
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. For. Glob. Change , 21 February 2024
Sec. Forest Disturbance
Volume 7 - 2024 | https://doi.org/10.3389/ffgc.2024.1293439
 Han Zhang1†
Han Zhang1† Xian Li1†
Xian Li1† Shengqiang Wang1Chenyang Jiang1Yuhong Cui1Rongyuan Fan1
Shengqiang Wang1Chenyang Jiang1Yuhong Cui1Rongyuan Fan1 Yahui Lan1
Yahui Lan1 Qianchun Zhang1
Qianchun Zhang1 Shaoming Ye1,2*
Shaoming Ye1,2*Introduction: Cultivation of Chinese fir (Cunninghamia lanceolata) have alleviated timber shortages and mixed stands with Chinese fir and indigenous species represent a sustainable forestry model. Studying system nutrient balance and tree nutrient homeostasis can provide insights into the ecological advantages of Chinese fir mixed stands and guide the management of plantations.
Methods: Mixed Chinese fir plantations with two native broadleaf species (Michelia macclurei and Mytilaria laosensis) and pure Chinese fir stands were examined for our study. The responses in carbon (C), nitrogen (N), and phosphorus (P) distribution and their stoichiometric characterization in the tree–litter–soil system to stand changes were evaluated. In addition, the ecological stoichiometric homeostasis of leaves, branches, trunks, bark and roots was used to measure the trees’ adaptive capacity to stand changes.
Results: The results showed that the mixed stands of Michelia macclurei and Chinese fir significantly increased soil OC, TN, and TP, and improved the carbon sequestration and nutrient storage functions of the plantations. The mixed stands improved the litter mass and C:N and C:P to different degrees. The soil N and P imbalance reduced the leaf N:P, resulting in N limitation of different trees, while the principal component analysis showed that the improvement of soil TN in the mixed plantation alleviated the N limitation. In addition, mixed stands reduced N, P, and N:P homeostasis in branch, trunk, and bark of some Chinese fir trees, whereas mixed species showed flexibility in leaf N:P homeostasis.
Discussion: Therefore, the selection of mixed species for mixed forests is a critical factor to consider when creating mixed plantations. These results contribute to our understanding of the ecological stoichiometry of fir plantations and are of considerable importance for the sustainable development of plantations as well as for the response to global climate change.
Chinese fir [Cunninghamia lanceolata (Lamb.) Hook] is a common species for afforestation in China (Zhou B. et al., 2020), and it greatly relieves the industrial demand for timber. Simultaneously, in accordance with the United Nations Sustainable Development Goals for 2023, the extensive establishment of plantation forests increases carbon storage in soils and biomass, thereby inhibiting global warming (Laganière et al., 2010; He et al., 2013); plantation forests also regulate local microclimates, mitigate land desertification, and inhibit the loss of biodiversity (Robin, 2008; Peng et al., 2014; Miao et al., 2015). Therefore, the sustainable development of planted forests is closely linked to the sustainable development of the Earth. However, the current widely practiced pure-stand model of Chinese fir cultivation has many ecological disadvantages, including reduced yield (Farooq et al., 2019), land degradation (Guan et al., 2015; Selvaraj et al., 2017), and reduced diversity of understory (Hou et al., 2021) caused by monoculture and short rotation periods. These issues significantly impact the sustainable development of Chinese fir plantations and hinder the alleviation of the triple planetary crisis of climate change, pollution and biodiversity loss. Recently, mixed Chinese fir plantations have been demonstrated to contribute understory vegetation and soil ecology and increase plantation productivity (Wang et al., 2010; Wang S.Q. et al., 2021; Feng et al., 2022) and are effective ways to achieve sustainable plantation development. Research on planted mixed plantations will not only help alleviate the energy crisis, but also make us more optimistic in dealing with global climate change.
Carbon (C), nitrogen (N), and phosphorus (P) are key factors in maintaining the stability and sustainability of forest ecosystems as they are energy sources and essential nutrients for plants and microorganisms (Fan et al., 2015). Soil and tree C serve as the main carbon sinks on land (Pugh et al., 2019), and soil N and P support plant growth (Wieder et al., 2015). The basal cycle of carbon and nutrients between plant and soil is achieved through litter return and root uptake (Bardgett et al., 2014; Handa et al., 2014). In this process, stoichiometric ratios regulate the distribution and balance of carbon and different nutrients among ecosystem components, profoundly affecting the ecosystem stability and development of plantations (Wang Z.C. et al., 2021). For example, P deficiency in trees leads to increased C:P and N:P, limiting tree growth (Zhang H. et al., 2018). Second, leaf N:P reflects the nutrient limitation of the plant communities: leaf N:P less than 14 indicates that the plant community is N-limited, N:P greater than 16 reflects P limitation, and a combination of N and P limitation is likely to be present when the N:P ratio is 14–16 (Koerselman and Meuleman, 1996). And changes in soil fertility and nutrient mineralization affect soil C:N and C:P (Gan et al., 2020; Luo et al., 2020). Meanwhile, some studies found high C:N and C:P inhibited litter decomposition (Mooshammer et al., 2012). Ecological stoichiometry also reflects the energy and nutrient linkages between ecosystem components (Zechmeister-Boltenstern et al., 2015; Wang and Zheng, 2020). It has been suggested that strong correlations between plant, litter and soil C:N:P stoichiometry explain how nutrient changes in ecological components are influenced by other components (Yang et al., 2018; Chen et al., 2022). However, the ecological advantages of mixed plantations based on tree–litter–soil system stoichiometry have not been clearly elucidated, including changes in C, N, and P distribution and balance as well as tree nutrient limitation by mixed stands.
Homeostasis is essential to the ecological stoichiometry concept, and in a tree–litter–soil system, stoichiometric homeostasis displays the potential of trees to keep the relative elemental stability in their bodies independent of soil nutrient alters (Sterner and Elser, 2002; Wang K. et al., 2021). Specifically, plants that do not vary with soil nutrients are considered to be strictly homeostatic plants, while plants that show changes in element with variations in soil elemental contents are classified as nonhomeostatic (Persson et al., 2010). Because nutrient acquisition and utilization capacities vary among tree species, they exhibit open or conservative nutrient acquisition strategies, which determine the adaptive capacity of different species (Yu et al., 2015). It is generally accepted that higher stoichiometric homeostasis helps plants adapt to more complex or difficult environments (Li et al., 2018; Wei et al., 2021), which is important for maintaining ecosystem function and stability (Yu et al., 2010). However, the nutrient requirements of plant organs vary according to their functions (Minden and Kleyer, 2014). Thus, exploring homeostasis at the organ level improves our understanding of tree response to environmental and resource changes.
Based on the above background, we measured the distribution and linkages among C, N, and P contents and their stoichiometric ratios in trees, litter layers, and soils in pure and two mixed Chinese fir stands. The differences in stoichiometric homeostasis among different stands and tree species was analyzed. Meanwhile, the fractions of trees (leaves, branches, trunks, bark and roots), litter (undecomposed leaves, partially decomposed leaves, and cladoptosis) and soil (0–20 cm and 20–40 cm) were also considered in this study. The objectives of this study were to determine (1) whether the creation of two mixed plantations could increase the elemental C, N, and P content of the tree-litter-soil system or any component; (2) whether mixed plantations resulted in an imbalance in the stoichiometry of elements C, N, and P in Chinese fir plantations; and (3) whether, at the organ level, mixed plantations contributed to stoichiometric homeostasis within the fir and mixed species. These objectives will provide a better understanding of the status of Chinese fir plantations and inform plantation management and responses to global climate change.
This study was conducted at the Tropical Forestry Experimental Center of the Chinese Academy of Forestry Sciences (106°41′–106°59′ E, 21°57′–22°16′ N) located in the west of Chongzuo City, Guangxi Zhuang Autonomous Region (Figure 1). The area has a subtropical monsoon climate. The region has annual sunshine duration of 1,218–1,620 h and average annual precipitation of 1,200–1,500 mm as well as the annual average temperature of 20.5–21.7°C, with a maximum temperature of 40.3°C and a minimum temperature of −1.3°C. Moreover, the annual accumulated temperature above 10°C is 6,000–7,600°C. The area is dominated by hills, with slopes of 20–30°. The soil parent material is mainly granite. The soil is mainly red loam and latosol with pH 4.8–5.5, with a thick topsoil layer and high fertility (Li et al., 2019). The main afforestation species in the region are Cunninghamia lanceolata and Pinus massoniana. In addition, Castanopsis hystrix, Betula alnoides, Mytilaria laosensis, Michelia macclurei, Dalbergia odorifera and other valuable broadleaf species have been recently cultivated as forestry species. The understory plants mainly include Evodia lepta, Rhodomyrtus tomentosa, Rhus chinensis, Rubus alceaefolius, Blechnum orientale, and Adiantum flabellulatum.
