 Ping Liu1,2
Ping Liu1,2 Shiyu Hu
Shiyu Hu Hongxu Wei
Hongxu Wei Wenting He
Wenting He
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. For. Glob. Change , 19 October 2023
Sec. Temperate and Boreal Forests
Volume 6 - 2023 | https://doi.org/10.3389/ffgc.2023.1272477
This article is part of the Research Topic Forest Ecosystems in Mountain Regions: Conditions, Risks and Impacts View all 13 articles
The frequency and severity of drought events are increasing under a changing climate, trees of different stand ages respond differently to drought events, which has a great impact on the stability of forest ecosystems. In this study, we measured radial growth (RG) in cored trees from 49 forests including young stands (20–30a) and middle-aged stands (31–50a) of Pinus sylvestris var. mongolica plantations in a semi-arid area of western Liaoning, China. We evaluated the response of RG of P. sylvestris to long-term climate, and calculate three response indicators: resistance (Rt), recovery (Rc) and resilience (Rs), so as to measure the growth response of trees to drought events. Results showed that a negative correlation was detected between RG of young stands and the monthly highest temperature (MHT) in April and May. Positive correlations were found between RG of young stands and the monthly lowest temperature (MLT) across periods from September to November, when RG of young stands was also positively correlated with Palmer Drought Severity Index (PDSI) across whole-years. There was a positive correlation between RG of middle-aged stands and MLT in September, and PDSI from June to December. After the first drought event, most pine trees recovered their RG (Rc > 1, Rs > 1). However, after three consecutive drought events, Rt, Rc, and Rs of pine trees decreased significantly (p < 0.05), and Rt and Rs were less than 1. In summary, younger pine trees are more sensitive to climate change, and spring drought is more inhibitory to growth of pine trees than high summer temperatures. Pine trees have a compensation effect after experiencing drought events, but the cumulative effect of multiple drought events will gradually offset the compensation effect of trees and eventually decline in pine tree growth, while the resistance and resilience of young stands after continuous drought events were significantly lower than those of middle-aged stands, and have a higher risk of death.
In recent decades, with global warming, drought events have become more frequent and drought severity has increased (Allen et al., 2010; Field et al., 2012), which has become a serious challenge for forest ecosystems (Bolte et al., 2009; Lindner et al., 2010). Studies have shown that forests are adaptive to drought events and can achieve self-regulation within the corresponding range (Adams, 2009; Lloret et al., 2011; Longo et al., 2018). However, extreme drought events beyond the range of adaptation can affect tree survival and thus the productivity and biodiversity of forest ecosystems (Bolte et al., 2009; van Mantgem et al., 2009; Allen et al., 2010). Therefore, an accurate assessment of the adaptability of trees to drought events is helpful for understanding and predicting the survival risk of forests under future climate change, which is of great importance for maintaining sustainable forest management (Oliver et al., 2015).
Dendrochronology is an important method for studying the response of tree radial growth (RG) to climate change with temporal certainty and annual continuity (Martín-Benito et al., 2008; Fritts, 2012). Studies have shown that temperature and precipitation are important drivers of tree growth. In both tropical and cold regions, low temperature in winter, rainfall during the growing season, and snowfall in winter promote tree growth (Bigelow et al., 2014; Yuanqiao et al., 2021; Jing et al., 2022; Ning et al., 2023). High summer temperatures often have a significant inhibitory effect on RG of trees (Gantois, 2022; Jing et al., 2022). To quantify the response of RG to climate change, Lloret et al. (2011) proposed three indicators resistance (Rt), recovery (Rc) and resilience (Rs) to evaluate the tolerance of trees to drought events. Studies have shown that forests with low stand density have stronger resistance and recovery to drought events than forests with high stand density, and reducing stand density contributes to tree growth (Hollunder et al., 2021; Ovenden et al., 2021). Trees in different ecological environments respond differently to drought events. Compared to arid areas, trees in humid areas generally grow better and recover faster (Gazol et al., 2017; Longo et al., 2018). The Rt, Rc, and Rs of different tree species to drought events also differ (Zang et al., 2014; Vitali et al., 2017), but there is still controversy about the response of the same tree species of different sizes to drought events: Some studies have found that large trees have a poor recovery capacity, while small trees have a stronger adaptability to dry environments (Pretzsch et al., 2018; Trugman et al., 2018), but other studies have shown that small trees are more susceptible (Colangelo et al., 2017; Versace et al., 2022). Age, as an important ecological indicator of forest community structure, affects the growth status of trees and the assessment of forest ecosystems (Law et al., 2001; Song and Woodcock, 2003; Tang et al., 2017). Trees of different ages respond differently to drought events and also affect the stability of forest ecosystems. The high mortality of small trees is likely to alter the trajectory of ecosystem succession, while the high mortality of large trees will disrupt the balance of the ecosystem (Lucas-Borja et al., 2021). In addition, some studies have found that some trees recover quickly after experiencing drought events and even exceed the original growth level, but some trees cannot recover to the previous growth level and are still affected by the legacy effect of drought events for a long time (Serra-Maluquer et al., 2018; Kannenberg et al., 2019). The ability of trees to recover quickly from drought events has a significant impact on forest ecosystems.
In order to prevent and control dust storms, soil desertification, soil erosion, etc., China started to implement the Three-North Shelter Forest Program in the late 1970s (Zhang et al., 2016). Pinus sylvestris has become one of the main afforestation tree species in the shelter forest due to its excellent characteristics of cold resistance, drought resistance, barren resistance, suitable sandy soil and fast growth. Zhanggutai Town, Zhangwu County, western Liaoning Province, is the first area in China to introduce P. sylvestris, and it is also the most important experimental demonstration base for sand-fixing afforestation of P. sylvestris (Yuzhang, 1990). After that, the introduction area of P. sylvestris continued to expand southward and westward, covering the entire “Three North” project area. Therefore, whether the widely planted P. sylvestris plantation can grow healthily directly affects the effectiveness of afforestation, windbreak and sand fixation in western Liaoning. As a typical semi-arid area in China, western Liaoning may face more severe climate challenges in the future, and trees will be exposed to multiple drought events (Allen et al., 2010; Zuidema et al., 2022). Therefore, it is very important to explore the response characteristics of P. sylvestris plantations of different stand ages to climatic factors, and to study the response ability of P. sylvestris of different stand ages to continuous multiple drought events. It is conducive to maintaining the stability of forest ecosystems in the context of climate change, improving forest management and forecasting, and understanding forest dynamics in the context of future global change.
In this study, P. sylvestris plantations in the semi-arid area of western Liaoning were taken as the research object, and the long-term climate change and sudden extreme drought events were combined to evaluate the differences in growth dynamic characteristics of P. sylvestris plantations at different stand ages under different climatic conditions. We hypothesized that: (1) RG of P. sylvestris in different age stands responds differently to climatic factors; (2) drought events have legacy effect and cumulative effect; (3) the Rt, Rc and Rs of P. sylvestris in different age stands were significantly different.
The study area is located in Zhangwu County, Liaoning Province, in the southeastern edge of Khorchin Sandy Land, which is a typical semi-arid area. The geographical location is between 121° 53 ‘and 122° 58 ‘east longitude, 42° 07 ‘and 42° 51 ‘north latitude, with an average altitude of 255 m. The main feature of the landform is that the dunes overlap each other, and there are wind erosion lowlands between the hills. It belongs to the temperate continental monsoon climate, and the main climate characteristics are drought and wind. The representative plants are Pinus sylvestris, Pinus tabuliformis, Ulmus pumila, etc. The meteorological data of the last decades show that the extreme lowest temperature is −36.3°C, the extreme highest temperature is 38.4°C, and the average annual temperature is 7.93°C. The hottest season appears in July, with an average temperature of 24.2°C. The coldest month is January, with an average temperature of −11.5°C (Figure 1). The average annual precipitation is 570.8 mm, the maximum annual precipitation is 1,390 mm, and the minimum annual precipitation is 295.1 mm. The seasonal distribution of precipitation is uneven, and the annual precipitation is concentrated in June, July and August. There is less snow in winter, spring precipitation is low, and spring drought is common (Figure 2).
