 Dehuang Zhu
Dehuang Zhu Yan Zhou
Yan Zhou Suhong Peng
Suhong Peng Weiping Hua
Weiping Hua Bin Luo
Bin Luo Dafeng Hui
Dafeng Hui- 1College of Ecology and Resources Engineering, Wuyi University, Nanping, China
- 2Wuyishan National Park Scientific Research Monitoring Center, Nanping, China
- 3College of Chemistry and Bioengineering, Yichun University, Yichun, China
- 4Department of Biological Sciences, Tennessee State University, Nashville, TN, United States
Background: Ecological stoichiometric characteristics of carbon (C), nitrogen (N), phosphorus (P), and potassium (K) serve as crucial indicators of nutrient cycling and limitation in terrestrial ecosystems. However, our current understanding of stoichiometric characteristics in subtropical forests and their response to different climate conditions is still limited.
Methods: We selected six altitudes ranging from 700 m to 1,200 m to simulate different climate conditions of an evergreen broadleaf forest in Wuyi Mountain, Fujian Province, China. We investigated C, N, P, and K stoichiometry and homeostasis in the green leaves, newly senesced leaf litter (fresh litter), and soil of this forest.
Results: Leaf P and K levels showed a decline with increasing altitude. Notably, the stoichiometric ratios in different components exhibited a bimodal distribution along the altitudinal gradient. Additionally, a decline trend of N resorption efficiencies was observed as altitude increased. Moreover, weak homeostasis was observed in P and K in green leaves. These findings highlighted the significant impact of altitude on the stoichiometry in evergreen broadleaf forest. This study also contributed to our understanding of the nutrient cycling mechanism and plant growth strategies of evergreen forests under different climate conditions.
1 Introduction
Mountains provide a unique and ideal platform for studying plant adaptation strategies to environmental changes. Climatic factors such as temperature, solar radiation, and precipitation vary along the altitudinal gradient (Beniston, 2003; Huber et al., 2007; Korner, 2007). Therefore, the altitudinal gradient has been received widely attention (Körner, 1998; Wang et al., 2018; Hou et al., 2019). Environmental conditions such as temperature and precipitation vary at different altitudes, and have direct impacts on plant growth by regulating the nutrient status. Therefore, studying the nutrient status of forest ecosystem along altitude can provide valuable insights into the nutrient cycling under different climate conditions (Sheikh et al., 2020; Kumar et al., 2021).
Nutrient cycling plays a crucial role in forest ecosystems, as it forms a plant leaf-litter-soil continuum (Legout et al., 2020; Lin et al., 2021). The cycling of elemental nutrients in forest ecosystems is regulated by biological and abiotic factors, such as tree species diversity and acid rain (De Groote et al., 2018; Hu et al., 2019). However, the impact of altitude on nutrient cycling within the plant leaf-litter-soil continuum still has not been well investigated (Kumar et al., 2021; Li et al., 2022). While altitude has been used as a proxy for studying different climate conditions effect on nutrient cycling, current studies have mainly focused on the nutrient characteristics and limitation of plant (Yu et al., 2014; Du et al., 2016), and the nutrient status of soil and leaf (Müller et al., 2017; Hou et al., 2019; Sheikh et al., 2020). However, these studies provide limited insights into our comprehensive understanding of nutrient cycling under different climate conditions. Therefore, it is crucial to conduct further studies investigating the processes involved in nutrient cycling. This will enhance our understanding of plant growth strategies and contribute to a more comprehensive understanding of nutrient cycling. Ecological stoichiometry, which characterizes the nutritional status of forest ecosystems, serves as a valuable tool for studying nutrient cycling.
Ecological stoichiometry is used to explore allocation of nutrient in organisms and cycling of element in plant leaf, above-ground litter (i.e., fresh litter and semi-decomposed litter), and soil components (Sterner and Elser, 2002; Zhang et al., 2019). Ratios of elements are commonly used to assess nutrient limitation and coupling relationships across these different components (Yang et al., 2020; Wang et al., 2021). Nevertheless, the nutrient stoichiometry of green leaves, fresh litter, and soil in forest ecosystems can be significantly altered by altitudinal variations, which lead to changes in soil microclimates (Chen et al., 2022). Studies have demonstrated that foliar C:N (carbon: nitrogen) and C:P (carbon: phosphorus) increase with altitude, while foliar N and P concentrations decrease (Müller et al., 2017). The C, N, and P contents, as well as C:N:P stoichiometry of leaf litter, exhibit wide variations along the altitudinal gradient, displaying distinct trends depending on species composition at different altitudes (Li et al., 2022; He et al., 2023). Soil available N tends to increase with altitude, whereas soil available P and total P concentrations in plant tissues decrease (Wang et al., 2018). Soil organic C generally decreases with altitude (Hou et al., 2019; Sheikh et al., 2020; Mganga et al., 2022), while soil nutrients demonstrate either an unimodal response (Nie et al., 2023) or an increasing trend (He et al., 2021) along the altitudinal gradient. Soil P fractions generally reach a maximum at mid-altitude due to the presence of more active microorganism that accelerate P weathering (Li et al., 2023). Additionally, soil available P was highly correlated with leaf P along the altitudinal gradient (Li et al., 2023). Unlike C, N, and P, the investigation of potassium (K) and its relationships with other elements has been relatively limited (Zhang et al., 2021; Sun et al., 2022), especially at different altitudes (Zhang et al., 2019). However, K is a vital element for the plant growth and its levels vary with different environments (Zhang et al., 2019; Li et al., 2022). Therefore, studying C, N, P, K, and their stoichiometric ratios across green leaves, leaf litter, and soil components is essential for understanding nutrient cycling and the responses of forest ecosystems to different climate conditions.
Stoichiometric homeostasis is a fundamental concept in ecological stoichiometry, and it encompasses nutrient cycling and physiological and biochemical adaptation of plant in response to environment (Sterner and Elser, 2002; Hessen et al., 2004). The organisms show strict homeostasis when their stoichiometric compositions remain unchanged with variations in stoichiometric resources; otherwise, it is considered non-homeostasis (Sterner and Elser, 2002). Stoichiometric homeostasis has been observed in diverse ecosystems, including grass ecosystems (Yu et al., 2011), wetlands (Demars and Edwards, 2007), and dry plantations (Li et al., 2022). Plants show homeostasis characteristics in response to nutrient limitation, such as N-or P-limitation at different growth stages in forests (Bai et al., 2019; Wang et al., 2019). They adopted conservation strategies for resource utilization, exhibiting lager homeostasis (Yu et al., 2011). Stoichiometric homeostasis can reflect soil nutrient limitation and significantly affect plant growth (Yu et al., 2015). However, stoichiometric homeostasis has received relatively less attention, particularly in mountains. Furthermore, different climate conditions can impact soil nutrient status through various mechanisms. Consequently, the response of leaf stoichiometry to soil nutrient availability along the altitude gradient remains unclear.
Nutrient resorption efficiency is a key parameter for understanding nutrient cycling and ecosystem balance. It helps to characterize nutrient allocation and strategies under environmental changes (Milla et al., 2005). Nutrient resorption refers to the uptake of nutrients from senesced leaves and nutrient allocation and utilization of those nutrients (Chen et al., 2021a; Wang et al., 2022). Previous studies have found that P and K resorption efficiencies are significantly higher than N resorption efficiency in the nutrient-poor soils of a tropical forest (Urbina et al., 2021). This suggest that the soil nutrient availability directly influences nutrient allocation in plants. Climate factors also play a role in leaf nutrient resorption. For example, P resorption efficiency tends to be higher than N resorption efficiency in Chinese fir plantation under different climate conditions (Tong et al., 2021). While nutrient status in mountain ecosystems has been investigated, few studies have been done on nutrient resorption efficiencies in these ecosystems under different climate conditions. Therefore, further investigations are necessary to understand the stoichiometric characteristics, homeostasis, and leaf nutrient resorption efficiencies in mountain forest ecosystems.
Wuyi Mountain, located within Wuyishan National Park, is one of the first five national parks established in China. This subtropical primary forest ecosystem is widely recognized for its extensive and representative distribution. It serves as a sanctuary for unique biodiversity and displays distinct vertical vegetational zonation (You et al., 2017). Given its exceptional characteristics, Wuyi Mountain is an ideal study site for investigating nutrient status of forests ecosystem in responses to different climate conditions. In this study, we formulated the following hypotheses: (1) Stoichiometric characteristics of different components are influenced by altitude. (2) Leaf nutrient resorption efficiencies are affected by altitude due to variation in temperature and humidity. (3) Nutrient resorption efficiencies are directly regulated by soil and leaf nutrients. The findings of this study will contribute to our scientific understanding of the nutrient status of evergreen broadleaf forests.
2 Study area and methods
2.1 Study sties
The study site is located in Wuyishan National Park of the northern Fujian Province, China. The park covers an area of 1,001.41 km2. The area belongs to a typical subtropical monsoon climate. The mean annual temperature is 17–19°C, mean annual precipitation is 1,684–1,780 mm, and the frost-free period is 234 days. The Park consists of subtropical evergreen broadleaved forests and central subtropical evergreen broadleaved forests, with Castanopsis eyrei, Castanopsis carlesii, Schima superba, Rhododendron ovatum, Quercus phillyraeoides, and Rhododendron latoucheae as dominant species. The original forest area in the park is 210.70 km2, as reported by the Science and Technology Commission of Fujian Province in 1993. One prominent feature is the vertical belt spectrum of vegetation, including evergreen broadleaved forest, coniferous and broad-leaved mixed forest, temperate coniferous forest, moss dwarf forest of middle mountain, and meadow of middle mountain. Wuyi Mountain has a variety of soil types, including mountain red soil, yellow red soil, yellow soil, and mountain meadow soil.