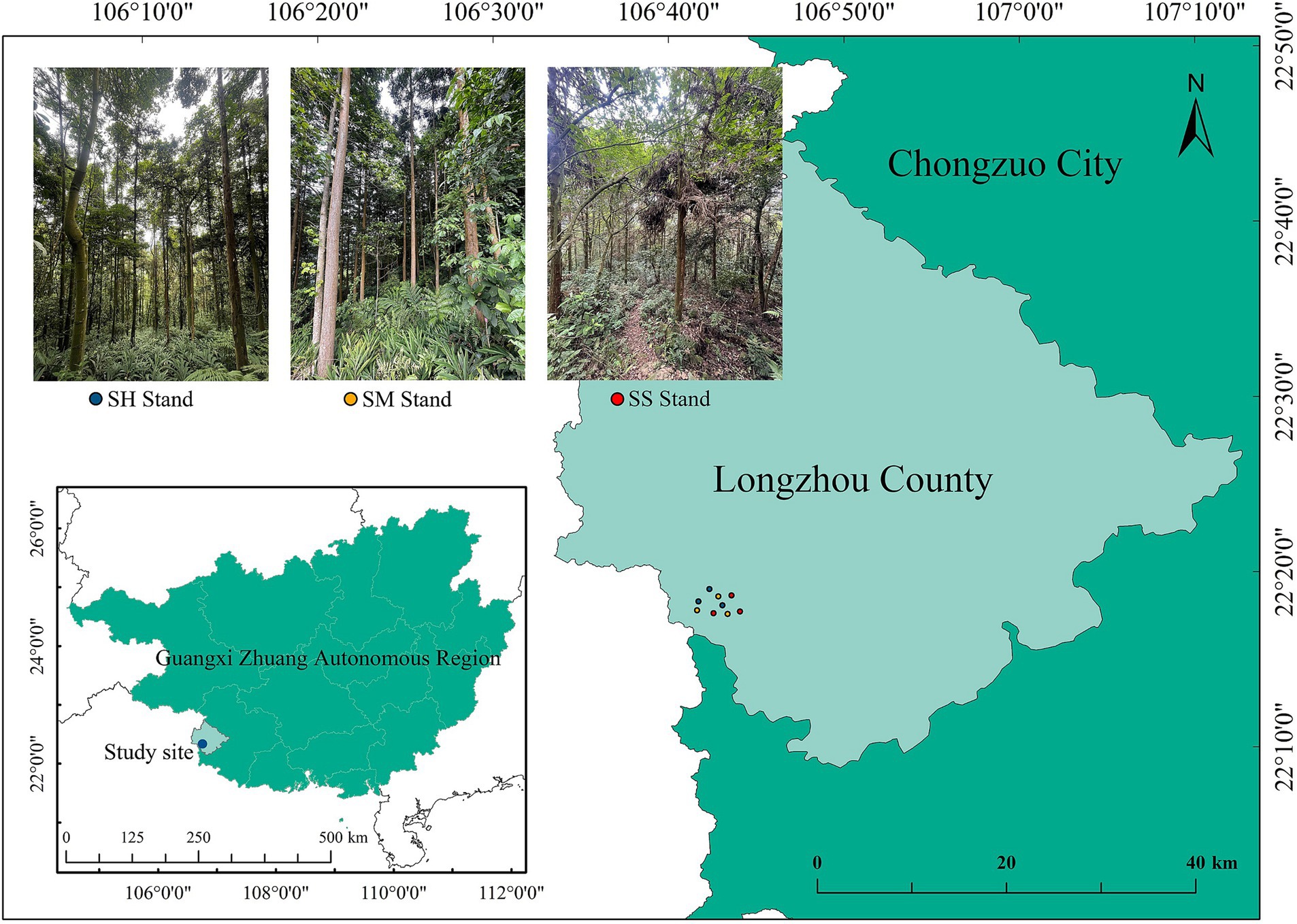
Figure 1. Location of the study area and conditions of the mixed and pure Chinese fir plantations.
The study was conducted in logging sites of pure Chinese fir plantation forests. These areas were originally pure Chinese fir plantations that had been forested, resulting in the near-complete destruction of the native vegetation due to prolonged and intense anthropogenic disturbances (He et al., 2023). The Chinese fir—Michelia macclurei mixed stands (SH stand), Chinese fir—Mytilaria laosensis mixed stands (SM stand) and pure Chinese fir stands (SS stand) selected for this study were established in March 1992 with plant spacing of 3 m × 2 m. The Chinese fir—Michelia macclurei and Chinese fir—Mytilaria laosensis mixed stands were planted in a 3:1 ratio of Chinese fir to mixed species, and the Chinese fir—Michelia macclurei mixed stands were managed to ensure tree survival at the beginning of the planting period, while the Chinese fir—Mytilaria laosensis mixed stands were cultivated to near maturity. The pure Chinese fir stands were subjected to 3 years of management to prevent pests and diseases as well as thinned (the 20% Chinese fir was cut down) in 2008; they were not subjected to any human disturbances after that. The three stands have not been affected by major pests and natural geological hazards thus far (Li Y. et al., 2021; Wang S.Q. et al., 2021). A standard quadrat (20 m × 20 m) was established in different plots (approximately 1 ha) of the Chinese fir-Michelia macclurei mixed stands, Chinese fir-Mytilaria laosensis mixed stands and pure Chinese fir stands in October 2020. Three replicates were set up in each stand, with a total of nine standard quadrats. To avoid spatial self-correlation, the plots were located at a distance of 800 m from each other and not less than 50 m from the stand edge (He et al., 2023). The basic information and soil physical and chemical properties of the standard quadrats are listed in Tables 1, 2.
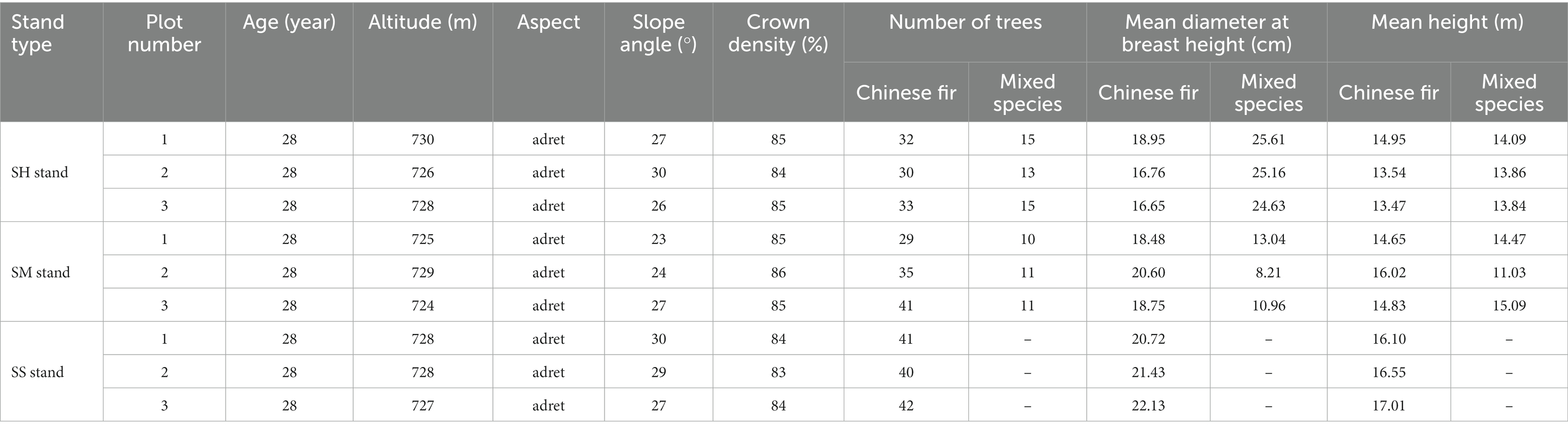
Table 1. Basic information for the plots among different Chinese fir stands.
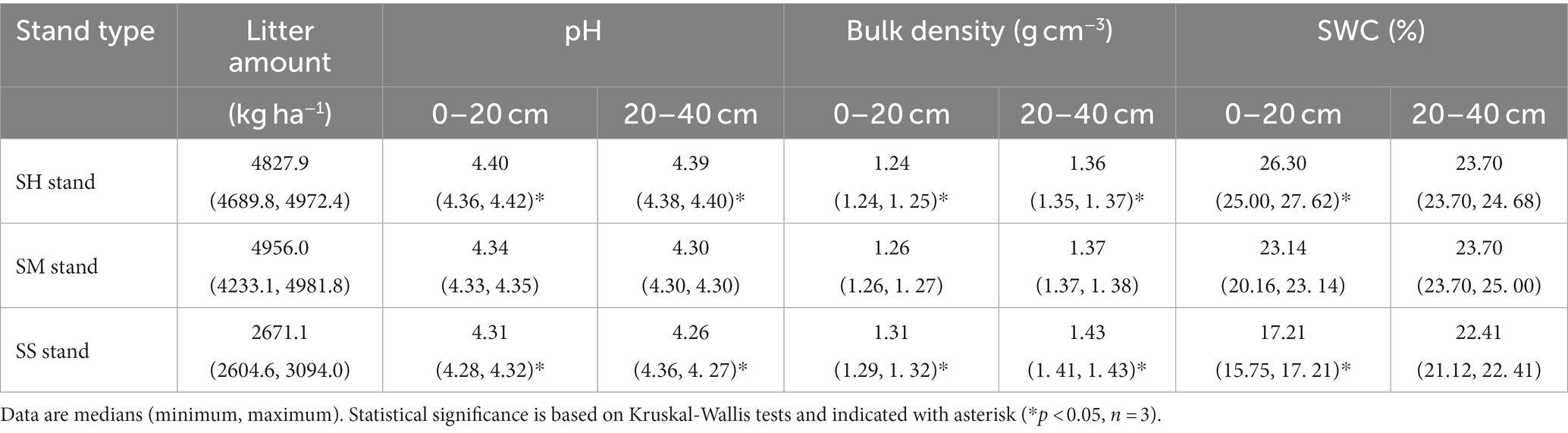
Table 2. Litter and soil characteristics among different Chinese fir plantations.
