit®, Panel (B) showcases Pheroprax A, and Panel (C) delineates the customized anti-attractant. A total ion chromatogram is utilized for the contour plot. When necessary, different colors are employed to highlight characteristic masses of co-eluting compounds. Specifically, trace amounts of trans-conophthorin are marked with the mass spectra of m/z 84 in orange, 87 in green, 97 in blue, 112 in black, and 156 (molecular ion) in gold color. Additionally, trans-thujanol is indicated with m/z 93 in orange, 121 in green, 136 in blue, and 154 in black. Major signals in the chromatograms were identified as components of the dispenser, aligning with the formulae published in the respective MSDS, or in Jakuš et al. (2024).
The total trap catches of I. typographus measured for all treatments (19,340 beetles) were incomparably larger than the total catches of T. formicarius (25 beetles) (p = 0.013). The number of both I. typographus and T. formicarius catches significantly differed among six treatment types (Supplementary Table S2). Traps baited with I. typographus pheromone caught significantly more I. typographus (PhI) than traps baited with T. formicarius attractant (TA) (Figure 2A; Supplementary Table S3). Correspondingly, larger numbers of I. typographus were caught when PhI was added to TA than when traps were baited with only TA. Conversely, traps baited with TA and PhI did not catch increased numbers of I. typographus compared to traps baited with PhI alone. Similarly, we did not observe significant differences in I. typographus catches between traps baited with I. typographus anti-attractant (AI) and AI + TA. However, the number of catches were significantly higher in traps baited with PhI, TA and AI, compared to traps baited with AI and TA + AI alone (Supplementary Table S3). Traps baited with both species' attractants did not catch significantly larger numbers of I. typographus when AI was added to the traps. We identified significantly smaller number of I. typographus catches in traps baited with anti-attractant and anti-attractant coupled with T. formicarius attractant than in traps lured with the pheromone of I. typographus and attractant of T. formicarius. However, there was no significant difference in I. typographus catches observed in traps lured with the pheromone of I. typographus and attractant for T. formicarius, and traps in which both dispensers were coupled with anti-attractant (Figure 2A; Supplementary Table S3).
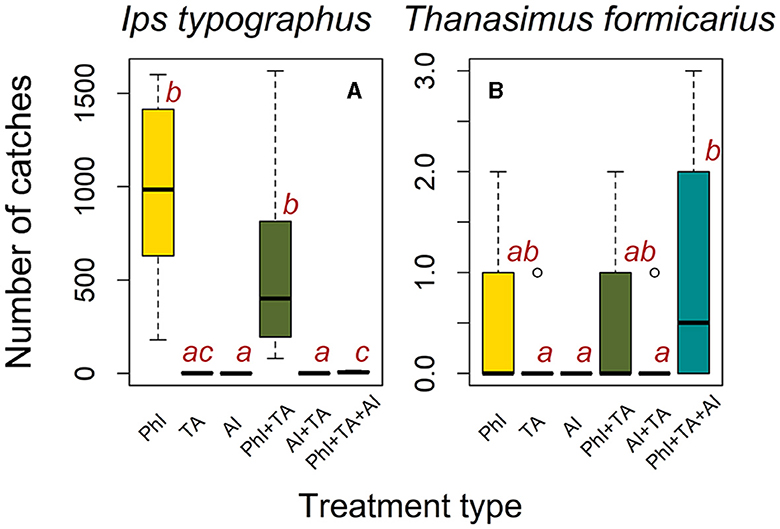
Figure 2. Boxplots showing the numbers of I. typographus (A) and T. formicarius (B) catches in Kostelec nad Černými lesy study area among six treatment types indicated by color. Definitions are PhI, I. typographus pheromone; TA, T. formicarius attractant; AI, I. typographus anti-attractant. The differences in catches among treatment types were checked using the Kruskal-Wallis rank sum test. Pairwise comparisons were performed using Dunn's multiple comparison (post-hoc) test with Holm's adjustment of p-values for multiple comparisons. Compact letter display (lowercase letters “a”–“c” and their combinations) indicates (in)significance in catches between treatment pairs at 0.05 significance level. If any two given treatment types within a panel share at least one common letter, the number of catches does not significantly differ between them. Conversely, if any two given treatment types within a panel do not share any common letters, the number of catches significantly varies between them. The horizontal lines inside the boxplots correspond to the median catch values. The boxes display the interquartile range, which represents the middle 50% of the data. The error bars are the 95% confidence intervals.
T. formicarius catches did not vary significantly among the treatments, except for the catches recorded in traps treated with I. typographus anti-attractant vs. traps treated with I. typographus pheromone, attractant for T. formicarius, and anti-attractant (Figure 2B; Supplementary Table S3), for which the highest number of T. formicarius catches was observed. The count of beetles in traps treated with both I. typographus pheromone, attractant for T. formicarius, and anti-attractant exceeded the corresponding value in traps without anti-attractant, yet the difference was statistically insignificant (Figure 2B; Supplementary Table S3). Traps baited with the attractant for T. formicarius (TA) failed to capture either T. formicarius or I. typographus.
