 Siyu Xie
Siyu Xie Tao Yan1
Tao Yan1
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. For. Glob. Change , 25 March 2024
Sec. Temperate and Boreal Forests
Volume 7 - 2024 | https://doi.org/10.3389/ffgc.2024.1343730
This article is part of the Research Topic Forest Ecosystems in Mountain Regions: Conditions, Risks and Impacts View all 13 articles
Understanding the relationship between tree growth and environmental conditions is essential to elucidating the impact of global climate change on forest ecosystems. We used the dendrochronology method to examine the growth sensitivity of a typical conifer to climate change in mountain forests of Central Hengduan Mountain. The study involved the establishment of tree ring width chronologies of Pinus yunnanensis in both Haba Snow Mountain (HB) and Yulong Snow Mountain (YL) in northwestern Yunnan, enabling the detection of the relationship between its radial growth and climates, i.e., monthly total precipitation, monthly temperatures (average minimum, mean and maximum) and monthly Palmer Drought Severity Index (PDSI). Response function and redundancy analysis (RDA) were used to identify correlations between climate variables and radial growth, and moving interval analysis was applied to determine the stability of climate-growth relationship. The findings demonstrated that the growth of P. yunnanensis had similar response patterns to climate change at two sites, exhibiting growth synchronization and common signals. Specifically, the radial growth of P. yunnanensis was negatively correlated with May temperature, while temperature in current October significantly promoted radial growth. Precipitation in June was the common climate variable with inverse effects between two sites, with positive impacts on YL and negative impacts on HB. The results of moving interval analysis were consistent with response function and RDA, presenting significant correlations in many years for those climatic variables significantly affecting tree growth. Stability analysis also revealed that the climate-growth relationship could fluctuate over a small range of time scales, induced by an abrupt change in climate. A forecast of strengthen in growth of P. yunnanensis forests was expected, since increases in precipitation and temperature of most months would benefit tree growth, and negative impacts of May temperature would be offset by the increase of precipitation in the corresponding month. These results could provide a basis for developing sustainable strategies of forest management under the climate change.
Global surface temperature will continue to increase until at least mid-century. Global warming will exceed 1.5°C to 2°C during the 21st century unless deep reduction measures of CO2 and other greenhouse gas emissions will be carried out in the coming decades (IPCC, 2021). In southwestern China, the climate has been warming and drying, with a tendency of intensification since the beginning of the 21st century (Su et al., 2014). Climate change has a massive impact on the structure and function of terrestrial ecosystems. For forest ecosystems, the largest component of the terrestrial ecosystem, climate change has greatly influenced its composition, structure, productivity, carbon storage capability, and ecological function (Zhang et al., 2017; Fan et al., 2019). Tree radial growth is sensitive to climate variation and mainly affected by climate factors, thus, a better understanding of climate effects on tree growth will help predict growth dynamics of forests under global climate change.
Climate warming is expected to enhance tree growth at alpine treelines (Camarero et al., 2021). Nevertheless, these positive influences need to be considered alongside water availability (Martínez-Vilalta et al., 2008), as high temperatures may lead to an increase in the frequency and intensity of droughts. If water availability is insufficient, the impact of drought stress on tree physiological processes will worsen, and consequently lead to a reduction of tree growth (Aber et al., 2001). Recent studies have revealed that droughts widely affected tree and forests growth over the world and even determined the survival of trees. For instance, a study found that consecutive hotter droughts posed a threat to forests under climate change in Central European floodplain forests, even with comparably high levels of water supply (Schnabel et al., 2022). Years with more intense drought corresponded with larger growth reductions and increased tree mortality rates in California forests, North America (Young et al., 2017; Bohner and Diez, 2021). Studies in Tibetan Plateau have shown that water deficit was the main factor controlling the resistance of juniper trees and higher drought frequency caused the decline of junipers’ growth, which was due to a lower tolerance to extreme drought events (Fang et al., 2021). In South America, predicted increases in drought frequency and intensity can have negative consequences for the functioning of the Amazon Forest (Van Passel et al., 2022).
Tree ring width is commonly used as a tree ring parameter and a measure of tree increment, which has contributed to the largest proportion of dendroclimatological research worldwide (He et al., 2019). Due to its precision as a proxy of recording climate information up to the millennium scale (Shen et al., 2020), it has been a useful tool in evaluating growth sensitivity to climate change and assessing the impact of future climate change on the radial growth of tree species and forests (Huang et al., 2010).
The Central Hengduan Mountains (CHM) is located in southeastern margin of Tibetan Plateau, which is a sensitive region to global climate change (Zhang et al., 2020). It is rich in biodiversity and covered by extensive forests composed of various species, covering their altitude distributional range and forming different treeline types. Therefore, the area is considered an excellent location to conduct a dendroclimatology study (Fan et al., 2009). Many efforts have been made to reconstruct historical climate and to find key climate factors affecting tree growth by using tree ring data of typical coniferous tree species in CHM, such as for Abies georgei (Fan et al., 2009; Liang et al., 2010; Panthi et al., 2018), Larix potaninii (Zhang et al., 2017¸ 2020), Picea brachytyla (Fan et al., 2008; Li et al., 2012; Yue et al., 2022), Picea likiangensis (Wang et al., 2018; Yu et al., 2018; Du et al., 2020), and Tsuga dumosa (Guo G. et al., 2009; Li et al., 2011; Aryal et al., 2020). These studies found that tree radial growth was influenced by both temperature and precipitation, but the growth response pattern was depended on species characteristics and site conditions. These studies provide an opportunity to understand climate-growth relationships for tree species and forests in high-altitude areas, and their connection to large-scale climate models (He et al., 2019).
Pinus yunnanensis forests are mainly distributed in Yunnan-Guizhou Plateau and in the southeastern edge of the Tibetan Plateau, and are important timber forests in southwestern China (Fu et al., 1999). P. yunnanensis is a shade-intolerant and deep-rooted coniferous species growing at altitudes between 1800 to 3,200 m a.s.l. in CHM (Deng et al., 2014; Shen et al., 2020). The species has strong adaptability to arid climate and barren soil, with red soil as the representative soil type. It grows best on well-drained north slopes with adequate light. The concurrent climate conditions in CHM are consistent with the climate background of global warming, which makes this typical species particularly interesting for studying the growth response to climate change. Tree ring studies of P. yunnanensis have been carried out at high (Yang et al., 2018, 2022), medium (Shen et al., 2020), and low altitudes in southwestern China (Sun et al., 2020). However, its growth responses to climate change in CHM is not well understood.
In this paper, we utilized dendrochronology methods to study the relationship between the radial growth of P. yunnanensis and climate by establishing tree ring width chronologies and detecting its growth response to climate factors on Haba Snow Mountain (HB) and Yulong Snow Mountain (YL) in CHM. We aim to (1) identify the critical climate factors affecting the growth of P. yunnanensis in the area, and (2) analyze the temporal stability of climate-growth relationships to determine whether the growth response to annual climate factors is smooth over time. It was hypothesized that temperature and precipitation interacted temporally, and negative impacts of temperature on growth were expected in dry periods. Furthermore, we explored the impact of future climate change on P. yunnanensis forests growth based on climate-growth relationships results and climate models.
HB with the peak of 5,396 m a.s.l. and YL with the peak of 5,596 m a.s.l. are two typical snow-capped mountains in the Central Hengduan Mountain (CHM), separated by the Jinsha River (Figure 1). Different vegetation types are formed along the altitudinal gradient in CHM. The dry valleys are usually covered by scrublands. Warm-temperate coniferous forests, dominated by P. yunnanensis and Pinus armandii, are mainly distributed at altitudes between 1,000 m a.s.l. and 3,300 m a.s.l. Pinus densata is a dominant species at altitudes of 2,600 to 3,500 m a.s.l. on the southern slope. Cold-temperate coniferous forests are dominated by spruce (P. brachytyla, P. likiangensis), fir (A. georgei, Abies forrestii) and larch species (L. potaninii), at elevations ranging from 3,000 m a.s.l. to 4,200 m a.s.l. (Fan et al., 2009).
