 Shalini Oogathoo
Shalini Oogathoo Louis Duchesne
Louis Duchesne Daniel Houle4
Daniel Houle4- 1Centre d’Étude de la Forêt, Université du Québec à Montréal, Montréal, QC, Canada
- 2Data Science Laboratory, Université du Québec (TÉLUQ), Montréal, QC, Canada
- 3Direction de la recherche forestière, Ministère des Ressources naturelles et des Forêts du Québec, Quebec City, QC, Canada
- 4Science and Technology Branch, Environment Canada and Climate Change, Montreal, QC, Canada
Forest ecosystems have a major role in sequestering atmospheric CO2 and as such, their resilience is of upmost importance. In the boreal forest, trees grow only during a short period when air temperature is favourable. During winter, trees have specific mechanisms to survive in the cold air temperature. In order to understand the response of trees to a changing climate, this study assessed the influence of environmental variables on three phases of tree radial variation (i.e., growth, shrinkage and expansion) during three periods of the year (i.e., growing season, freeze–thaw period, and winter). The three phases were extracted from stem radial variation measured for as much as 11 years on 12 balsam fir [Abies balsamea (L.) Mill.] trees located in a cold and humid boreal forest of eastern Canada. The random forest algorithm was used to model each phase during each period. Our results show that tree growth increased with high precipitation and high relative humidity. Stem shrinkage was affected mostly by solar radiation, precipitation and vapour pressure deficit during the growing season and was likely caused by tree transpiration. During both the freeze–thaw and winter season periods, stem shrinkage increased with decreasing air temperature. During the growing season, stem expansion was related to 1-day-lag solar radiation and 1-day-lag vapour pressure deficit, which are the same variables associated with shrinkage the preceding day. Stem expansion increased with increasing air temperature and relative humidity during the freeze–thaw and winter season periods, respectively. This study shows that sink-driven tree growth is promoted mostly under humid conditions while antecedent dry and warm conditions are required during the growing season for trees to assimilate carbon through photosynthesis.
1 Introduction
Trees in different biomes are adversely affected by a warmer and drier climate, induced directly by climate change, thus leading to increased tree mortality across the globe (Allen et al., 2010). Trees mitigate climate change by acting as a carbon sink and thus controlling atmospheric CO2 concentrations (Bonan, 2008). To ensure the conservation of trees and forest land across the globe, it is vital to understand the mechanisms that influence tree growth as a means to better anticipate their response to climate change.
Throughout the year, tree stem diameter is subjected to important variations on a daily basis due to multiple processes including tree growth as well as non-growth-related stem shrinkage and stem expansion (Zweifel, 2016). While tree growth is restricted to a short and warm period in summer (mostly from June to August) in the boreal forest (Rossi et al., 2008), stems shrink and expand throughout the whole year. Both stem shrinkage and expansion are caused mainly by variation in water content and slightly by variation in temperature (Sevanto et al., 2005). The mechanisms causing both shrinkage and expansion due to variation in water content in the boreal forest differ based on the period of the year. Stem shrinkage/expansion are caused by: (1) dehydration/rehydration of trees during the growing season, (2) frost/thaw -induced bark shrinkage/expansion during freeze–thaw period, and (3) hygroscopic shrinkage/expansion throughout the year, respectively. Dehydration and rehydration during the growing season occur when transpiration exceeds the soil water uptake and vice-versa, respectively (Zweifel et al., 2001). Frost-induced bark shrinkage occurs when ice is formed in xylem and apoplastic spaces, leading to a decrease in water potential and thus movement of water from the inner bark towards the xylem (Zweifel and Häsler, 2000). As for the thaw-induced bark expansion, there is movement of water from the xylem towards the bark due to melting of the ice present in the xylem (Zweifel and Häsler, 2000). The hygroscopic shrinkage/expansion is caused by evaporation/absorption of water from/by the bark to/from the surrounding air during low/high atmospheric relative humidity (Zweifel et al., 2000; Ilek et al., 2017; Delapierre et al., 2023). The stem shrinkage and expansion caused by variation in temperature is minimal, and the thermal expansion is lower in dry wood than in fresh/wet wood (Sevanto et al., 2005; Zhao et al., 2016; Goli et al., 2019) and lower in the radial direction than in the tangential direction (Espinoza-Herrera et al., 2020).
Both dehydration and rehydration occur for the survival of trees under a cold climate (Zweifel and Häsler 2000). For example, trees undergo a period of pre-dormancy (hardening stage) during autumn when air temperature decreases below the freezing point, followed by a true dormancy period (cold-hardy stage) during winter, and a post-dormancy period (dehardening stage) during spring when air temperature increases (Havranek and Tranquillini, 1995). During the hardening stage, trees adopt either a deep supercooling or an extracellular freezing strategy for frost resistance (in order to avoid cellular damage of living cells) depending on species (George et al., 1982; Havranek and Tranquillini, 1995; Lintunen et al., 2015), where the extracellular freezing strategy leads to the dehydration of the living cells in the stem and subsequent stem shrinkage. However, trees may also undergo freezing stress that leads to: (1) thaw-expansion embolism (Sperry and Sullivan, 1992; Mayr and Sperry, 2010); or (2) frost-induced cellular damage (Lintunen et al., 2015), with the latter leading to an irreversible stem shrinkage in the freeze-stressed trees (Lintunen et al., 2016). During the dehardening stage, rehydration occurs which may be part of the repairing mechanisms by trees (Havranek and Tranquillini, 1995). The response of the three stages (hardening, cold-hardy and dehardening) to their changing environment under climate change may be greatly altered and eventually may have a negative impact on trees. For example, an early winter season will expose trees with insufficient pre-dormancy period to extreme freezing stress that can be damaging to trees, leading to irreversible shrinkage (Lintunen et al., 2016). Moreover, trees will not terminate their true dormancy period in a warmer winter due to their own endogenous rhythms, affecting tree recovery (Havranek and Tranquillini, 1995). Similarly, an earlier summer may disrupt the gradual restoration of physiological activity of trees that normally takes place during the post-dormancy period (Havranek and Tranquillini, 1995). Due to the high importance of tree growth during summer as well as both stem shrinkage and stem expansion in tree survival during winter (including their varying causal mechanisms based on the period of the year), there is a necessity to study all three phases (growth, shrinkage and expansion) for different periods of the year.
Studies evaluating the impact of environmental variables on tree growth showed that precipitation and relative humidity had the highest impact (Urrutia-Jalabert et al., 2015; He et al., 2016; Hu and Fan, 2016; Wang et al., 2016; Güney et al., 2020a). As for shrinkage, few studies have been done to evaluate the impact of environmental variables. Minimum air temperature was found to be strongly associated with maximum shrinkage during winter (Cocozza et al., 2009), while solar radiation, vapour pressure deficit and maximum air temperature had a higher impact on shrinkage during the growing season (Urrutia-Jalabert et al., 2015). To our knowledge, almost no work has been done explicitly on the expansion phase. Although Güney et al. (2020b) found vapour pressure deficit to increase tree recovery (rehydration) from tree water deficit, they did not analyse the expansion phase explicitly and for separate periods of the year.
In this study, we used the extracted tree growth, stem shrinkage and stem expansion phases based on the Zero-Growth (ZG) approach (Zweifel et al., 2016) to evaluate the impact of environmental variables on these phases. The three phases were extracted from stem radial variation measured in 12 balsam fir trees [Abies balsamea (L.) Mill.] for a period of as long as 11 years. Models were built using random forests for the three phases over four periods (the entire year, the growing season, the freeze–thaw period, and the winter season) with selected environmental variables. Our objectives for this study were to: (1) determine the relative importance of each environmental variables on each phase for each period; and (2) evaluate the relationship of key environmental variables with each phase. We first hypothesised that high precipitation and high relative humidity are the main drivers of tree growth during the growing season. Our second hypothesis was that VPD and PAR drive dehydration and the lagged VPD and lagged PAR drive rehydration during the growing season. Finally, we hypothesised that the control of stem shrinkage and stem expansion during the freeze–thaw period and winter season is mainly driven by minimum air temperature and daily minimum air temperature variation.
2 Methodology
2.1 Study area
The experimental site (an area of 3,600 m2) is in the boreal forest of the Laurentian Mountains in Quebec, Canada (47°19′31.36″ N, 71°07′43.15″ W, 740–760 m.a.s.l). It is an even-aged forest stand dominated by balsam fir, covering 93% of total basal area (i.e., 43 m2 ha−1 in 2019). It also has a small portion of paper birch (Betula papyrifera Marsh.) and white spruce [Picea glauca (Moench) Voss]. The forest stand had a stem density of 1,958 trees ha−1 (diameter at breast height ≥ 9 cm) and a quadratic mean diameter of 168 mm in 2019. The diameter at breast height (DBH) of the instrumented trees ranged from 14.0 to 22.8 cm in 2009. The site is characterised by sandy till soils overlaying a Precambrian charnockitic gneiss bedrock and has a slope of 15%. The average annual air temperature and precipitation at the site are −0.18°C and 1,460 mm for the period 1981–2010, respectively. The average minimum temperature and average maximum temperature at the experimental site were − 32.7 and 28.0°C, respectively, during the period 1999–2020.