The results of the tree protection experiment that was conducted in the Libavá military forest part of the study area indicate that the largest number of bark beetle-killed trees was observed for individuals treated with TA (Figure 3). Actually, all trees treated with the attractant developed for T. formicarius were attacked and killed by I. typographus. The number of these trees significantly differed from the mortality rates recorded for the rest of the treatment types (Supplementary Table S4). Bark beetles did not attack individuals treated with AI and AI+TA, and also they did not colonize the untreated trees (i.e., control samples).
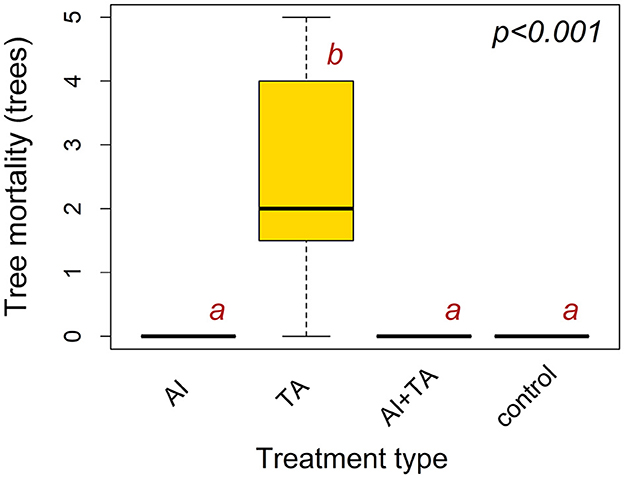
Figure 3. Boxplots showing tree mortality during the tree protection experiment, conducted in the Libavá military forest part of the study area, by treatment type. The y-axis represents the number of bark beetle-killed Norway spruce trees. Definitions: AI is I. typographus anti-attractant, TA is T. formicarius attractant, control designates the absence of treatment. The differences in tree mortality among treatment types were checked using the Kruskal-Wallis rank sum test. Pairwise comparisons were performed using Dunn's multiple comparison (post-hoc) test with Holm's adjustment of p-values for multiple comparisons. Compact letter display (lowercase letters “a”, “b”) indicates (in)significance in tree mortality between the treatment pairs at 0.05 significance level. If any given two treatment types do not share a common letter, the number of bark beetle killed trees significantly differs between them. The horizontal lines inside the boxplot correspond to the median tree mortality value. The box displays the interquartile range, which represents the middle 50% of the data. The error bars are the 95% confidence intervals.
In total, we caught a significantly larger number of I. typographus beetles than T. formicarius beetles, which approximately corresponds to the balance between predators and their prey existing in natural bark beetle communities (Reeve, 1997; Turchin et al., 1999). The T. formicarius/I. typographus ratio observed by us (1/774) and based on the number of catches in the baited traps is consistent with previously published empirical evidence on the same species (Warzée et al., 2006). Previous research (Warzée et al., 2006) has indicated that the proportion of pines within a 500-m radius significantly influences these ratios in traps. In this study, our objective was not to quantify the effects of stand composition on the abundance of predator/prey ratios. Given that our experimental plots were situated in a spruce-monodominated forest, with P. abies comprising nearly 100% of the trees, we aimed to ensure homogeneous experimental conditions, including consistent proportions of host trees, across all plots. Consequently, drawing conclusions about the impact of stand composition on predator/prey ratios and the number of beetle catches would be hindered by the lack of available data resulting from our experimental settings.
Contrasting our expectations, the results of the trapping experiment conducted in the Norway spruce stands in the Kostelec nad Černými lesy part of the study area indicate that traps treated with the I. typographus pheromone and attractant for T. formicarius caught smaller, though insignificantly, numbers of I. typographus compared to the traps treated with I. typographus pheromone alone (Figure 2A). Interestingly, the number of T. formicarius catches also did not vary significantly between the traps treated with the I. typographus pheromone and attractant for T. formicarius, and traps baited with I. typographus pheromone alone (Figure 2B). A larger sample size may be needed in order to detect a more pronounced number of the clerid beetle caught in traps baited with I. typographus pheromone and attractant for T. formicarius. The number of I. typographus catches varied more often among the treatments than the number of T. formicarius catches, presumably due to an inherently smaller overall population density that is commonly observed for T. formicarius, being a natural enemy of I. typographus (Warzée et al., 2006).