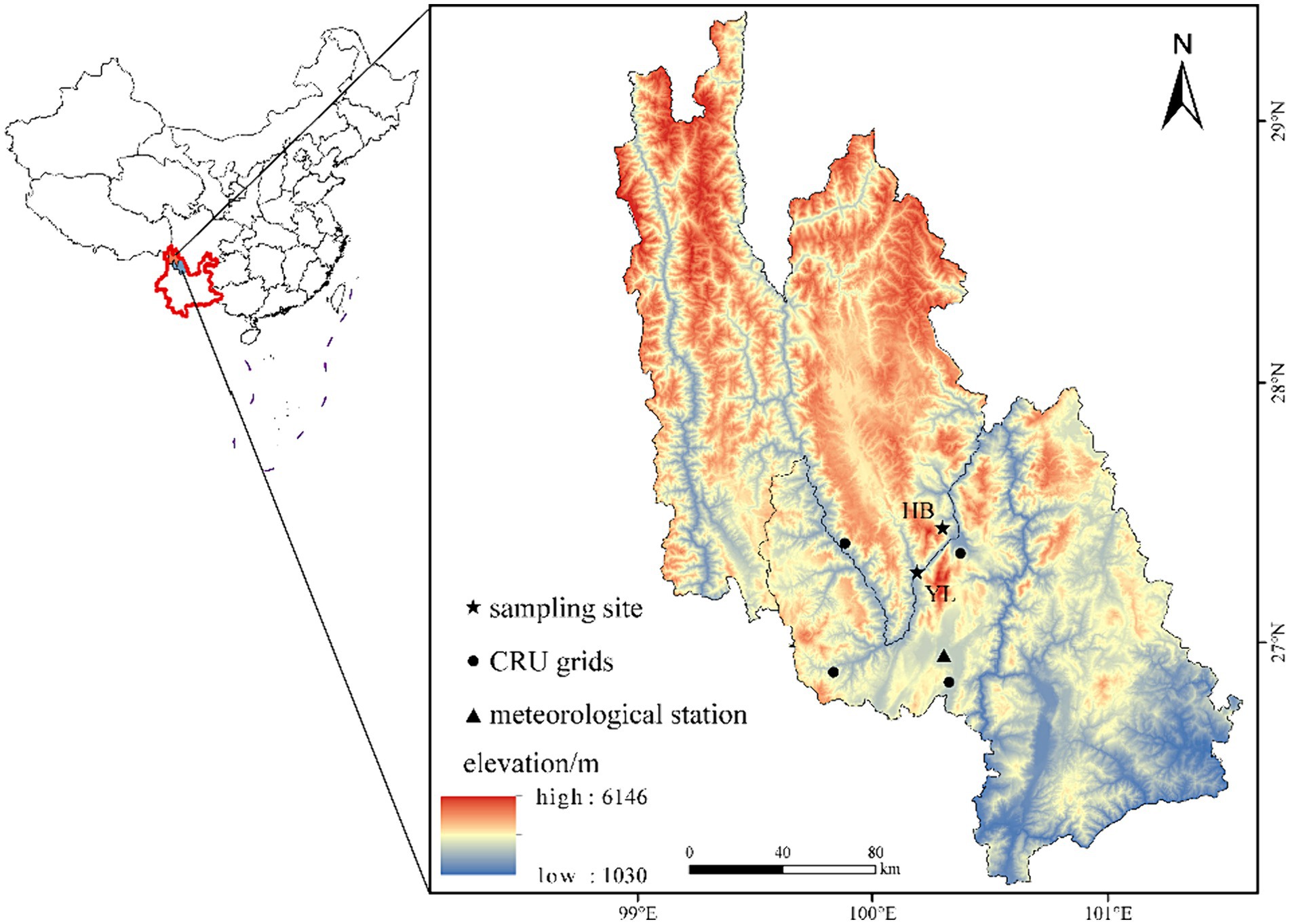
Figure 1. The location of sampling sites and meteorological station. HB: Haba Snow Mountain; YL: Yulong Snow Mountain; the same below.
Although vegetation types along the altitudinal gradient were similar in both HB and YL, the distribution range of P. yunnanensis was a bit different. The species is mainly distributed from 2000 m a.s.l. to 3,500 m a.s.l. in HB, while the distribution of P. yunnanensis in YL ranged from 2000 m a.s.l. to 3,000 m a.s.l. In HB, the soil texture is predominantly loamy, distributed from 1800 to 3,450 m a.s.l., transitioning from red soils to yellow and then brown soils. Above 3,900 m a.s.l., podzolic soil and meadow soil prevail. In YL, the soil texture is dominated by loam and clay, with a gradual transition from clay and loamy clay to sandy clay, sandy loam, and loam as the altitude increases from low to high.
The climate in CHM is strongly influenced by the southeast monsoon and southwest monsoon, with the monsoon climate characteristics of simultaneous rain and heat, and distinct dry and wet seasons. According to the meteorological data (1951–2021) of Lijiang meteorological station (26°51′N, 100°13′E, 2380.9 m a.s.l., Figure 2), annual mean temperature was 12.97°C, while annual maximum temperature (23.92°C) and annual minimum temperature (−0.11°C) appeared in June and January, respectively. The annual total precipitation was 988.93 mm, mainly occurring in summer, as the precipitation from June to September accounted for 75% of the total precipitation. Over the last three decades, the climate of the study area has been characterized by noticeable warming and drying, with a significant increasing trend in temperature (particularly during the period from 1990 to 2016) and a decreasing trend in Palmer Drought Severity Index (PDSI) between 1951 and 2021, while precipitation did not show a significant decreasing trend (Figure 3).
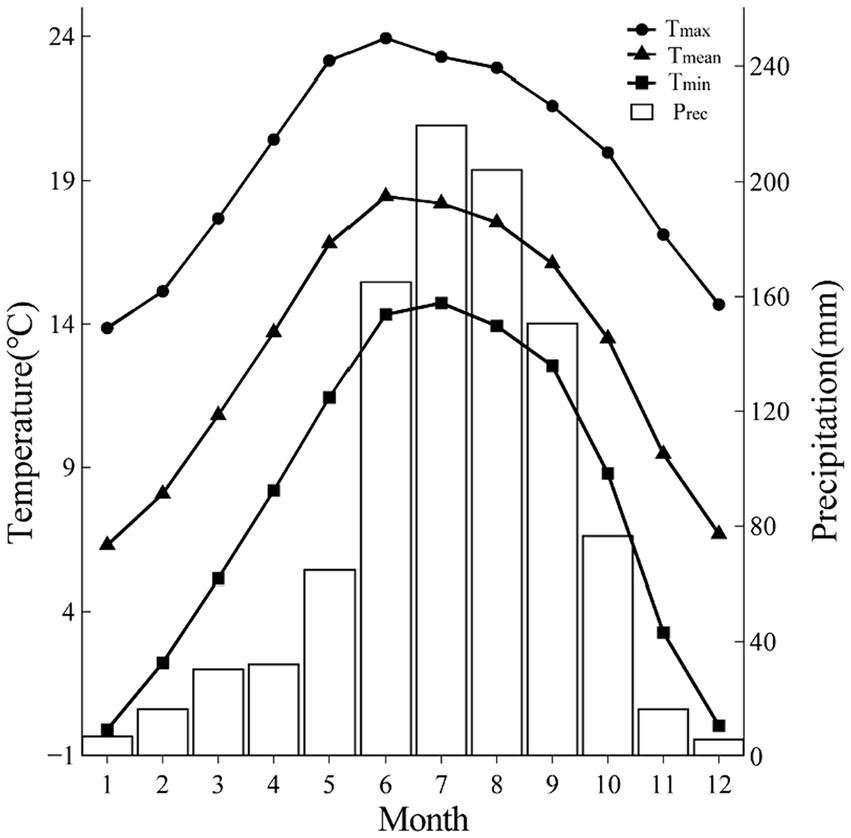
Figure 2. The monthly climatic characteristics at Lijiang Meteorological Station (1951–2021). Prec, monthly total precipitation; Tmin, monthly mean minimum temperature; Tmean, monthly mean temperature; Tmax, monthly mean maximum temperature.