In each standard quadrat, a 5-point sampling method was used to collect 1 m2 of existing litter from the ground in a “Z” pattern, and the litter was divided into undecomposed leaves, partially decomposed leaves and cladoptosis. Soil samples were collected at the same positions as the litter samples, 40 cm soil profile was dug out with a spade and collected at depths of 0 ~ 20 cm and 20 ~ 40 cm. The soil from 5 points in each sample plot was mixed in equal mass proportions to obtain a composite soil sample for that standard sample plot. Also, the soil was sampled with a ring knife for determining the soil properties. One healthy tree with an average diameter at breast height (at 1.3 m) was selected from the three standard samples of each stand for plant sample collection (in mixed stands, one species of each was selected) for a total of three replicates. Mature leaves and branches were cut from different aspects and heights of the trees, and roots of similar diameters (<2 mm) were dug out from different orientations below the canopy. Trunk samples were collected using growth cones at different locations (top, middle, and bottom), and bark were taken at the same location. After the soil has dried naturally, stones and broken roots are removed with a 5 mm sieve. Samples of plants and litter were dried at 70°C. Finally, soil, litter and plant samples were ground and used to measure C, N, and P content.
Plant and litter samples were digested using H2SO4-H2O2, and soil samples were digested using H2SO4-HClO4. Organic carbon (OC) content was measured with reference to the potassium dichromate oxidation spectrophotometric method described by Nelson and Sommers (1996), total nitrogen (TN) content was calculated by the micro-Kjeldahl method of Bremner (1996), and total phosphorus (TP) content was determined using the molybdenum antimony anticolorimetric method according to Bo et al. (2020). A pH meter was used to calculate soil pH (water:soil = 2.5:1, v:v).
The dynamic equilibrium coefficient (H) reflects the stoichiometric dynamics of organisms and is derived from the formula of Persson et al. (2010):
where y is the N and P content (g kg−1) and N:P ratio in the leaves, branches, trunks, bark and roots of trees, x is the corresponding N and P content (g kg−1) and N:P ratio in the soil, c is a constant, and H is the equilibrium adjustment coefficient. Because the expected slope is equal to or greater than 0, a one-tailed test at α = 0.1 is used. Since strictly homeostatic organisms have a large H, we use a regression slope of 1/H in all analyses (Persson et al., 2010). When no significant level (p > 0.1) was reached in the correlation of regression, 1/H was set to 0 and the species was considered to follow strict homeostasis (internal environmental balance) (Wang K. et al., 2021; Wang S.Q. et al., 2021; Wang Z.C. et al., 2021; Bagedeng et al., 2022). The species was considered in a flexible homeostasis when the regression relationship was significant (p < 0.1), and the states can be categorized as steady state (0 < | 1/H | <0.25), weak steady state (0.25 < | 1/H | < 0.5), weak plasticity (0.5 < | 1/H | < 0.75), plastic (0.75 < | 1/H | < 1) or unsteady (| 1/H | ≥ 1).
First, the normal distribution and variance of the data were tested. Kruskal-Wallis tests were then used to analyze changes in C, N, and P among stands, organs, litter fractions, and soil layers. Principal component analysis (PCA) was used to explore the relationships among system components.
In the 0–20 cm soil, the median OC and TN contents of the SH stands were 34.65 g kg−1 and 1.46 g kg−1, significantly higher than the OC (22.93 g kg−1) and TN (1.18 g kg−1) contents of the pure fir stands (Kruskal-Wallis tests). Similarly, in the 20–40 cm soil, the SH stands had median OC, TN, and TP contents of 23.78 g kg−1, 1.22 g kg−1, and 0.43 g kg−1, respectively, which were significantly higher than the OC (8.44 g kg−1), TN (0.69 g kg−1), and TP (0.28 g kg−1) contents of the pure fir stands (Kruskal-Wallis tests). While SM stands were not dramatically different from pure Chinese fir stands (Figures 2A–C). Among the trees, the median of OC contents in branch (476.00 g kg−1), trunk (489.33 g kg−1), and root (461.37 g kg−1) of Chinese fir in the SH stand were significantly lower than those in the SM stand (branch: 569.89 g kg−1, trunk: 566.89 g kg−1 and root: 488.52 g kg−1), but none of the OC contents of Chinese fir in the two mixed stands differed significantly from those of Chinese fir in the SS stand (Figure 2D). Meanwhile, the median of leaf TN content was significantly higher (p < 0.05, Kruskal-Wallis tests) than that of the trunk for all tree species in the three stands (Figure 2E). In addition, there was a significant difference between leaf (median: 4.14 g kg−1) and trunk (median: 2.50 g kg−1) TP content of Michelia macclurei in SH stands (Figure 2F). For litter, significant differences were found only between OC content of partially decomposed leaves and TP content of cladoptosis in SM and SS stands, as well as between TN content of partially decomposed leaves in SH and SM stands (Figures 2G–I).
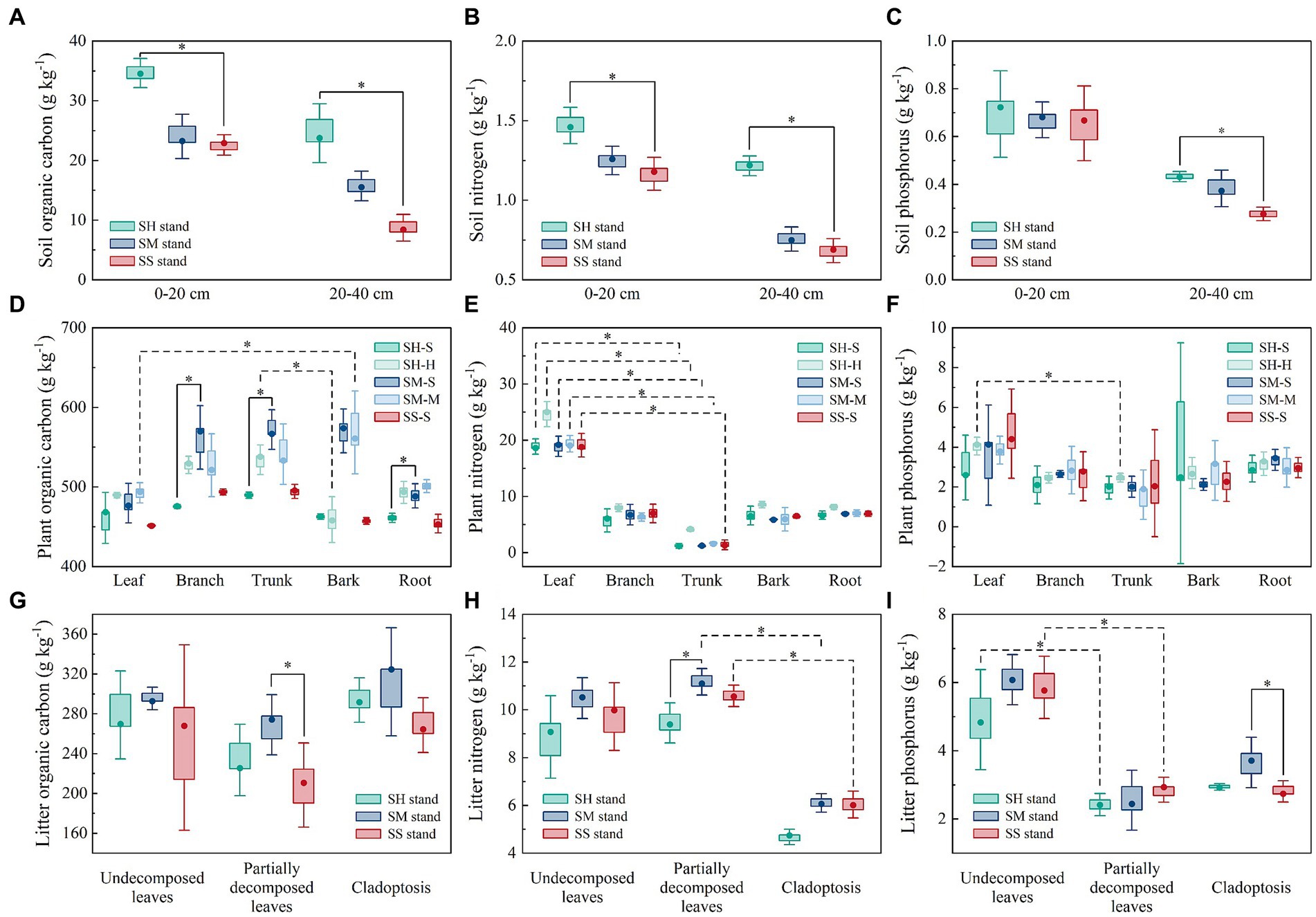
Figure 2. The C, N, and P content in soil, plant and litter of mixed and pure Chinese fir stands. Statistical significance is based on Kruskal-Wallis tests and indicated with asterisk (*). The box limits the 25–75th percentile range, the dots within the boxes are medians, the whiskers indicate maximum and minimum values. Solid black line indicate that the data are significantly different among different stand types (p < 0.05, n = 3), and black dotted line indicate that the data are significantly different among components (p < 0.05, n = 3). SH stand: the Chinese fir–Michelia macclurei mixed stands. SM stand: the Chinese fir–Mytilaria laosensis mixed stands. SS stand: pure Chinese fir stands.