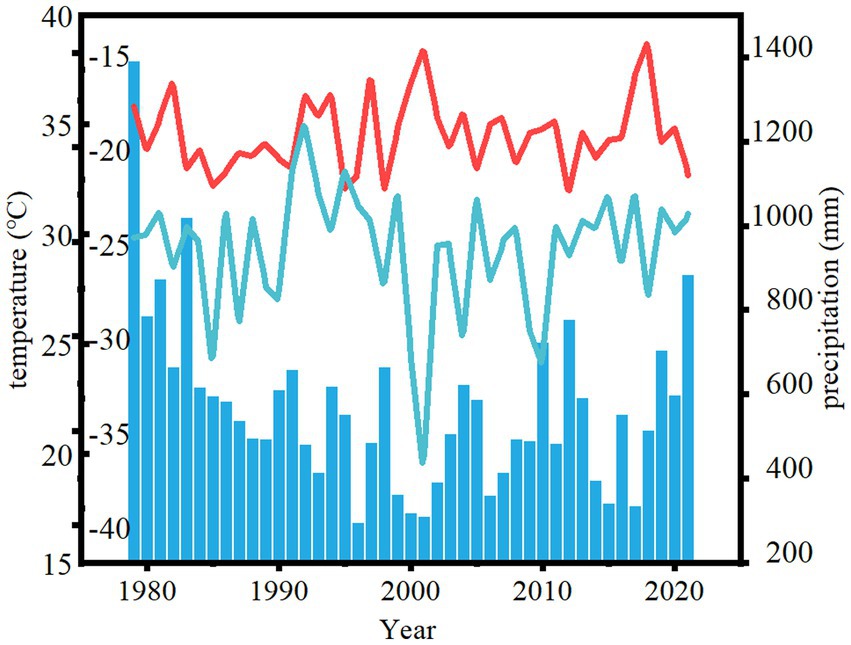
Figure 1. Precipitation and temperature changes in Zhangwu County from 1979 to 2021. The red line indicates the highest temperature of the year, the blue line indicates the lowest temperature of the year, and the bar chart indicates the total annual precipitation.
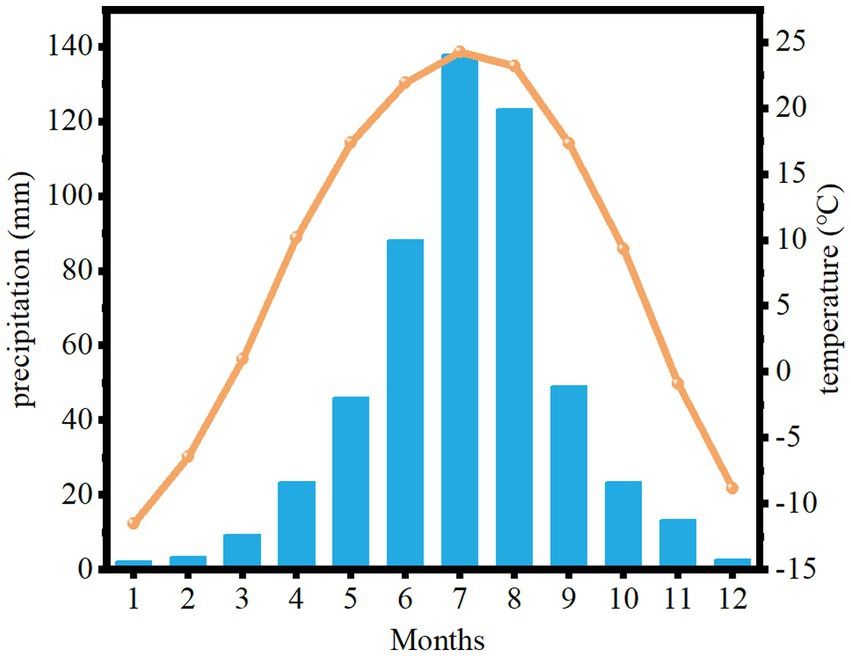
Figure 2. Monthly average precipitation and temperature changes in Zhangwu County from 1979 to 2021. The orange line represents the monthly average temperature, and the bar chart represents the monthly average precipitation.
The sample plot of P. sylvestris artificial shelter forest is located in the experimental forest farm of Shenyang Agricultural University in Aer Township, Zhangwu County. According to the National Forestry Administration (2017), in the artificial shelter forest of P. sylvestris, young stands (≤ 30a) and middle-aged stands (31a ~ 50a) with basically the same site conditions (Supplementary Table S1), the dominant trees with straight trunks, no pests and diseases, no scars, and good growth were selected. The diameter at breast height, tree height, and crown width of each tree were measured by breast height rule, height meter, and tape measure (Table 1). To study RG of tree rings, we used an increment borer with a diameter of 5.15 mm to drill two tree cores at breast height (1.3 m) of each tree. When drilling, the increment borer is perpendicular to the trunk, avoiding trunk deformation and depression, and only the core without breakage is retained.
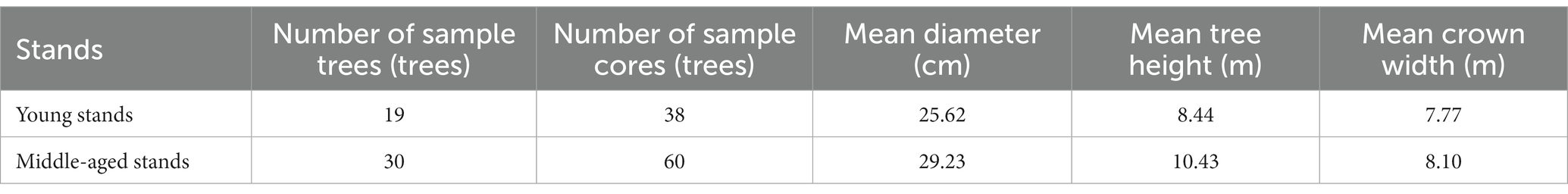
Table 1. Basic information of Pinus sylvestris var. mongolica in different agestands.
The tree core was loaded into a special plastic tube, numbered, and returned to the laboratory. In the laboratory, the tree core was fixed in a wooden trough with white emulsion and paper tape for natural air drying. After air drying, the tree rings were polished in the laboratory with 240, 320, 400, and 600 mesh sandpaper in turn in the laboratory until the bright and clear tree ring contour could be seen under the microscope. The polished tree ring was placed under the microscope for preliminary marking and dating. Then the width of each annual ring is measured with a LINTAB. Six measuring instrument with an accuracy of 0.001 mm. Finally, the COFECHA software was used to check and correct the measurement and dating results in order to ensure the accuracy of the dating, to remove the sequence of poor quality, and finally 98 cores were kept, including 38 in the young stands and 60 in the middle-aged stands.
To eliminate the influence of the trees’ own physiological factors, we used negative exponential curve or linear regression to fit the growth trend of the trees and detrend the ring-width series in the ARSTAN program, and the mean standard chronologies for each age stand were calculated as the bi-weight robust mean of the detrended individual series. Finally, the standard chronology of each age stand was obtained and used to analyze the response of stand growth to climate. Express population signal (EPS) values of the two stand ages were greater than 0.85, mean sensitivity (MS)of young stands and middle-aged stands were 0.211 and 0.208, respectively. And serial correlation were 0.505 and 0.449, respectively. In addition, to more accurately quantify the growth of individual P. sylvestris, we used the tree-ring width to calculate the basal area increment (BAI) of each sample tree to better analyze the response of its RG to extreme drought events. The calculation formula is as follows:
In the formula, Rt and Rt-1 are the radius of the tree corresponding to year t and year t-1, respectively. The tree radius is calculated from the tree-ring width.
The daily lowest temperature, daily highest temperature, daily average temperature and daily precipitation data were obtained from the Zhangwu County Meteorological Bureau (42 ° 38 ‘N, 122 ° 55 ‘E) closest to the sampling site. In addition, the Palmer Drought Severity Index (PDSI) grid data are obtained using the data sharing network1 of the KNMI Climate Explorer. PDSI data set is CRU self-calibrated data with a spatial resolution of 0.5° × 0.5°. Compared to other drought indices, PDSI takes into account temperature, precipitation, soil moisture, and water evaporation (Palmer, 1965). Since it was proposed by Palmer in 1965, it has been used as one of the most widely used indices for the study and detection of meteorological drought (Sun et al., 2012; Yan et al., 2014; Smerdon et al., 2015). In order to analyze the differences in the response of growth of P. sylvestris plantations of different stand ages to climate, this paper selected the public period of growth of P. sylvestris plantations for growth-climate response analysis, that is, 1980–2021.