Surprisingly, traps treated solely with the attractant designed for T. formicarius failed to capture either T. formicarius or I. typographus. However, in traps treated with both attractants (I. typographus pheromone and T. formicarius attractant) and anti-attractant, no I. typographus were caught, while the highest numbers of T. formicarius were observed (Figure 2). The absence of I. typographus catches in the combined treatment traps may be attributed to the strong repellent effects of tree-based anti-attractants comprising the anti-attractant dispenser (Jakuš et al., 2024). The deterrent effect might have outweighed the luring effect of the I. typographus attractant, despite the latter presumably being enhanced by the presence of Ie, Id, and MB—compounds constituting the T. formicarius attractant dispenser. While, to our knowledge, there is no evidence in the literature in support of T. formicarius attraction to the compounds comprising the anti-attractant, previous studies have reported catches of Thanasimus dubius in traps baited with eucalyptol (Munro et al., 2020), a compound known to act as a repellent for I. typographus (Andersson et al., 2010; Binyameen et al., 2014), and a key component of our anti-attractant mixture. The results of our trapping experiment demonstrate the considerable potential of the combined use of an attractant for T. formicarius and anti-attractants in tree protection.
We recorded tree mortality only in trees baited with attractant for T. formicarius and trees in their proximity, which contradicts our expectations (Figure 3). The influence of T. formicarius on the population dynamics of I. typographus was reported to be substantial (Mills, 1985, 1986; Weslien, 1992; Weslien and Regnander, 1992), primarily owing to its considerable reproductive capacity (106–162 eggs per female) and its significant voracity both in the adult stage (consuming 0.86 to 2–3 adult I. typographus day−1) (Weslien and Regnander, 1992; Faccoli and Stergulc, 2004) and during the larval stage (preying upon 44–57 larvae throughout its entire larval life) (Mills, 1985; Hérard and Mercadier, 1996; Dippel et al., 1997). Thus, we anticipated that the attractant of T. formicarius will not attract I. typographus and if any I. typographus lands on tree, it will be killed by the elevated numbers of T. formicarius, functioning as a predator. The mortality of individuals treated with attractant designed for T. formicarius was possibly caused by a synergistic effect of spruce primary attractants and the components of T. formicarius attractant (2-methyl-3-buten-2-ol, ipsenol, ipsdienol) on I. typographus colonization behavior. To the best of our knowledge, the previously published literature does not provide evidence for a spruce mortality increase in response to such a mixture of compounds. However, traps baited with T. formicarius attractant did not catch either of the beetles, supposedly due to the low population densities of the clerid beetle and potentially higher comparative attractiveness of the traps baited with both I. typographus pheromone and T. formicarius attractant. The absence of tree mortality observed among the control trees in our experiment may be attributed to the relatively low bark beetle population in the study area, notwithstanding the fact that it is sufficient to cause mortality among individuals treated with T. formicarius attractant.
We observed that trees treated with a combination of anti-attractant and T. formicarius attractant were not affected by the bark beetles (Figure 3). This outcome suggests that the repelling effect of the anti-attractant, composed of green leaf and non-host volatiles (1-hexanol, 1-octen-3-ol, 3-octanol, eucalyptol, trans-thujanol, and trans-conophthorin), could be strong enough to overwhelm the attractiveness of the compounds that are present in the T. formicarius attractant. The absence of variation in tree mortality rates among the control trees, trees treated with anti-attractants, or trees treated with a combination of anti-attractant and T. formicarius attractant, hinders a comprehensive understanding of the effects of the combined treatment. Results from trapping experiments suggest that the combined treatment could held the greatest potential for improving tree protection methods. To unravel the mechanisms underlying the efficacy of the combined treatment, further experiments employing pheromone traps would be required that are aimed at identification of the optimal composition of dispensers with compounds repelling I. typographus and attracting T. formicarius.
The number of replications (7) in the tree protection experiment was largely constrained by the availability of suitable forest edges in the study area. We acknowledge that the number of replications used in this study may be perceived as relatively small. However, it is comparable to the number of replications used in similar tree protection experiments conducted in spruce stands in the field of chemical ecology of bark beetles, typically ranging from 10 to 25 (Christiansen and Krokene, 1999; Graves et al., 2008; Mageroy et al., 2020). Our results showed statistically significant differences in the number of bark beetle-killed trees among the treatment types, suggesting that the number of replications implemented can be deemed adequate. Considering the observed pattern of bark beetle attacks experienced exclusively by the individuals treated with T. formicarius attractant, we assume that increasing the number of replications would be unlikely to alter the statistical significance of our findings.
The second limitation of our tree protection experiment is the absence of a variant involving the attachment of I. typographus pheromone dispensers to the spruce trees. Application of this treatment would likely result in augmented numbers of bark beetle attacks on all treated trees, a pattern reported in previous studies, especially in forest edge conditions exposed to relatively high bark beetle pressure (Mulock and Christiansen, 1986; Weslien et al., 1989; Hübertz et al., 1991). The potential outcomes of using I. typographus pheromone dispensers are predictable and suggest significant infestations, including the possibility of widespread I. typographus proliferation throughout the entire stand, which could disrupt the homogeneous experimental conditions established for the rest of the treatments. Additionally, obtaining permission from the local authorities to apply such treatment would be challenging, if not impossible. Overall, implementing this variant correctly would require significant alterations to the experimental design, including a substantial increase in the spacing between experimental trees, which was unfeasible due to constrained availability of suitable forest edges in the study area.