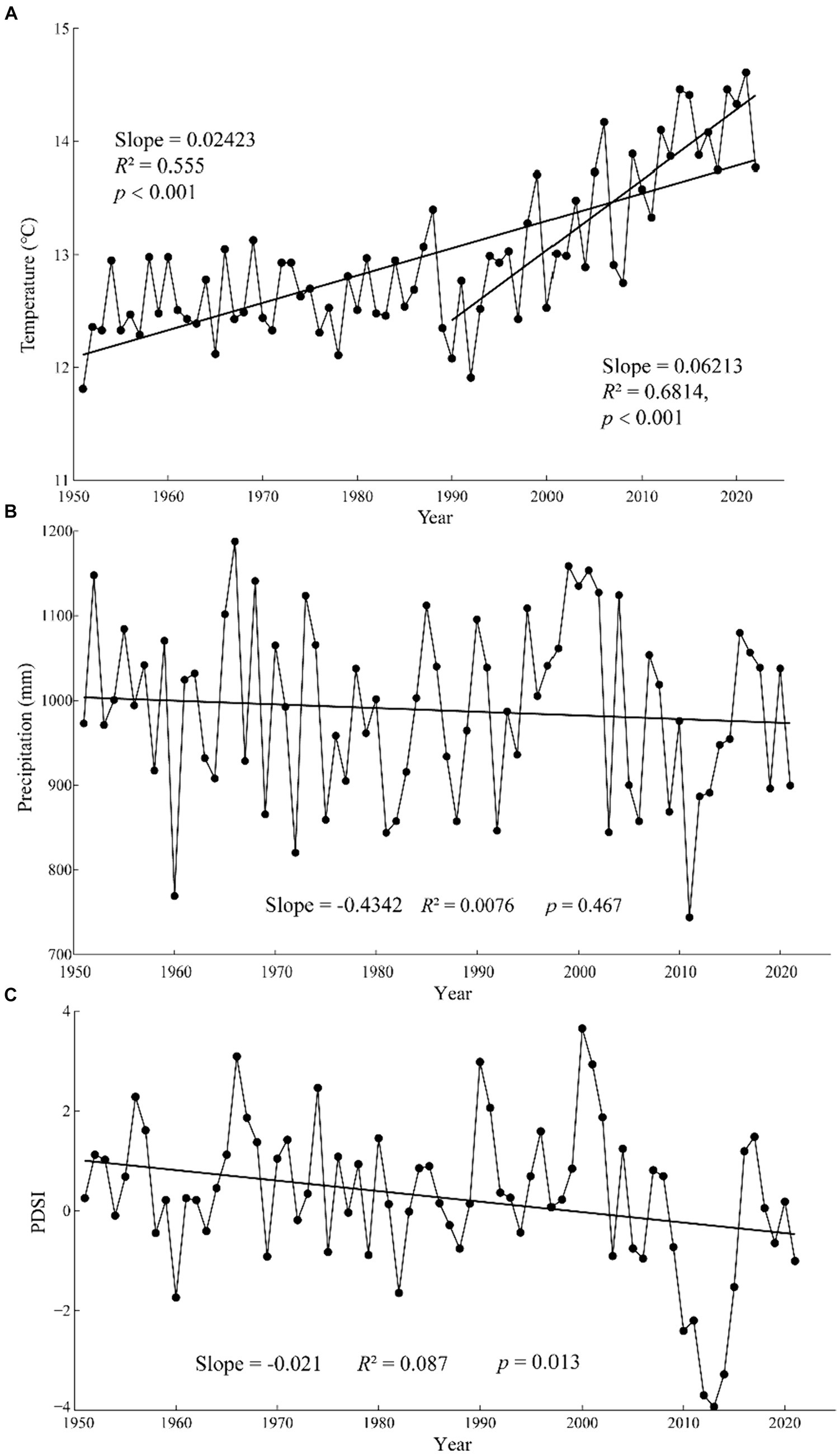
Figure 3. Annual trends of mean temperature (A), precipitation (B) and Palmer Drought Severity Index (PDSI) (C) at Lijiang Meteorological Station (1951–2021).
We sampled mature and healthy trees without insect damage at undisturbed forest communities at two adjacent sites with similar climate conditions and growth environments (Table 1). The cores were drilled at the trees’ breast height (1.3 m) with a 5.15-mm-diameter increment borer, one or two cores were extracted per tree from the opposite direction parallel to the contour to avoid growth reaction to the slope. Obtained cores were labelled and placed in plastic straws. In the laboratory, the cores were put into wooden tanks by using white latex and tape. The samples were polished by sandpaper with gradually finer particle sizes (200 mesh to 1,000 mesh) until the growth rings were visible. Then, the cores were visual cross-dating under a microscope and scanned by EPSON (Expression11000XL) scanner. The scanner parameters were set to 24-bit full-color professional mode image types, with a resolution of 2000 dpi. The dated cores were measured using the CDendro and CooRecorder ver. 9.8.1 software at a precision of 0.001 mm to obtain the ring widths. Furthermore, the results of cross-dating were examined by the COFECHA program (Holmes, 1983), and those cores with inadequate quality were removed based on the testing results. Ultimately, 62 trees from 112 cores remained for the further analysis.
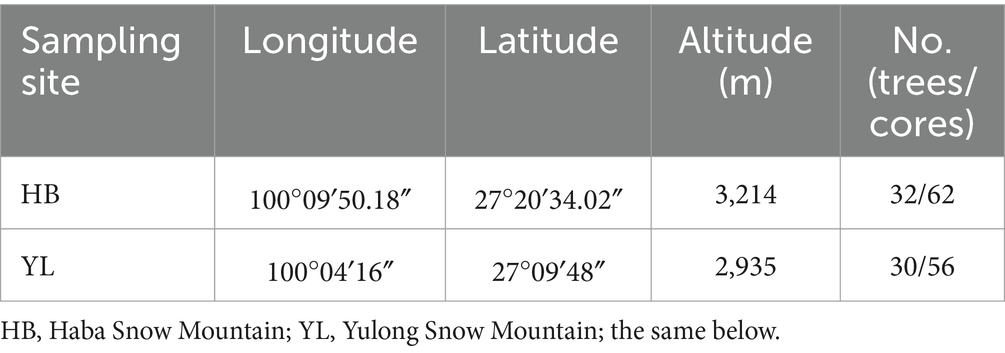
Table 1. Sampling sites information.
To remove the growth trend caused by genetic factors or individual interference while preserving more climate signals, a negative-exponential curve function was applied to standardize the ring width series. Residual chronologies (Figure 4) were developed after autoregressive modelling performed on per standardized series to remove potential autocorrelation and retain high-frequency climate change. Further, all residual series were averaged on a site-by-site basis utilizing the bi-weight robust mean (Cook, 1985), to minimize the influence of outliers. All procedures were executed using the ARSTAN Program (Cook, 1985) and two residual chronologies were generated with chronological eigenvalue (Table 2), including mean sensitivity (MS), signal-to-noise ratio (SNR), standard deviation (SD), and expressed population signal (EPS), etc.
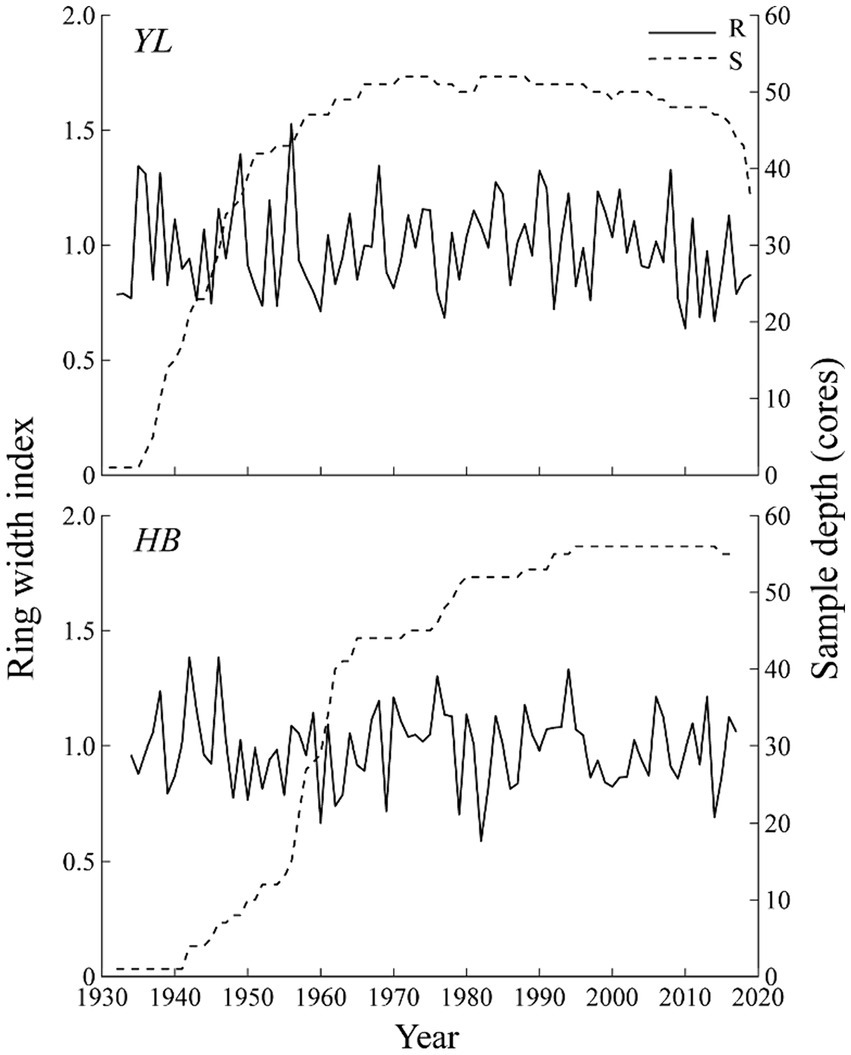
Figure 4. Residual chronologies of tree ring width for two sampling sites. R, ring width index; S, sample depth.