2.2 Experimental design
In Wuyi mountain, evergreen broadleaved forest is mainly distributed below 1,200 m, and the study area with less human disturbance was selected above 700 m. In September 2022, the plot sites were set at 700 m, 800 m, 900 m, 1,000 m, 1,100 m, and 1,200 m in the evergreen broadleaved forest (Supplementary Figure S1; Supplementary Table S1). Four 10 m × 10 m quadrats were established at each site, with a total of 24 quadrats a total area of 2,400 m2. Geographic information such as longitude, latitude, slope, slope position, and slope direction at each site were recorded (Supplementary Table S1), and a vegetation survey was conducted. The dominate trees in the plots included C. eyrei.
To analyze leaf nutrient resorption, green leaves and fresh leaf litter of dominate species of tree layer were collected at each altitude. The mature green leaves were also collected from three dominant trees in each plot. In the middle of each quadrat, a leaf litter collector (1 × 1 m2) was set for collecting fresh litter of dominant species for the chemical element analysis from September 3rd to December 31st in 2022. Four litter collectors were set at each altitude, with a total of 24 litter collectors. In addition, three soil samples were taken at depths of 0–20 cm and 20–40 cm using a soil drill from each quadrat. A total of 72 green leaf samples, 72 fresh litter samples, and 144 soil samples were collected, respectively. The green leaf and fresh litter samples were dried at 80°C for 48 h to keep the constant weight, while the soil samplings were naturally air-dried. The green leaves, fresh litter, and soil were filtered through a 2 mm sieve and then packed in plastic self-sealing bag for chemical element determination.
2.3 Functional trait and chemical element measurement
The total C, N, P, and K of green leaves, fresh litter, and soil were analyzed by the potassium dichromate KCr2O7, the Kjeldahl method, the digestion with H2SO4–H2O2 solution, and the atomic flame photometry, respectively (Jones, 2001). Available N (AN), available P (AP), and available K (AK) of soil were determined by the ultraviolet spectrophotometry, the NaHCO3 extraction molybdenum-antimony colorimetry, and the atomic flame photometry, respectively (Jones, 2001). pH of the soil was analyzed by a pH meter (PHS-3E, Leici, China).
2.4 Data analysis
The leaf nutrient resorption efficiencies were calculated by the following equation (Vergutz et al., 2012; Eq. 1):
where NuRE represents the N (NRE), P (PRE), and K (KRE) resorption efficiencies. Nusenesced and Nugreen are N, P, or K contents of the fresh litter and green leaves along different altitudes, respectively. MLCF is the mass loss factor during the leaf senescence (van Heerwaarden et al., 2003), with the vlaues of 0.780 and 0.745 for evergreen and confiers tree species, respectively (Vergutz et al., 2012).
The stoichiometric homeostasis of the green leaves was quantified by a homeostasis coefficient (Eq. 2):
where y is the contents or stoichiometric ratio of leaf C, N, P, or K. x represents the contents or stoichiometric ratio of soil C, N, P, or K. H represents the homeostasis coefficient. The parameters a and H are estimated using linear regression. The homeostasis characteristics are judged based on H: 1 < H < 1.3 indicates “plastic,” 1.3 < H < 2 indicates “weak plastic,” 2 < H < 4 indicates “weak homeostatic,” and H > 4 indicates “homeostatic” (Persson et al., 2010).
The C, N, P, and K contents and their stoichiometric ratios of green leaves, fresh litter, and soil at a given altitude were calculated using three samples in each qudrate of four plots at each of the six altitudes. Mean and standard error were presented in this study. The contents of C, N, P, and K and stoichimetric ratios of different components at the same altitude and the same component at different altitudes, were tested by ANOVA. The least significant difference (LSD) method was applied for multiple comparison when a significant effect was detected. Leaf nutrient resorption efficiencies at different altitudes were also analyzed by ANOVA. To investigat the effects of altitude on leaf nutrient resorption efficiencies, linear regression models were used. Redundancy analysis (RDA) was performed to visualize the effects of nutrients in different components on leaf nutrient resorption efficiencies. Pearson correlation coefficient were utilized to examine the relationships between leaf nutrient resorption efficiencies and soil nutrients, green leaves and fresh litter nutrients, and stoichiometric ratios. All data were checked for homogeneity and normality before analysis. All data analyses and figures were carried out using R version 4.2.1 (R Core Team, 2020).
3 Results
3.1 C, N, P, and K contents and stoichiometric characteristics
The nutrient contents and stoichiometric ratios within the plant leaf, litter, and soil exhibited altitudinal variations. Green leaf N and P decreased first, then increased, and finally decreased again with the increase of altitude. However, leaf C did not change significantly with altitude (Figure 1). Leaf K content also decreased with increasing of altitude, while fresh litter C content significantly increased with altitude. The highest value of fresh litter N (14.72 mg/g) was observed at 1200 m, which was significantly higher than at other altitudes. Fresh litter P and K contents decreased significantly with altitude. The contents of SOC and total N showed double peaks, occurring at 800 m (SOC: 10.32 mg/g, total N: 3.98 mg/g) and 1,100 m (SOC: 11.16 mg/g, total N: 3.78 mg/g), respectively. Soil total P content significantly initially decreased and then increased with altitude. The ratios of C:P and N:P of green leaves, and the C:P ratio of the litter increased first and then decreased with altitude (Supplementary Table S2).
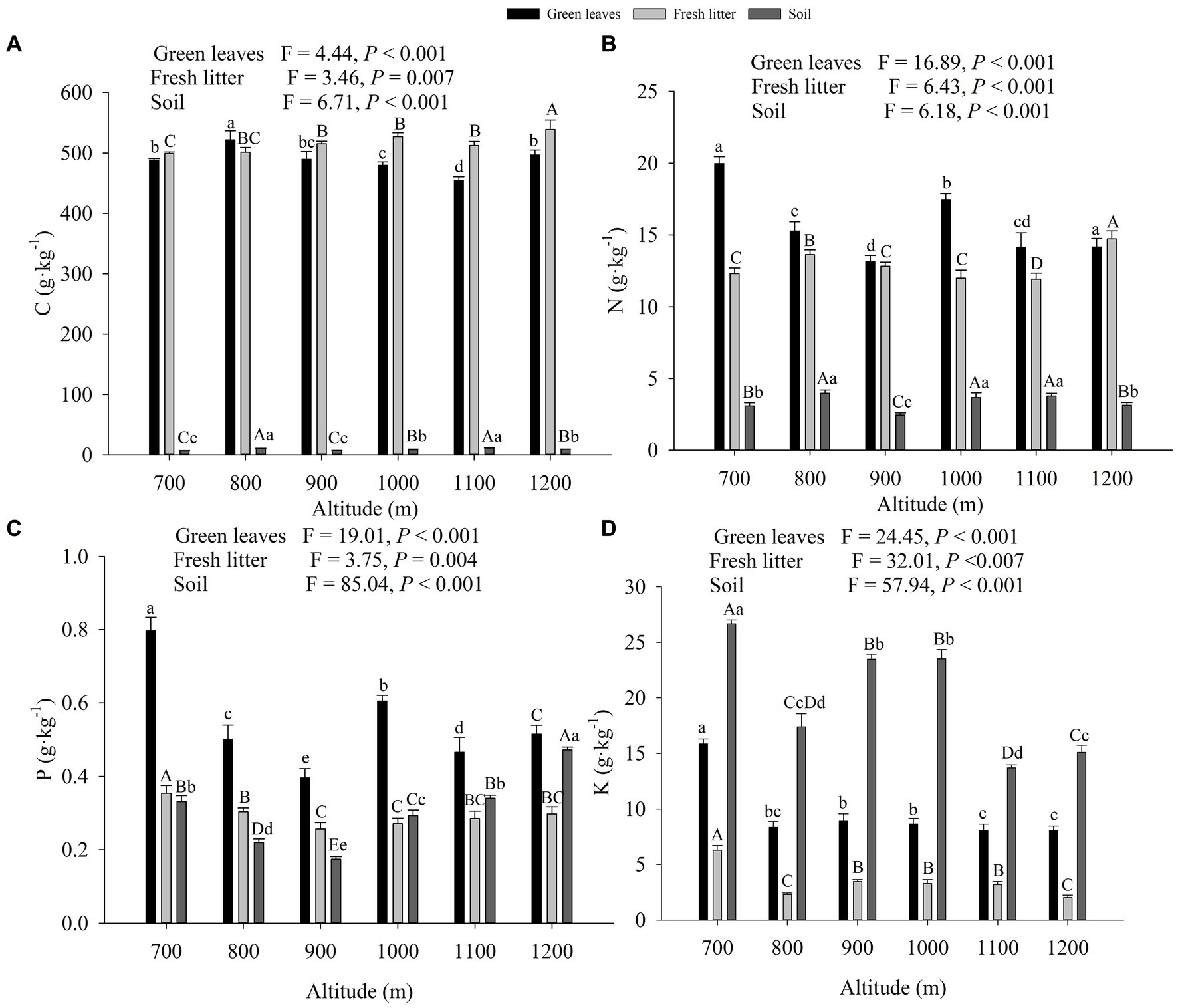
Figure 1. The mean (±SE) contents of C, N, P, and K of green leaves, fresh litter, and soil in Wuyi Mountain along an altitudinal gradient. Different lowercase letters, capital letters, and capital and lowercase letters indicate significant differences for green leaves, fresh litter, and soil among different altitudes, respectively (p < 0.05, n = 72).
In Wuyi Mountain, fresh litter C content was significantly higher than those of green leaves and soil (Table 1). N content ranked in the following order: Green leaves > Fresh litter > Soil (0–20 cm) > Soil (20–40 cm). Green leaves had the highest P (0.54 ± 0.02 mg/g) and K (9.55 ± 0.37 mg/g) contents compared to the other components. Soil AP and AK at a depth of 0–20 cm were significantly higher than those at the depth of 20–40 cm. The stoichiometric ratios of fresh litter were significantly higher than those of green leaves and soil.