We have made every effort to maintain homogeneous experimental conditions, employing the maximum number of replications feasible in our study area, and applying all permitted treatment types that would not potentially bias the outcome. We argue that the absence of tree mortality in all other groups, except for the individuals treated with T. formicarius attractant, could be explained by the luring effect of the T. formicarius attractant, coupled with host tree volatiles, on bark beetle behavior. Contrary to our expectations, this effect could overwhelm the anticipated predatory activity of the increased numbers of T. formicarius presumed to be lured by the attractant designed for this species. Individuals treated with attractant for T. formicarius seem to be more attractive for I. typographus than stressed untreated forest edge trees that escaped infestation.
Our experiments demonstrate the potential to enhance the efficacy of anti-attractant treatments for tree protection by combining I. typographus anti-attractant dispensers with attractants designed for T. formicarius. Another implication of our findings for enhancing forest management practices suggests that attractant dispensers designed for T. formicarius should not be employed to boost predator numbers in the absence of simultaneously applied I. typographus anti-attractant, which repels bark beetles. The green leaf and non-host volatiles emitted by the anti-attractant act as deterrents for bark beetles (Zhang and Schlyter, 2003, 2010; Unelius et al., 2014). The deterrent effect could outweigh the attractive influence of the T. formicarius attractant combined with host tree volatiles on I. typographus aggregation behavior, as suggested this study. However, we contend that additional development and field testing of semiochemical mixtures are necessary to clarify the potential effects of the attractants designed for both beetle species, I. typographus anti-attractant, and host and non-host volatiles on the beetles' behavior. Specifically, for the enhancement of forest protection measures, it would be advantageous to investigate how the behavior of I. typographus and T. formicarius under field conditions modifies, with different proportions of constitutive compounds in admixtures, varying bark beetle population densities, and predator–prey ratios.
The application of combined dispensers may prove particularly effective in the later stages of bark beetle gradation, where higher population densities of bark beetle predators are anticipated (Weslien, 1994). This approach may be beneficial in localities near unmanaged areas, where we expect a higher population of bark beetle predators compared to managed stands (Weslien and Schroeder, 1999). Additionally, ThanasiWit®, a T. formicarius dispenser employed in our experiment, can be used for attracting the clerid beetles to localized areas of infestation and to wood stacks in order to diminish bark beetle populations, as well as to prevent the predator from being caught in pheromone traps. One possible direction for further development of an improved dispenser could involve using compounds that have not demonstrated any potential attraction to I. typographus but are attractive to T. formicarius. According to Zuhlke and Mueller (2008), chemical compounds comprising the pheromones of various bark beetle species, including those attacking broad-leaved trees, could potentially be used to attract T. formicarius to its prey, I. typographus, without simultaneously elevating the risk of bark beetle infestations in host trees.
Our trapping experiment has shown that traps baited with a combination of anti-attractant for I. typographus and attractant for T. formicarius did not catch any I. typographus. Concurrently, these traps caught the highest numbers of T. formicarius specimens. This synergistic combination indicates promising potential for enhancing tree protection measures. However, the observed mortality of Norway spruce, exclusively in trees treated with attractant dispensers designed for T. formicarius, highlights potential risks associated with such applications. This suggests that further investigation is necessary to optimize the composition and compound proportions of the combined dispenser.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
NK: Data curation, Formal analysis, Investigation, Software, Visualization, Writing—original draft, Writing—review & editing. JB: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Writing—original draft, Writing—review & editing. JH: Investigation, Visualization, Writing—original draft, Writing—review & editing. JK: Conceptualization, Data curation, Investigation, Methodology, Writing—review & editing. BD: Investigation, Visualization, Writing—review & editing. RJ: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Writing—original draft, Writing—review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was supported by grants “Development of integrated modern and innovative diagnostic and protection methods of spruce stands with the use of semiochemicals and methods of molecular biology”, No. QK1910480 funded by the Ministry of Agriculture of Czech Republic; “Effects of host and non-host volatiles on pheromone attraction of the beech bark beetle Taprorychus bicolor” funded by internal grant agency of the Faculty of Forestry and Wood Sciences, Czech University of Life Sciences Prague; “Norway spruce (Picea abies) resistance to bark beetle attack in condition of changing climate”, VEGA−2/0155/22 funded by the Scientific Grant Agency of the Ministry of Education, Science, Research, and Sport of the Slovak Republic and the Slovak Academy of Sciences.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2024.1383672/full#supplementary-material
1. ^Available online at: https://www.vojujezd-libava.cz/vismo/dokumenty2.asp?u=9342&id_org=9342&id=3381 (accessed January 20, 2024).