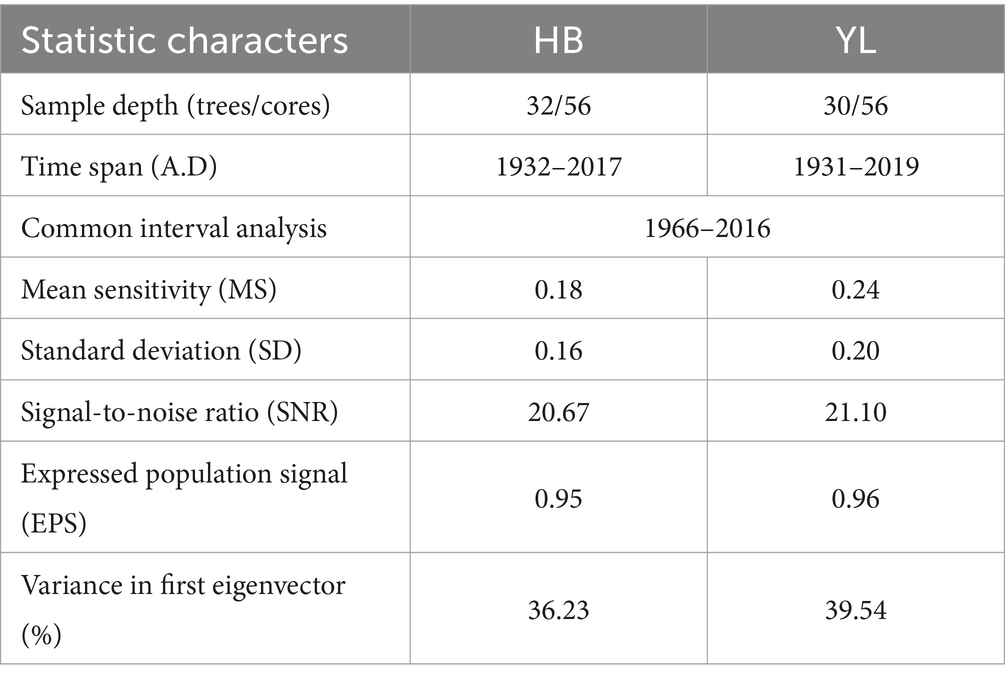
Table 2. Statistics of tree ring width residual chronologies.
The MS quantifies the degree of inter-annual variation in ring width, serving as an indicator of the sensitivity of tree growth to climate change. A higher value indicates a greater inter-annual variation in ring width, suggesting that the trees are highly responsive to environmental changes. The SNR serves as an indicator of the proportion of climatic signals to non-climatic noise within the ring width data. A high SNR signifies that the ring-width data contains a richer climatic information. The SD is a statistical measure that assesses the dispersion of tree ring width data, reflecting the variation in ring width across different growth years. The EPS is a statistical metric that assesses the representativeness of a sample. A high value of EPS means that the sample data are well represented and can be used to infer growth and climate change in broader regions.
Monthly mean maximum temperature (Tmax), monthly mean temperature (Tmean), and monthly mean minimum temperature (Tmin), sourced from the neareast Lijiang Meteorological Station (26°50′N, 100°13′E, 2,382 m a.s.l), were acquired through the National Oceanic and Atmospheric Administration (NOAA).1 Precipitation data were obtained from the Climatic Research Unit (CRU TS v. 4.07; https://crudata.uea.ac.uk/cru/data/hrg) at a resolution of 0.5° × 0.5° (Harris et al., 2020). The PDSI is a comprehensive meteorological drought index that considers precipitation, temperature, evapotranspiration, and drought duration (Alley, 1985). The PDSI data was extracted from the monthly data set compiled by the global CRU grids2 at 0.5° × 0.5° spatial resolution (Dunn et al., 2022). Its index ranges from −4 (indicating extremely dry conditions) to +4 (indicating extremely wet conditions), where a smaller value represents a more severe relative drought.
To capture the lag effect of climate on tree radial growth (Fritts et al., 1965), each tree ring was paired with 14 months of climate data spanning from the previous year’s September through the following October. The climate timeseries interval was from 1987 to 2017 for the data analysis. Five climate variables were selected for analyzing relation to tree radial growth, including Tmean, Tmax, Tmin, the monthly total precipitation, and PDSI.
The response function was applied to analyze correlations between climate variables and chronologies by using DendroClim2002 (Biondi and Waikul, 2004). The coefficients of the response function are multivariate estimates from a principal component regression model (Morzuch and Ruark, 1991) and the method solves the inter-correlation problems among climatic variables (Fritts et al., 1971). To further identify common climate variables affecting the radial growth of P. yunnanensis, the redundancy analysis (RDA) was also conducted using CANOCO 5.0 software. RDA is the canonical form of principal component analysis (PCA) and constitutes the direct extension of multiple regressions applied to multivariate data (Legendre and Legendre, 1998). In RDA, the ordination axes are constrained to be linear combinations of supplied environmental variables (ter Braak, 1994), and it is an effective method in quantifying the relationship between tree-ring width and climate variables (Girardin et al., 2004; Drobyshev et al., 2013; Zhang et al., 2020). Like the response function, Tmean, Tmax, Tmin, the monthly total precipitation, and PDSI were selected for RDA. Following a Monte Carlo permutation test involving 999 random permutations, only those climate variables showing significant correlations (p < 0.05) with the two residual chronologies were presented in the RDA.
For tracing the dynamic relationship over time between the annual ring width index and climate, the Moving Forward method (with a window of 36 years) in the Evolutionary and Moving Response and Correlation module of DendroClim 2002 software was also applied to calculate temporal stability of relationships between chronologies and climate variables. The Special Report on Emissions Scenarios (SRES) B2 scenario was considered as future changes in temperature and precipitation, it was published by the Intergovernmental Panel on Climate Change to provide an assessment of the climate change. The graphs were plotted using the ggplot2 program package in R.
Both residual chronologies with high values of SNR and variance in first eigenvector contained large climate information (Table 2). Tree growing in YL with greater MS and SD showed higher interannual fluctuations than those in HB (Table 2). The high EPS (>0.85) confirmed that the sampled population for each site was well-represented, and two residual chronologies could reflect the overall growth characteristics of P. yunnanensis in the study area and then the chronologies could be applied to conduct correlation analysis with climate variables.
We showed correlations between radial growth and climate variables in Figure 5. In HB, the radial growth of P. yunnanensis was significantly associated with Tmax in May and precipitation in February, by showing negative and positive correlations, respectively. In addition, PDSI in the previous September negatively affected P. yunnanensis growth. In YL, the radial growth was significantly and negatively correlated with May and June temperatures (both Tmean and Tmax), and significantly and positively correlated with Tmean and Tmin of the current October. While P. yunnanensis radial growth was found to be positively correlated with precipitation in May and June, and previous October. For PDSI, significant and positive correlations were found in June and July.