In order to exclude the reduction in tree growth caused by non-drought and to more accurately select drought events, we combined PDSI with the trend of change in tree basal area growth. We define that if the annual average PDSI is moderate drought and below (Supplementary Table S2), and the BAI of trees has a significant downward trend, the year is a drought year, that is, a drought event (Figures 3A,B), and defined drought events lasting more than 2 years as extreme drought events. Finally, the drought events in the study area since the 1980s were determined to be 1981, 1982, 1983, 1989, 2000, 2001, 2002, 2003, 2004, 2007, 2008, 2009, and 2015 (Figures 3A,B). In order to compare the drought events of different stand ages in the same period, the drought years 2000, 2001, 2002, 2003, 2004, 2007, 2008, 2009, and 2015 were selected during the public period of young and middle-aged stands. In fact, after the severe spring, summer, fall and winter drought in 2000, the study area was still a four-season continuous drought in 2001, 2002, 2003, and 2004. Therefore, 2000–2004 was considered as an extreme drought event for 60 consecutive months, and 2007, 2008 and 2009 were also extreme drought events for 36 consecutive months. Therefore, three drought events (2000–2004, 2007–2009, 2015) occurred in the last 20 years (2000–2020) during the growth of trees in the study area were selected for the study (Figures 3A,B).
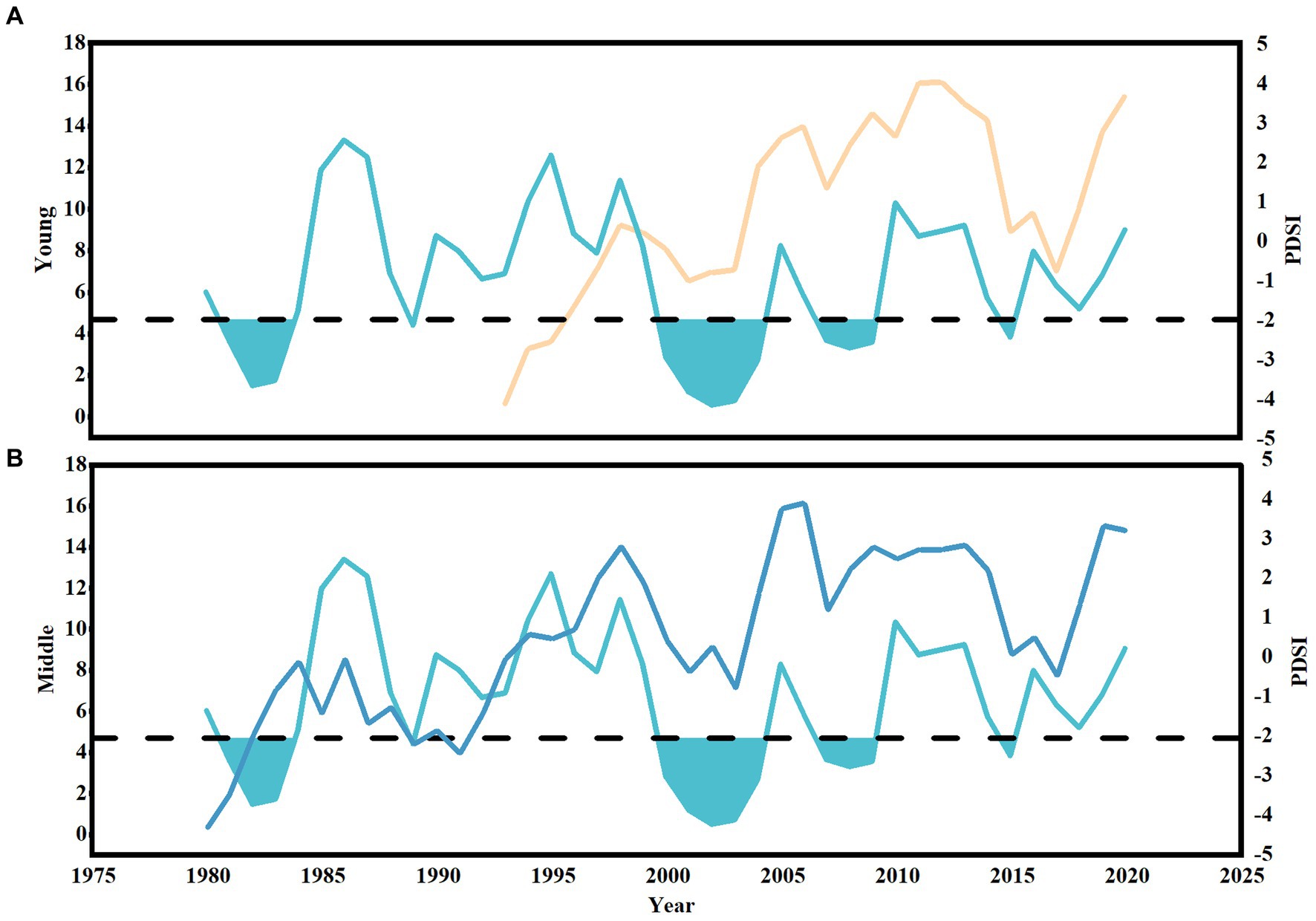
Figure 3. Drought event screening. The changes of BAI growth and annual average Palmer Drought Severity Index (PDSI) with time in young stands (A) and middle-aged stands (B). The yellow and dark blue lines represent the annual BAI growth of young and middle-aged Pinus sylvestris var. mongolica, respectively. The light blue line represents the annual average PDSI value change, and the blue filling part is the year of drought event.
Following the method of Lloret et al. (2011), we quantified the response of RG of trees of different stand ages to drought stress in different indicators: Rt, Rc and Rs. The calculation formula is as follows:
Among them, Gprev and Gpost are the BAI of trees in 3 years before and after the drought event, Gd is the BAI in the drought year, and if the drought event exceeds 1 year, it is the average BAI during the drought event. Rt quantifies the intensity of tree growth inhibition in the drought event (Gd) relative to the predrought event (Gprev). The higher the value, the stronger the resistance of trees. When Rt = 1, it indicates that the drought event does not limit the growth of trees. Rt > 1 indicates that the growth of trees in drought years is better than in non-drought years, and the resistance of trees is strong. Rt < 1 indicates that tree growth decreases during drought events. Rc is defined as the ratio of the BAI of trees after the drought event (Gpost) to that in the year of the drought event (Gd). Rc = 1 means that trees grow normally after the drought event and are not affected. Rs is defined as the ratio of tree growth after the drought event (Gpost) to that in the year before the drought event (Gprev). Rs = 1 indicates that trees are not affected by drought events and return to their original growth level. Rs > 1 indicates that the growth rate of trees exceeds the growth rate before drought events and that tree resilience is strong. Rs < 1 indicates that trees do not return to the growth level before drought events (Lloret et al., 2011). Considering the ‘legacy effect’ of trees after drought, the reference period before and after drought was set to 3 years in this study. All these indicators were calculated at the individual tree level.
Pearson correlation analysis was used to test the correlation between the ring-width index (RWI) of P. sylvestris and climatic factors (monthly lowest temperature, MLT; monthly highest temperature, MHT; monthly average temperature, MAT; monthly precipitation; monthly PDSI). Considering the lag effect of climate, the climate data of the previous year and the current year were selected. Single factor analysis of variance and significance test were used to compare the differences in resistance, recovery, resilience among different ages of stands of P. sylvestris. All statistical analysis levels: α = 0.05. All statistical analyses were performed using SPSS Statistics 26.
Ring width index (RWI) represents the growth of tree rings. It can be seen from Figure 4 that the RWI of young and middle-aged stands are basically the same from 1995 to 2021. The RWI of the two ages of stands decreased significantly in 2000–2004, 2007–2009, 2015 and 2018, indicating that RG of P. sylvestris plantation may be affected by extreme climate change.