Andersson, M. N., Larsson, M. C., Blaženec, M., Jakuš, R., Zhang, Q. H., and Schlyter, F. (2010). Peripheral modulation of pheromone response by inhibitory host compound in a beetle. J. Exp. Biol. 213, 3332–3339. doi: 10.1242/jeb.044396
Bakke, A. (1977). Field response to a new pheromonal compound isolated from Ips typographus. Naturwisenschaften 64, 98–99. doi: 10.1007/BF00437364
Bakke, A., and Kvamme, T. (1981). Kairomone response in Thanasimus predators to pheromone components of Ips typographus. J. Chem. Ecol. 7, 305–312. doi: 10.1007/BF00995753
Binyameen, M., Jankuvová, J., Blaženec, M., Jakuš, R., Song, L., Schlyter, F., et al. (2014). Co-localization of insect olfactory sensory cells improves the discrimination of closely separated odour sources. Funct. Ecol. 28, 1216–1223. doi: 10.1111/1365-2435.12252
Birgersson, G., and Leufvén, A. (1988). The influence of host tree response to Ips typographus and fungal attack on production of semiochemicals. Insect Biochem. 18, 761–770. doi: 10.1016/0020-1790(88)90098-4
Birgersson, G., Schlyter, F., Bergström, G., and Löfqvist, J. (1988). Individual variation in aggregation pheromone content of the bark beetle Ips typographus. J. Chem. Ecol. 14, 1737–1761. doi: 10.1007/BF01014641
Birgersson, G., Schlyter, F., Lofqvist, J., and Bergstrom, G. (1984). Quantitative variation of pheromone components in spruce bark beetle Ips typographus from different attack phases. J. Chem. Ecol. 10, 1029–1055. doi: 10.1007/BF00987511
Blažytė-Čereškienė, L., Apšegaite, V., Radžiute, S., Mozuraitis, R., Buda, V., and Pečiulyte, D. (2016). Electrophysiological and behavioral responses of Ips typographus (L.) to trans-4-thujanol—a host tree volatile compound. Ann. For. Sci. 73, 247–256. doi: 10.1007/s13595-015-0494-5
Buras, A., Rammig, A. S., and Zang, C. (2020). Quantifying impacts of the 2018 drought on European ecosystems in comparison to 2003. Biogeosciences 17, 1655–1672. doi: 10.5194/bg-17-1655-2020
Christiansen, E., and Krokene, P. (1999). Can Norway spruce trees be ‘vaccinated' against attack by Ips typographus? Agric. For. Entomol. 1, 185–187. doi: 10.1046/j.1461-9563.1999.00024.x
Deganutti, L., Biscontin, F., Bernardinelli, I., and Faccoli, M. (2023). The semiochemical push-and-pull technique can reduce bark beetle damage in disturbed Norway spruce forests affected by the Vaia storm. Agric. For. Entomol. 26, 115–125. doi: 10.1111/afe.12600
Dippel, C., Heidger, C., Nicolai, V., and Simon, M. (1997). The influence of four different predators on bark beetles in European forest ecosystems (Coleoptera: Scolytidae). Ent. Gen. 21, 161–175. doi: 10.1127/entom.gen/21/1997/161
Etxebeste, I., Álvarez, G., Pérez, G., and Pajares, J. A. (2012). Field response of the six-toothed pine bark beetle, Ips sexdentatus (Col.: Curculionidae, Scolytinae), to pheromonal blend candidates. J. Appl. Entom. 136, 431–444. doi: 10.1111/j.1439-0418.2011.01682.x
Faccoli, M., and Stergulc, F. (2004). Ips typographus (L.) pheromone trapping in south Alps: spring catches determine damage thresholds. JEN 128, 307–311. doi: 10.1111/j.1439-0418.2004.00848.x
Fettig, C. J., and Hilszczański, J. (2015). Management Strategies for Bark Beetles in Conifer Forests, in: Bark Beetles: Biology and Ecology of Native and Invasive Species. Cambridge, MA: Academic Press, 555–584. doi: 10.1016/B978-0-12-417156-5.00014-9
Graves, A. D., Holsten, E. H., Ascerno, M. E., Zogas, K. P., Hard, J. S., Huber, D. P. W., et al. (2008). Protection of spruce from colonization by the bark beetle, Ips perturbatus, in Alaska. For. Ecol. Man. 256, 1825–1839. doi: 10.1016/j.foreco.2008.07.008
Hansen, K. (1983). Reception of bark beetle pheromone in the predaceous clerid beetle, Thanasimus formicarius (Coleoptera: Cleridae). J. Comp. Physiol. 150, 371–378. doi: 10.1007/BF00605026
Hérard, F., and Mercadier, G. (1996). Natural enemies of Tomicus piniperda and Ips acuminatus (Col., Scolytidae) on Pinus sylvestris near Orléans, France: temporal occurrence and relative abundance, and notes on eight predatory species. Entomophaga 41, 183–210. doi: 10.1007/BF02764245