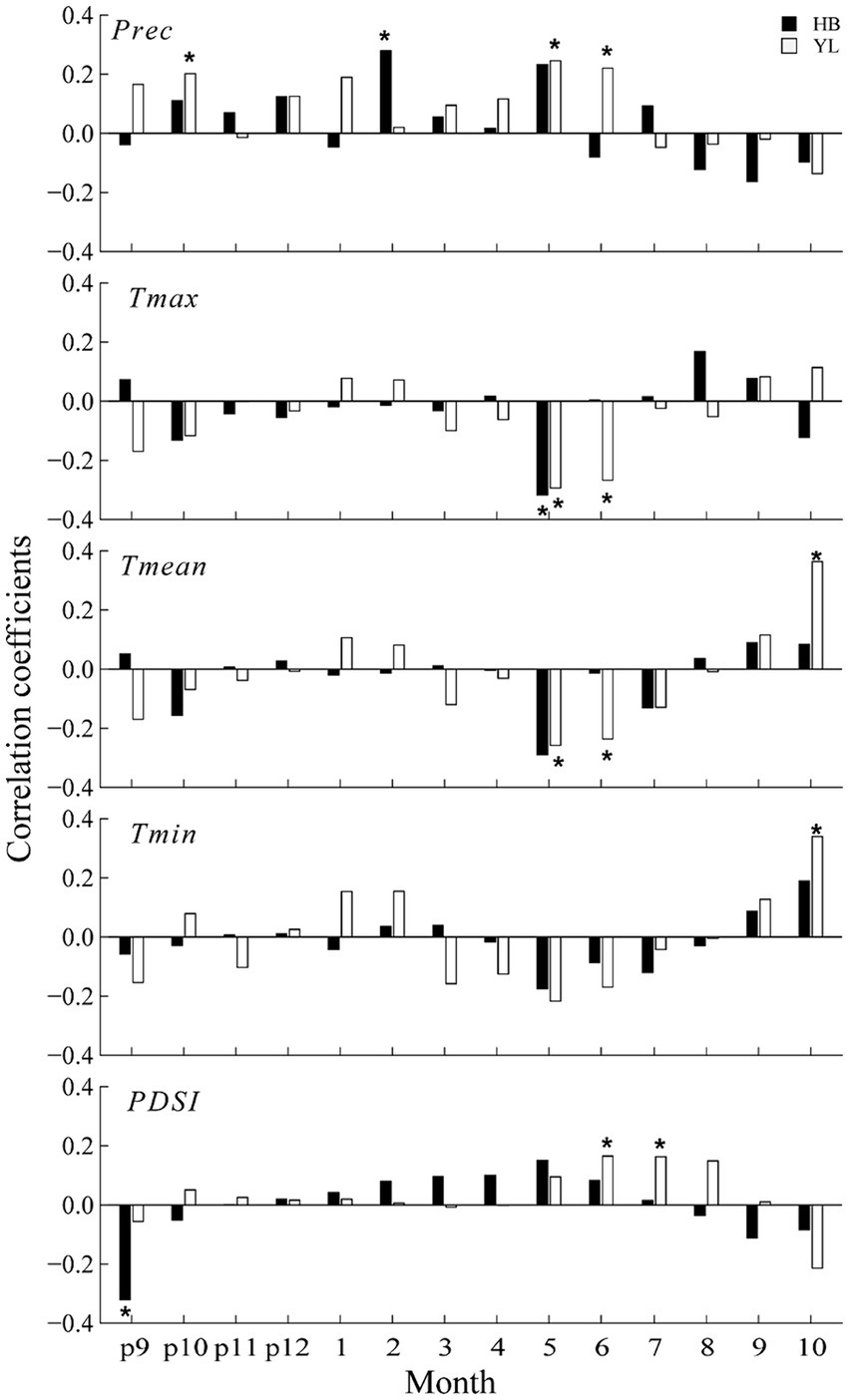
Figure 5. Correlation coefficients between residual chronologies and climate variables during the periods between 1966 and 2016. * indicates a significant correlation at p < 0.05. p: previous year, the same below.
The results of RDA (Figure 6) revealed that three climate variables significantly affected the radial growth of P. yunnanensis. Specifically, May Tmean, current October Tmean, and June precipitation explained 17.9, 6.7, and 4.7% of the radial growth, respectively. The first axis and the second axis accounted for 24.49 and 4.85% of the response variables, respectively. May Tmean exerted the most considerable influence on radial growth with negative impacts at both sites, while current October Tmean positively affected the radial growth. June precipitation presented inverse impacts on tree growth at two sites, by showing negative and positive influences in HB and YL, respectively.
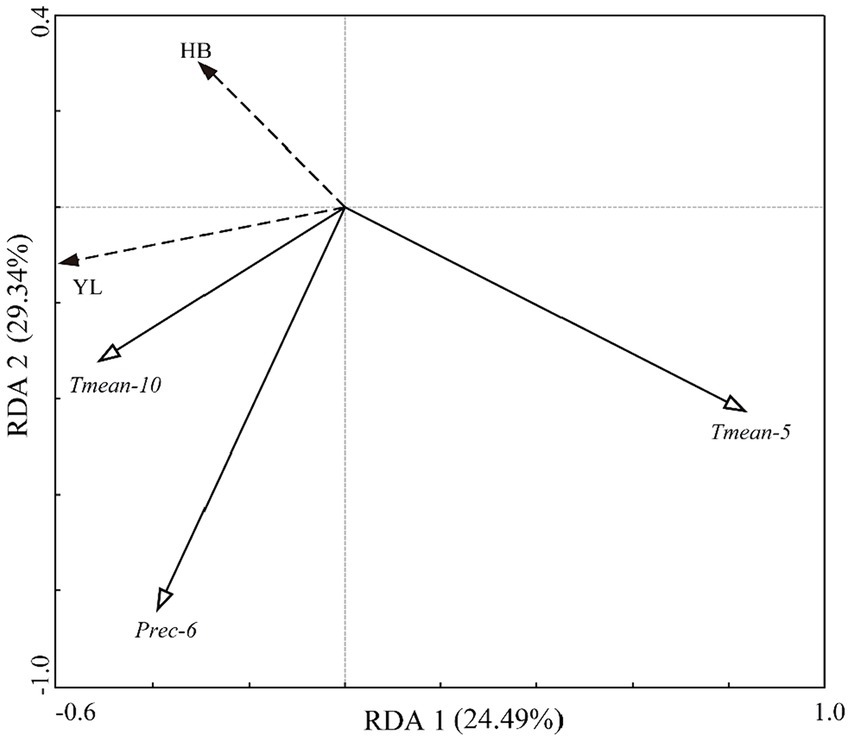
Figure 6. The redundancy analysis (RDA) between chronologies and the climate variables (1966–2016). Only significant (p < 0.05) climate variables were presented. Tmean-10, mean temperature in current October; Tmean-5, mean temperature in current May; Prec-6, precipitation in current June.
The moving response analysis results (Figures 7A,C,E) showed that the growth response of P. yunnanensis to precipitation in May remained relatively stable in HB, by showing positive correlations over the decade with significant periods between 1987 and 1999. The negative response to May temperature reached a significant level before 2000 (p < 0.05) and then weakened. The drought condition in previous September was likely to affect P. yunnanensis growth, as the negative influences of PDSI strengthened in recent 20 years.
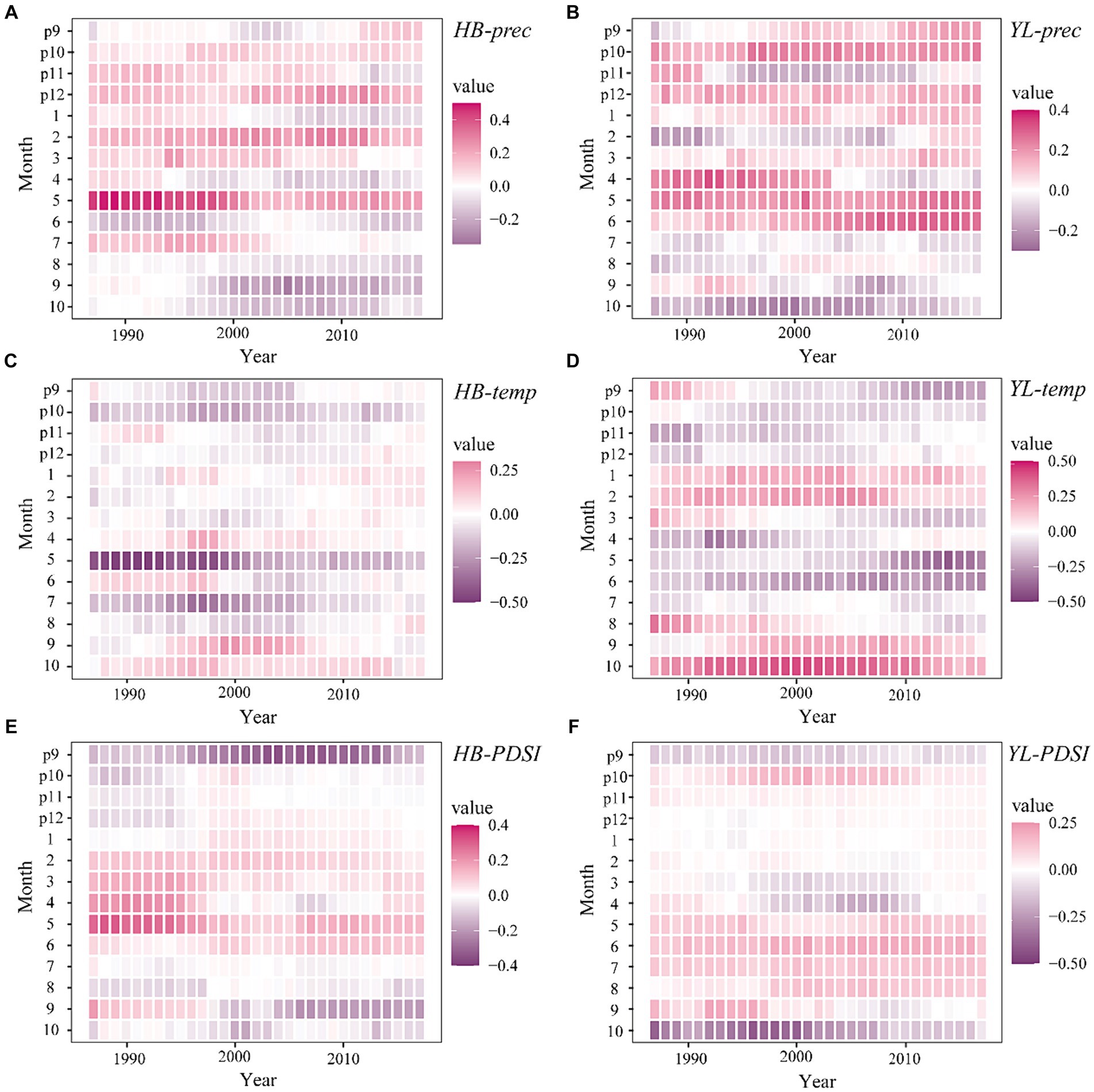
Figure 7. Moving interval analysis between chronologies and the monthly total precipitation (prec) (A), monthly mean temperature (temp) (C) and Palmer Drought Severity Index (PDSI) (E) in HB, and (prec) (B), (temp) (D), (PDSI) (F) in YL from previous September to current October (1987–2017). p: previous year.