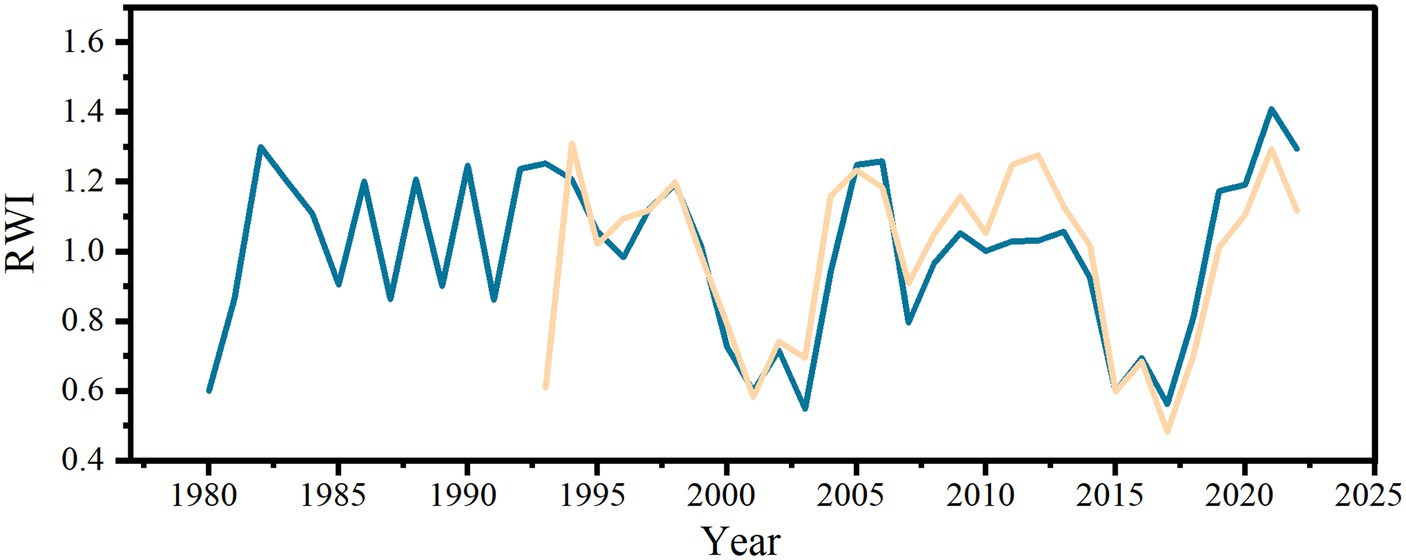
Figure 4. Change of ring-width index (RWI) of young and middle-aged stands The yellow line represents the RWI of young stands, and the blue line represents the RWI of middle-aged stands.
The RWI of young stands was positively correlated with MLT in November of the previous year and September–November of the current year (Figure 5A), and negatively correlated with MHT in September of the previous year, April and May of the current year (Figure 5B). It was positively correlated with precipitation in August of the previous year, July and November of the current year (Figure 5D), and positively correlated with PDSI in 12 months of the current year (Figure 5E). The RWI of the middle-aged stands was positively correlated with MLT in October of the previous year (Figure 5A), but not correlated with MHT (Figure 5B). It was negatively correlated with precipitation in December of the previous year (Figure 5D), and positively correlated with PDSI from June to December of the current year (Figure 5E).
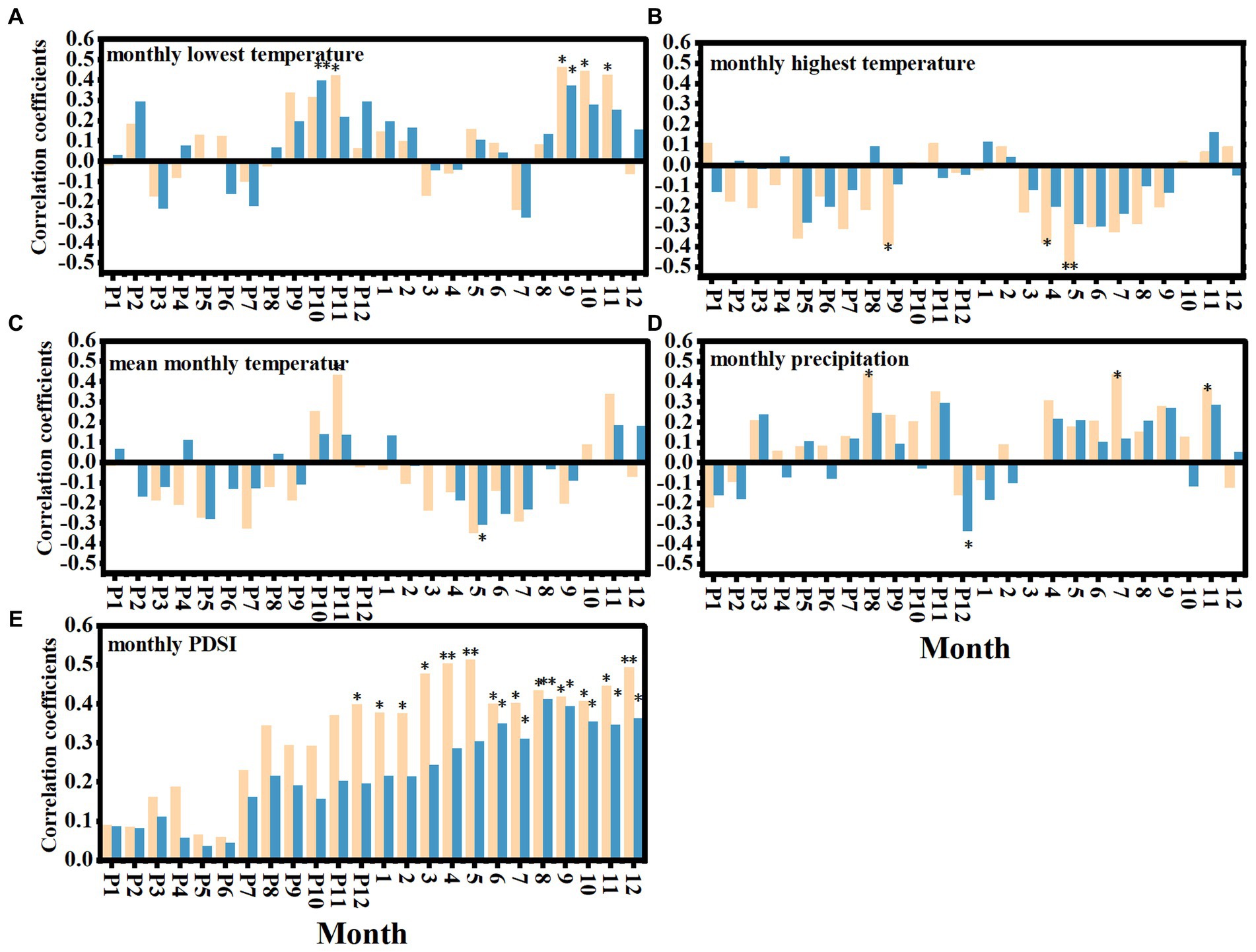
Figure 5. Correlation between different age stands and climatic factors. The correlation between different age stands and monthly lowest temperature (MLT) (A), monthly highest temperature(MHT) (B), monthly average temperature (MAT) (C), monthly precipitation (D), monthly average PDSI (E). The yellow and blue bar charts represent the correlation coefficients between the RWI of young and middle-aged stands and climatic factors, respectively. The letter P represents the month of the previous year, * indicates that there are significant differences between different age stands (p < 0.05), ** indicates that there are extremely significant differences between different age stands (p < 0.01).