Hübertz, H., Larsen, J. R., and Bejer, B. (1991). Monitoring spruce bark beetle (Ips typographus (L.)) populations under non-epidemic conditions. Scand. J. For. Res. 6, 217–226. doi: 10.1080/02827589109382663
Hulcr, J., Ubik, K., and Vrkoc, J. (2006). The role of semiochemicals in tritrophic interactions between the spruce bark beetle Ips typographus, its predators and infested spruce. J. Appl. Entomol. 130, 275–283. doi: 10.1111/j.1439-0418.2006.01069.x
Jakuš, R., Modlinger, R., Kašpar, J., Majdak, A., Blaženec, M., Korolyova, N., et al. (2022). Testing the efficiency of the push-and-pull strategy during severe Ips typographus outbreak and extreme drought in Norway spruce stands. Forests 13:2175. doi: 10.3390/f13122175
Jakuš, R., Schlyter, F., Zhang, Q. H., Blaženec, M., Vaverčák, R., Grodzki, W., et al. (2003). Overview of development of an anti-attractant based technology for spruce protection against Ips typographus: from past failures to future success. Anzeiger Schadlingskd 76, 89–99. doi: 10.1046/j.1439-0280.2003.03020.x
Jakuš, R., Trubin, A., Singh, V. V., Zabihi, K., Jirošová, A., Modlinger, R., et al. (2024). Spruce protection against Ips typographus with anti-attractant blend of tree-based semiochemicals: from small experimental plots to stand scales. Forests. 15:356. doi: 10.3390/f15020356
Jirošová, A., Kalinová, B., Modlinger, R., Jakuš, R., Unelius, C. R., Blaženec, M., et al. (2022). Anti-attractant activity of (+)-trans-4-thujanol for Eurasian spruce bark beetle Ips typographus: novel potency for females. Pest Manag. Sci. 78, 1992–1999. doi: 10.1002/ps.6819
Kalinová, B., Brízová, R., Knížek, M., Turčáni, M., and Hoskovec, M. (2014). Volatiles from spruce trap-trees detected by Ips typographus bark beetles: chemical and electrophysiological analyses. Arthropod. Plant Interact. 8, 305–316. doi: 10.1007/s11829-014-9310-7
Koçoglu, N., and Özcan, G. E. (2018). Feeding preferences of the rearing of Thanasimus formicarius (L.) (Coleoptera, Cleridae). Alinteri Zirai Bilimler Dergisi 33, 215–220. doi: 10.28955/alinterizbd.449574
Mageroy, M. H., Christiansen, E., Långström, B., Borg-Karlson, A. K., Solheim, H., Björklund, N., et al. (2020). Priming of inducible defenses protects Norway spruce against tree-killing bark beetles. Plant Cell Environ. 43, 420–430. doi: 10.1111/pce.13661
Meshkova, V. L., Ridkokasha, A. D., Omelich, A. R., and Baturkin, D. O. (2021). The first results of the biological control of Ips sexdentatus using Thanasimus formicarius in Ukraine. For. For. Melior. 138, 91–96. doi: 10.33220/1026-3365.138.2021.91
Millar, C. I., and Stephenson, N. L. (2015). Temperate forest health in an era of emerging megadisturbance. Science 349, 823–826. doi: 10.1126/science.aaa9933
Mills, N. J. (1985). Some observations on the role of predation in the natural regulation of Ips typographus populations. Z. Angew. Entomol. 99, 209–215. doi: 10.1111/j.1439-0418.1985.tb01980.x
Mills, N. J. (1986). A preliminary analysis of the dynamics of within tree populations of Ips typographus (L.) (Col.: Scolytidae). J. Appl. Entomol. 102, 402–416. doi: 10.1111/j.1439-0418.1986.tb00938.x
Mulock, P., and Christiansen, E. (1986). The threshold of succesful attack by Ips typographus on Picea abies: a field experiment. For. Ecol. Manage. 14, 125–132. doi: 10.1016/0378-1127(86)90097-6
Munro, H. L., Gandhi, K. J. K., Barnes, B. F., Montes, C. R., Nowak, J. T., Shepherd, W. P., et al. (2020). Electrophysiological and behavioral responses Dendroctonus frontalis and D. terebrans (Coleoptera: Curculionidae) to resin odors of host pines (Pinus spp.). Chemoecology 30, 215–223. doi: 10.1007/s00049-020-00311-7
Pirtskhalava-Karpova, N., Trubin, A., Karpov, A., and Jakuš, R. (2024). Drought initialised bark beetle outbreak in Central Europe: meteorological factors and infestation dynamic. For. Ecol. Manage. 554:121666. doi: 10.1016/j.foreco.2023.121666
Reeve, J. D. (1997). Predation and bark-beetle dynamics. Oecologia 112, 48–54. doi: 10.1007/s004420050282
Rudinský, J. A., Novák, V., and Švihra, P. (1971). Attraction of the Bark Beetle Ips typographus L. to terpenes and a male-produced pheromone. Zeitschrift Angew. Entomol. 67, 179–188. doi: 10.1111/j.1439-0418.1971.tb02112.x