In YL, the response of P. yunnanensis to precipitation (positive) and temperature (negative) in current May, temperature in current October (positive), and PDSI (negative) in current October were stable, by showing significant correlations at many years. For current June, the negative impacts of temperature and positive effects of precipitation on tree growth were enhanced after 1992. In Addition, the positive response to PDSI in current June and July fluctuated due to climate change and reached a significant level from 2012 to 2016 (Figures 7B,D,F).
Both response function analysis and RDA revealed that high temperature (Tmax) in May restricted the radial growth of P. yunnanensis at two sites (Figures 5, 6). In general, temperatures rose rapidly during the early growing season (April to May), but the region does not experience a rapid increase in precipitation during these months (Figure 2). This may cause drought stress for tree growth induced by the acceleration of plant transpiration and soil moisture evaporation, which lead to insufficient water supply (Denmead and Shaw, 1962). Similar results have been reported in other conifers in CHM, such as the A. georgei in HB (Sun et al., 2021), A. forrestii and P. likiangensis in YL (Yang et al., 2022), and T. dumosa in YL (Guo G. et al., 2009), indicating the importance of sufficient moisture condition of this period on tree growth in the area. This water-demand effect was also supported by the results of response function analysis, by showing positive relation to May precipitation (Figure 5). High precipitation could compensate for the loss of soil water caused by rapid increase in temperature with high evaporation, consequently alleviate the plant transpiration and avoid stomatal closure. Moreover, the increase in precipitation could supply soil moisture for xylem cell production and the onset of xylogenesis (Shen et al., 2020) during the cambium active period of tree growth in the early growing season (Rossi et al., 2016).
Current October Tmean positively affected the radial growth of P. yunnanensis (Figure 6), suggesting the importance of the impact of post-growing season temperature on tree growth. During the post-growing season, cambial activity tended to be weakened (Gričar et al., 2007). However, elevated temperatures could stimulate photosynthesis and generate nutrients for tree growth, and prolong the growing season, consequently leading to the formation of wider annual ring widths (Dawadi et al., 2013). Similar results have also been found in the growth of A. georgei (Sun et al., 2021) in HB and Abies fargesii (Kharal et al., 2017) in adjacent Manan River Valley of central Himalayas.
The common climate variables with inverse effects between two sites were also observed. Excessive precipitation in June had negative impacts on P. yunnanensis growth in HB. Tree began to fast growth in June on HB and would benefit from high temperature for photosynthesis (Zhang et al., 2020). However, greater precipitation is generally combined with enhanced cloudiness, reduced solar radiation input, and lower temperatures, which might decrease the photosynthetic process (Fan et al., 2019). The constraints of early summer precipitation on tree growth have also been reported in previous studies in four conifers (Zhang et al., 2020), P. yunnanensis (Shen et al., 2020), and A. georgei (Yin et al., 2018) in CHM. In comparison with HB, the sampling site is about 300 m lower in YL, water availability was more important than temperature at lower elevation sites (Fan et al., 2009). Xylem cell production benefits from energy and soil moisture during the cambium active phase (Liang et al., 2014) and enough precipitation can satisfy the water demand of the tree growth in the growing season. This was also demonstrated by growth reaction to PDSI in YL, presenting significant and positive correlations with PDSI in current June (Figure 5).
Furthermore, the observed inverse response can be attributed to variations in soil conditions. The soil is relatively infertile with low organic matter due to steep slopes and shallow soil layers in HB (Su and Wang, 2020), elevated precipitation may trigger soil erosion and therefore endanger its growth. While in YL, the soil is comprised of loam and clay with a higher organic matter content, abundant precipitation may create moist soil conditions for tree growth (Guo L. N. et al., 2009).
The radial growth of P. yunnanensis was affected by winter precipitation in HB (Figure 5). The rise in February precipitation facilitated the formation of a thicker snow cover, effectively shielding roots from potential damage induced by low temperatures and harsh winds at higher altitudes (Zhang et al., 2020). As the snow melted, the heightened soil moisture further supported roots in optimizing water and nutrient uptake, ultimately promoting tree growth (Su and Wang, 2020). The positive effects of winter temperature on tree growth were also highlighted by previous studies at high elevations in the area (Fan et al., 2009; Zhang et al., 2018).
Favorable soil moisture conditions in July promoted the radial growth of P. yunnanensis in YL, since adequate soil moisture could accelerate nutrient dissolution and transport, as well as facilitate root absorption, enriching trees with essential nutrients and alleviating drought stress. A similar response pattern to soil moisture was observed by Sun et al. (2020) in Hengduan Mountains, where P. yunnanensis tree rings were positively correlated with summer moisture availability.
In general, the results of moving interval analysis approved the findings obtained through response function and RDA, by showing significant correlations in many years for those months (e.g., May, June, July, and October), which presented significant relations to tree growth. Furthermore, the stability of climate-growth relationships exhibited variability on an interannual scale. For example, the strongest correlation between tree growth and May temperature in HB was detected from 1990 to 2000, then the strength of the relationship became weakened after the 21st century. This may be attributed to the noticeable temperature surge in Lijiang during that 10-year period (Figure 3), correspondingly, there was a notable strengthening of the positive correlation between growth and May precipitation during this period. The increased precipitation seemed to alleviate the drought stress caused by the sharp temperature rise. Similar results were also presented by the relationship between tree growth and June precipitation, temperature, and PDSI in YL, and other studies that showed the stability of climate-growth relationships varied with warming periods and abrupt climatic changes in the region (Shen et al., 2020).
According to the future climate simulation under the Special Report on Emissions Scenarios (SRES) B2 scenario, the mean temperature in spring, summer, autumn, and winter in Southwest China is projected to increase by 2.6°C, 3.1°C, 2.7°C, and 3.1°C, and precipitation will relatively increase by 8, 7, 6, and 8% in 2071–2,100 (Xu et al., 2005). Incorporating the climate response of tree growth and the prediction of future climate conditions, the productivity of P. yunnanensis forests would likely be enhanced. The radial growth of P. yunnanensis will benefit from increases in precipitation since precipitation is all positively correlated with tree growth (except June precipitation in HB). Although higher temperature in spring and early summer will inhibit tree growth, increases in precipitation of the corresponding seasons is likely to offset the negative impact. Therefore, the growth of P. yunnanensis forests was expected to enhance under future climate change in the area.