2.2 Dendrometric data
With the recent advent of electronic (point/band) dendrometers, stem radial/diameter variations can be measured continuously with a high precision and at a very fine temporal resolution (daily, hourly, and sub-hourly), thus allowing to measure the individual phases (growth, shrinkage, and expansion). Daily stem diameter variations have been used mostly to extract seasonal parameters (beginning and end of growth, growth rate, etc.) by fitting the dendrometer data with different mathematical models, such as the Gompertz equation (Rossi et al., 2006; Duchesne et al., 2012; Miller et al., 2022). The finely resolved sub-hourly dendrometer data have been decomposed into different phases to study stem radius increment, shrinkage/contraction and recovery/expansion, known as the stem circadian-cycle approach (Downes et al., 1999; Deslauriers et al., 2003), or tree growth and water deficit, known as the zero-growth (ZG) approach (Zweifel et al., 2016). In the stem circadian approach, the stem radius increment phase is extracted based on the previous maximum stem radius observation obtained over a 24-h period, whereas the ZG approach partitions the stem diameter variation into growth period and tree water deficit period based on the assumption of no growth during stem shrinkage. In the stem circadian approach, the stem radius increment phase may contain repeated measures of growth during the growing season (calculated based on every 24-h period), whereas in the ZG approach, the growth phase has no repeated measures of growth as it is determined based on the maximum of previous stem radius records (which is not limited to the 24-h period). Many studies have evaluated the impact of climate/environmental variables on tree growth, but most of them adopted the stem circadian-cycle approach (Urrutia-Jalabert et al., 2015; He et al., 2016; Hu and Fan, 2016; Wang et al., 2016; Güney et al., 2020a) while a small number of studies used the ZG approach (Xue et al., 2022; Dulamsuren et al., 2023). Although the ZG approach has been increasingly used in the past few years to extract tree growth and water deficit phases (Dietrich et al., 2018; Barraclough et al., 2020; Pappas et al., 2020; Oogathoo et al., 2022, 2023; Xue et al., 2022; Dulamsuren et al., 2023), these studies did not evaluate the impact of environmental variables on each phase.
In this study, stem radial variation of the 12 selected balsam fir trees was measured at a temporal resolution of 15-min using point-dendrometers (model DR1, Ecomatik, Munich, Germany) in a soil warming experiment. The dendrometers were orientated to the south and positioned at 2 m above the ground (to ensure the instruments being above snowpack during winter) in 2009. Prior to installation of the point-dendrometer, the surface of the bark was scraped slightly to remove the dead bark. The soil warming experiment was initiated in the autumn 2008 (D’Orangeville et al., 2013a). It was carried out in three blocks laid out along the slope (for slope effect), with each block having two sub-blocks and each subblock having two plots of 12 m × 12 m each. In each plot, one healthy dominant/co-dominant balsam fir tree was selected. The soil warming treatment was allocated randomly in six of the 12 plots, whereby cables (heating and nonheating) were buried in a spiral pattern at 5–10 cm depth (at the interface of forest floor organic and mineral soil layers) around each selected tree. The cables were placed 0.9–2.5 m away from the base of the tree and at a spacing of 30 cm.
Due to the invasion and outbreak of the insect hemlock looper (Lambdina fiscellaria, Guenée) in the study area which reached its peak in 2012, five of the 12 instrumented trees were defoliated and died in 2013. The trees were also affected by a short drought in July 2012 (Houle et al., 2016). In addition, monitoring of three trees was discontinued due to malfunctioning of the dendrometers or the potentiometer required re-adjustment. In total, the stem radial variation data are available from 2009 to 2020 for the majority of the instrumented trees, encompassing precisely 92 complete years of dendrometer data. The monitored trees from this soil warming experiment were previously used for studying xylogenesis, soil ion fluxes, growth, phenology as well as seasonal and diel patterns of growth and water status dynamics (D’Orangeville et al., 2013a,b,c; Oogathoo et al., 2022, 2023).
2.3 Environmental data
Climate variables were measured continuously on an hourly basis from 1999 to 2020 at an open location, 200 m from the study area. Precipitation (P) was measured at a height of 2.3 m with a rain gauge located nearby (35–1,558, Fisher and Porter, Albany, NY, United States), while photosynthetically active radiation (PAR) was measured at a height of 8 m (LI190SB, Campbell Scientific, Logan, UT). Air temperature (T) and relative humidity (RH) were measured at a height of 3 m (HMP35CF, Campbell Scientific, Logan, UT), while wind speed (WS) was measured at the top (11 m high) of a straight tubular tower (013A, Campbell Scientific, Logan, UT). Vapour pressure deficit (VPD) was calculated using air temperature and relative humidity (Campbell and Norman, 1998). Sunshine hours (SH) was calculated using latitude and day of the year (Honsberg and Bowden, 2019). The daily cumulative degree days (DD) was calculated using the cumulative positive temperature difference (daily average air temperature and a base air temperature of 5°C) from the beginning to the end of the year for all study years.
Both soil temperature (401, YSI, Yellow Springs, OH) and soil water content (CS615, Campbell Scientific, Logan, UT) were measured daily since 1999 in four soil profiles at three soil depths corresponding to three distinct horizons (humus layer, 2 cm above the mineral soil surface; B horizon, 22 cm below the mineral soil surface; and C horizon C, 81 cm below the mineral soil surface). The daily mean soil temperature (ST) and daily mean soil water (SWC) for each soil horizon (Hum, HorB, HorC) were calculated from the average of the four soil profiles.
For both precipitation and sunshine hours, the daily sum was used, while for air temperature and vapour pressure deficit, the daily minimum and daily maximum were used, respectively. For the other variables (i.e., PAR, RH, WS, and ST.Hum), the daily mean values were used. In addition, for all these variables (P, T, VPD, SH, PAR, RH, WS, and ST.Hum), the daily variation (dif) and lag of up to 7 days (1 day lag—Lag1, 2 days lag—Lag2, 3 days lag—Lag3, 4 days lag—Lag4, 5 days lag—Lag5, 6 days lag—Lag6, and 7 days lag—Lag7) were calculated. For soil temperature (ST) in the B and C horizons, and soil water (SWC) in all three horizons, only daily differences were used.
2.4 Characterisation of growth, shrinkage and expansion from dendrometric data
Stem radial variation was segregated into three phases, namely irreversible tree growth, stem shrinkage and stem expansion (Oogathoo et al., 2023), using the ZG approach (Zweifel et al., 2016). The growth phase begins when the previous maximum stem radius is exceeded and ends when stem shrinkage begins, while the shrinkage phase occurs when stem radius decreases below a preceding maximum and the expansion phase is when stem radius increases above a preceding minimum but does not exceed a preceding maximum (Oogathoo et al., 2023). While the ZG approach is being used extensively to extract growth period, it may not be always accurate. For example, the beginning of the growing season may be much earlier than estimated if there is substantial cellular damage in trees due to frost that led to partial irreversible stem shrinkage during winter (Lintunen et al., 2015).
The stem radial variation is the difference in the stem radius between two time-intervals (15 min), where both growth and expansion have a positive stem radial variation, while shrinkage has a negative stem radial variation. For each phase, the stem radial variation was cumulated for each day. In the absence of any phase for certain days or periods (e.g., growth phase in winter), a value of zero was assigned.
2.5 Analyses of growth, shrinkage and expansion phases
The influence of environmental variables, individual trees and the soil warming treatment on the three phases was analysed for four specific periods, namely the entire year, the growing season, the freeze–thaw period (includes both spring and autumn) and the winter (excluding freeze–thaw events) season using Random Forests algorithm (Breiman, 2001). The purpose of analysing the three phases for three periods besides the entire year is to avoid masking important variables and to identify the most important ones influencing each phase for each period. The growing season was extracted when the growth phase begins and ends for each tree and for each year separately. The freezing period was days when minimum air temperature is below −5°C (Zweifel and Häsler, 2000) and stem shrinkage was high (< −0.2 μm). The thaw period was days when minimum air temperature was between −12 and + 10°C (Zweifel and Häsler, 2000) and stem expansion was high (> +0.2 μm). The extracted freeze and thaw periods were combined to obtain the freeze–thaw period (Figures 1A,C). The winter season was days when minimum air temperature was below −5°C and both stem shrinkage and stem expansion were low (< |0.2| μm; Figures 1B,D). The stem radial variation at 0.2 μm was chosen as a threshold to separate the winter period from freeze/thaw events as the trees exhibit most of the time a maximum of |0.2| μm stem radial variation during winter for all the years (Figure 1), which may be attributed to hygroscopic shrinkage/expansion of bark.