Schiebe, C.h., Blaženec, M., Jakuš, R., Unelius, C. R., and Schlyter, F. (2011). Semiochemical diversity diverts bark beetle attacks from Norway spruce edges. J. Appl. Entomol. 135, 726–737. doi: 10.1111/j.1439-0418.2011.01624.x
Schiebe, C.h., Unelius, C. R., Ganji, S., Binyameen, M., Birgersson, G., and Schlyter, F. (2019). Styrene, (+)-trans-(1R,4S,5S)-4-thujanol and oxygenated monoterpenes related to host stress elicit strong electrophysiological responses in the bark beetle Ips typographus. J. Chem. Ecol. 45, 474–489. doi: 10.1007/s10886-019-01070-8
Schlyter, F., Birgersson, G., Byers, J. A., and Bakke, A. (1992). The aggregation pheromone of Ips duplicatus and its role in competitive interactions with I. typographus (Coleoptera: Scolytidae). Chemoecology 3, 103–112. doi: 10.1007/BF01370137
Schlyter, F., Löfqvist, J., and Byers, J. (1987). Behavioural sequence in the attraction of the bark beetle Ips typographus to pheromone sources. Physiol. Entomol. 12, 185–196. doi: 10.1111/j.1365-3032.1987.tb00741.x
Schroeder, L. M., and Lindelöw, Å. (1989). Attraction of scolytids and associated beetles by different absolute amounts and proportions of α-pinene and ethanol. J. Chem. Ecol. 15, 807–817 doi: 10.1007/BF01015179
Schroeder, L. M. (1996). Interactions between the predators Thanasimus formicarius (Col.: Cleridae) and Rhizophagus depressus (Col: Rhizophagidae), and the bark beetle Tomicus Piniperda (Col.: Scolytidae). Entomophaga 41, 63–75. doi: 10.1007/BF02893294
Schroeder, L. M. (1999). Population levels and fight phenology of bark beetle predators in stands with and without previous infestations of the bark beetle Tomicus piniperda. For. Ecol. Manage. 123, 31–40. doi: 10.1016/S0378-1127(99)00014-6
Schroeder, L. M. (2003). Differences in responses to α-pinene and ethanol, and flight periods between the bark beetle predators Thanasimus femoralis and T. formicarius (Col.: Cleridae). For. Ecol. Manage. 177, 301–311. doi: 10.1016/S0378-1127(02)00441-3
Senf, C., and Seidl, R. (2018). Natural disturbances are spatially diverse but temporally synchronized across temperate forest landscapes in Europe. Glob. Chang. Biol. 24, 1201–1211. doi: 10.1111/gcb.13897
Sun, X.-L., Qing-Yin, Y., Sweeney, J. D., and Chang-Qi, G. (2006). A review: chemical ecology of Ips typographus (Coleoptera, Scolytidae). J. For. Res. 17, 65–70. doi: 10.1007/s11676-006-0016-2
Tolasz, R., Míková, T., Valeriánová, A., and Voženílek, V. (2007). Climate Atlas of Czechia, 1st Edn. Olomouc: Ceský Hydrometeorologický Ústav, Univerzita Palackého Praha, 255.
Tømmerås, B. Å. (1985). Specialization of the olfactory receptor cells in the bark beetle Ips typographus and its predator Thanasimus formicarius to bark beetle pheromones and host tree volatiles. J. Compar. Physiol. A 157, 335–341. doi: 10.1007/BF00618123
Turchin, P., Taylor, A. D., and Reeve, J. D. (1999). Dynamical role of predators in population cycles of a forest insect: an experimental test. Science 285, 1068–1071. doi: 10.1126/science.285.5430.1068
Unelius, C. R., Schiebe, C., Bohman, B., Andersson, M. N., and Schlyter, F. (2014). Non-host volatile blend optimization for forest protection against the European spruce bark beetle, Ips typographus. PLoS ONE 9:e85381. doi: 10.1371/journal.pone.0085381
Vité, J. P., Bakke, A., and Renwick, J. A. A. (1972). Pheromones in Ips (Coleoptera: Scolytidae): occurrence and production. Can. Entomol. 104, 1967–1975. doi: 10.4039/Ent1041967-12
Warzée, N., Gilbert, M., and Grégoire, J. C. (2006). Predator/prey ratios: a measure of bark-beetle population status influenced by stand composition in different French stands after the 1999 storms. Ann. For. Sci. 63, 301–308. doi: 10.1051/forest:2006009
Wegensteiner, R., Wermelinger, B., and Herrmann, M. (2015). “Natural enemies of bark beetles: predators, parasitoids, pathogens, and nematodes,” in Bark Beetles: Biology and Ecology of Native and Invasive Species, eds. F. E. Vega, and R. W. Hofstetter (Cambridge, MA: Academic Press), 247–304.