We studied the climate-growth relationships of P. yunnanensis in CHM and predicted the growth of its predominant forest. Both temperature and precipitation affected its growth, and tree growth showed common and site-specific responses to climate change. Moisture conditions in May would be the most critical factor influencing its growth, while June precipitation and current October temperature were other two common factors affecting P. yunnanensis growth. Although intense warming of spring and early summer months in the future may exacerbate drought stress on forests, we believe that a high percentage of increased precipitation would offset insufficient water supply induced by warming. According to the results, the radial growth of P. yunnanensis may increase under future climate conditions in our study area. Further studies on species’ drought response mechanism are required and sustainable strategies of forest management should be focused on improving resistance to drought.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
SX: Validation, Writing – review & editing, Writing – original draft, Visualization, Software, Conceptualization. TY: Writing – review & editing, Methodology, Investigation, Data curation. XS: Writing – review & editing, Methodology, Data curation. HC: Writing – review & editing, Methodology, Data curation. MS: Writing – review & editing, Methodology, Investigation, Data curation. YZ: Writing – review & editing, Validation, Supervision, Resources, Methodology, Conceptualization.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research was supported by Agricultural Joint Project of Yunnan Province Science and Technology Department (No. 202101BD070001-098).
We are very grateful to staffs of two Nature Reserve Administration for their great support for our experiment.
XS was employed by Anji Yinjiang Agriculture Development Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aber, J., Neilson, R. P., McNulty, S., Lenihan, J. M., Bachelet, D., and Drapek, R. J. (2001). Forest processes and global environmental change: predicting the effects of individual and multiple stressors. Bioscience 51, 735–751. doi: 10.1641/0006-3568(2001)051[0735,FPAGEC]2.0.CO;2
Alley, W. M. (1985). The palmer drought severity index as a measure of hydrological drought. J. Am. Water Resour. Assoc. 21, 105–114. doi: 10.1111/j.1752-1688.1985.tb05357.x
Aryal, S., Gaire, N. P., Pokhrel, N. R., Rana, P., Sharma, B., Kharal, D. K., et al. (2020). Spring season in western Nepal Himalaya is not yet warming: a 400-year temperature reconstruction based on tree-ring widths of Himalayan hemlock (Tsuga dumosa). Atmos. 11:132. doi: 10.3390/atmos11020132
Biondi, F., and Waikul, K. (2004). DENDROCLIM2002: A C++ program for statistical calibration of climate signals in tree-ring chronologies. Comput. Geosci. 30, 303–311. doi: 10.1016/j.cageo.2003.11.004
Bohner, T., and Diez, J. (2021). Tree resistance and recovery from drought mediated by multiple abiotic and biotic processes across a large geographic gradient. Sci. Total Environ. 789:147744. doi: 10.1016/j.scitotenv.2021.147744
Camarero, J. J., Gazol, A., Sánchez-Salguero, R., Fajardo, A., McIntire, E. J. B., and Liang, E. (2021). Tree growth and treeline responses to temperature: different questions and concepts. Glob. Chang. Biol. 27, e13–e14. doi: 10.1111/gcb.15728
Cook, E. R. (1985). A time series analysis approach to tree ring standardization. University of Arizona, Tucson, AZ, USA.
Dawadi, B., Liang, E., Tian, L., Devkota, L. P., and Yao, T. (2013). Pre-monsoon precipitation signal in tree rings of timberline Betula utilis in the Central Himalayas. Quat. Int. 283, 72–77. doi: 10.1016/j.quaint.2012.05.039
Deng, X. Q., Huang, B. L., Wen, Q. Z., Hua, C. L., Tao, J., and Zheng, J. X. (2014). Dynamic of Pinus yunnanensis forest resources in Yunnan. J. Nat. Resour. 29, 1411–1419. doi: 10.11849/zrzyxb.2014.08.013
Denmead, O. T., and Shaw, R. H. (1962). Availability of soil water to plants as affected by soil moisture content and meteorological conditions. Agron. J. 54, 385–390. doi: 10.2134/agronj1962.00021962005400050005x
Drobyshev, I., Gewehr, S., Berninger, F., and Bergeron, Y. (2013). Species specific growth responses of black spruce and trembling aspen may enhance resilience of boreal forest to climate change. J. Ecol. 101, 231–242. doi: 10.1111/1365-2745.12007
Du, Q., Rossi, S., Lu, X., Wang, Y., Zhu, H., Liang, E., et al. (2020). Negative growth responses to temperature of sympatric species converge under warming conditions on the southeastern Tibetan plateau. Trees 34, 395–404. doi: 10.1007/s00468-019-01924-4
Dunn, R. J. H., Aldred, F., Gobron, N., Miller, J. B., Willett, K. M., Ades, M., et al. (2022). Global climate. Bull. Am. Meteorol. Soc. 103, S11–S142. doi: 10.1175/BAMS-D-22-0092.1
Fan, Z. X., Bräuning, A., and Cao, K. F. (2008). Annual temperature reconstruction in the central Hengduan Mountains, China, as deduced from tree rings. Dendrochronologia 26, 97–107. doi: 10.1016/j.dendro.2008.01.003
Fan, Z. X., Bräuning, A., Cao, K. F., and Zhu, S. D. (2009). Growth-climate responses of high-elevation conifers in the central Hengduan Mountains, southwestern China. For. Ecol. Manag. 258, 306–313. doi: 10.1016/j.foreco.2009.04.017
Fan, Z. X., Bräuning, A., Fu, P. L., Yang, R. Q., Qi, J. H., Grießinger, J., et al. (2019). Intra-annual radial growth of Pinus kesiya var. langbianensis is mainly controlled by moisture availability in the Ailao Mountains, southwestern China. Forests 10:899. doi: 10.3390/f10100899
Fang, O., Zhang, Q. B., Vitasse, Y., Zweifel, R., and Cherubini, P. (2021). The frequency and severity of past droughts shape the drought sensitivity of juniper trees on the Tibetan plateau. For. Ecol. Manag. 486:118968. doi: 10.1016/j.foreco.2021.118968
Fritts, H. C., Blasing, T. J., Hayden, B. P., and Kutzbach, J. E. (1971). Multivariate techniques for specifying tree-growth and climate relationships and for reconstructing anomalies in paleoclimate. J. Appl. Meteorol. 10, 845–864. doi: 10.1175/1520-0450(1971)010<0845:MTFSTG>2.0.CO;2
Fritts, H. C., Smith, D. G., Cardis, J. W., and Budelsky, C. A. (1965). Tree-ring characteristics along a vegetation gradient in northern Arizona. Ecology 46, 393–401. doi: 10.2307/1934872
Fu, L. G., Li, N., and Robert, R. M. (1999). “Pinus” in Flora of China. eds. Z. Y. Wu and P. H. Raven (Beijing: Science Press), 11–25.
Girardin, M. P., Tardif, J., Flannigan, M. D., and Bergeron, Y. (2004). Multicentury reconstruction of the Canadian drought code from eastern Canada and its relationship with paleoclimatic indices of atmospheric circulation. Clim. Dyn. 23, 99–115. doi: 10.1007/s00382-004-0417-x
Gričar, J., Zupančič, M., Čufar, K., and Primož, O. (2007). Regular cambial activity and xylem and phloem formation in locally heated and cooled stem portions of Norway spruce. Wood Sci. Technol. 41, 463–475. doi: 10.1007/s00226-006-0109-2
Guo, L. N., He, Z. J., Long, X. Z., Wang, J. Z., Wang, L. D., and Li, C. X. (2009). Genetic characteristics and taxonomy of soil in the Yulong Snow Mountain. J. South. Agric. 40, 1177–1183.
Guo, G., Li, Z. S., Zhang, Q. B., Ma, K. P., and Mu, C. (2009). Dendroclimatological studies of Picea Likiangensis and Tsuga Dumosa in Lijiang, China. IAWA J. 30, 435–441. doi: 10.1163/22941932-90000230
Harris, I., Osborn, T. J., Jones, P., and Lister, D. (2020). Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 7:109. doi: 10.1038/s41597-020-0453-3
He, M., Yang, B., Bräuning, A., Rossi, S., Ljungqvist, F. C., Shishov, V., et al. (2019). Recent advances in dendroclimatology in China. Earth Sci. Rev. 194, 521–535. doi: 10.1016/j.earscirev.2019.02.012
Holmes, R. L. (1983). Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 43, 69–78.
Huang, J., Tardif, J. C., Bergeron, Y., Denneler, B., Berninger, F., and Girardin, M. P. (2010). Radial growth response of four dominant boreal tree species to climate along a latitudinal gradient in the eastern Canadian boreal forest. Glob. Chang. Biol. 16, 711–731. doi: 10.1111/j.1365-2486.2009.01990.x
IPCC (2021). “Summary for Policymakers” in Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. eds. V. Masson-Delmotte, P. Zhai, A. Pirani, S. L. Connors, C. Péan, and S. Berger, et al. (Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press), 3–32.