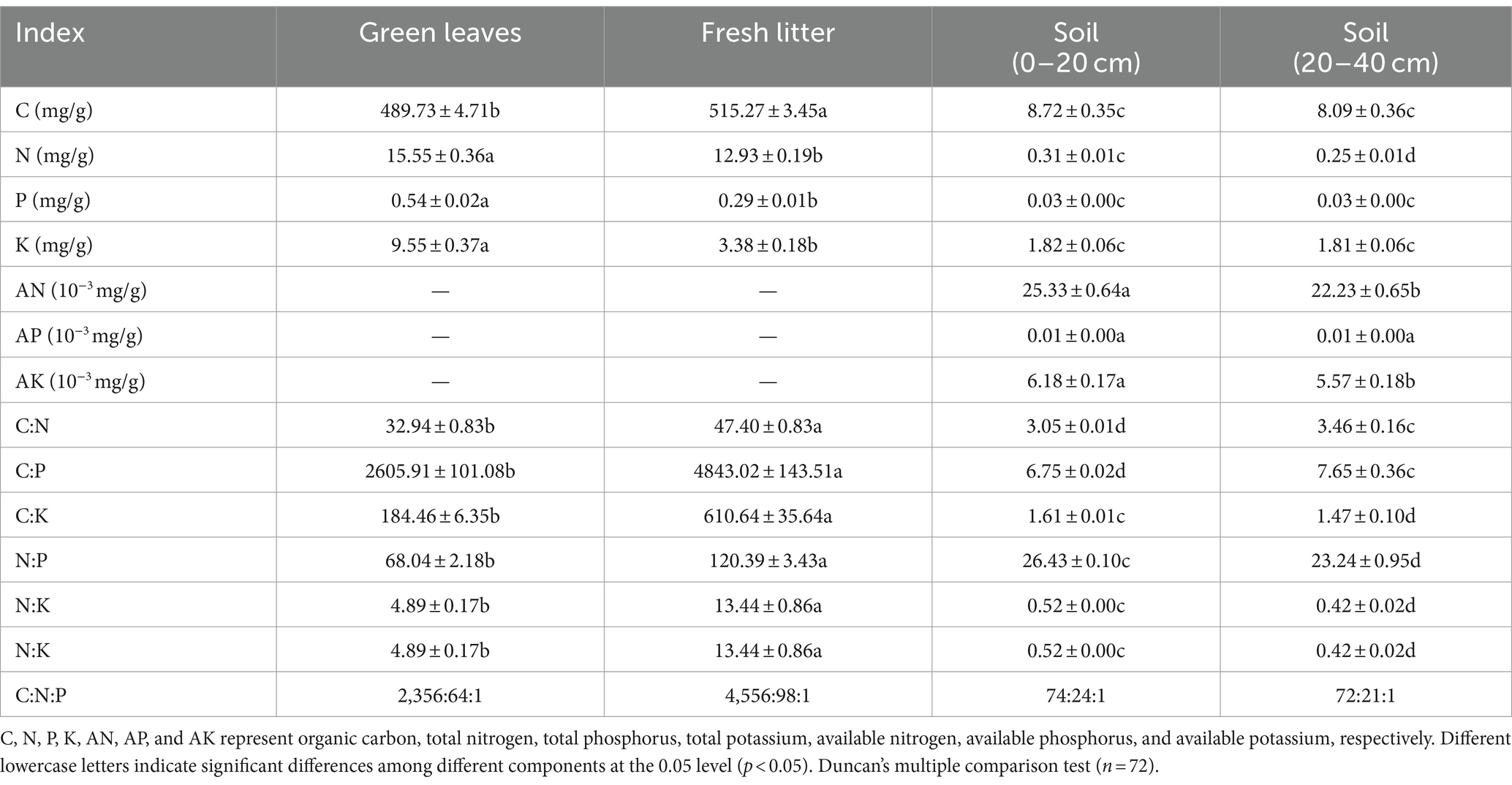
Table 1. N, P, and K contents and their stoichiometric ratios of green leaves, fresh litter, and soil in Wuyi Mountain (mean ± se of six altitudes).
3.2 Nutrient resorption efficiencies and ecological stoichiometric homeostasis
Nitrogen resorption efficiency decreased from 51.75% at 700 m to 17.50% at 1,200 m (Figure 2). The phosphorous resorption efficiencies at 700 m and 1,000 m were significantly higher than at other altitudes. The potassium resorption efficiency at 1200 m was the highest, and the K use efficiency at 800 m was also significantly higher compared to the other altitudes. NRE showed had a significant negative correlation with altitude (p < 0.05; Figure 3). Conversely, PRE and KRE did not show a significant relationship with altitude.
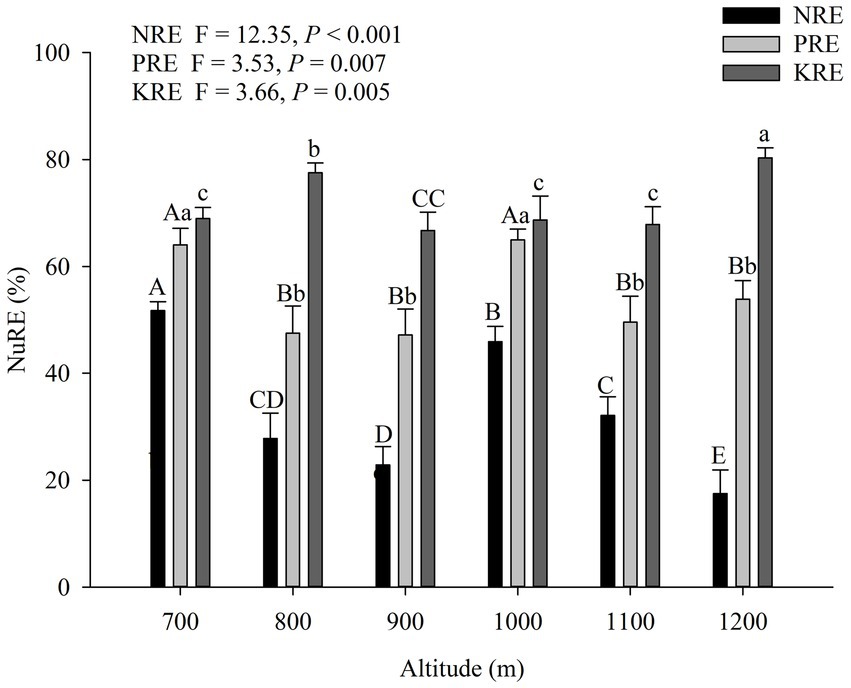
Figure 2. The mean (±SE) nutrient resorption efficiencies (NuRE) in Wuyi Mountain along an altitudinal gradient. Different letters indicate significant differences among different altitudes for the same index at the 0.05 level (p < 0.05).
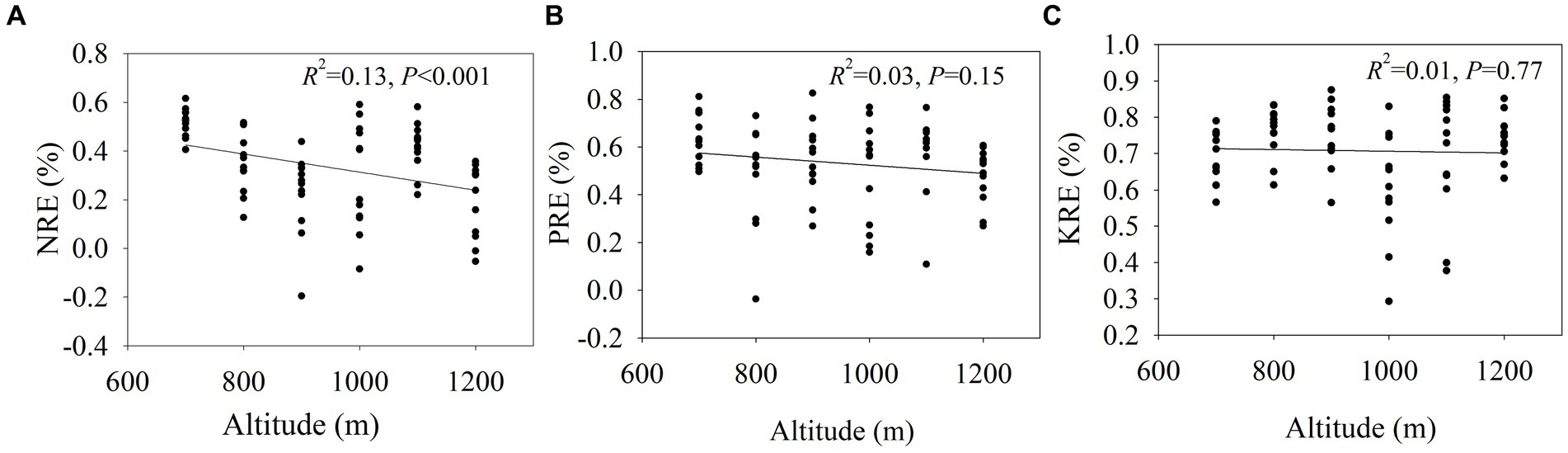
Figure 3. The linear regression between nutrient resorption efficiencies (NuRE) and altitude in the Wuyi Mountain.
In Wuyi Mountain, green leaves at altitudes ranging from 700 m to 1,200 m exhibited stoichiometric homeostasis (Table 2). The homeostasis coefficients of green leaves to soil P, K, C:N, C:K, and N:K were 3.92, 1.90, 2.65, 3.17, and 3.13, respectively. These coefficients determined as “weak homeostatic” with except of soil K for weak plastic. Other indexes did not demonstrate homeostatic characteristics.