The C:P of the 20–40 cm soil were significantly higher in the SH stand (median: 55.94) than in the SS stand (median: 31.84) (Figure 3B). In the 20–40 cm soil, there is a significant difference in N:P between the SH stands (median: 2.80) and the SM stands (median: 2.08) (Figure 3B). The medians of leaf C:N of all tree species in the three stands was significantly lower than while N:P was significantly higher than their trunk. Meanwhile, the median of leaf C:p (118.73) of Michelia macclurei in SH stand was significantly lower than that of branch (217.36) and trunk (226.85), as well as leaf C:p (136.83) of Mytilaria laosensis in SH stand was significantly lower than that of trunk (514.13) (Figures 3D–F). The median of Cladoptosis C:N was significantly higher in SH stands while C:P was significantly lower in SS stands. Partially decomposed leaf fractions of all three stand litter had significantly lower C:N medians than cladoptosis (Figure 3G). Undecomposed leaf C:P was significantly lower than cladoptosis in both SH and SS stands, whereas partially decomposed leaf C:P was significantly lower than partially decomposed leaf in the SM stand (Figure 3G). In addition, the median of undecomposed leaf N:P was significantly lower than that of partially decomposed leaves in the SS stand (Figure 3I).
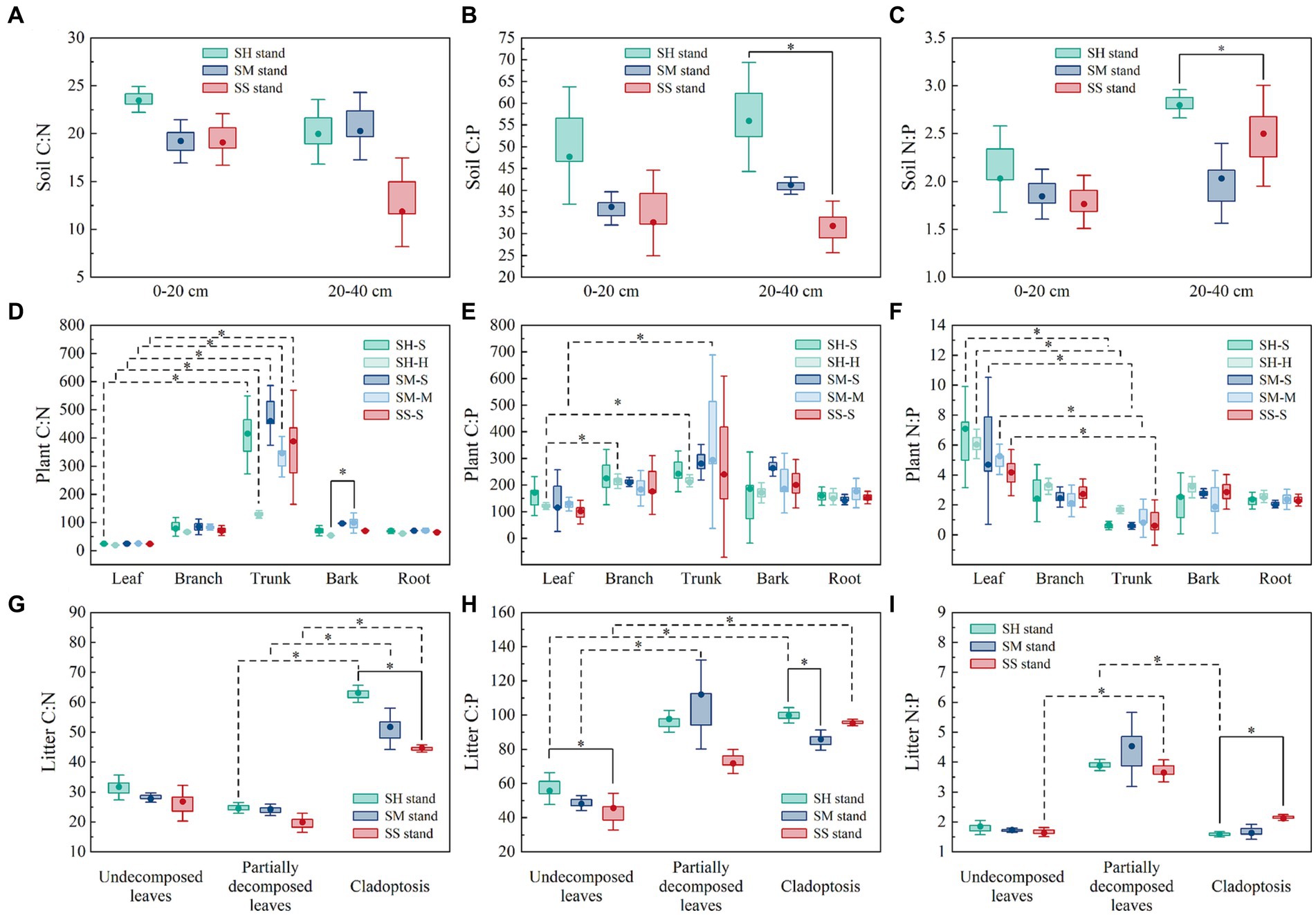
Figure 3. The C:N, C:P and N:P in soil, plant and litter of mixed and pure Chinese fir stands. Statistical significance is based on Kruskal-Wallis tests and indicated with asterisk (*). The box limits the 25–75th percentile range, the dots within the boxes are medians, the whiskers indicate maximum and minimum values. Solid black line indicate that the data are significantly different among different stand types (p < 0.05, n = 3), and black dotted line indicate that the data are significantly different among components (p < 0.05, n = 3). SH stand: the Chinese fir–Michelia macclurei mixed stands. SM stand: the Chinese fir–Mytilaria laosensis mixed stands. SS stand: pure Chinese fir stands.
The PCA results showed that the three stand species were clearly distinguished in the PCA plot and that the difference in elemental content and stoichiometric ratio between stands was much greater than that between species. There was an obvious positive correlation between the OC, TN and TP contents in different soil layers. Moreover, the soil elemental contents had a strong positive relationship with the plant TN content and the TP content of branches and bark, but the soil elemental contents had an obvious negative correlation with the TN content of litter. The plant OC and TP contents were positively correlated with those in litter (Figure 4A). For the stoichiometric ratio, there was a strong correlation between soil indexes as a whole. The soil stoichiometric ratios had a strong positive correlation with plant N:P and a negative correlation with plant C:N. The stoichiometric ratio of litter components was weakly correlated with plants and soils, except for plant leaf and root C:P (Figure 4B).
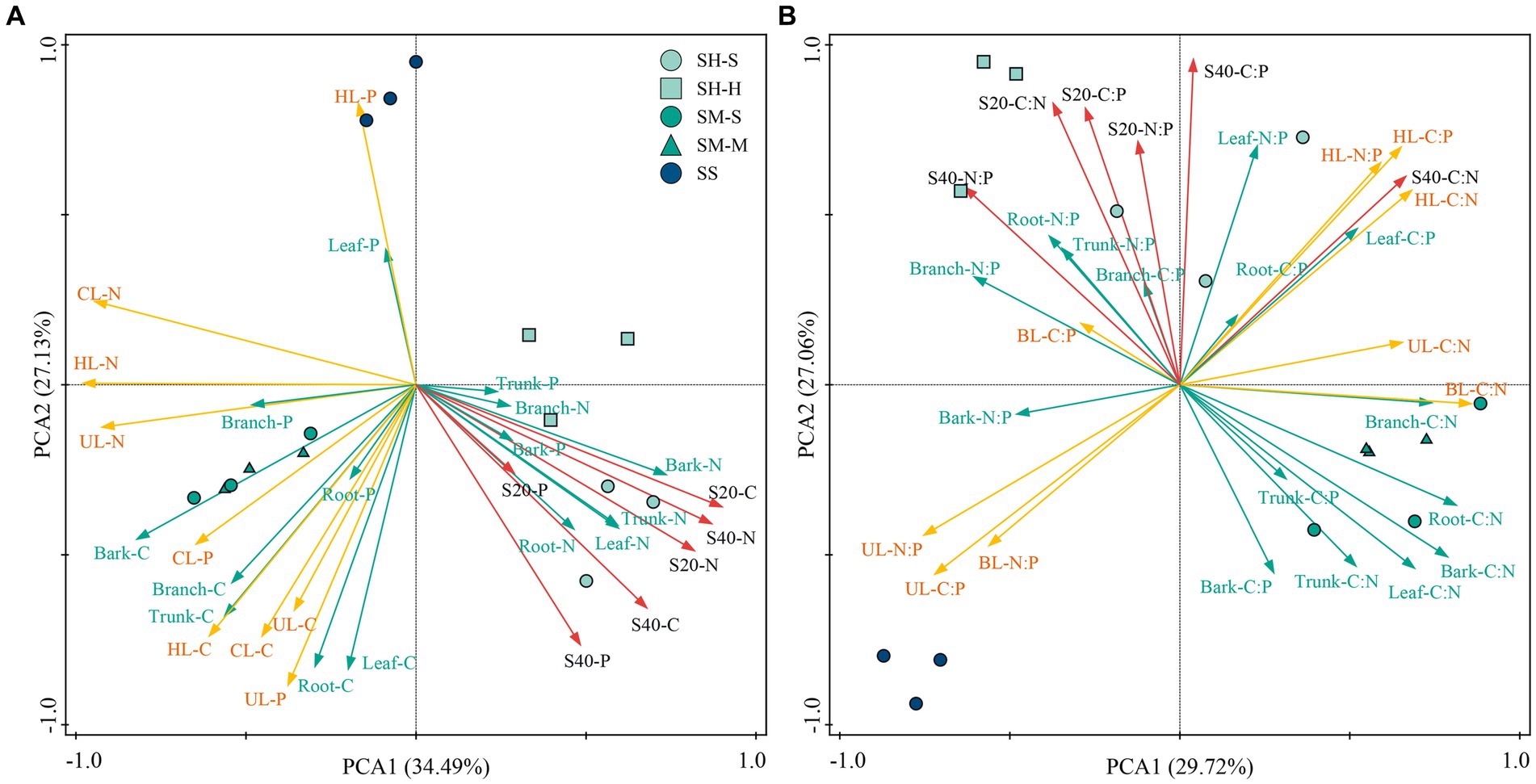
Figure 4. Principal component analysis of nutrient (A) and stoichiometric (B) ratios in plant-litter-soil systems. SH-S, Chinese fir in SH stand; SH-H, Michelia macclurei in SH stand; SM-S, Chinese fir in SM stand; SM-M, Mytilaria laosensis in SM stand; SS, Chinese fir in pure stand. C:N: Stoichiometric ratio of carbon to nitrogen, C:P: Stoichiometric ratio of carbon to phosphorus and N:P: Stoichiometric ratio of nitrogen to phosphorus. Red arrows, green arrows and yellow arrows indicate soil, plant and litter components, respectively. S20: 0-20 cm soil layer, S40: 20-40 cm soil layer. UL: Undecomposed leaves, HL: Partially decomposed leaves, CL: Cladoptosis.