Kharal, D. K., Thapa, U. K., George, S. S., Meilby, H., Rayamajhi, S., and Bhuju, D. R. (2017). Tree-climate relations along an elevational transect in Manang Valley, Central Nepal. Dendrochronologia 41, 57–64. doi: 10.1016/j.dendro.2016.04.004
Li, Z., Shi, C., Liu, Y., Zhang, J., and Ma, K. (2011). Winter drought variations based on tree-ring data in Gaoligong Mountain, northwestern Yunnan, China, A. D. 1795-2004. Pak. J. Bot. 43, 2469–2478.
Li, Z. S., Zhang, Q. B., and Ma, K. (2012). Tree-ring reconstruction of summer temperature for A.D. 1475-2003 in the central Hengduan Mountains, northwestern Yunnan, China. Clim. Chang. 110, 455–467. doi: 10.1007/s10584-011-0111-z
Liang, E., Dawadi, B., Pederson, N., and Eckstein, D. (2014). Is the growth of birch at the upper timberline in the Himalayas limited by moisture or by temperature? Ecology 95, 2453–2465. doi: 10.1890/13-1904.1
Liang, E., Wang, Y., Xu, Y., Liu, B., and Shao, X. (2010). Growth variation in Abies georgei var. smithii along altitudinal gradients in the Sygera Mountains, southeastern Tibetan plateau. Trees 24, 363–373. doi: 10.1007/s00468-009-0406-0
Martínez-Vilalta, J., López, B. C., Adell, N., Badiella, L., and Ninyerola, M. (2008). Twentieth century increase of Scots pine radial growth in NE Spain shows strong climate interactions: recent changes in scots pine radial growth. Glob. Chang. Biol. 14, 2868–2881. doi: 10.1111/j.1365-2486.2008.01685.x
Morzuch, B. J., and Ruark, G. A. (1991). Principal components regression to mitigate the effects of multicollinearity. For. Sci. 37, 191–199. doi: 10.1093/forestscience/37.1.191
Panthi, S., Bräuning, A., Zhou, Z. K., and Fan, Z. X. (2018). Growth response of Abies georgei to climate increases with elevation in the central Hengduan Mountains, southwestern China. Dendrochronologia 47, 1–9. doi: 10.1016/j.dendro.2017.11.001
Rossi, S., Anfodillo, T., Čufar, K., Cuny, H. E., Deslauriers, A., Fonti, P., et al. (2016). Pattern of xylem phenology in conifers of cold ecosystems at the northern hemisphere. Glob. Change Biol. Bioenergy. 22, 3804–3813. doi: 10.1111/gcb.13317
Schnabel, F., Purrucker, S., Schmitt, L., Engelmann, R. A., Kahl, A., Richter, R., et al. (2022). Cumulative growth and stress responses to the 2018-2019 drought in a European floodplain forest. Glob. Chang. Biol. 28, 1870–1883. doi: 10.1111/gcb.16028
Shen, J. Y., Li, Z. S., Gao, C. J., Li, S. F., Huang, X. B., Lang, X. D., et al. (2020). Radial growth response of Pinus yunnanensis to rising temperature and drought stress on the Yunnan plateau, southwestern China. For. Ecol. Manag. 474:118357. doi: 10.1016/j.foreco.2020.118357
Su, H., and Wang, P. (2020). Vertical differentiation of soil organic matter in Haba Snow Mountain nature reserve. Yunnan Geogr. Environ. Res. 32, 65–76.
Su, X. C., Wang, L., Li, Q. L., and Teng, C. (2014). Study of surface dry and wet conditions in Southwest China in recent 50 years. J. Nat. Resour. 29, 104–116.
Sun, L., Cai, Y., Zhou, Y., Shi, S., Zhao, Y., Gunnarson, B. E., et al. (2020). Radial growth responses to climate of Pinus yunnanensis at low elevations of the Hengduan Mountains, China. Forests 11:1066. doi: 10.3390/f11101066
Sun, M., Li, J., and Zhang, Y. (2021). Climate-growth relations of Abies georgei along an altitudinal gradient in Haba Snow Mountain, southwestern China. Forests 12:1569. doi: 10.3390/f12111569
ter Braak, C. J. F. (1994). Canonical community ordination. Part I: basic theory and linear methods. Écoscience 1, 127–140. doi: 10.1080/11956860.1994.11682237
Van Passel, J., De Keersmaecker, W., Bernardino, P. N., Jing, X., Umlauf, N., Van Meerbeek, K., et al. (2022). Climatic legacy effects on the drought response of the Amazon rainforest. Glob. Chang. Biol. 28, 5808–5819. doi: 10.1111/gcb.16336
Wang, Y., Zhang, Y., Fang, O., and Shao, X. (2018). Long-term changes in the tree radial growth and intrinsic water-use efficiency of Chuanxi spruce (Picea likiangensis var. balfouriana) in southwestern China. J. Geog. Sci. 28, 833–844. doi: 10.1007/s11442-018-1508-7
Xu, Y. L., Huang, X. Y., Zhang, Y., Lin, W. T., and Lin, E. D. (2005). Statistical analyses of climate change scenarios over China in the 21st century. Adv. Clim. Chang. Res. 1, 80–97.
Yang, R. Q., Fan, Z. X., Li, Z. S., and Wen, Q. Z. (2018). Radial growth of Pinus yunnanensis at different elevations and their responses to climatic factors in the Yulong Snow Mountain, Northwest Yunnan, China. Acta Ecol. Sin. 38, 8983–8991. doi: 10.5846/stxb201805311214
Yang, R. Q., Fu, P. L., Fan, Z. X., Panthi, S., Gao, J., Niu, Y., et al. (2022). Growth-climate sensitivity of two pine species shows species-specific changes along temperature and moisture gradients in Southwest China. Agric. For. Meteorol. 318:108907. doi: 10.1016/j.agrformet.2022.108907
Yin, D. C., Xu, D., Tian, K., Xiao, D., Zhang, W., Sun, D., et al. (2018). Radial growth response of Abies georgei to climate at the upper timberlines in central Hengduan Mountains, southwestern China. Forests 9:606. doi: 10.3390/f9100606
Young, D. J. N., Stevens, J. T., Earles, J. M., Moore, J., Ellis, A., Jirka, A. L., et al. (2017). Long-term climate and competition explain forest mortality patterns under extreme drought. Ecol. Lett. 20, 78–86. doi: 10.1111/ele.12711
Yu, J., Liu, Q., Meng, S., Zhou, G., Shah, S., and Xu, Z. (2018). Summer temperature variability inferred from tree-ring records in the central Hengduan Mountains, southeastern Tibetan plateau. Dendrochronologia 51, 92–100. doi: 10.1016/j.dendro.2018.08.004
Yue, W., Zhao, X., Chen, F., Khan, A., and Gao, Z. (2022). Reconstructed precipitation in the Lohit River basin, southern Tibetan plateau since 1720 CE and its weak linkages with monsoon-season Brahmaputra discharge. Int. J. Climatol. 42, 3793–3808. doi: 10.1002/joc.7445
Zhang, Y., Cao, R. J., Yin, J., Tian, K., Xiao, D. R., Zhang, W. G., et al. (2020). Radial growth response of major conifers to climate change on Haba Snow Mountain, southwestern China. Dendrochronologia 60:125682. doi: 10.1016/j.dendro.2020.125682
Zhang, Y., Yin, D. C., Sun, M., Wang, H., Tian, K., Xiao, D. R., et al. (2017). Variations of climate-growth response of major conifers at upper distributional limits in Shika Snow Mountain, northwestern Yunnan plateau, China. Forests 8:377. doi: 10.3390/f8100377
Keywords: tree ring, conifer, dendroclimatology, climate change, climate-growth relationships
Citation: Xie S, Yan T, Sun X, Chen H, Sun M and Zhang Y (2024) Radial growth response of Pinus Yunnanensis to climate in high mountain forests of northwestern Yunnan, southwestern China. Front. For. Glob. Change. 7:1343730. doi: 10.3389/ffgc.2024.1343730
Edited by:
Lucian Dinca, Retired, Brasov, RomaniaReviewed by:
Maciej Pach, University of Agriculture in Krakow, PolandCopyright © 2024 Xie, Yan, Sun, Chen, Sun and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yun Zhang, emhhbmd5dW5jb29sQDE2My5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.