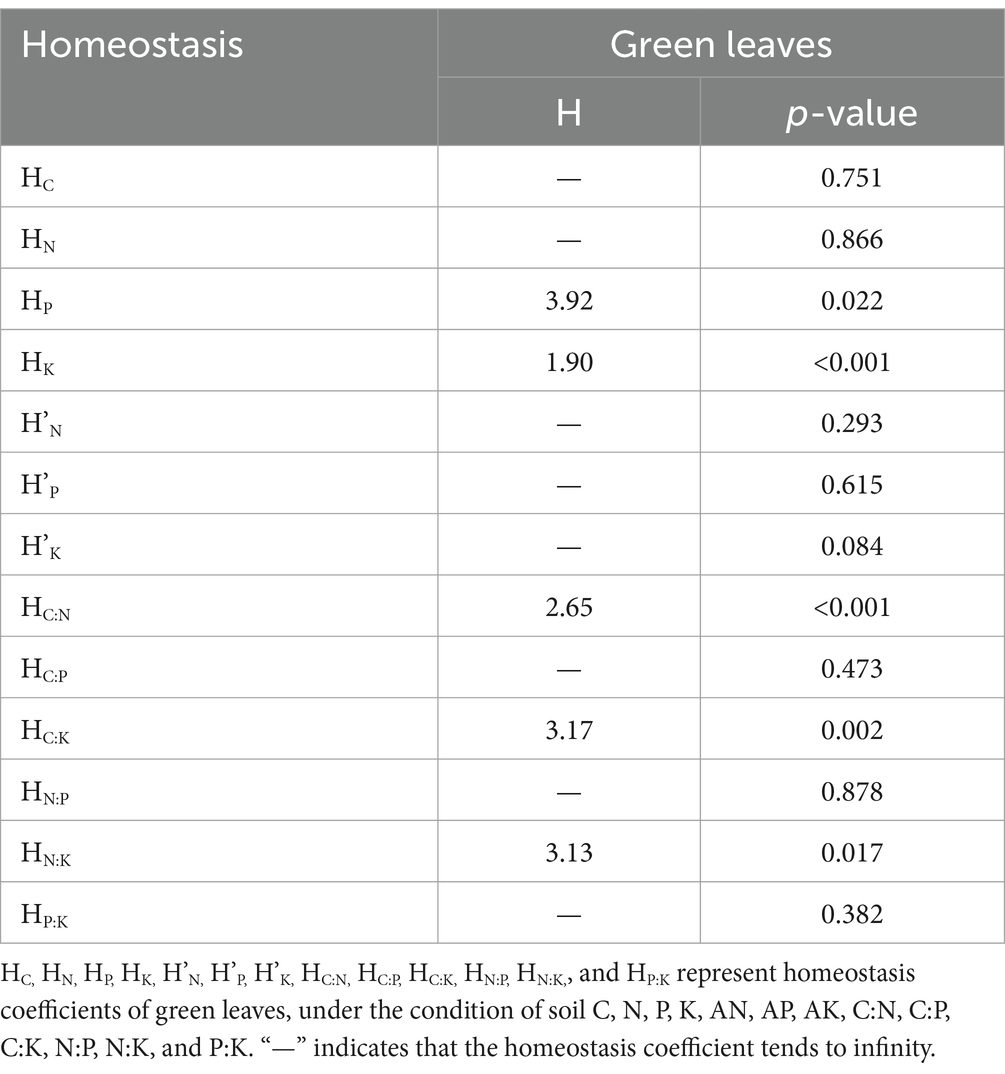
Table 2. Homeostasis coefficients of green leaves in Wuyi mountain.
3.3 Relationships between nutrient contents, stochiometric characteristics, and nutrient resorption efficiencies
With the exception of fresh litter N and P, N and K, there were significant positive relationships among N, P, and K of the green leaves and fresh litter (p < 0.01) (Figure 4). In the soil component, soil C, N, and P were significantly positively correlated (p < 0.01 or p < 0.05), while soil K had significant negative relationships with soil C, soil N, and soil P. Additionally, there were significant positive relationships between soil AN, AP, and AK (p < 0.01 or p < 0.05), and they also significantly correlated with soil N. Soil K exhibited a positive correlation with green leaves N, P, and K, but a negative correlation with fresh litter N.
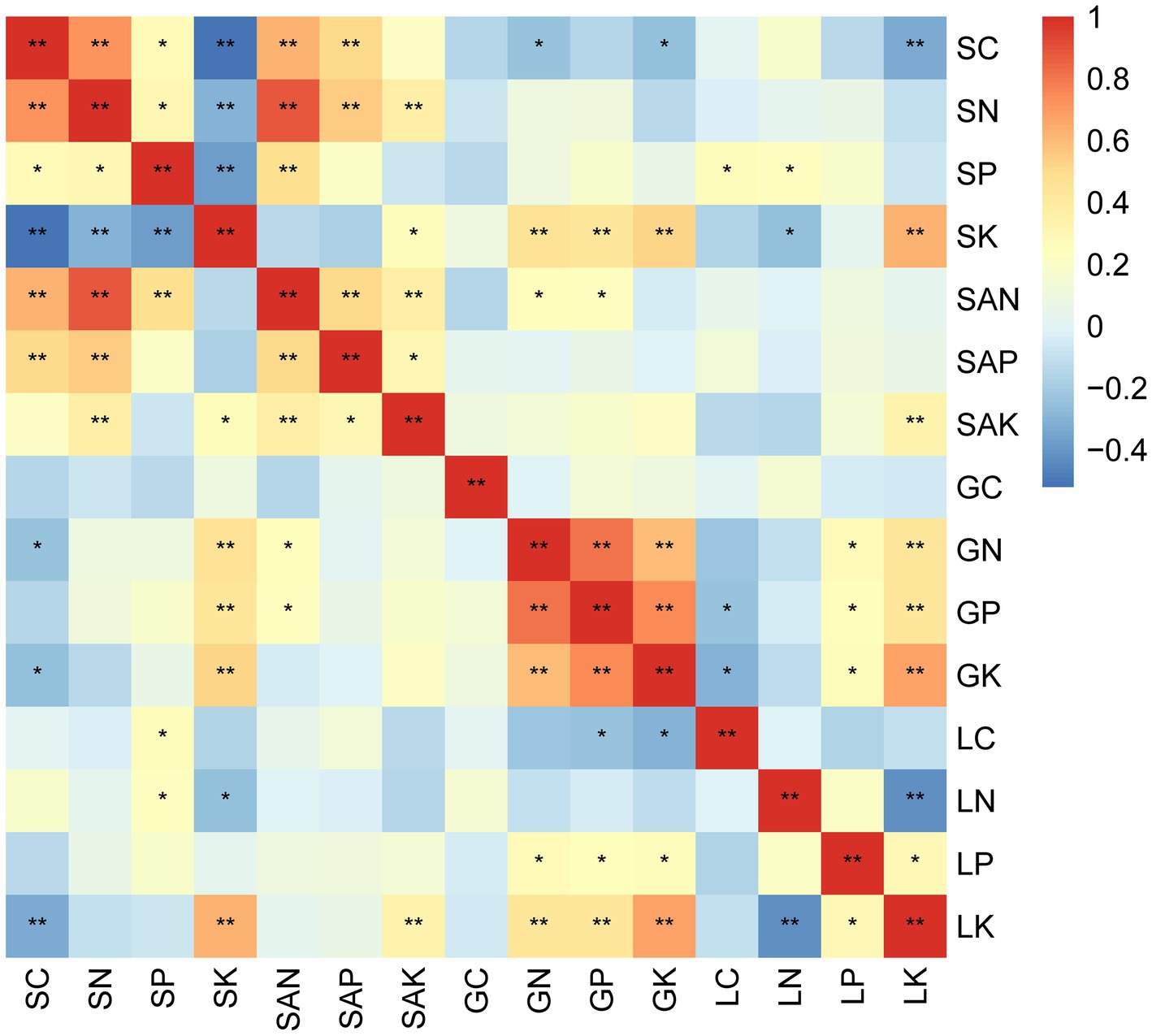
Figure 4. The heatmap of Pearson’s correlation coefficients of stoichiometric characteristics among “plant leaf-litter-soil” in Wuyi Mountain. G, L, and S indicate green leaves, fresh litter, and soil, respectively. SAN, SAP, and SAK represent available nitrogen, available phosphorus, and available potassium in the soil, respectively. * and ** indicate a significant correlation at the 0.05 (p < 0.05) and 0.01 (p < 0.01) level.
The first and second axis of RDA explained 31.09 and 2.99% of total variation in the leaf nutrient resorption efficiencies, respectively (Figure 5). The RDA analysis revealed significant correlations between leaf nutrient resorption efficiencies and various factors, including soil available N (F = 4.55, p = 0.017), contents of P (F = 5.89, p = 0.011), and K (F = 7.50, p = 0.006) in the fresh litter. Notably, these effects were more pronounced at low altitudes than at high altitudes. Furthermore, the analysis showed significant relationships between specific nutrients and nutrient resorption efficiencies. Moreover, the N, P, and K contents in the green leaves, as well as P and K in the fresh litter, were found to have significant and positive effects on NRE, while soil AK was also significantly and positively correlated with NRE (Figure 6). The P and K contents in the green leaves, as well as K in the fresh litter, had significantly positive effects on PRE (p < 0.01). Moreover, nutrient contents in the green leaves and fresh litter components had no influence on KRE.