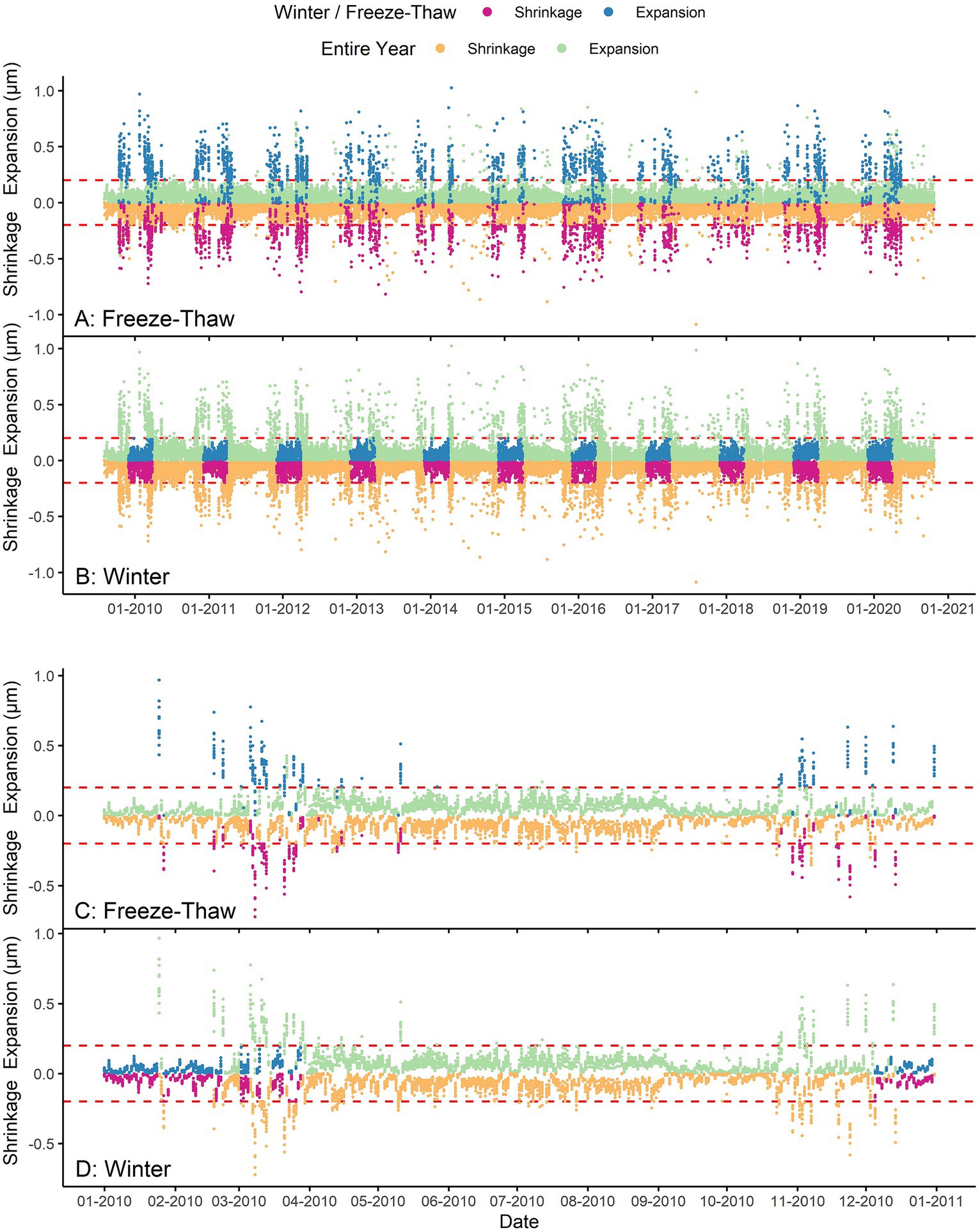
Figure 1. Illustration of the expansion (μm) and shrinkage (μm) phases during the freeze–thaw and winter periods for all the years (2009–2020; A,B) and for 2010 (C,D). The horizontal dashed red line is the expansion/shrinkage at ±0.2 μm used for the separation of the freeze–thaw and winter periods.
2.5.1 Running the random forests algorithm
Four databases (one for each period) consisting of the three phases together with all the trees, treatment (six control trees, six heated trees) and environmental variables (including the daily difference and the lags) described above were first set up. Since the cumulated daily value of the phases from all the trees were assembled in one database, each tree was incorporated as a dummy variable in the database to assess their individual impact on the phases and to highlight for any difference (if present) among the trees. Second, the Random Forests algorithm in the Boruta package in R Studio (Kursa and Rudnicki, 2010) was used to model each phase for each period in order to select all the predictor variables that are statistically important (p-value = 0.01), and only the Boruta selected variables were retained. Third, the Random Forest algorithm in the caret package in R Studio (Kuhn, 2008) was used to model (include fine-tuning) each phase by using only the Boruta selected variables, and to evaluate the precision and error associated with the model. The model was fine-tuned by changing the parameters (mtry and minimum node size) used in Random Forests in order to have a better fit. Finally, the best parameters obtained from the fine-tuning were used to run the final Random Forest model for each phase for each period.
2.5.2 Statistical analyses of the random forest models
Each final Random Forests model was evaluated using the following statistical tests: regression coefficient (R2), mean absolute error (MAE), and mean bias error (MBE). The relative importance of all the variables in each phase for the entire year and for the three periods were also determined using the caret package in R Studio. The ggRandomForests package in R studio (Ehrlinger, 2015) was used to develop partial dependence plots for the three periods (growing season, freeze–thaw period, and winter season) as a means to analyse the influence of the eight environmental variables (P, RH, VPD, PAR, SH, T, T.dif, and RH.dif) on each phase. These variables were selected because they were ranked the most important in the relative importance analysis for each Random Forest model.
3 Results
3.1 Growth, shrinkage, and expansion over the entire year and by period
The final Random Forest (RF) models for each phase for the entire year were robust, with high R2 (growth—0.71; shrinkage—0.81; expansion—0.83; Figure 2). For the growing season period, the robustness of the RF growth model was comparable to the entire year period (R2: 0.68), although there was less error and bias (lower MBE and MAE) for the entire year period. However, during the growing season period, the robustness of the RF shrinkage and expansion models was lower (R2: 0.59 and 0.54, respectively).
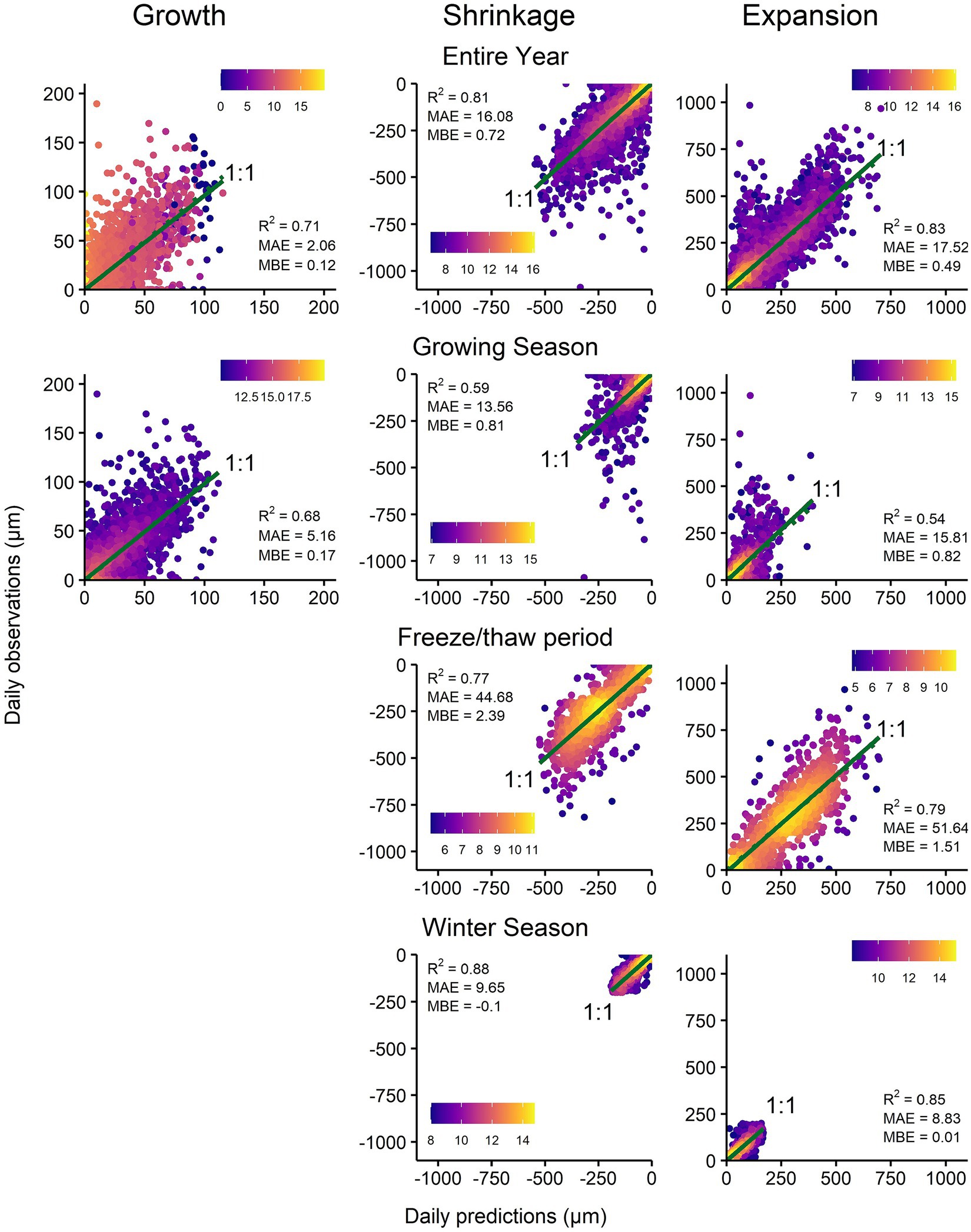
Figure 2. Observed vs. predicted growth, shrinkage and expansion during the entire year, growing season, freeze–thaw period, and winter season. MAE: Mean absolute error; MBE: Mean bias error.