There was no significant difference in Rt between the first two drought events in the two stand ages, and it was significantly lower than the first two in the third drought event (Figures 6A,D). Rc in the first drought event was significantly higher than that in the second and third drought events, but there was no difference between the latter two (Figures 6B,E). The Rs of the young stands decreased significantly in the three drought events, and the Rs of the latter two drought events of the middle-aged stands were significantly lower than that of the first drought event, but there was no significant difference between the latter two (p < 0.05) (Figures 6C,F). During the first drought event, the average value of Rt in the young stands was greater than 1, and the average values of Rc and Rs in the two-aged stands were also greater than 1. However, after three drought events, only the average value of RC was greater than 1, while the average values of Rt and Rs were less than 1, indicating that most trees can recover radial growth level after the first drought. However, multiple consecutive drought events have a serious impact on the resistance and resilience of trees. Although the growth after the drought event will be better than that of the year in which the drought occurred, it will not return to the growth level before the drought event. Multiple drought events have seriously reduced the radial growth level of trees.
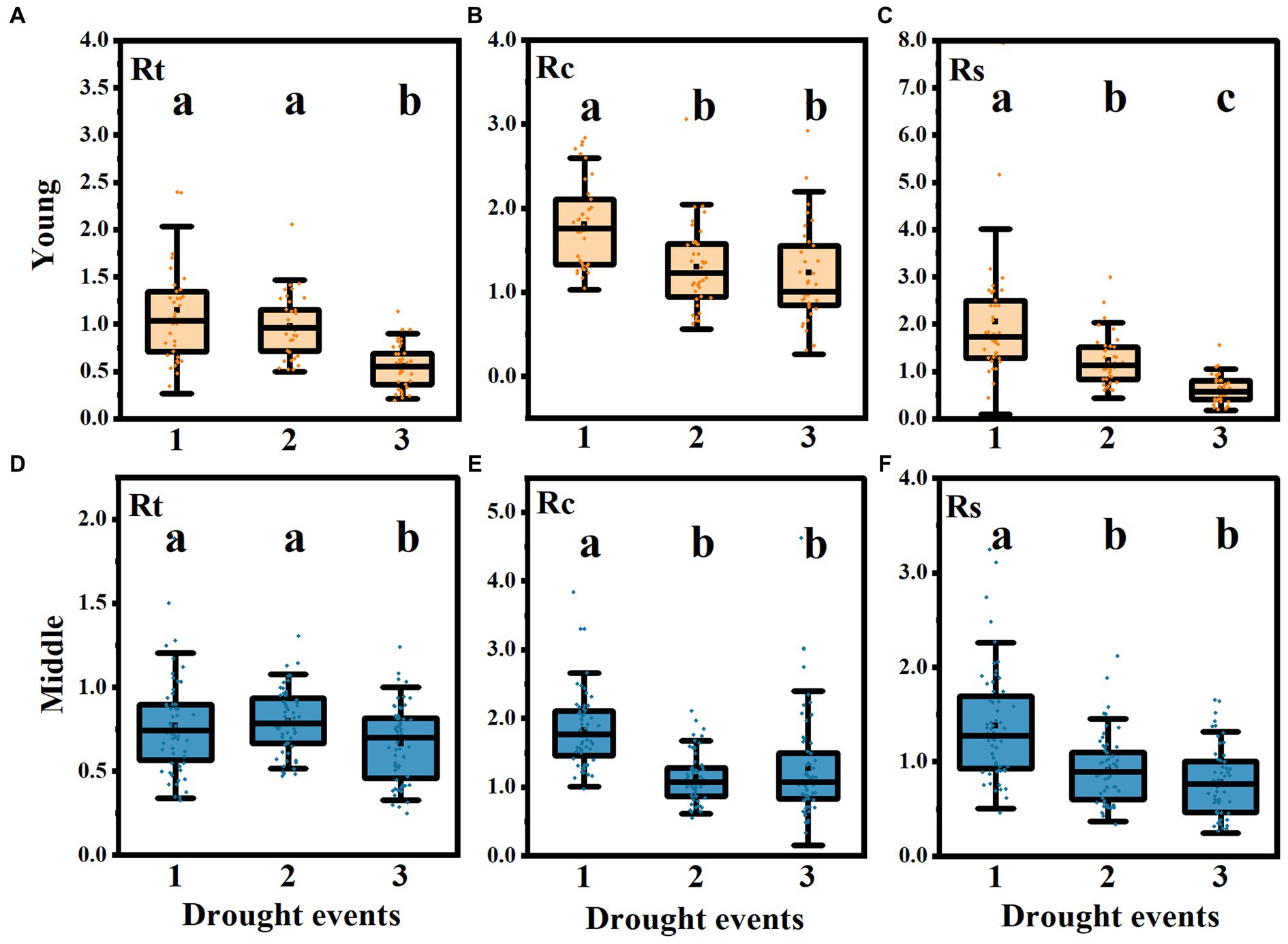
Figure 6. Difference analysis of Rt, Rc and Rs under different drought events. The changes of resistance (Rt) (A), recovery (Rc) (B), and resilience (Rs) (C) in young group and Rt (D), Rc (E), Rs (F) in middle-aged stands in three drought events. Black dots represent the median; the bottom and top of the box represent the first and third quartiles, respectively. The extension line represents the most extreme data points in the 1.5 times quartile range (the same below). Different letters indicated that there were significant differences between drought events (p < 0.05).
The average Rc values of the young stands and the middle-aged stands after the three drought events were 1.23 and 1.27 (p < 0.05), respectively, and there was no significant difference between the two stands (Figure 7B). The average Rt values of the young stands and the middle-aged stands after the three drought events were 0.55 and 0.66 (p < 0.05), respectively, and the average Rs values were 0.61 and 0.78 (p < 0.05), respectively, and there were significant differences in Rt and Rs between the two stands.
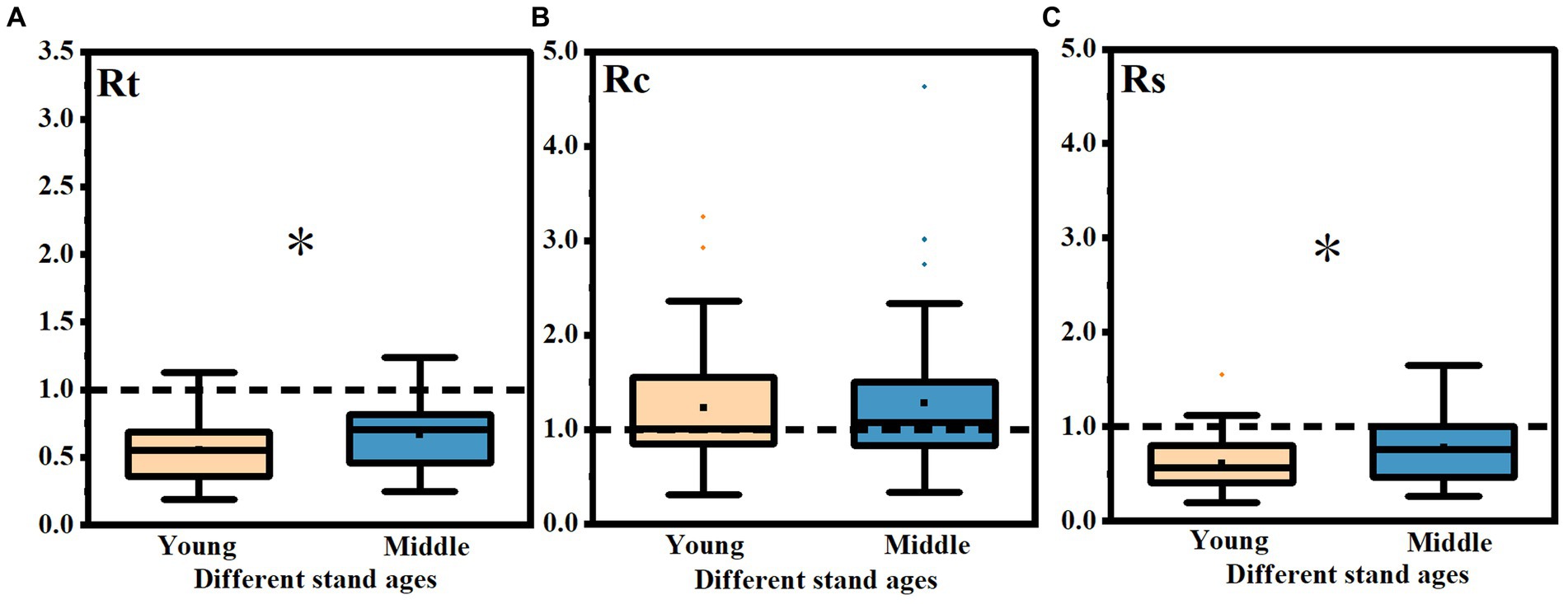
Figure 7. Difference analysis of Rt, Rc and Rs in different age stands after three drought events. The differences of Rt (A), Rc (B) and Rs (C) in different age stands after three drought events. * indicated that there were significant differences between different age stands (p < 0.05).