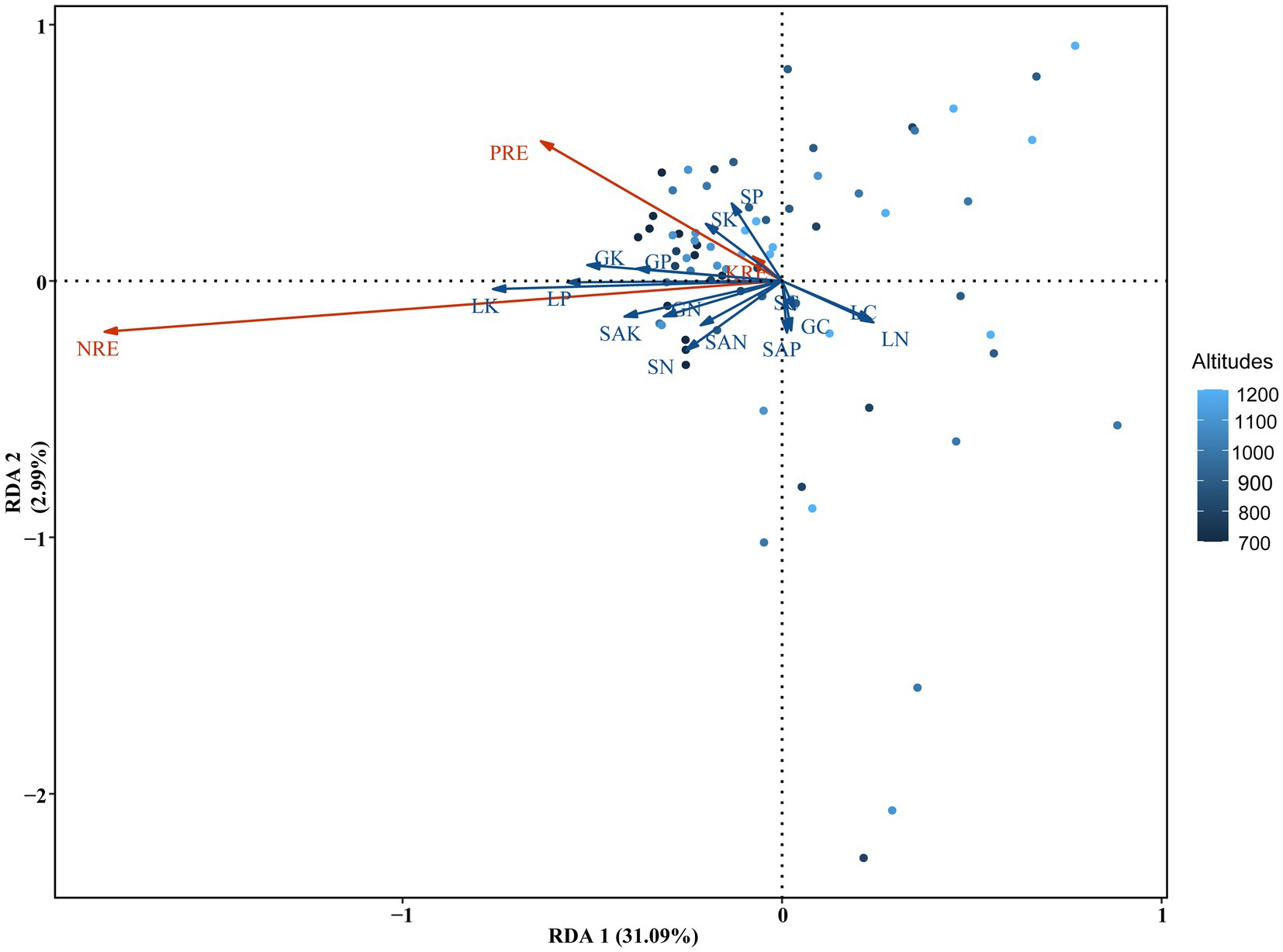
Figure 5. Redundancy analysis (RDA) of NuRE, soil nutrients contents, green leaves nutrient contents, and fresh litter nutrients contents in evergreen broadleaf forests along the altitudes in Wuyi Mountain. G, L, and S indicate green leaves, fresh litter, and soil, respectively. SAN, SAP, and SAK represent available nitrogen, available phosphorus, and available potassium in the soil, respectively.

Figure 6. The heatmap of Pearson’s correlation coefficients between nutrient resorption efficiencies and nutrient contents in the different components in Wuyi Mountain. G, L, and S indicate green leaves, fresh litter, and soil, respectively. SAN, SAP, and SAK represent available nitrogen, available phosphorus, and available potassium in the soil, respectively. * and ** indicate a significant correlation at the 0.05 (p < 0.05) and 0.01 (p < 0.01) level.
4 Discussion
4.1 Variations of nutrient stoichiometric characteristics along the altitudinal gradient
The C:N:P ratios in green leaf, fresh litter, surface soil (0–20 cm), and sub-soil (20–40 cm) of the evergreen broadleaf forest was 2,356:64:1, 4,556:98:1, 74:24:1, and 72:21:1, respectively. These ratios were higher than those of nine natural forest ecosystems from cold-temperature to tropical forests in China (Zhang et al., 2018).
The higher C content of the evergreen broadleaf forests in Wuyi Mountain that of plants globally (Elser et al., 2000a) suggested that green leaves of the evergreen broadleaf forests possessed a high C capacity. Interestingly, we also observed that a decreasing trend in leaf C content with increasing altitude. This finding contradicted the results reported by Yu et al. (2014). The inconsistence could be explained by the low temperature at high altitudes, limited soil microbial activity, and the slow turnover of ecosystem C. Therefore, the plant has low leaf C content at high altitudes. At low altitudes, accumulation of humus leads to an increasing of leaf C content. Our finding did not support the physiological acclimation mechanism (Weih and Karlsson, 2001). Plants at higher altitudes experienced higher environmental stress, leading to lower ability to produce non-structural carbohydrate.
In this study, we found that the highest leaf N content was observed at the lowest latitude, which could be attributed to lower decomposition at high altitude (Hobbie et al., 2000). This finding was in agreement with a previous report (Liu et al., 2021). The mean N and P contents of green leaves of plants ranging from 700 m to 1,200 m in Wuyi Mountain were 15.55 mg/g and 0.54 mg/g, respectively, which were lower than the global (N: 20.1 mg/g, P: 1.99 mg/g) and China’s flora (N: 20.2 mg/g, P: 1.5 mg/g) (Elser et al., 2000a; Han et al., 2005). Overall, leaf N and P showed a decreasing trend with altitude, supporting previous studies (Müller et al., 2017; Wang et al., 2018). These results indicated the direct influence of different climate conditions on plant photosynthesis, especially at low altitudes where leaf N in the evergreen broadleaf forests could increase (Forrester et al., 2019). Additionally, the N:P ratio in the green leaves was extremely high, much higher than 16. As leaf N:P > 16, the plants are P-limited (Koerselman and Meuleman, 1996), implying intense P limitation in the evergreen broadleaf forests in Wuyi Mountain due to P deficient in the soil. This finding confirmed what had been observed in tropical region (Mo et al., 2019; van Breugel et al., 2019). Interesting, P limitation was most pronounced at 900 m, where distribute Quercus phillyraeoides with shade-tolerance. This result differed from a previous report that suggested N limitation for Q. phillyraeoides (Xie et al., 2017), due to the inclusion of different land use types in the studies.
The leaf K content of the evergreen broadleaf forests in this study was lower than the average of 78 typical ecosystems in China (18.30 mg/g) (Li et al., 2021). We observed higher leaf K content at lower altitudes, this finding was not supported by a previous study that reported increased K uptake for plant growth in cold temperature condition (Zorb et al., 2014). This further confirmed that environmental changes, especially different climate conditions, could affect leaf traits (Richards and Damschen, 2022). Leaf C:N and C:P showed bimodal responses with altitude, contrasting the typical pattern of increasing leaf C:N and C:P with altitude (Müller et al., 2017). In the evergreen broadleaf forests of Wuyi Mountain, the leaf C:N ratio along the altitudinal gradient was higher than the global average for terrestrial plants (Elser et al., 2000b). Similarly, leaf C:P at different altitudes was higher than both the global and China flora averages (Elser et al., 2000b; Tang et al., 2018).
Compared to green leaves, the nutrient contents of fresh litter showed a similar trend with altitude. The C content of fresh litter in this study was higher than that of coniferous and broadleaved forests in China (He et al., 2013; Bai et al., 2019). In Wuyi Mountain, fresh litter C:N was higher at higher altitudes, suggesting lower decomposition due to colder temperature at those altitudes (Enríquez et al., 1993). Litter C:P and N:P ratios also serve as indicators of decomposition, with negative relationships between these ratios and decomposition rates (Shen et al., 2021). Our study found that plants at higher altitudes had higher fresh litter C:P and N:P ratios, implying potential lower decomposition rate in these region. Previous studies have showed that litter is susceptible to P limitation when the N:P exceeds 25 (Güsewell and Verhoeven, 2006). In our study, fresh litter N:P was higher than 25, implying P deficiency in litter decomposition within the evergreen broadleaf forests. Additionally, when the soil experienced P limitation at different altitudes, plants responded by increasing N contents. Moreover, both green leaves and litter exhibited higher C:P and C:K ratios. These findings suggest that plants adopt this strategy to offset P deficiency for plant growth.
According to classification standard for the second national soil survey in China, the soil in Wuyi Mountain displayed a low SOC (< 20 mg/g) at the different altitudes, but had high N (> 2 mg/g) levels. The soil P content indicated a deficiency, especially at 900 m. Interestingly, our study showed that soil N and P contents decreased with altitude, supporting for the results observed by Kumar et al. (2021). Furthermore, the soil C:N ratio was higher at higher altitudes, corroborating a previous report (Bing et al., 2015). The ratios of C:N and C:P in the soil were lower than those of global (C:N 14.3, C:P 186) and China’s soil averages (C:N 11.9, C:P 61) (Cleveland and Liptzin, 2007; Tian et al., 2009). However, the soil N:P ratios were higher than those observed in China’s soil (Tian et al., 2009). Different altitudes have different soil stoichiometry, this might be affected by different vegetation types (Su et al., 2019). These findings supported our first hypothesis.