The stoichiometric homeostasis analysis of different tree organs in different species showed that most plant organs in mixed and pure stands followed strict N and P homeostasis (p > 0.1), especially the leaf and root of Chinese fir and the root of mixed species (Table 3). It’s important to noting that a weak plasticity of P and a steady-state of N:P (p < 0.1) existed in the Michelia macclurei trunk, while other nonhomeostatic organs showed unsteady states (Table 3).

Table 3. The regression slopes (1/H) of relationships between plant and soil nutrient stoichiometry of mixed and pure Chinese fir stands.
In this study, the 0–40 cm soil OC, TN contents and 20–40 cm soil TP contents were significantly higher in the SH stand than in the SS stand (Figures 2A–C), which was consistent with the global trend (Guo et al., 2023). And the soil OC, TN and TP contents of the SM stand were not significantly higher than those of the pure stand (Figures 2A–C), indicating that the effect of mixed forest on soil nutrient improvement was highly dependent on tree species. The improvement in soil C, N, and P contents in the mixed stands can be attributed to the below reasons. First, the soil carbon and nutrient content advantages of mixed stands mainly depend on litter yield and quality advantages and the litter increased decomposition rate caused by the mixing of litter from different tree species (Wang et al., 2008, 2009; Zhao et al., 2013), which is accordance with the higher litter mass of mixed stands in this study (Table 2). Second, the improvement in soil chemical and physical properties (pH, aggregate structure, etc.) in mixed stands leads to differences in soil nutrient content (Zhou L. et al., 2020; Li et al., 2022). In addition, mixed tree species can alter the abundance, composition and activity of different functional microflora in soil, resulting in metabolic and ecological functional variation in soil microorganisms, which in turn influence soil carbon and nutrients fixation and cycling (Xia et al., 2019; Zhang et al., 2021; Ding et al., 2022). For example, Zhang et al. (2021) found that mixed stands of Cunninghamia lanceolata and Phoebe bournei increased the number of soil potential diazotrophic Firmicutes (i.e., Bacillales) and the abundance of soil N-fixing and nitrate-reducing related genes (NnifH and nrfA) compared to pure Cunninghamia lanceolata stands, which thereby increasing soil N fixation and cycling. Another study also showed that species mixing increased the relative abundance of K-strategy bacteria (Acidobacteria phylum) and decreased the relative abundance of r-strategy bacteria (Ascomycetes and Anaplasma phylum), which had an effect on soil C mineralization rates (Zhang X. et al., 2018). The soil pH and moisture content of the mixed stands were greater than in pure stands in this study, and the lower soil bulk density of mixed stands (Table 2) indicated that the soil in these stands was looser, which promote the organic matter decomposition by providing a more appropriate environment for soil microbial colonization and activity (Tao et al., 2017; Dai et al., 2021).
The content and distribution of C, N and P in plants reflect plant physiological functions and adaptability. In this study, leaves possessed the lowest OC content and the highest TN and TP content among the organs, while the trunk showed the opposite trend to the leaves (Figure 3). This suggests that plants need to allocate more nitrogen and phosphorus to satisfy their leaf building activities and sustain photosynthesis (Chen et al., 2013), while more carbohydrates are used to build trunks for support and carbon storage (Kraenzel et al., 2003). In addition, the C content of Chinese fir leaves in the present study decreased compared to the means of subtropical Chinese fir leaf carbon content (485.49 mg g−1), while the N and P contents were significantly higher than the average values of N (13.23 mg g−1) and P (1.24 mg g−1) in subtropical pure Chinese fir stands reported by Tong et al. (2021a,b). This means that the Chinese fir in this area has greater access to resources and higher growth rates than that in other areas. Although this study showed less soil N content than that reported by Tong et al. (2021b), according to the PCA results (Figure 4A), the increase in soil P content promoted an uptake of soil N by Chinese fir (Chen et al., 2015). This study also revealed that OC content of branch, trunk and root of Chinese fir was significantly increased in SM stands compared to SH stands, which increased the carbon sequestration potential of the plantation (Figure 2D). Furthermore, the TP contents of undecomposed leaves were all higher than those of partially decomposed leaves, which is due to the direct release of P elements during the decomposition process of Chinese fir rather than aggregation followed by release of P elements (Zhang et al., 2020).
Ecological stoichiometry reflects the elemental balance and linkages among different ecosystem components and is a valuable tool to study the forests element status (Zechmeister-Boltenstern et al., 2015). This result showed that soil stoichiometric ratios were enhanced to varying degrees in SH stands compared to pure stands (Figures 3A–C), which resulted from an enhancement of C and N concentrations of the various soil layers in the SH stands, consistent with the results of Wang Z.C. et al. (2021) for Chinese fir—Phoebe bournei mixed stands. It is generally accepted that higher C:N and C:P of litter determines its lower decomposition rate (Sariyildiz and Anderson, 2003; Wang Z.C. et al., 2021). The findings showed that mixed stands litter C:N and C:P was greater compared to pure stands to varying degrees (Figures 3A–C). Therefore, the quantity of litter was the primary driver of soil differences between mixed and pure stands (Table 2). In addition, C:N and C:P of litter leaves were overall lower than that of cladoptosis (Figures 3G–I), suggesting that leaf fractions in litter may be an important contributor to the fast conversion of plant to soil nutrients (Zhang et al., 2020).
In this study, Chinese fir stoichiometric ratios were less affected by stand change, but the stoichiometric ratios differed more significantly among organs (Figures 3D–F). Additionally, Leaf N:P below 14, showing that all stand species were relatively restricted by nitrogen and that pure stands were more severely restricted than mixed stands (Koerselman and Meuleman, 1996; Zhang et al., 2015). The findings suggest that although the soil TN content of mixed stands increased, it only alleviated but did not change the relative N limitation of the trees. The nutrient balance of trees is closely related to the nutrient status of the soil. A comparison with other relevant studies revealed that the soil TN content in this study was below that reported in other studies for subtropical pure Chinese fir plantations, usually higher than 1.5 g kg−1, while the TP content in this research was higher than the average from other studies (approximately 0.3 g kg−1) (Selvalakshmi et al., 2018; Duan et al., 2019; Li J.B. et al., 2021). That led to an imbalance in soil N and P, which was the main cause of the overall nitrogen limitation of the trees in this study (Figures 2B,C). Besides, the average values of C:N (24.52), C:p (133.26), and N:p (5.44) of Chinese fir leaves in the three stands were less than those of Chinese fir leaf C:N (37.60), C:p (428.73), and N:p (11.49) values in subtropical China reported by Tong et al. (2021a,b), and the leaf stoichiometric ratios in this study were lower than those of global forest tree and plant leaves (McGroddy et al., 2004; Zechmeister-Boltenstern et al., 2015). This suggests that trees at this site possess high N and P utilization efficiencies (Bai et al., 2019). A fast growing species, according to the growth rate hypothesis, generally has lower C:N, C:P, and N:P (Rivas-Ubach et al., 2012). The present study site is situated in low subtropical latitudes, where the relatively high light and temperature conditions support rapid growth, may have contributed to the relatively low stoichiometric ratios (Reich and Oleksyn, 2004).