The Rt and Rs of the young stands were significantly lower than those of the middle-aged stands (Figures 7A,C), indicating that the resistance and resilience of the young stands were worse, and the adaptability of the middle-aged stands to extreme drought was better.
Although the overall growth trends of young and middle-aged P. sylvestris are similar (Figure 4), the correlation between the two stand ages and climate is not the same. The RWI of young stands was negatively correlated with MHT in April and May of the current year, while the RWI of middle-aged stands was not correlated with the MHT. The RWI of the young stands was positively correlated with precipitation in August of the previous year, July and November of the current year, while the RWI of the middle-aged stands was only negatively correlated with precipitation in December of the previous year. The RWI of the young stands was positively correlated with PDSI of the 12 months of the current year, and the RWI of the middle-aged stands was positively correlated with PDSI only from June to December of the current year. The above results indicate that the young stands are more sensitive to high temperature, precipitation and PDSI than the middle-aged stands (Figure 5), which supports our first hypothesis and is consistent with the results of many studies (vieira et al., 2006; kontera et al., 2016; Christopoulou et al., 2022). Arco Molina et al. (2016) and Gurskaya and Shiyatov (2006) found that the bark of trees thickened with age, resulting in a stronger insulation and protective capacity, thereby reducing the sensitivity of older trees to climate change. In addition, the soil desertification in the semi-arid area of western Liaoning is severe, the surface water retention capacity is low, the root system of young stands is shallow, and less groundwater available than in middle-aged forests. When the temperature increases and precipitation decreases, the available water of young trees is less, which also explains why young stands are more sensitive to climate change (Haijun, 2015). The RWI of young stands was negatively correlated with MHT in April and May in the spring of the year. This may be due to the fact that in the semi-arid area of western Liaoning, although the temperature was relatively high in June, July and August, it concentrated more than 66% of the annual precipitation (Figure 2), which can alleviate the inhibitory effect of high temperature on tree growth. In April and May, the precipitation is less, but the temperature is second only to June, July and August, which is more prone to spring drought (Mingshu et al., 2016). The increase in temperature will increase the evaporation of soil moisture and increase the evapotranspiration of trees. If the precipitation is not timely, or the precipitation is low and the soil moisture is not sufficient, the trees are susceptible to drought stress. Therefore, the high temperature and low precipitation in April and May inhibited the growth of young stands more than that in June, July and August. Similarly, the growth of young stands was positively correlated with MLT in the fall of the year, and the growth of middle-aged stands was positively correlated with MLT in September of the year. It was also because the low temperature reduced the transpiration of trees and reduced the evaporation of water, which was beneficial to the growth of trees. This is similar to the spring drought of pines in France (Merlin et al., 2015). However, the study of Peng et al. (2018) showed that pine trees were subjected to high temperature stress in summer, which was different from our research results. The possible reason is that its research is located in the humid climate zone of central China, and the annual rainfall is abundant. Even if the temperature rises, the trees still have enough water to maintain normal growth and are not affected by drought.
In the growth process of P. sylvestris, there were multiple growth reductions in the young and middle-aged stands (Figure 4), and its growth reduction corresponded to the reduction of PDSI (Figure 3), indicating that drought events had a certain negative effect on the growth of trees. However, our study found that most trees recovered their radial growth level 3 years after the first drought event (RS > 1, Figure 6), which is different from the first part of our second hypothesis. This may be due to the compensation effect of trees (Arend et al., 2016). Compensatory growth is a typical physiological response or domestication strategy of trees to compensate for losses during drought. When the drought event ends and growth conditions become favorable, trees will grow faster to compensate for the lack of drought, which also shows that trees have some resistance and resilience. Compensatory growth effects are widespread in forests around the world (Granda et al., 2013; Bose et al., 2021; Ovenden et al., 2021), which also shows that many studies are consistent with our findings. However, our study also found that when pine trees experienced multiple consecutive drought events, their resistance and resilience decreased rapidly, especially after the third drought event, almost all pine trees could not recover to the pre-drought level (Rt < 1, Rs < 1, Figure 6), which is consistent with the research conclusions of Serra-Maluquer et al. (2018) and Mueller et al. (2005), which is also consistent with the cumulative effect of our second hypothesis. Serra-Maluquer et al. (2018) and Mueller et al. (2005) believe that the failure to recover to the previous level after multiple drought events is related to the previous drought events. The compensation effect of trees is not infinite. The cumulative effect of continuous drought events will offset the compensation effect of drought events and damage the resistance of species to disasters and the ability to resilience after drought. We speculate that this phenomenon is caused by drought-induced xylem cavitation damage in trees. This change reduces the available water in trees, and continuous drought events aggravate the hydraulic degradation caused by xylem cavitation. Even if the drought event passes, the hydraulic degradation is irreversible and has long-term effects, eventually preventing the tree from returning to its original growth level (Anderegg et al., 2013). At the same time, we also found that although the third drought event was only a moderate drought, its severity and duration were much smaller than the first and second extreme droughts, but the Rs was still significantly reduced in the young groups, indicating that the cumulative effect of the drought events was very severe. We speculate that the resilience of trees will be more affected when drought events occur again. With the increase of drought events in the future, the cumulative effect will continue to increase until the trees die and the forests decline on a large scale.
After three events, there were significant differences in Rt and Rs between the two stand ages (Figure 7), which was consistent with our third hypothesis. The Rt and Rs of the middle-aged stands were significantly higher than those of the young stands, indicating that the middle-aged stands had stronger adaptability to extreme drought events, which was consistent with the results of Colangelo et al. (2017) and Versace et al. (2022), which is also consistent with our first part of the conclusion. Studies have shown that with the increase of tree age, the greater the dependence on water storage in the body, the stronger the water storage capacity of trees. During the drought period, the water that trees can absorb from the soil is limited, so the water stored in trees is very important. The more water stored in trees, the less affected by drought events (Phillips et al., 2003; Scholz et al., 2011). In addition, the stomatal closure ability of large trees is often better than that of small trees, which helps to reduce the risk of water evaporation and xylem cavitation during drought (Niinemets, 2002), which may also explain why the resistance and resilience of middle-aged stands are greater than those of young stands. However, the study by Loïc et al. (2021) shows that the age of trees has a negative effect on their resilience, contrary to our findings. This may be due to the different study sites. Their research site is located in Switzerland, which has a temperate maritime climate. Unlike semi-arid areas, where trees rely mainly on their own stored water during droughts, Switzerland has a mild climate and high annual precipitation. The water-holding capacity of the soil is better than that of the desertified sandy land. Even during droughts, groundwater is more abundant and trees are less dependent on stored water in the body. In the same stand, small trees were shaded by large trees, which reduced water evaporation (pretzsch et al., 2018; Loïc et al., 2021), so small trees were less affected in his study.
Based on the method of dendrochronology, this study investigated the response of P. sylvestris plantations in different age stands to climate change and drought events, and drew the following conclusions: Young stands display a greater sensitivity to climate change and drought events compared to middle-aged stands. In the semi-arid area, the spring drought has a more significant inhibitory impact on the growth of pine trees compared to the high summer temperatures. While pine trees have some compensation effect after drought events, the cumulative effect of repeated drought events will gradually offset the trees’ compensation effect, leading to reduced growth and death in some cases. Global warming has resulted in more frequent drought events. As a consequence, young trees are at a higher risk of death compared to middle-aged trees. In order to prevent the decay or death of trees, it is necessary to focus on the growth of young trees. In the process of forest construction, we should pay attention to the age structure of the stand, strengthen manual management and tending, and create a forest ecological environment with strong resistance and strong recovery ability.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
PL: Writing – review & editing, Conceptualization. SH: Investigation, Writing – original draft, Conceptualization. HW: Writing – review & editing, Conceptualization. WH: Investigation, Writing – review & editing. YZ: Investigation, Writing – review & editing. YW: Writing – review & editing, Conceptualization.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was funded by the Science and Technology Program of Liaoning Province (No. 2021JH2/10200007) and the Key Technology Research and Demonstration Project of Desertification Control in Northwest Liaoning.