4.2 Effects of altitude on nutrient resorption efficiencies
The resorption efficiency of N in evergreen broadleaf forests in Wuyi Mountain significantly decreased with altitude, while P and K resorption efficiencies did not show obvious changes. These variations in nutrient absorption were attributed to the varying biological and metabolic capacities of different species at different altitudes (Wang et al., 2022; Sun et al., 2023). Those results partially supported our second hypothesis. In addition, NRE in this study was lower than that of the global plants (50.2%) (Vergutz et al., 2012; Chen et al., 2021b), which was consistent with previous studies (Zhu et al., 2022). Similarly, PRE was lower than that the global plant (58.4%) except at 700 m and 1,000 m. In this study, KRE was found to be higher than 56.1%. However, this value was lower than that of natural forests in this region (Zhu et al., 2022). This suggested that the evergreen broadleaf forests in Wuyi Mountain had an efficient nutrient absorption capacity from the soil, even from the senesced leaves. This adaption improved the plant growth by alleviating nutrient deficiency. According to the relative resorption hypothesis, the relative nutrient resorption efficiency (NRE-PRE) provided insight into potential nutrient limitation (Han et al., 2013). The findings in Wuyi Mountain also indicated a P-limitation, as the relative efficiency was less than 0. This further implied that the plants resorb more P from senesced leaves than N (Müller et al., 2017). Additionally, when litter N content exceeded 10 mg/g or P surpassed 0.8 mg/g, suggesting incompletely absorbed of litter N and P. Conversely, complete absorption of litter N and P occurred when litter N content and P content was less than 7 mg/g and 0.5 mg/g, respectively (Killingbeck, 1996; Yu et al., 2010). Furthermore, there were significant correlations between fresh litter N:P and N and P contents of the green leaves. These findings demonstrated the direct impact of litter stoichiometry on the nutrient limitation of evergreen broadleaf forests at different altitudes.
The positive relationships between nutrient resorption efficiencies and the nutrient contents of green leaves and fresh litter were observed, consistent with the findings of Zhu et al. (2022). However, this finding contradicted the results reported by Tong et al. (2021), suggesting that variations in environmental conditions lead to different results. Furthermore, these findings implied that higher levels of leaf nutrients could potentially enhance nutrient resorption. Moreover, this study revealed a pattern where lower altitudes associated with higher C and N contents in green leaves, but lower fresh litter C and N contents. This results also indicated heightened decomposition rates at lower altitudes may result in increased leaf resorption efficiencies.
In this study, positive correlations were observed between NRE, PRE, and the levels of P and K in green leaves. Furthermore, positive relationships were found between NRE, PRE, and K in the fresh litter. These findings suggested that trees employed different nutrient strategies in response to low temperature, high solar radiation, and a humid environment (Reich and Oleksyn, 2004). Overall, altitude was found to have significant effects on leaf nutrient resorption efficiencies and nutrient limitation in the forests.
4.3 Stoichiometric homoeostasis and allocation of nutrition
Stoichiometric homoeostasis could reflect an organism’s physiological adjustments in response to fluctuating environments (Sterner and Elser, 2002; Yu et al., 2010). This study aimed to investigate the maintenance of homeostasis in green leaves concerning variations in soil P, K, C:N, C:K, and N:K. Those observations suggested that the plants at different altitudes were able to maintain relative stable levels of P and K under changing environment, as previous studies have confirmed (Sterner and Elser, 2002). Our findings highlighted the contribution of homeostasis in the evergreen broadleaf forests to nutrient balance in harsh environments (Blouin et al., 2012).
In Wuyi Mountain, higher nutrient contents were allocated to the green leaves of the evergreen broadleaf forests compared to other components. This can be attributed to the fact that green leaves are the most active organ, requiring higher nutrients levels to support essential physiological activities such as photosynthesis, respiration, and metabolic activity (Zhu et al., 2020; Li et al., 2022). Furthermore, our investigation revealed that nutrient contents in the fresh litter were higher than that those in the soil, suggesting a slow return of nutrients from the fresh litter to the soil. This finding further affirms that lower decomposition at higher altitudes delays the nutrient flow process. In summary, our results provided evidence that different climate conditions directly impacts the nutrient status of forest ecosystems (Kumar et al., 2021).
In term of C, there were no significant coupling relationship for the contents among the components of green leaves, fresh litter, and soil. However, soil available N and available K were significantly correlated with green leaves and fresh litter, which is consistent with previous studies (Liu and Wang, 2021; Li et al., 2022). While there were no significant relationships among N contents in different components, the N content in the green leaves was higher than that in the fresh litter and soil, as evidenced by the higher NRE compared to CRE at different altitudes. As for the P nutrient, there were significant coupling relationships between the contents in the green leaves and fresh litter, indicating that nutrients levels in green leaves were directly influenced by the litter nutrient status during the P cycling (Wang and Zheng, 2020). Similarly, there were positive relationships for K content among the components of green leaves, fresh litter, and soil, suggesting that K is transported from green leaves to fresh litter and then stored in soil. Furthermore, C was fixed in green leaves and litter in this study, while N was absorbed into the soil through litter decomposition at different altitudes. P and K were stored in green leaves and soil, respectively. Interestingly, trees at lower altitudes utilized less K in order to obtain more N and P for metabolic activities, as compared to those at higher altitudes. These findings supported the third hypothesis.
5 Conclusion
Based on a field investigation conducted at six altitudes of Wuyi Mountain, this research provides a comprehensive evaluation of the stoichiometric characteristics in the plant green leaves, fresh litter, and soil along the altitudinal gradient. The research revealed significant variations in nutrient content and stoichiometry among different components. The latitude had a significant impact on NRE, resulting in the vulnerability of N content in the green leaves and fresh litter at different altitudes. While PRE and KRE showed no significant relationship with altitude, higher altitudes exhibited increased P and K levels in green leaves and fresh litter compared to lower altitudes, suggesting their weak plasticity. Furthermore, the study observed low nutrient resorption efficiencies at higher altitudes, implying a delay in element cycling due to cold climate hinders nutrient flow. The study also found that nutrient levels in green leaves and fresh litter directly regulated nutrient resorption efficiencies. Additionally, the study identifies severe P deficiency of the evergreen broadleaf forest in Wuyi Mountain. The significant coupling relationships between P and K contents in green leaves, fresh litter, and soil, implying that P and K cycling occur simultaneously in the leaf-litter-soil continuum. These findings are of great significance for understanding the mechanism of nutrient cycling and plant adaptation strategies in response to different climate conditions.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding authors.
Author contributions
DZ: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. YZ: Data curation, Funding acquisition, Investigation, Writing – review & editing. SP: Formal analysis, Investigation, Writing – review & editing. WH: Data curation, Funding acquisition, Investigation, Writing – review & editing. BL: Data curation, Investigation, Writing – review & editing. DH: Conceptualization, Funding acquisition, Methodology, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by the Natural Science Foundation of Fujian Province, China (grant number 2021 J05248 and 2021 J011141), Forestry Science and Technology Promotion Project of Fujian Province, China (grant number 2022FKJ08), and Science Foundation Project for Talented New Faculty at Wuyi University (grant number YJ202103). Dafeng Hui was supported by the US NSF and USDA projects.