The ecological stoichiometry of a plant reflects its ability to maintain homeostasis and adapt to environmental changes. It is generally accepted that autotrophs have flexible homeostasis, which can help them cope with changing habitats (Sterner and Elser, 2002; Persson et al., 2010). However, several recent studies have noted the complex status of homeostasis in plants (Wang et al., 2018; Peng et al., 2021; Bagedeng et al., 2022). It has been discovered that different organs of trees exhibit different homeostasis, which is attributed to fundamental trade-offs in plant nutrient investments (Yu et al., 2010; Wang et al., 2019). Moreover, homeostasis is closely related to the physiological functions of plant organs, with more active organs possessing more stable homeostasis (Zhang J.H. et al., 2018; Zhao et al., 2020). Our conclusions are in line with previous studies. Our study revealed that leaves and roots of Chinese fir and mixed species exhibit strict N and P homeostasis, which ensures organ functions (Aerts and Chapin, 2000; Wang et al., 2018). In fact, some research suggest that plant homeostasis and tree species are closely related, reflecting differences in environmental adaptability among tree species, i.e., species with greater degrees of homeostasis have better adaptive capacity to habitat modification (Wang K. et al., 2021; Ci et al., 2022). The results showed that compared with the SS stand, the N of the trunk and bark and the P of the trunk of Chinese fir in the SH stand, as well as the N of the branch, the P of the trunk and the N:P of the branch in the SM stand, changed from a strict homeostasis to an unsteady (Table 3). This change reflects the adaptation of Chinese fir to changes in N and P and the balance of N and P nutrients in mixed plantation environments (Wang K. et al., 2021). It is important to note that the leaf N:P of Michelia macclurei and Mytilaria laosensi exhibit “steady state” and “unsteady state” respectively (Table 3). This finding contrasts with most previous studies, which suggest that leaves maintain strict nutrient homeostasis to ensure normal photosynthesis, a process essential for plant growth and development (Aerts and Chapin, 2000; Wang et al., 2018; Wang K. et al., 2021). The above results exceeded our expectation that mixed stands promote nutrient homeostasis in trees, where Chinese fir and mixed species showed nonhomeostasis in branch, trunk, and bark, which seems to indicate that different tree species in mixed stands prefer a more conservative nutrient allocation strategy for branch, trunk, and bark (Peng et al., 2021; Wang K. et al., 2021). And our results also indicated the presence of lower N and P concentrations in these several organs (Figures 2E,F). Furthermore, the mixed species involved in this study, Michelia macclurei and Mytilaria laosensi, are both native broadleaf species, and their larger leaf area and light requirements relative to conifers may have led them to adopt more flexible strategies for leaf nutrient homeostasis in mixed plantations.
The new insights on tree homeostasis facilitate the selection of appropriate native species for converting pure plantations into mixed stands that are adapted to local conditions. In accordance with the sustainable development goals for Chinese fir plantations and the 2023 global sustainable development goals, ecologically friendly mixed species with high adaptive capacity, high productivity, high nutrient use efficiency, and large carbon sequestration (soil and biomass) will be preferred in the future (Liu et al., 2018). The study indicates that Michelia macclurei possesses higher mean diameter at breast height (Table 1), more stable N:P homeostasis (Table 3), and higher soil and plant carbon sinks compared to Mytilaria laosensi (Figures 2A,D). Therefore, Michelia macclurei may be a better choice for building mixed Chinese fir plantations.
The nutrient status of plantation systems is closely linked to global sustainable development. The study found that the impact of mixed forests on nutrient enhancement in the tree-litter-soil system varied depending on the stand. Specifically, SH stands significantly increased soil OC, TN, and TP content, while SM mixed stands increased the OC of partially decomposed leaves and the TP content of fallen branches. This improved the carbon sequestration function and nutrient storage capacity of plantations. As a result, it slowed down the global warming process and reduced nutrient loss and water pollution. Imbalances in soil N and P elements lead to potential relative N limitation of trees, while mixed plantations alleviate the degree of N limitation, promote aboveground tree growth, and increase carbon sinks. Besides, mixed stands reduced N, P, and N:P homeostasis in some tree organs, reflecting the adaptation of nutrient allocation strategies of different tree species to mixing. This finding provides a breakthrough for near-natural management of plantations. Selecting more suitable species for forestation is key to achieving sustainable development of plantations. Combining plant, litter, and soil C, N, and P performance and stoichiometry, mixed stands of Michelia macclurei and Chinese fir are considered to be a better measure to cope with the current global climate change and to achieve the goal of sustainable development. Additionally, evaluating the sustainability of mixed forests from an ecological perspective is an urgent task for the future.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
HZ: Conceptualization, Methodology, Visualization, Writing – original draft. XL: Data curation, Investigation, Writing – original draft. SW: Formal analysis, Writing – review & editing. CJ: Investigation, Writing – original draft. YC: Investigation, Writing – original draft. RF: Investigation, Writing – original draft. YL: Visualization, Writing – original draft. QZ: Supervision, Writing – original draft. SY: Funding acquisition, Project administration, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the National Natural Science Foundation of China (grant nos. 32260382 and 31460196).
The insightful comments from American Journal Experts (www.aje.cn) helped us to improve the grammar and phrasing of this manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2024.1293439/full#supplementary-material
Aerts, R., and Chapin, F. S. (2000). The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns. Adv. Ecol. Res. 30, 1–67. doi: 10.1016/S0065-2504(08)60016-1
Bagedeng, X., Xia, G., Lin, T., Xu, Z., and Wang, Y. (2022). Analysis of nutrient resorption efficiency and homeostasis of four tree species in kanas natural forest, Xinjiang, China. Front. Ecol. Evol. 10:1046444. doi: 10.3389/fevo.2022.1046444
Bai, X. J., Wang, B. R., An, S. S., Zeng, Q. C., and Zhang, H. X. (2019). Response of forest species to C:N:P in the plant-litter-soil system and stoichiometric homeostasis of plant tissues during afforestation on the loess plateau, China. Catena 183:104186. doi: 10.1016/j.catena.2019.104186
Bardgett, R. D., Mommer, L.De, and Vries, F. T. (2014). Going underground: root traits as drivers of ecosystem processes. Trends Ecol. Evol. 29, 692–699. doi: 10.1016/j.tree.2014.10.006
Bo, F. J., Zhang, Y. X., Chen, H. Y. H., Wang, P. G., Ren, X. M., and Guo, J. P. (2020). The C:N:P stoichiometry of planted and natural larix principis-rupprechtii stands along altitudinal gradients on the loess plateau, China. Forests 11:363. doi: 10.3390/f11040363
Bremner, J. M. (1996). “Nitrogen-total” in Part 3. Chemical methods. ed. D. L. Sparks , Vol. 5 (Madison, WI: ASA and SSSA), 1085–1121.