The shrinkage and expansion RF models during the freeze–thaw period were more robust than during the growing season period (R2: 0.77 and 0.79, respectively), but the errors were highest in the former (MAE: 44.68 and 51.64, respectively; MBE: 2.39 and 1.51, respectively) compared to all other periods. The shrinkage and expansion RF models during the winter period had the highest R2 among all the periods (0.88 and 0.85, respectively) as well as the lowest errors (MAE: 9.65 and 8.83, respectively) and biases (MBE: −0.1 and 0.01, respectively). There was no growth during the freeze–thaw and winter periods.
3.2 Control of environmental variables on each phase for each period
The most important variables influencing each phase differs for each of the periods (Figure 3). There is also often one environmental variable that exerts much more control than any of the other variables and in turn, other variables exert a much smaller influence. For growth, P, RH, and PAR were the three most significant variables for both the entire year and the growing season. While all the SH related variables exerted a large influence (in the following order P, RH, and PAR) for the entire year period, DD was the fourth variable included in the model, followed by SH during the growing season period. However, for the entire year period, DD was not among the 15 most important variables influencing growth.
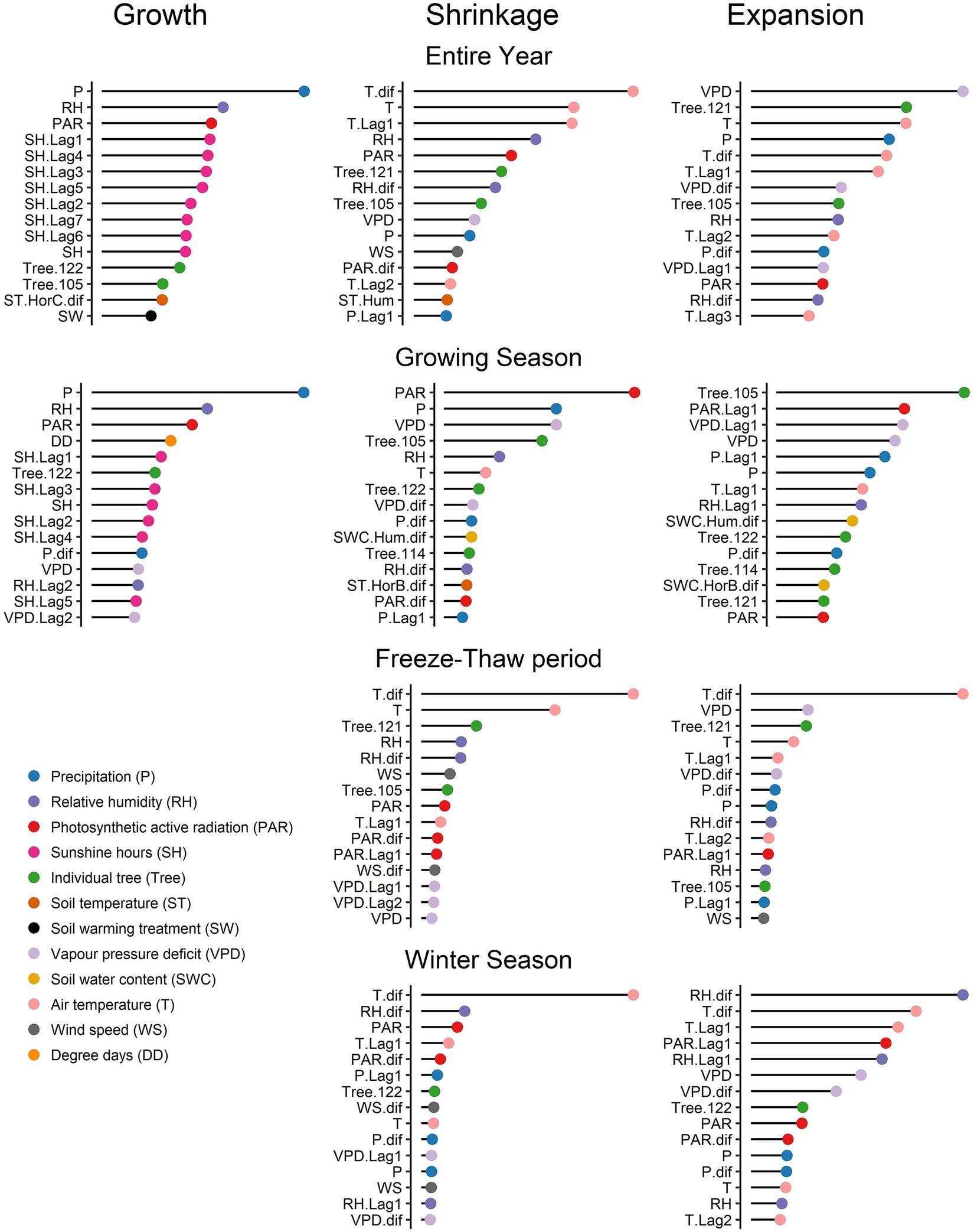
Figure 3. Relative influence of all variables for each phase (growth, shrinkage, and expansion) during the entire year, growing season, freeze–thaw period, and winter season.
For shrinkage, the daily air temperature variation (T.dif) was the most influential environmental variable for the entire year, freeze–thaw, and winter season periods, followed by air temperature (T) for the entire year and freeze–thaw periods, and daily relative humidity variation (RH.dif) for the winter season. During the growing season period, PAR followed by P and VPD were the environmental variables that influenced shrinkage the most.
For expansion, VPD and T.dif were the most influential environmental variables for the entire year and freeze–thaw period, respectively. During the winter season, RH.dif was the most significant variable, followed by T.dif, while VPD was ranked sixth. This shows that daily air temperature variation had a greater impact on expansion during the freeze–thaw period, while variation in daily relative humidity had a higher influence on expansion during the winter period. During the growing season period, the previous days PAR and VPD together with current day VPD had a higher impact on expansion.
3.2.1 Influence of environmental variables on each phase for each period
Majority of environmental variables had most important effects on the phases compared to their lags, but the lags of some variables also affected the phases. Additionally, the influence of most environmental variables on each phase (growth, shrinkage and expansion) depended on a threshold value.
During the growing season period, P was the most important compared to all lagged P for growth, and the latter increased with increasing P (up to 20 mm; Figure 4). Other variables (including their specific lags) had important effects on growth. Growth increased when RH and RH.Lag1 were > 75%, but it decreased with increasing RH.Lag2. Similarly, growth decreased with increasing VPD (up to 0.5 kPa), but it increased when VPD.Lag2 increased beyond 1.5 kPa. Growth decreased with increasing PAR (up to 150 μmol m−2 s−1), but it increased when both PAR.Lag2 and PAR.Lag3 increased beyond 200 μmol m−2 s−1. When SH and SH.Lag2 were greater than 15 h and when T and T.Lag1 were > 0°C, growth increased. As for shrinkage, all the six variables had the greatest impact compared to their respective lags during the growing season. Shrinkage increased with increasing P, decreasing RH, increasing PAR and when T was <0°C. Shrinkage decreased when VPD increased, reaching a minimum shrinkage value at 0.5 kPa, then increased again when VPD was >0.5 kPa. The effect of SH was negligible. As for expansion, it increased with increasing P and P.Lag1 and increasing PAR and PAR.Lag1. Expansion decreased slightly when RH.Lag1 was >75%, but it increased greatly when RH.Lag2 was <50%. While expansion increased with increasing VPD and VPD.Lag1, it was highest when VPD.Lag1 was almost zero. Expansion increased when T.Lag1 and T.Lag2 were < ~0°C, while the effect of SH was negligible.