Wehnert, M., and Müller, M. (2012). ‘Allochthonous Kairomones' in stands of European beech (Fagus sylvatica) – Approach for nature-based bark beetle management with clerid beetles (Thanasimus spp.). Biol. Control 62, 16–23. doi: 10.1016/j.biocontrol.2012.03.003
Wermelinger, B. (2004). Ecology and management of the spruce bark beetle Ips typographus- A review of recent research. For. Ecol. Manage. 202, 67–82. doi: 10.1016/j.foreco.2004.07.018
Weslien, J. (1992). The arthropod complex associated with Ips typographus (L.) (Col., Scolytidae): species composition, phenology, and impact on bark beetle productivity. Entomol. Fenn. 3, 205–213. doi: 10.33338/ef.83730
Weslien, J. (1994). Interactions within and between species at different densities of the bark beetle Ips typographus and its predator Thanasimus formicarius. Entomol. Exp. Appl. 71, 133–143. doi: 10.1111/j.1570-7458.1994.tb01779.x
Weslien, J., Annila, E., Bakke, A., Bejer, B., Eidmann, H. H., Narvestad, K., et al. (1989). Estimating risks for spruce bark beetle (Ips typographus (L.)) damage using pheromone-baited traps and trees. Scand. J. For. Res. 4, 87–98. doi: 10.1080/02827588909382549
Weslien, J., and Regnander, J. (1992). The influence of natural enemies on brood production in Ips typographus (Col.: Scolytidae) with special reference to egg-laying and predation by Thanasimus formicarius (Col.: Cleridae). Entomophaga 37, 333–342. doi: 10.1007/BF02372435
Weslien, J., and Schroeder, L. M. (1999). Population levels of bark beetles and associated insects in managed and unmanaged spruce stands. For. Ecol. Manage. 115, 267–275. doi: 10.1016/S0378-1127(98)00405-8
Xie, S. A., and Lv, S. J. (2013). Effect of different semiochemicals blends on spruce bark beetle, Ips typographus (Coleoptera: Scolytidae). Entomol. Sci. 16, 179–190. doi: 10.1111/j.1479-8298.2012.00555.x
Zhang, Q. H., and Schlyter, F. (2003). Redundancy, synergism, and active inhibitory range of non-host volatiles in reducing pheromone attraction in European spruce bark beetle Ips typographus. Oikos 101, 299–310. doi: 10.1034/j.1600-0706.2003.111595.x
Zhang, Q. H., and Schlyter, F. (2010). Inhibition of predator attraction to kairomones by non-host plant volatiles for herbivores: a bypass-trophic signal. PLoS ONE 5:e11063. doi: 10.1371/journal.pone.0011063
Zhang, Q. H. E., Birgersson, G., Zhu, J., Löfstedt, C., Löfqvist, J., and Schlyter, F. (1999). Leaf volatiles from nonhost deciduous trees: Variation by tree species, season and temperature, and electrophysiological activity in Ips typographus. J. Chem. Ecol. 25, 1923–1943. doi: 10.1023/A:1020994119019
Zhao, T., Ganji, S., Schiebe, C., Bohman, B., Weinstein, P., Krokene, P., Borg-Karlson, A. K., and Unelius, C. R. (2019). Convergent evolution of semiochemicals across Kingdoms: bark beetles and their fungal symbionts. Isme J. 13, 1535–1545. doi: 10.1038/s41396-019-0370-7
Keywords: bark beetle, Picea abies, natural enemy, ipsdienol, ipsenol, drought, climate change, semiochemicals
Citation: Korolyova N, Bláha J, Hradecký J, Kašpar J, Dvořáková B and Jakuš R (2024) Mitigating Norway spruce mortality through the combined use of an anti-attractant for Ips typographus and an attractant for Thanasimus formicarius. Front. For. Glob. Change 7:1383672. doi: 10.3389/ffgc.2024.1383672
Received: 07 February 2024; Accepted: 22 April 2024;
Published: 16 May 2024.
Edited by:
Milica Zlatkovic, University of Novi Sad, SerbiaReviewed by:
Jovan Dobrosavljević, University of Belgrade, SerbiaCopyright © 2024 Korolyova, Bláha, Hradecký, Kašpar, Dvořáková and Jakuš. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nataliya Korolyova, a29yb2x5b3ZhLm5AY3plY2hnbG9iZS5jeg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.