Acknowledgments
We thank Fan Yang, Qiaolan Song, Liuxian He, Peiying Li, Wenjun Lin, Linya Huang, Fan Chen, Shilong Li, Jianfei Liao, and other people for their assistance in the field and laboratory.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2024.1331623/full#supplementary-material
References
Bai, X., Wang, B., An, S., Zeng, Q., and Zhang, H. (2019). Response of forest species to C:N:P in the plant-litter-soil system and stoichiometric homeostasis of plant tissues during afforestation on the loess plateau, China. Catena 183, 104186–104194. doi: 10.1016/j.catena.2019.104186
Beniston, M. (2003). Climatic change in mountain rRegions: a review of possible impacts. Clim. Chang. 59, 5–31. doi: 10.1007/978-94-015-1252-7_2
Bing, H., Wu, Y., Zhou, J., Sun, H., Luo, J., Wang, J., et al. (2015). Stoichiometric variation of carbon, nitrogen, and phosphorus in soils and its implication for nutrient limitation in alpine ecosystem of eastern Tibetan plateau. J. Soil Sediment 16, 405–416. doi: 10.1007/s11368-015-1200-9
Blouin, M., Mathieu, J., and Leadley, P. W. (2012). Plant homeostasis, growth and development in natural and artificial soils. Ecol. Complex. 9, 10–15. doi: 10.1016/j.ecocom.2011.11.001
Chen, B., Chen, L., Jiang, L., Zhu, J., Chen, J., Huang, Q., et al. (2022). C:N:P stoichiometry of plant, litter and soil along an elevational gradient in subtropical forests of China. Forests 13:372. doi: 10.3390/f13030372
Chen, H., Reed, S. C., Lu, X., Xiao, K., Wang, K., and Li, D. (2021a). Coexistence of multiple leaf nutrient resorption strategies in a single ecosystem. Sci. Total Environ. 772:144951. doi: 10.1016/j.scitotenv.2021.144951
Chen, H., Reed, S. C., Lü, X., Xiao, K., Wang, K., and Li, D. (2021b). Global resorption efficiencies of trace elements in leaves of terrestrial plants. Funct. Ecol. 35, 1596–1602. doi: 10.1111/1365-2435.13809
Cleveland, C. C., and Liptzin, D. (2007). C:N:P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 85, 235–252. doi: 10.1007/s10533-007-9132-0
De Groote, S. R. E., Vanhellemont, M., Baeten, L., De Schrijver, A., Martel, A., Bonte, D., et al. (2018). Tree species diversity indirectly affects nutrient cycling through the shrub layer and its high-quality litter. Plant Soil 427, 335–350. doi: 10.1007/s11104-018-3654-1
Demars, B. O. L., and Edwards, A. C. (2007). Tissue nutrient concentrations in freshwater aquatic macrophytes: high inter-taxon differences and low phenotypic response to nutrient supply. Freshw. Biol. 52, 2073–2086. doi: 10.1111/j.1365-2427.2007.01817.x
Du, B., Ji, H., Peng, C., Liu, X., and Liu, C. (2016). Altitudinal patterns of leaf stoichiometry and nutrient resorption in Quercus variabilis in the Baotianman Mountains, China. Plant Soil 413, 193–202. doi: 10.1007/s11104-016-3093-9
Elser, J. J., Fagan, W. F., Denno, R. F., Dobberfuhl, D. R., Folarin, A., Huberty, A., et al. (2000a). Nutritional constraints in terrestrial and freshwater food webs. Nature 408, 578–580. doi: 10.1038/35046058
Elser, J. J., Sterner, R. W., Gorokhova, E. A., Fagan, W. F., Markow, T. A., Cotner, J. B., et al. (2000b). Biological stoichiometry from genes to ecosystems. Ecol. Lett. 3, 540–550. doi: 10.1046/j.1461-0248.2000.00185.x
Enríquez, S., Duarte, C. M., and Sand-Jensen, K. (1993). Patterns in decomposition rates among photosynthetic organisms: the importance of detritus C: N: P content. Oecologia 94, 457–471. doi: 10.1007/BF00566960
Forrester, D. I., Rodenfels, P., Haase, J., Härdtle, W., Leppert, K. N., Niklaus, P. A., et al. (2019). Tree-species interactions increase light absorption and growth in Chinese subtropical mixed-species plantations. Oecologia 191, 421–432. doi: 10.1007/s00442-019-04495-w
Güsewell, S., and Verhoeven, J. T. (2006). Litter N: P ratios indicate whether N or P limits the decomposability of graminoid leaf litter. Plant Soil 287, 131–143. doi: 10.1007/s11104-006-9050-2
Han, W., Fang, J., Guo, D., and Zhang, Y. (2005). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 168, 377–385. doi: 10.1111/j.1469-8137.2005.01530.x
Han, W., Tang, L., Chen, Y., and Fang, J. (2013). Relationship between the relative limitation and resorption efficiency of nitrogen vs phosphorus in woody plants. PLoS One 8:e83366. doi: 10.1371/journal.pone.0083366
He, X., Chu, C., Yang, Y., Shu, Z., Li, B., and Hou, E. (2021). Bedrock and climate jointly control the phosphorus status of subtropical forests along two elevational gradients. Catena 206:105525. doi: 10.1016/j.catena.2021.105525
He, P., Lu, J., Ren, Y., Li, J., Hou, L., Deng, X., et al. (2023). Altitude and slope aspects as the key factors affecting the change of C: N: P stoichiometry in the leaf-litter-soil system of alpine timberline. Geroderma Reg. 32:e00602. doi: 10.1016/j.geodrs.2022.e00602
He, Y., Qin, L., Li, Z., Liang, X., Shao, M., and Tan, L. (2013). Carbon storage capacity of monoculture and mixed-species plantations in subtropical China. Forest Ecol. Manag. 295, 193–198. doi: 10.1016/j.foreco.2013.01.020
Hessen, D. O., Ågren, G. I., Anderson, T. R., Elser, J. J., and de Ruiter, P. C. (2004). Carbon sequestration in ecosystems: the role of stoichiometry. Ecology 85, 1179–1192. doi: 10.1890/02-0251
Hobbie, S., Schimel, J., Trumbore, S., and Randerson, J. (2000). Controls over carbon storage and turnover in high-latitude soils. Glob. Chang. Biol. 6, 196–210. doi: 10.1046/j.1365-2486.2000.06021.x
Hou, Y., He, K., Chen, Y., Zhao, J., Hu, H., and Zhu, B. (2019). Changes of soil organic matter stability along altitudinal gradients in Tibetan alpine grassland. Plant Soil 458, 21–40. doi: 10.1007/s11104-019-04351-z
Hu, X., Wu, A., Wang, F., and Chen, F. (2019). The effects of simulated acid rain on internal nutrient cycling and the ratios of Mg, Al, Ca, N, and P in tea plants of a subtropical plantation. Environ. Monit. Assess. 191:99. doi: 10.1007/s10661-019-7248-z
Huber, E., Wanek, W., Gottfried, M., Pauli, H., Schweiger, P., Arndt, S. K., et al. (2007). Shift in soil–plant nitrogen dynamics of an alpine–nival ecotone. Plant Soil 301, 65–76. doi: 10.1007/s11104-007-9422-2
Jones, J.B. (2001). Laboratory guide for conducting soil tests and plant analysis. CRC Press: New York, NY, USA.
Killingbeck, K. T. (1996). Nutrients in senesced leaves: keys to the search for potential resorption and resorption proficiency. Ecology 77, 1716–1727. doi: 10.2307/2265777
Koerselman, W., and Meuleman, A. F. (1996). The vegetation N: P ratio: a new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 33, 1441–1450. doi: 10.2307/2404783
Körner, C. (1998). A re-assessment of high elevation treeline positions and their explanation. Oecologia 115, 445–459. doi: 10.1007/s004420050540
Korner, C. (2007). The use of 'altitude' in ecological research. Trends Ecol. Evol. 22, 569–574. doi: 10.1016/j.tree.2007.09.006
Kumar, A., Kumar, M., Pandey, R., ZhiGuo, Y., and Cabral-Pinto, M. (2021). Forest soil nutrient stocks along altitudinal range of Uttarakhand Himalayas: An aid to nature based climate solutions. Catena 207:105667. doi: 10.1016/j.catena.2021.105667
Legout, A., Hansson, K., van der Heijden, G., Laclau, J. P., Mareschal, L., Nys, C., et al. (2020). Chemical fertility of forest ecosystems. Part 2: towards redefining the concept by untangling the role of the different components of biogeochemical cycling. Forest Ecol. Manag 461:117844. doi: 10.1016/j.foreco.2019.117844
Li, Y., Dong, X., Yao, W., Han, C., Sun, S., and Zhao, C. (2022). C, N, P, K stoichiometric characteristics of the “leaf-root-litter-soil” system in dryland plantations. Ecol. Indic. 143:109371. doi: 10.1016/j.ecolind.2022.109371
Li, X., He, N., Xu, L., Li, S., and Li, M. (2021). Spatial variation in leaf potassium concentrations and its role in plant adaptation strategies. Ecol. Indic. 130:108063. doi: 10.1016/j.ecolind.2021.108063
Li, J., Wu, B., Zhang, D., and Cheng, X. (2023). Elevational variation in soil phosphorus pools and controlling factors in alpine areas of Southwest China. Geoderma 431:116361. doi: 10.1016/j.geoderma.2023.116361
Lin, H., Li, Y., Bruelheide, H., Zhang, S., Ren, H., Zhang, N., et al. (2021). What drives leaf litter decomposition and the decomposer community in subtropical forests-the richness of the above-ground tree community or that of the leaf litter? Soil Biol. Biochem. 160:108314. doi: 10.1016/j.soilbio.2021.108314
Liu, R., and Wang, D. (2021). C:N:P stoichiometric characteristics and seasonal dynamics of leaf-root-litter-soil in plantations on the loess plateau. Ecol. Indic. 127:107772. doi: 10.1016/j.ecolind.2021.107772
Liu, D., Zheng, D., Xu, Y., Chen, Y., Wang, H., Wang, K., et al. (2021). Changes in the stoichiometry of Castanopsis fargesii along an elevation gradient in a Chinese subtropical forest. Peerj 9:e11553. doi: 10.7717/peerj.11553
Mganga, K. Z., Sietiö, O. M., Meyer, N., Poeplau, C., Adamczyk, S., Biasi, C., et al. (2022). Microbial carbon use efficiency along an altitudinal gradient. Soil Bio. Biochem. 173:108799. doi: 10.1016/j.soilbio.2022.108799
Milla, R., Castro-Díez, P., Maestro-Martínez, M., and Montserrat-Martí, G. (2005). Does the gradualness of leaf shedding govern nutrient resorption from senescing leaves in Mediterranean woody plants? Plant Soil 278, 303–313. doi: 10.1007/s11104-005-8770-z
Mo, Q., Li, Z., Sayer, E. J., Lambers, H., Li, Y., Zou, B. I., et al. (2019). Foliar phosphorus fractions reveal how tropical plants maintain photosynthetic rates despite low soil phosphorus availability. Funct. Ecol. 33, 503–513. doi: 10.1111/1365-2435.13252
Müller, M., Oelmann, Y., Schickhoff, U., Böhner, J., and Scholten, T. (2017). Himalayan treeline soil and foliar C:N:P stoichiometry indicate nutrient shortage with elevation. Geoderma 291, 21–32. doi: 10.1016/j.geoderma.2016.12.015
Nie, K., Xu, M., and Zhang, J. (2023). Changes in soil carbon, nitrogen, and phosphorus in Pinus massoniana forest along altitudinal gradients of subtropical karst mountains. Peerj 11:e15198. doi: 10.7717/peerj.15198
Persson, J., Fink, P., Goto, A., Hood, J. M., Jonas, J., and Kato, S. (2010). To be or not to be what you eat: regulation of stoichiometric homeostasis among autotrophs and heterotrophs. Oikos 119, 741–751. doi: 10.1111/j.1600-0706.2009.18545.x
R Core Team (2020). R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
Reich, P. B., and Oleksyn, J. (2004). Global patterns of plant leaf N and P in relation to temperature and latitude. P. Natl. Acad. Sci. USA 101, 11001–11006. doi: 10.1073/pnas.0403588101
Richards, J. H., and Damschen, E. I. (2022). Leaf economics in a three-dimensional environment: testing leaf trait responses in vascular epiphytes to land use, climate, and tree zone. Funct. Ecol. 36, 727–738. doi: 10.1111/1365-2435.13978
Science and Technology Commission of Fujian Province, CeoWNR. (1993). Collection of scientific investigation reports of Wuyishan nature reserve. Fujian science and technology press, Fuzhou, China.