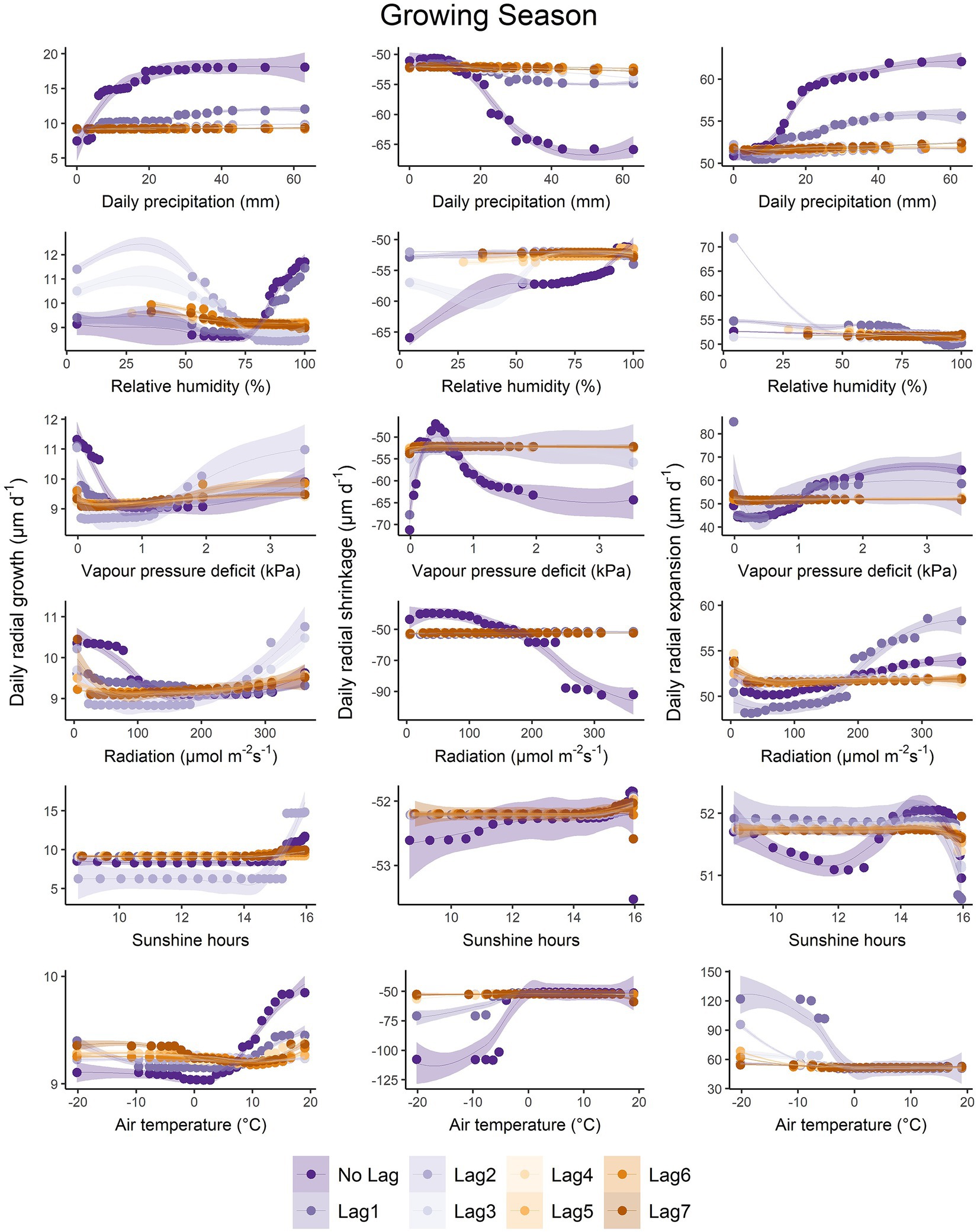
Figure 4. The partial dependence plot of daily precipitation (mm), relative humidity (%), vapour pressure deficit (kPa), photosynthetic active radiation (μmol m−2 s−1), sunshine hours (h), and air temperature (°C) for each phase during the growing season.
During the freeze–thaw period, RH and T had an important impact on shrinkage, where shrinkage decreased when RH was > ~80% and with increasing T above −10°C up to 0°C, reaching a minimum value at 0°C, then increased again when T was >0°C (Figure 5). As for the other variables, shrinkage increased with increasing P.Lag7, increasing lagged VPD (Lag2, Lag3 and Lag1) and increasing PAR and PAR.Lag1. Shrinkage also increased with decreasing RH variation and decreasing T variation. As for expansion, it increased with increasing P and when RH was > ~80%. Expansion also increased with increasing VPD up to 0.5 kPa, then remained almost constant. While expansion decreased with increasing PAR (up to 50 μmol m−2 s−1) then increased again (up to 200 μmol m−2 s−1) for PAR, it decreased when PAR.Lag1 and PAR.Lag2 were above 200 μmol m−2 s−1. High SH (> 12 h), and high T and T.Lag1 (> ~ −10°C) increased expansion. Expansion also increased with increasing T variation, while the effect of RH.variation was negligible.
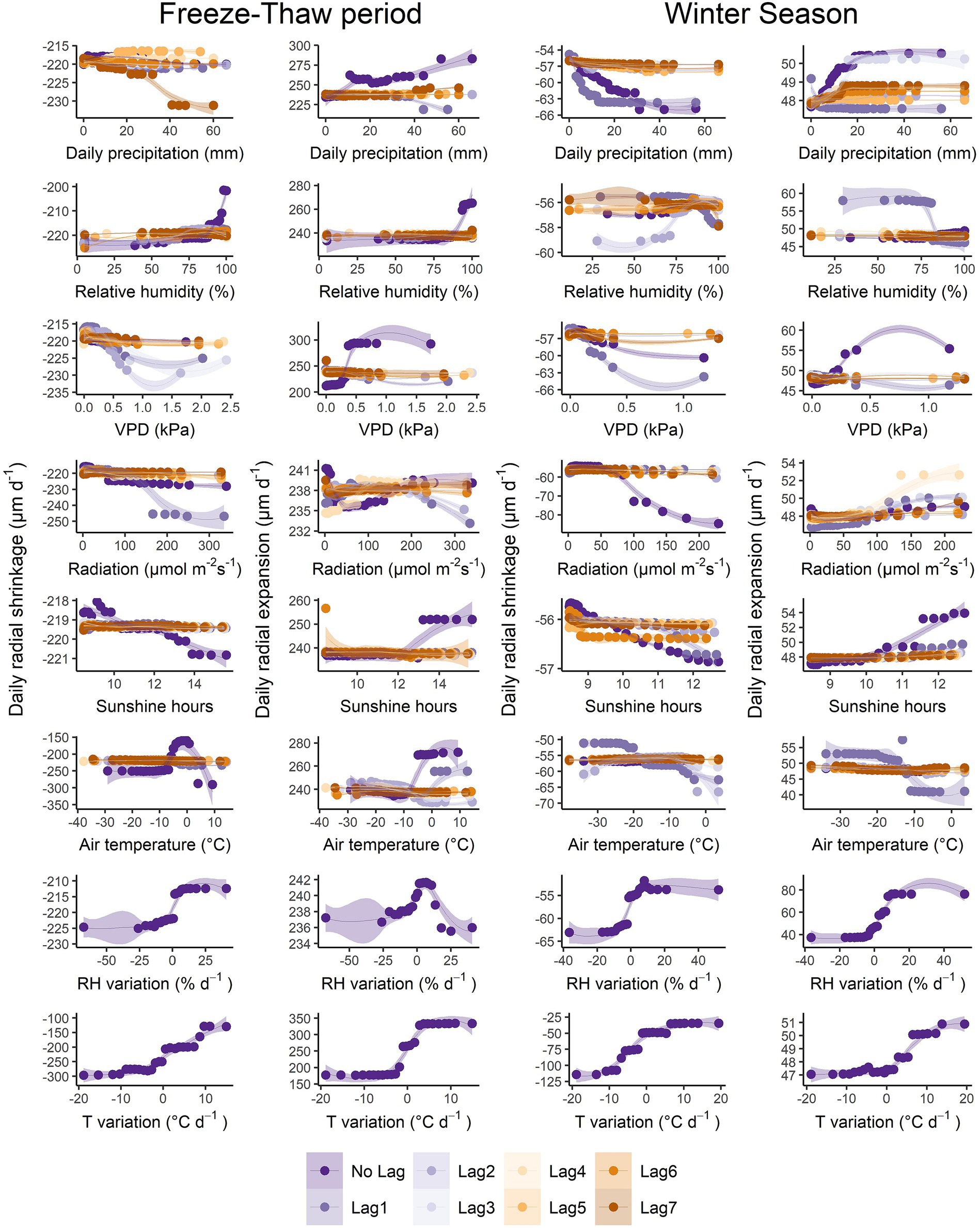
Figure 5. The partial dependence plot of daily precipitation (mm), relative humidity (RH, %), VPD (vapour pressure deficit, kPa), photosynthetic active radiation (μmol m−2 s−1), sunshine hours (h), air temperature (T, °C), RH variation (% day−1), and T variation (°C day−1) for each phase during the freeze–thaw period and winter season.