Chen, B., Chen, Y. Y., Jiang, L., Zhu, J., Chen, J. J., Huang, Q. R., et al. (2022). C:N:P stoichiometry of plant, litter and soil along an elevational gradient in subtropical forests of China. Forests 13:372. doi: 10.3390/f13030372
Chen, Y. H., Han, W. X., Tang, L. Y., Tang, Z. Y., and Fang, J. Y. (2013). Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form. Ecography 36, 178–184. doi: 10.1111/j.1600-0587.2011.06833.x
Chen, F. S., Niklas, K. J., Liu, Y., Fang, X. M., Wan, S. Z., and Wang, H. M. (2015). Nitrogen and phosphorus additions alter nutrient dynamics but not resorption efficiencies of Chinese fir leaves and twigs differing in age. Tree Physiol. 35, 1106–1117. doi: 10.1093/treephys/tpv076
Ci, H., Guo, C., Tuo, B., Zheng, L. T., Xu, M. S., Sai, B. L., et al. (2022). Tree species with conservative foliar nutrient status and strong phosphorus homeostasis are regionally abundant in subtropical forests. J. Ecol. 110, 1497–1507. doi: 10.1111/1365-2745.13884
Dai, W. W., Peng, B., Liu, J., Wang, C., Wang, X., Jiang, P., et al. (2021). Four years of litter input manipulation changes soil microbial characteristics in a temperate mixed forest. Biogeochemistry 154, 371–383. doi: 10.1007/s10533-021-00792-w
Ding, K., Zhang, Y. T., Yrjala, K., Tong, Z. K., and Zhang, J. H. (2022). The introduction of Phoebe bournei into Cunninghamia lanceolata monoculture plantations increased microbial network complexity and shifted keystone taxa. Forest. Ecol. Manag. 509:120072. doi: 10.1016/j.foreco.2022.120072
Duan, A. G., Lei, J., Hu, X. Y., Zhang, J. G., Du, H. L., Zhang, X. Q., et al. (2019). Effects of planting density on soil bulk density, pH and nutrients of unthinned Chinese fir mature stands in south subtropical region of China. Forests 10:351. doi: 10.3390/f10040351
Fan, H. B., Wu, J. P., Liu, W. F., Yuan, Y. H., Hu, L., and Cai, Q. K. (2015). Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant Soil 392, 127–138. doi: 10.1007/s11104-015-2444-2
Farooq, T. H., Yan, W., Rashid, M. H. U., Tigabu, M., Gilani, M. M., Zou, X. H., et al. (2019). Chinese fir (Cunninghamia Lanceolata) a green gold of China with continues decline in its productivity over the successive rotations: a review. Appl. Ecol. Env. Res. 17, 11055–11067. doi: 10.15666/aeer/1705_1105511067
Feng, Y. H., Schmid, B., Loreau, M., Forrester, D. I., Fei, S. L., Zhu, J. X., et al. (2022). Multispecies forest plantations outyield monocultures across a broad range of conditions. Science 376:865-+. doi: 10.1126/science.abm6363
Gan, H. Y., Schoning, I., Schall, P., Ammer, C., and Schrumpf, M. (2020). Soil organic matter mineralization as driven by nutrient stoichiometry in soils under differently managed forest stands. Front. For. Glob. Chang. 3:99. doi: 10.3389/ffgc.2020.00099
Guan, F. Y., Tang, X. L., Fan, S. H., Zhao, J. C., and Peng, C. (2015). Changes in soil carbon and nitrogen stocks followed the conversion from secondary forest to Chinese fir and Moso bamboo plantations. Catena 133, 455–460. doi: 10.1016/j.catena.2015.03.002
Guo, J. H., Feng, H. L., Mcnie, P., Liu, Q. Y., Xu, X., Pan, C., et al. (2023). Species mixing improves soil properties and enzymatic activities in Chinese fir plantations: a meta-analysis. Catena 220:106723. doi: 10.1016/j.catena.2022.106723
Handa, I. T., Aerts, R., Berendse, F., Berg, M. P., Bruder, A., Butenschoen, O., et al. (2014). Consequences of biodiversity loss for litter decomposition across biomes. Nature 509:218-+. doi: 10.1038/nature13247
He, Y. J., Qin, L., Li, Z. Y., Liang, X. Y., Shao, M. X., and Tan, L. (2013). Carbon storage capacity of monoculture and mixed-species plantations in subtropical China. Forest. Ecol. Manag. 295, 193–198. doi: 10.1016/j.foreco.2013.01.020
He, Y., Zhang, Q., Jiang, C., Lan, Y., Zhang, H., and Ye, S. (2023). Mixed planting improves soil aggregate stability and aggregate-associated C-N-P accumulation in subtropical China. Front. For. Glob. Chang. 6:e1141953. doi: 10.3389/ffgc.2023.1141953
Hou, L. Y., Zhang, Y. Q., Li, Z. C., Shao, G. D., Song, L. G., and Sun, Q. W. (2021). Comparison of soil properties, understory vegetation species diversities and soil microbial diversities between Chinese fir plantation and close-to-natural forest. Forests 12:632. doi: 10.3390/f12050632
Koerselman, W., and Meuleman, A. F. M. (1996). The vegetation N:P ratio: a new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 33, 1441–1450. doi: 10.2307/2404783
Kraenzel, M., Castillo, A., Moore, T., and Potvin, C. (2003). Carbon storage of harvest-age teak (Tectona grandis) plantations. Panama. Forest. Ecol. Manag. 173, 213–225. doi: 10.1016/S0378-1127(02)00002-6
Laganière, J., Angers, D. A., and Paré, D. (2010). Carbon accumulation in agricultural soils after afforestation: a meta-analysis. Glob. Chang. Biol. 16, 439–453. doi: 10.1111/j.1365-2486.2009.01930.x
Li, J. B., Cao, X. Y., Wang, Y. R., Yan, W. D., Peng, Y. Y., and Chen, X. Y. (2021). Effects of thinning on soil nutrients in a chronosequence of Chinese fir forests in subtropical China. Ann. For. Res. 64, 147–158. doi: 10.15287/afr.2020.2206
Li, Y. F., Li, Q. Y., Liu, H. S., Li, S. J., Wang, Y. J., and Liu, Y. (2018). Ecological stoichiometry-based study of the influence of soil saline-alkali stress on nutrient homeostasis in L. chinensis. Ecotox. Environ. Saf. 165, 243–249. doi: 10.1016/j.ecoenv.2018.09.020
Li, X., Liao, L., Yang, H., Huang, Y., Li, Y., and Ye, S. (2019). Spatial pattern of woody plants in artificial mixed forest of Cunninghamia lanceolata and Mytilaria laosensis. Acta Botan. Boreali-Occiden. Sin. 39, 1842–1850. doi: 10.7606/j.issn.1000-4025.2019.10.1842
Li, W. Q., Shen, F. X., Liu, Y. J., Li, Z. W., Jiang, J. F., Li, Q., et al. (2022). Soil depth determine the ecological stoichiometry of soil aggregates after returning ancient rice terraces to forest. Catena 219:106587. doi: 10.1016/j.catena.2022.106587
Li, Y., Xu, J., Wang, H., Nong, Y., Sun, G., Yu, S., et al. (2021). Long-term effects of thinning and mixing on stand spatial structure: a case study of Chinese fir plantations. Iforest 14, 113–121. doi: 10.3832/ifor3489-014
Liu, C. L. C., Kuchma, O., and Krutovsky, K. V. (2018). Mixed-species versus monocultures in plantation forestry: development, benefits, ecosystem services and perspectives for the future. Glob. Ecol. Conserv. 15:e400419. doi: 10.1016/j.gecco.2018.e00419
Luo, X. Z., Hou, E. Q., Chen, J. Q., Li, J., Zhang, L. L., Zang, X. W., et al. (2020). Dynamics of carbon, nitrogen, and phosphorus stocks and stoichiometry resulting from conversion of primary broadleaf forest to plantation and secondary forest in subtropical China. Catena 193:104606. doi: 10.1016/j.catena.2020.104606
Mcgroddy, M. E., Daufresne, T., and Hedin, L. O. (2004). Scaling of C: N: P stoichiometry in forests worldwide: implications of terrestrial Redfield-type ratios. Ecology 85, 2390–2401. doi: 10.1890/03-0351
Miao, L. J., Moore, J. C., Zeng, F. J., Lei, J. Q., Ding, J. L., He, B., et al. (2015). Footprint of research in desertification management in China. Land Degrad. Dev. 26, 450–457. doi: 10.1002/ldr.2399
Minden, V., and Kleyer, M. (2014). Internal and external regulation of plant organ stoichiometry. Plant Biol. 16, 897–907. doi: 10.1111/plb.12155
Mooshammer, M., Wanek, W., Schnecker, J., Wild, B., Leitner, S., Hofhansl, F., et al. (2012). Stoichiometric controls of nitrogen and phosphorus cycling in decomposing beech leaf litter. Ecology 93, 770–782. doi: 10.1890/11-0721.1
Nelson, D. W., and Sommers, L. E. (1996). “Total carbon, organic carbon and organic matter” in Part 3. Chemical methods. ed. D. L. Sparks , vol. 5 (Madison, WI: ASA and SSSA), 961–1010.
Peng, S. S., Piao, S. L., Zeng, Z. Z., Ciais, P., Zhou, L. M., Li, L. Z. X., et al. (2014). Afforestation in China cools local land surface temperature. Proc. Natl. Acad. Sci. U. S. A. 111, 2915–2919. doi: 10.1073/pnas.1315126111
Peng, C., Tu, J., Yang, M., Meng, Y., Li, M. Q., and Ai, W. S. (2021). Root stoichiometric dynamics and homeostasis of invasive species Phyllostachys edulis and native species Cunninghamia lanceolata in a subtropical forest in China. J. Forestry. Res. 32, 2001–2010. doi: 10.1007/s11676-020-01253-0
Persson, J., Fink, P., Goto, A., Hood, J. M., Jonas, J., and Kato, S. (2010). To be or not to be what you eat: regulation of stoichiometric homeostasis among autotrophs and heterotrophs. Oikos 119, 741–751. doi: 10.1111/j.1600-0706.2009.18545.x
Pugh, T. A. M., Lindeskog, M., Smith, B., Poulter, B., Arneth, A., Haverd, V., et al. (2019). Role of forest regrowth in global carbon sink dynamics. Proc. Natl. Acad. Sci. U. S. A. 116, 4382–4387. doi: 10.1073/pnas.1810512116
Reich, P. B., and Oleksyn, J. (2004). Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. U. S. A. 101, 11001–11006. doi: 10.1073/pnas.0403588101
Rivas-Ubach, A., Sardans, J., Perez-Trujillo, M., Estiarte, M., and Penuelas, J. (2012). Strong relationship between elemental stoichiometry and metabolome in plants. Proc. Natl. Acad. Sci. U. S. A. 109, 4181–4186. doi: 10.1073/pnas.1116092109
Robin, L. C. (2008). Beyond deforestation: restoring forests and ecosystem services on degraded lands. Science 320, 1458–1460. doi: 10.1126/science.1155365
Sariyildiz, T., and Anderson, J. M. (2003). Interactions between litter quality, decomposition and soil fertility: a laboratory study. Soil Biol. Biochem. 35, 391–399. doi: 10.1016/S0038-0717(02)00290-0
Selvalakshmi, S., Vasu, D., Zhijun, H., Guo, F., and Ma, X. Q. (2018). Soil nutrients dynamics in broadleaved forest and Chinese fir plantations in subtropical forests. J. Trop. For. Sci. 30, 242–251. doi: 10.26525/jtfs2018.30.2.242251
Selvaraj, S., Duraisamy, V., Huang, Z. J., Guo, F. T., and Ma, X. Q. (2017). Influence of long-term successive rotations and stand age of Chinese fir (Cunninghamia lanceolata) plantations on soil properties. Geoderma 306, 127–134. doi: 10.1016/j.geoderma.2017.07.014
Sterner, R. W., and Elser, J. J. (2002). Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton, NJ: Princeton University Press.