Sheikh, M. A., Kumar, M., Todaria, N. P., and Pandey, R. (2020). Biomass and soil carbon along altitudinal gradients in temperate Cedrus deodara forests in central Himalaya, India: implications for climate change mitigation. Ecol. Indic. 111:106025. doi: 10.1016/j.ecolind.2019.106025
Shen, Y., Tian, D., Hou, J., Wang, J., Zhang, R., Li, Z., et al. (2021). Forest soil acidification consistently reduces litter decomposition irrespective of nutrient availability and litter type. Funct. Ecol. 35, 2753–2762. doi: 10.1111/1365-2435.13925
Sterner, R.W., and Elser, J.J. (2002). Ecological stoichiometry: The biology of elements from molecules to the biosphere. Princeton university press, Princeton, NJ.
Su, L., Du, H., Zeng, F., Peng, W., Rizwan, M., Núñez-Delgado, A., et al. (2019). Soil and fine roots ecological stoichiometry in different vegetation restoration stages in a karst area, Southwest China. J. Environ. Manag. 252:109694. doi: 10.1016/j.jenvman.2019.109694
Sun, X., Li, D., Lü, X., Fang, Y., Ma, Z., Wang, Z., et al. (2023). Widespread controls of leaf nutrient resorption by nutrient limitation and stoichiometry. Funct. Ecol. 37, 1653–1662. doi: 10.1111/1365-2435.14318
Sun, Y., Wang, C., Chen, X., Liu, S., Lu, X., Chen, H. Y. H., et al. (2022). Phosphorus additions imbalance terrestrial ecosystem C:N:P stoichiometry. Glob. Chang. Biol. 28, 7353–7365. doi: 10.1111/gcb.16417
Tang, Z., Xu, W., Zhou, G., Bai, Y., Li, J., Tang, X., et al. (2018). Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China's terrestrial ecosystems. Proc. Natl. Acad. Sci. 115, 4033–4038. doi: 10.1073/pnas.1700295114
Tian, H., Chen, G., Zhang, C., Melillo, J. M., and Hall, C. A. S. (2009). Pattern and variation of C:N:P ratios in China’s soils: a synthesis of observational data. Biogeochemistry 98, 139–151. doi: 10.1007/s10533-009-9382-0
Tong, R., Zhou, B., Jiang, L., Ge, X., and Cao, Y. (2021). Spatial patterns of leaf carbon, nitrogen, and phosphorus stoichiometry and nutrient resorption in Chinese fir across subtropical China. Catena 201, 105221–105229. doi: 10.1016/j.catena.2021.105221
Urbina, I., Grau, O., Sardans, J., Margalef, O., Peguero, G., Asensio, D., et al. (2021). High foliar K and P resorption efficiencies in old-growth tropical forests growing on nutrient-poor soils. Ecol. Evol. 11, 8969–8982. doi: 10.1002/ece3.7734
van Breugel, M., Craven, D., Lai, H. R., Baillon, M., Turner, B. L., and Hall, J. S. (2019). Soil nutrients and dispersal limitation shape compositional variation in secondary tropical forests across multiple scales. J. Ecol. 107, 566–581. doi: 10.1111/1365-2745.13126
van Heerwaarden, L. M., Toet, S., and Aerts, R. (2003). Current measures of nutrient resorption efficiency lead to a substantial underestimation of real resorption efficiency: facts and solutions. Oikos 101, 664–669. doi: 10.1034/j.1600-0706.2003.12351.x
Vergutz, L., Manzoni, S., Porporato, A., Novais, R. F., and Jackson, R. B. (2012). Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol. Monogr. 82, 205–220. doi: 10.1890/11-0416.1
Wang, J., Hui, D., Liu, Z., Lin, Y., Wang, F., Long, J., et al. (2022). Leaf nutrient resorption differs among canopy and understory plant species in subtropical Eucalyptus and Acacia plantations. Land Degrad. Dev. 33, 1662–1676. doi: 10.1002/ldr.4254
Wang, A., Wang, X., Tognetti, R., Lei, J., Pan, H., Liu, X., et al. (2018). Elevation alters carbon and nutrient concentrations and stoichiometry in Quercus aquifolioides in southwestern China. Sci. Total Environ. 622, 1463–1475. doi: 10.1016/j.scitotenv.2017.12.070
Wang, J., Wang, J., Wang, L., Zhang, H., Guo, Z., Geoff Wang, G., et al. (2019). Does stoichiometric homeostasis differ among tree organs and with tree age? Forest Ecol. Manag. 453:117637. doi: 10.1016/j.foreco.2019.117637
Wang, K., Zhang, R., Song, L., Yan, T., and Na, E. (2021). Comparison of C:N:P stoichiometry in the plant-litter-soil system between poplar and elm plantations in the Horqin Sandy land, China. Front. Plant Sci. 12:655517. doi: 10.3389/fpls.2021.655517
Wang, Z., and Zheng, F. (2020). Ecological stoichiometry of plant leaves, litter and soils in a secondary forest on China’s Loess Plateau. PeerJ 8:e10084. doi: 10.7717/peerj.10084
Weih, M., and Karlsson, P. S. (2001). Growth response of Mountain birch to air and soil temperature: is increasing leaf-nitrogen content an acclimation to lower air temperature? New Phytol. 150, 147–155. doi: 10.1046/j.1469-8137.2001.00078.x
Xie, H., Yu, M., and Cheng, R. (2017). Effects of light intensity variation on nitrogen and phosphorus contents, allocation and limitation in five shade-enduring plants. Chin. J. Plant Ecol. 41, 559–569. doi: 10.17521/cjpe.2016.0248
Yang, Y., Liang, C., Wang, Y., Cheng, H., An, S., and Chang, S. X. (2020). Soil extracellular enzyme stoichiometry reflects the shift from P-to N-limitation of microorganisms with grassland restoration. Soil Biol. Biochem. 149:107928. doi: 10.1016/j.soilbio.2020.107928
You, W., Lin, L., Wu, L., Ji, Z., Yu, J., Zhu, J., et al. (2017). Geographical information system-based forest fire risk assessment integrating national forest inventory data and analysis of its spatiotemporal variability. Ecol. Indic. 77, 176–184. doi: 10.1016/j.ecolind.2017.01.042
Yu, Q., Chen, Q., Elser, J. J., He, N., Wu, H., Zhang, G., et al. (2010). Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability. Ecol. Lett. 13, 1390–1399. doi: 10.1111/j.1461-0248.2010.01532.x
Yu, Q., Elser, J. J., He, N., Wu, H., Chen, Q., Zhang, G., et al. (2011). Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland. Oecologia 166, 1–10. doi: 10.1007/s00442-010-1902-z
Yu, D., Wang, Q., Liu, J., Zhou, W., Qi, L., Wang, X., et al. (2014). Formation mechanisms of the alpine Erman’s birch (Betula ermanii) treeline on Changbai Mountain in Northeast China. Trees 28, 935–947. doi: 10.1007/s00468-014-1008-z
Yu, Q., Wilcox, K., Pierre, K. L., Knapp, A. K., Han, X., and Smith, M. D. (2015). Stoichiometric homeostasis predicts plant species dominance, temporal stability, and responses to global change. Ecology 96, 2328–2335. doi: 10.1890/14-1897.1
Zhang, A., Li, X., Wu, S., Li, L., Jiang, Y., Wang, R., et al. (2021). Spatial pattern of C:N:P stoichiometry characteristics of alpine grassland in the Altunshan Nature Reserve at North Qinghai-Tibet Plateau. Catena 207:105691. doi: 10.1016/j.catena.2021.105691
Zhang, J., Zhao, N., Liu, C., Yang, H., Li, M., Yu, G., et al. (2018). C:N:P stoichiometry in China's forests: from organs to ecosystems. Funct. Ecol. 32, 50–60. doi: 10.1111/1365-2435.12979
Zhang, Q., Zhou, J., Li, X., Yang, Z., Zheng, Y., Wang, J., et al. (2019). Are the combined effects of warming and drought on foliar C:N:P:K stoichiometry in a subtropical forest greater than their individual effects? Forest Ecol. Manag. 448, 256–266. doi: 10.1016/j.foreco.2019.06.021
Zhu, D., Hui, D., Wang, M., Yang, Q., and Yu, S. (2020). Light and competition alter leaf stoichiometry of introduced species and native mangrove species. Sci. Total Environ. 738:140301. doi: 10.1016/j.scitotenv.2020.140301
Zhu, D., Peng, S., Wang, J., and Hui, D. (2022). Responses of nutrient resorption to human disturbances in Phoebe bournei forests. Forests 13:905. doi: 10.3390/f13060905
Keywords: stoichiometry, altitude, nutrient resorption efficiency, nutrient cycling, evergreen broadleaf forests
Citation: Zhu D, Zhou Y, Peng S, Hua W, Luo B and Hui D (2024) Impacts of altitude on plant green leaf, fresh litter, and soil stoichiometry in subtropical forests. Front. For. Glob. Change. 7:1331623. doi: 10.3389/ffgc.2024.1331623
Edited by:
Xiankai Lu, South China Botanical Garden (CAS), ChinaCopyright © 2024 Zhu, Zhou, Peng, Hua, Luo and Hui. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dehuang Zhu, emh1ZGg1QG1haWwyLnN5c3UuZWR1LmNu; Dafeng Hui, ZGh1aUBUbnN0YXRlLmVkdQ==