During winter, shrinkage increased with increasing P and P.Lag1, increasing VPD and more importantly with VPD.Lag1, increasing PAR, increasing SH, decreasing RH variation and more importantly with decreasing T variation (Figure 5). Shrinkage increased when RH.Lag1 was > ~80% and when RH.Lag2 was < ~80%, and with increasing T.Lag1, especially above −10°C. Expansion also increased with increasing P and P.Lag3, VPD, PAR (especially Lag4), SH, T variation and more importantly with RH variation. Expansion decreased when RH.Lag1 was > ~80% and when T.Lag1 was > ~ −20°C.
4 Discussion
4.1 Growth
Our results showed that P, RH and PAR were the main environmental variables that favour growth, either during the growing season or during the entire year, thus confirming our first hypothesis of high precipitation and high relative humidity being the main drivers of tree growth. Other studies using the stem circadian-cycle approach to determine the stem radial increment also found that P and RH are the main variables positively influencing growth (Urrutia-Jalabert et al., 2015; He et al., 2016; Hu and Fan, 2016; Wang et al., 2016; Güney et al., 2020a). However, using daily amplitude to investigate the effect of climate variables on tree growth, King et al. (2013) suggested that conventional methods possibly overstate the influence of precipitation and relative humidity on tree growth because the fluctuating stem diameter variations incorporate both radial growth and tree water dynamics. In our study, we extracted the growth phase from the stem radial variation, thus excluding tree water dynamics, and we still found P and RH to be the main environmental drivers of growth. Though antecedent favourable conditions (dry and sunny days) for photosynthesis are essential for trees to assimilate sufficient carbon for subsequent growth, it is nevertheless important to emphasise that tree growth is mostly sink-driven (tissue-growth) than source-driven (photosynthesis), where the tissue-growth is limited by water, temperature, and nutrients (Körner, 2015). This study showed that the sink-driven growth is favoured under humid conditions.
Growth was greater with P > 20 mm and RH > 75% as well as with PAR < 100 μmol m−2 s−1, implying that growth occurred mostly at night (Zweifel et al., 2021; Oogathoo et al., 2023), although growth can also be observed during the day at the favour of periods with a few consecutive rainy days. Although VPD was not the main environmental driver of growth, growth was greater when the evaporative demand was low (VPD < 0.5 kPa), which also corresponds to the high RH. Growth was also greater with greater day length (SH > 14 h), which is a finding that agrees well with Oogathoo et al. (2023).
4.2 Stem shrinkage and stem expansion
Although the RF model gave approximately the same results for the growth phase for entire year and the growing season, this was not the case for shrinkage and expansion phases for which the results differ between the various periods. The model for the entire year was greatly influenced by the large stem diameter variation that occurred during freeze–thaw and winter and thus masked the true variables influencing the phases during each period, particularly for the growing season for which stem shrinkage and expansion are linked to the tree water status. This shows the importance of partitioning the entire year into three periods to analyse the phases for each period separately, allowing to better understand the variables influencing the tree water status during the different phases and during the growing season in particular. Although the term shrinkage and expansion are used for all the periods, they are mostly associated with dehydration and rehydration during the growing season.
4.2.1 Stem shrinkage
During the growing season, PAR followed by P and VPD influenced stem shrinkage, thus confirming our second hypothesis for PAR and VPD while the effect of P was unexpected. Shrinkage increased greatly (~60 μm day−1) with increasing PAR, which indicates that dehydration occurs mostly when tree transpiration occurs during the day. Shrinkage decreased initially with increasing VPD, reaching a minimum at 0.5 kPa, then it increased moderately (~ 20 μm day−1) with further increase in VPD. The initial decrease in shrinkage might be due to the amount of soil water uptake being greater than the amount of water lost, while the eventual increase in shrinkage was due to higher amount of water being lost by the trees. Transpiration, which causes dehydration during the growing season (Zweifel et al., 2001), was also previously found to be highly correlated with PAR and VPD at the same site (Oogathoo et al., 2020). Besides losing water via transpiration at high VPD, trees may also have lost water via evaporation of the bark’s hygroscopic water under drier surrounding air (Oberhuber et al., 2020), thus leading to further shrinkage. While stem shrinkage was found to increase with increasing PAR and VPD during the growing season in a coastal temperate forest (Urrutia-Jalabert et al., 2015), increasing mean and maximum air temperature were also important variables. In our study, results are based on minimum air temperature, and this could be a reason why it was not among the main variables influencing shrinkage during the growing season. Although minimum air temperature is important for the other periods (i.e., freeze–thaw and winter), maximum air temperature might have a greater impact on dehydration during the growing season because it is closely associated with transpiration (following VPD and PAR) compared to minimum air temperature (Oogathoo et al., 2020). Unexpectedly, shrinkage also increased slightly (magnitude of ~15 μm day−1) with increasing P. The increased shrinkage occurred mostly when P was above 20 mm, which are high rainfall events that could have occurred early in the morning and thus allowed transpiration during the day. On the other hand, these high rainfall events could be associated to thunderstorms in summer that occur during a short duration (few hours), which can lead to high transpiration afterwards under clearer skies.
During the freeze–thaw period, both the T variation and T were the main variables controlling stem shrinkage, while only T variation had a major impact on stem shrinkage during winter, thus confirming only partially our third hypothesis. During the freeze–thaw period, Tree.121 was the third most important variable influencing shrinkage. This could be due to the large variations in stem diameter observed during the freeze–thaw period (in autumn and spring) for this particular tree (see Supplementary Figure S9 in Oogathoo et al., 2022). Stem shrinkage, which can be caused by frost-induced bark shrinkage (Zweifel and Häsler, 2000), increased greatly (~ 100 μm day−1) when air temperature decreased below an apparent threshold of −2 to 0°C, then remained more or less stable below ~ − 10°C in the freeze–thaw period (Figure 5). Stem shrinkage also increased greatly when air temperature increased above 0°C during the freeze–thaw period, but this increase might be due to transpiration or evaporation of the bark hygroscopic water (Figure 5). A decreasing daily air temperature variation (T.dif < 0°C day−1) led to an increase in shrinkage during both freeze–thaw and winter periods (Figure 5), which was also reported by Zweifel and Häsler (2000), where stem shrinkage occurs when air temperature falls below −5°C. Cocozza et al. (2009) also reported that stem shrinkage was linked to minimum air temperature, where trees shrink when air temperature decreases below a thermal threshold that varies with tree species. Other than T variation, RH variation was the next most important variable (but with much less significance) controlling stem shrinkage during winter, where shrinkage increased when RH decreased (<80%; Figure 5). This could be due to evaporation of hygroscopic water from the bark under the drier surrounding air (Lövdahl and Odin, 1992; Oberhuber et al., 2020).
4.2.2 Stem expansion
In our results, Tree.105, the preceding day PAR, VPD and P, together with the present day VPD and P, were the main variables influencing expansion during the growing season (Figure 3), confirming our second hypothesis of lagged PAR and lagged VPD influencing expansion. These observations are in good agreement with the concept that, during the growing season, rehydration is a response proportional to the extent of the preceding dehydration. For example, if a tree is dehydrated by a certain amount for 1 day or continuously for many days, it will rehydrate by the same amount to reach its initial capacity (which depends on the plant capacitance) when conditions are favourable for rehydration.
The high influence of Tree.105 on expansion might be due to its large stem diameter variation during the growing season. Expansion increased with increasing PAR.Lag1 as the latter triggered transpiration the preceding day and led to possible subsequent soil water uptake or redistribution of internal water stores (Figure 4). However, during transpiration, the amount of water transpired to the atmosphere might have been less than the amount of water uptake, leading to an overall expansion on the current day. Moreover, expansion was also very high when VPD during the precedent day was almost zero, which could be due to trees that did not lose much water via transpiration on the previous day and thus to more water being stored in the stem. In respect to the influence of increasing present day VPD (up to 1.5 kPa; Figure 4), water uptake from the soil is used to meet the evaporative demand and thus tree rehydration. Increasing P on the precedent and present days increased the soil water availability and the amount of water retained on the branches/leaves, and as such, trees might have expanded due to water uptake from the soil or via tree branches directly (Mayr et al., 2014). Although these variables affected stem expansion the most during the growing season in our study, this may differ based on tree species. Güney et al. (2020b), for example, reported that high VPD led to faster tree recovery (rehydration) in one conifer species (Cedrus libani A. Rich.), but P and high SWC were required for another conifer species (Juniperus excelsa M. Bieb.) in a Mediterranean forest, a much drier environment than our study site, which may also explain the differences.