Tao, J. M., Liu, X. D., Liang, Y. L., Niu, J. J., Xiao, Y. H., Gu, Y. B., et al. (2017). Maize growth responses to soil microbes and soil properties after fertilization with different green manures. Appl. Microbiol. Biot. 101, 1289–1299. doi: 10.1007/s00253-016-7938-1
Tong, R., Cao, Y. N., Zhu, Z. H., Lou, C. Y., Zhou, B. Z., and Wu, T. G. (2021a). Solar radiation effects on leaf nitrogen and phosphorus stoichiometry of Chinese fir across subtropical China. For. Ecosyst. 8:62. doi: 10.1186/s40663-021-00344-6
Tong, R., Zhou, B. Z., Jiang, L. N., Ge, X. G., and Cao, Y. H. (2021b). Spatial patterns of leaf carbon, nitrogen, and phosphorus stoichiometry and nutrient resorption in Chinese fir across subtropical China. Catena 201:105221. doi: 10.1016/j.catena.2021.105221
Wang, Z. C., He, G. X., Hou, Z. H., Luo, Z., Chen, S. X., Lu, J., et al. (2021). Soil C:N:P stoichiometry of typical coniferous (Cunninghamia lanceolata) and/or evergreen broadleaved (Phoebe bournei) plantations in South China. Forest. Ecol. Manag. 486:118974. doi: 10.1016/j.foreco.2021.118974
Wang, S. Q., Huang, Y. Z., and Ye, S. M. (2021). Distribution of organic carbon and nutrients in soil aggregates under different stand types of Cunninghamia lanceolata in southern Guangxi of China. J. Soil. Sci. Plant. Nut. 67, 427–438. doi: 10.1080/00380768.2021.1932585
Wang, J. Y., Wang, J. N., Guo, W. H., Li, Y. G., Wang, G. G., and Wu, T. G. (2018). Stoichiometric homeostasis, physiology, and growth responses of three tree species to nitrogen and phosphorus addition. Trees Struct. Funct. 32, 1377–1386. doi: 10.1007/s00468-018-1719-7
Wang, Q. K., Wang, S. L., and Huang, Y. (2008). Comparisons of litterfall, litter decomposition and nutrient return in a monoculture Cunninghamia lanceolata and a mixed stand in southern China. Forest. Ecol. Manag. 255, 1210–1218. doi: 10.1016/j.foreco.2007.10.026
Wang, Q. K., Wang, S. L., and Huang, Y. (2009). Leaf litter decomposition in the pure and mixed plantations of Cunninghamia lanceolata and Michelia macclurei in subtropical China. Biol. Fert. Soils. 45, 371–377. doi: 10.1007/s00374-008-0338-7
Wang, J. N., Wang, J. Y., Wang, L., Zhang, H., Guo, Z. W., Wang, G. G., et al. (2019). Does stoichiometric homeostasis differ among tree organs and with tree age? Forest. Ecol. Manag. 453:117637. doi: 10.1016/j.foreco.2019.117637
Wang, S. L., Zhang, W. D., and Sanchez, F. (2010). Relating net primary productivity to soil organic matter decomposition rates in pure and mixed Chinese fir plantations. Plant Soil 334, 501–510. doi: 10.1007/s11104-010-0400-8
Wang, K., Zhang, R. S., Song, L. N., Yan, T., and Na, E. H. (2021). Comparison of C:N:P stoichiometry in the plant-litter-soil system between poplar and elm plantations in the Horqin Sandy land, China. Front. Plant Sci. 12:655517. doi: 10.3389/fpls.2021.655517
Wang, Z. F., and Zheng, F. L. (2020). Ecological stoichiometry of plant leaves, litter and soils in a secondary forest on China's loess plateau. Peerj 8:e10084. doi: 10.7717/peerj.10084
Wei, Y. J., Dang, X. H., Wang, J., Gao, J. L., and Gao, Y. (2021). Response of C:N:P in the plant-soil system and stoichiometric homeostasis of Nitraria tangutorum leaves in the oasis-desert ecotone, Northwest China. J. Arid. Land 13, 934–946. doi: 10.1007/s40333-021-0019-z
Wieder, W. R., Cleveland, C. C., Smith, W. K., and Todd-Brown, K. (2015). Future productivity and carbon storage limited by terrestrial nutrient availability. Nat. Geosci. 8, 441–444. doi: 10.1038/Ngeo2413
Xia, Z. C., Yu, L., He, Y., Korpelainen, H., and Li, C. Y. (2019). Broadleaf trees mediate chemically the growth of Chinese fir through root exudates. Biol. Fert. Soils 55, 737–749. doi: 10.1007/s00374-019-01389-0
Yang, Y., Liu, B. R., and An, S. S. (2018). Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of northern China. Catena 166, 328–338. doi: 10.1016/j.catena.2018.04.018
Yu, Q., Chen, Q., Elser, J. J., He, N., Wu, H., Zhang, G., et al. (2010). Ecol. Lett. 13, 1390–1399. doi: 10.1111/j.1461-0248.2010.01532.x
Yu, Q., Wilcox, K., La Pierre, K., Knapp, A. K., Han, X. G., and Smith, M. D. (2015). Stoichiometric homeostasis predicts plant species dominance, temporal stability, and responses to global change. Ecology 96, 2328–2335. doi: 10.1890/14-1897.1
Zechmeister-Boltenstern, S., Keiblinger, K. M., Mooshammer, M., Penuelas, J., Richter, A., Sardans, J., et al. (2015). The application of ecological stoichiometry to plant-microbial-soil organic matter transformations. Ecol. Monogr. 85, 133–155. doi: 10.1890/14-0777.1
Zhang, Y. T., Ding, K., Yrjala, K., Liu, H. Y., Tong, Z. K., and Zhang, J. H. (2021). Introduction of broadleaf species into monospecific Cunninghamia lanceolata plantations changed the soil Acidobacteria subgroups composition and nitrogen-cycling gene abundances. Plant Soil 467, 29–46. doi: 10.1007/s11104-021-05014-8
Zhang, J., Li, J., Fan, Y., Mo, Q., Li, Y., Li, Y., et al. (2020). Effect of nitrogen and phosphorus addition on litter decomposition and nutrients release in a tropical forest. Plant Soil 454, 139–153. doi: 10.1007/s11104-020-04643-9
Zhang, X., Liu, S., Huang, Y., Fu, S., Wang, J., Ming, A., et al. (2018). Tree species mixture inhibits soil organic carbon mineralization accompanied by decreased r-selected bacteria. Plant Soil 431, 203–216. doi: 10.1007/s11104-018-3755-x
Zhang, H., Wang, J. N., Wang, J. Y., Guo, Z. W., Wang, G. G., Zeng, D. H., et al. (2018). Tree stoichiometry and nutrient resorption along a chronosequence of Metasequoia glyptostroboides forests in coastal China. Forest. Ecol. Manag. 430, 445–450. doi: 10.1016/j.foreco.2018.08.037
Zhang, J. H., Zhao, N., Liu, C. C., Yang, H., Li, M. L., Yu, G. R., et al. (2018). C:N:P stoichiometry in China's forests: from organs to ecosystems. Funct. Ecol. 32, 50–60. doi: 10.1111/1365-2435.12979
Zhang, W., Zhao, J., Pan, F. J., Li, D. J., Chen, H. S., and Wang, K. L. (2015). Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in Southwest China. Plant Soil 391, 77–91. doi: 10.1007/s11104-015-2406-8
Zhao, L., Hu, Y. L., Lin, G. G., Gao, Y. C., Fang, Y. T., and Zeng, D. H. (2013). Mixing effects of understory plant litter on decomposition and nutrient release of tree litter in two plantations in Northeast China. PLoS One 8:e76334. doi: 10.1371/journal.pone.0076334
Zhao, N., Yu, G. R., Wang, Q. F., Wang, R. L., Zhang, J. H., Liu, C. C., et al. (2020). Conservative allocation strategy of multiple nutrients among major plant organs: from species to community. J. Ecol. 108, 267–278. doi: 10.1111/1365-2745.13256
Zhou, B., Peng, D., Zhao, Q. X., Yangnan, S. Y., Yang, S. Q., Yang, F., et al. (2020). Improvements in timber production of Chinese fir (Cunninghamia lanceolata) per unit forest area in China via tree breeding: status and challenges. Dendrobiology 83, 43–51. doi: 10.12657/denbio.083.004
Keywords: Chinese fir, mixed plantation, tree-litter-soil system, ecological stoichiometry, stoichiometric homeostasis
Citation: Zhang H, Li X, Wang S, Jiang C, Cui Y, Fan R, Lan Y, Zhang Q and Ye S (2024) Tree–litter–soil system C:N:P stoichiometry and tree organ homeostasis in mixed and pure Chinese fir stands in south subtropical China. Front. For. Glob. Change. 7:1293439. doi: 10.3389/ffgc.2024.1293439
Edited by:
Mehebub Sahana, The University of Manchester, United KingdomReviewed by:
Lydie-Stella Koutika, CRDPI, Republic of CongoCopyright © 2024 Zhang, Li, Wang, Jiang, Cui, Fan, Lan, Zhang and Ye. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shaoming Ye, eXNoYW9taW5nQDE2My5jb20=
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.