We are very grateful to the staff of the forest farm for their great support for our experiment.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2023.1272477/full#supplementary-material
Adams, J. (2009). Vegetation-climate interaction: How plants make the global environment. Berlin, New York, Chichester: Springer in association with Praxis.
Allen, C. D., Macalady, A. K., Chenchouni, H., Bachelet, D., McDowell, N., Vennetier, M., et al. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 259, 660–684. doi: 10.1016/j.foreco.2009.09.001
Anderegg, W. R. L., Plavcová, L., Anderegg, L. D. L., Hacke, U. G., Berry, J. A., and Field, C. B. (2013). Drought's legacy: multiyear hydraulic deterioration underlies widespread aspen forest die-off and portends increased future risk. Glob. Change Biol. 19, 1188–1196. doi: 10.1111/gcb.12100
Arco Molina, J. G., Domínguez, D. P., Roig, F. A., and Martín, A. H. (2016). Tree age and bark thickness as traits linked to frost ring probability on araucaria araucana trees in northern Patagonia. Dendrochronologia 37, 116–125. doi: 10.1016/j.dendro.2016.01.003
Arend, M., Sever, K., Pflug, E., Gessler, A., and Schaub, M. (2016). Seasonal photosynthetic response of european beech to severe summer drought: limitation, recovery and post-drought stimulation. Agric. For. Meteorol. 220, 83–89. doi: 10.1016/j.agrformet.2016.01.011
Bigelow, S. W., Papaik, M. J., Caum, C., and North, M. P. (2014). Faster growth in warmer winters for large trees in a mediterranean-climate ecosystem. Clim. Chang. 123, 215–224. doi: 10.1007/s10584-014-1060-0
Bolte, A., Ammer, C., Lof, M., Madsen, P., Nabuurs, G., Schall, P., et al. (2009). Adaptive forest management in central europe: climate change impacts, strategies and integrative concept. Scand. J. For. Res. 24, 473–482. doi: 10.1080/02827580903418224
Bose, A. K., Scherrer, D., Camarero, J. J., Ziche, D., Babst, F., Bigler, C., et al. (2021). Climate sensitivity and drought seasonality determine post-drought growth recovery of quercus petraea and quercus robur in europe. Sci. Total Environ. 784:147222. doi: 10.1016/j.scitotenv.2021.147222
Christopoulou, A., Sazeides, C. I., and Fyllas, N. M. (2022). Size-mediated effects of climate on tree growth and mortality in mediterranean brutia pine forests. Sci. Total Environ. 812:151463. doi: 10.1016/j.scitotenv.2021.151463
Colangelo, M., Camarero, J. J., Borghetti, M., Gazol, A., Gentilesca, T., and Ripullone, F. (2017). Size matters a lot: drought-affected italian oaks are smaller and show lower growth prior to tree death. Front. Plant Sci. 8:135. doi: 10.3389/fpls.2017.00135
Field, C. B., Barros, V., Stocker, T. F., and Dahe, Q. (2012). Managing the risks of extreme events and disasters to advance climate change adaptation: Special report of the intergovernmental panel on climate change. Cambridge: Cambridge University Press.
Gantois, J. (2022). New tree-level temperature response curves document sensitivity of tree growth to high temperatures across a US-wide climatic gradient. Glob. Chang. Biol. 28, 6002–6020. doi: 10.1111/gcb.16313
Gazol, A., Camarero, J. J., Anderegg, W., and Vicente-Serrano, S. (2017). Impacts of droughts on the growth resilience of northern hemisphere forests. Glob. Ecol. Biogeogr. 26, 166–176. doi: 10.1111/geb.12526
Granda, E., Camarero, J. J., Gimeno, T. E., Martinez-Fernandez, J., and Valladares, F. (2013). Intensity and timing of warming and drought differentially affect growth patterns of co-occurring mediterranean tree species. Eur. J. Forest Res. 132, 469–480. doi: 10.1007/s10342-013-0687-0
Gurskaya, M. A., and Shiyatov, S. G. (2006). Distribution of frost injuries in the wood of conifers. Russ. J. Ecol. 37, 7–12. doi: 10.1134/S1067413606010024
Haijun, Z. (2015). Vigorously promote agricultural mechanization to improve rural ecological environment. Agri. Technol. 9, 46–48. doi: 10.11974/nyyjs.20150932011
Hollunder, R. K., Mariotte, P., Carrijo, T. T., Holmgren, M., Luber, J., Stein-Soares, B., et al. (2021). Topography and vegetation structure mediate drought impacts on the understory of the south american Atlantic forest. Sci. Total Environ. 766:144234. doi: 10.1016/j.scitotenv.2020.144234
Jing, M., Zhu, L., Liu, S., Cao, Y., Zhu, Y., and Yan, W. (2022). Warming-induced drought leads to tree growth decline in subtropics: evidence from tree rings in Central China. Front. Plant Sci. 13:964400. doi: 10.3389/fpls.2022.964400
Kannenberg, S. A., Maxwell, J. T., Pederson, N., D'Orangeville, L., Ficklin, D. L., and Phillips, R. P. (2019). Drought legacies are dependent on water table depth, wood anatomy and drought timing across the eastern US. Ecol. Lett. 22, 119–127. doi: 10.1111/ele.13173
Kontera, O., Buentgenb, U., Carrere, M., Timonenf, M., and Espera, J. (2016). Climate signal age effects in boreal tree-rings: lessons to be learned for paleoclimatic reconstructions. Quat. Sci. Rev. 142, 164–172. doi: 10.1016/j.quascirev.2016.04.020
Law, B., Thornton, P., Irvine, J., Anthoni, P., and Van Tuyl, S. (2001). Carbon storage and fluxes in ponderosa pine forests at different developmental stages. Glob. Chang. Biol. 7, 755–777. doi: 10.1046/j.1354-1013.2001.00439.x
Lindner, M., Maroschek, M., Netherer, S., Kremer, A., Barbati, A., Garcia-Gonzalo, J., et al. (2010). Climate change impacts, adaptive capacity, and vulnerability of european forest ecosystems. For. Ecol. Manag. 259, 698–709. doi: 10.1016/j.foreco.2009.09.023
Lloret, F., Keeling, E. G., and Sala, A. (2011). Components of tree resilience: effects of successive low-growth episodes in old ponderosa pine forests. Oikos 120, 1909–1920. doi: 10.1111/j.1600-0706.2011.19372.x
Loïc, G., Forrester, D. I., Bottero, A., Andreas, R., and Mathieu, L. (2021). Tree neighbourhood diversity has negligible effects on drought resilience of european beech, silver fir and Norway spruce. Ecosystems 24, 20–36. doi: 10.1007/s10021-020-00501-y
Longo, M., Knox, R. G., Levine, N. M., Alves, L. F., Bonal, D., Camargo, P. B., et al. (2018). Ecosystem heterogeneity and diversity mitigate amazon forest resilience to frequent extreme droughts. New Phytol. 219, 914–931. doi: 10.1111/nph.15185
Lucas-Borja, M. E., Bose, A. K., Andivia, E., Candel-Pérez, D., Plaza-Álvarez, P. A., and Linares, J. C. (2021). Assessing tree drought resistance and climate-growth relationships under different tree age classes in a Pinus nigra Arn ssp. salzmannii forest. Forests 12:1161. doi: 10.3390/f12091161
Martín-Benito, D., Cherubini, P., del Río, M., and Cañellas, I. (2008). Growth response to climate and drought in Pinus nigra Arn. Trees of different crown classes. Trees 22, 363–373. doi: 10.1007/s00468-007-0191-6
Merlin, M., Perot, T., Perret, S., Korboulewsky, N., and Vallet, P. (2015). Effects of stand composition and tree size on resistance and resilience to drought in sessile oak and scots pine. For. Ecol. Manag. 339, 22–33. doi: 10.1016/j.foreco.2014.11.032
Mingshu, C., Baoli, S., and Ke, S. (2016). Analysis of causes of spring drought in Fuxin area and defense countermeasures. Friends of Farmers Getting Rich. 2:294. doi: 10.3969/j.issn.1003-1650.2016.02.280
Mueller, R. C., Scudder, C. M., Porter, M. E., Trotter, R. T. III, Gehring, C. A., and Whitham, T. G. (2005). Differential tree mortality in response to severe drought: evidence for long-term vegetation shifts. J. Ecol. 93, 1085–1093. doi: 10.1111/j.1365-2745.2005.01042.x
National Forestry Administration (2017). Age-class and age-group of main tree species. Beijing: China Standard Press.