During the freeze–thaw period, T variation was by far the most important driver of expansion, followed by VPD, and Tree.121 (Figure 3). An increase in T variation led to an increase in expansion (Figure 5), which is believed to be due to a thaw-induced bark expansion (Zweifel and Häsler, 2000). Stem expansion increased with increasing VPD up to 0.5 kPa (Figure 5), but the effect of VPD on expansion was small and comparable to the rest of the variables. This shows that the higher evaporative demand did not have a greater impact on expansion during the freeze–thaw period compared to the T variation. The influence of Tree.121 (3rd in rank of importance) could again be due to the large stem diameter variation that occurred during the freeze–thaw period as mentioned above. Contrary to our third hypothesis, RH variation had the most significant effect on stem expansion during the winter season, followed by T variation. Stem expansion was greater with increasing RH variation (RH.dif > 0% day−1) because high humidity from the air is absorbed by the bark (Oberhuber et al., 2020), and with increasing T variation (T.dif > ~2°C day−1; Figure 5) caused by a thaw-induced bark expansion (Zweifel and Häsler, 2000).
5 Conclusion
In this study, the modelling of each phase of tree radial variation for each period using Random Forests algorithm allowed us to identify the most important environmental variables affecting them. Our main finding is that tree growth mainly occurs when precipitation and relative humidity are elevated. Stem shrinkage in the growing season occurred during daytime (high PAR) with high precipitation and high/low evaporative demand (high/low VPD). Stem expansion was increased during high solar radiation (PAR.Lag1), either low or high vapour pressure deficit (VPD.Lag1 and VPD) and high precipitation (P.Lag1 and P) during the growing season. While Lag1 of PAR, VPD and P influenced expansion, it is the same variables that influenced shrinkage the day before. This result suggests that expansion is a direct response of prior shrinkage under favourable conditions, but this merits further investigations. During both freeze–thaw and winter season periods, shrinkage increased with decreasing daily air temperature variation. Expansion increased with increasing daily air temperature variation during the freeze–thaw period and with increasing daily relative humidity variation during the winter season.
While trees have to assimilate enough carbon during antecedent sunny days when conditions are favourable for photosynthesis, it is important to emphasise that the sink-driven tree growth occurs mostly under humid conditions at night. Our findings also showed that shrinkage and expansion during the growing period occur through different temporal patterns. For instance, it can be characterised by pronounced daily variation during many consecutive sunny days, but each phase can also follow a long increasing or decreasing trend that can last for a few consecutive days. During these contrasting periods, the influence of the controlling variables may differ, which deserves further consideration in future studies.
Data availability statement
The data analysed in this study are subject to the following licences/restrictions: the dataset will be made available upon request. Requests to access these datasets should be directed to c2hhbGluaS5vb2dhdGhvb0BnbWFpbC5jb20=; TG91aXMuRHVjaGVzbmVAbXJuZi5nb3V2LnFjLmNh.
Author contributions
SO: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Visualization, Writing – original draft. LD: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Validation, Visualization, Writing – review & editing. DH: Conceptualization, Investigation, Methodology, Validation, Writing – review & editing. DK: Conceptualization, Validation, Writing – review & editing. NB: Conceptualization, Funding acquisition, Project administration, Validation, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This project was made possible through Alliance funding (no. ALLRP 570411-21, NB as project leader) provided by the Natural Sciences and Engineering Research Council of Canada, which was coupled with a Contrat de service de recherche forestière obtained by NB from the Ministère des Ressources naturelles et des Forêts (MRNF) as part of research project no. 142959361 with LD as the government lead. Field work was also funded by the MRNF through project no. 142332064 to LD as well as within the framework of Action 3.6.1.1b of the implementation plan of Québec’s Plan pour une économie verte 2030, which aims to acquire knowledge about climate change adaptation.
Acknowledgments
We thank the field technicians at the Direction de la recherche forestière for experimental setup, maintaining equipment and collecting the data. We also thank the three reviewers for their constructive feedback on the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Allen, C. D., Macalady, A. K., Chenchouni, H., Bachelet, D., McDowell, N., Vennetier, M., et al. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 259, 660–684. doi: 10.1016/j.foreco.2009.09.001
Barraclough, A. D., Cusens, J., Zweifel, R., and Leuzinger, S. (2020). Environmental drivers of stem radius change and heterogeneity of stem radial water storage in the mangrove Avicennia marina (Forssk.) Vierh. Agric. For. Meteorol. 280:107764. doi: 10.1016/j.agrformet.2019.107764
Bonan, G. B. G. B. (2008). Forests and climate change: forcings, feedbacks, and the climate benefits of forests. Science 320, 1444–1449. doi: 10.1126/science.1155121
Breiman, L. (2001). An introduction to environmental biophysics. Springer Science & Business Media, New York.
Campbell, G. S., and Norman, J. M. (1998). An Introduction to Environmental Biophysics. Springer Science & Business Media.
Cocozza, C., Lasserre, B., Giovannelli, A., Castro, G., Fragnelli, G., and Tognetti, R. (2009). Low temperature induces different cold sensitivity in two poplar clones (Populus× canadensis Mönch ‘I-214’and P. deltoides marsh.‘Dvina’). J. Exp. Bot. 60, 3655–3664. doi: 10.1093/jxb/erp212
D’Orangeville, L., Côté, B., Houle, D., Morin, H., and Duchesne, L. (2013a). Increased soil temperature and atmospheric N deposition have no effect on the N status and growth of a mature balsam fir forest. Biogeosciences 10, 4627–4639. doi: 10.1007/s00468-013-0899-4
D’Orangeville, L., Côté, B., Houle, D., and Whalen, J. (2013b). Reduced mineralizable carbon in a boreal forest soil after three years of artificial warming. Can. J. Soil Sci. 93, 567–572. doi: 10.4141/cjss2013-046
D’Orangeville, L., Côté, B., Houle, D., and Whalen, J. (2013c). Reduced mineralizable carbon in a boreal forest soil after three years of artificial warming. Can. J. Soil Sci 93, 567–572.
Delapierre, F., Fonti, P., Lischke, H., and Moos, C. (2023). A method to quantify and account for the hygroscopic effect in stem diameter variations. Front. For. Glob. Change 6:1167542. doi: 10.3389/ffgc.2023.1167542
Deslauriers, A., Morin, H., Urbinati, C., and Carrer, M. (2003). Daily weather response of balsam fir (Abies balsamea (L.) mill.) stem radius increment from dendrometer analysis in the boreal forests of Québec (Canada). Trees 17, 477–484. doi: 10.1007/s00468-003-0260-4
Dietrich, L., Zweifel, R., and Kahmen, A. (2018). Daily stem diameter variations can predict the canopy water status of mature temperate trees. Tree Physiol. 38, 941–952. doi: 10.1093/treephys/tpy023
Downes, G., Beadle, C., and Worledge, D. (1999). Daily stem growth patterns in irrigated Eucalyptus globulus and E. nitens in relation to climate. Trees 14:102. doi: 10.1007/s004680050214
Duchesne, L., Houle, D., and D’Orangeville, L. (2012). Influence of climate on seasonal patterns of stem increment of balsam fir in a boreal forest of Québec, Canada. Agric. For. Meteorol. 162-163, 108–114. doi: 10.1016/j.agrformet.2012.04.016
Dulamsuren, C., Coners, H., Leuschner, C., and Hauck, M. (2023). Climatic control of high-resolution stem radius changes in a drought-limited southern boreal forest. Trees 37, 797–810. doi: 10.1007/s00468-022-02384-z
Ehrlinger, J. (2015). Ggrandomforests: visually exploring a random forest for regression. arXiv [Preprint]. doi: 10.48550/arXiv.1501.07196
Espinoza-Herrera, R., Olmos, L., Trujillo, I. A., and Garnica-Gonzalez, P. (2020). Study of thermal expansion and compression strength of three wood species from plantations. Cerne 26, 256–264. doi: 10.1590/01047760202026022723
George, M. F., Becwar, M. R., and Burke, M. J. (1982). Freezing avoidance by deep undercooling of tissue water in winter-hardy plants. Cryobiology 19, 628–639. doi: 10.1016/0011-2240(82)90192-4
Goli, G., Becherini, F., Di Tuccio, M. C., Bernardi, A., and Fioravanti, M. (2019). Thermal expansion of wood at different equilibrium moisture contents. J. Wood Sci. 65, 1–7. doi: 10.1186/s10086-019-1781-9
Güney, A., Gülsoy, S., Şentürk, Ö., Niessner, A., and Küppers, M. (2020a). Environmental control of daily stem radius increment in the montane conifer Cedrus libani. J. For. Res. 31, 1159–1171. doi: 10.1007/s11676-019-00983-0
Güney, A., Zweifel, R., Türkan, S., Zimmermann, R., Wachendorf, M., and Güney, C. O. (2020b). Drought responses and their effects on radial stem growth of two co-occurring conifer species in the Mediterranean mountain range. Ann. For. Sci. 77, 1–16. doi: 10.1007/s13595-020-01007-2
Havranek, W. M., and Tranquillini, W. (1995). “Physiological processes during winter dormancy and their ecological significance” in Ecophysiology of Coniferous Forests Eds. William K. Smith, Thomas M. Hinckley, (Academic Press), 95–124.
He, M., Yang, B., Wang, Z., Bräuning, A., Pourtahmasi, K., and Oladi, R. (2016). Climatic forcing of xylem formation in Qilian juniper on the northeastern Tibetan plateau. Trees 30, 923–933. doi: 10.1007/s00468-015-1333-x
Honsberg, C.B., and Bowden, S.G. (2019). Calculation of Solar Insolation. Photovoltaics Education Website. Available at: https://www.pveducation.org/pvcdrom/properties-of-sunlight/calculation-of-solar-insolation (Accessed November 1, 2023).
Houle, D., Lajoie, G., and Duchesne, L. (2016). Major losses of nutrients following a severe drought in a boreal forest. Nat. Plants 2, 16185–16187. doi: 10.1038/nplants.2016.187
Hu, L., and Fan, Z. (2016). Stem radial growth in response to microclimate in an Asian tropical dry karst forest. Acta Ecol. Sin. 36, 401–409. doi: 10.1016/j.chnaes.2016.09.005
Ilek, A., Kucza, J., and Morkisz, K. (2017). Hygroscopicity of the bark of selected forest tree species. iForest 10, 220–226. doi: 10.3832/ifor1979-009
King, G., Fonti, P., Nievergelt, D., Büntgen, U., and Frank, D. (2013). Climatic drivers of hourly to yearly tree radius variations along a 6 C natural warming gradient. Agric. For. Meteorol. 168, 36–46. doi: 10.1016/j.agrformet.2012.08.002
Körner, C. (2015). Paradigm shift in plant growth control. Curr. Opin. Plant Biol. 25, 107–114. doi: 10.1016/j.pbi.2015.05.003
Kuhn, M. (2008). Building predictive models in R using the caret package. J. Stat. Softw. 28, 1–26. doi: 10.18637/jss.v028.i05
Kursa, M. B., and Rudnicki, W. R. (2010). Feature selection with the Boruta package. J. Stat. Softw. 36, 1–13. doi: 10.18637/jss.v036.i11
Lintunen, A., Paljakka, T., Lindfors, L, and Hölttä, T (2016). “Tree water relations during winter” in 6th PEEX meeting (p. 309).
Lintunen, A., Paljakka, T., Riikonen, A., Lindén, L., Lindfors, L., Nikinmaa, E., et al. (2015). Irreversible diameter change of wood segments correlates with other methods for estimating frost tolerance of living cells in freeze-thaw experiment: a case study with seven urban tree species in Helsinki. Ann. For. Sci. 72, 1089–1098. doi: 10.1007/s13595-015-0516-3
Lövdahl, L., and Odin, H. (1992). Diurnal changes in the stem diameter of Norway spruce in relation to relative humidity and air temperature. Trees 6, 245–251. doi: 10.1007/BF00224344
Mayr, S., Schmid, P., Laur, J., Rosner, S., Charra-Vaskou, K., Dämon, B., et al. (2014). Uptake of water via branches helps timberline conifers refill embolized xylem in late winter. Plant Physiol. 164, 1731–1740. doi: 10.1104/pp.114.236646
Mayr, S., and Sperry, J. S. (2010). Freeze–thaw-induced embolism in Pinus contorta: centrifuge experiments validate the ‘thaw-expansion hypothesis’ but conflict with ultrasonic emission data. New Phytol. 185, 1016–1024. doi: 10.1111/j.1469-8137.2009.03133.x
Miller, T. W., Stangler, D. F., Larysch, E., Honer, H., Seifert, T., and Kahle, H.-P. (2022). A methodological framework to optimize models predicting critical dates of xylem phenology based on dendrometer data. Dendrochronologia 72:125940. doi: 10.1016/j.dendro.2022.125940
Oberhuber, W., Sehrt, M., and Kitz, F. (2020). Hygroscopic properties of thin dead outer bark layers strongly influence stem diameter variations on short and long time scales in scots pine (Pinus sylvestris L.). Agric. For. Meteorol. 290:108026. doi: 10.1016/j.agrformet.2020.108026
Oogathoo, S., Duchesne, L., Houle, D., and Kneeshaw, D. (2022). Characterizing seasonal radial growth dynamics of balsam fir in a cold environment using continuous Dendrometric data: a case study in a 12-year soil warming experiment. Sensors 22:5155. doi: 10.3390/s22145155
Oogathoo, S., Duchesne, L., Houle, D., Kneeshaw, D., and Bélanger, N. (2023). Seasonal, monthly, daily, and diel growth, and water status dynamics of balsam fir in a cold and humid boreal environment. Forests 14:802. doi: 10.3390/f14040802
Oogathoo, S., Houle, D., Duchesne, L., and Kneeshaw, D. (2020). Vapour pressure deficit and solar radiation are the major drivers of transpiration of balsam fir and black spruce tree species in humid boreal regions, even during a short-term drought. Agric. For. Meteorol. 291:108063. doi: 10.1016/j.agrformet.2020.108063
Pappas, C., Peters, R. L., and Fonti, P. (2020). Linking variability of tree water use and growth with species resilience to environmental changes. Ecography 43, 1386–1399. doi: 10.1111/ecog.04968
Rossi, S., Deslauriers, A., Anfodillo, T., Morin, H., Saracino, A., Motta, R., et al. (2006). Conifers in cold environments synchronize maximum growth rate of tree-ring formation with day length. New Phytol. 170, 301–310. doi: 10.1111/j.1469-8137.2006.01660.x
Rossi, S., Deslauriers, A., Griçar, J., Seo, J. W., Rathgeber, C. B., Anfodillo, T., et al. (2008). Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 17, 696–707. doi: 10.1111/j.1466-8238.2008.00417.x
Sevanto, S., Hölttä, T., Hirsikko, A., Vesala, T., and Nikinmaa, E. (2005). Determination of thermal expansion of green wood and the accuracy of tree stem diameter variation measurements. Boreal Environ. Res. 10:437.
Sperry, J. S., and Sullivan, J. E. (1992). Xylem embolism in response to freeze-thaw cycles and water stress in ring-porous, diffuse-porous, and conifer species. Plant Physiol. 100, 605–613. doi: 10.1104/pp.100.2.605
Urrutia-Jalabert, R., Rossi, S., Deslauriers, A., Malhi, Y., and Lara, A. (2015). Environmental correlates of stem radius change in the endangered Fitzroya cupressoides forests of southern Chile. Agric. For. Meteorol. 200, 209–221. doi: 10.1016/j.agrformet.2014.10.001
Wang, W., Zhang, F., Yuan, L., Wang, Q., Zheng, K., and Zhao, C. (2016). Environmental factors effect on stem radial variations of Picea crassifolia in Qilian Mountains, northwestern China. Forests 7:210. doi: 10.3390/f7100210
Xue, F., Jiang, Y., Dong, M., Wang, M., Ding, X., Yang, X., et al. (2022). Different drought responses of stem water relations and radial increments in Larix principis-rupprechtii and Picea meyeri in a montane mixed forest. Agric. For. Meteorol. 315:108817. doi: 10.1016/j.agrformet.2022.108817
Zhao, L., Jiang, J., and Lu, J. (2016). Effect of thermal expansion at low temperature on mechanical properties of birch wood. Cold Reg. Sci. Technol. 126, 61–65. doi: 10.1016/j.coldregions.2016.03.008
Zweifel, R. (2016). Radial stem variations–a source of tree physiological information not fully exploited yet. Plant Cell Environ. 39, 231–232. doi: 10.1111/pce.12613
Zweifel, R., Haeni, M., Buchmann, N., and Eugster, W. (2016). Are trees able to grow in periods of stem shrinkage? New Phytol. 211, 839–849. doi: 10.1111/nph.13995
Zweifel, R., and Häsler, R. (2000). Frost-induced reversible shrinkage of bark of mature subalpine conifers. Agric. For. Meteorol. 102, 213–222. doi: 10.1016/S0168-1923(00)00135-0
Zweifel, R., Item, H., and Häsler, R. (2000). Stem radius changes and their relation to stored water in stems of young Norway spruce trees. Trees 15, 50–57. doi: 10.1007/s004680000072
Zweifel, R., Item, H., and Häsler, R. (2001). Link between diurnal stem radius changes and tree water relations. Tree Physiol. 21, 869–877. doi: 10.1093/treephys/21.12-13.869
Keywords: point dendrometer, tree growth, stem shrinkage, stem expansion, boreal forest, environmental variables
Citation: Oogathoo S, Duchesne L, Houle D, Kneeshaw D and Bélanger N (2024) Precipitation and relative humidity favours tree growth while air temperature and relative humidity respectively drive winter stem shrinkage and expansion. Front. For. Glob. Change. 7:1368590. doi: 10.3389/ffgc.2024.1368590
Edited by:
Jason Vogel, University of Florida, United StatesReviewed by:
Jesús Julio Camarero, Spanish National Research Council (CSIC), SpainXianliang Zhang, Hebei Agricultural University, China
Copyright © 2024 Oogathoo, Duchesne, Houle, Kneeshaw and Bélanger. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shalini Oogathoo, c2hhbGluaS5vb2dhdGhvb0BnbWFpbC5jb20=