Niinemets, Ü. (2002). Stomatal conductance alone does not explain the decline in foliar photosynthetic rates with increasing tree age and size in Picea abies and Pinus sylvestris. Tree Physiol. 22, 515–535. doi: 10.1093/treephys/22.8.515
Ning, P., Zhang, M., Bai, T., Zhang, B., Yang, L., Dang, S., et al. (2023). Dendroclimatic response of pinus tabuliformis carr. Along an altitudinal gradient in the warm temperate region of China. Front. Plant Sci. 14:1147229. doi: 10.3389/fpls.2023.1147229
Oliver, T. H., Heard, M. S., Isaac, N. J. B., Roy, D. B., Procter, D., Eigenbrod, F., et al. (2015). Biodiversity and resilience of ecosystem functions. Trends Ecol. Evol. 30, 673–684. doi: 10.1016/j.tree.2015.08.009
Ovenden, T. S., Perks, M. P., Clarke, T.-K., Mencuccini, M., and Jump, A. S. (2021). Life after recovery: increased resolution of forest resilience assessment sheds new light on post-drought compensatory growth and recovery dynamics. J. Ecol. 109, 3157–3170. doi: 10.1111/1365-2745.13576
Palmer, W. C. (1965), Meteorological drought. Weather Bureau Research Paper 45, U.S. Dep. of Commerce, Washington, D. C.
Peng, J. F., Peng, K. Y., and Li, J. B. (2018). Climate-growth response of Chinese white pine (Pinus armandii) at different age groups in the Baiyunshan National Nature Reserve, Central China. Dendrochronologia 49, 102–109. doi: 10.1016/j.dendro.2018.02.004
Phillips, N. G., Ryan, M. G., Bond, B. J., McDowell, N. G., Hinckley, T. M., and Cermák, J. (2003). Reliance on stored water increases with tree size in three species in the pacific northwest. Tree Physiol. 23, 237–245. doi: 10.1093/treephys/23.4.237
Pretzsch, H., Schütze, G., and Biber, P. (2018). Drought can favour the growth of small in relation to tall trees in mature stands of Norway spruce and european beech. For. Ecosyst. 5, 1–19. doi: 10.1186/s40663-018-0139-x
Scholz, F. G., Phillips, N. G., Bucci, S. J., Meinzer, F. C., and Goldstein, G. (2011). “Hydraulic capacitance: biophysics and functional significance of internal water sources in relation to tree size” in Size-and age-related changes in tree structure and function (Dordrecht: Springer), 341–361.
Serra-Maluquer, X., Mencuccini, M., and Martínez-Vilalta, J. (2018). Changes in tree resistance, recovery and resilience across three successive extreme droughts in the northeast iberian peninsula. Oecologia 187, 343–354. doi: 10.1007/s00442-018-4118-2
Smerdon, J. E., Cook, B. I., Cook, E. R., and Seager, R. (2015). Bridging past and future climate across paleoclimatic reconstructions, observations, and models: A hydroclimate case study*. J. Clim. 28, 3212–3231. doi: 10.1175/JCLI-D-14-00417.1
Song, C., and Woodcock, C. E. (2003). A regional forest ecosystem carbon budget model: impacts of forest age structure and landuse history. Ecol. Model. 164, 33–47. doi: 10.1016/S0304-3800(03)00013-9
Sun, J., Liu, Y., Sun, B., and Wang, R. (2012). Tree-ring based PDSI reconstruction since 1853 AD in the source of the fenhe river basin, Shanxi province, China. Sci. China: Earth Sci. 55, 1847–1854. doi: 10.1007/s11430-012-4369-4
Tang, X., Li, H., Ma, M., Yao, L., Peichl, M., Arain, A., et al. (2017). How do disturbances and climate effects on carbon and water fluxes differ between multi-aged and even-aged coniferous forests? Sci. Total Environ. 599-600, 1583–1597. doi: 10.1016/j.scitotenv.2017.05.119
Trugman, A. T., Detto, M., Bartlett, M. K., Medvigy, D., Anderegg, W. R. L., Schwalm, C., et al. (2018). Tree carbon allocation explains forest drought-kill and recovery patterns. Ecol. Lett. 21, 1552–1560. doi: 10.1111/ele.13136
van Mantgem, P., Stephenson, N. L., Byrne, J. C., Daniels, L. D., Franklin, J. F., Fulé, P. Z., et al. (2009). Widespread increase of tree mortality rates in the western United States. Science 323, 521–524. doi: 10.1126/science.1165000
Versace, S., Bräuning, A., Cherubini, P., Febbraro, M. D., Häusser, M., Lombardi, F., et al. (2022). New evidence for population-specific responses to drought events from tree ring chronologies of pinus nigra ssp. laricio across the entire distribution range. Agric. For. Meteorol. 323:109076. doi: 10.1016/j.agrformet.2022.109076
Vieira, J., Campelo, F., and Nabais, C. (2006). Age-dependent responses of tree-ring growth and intra-annual density fluctuations of pinus pinaster to mediterranean climate. Trees 23, 257–265. doi: 10.1007/s00468-008-0273-0
Vitali, V., Büntgen, U., and Bauhus, J. (2017). Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in South-Western Germany. Glob. Chang. Biol. 23, 5108–5119. doi: 10.1111/gcb.13774
Yan, H., Wang, S., Lu, H., Yu, Q., Zhu, Z., Myneni, R. B., et al. (2014). Development of a remotely sensing seasonal vegetation-based palmer drought severity index and its application of global drought monitoring over 1982-2011. J. Geophys. Res. Atmos. 119, 9419–9440. doi: 10.1002/2014JD021673
Yuanqiao, L., Xiuchen, W., Huang, Y., Li, X., Fangzhong, S., Zhao, S., et al. (2021). Compensation effect of winter snow on larch growth in Northeast China. Clim. Chang. 164, 1–17. doi: 10.1007/s10584-021-02998-1
Yuzhang, Z. (1990). Liaoning Jilin, Heilongjiang Province, Zhangzi pine plantation investigation report. J. Liaoning For. Sci. Technol. 2, 52–57.
Zang, C., Hartl-Meier, C., Dittmar, C., Rothe, A., and Menzel, A. (2014). Patterns of drought tolerance in major european temperate forest trees: climatic drivers and levels of variability. Glob. Chang. Biol. 20, 3767–3779. doi: 10.1111/gcb.12637
Zhang, Y., Peng, C., Zhang, G., Liu, G., Chen, H., Wang, J., et al. (2016). Multiple afforestation programs accelerate the greenness in the ‘three north’ region of China from 1982 to 2013. Ecol. Indic. 61, 404–412. doi: 10.1016/j.ecolind.2015.09.041
Keywords: Pinus sylvestris var. mongolica, different stand ages, drought events, resistance, resilience, compensation effect, cumulative effect, climate–growth relations
Citation: Liu P, Hu S, Wei H, He W, Zhou Y and Wang Y (2023) Response of radial growth of Pinus sylvestris var. mongolica of different stand ages to climate and extreme drought events in the semi-arid region of western Liaoning, Northeast China. Front. For. Glob. Change. 6:1272477. doi: 10.3389/ffgc.2023.1272477
Edited by:
Andrei G. Lapenas, Albany State University, United StatesCopyright © 2023 Liu, Hu, Wei, He, Zhou and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yutao Wang, eXR3NzMwQHN5YXUuZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.